
Submitted:
12 June 2024
Posted:
14 June 2024
You are already at the latest version
Abstract
Hazelnut (Corylus avellana L.) is of great agroeconomic importance, prized for its nutritional value and antioxidant properties. Its yield and quality may differ due to factors such as season (S), locality (L) and cultivar (C). Our objective was to determine the dynamics of morphological features and antioxidant properties in nuts of Barcelona and Tonda di Giffoni (TDG) cultivars planted at two localities (Cunco and Perquenco) during three productive seasons (S1, S2 and S3). Barcelona grown in Perquenco in S1 and S3 (less precipitation, more solar radiation), performed better than in Cunco with respect to most morphological nut, kernel and shell features, and the same parameters generally had the same trends for TDG in S2 and S3. Among the seasons at the same localities, the differences between the predominant condition defects for both cultivars were in the decreasing order wrinkled/deformed kernels, double kernels, blank and mold ker-nels, with about 75% of hazelnuts without defect. In both cultivars, the shell had higher antioxi-dant properties that increased in S1 and S3 at both localities, and were consistently higher than in the kernels. Lipid peroxidation was higher in the kernel than shell in both cultivars and local-ities for S1 and S3. The results of this study suggest that the different hazelnut cultivar features including physical and chemical components depend on season and locality, which offers im-portant clues for all those interested in the hazelnut industry.
Keywords:
antioxidant activity
; morphological features
; oxygen radical absorbance capacity
; phenolic compounds
1. Introduction
Native from Central Europe and Western Asia, hazelnut (Corylus avellana L.) is an important commercial crop with an annual world production of approximately 1,255,700 tons (t) of shelled nuts with Turkey as the main producer (75%), followed by Italy (5%), USA (5%), Azerbaijan (4%) and Chile (4%) [1]. In Chile, the planted area comprises 37,000 hectares (ha) concentrated between El Maule and La Araucanía regions, with a total production of 52,000 t for the 2022/2023 season, becoming as the main producer in the Southern Hemisphere due to its advantage for off-season hazelnut production [2]. The most planted cultivars are Barcelona and TDG (Tonda di Giffoni) covering 90% of the surface, showing mean yields from 2500 to 3000 t ha−1 [2,3].
Morphological features such as industrial components of hazelnuts may differ due to weather, locality, cultivar, harvest season and even orchard management. Król et al. [4], examined the influence of cold climatic conditions in Poland and genotypic characteristics on cultivars such as Barceloński, Kataloński, Webba Cenny, Olbrzym z Halle, Cosford and Nottinghamski, determining that morphological features, such as weight, diameter and width differ genotypically. Whereas Manterola-Barroso et al. [5] reported that in TDG cultivar morphological features were affected by localities characterized by temperate climatic influences in foothills, obtaining greater nut length (21 mm), nut weight (3.1 g), kernel yield (42%), shell weight (1.8 g) and thickness (1.9 mm) than in orchard planted in central valley under 100 m.a.s.l. Moreover, these authors determined a locality effect concerning total phenolic content (TPC), radical scavenging activity (RSA) and antioxidant capacity (AC) and concluded that planting locality affected the antioxidant properties of shell within a season. In fact, those nuts produced in orchard planted in foothills showed a greater antioxidants component. Additionally, Gülsoy et al. [6] studied three altitude conditions in different localities on hazelnut kernel antioxidant at 100 (Yağlı and Sivri), 350 (Kara and Palaz) and 800 (Çakıldak) m a.s.l. and determined that cultivar and location together mainly affected phenolic compounds and antioxidant activity, however altitudes alone did not directly affect the antioxidant capacity of hazelnuts analyzed in the study. Meanwhile, Solar et al. [7] determined higher kernel TPC in cultivars Merveille de Bollwiller, Pauetet, Barcelona, Tonda Gentile delle Langhe (TGDL) and TDG cultivars grown in localities from Spain and Northern Italy, and lower in Slovenia and France. This may be due to higher solar irradiance values in northern latitudes, which increased TPC.
Although the significant influence of some factors on morphological features and some antioxidant properties has been demonstrated [3,4,5,6], there is a lack of research investigating the influence and adaptation of different hazelnut structures during different consecutive harvesting seasons, planting locality and cultivar. Therefore, the objective of our study was to determine the adaptive behavior of morphological features and antioxidant properties in kernel and shell of Barcelona and TDG cultivars planted in La Araucanía region during three productive seasons.
2. Materials and Methods
2.1. Experimental Sites and Plant Material
Experiments were performed in two commercial hazelnut orchards planted in Cunco (38°58′19″ S; 72°07′33″ W; 400 m a.s.l) and Perquenco (38°25′33″ S; 72°21′16″ W; 299 m a.s.l), La Araucanía region, Chile, characterized as temperate area. Nuts were harvested from cultivars Barcelona and TDG trees (nine to twelve years-old) during 2019/2020 (S1), 2020/2021 (S2) and 2021/2022 (S3) productive seasons. Orchard trees were established both in multi- and mono-stem systems, under regular rows in frames of 5 × 3.5 m (571 trees ha−1) and 5 × 3 m (667 trees ha−1) for Cunco and Perquenco respectively. In all studied season and orchards, the nut harvest was performed between March and April using two vacuums (V1200E, Cifarelli S.p.A, Voghera, Italy). Within harvest, nuts were gently transported to Laboratory of Plant Physiology and Nutrition for Fruit Crops (Faculty of Agricultural Sciences and Environment, Universidad de La Frontera) and disposed for stabilization at 40 °C by forced air oven (Heratherm OGS100, Thermo Scientific, Waltham, MA, USA) to reach 6% of humidity and stored at room temperature until analysis. The weather conditions of the hazelnut orchards at the time of the experiments are shown in Figure 1 (temperature, relative humidity, precipitation and solar radiation). Data were downloaded from Agromet INIA [8] weather stations installed in the locations evaluated.
2.2. Morphological Features with Industrial Value
A total of 12 experimental units were evaluated, where each experimental unit consisted of 100 stabilized hazelnuts. The following morphological features were measured in whole nuts, kernel and shells- length (mm), width (mm), thickness (mm), weight (g), nut roundness index (NRI), kernel roundness index (KRI), internal cavity (IC, mm) and kernel yield (%). To measure the length, thickness, width and IC, a digital caliper (CALDI-6MP, Truper, CDMX, Mexico) was used and an analytical balance for weight (Model BA2204B, Biobase Meihua Trading, Jinan, China).
The nut roundness index (NRI), kernel roundness index (KRI) and kernel yield (%) were calculated using the following equations [9]:
NRI = (nut width + nut thickness)/(2 × nut length)
KRI = (kernel width + kernel thickness)/(2 × kernel length)
Kernel yield (%) = (kernel weight/nut weight) × 100
The frequency of four major defects, including blank, wrinkled or misshapen kernels, double kernels and molds, in hazelnuts was evaluated. For this, a total of 30 nuts were randomly selected, broken and inspected for each season, locality and cultivar and each defect type was recorded.
2.3. Preparation and Extraction of Low Molecular Weight Antioxidants
All hazelnut nut samples that were analyzed in triplicate (n = 30) were ground separately in a grinder (Sindelen, MOL-165, Santiago, Chile) for 3 min. The shell samples were mechanically cracked using a traditional nutcracker and subsequently ground in an ultra-centrifugal mill (Retsch, model ZM 200, Haan, Germany), obtaining the shell dust sieved at <2 µm. The samples were stored in plastic bags at −80 °C until use.
Different extraction times were used for kernel and shell, since the extraction yield of phenolic compounds and the antioxidant capacity of these extracts depend on the structural diversity from which they are derived. Kernel samples (0.2 g) according to the protocol of [3] were homogenized in liquid nitrogen and immediately mixed with 2 mL 80% ethanol. The samples were then centrifuged at 13,000 rpm for 5 min at 4 °C in a microcentrifuge (Sciloc, model D3024R, NY, USA). Subsequently, the supernatant was recovered and transferred to amber Eppendorf tubes, to be stored at −20 °C until use.
On the other hand, the powdered shell samples (1 g) according to the protocol of [5] were transferred to a 15 mL falcon tube adding 10 mL 80% ethanol and disposed in a shaker incubator (Labwit, model ZWY-100D, Shanghai, China) at 180 rpm for 8 h at 37 °C. Extracts were centrifuged at 10,000 rpm for 10 min at 4 °C (5804 Eppendorf centrifuge, Hamburg, Germany) and subsequently, the supernatant was recovered and transferred to amber vial tubes and stored at −20 °C until analytical procedures.
2.4. Total Phenolic Compounds (TPC)
The determination of TPC was carried out using the Folin-Ciocalteu method [10] with minor modifications. A mix of 10 µL of ethanolic extract, 100 µL of Folin Ciocalteu reagent, 300 µL of 7% w/v sodium carbonate and 590 µL of deionized water was prepared. Afterwards, the solution was incubated at 20 °C for 30 min in the dark. Absorbance was measured against the reagent blank at a wavelength of 765 nm on a UV/VIS spectrophotometer (SP-8001, Metertech, Taipei, Taiwan) and absorbance data were analyzed and interpreted by interpolation using a gallic acid (GA) calibration curve (0 to 25 µg mL−1). All data were expressed as GA equivalents (GAE).
2.5. Free Radical Scavenging Activity (RSA)
To determine the RSA, the methodology proposed by Brand-Williams et al. [11] was adapted, where 100 µL of sample and 1.9 mL of radical DPPH was added per tube, the mixture was shaken vigorously for 30 s and disposed at room temperature for 30 min in the dark. The absorbance was measured against the reagent blank at 517 nm in a UV-Visible spectrophotometer (SP-8001, Metertech, Taipei, Taiwan) and absorbance data were analyzed and interpreted by interpolation using a Trolox (TE) calibration curve (0 to 1000 µM). All data are expressed as Trolox equivalents (μmol TE 100 g−1 DW).
2.6. Antioxidant Capacity (AC)
The ORAC in kernel and shell samples was evaluated using the methodology implemented by the Universidad de La Frontera No. 2021-A-8614 [5]. This procedure was performed in triplicate with a multimode reader (Synergy H1 Hybrid, Biotek, Winooski, VT, USA). Solutions of 2,2′-azobis(2-amidinopropane) dihydrochloride (AAPH) (153 mM) and 70 nM fluorescein were prepared. The methodology was carried out with the Gen5™ software. First, 25 µL of AAPH and 150 µL of fluorescein (70 nM) were preincubated at 37 °C for 15 min. Then 25 µL of each sample were loaded into black 96-well Jet BIOFIL microplates, the outer wells were filled with 250 µL of distilled H2O and incubated at 37 °C measuring their kinetics every minute for 60 s for a total of 2.5 h. Finally, data were calculated using a Trolox calibration curve (0 to 250 µM) and area under the curve (AUC). Data were expressed as Trolox equivalents (µmol TE 100 g−1 DW).
2.7. Lipid Peroxidation (LP)
As an indicator of oxidative stress, lipid peroxidation was determined in shell and kernel using the thiobarbituric acid-reactive substance assay (TBARS), according to the modified protocol of Du and Bramlage [12] by Meriño-Gergichevich et al. [3] for hazelnut. The analysis was carried out using a Synergy H1 Hybrid multimode reader plate reader, Biotek (Charlotte, VT, USA) taking three reading measurements at 440, 532 and 600 nm to correct the interference generated by the TBARS-sugar complexes. Results were expressed as nanomolecular equivalents of malondialdehyde (MDA) content per dry weight (nmol MDA g‒1 DW), a secondary product of polyunsaturated fatty acid oxidation.
2.8. Experimental Design and Statistical Analysis
The experiment was performed in a completely randomized factorial design where the factors were three seasons (S), two localities (L) and two cultivars (C) (3 × 2 × 2). All commercially relevant features measured (whole nut, kernel and shell features) were subjected to a normality test and a three-way analysis of variance (ANOVA). The values obtained for TPC, RSA, AC and LP in kernel and shell, were subjected to a normality test and a three-way analysis of variance (ANOVA) where factors were S, L and nut material (NM) as kernel or shell. The comparison of means according to the Tukey’s post-hoc test at significance level of p < 0.05. All determination were performed by triplicate (12 experimental units). To explain the relationships between morphological features, antioxidant properties and the variables studied, a Pearson correlation test and a Principal Component Analysis (PCA) performed in R version 4.2.1.
3. Results
3.1. Experimental Sites and Weather Conditions
In Cunco, the lowest mean temperature was recorded on October 1 (10.3 °C) and the highest on January 21 (16.1 °C) for S1. In the S2 and S3, the lowest was recorded on 14 October (10.4 and 10.7 °C) and the highest on 4 February (17.1 and 17.2 °C) (Figure 1A). The monthly relative humidity for S1 was 77.9% in November and the lowest in February was 70.7%. In the S2, the maximum was 81.4% in November and the minimum was 74.1% in January. In the S3, the maximum was 81.8% in March and the minimum was 68.8% in February (Figure 1B). The highest accumulated monthly precipitation in the S1 was 92 mm and the lowest was 10.6 mm. In the S2, the highest was 131.6 mm and the lowest was 9.2 mm. In the S3, the maximum was 109.9 mm and the minimum was 13 mm (Figure 1C). The lowest solar radiation was recorded on 19 March (12.9 and 10.4 MJ m2) and the highest on 12 December (18.9 and 16.2 MJ m2) for S1 and S3. At S2, the lowest was recorded on March 21 (10.8 MJ m2) and the highest on January 10 (16.1 MJ m2) (Figure 1D).
In Perquenco, the lowest mean temperature was recorded on 19 October (9.9, 9.6 and 9.8 °C) and the highest on 21 February (16.3, 16.7 and 16.8 °C) for S1, S2 and S3 (Figure 1E). The monthly relative humidity for S1 was 76.4% in November and the lowest in February was 65.6%. In S2, the maximum was 78.6% in October and the minimum was 69.6% in January. In S3, the maximum was 80.6% in October and the minimum was 65.1% in February (Figure 1F). The highest monthly cumulative precipitation in S1 was 39.9 mm and the lowest was 2.5 mm. In S2, the maximum was 65.5 mm and the minimum was 6.6 mm. In S3, the maximum was 52.8 mm and the minimum was 2.4 mm (Figure 1G). The lowest solar radiation was recorded on March 23 (16.5 and 16.8 MJ m2) and the highest on December 25 (27.3 and 27.7 MJ m2) for S1 and S3. In S2, the lowest was recorded on March 23 (16.9 MJ m2) and the highest on January 25 (25.9 MJ m2) (Figure 1H).
3.2. Morphological Features in Hazelnut
Table 1 shows morphological features of harvested hazelnut and shell for all studied seasons in both localities. A significant interaction SxLxC (three-way ANOVA) was found for whole nut length, thickness and width in addition to shell weight, with Barcelona grown in Perquenco (S1 and S3) having greater nut length (5 and 15%), thickness (14 and 22%), width (13 and 16%) and shell weight (29% and 15%), compared to trees grown in Cunco (p < 0.05). For nut weight and shell thickness a significant SxC interaction showed that Barcelona reached 17% and 20% higher nut weight and 7% and 7% higher thickness shell than TDG in Cunco for S1 and S3 (p < 0.05). For NRI, Barcelona harvested in Perquenco showed values closer to 1.0 for three seasons (SxL) with a high difference (7%) between localities (LxC) (p < 0.05). Cultivar TDG grown in Cunco showed a significant decrease in seasons S2 and S3 in the length (4%) thickness (9 and 4%), width (6 and 0.05%), weight (13 and 14%) and NRI (3 and 3%) of the nuts, also in shell weight (14 and 8%) with respect to those trees (Barcelona and TDG) harvested from the Perquenco orchard (p < 0.05).
Regarding kernel morphological features (Table 2), a significant SxC interaction was found. In S3 Barcelona kernels collected from Perquenco orchard showed significant increases in length, thickness and width of 18, 43 and 31% respectively, compared to Cunco. However, in S2 the only difference between the localities for Barcelona kernels was thickness (an increase of in Perquenco 10% compared to Cunco). Whereas for kernel yield a significant SxL interaction was determined for S2 and S3 which showed that TDG in Perquenco achieved 1% and 7% higher yield than Cunco (p < 0.05). As for the TDG planted in Perquenco, the highest averages were determined for the last two seasons studied (S2 and S3); particularly in S2, kernels had higher length (1.6%), thickness (4%), width (9%), weight (13%), KRI (9%), IC (13%) and grain yield (1%) compared to Cunco.
The hazelnut defects include both physiological and biotic constraints such as blank, wrinkled or misshapen kernel, double kernel and molds (Figure 2), considered as commercial quality indicators for hazelnut industry (Table 3). Wrinkled/deformed kernel were the most frequent defects observed in the S1 (10%) and S3 (13%) seasons in Barcelona and TDG at both localities. While for S2, 7% and 13% of Barcelona hazelnuts had wrinkled/deformed kernels at both localities, TDG had the highest number of moldy hazelnuts, 3% and 17% at both locations, respectively. In addition, an average of the three seasons for Barcelona revealed that the highest percentage of defects found for both localities were wrinkled/misshapen kernel (about 10%) and the lowest was mold (0.3 and 2.3%). Likewise, in TDG, the largest defects found were wrinkled/misshapen kernels (6.6% and 9%). Finally, the average number of hazelnuts without defect for Barcelona was 76% and TDG was 74%.
3.3. Antioxidant Properties in Hazelnut Kernels and Shells
3.3.1. Total Phenolic Compounds
Regarding TPC, a significant interaction between season, locality and nut material (SxLxNM) was determined for both cultivars Barcelona (p < 0.001) and TDG (p < 0.01) (Figure 3). The shell and kernel of Barcelona and TDG in Cunco were the highest in S1 and both declined in S2 and S3 (p < 0.001) (Figure 3A,B). However, in Perquenco in the S2, both shell and kernel TPC showed 57% and 64% increase compared to the S1 and S3 (p < 0.001).
3.3.2. Free Radical Scavenging Activity
For RSA, a significant SxLxNM interaction was determined for Barcelona (p < 0.001) and TDG (p < 0.001). The shell and kernel of Barcelona had 57% and 66% higher RSA in the Cunco locality for the S1 compared to S2 and S3 (p < 0.001). However, in Perquenco, both in Barcelona shell and kernel in the first two seasons RSA was higher in comparison with the S3 (p < 0.001) (Figure 4A). For TDG the highest RSA was also in S1 in Cunco, (p < 0.001) 59% and 37% for shell and kernel, respectively compared to S2 and S3. However, in Perquenco, similarly to Cunco locality Barcelona had higher RSA than TDG in S2 and S3. The high RSA values in Perquenco for Barcelona were detected for both S1 and S2 with a decline in S3 (p < 0.001) (Figure 4B).
3.3.3. Antioxidant Capacity
For AC, measured by ORAC, a significant SxLxNM interaction was determined in Barcelona (p < 0.001) and LxNM in TDG (p < 0.001). Barcelona had higher AC, which was evidenced with for Cunco and Perquenco shell samples with averages of 60% and 48% increase for seasons S1 and S3 (p < 0.001), compared to S2. Barcelona kernels showed higher AC in S3 (p < 0.001) in both localities evaluated with an average of 55% and 62%, respectively, compared to other seasons (Figure 5A). In TDG at the Cunco locality, the highest AC was in the S3 (p < 0.01). However, the AC of TDG kernels was 61% (S1) and 64% (S3), respectively, higher compared to the S2 (p < 0.001). However, in Perquenco in S3, the highest AC was 45% for shell and 61% for kernel (p < 0.001), respectively (Figure 5B).
3.3.4. Lipid Peroxidation
For LP, a significant SxLxNM interaction was found for Barcelona (p < 0.05) but not for TDG. The kernels showed 60% higher in LP than that determined in shell, for all studied seasons, being higher in Barcelona planted in Cunco (p < 0.001). Barcelona showed higher LP in kernels, which was more intense in Cunco with an average of 64% and 61% for the S1 and S3 seasons (p < 0.001) and in Perquenco with an average of 58% for the S3 season (p < 0.001), compared to the other seasons analyzed (Figure 6A). Likewise, in the TDG cultivar, the kernel was 76% higher in LP than that determined in the shell for all seasons evaluated (p < 0.001). However, in the kernel, seasons S1 and S3 presented the highest LP in 54% and 50%, respectively, compared to S2 (p < 0.001) (Figure 6B).
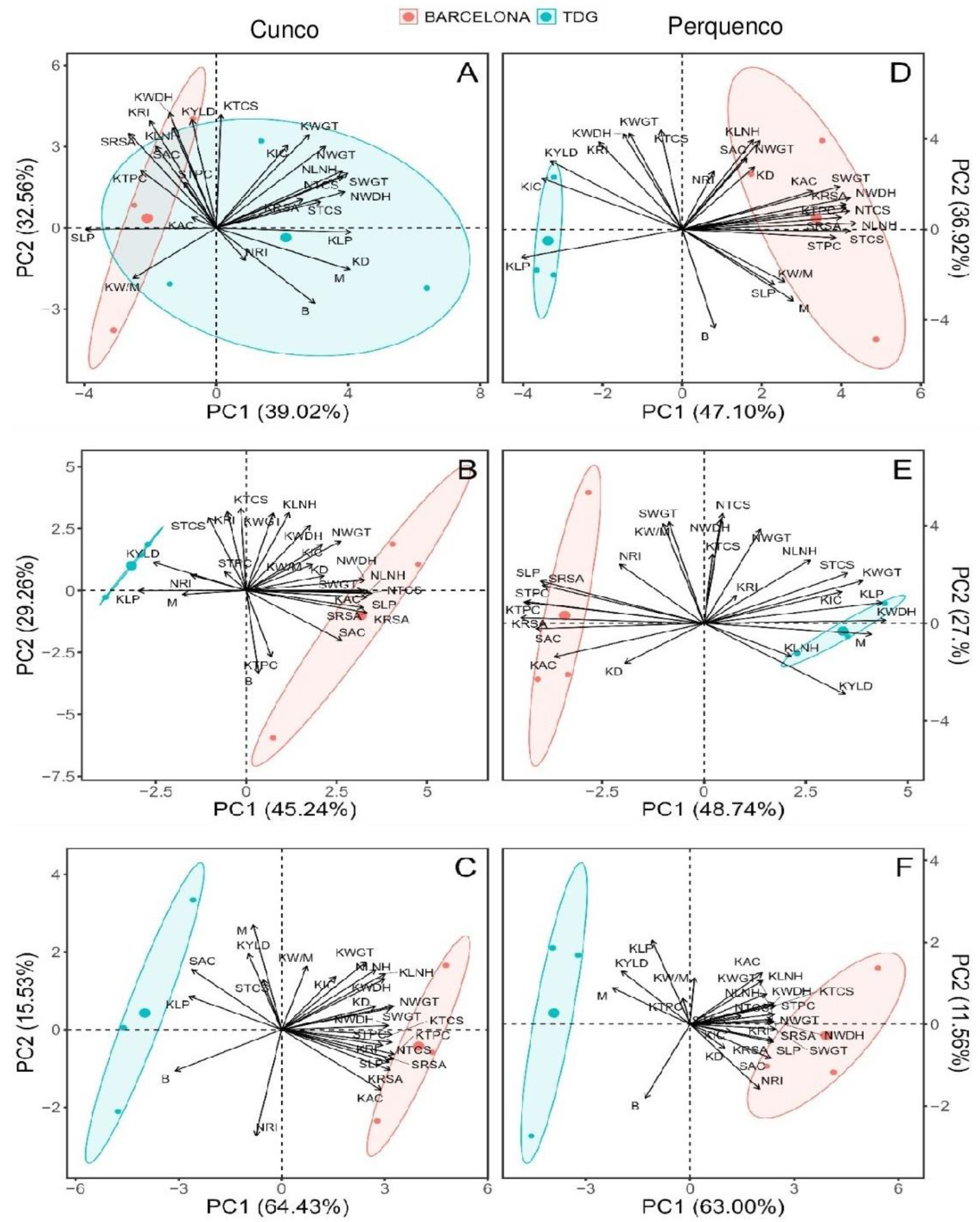
3.4. Principal Components Analysis
The data set was divided by season and subjected to principal component analysis (PCA). Confidence ellipses (group means) were generated by cultivar taking into account the effects of the factors analyzed on their morphological features and the antioxidant properties analyzed in hazelnut kernel and shell samples (Figure 7).
Regarding the components, it was observed that Barcelona grown in Cunco for S1 was mainly by higher precipitations which had positive correlations with kernel characteristics (yield, width, roundness, length, TPC) and shell (RSA, AC, TPC). While TDG influenced by low humidities showed positive correlations with most of the morphological features. However, for Barcelona in S2 positive correlations were found for NWDH, SWGT and NLNH. On the other hand, in S3 influenced by higher precipitation, positive correlations were found for KLNH, NWGT and SWGT. On the other hand, for TDG in S2 positive correlations were found only for KYLD and negative correlations with KLP. While in S3 it showed positive correlations in SAC and KLP and negative correlations with B.
However, Barcelona grown at Perquenco in S1 was influenced by lower temperatures and precipitations as well as higher radiation. It correlated positively with most of the morphological features and antioxidant properties in both kernel and shell. During that season, TDG correlated positively with KYLD and KIC. However, Barcelona grown at Perquenco in S2 correlated positively with SRSA, SLP, STPC and KTPC. While in that season, TDG correlated positively only with KWDH and negatively with M and KLNH. In S3, Barcelona under lower temperatures and precipitations showed positive correlations with KTCS, STPC and NWGT and negative correlations with SRSA, NWDH, SLP, SWGT, KRSA, SAC and NRI. In contrast, TDG showed positive correlations with KLP, KYLD and M in the same season.
Figure 7.
Principal components (PC) for the experimental variables determined in kernel and shell of the cultivars Barcelona and TDG. The graph shows the distribution of the experimental variables according to PC and grouped according to locality Cunco (A) S1, (B) S2, (C) S3, locality Perquenco (D) S1, (E) S2 and (F) S3. NLNH, nut length; NTCS, nut thickness; NWDH, nut width; NWGT, nut weight; KLNH, kernel length; KTCS, kernel thickness; KWDH, kernel width; KWGT, kernel weight; KIC, kernel internal cavity; SWGT, shell weight; STCS, shell thickness; KYLD, kernel yield; KRI, kernel roundness index; NRI, nut roundness index; SRSA, shell radical scavenging activity; KRSA, kernel radical scavenging activity; STPC, shell total phenolic compounds; KTPC, kernel total phenolic compounds; SAC, shell antioxidant capacity; KAC, kernel antioxidant capacity; SLP, shell lipid peroxidation; KLP, kernel lipid peroxidation; B, Blank; KW/M, kernel wrinkled/misshapen; KD, kernel double y M, Mold.
Figure 7.
Principal components (PC) for the experimental variables determined in kernel and shell of the cultivars Barcelona and TDG. The graph shows the distribution of the experimental variables according to PC and grouped according to locality Cunco (A) S1, (B) S2, (C) S3, locality Perquenco (D) S1, (E) S2 and (F) S3. NLNH, nut length; NTCS, nut thickness; NWDH, nut width; NWGT, nut weight; KLNH, kernel length; KTCS, kernel thickness; KWDH, kernel width; KWGT, kernel weight; KIC, kernel internal cavity; SWGT, shell weight; STCS, shell thickness; KYLD, kernel yield; KRI, kernel roundness index; NRI, nut roundness index; SRSA, shell radical scavenging activity; KRSA, kernel radical scavenging activity; STPC, shell total phenolic compounds; KTPC, kernel total phenolic compounds; SAC, shell antioxidant capacity; KAC, kernel antioxidant capacity; SLP, shell lipid peroxidation; KLP, kernel lipid peroxidation; B, Blank; KW/M, kernel wrinkled/misshapen; KD, kernel double y M, Mold.
4. Discussion
Hazelnut cultivars such as Barcelona and TDG are highly appreciated by the food industry mainly for the morphological features and antioxidant properties of their nuts [13,14], as these factors especially influence their price and exported volumes. The length, thickness, width, weight and NRI in the nut, varied significantly for Barcelona grown in Perquenco, under lower precipitation and higher radiation in S1 and S3 (Figure 1) with respect to the SxLxC interaction. These parameters were less affected by season in TDG at the Cunco locality for the seasons studied (Table 1). Our results are similar to what Özdemir and Akinci [15] and Ercisli et al. [16] reported for nut length (15.66–19.85 mm), thickness (16.17–17.28 mm), width (18.16–18.97 mm), weight (1.68–2.31 g) and NRI (0.94–1.06) in the same cultivars studied, indicating that the difference obtained in our study were attributed to individual characteristics among cultivars, as well as weather and growing conditions.
Regarding the thickness and weight of the shells in these cultivars, they were greater in Barcelona in Perquenco by 7% (1.59 mm) and 17% and 20% (1.78 g and 1.94 g) than in Cunco, under the influence of lower temperatures, humidity and precipitations and higher radiation for the S1 and S3 studied (Figure 1) with respect to the SxC interaction. Our study showed that shell thickness and weight were higher than those reported by Petriccione et al. [17], Delprete and Sesana [18], and Milošević and Milošević [19] who reported for cultivars Tonda Gentile Bilobata, TDG and TGDL, shell thickness and weight ranges from 1.17 to 1.30 mm and 1.15 g respectively. The same authors also indicated that differences between cultivar genetics, weather conditions and cultural practices directly affect in hazelnut morphological features. In addition, other morphological characteristics evaluated in kernels such as length, thickness and width for the Barcelona in Perquenco were affected by the season, showing significant increases in the last season (S3), characterized by low temperatures, in addition to higher precipitations and radiation (Figure 1), with respect to the SxC interaction.
Likewise, for TDG in Perquenco, SxL interaction was determined reaching greater length, thickness, width, weight, KRI, IC and yield, influenced by lower temperatures, humidities and precipitation, in addition to higher radiations for S2 and S3 (Figure 1) compared to the same cultivar planted in Cunco (Table 2). Our results obtained for both Barcelona and TDG are concomitant with studies conducted by Ferrão et al. [9], Özdemir and Akinci [15], Milošević and Milošević [19], who reported similar ranges of values for kernel length (11.19–17.06 mm), thickness (11.97–14.02 mm), width (12.60–16.05 mm), weight (0.81–1.76 g) and NRI (0.99–1.12). Moreover, the average kernel yield was similar to that reported by Ferrão et al. [9], Mehlenbacher et al. [20], Solar and Stampar [21]. Meriño-Gergichevich et al. [3] reported kernel yield ranges between 37-53%, similar to those reported in our study for Barcelona and TDG in both localities. However, previous studies by Mohammadzedeh et al. [22], Cristofori et al. [23], Ilić et al. [24] and Correia et al. [25] determined lower kernel yields (35%) than those recorded in our study, which is mainly due to the insufficient amount of water required for nut filling with kernel during kernel growth, therefore the lower the water supply, the lower the yield, this situation was similar to our observations in the S1 and S3 season for Barcelona grown in Perquenco, where a kernel yield of 37% was obtained (Table 2) as a consequence of low accumulated precipitations of around 22.36 mm and 29.95 mm accompanied by high solar radiation between October and March (Figure 1).
The differences in nut, shell and kernel morphological features in this study could be the result of the individual properties of the hazelnut cultivars, the locality, and the prevailing weather conditions in each season studied, as well as their interaction with the environment, including postharvest handling, as reported by Özdemir et al. [26], Özdemir and Akinci [15].
The predominant condition defects detected in the harvested nuts of Barcelona for all seasons were wrinkled/deformed kernel (average 9.8%), double kernel (average 6.5%), blank (average 3%) and mold incidence (average 1.3). While in TDG for all seasons were wrinkled/deformed kernel (average 7.8%), mold incidence (average 7.3%), blank (average 4.9%) and double kernel (average 2%) (Table 3). Abnormally small kernels were classified as wrinkled or deformed, which is a defect with low heritability being frequent in cultivars with large nut such as Barcelona, in which the highest incidence percentages were obtained for seasons S2 and S3 in both locations analyzed (Table 3). In addition, this condition could be considered more severe in years with abundant harvest, drought, herbicide damage or some other type of stress [27,28], which coincides with our results, since the seasons where this defect was most present were characterized by low temperatures and precipitations accompanied by higher radiation (Figure 1).
The incidence of moldy kernels is moderately heritable, moreover, this disorder is a serious defect that can substantially decrease shelf life [4,27,28]. This disorder has been associated with postharvest pathogenic fungi such as Cladosporium spp., Botrytis cinerea, Aspergillus spp., Penicillium spp., Diaporthe spp. and Trichotecium roseum in hazelnuts grown in southern Chile [29]. The mold incidence depends on favorable environmental factors for infection by phytopathogens, especially during flowering and fruit set [30,31,32,33]. However, in our study, the highest incidence of molds was detected in TDG grown in Perquenco corresponding to the last two seasons, in which low temperatures, humidity and precipitation were present, in addition to higher radiation during production (Figure 1).
While double kernels are undesirable due to reduced quality, size and increased irregular shapes, a ratio of 1 to 5% is considered acceptable [27]. Furthermore, this condition defect is primarily associated with the number of ovules present in the ovary, that is, as the number of ovules in the ovary increases, the number of double kernels also increases. Furthermore, double kernel frequency is largely a cultivar characteristic [27]. According to Beyhan and Marangoz [34], Palaz and Tombul cultivars contained 24% to 39% more double kernels compared to Barcelona (6.5%) and TDG (2%). While in this study the highest incidence of double kernels was for the cultivar Barcelona in both localities in S1 and S3 (Table 3). Blank nuts cause a significant crop loss in hazelnut production [35], and this is one of the most serious disorders, characterized by embryo failure due to seminal abortion, insufficient pollination or poor fertilization. The results obtained in this study indicate that the highest incidence of blank nuts for both cultivars was at both localities in the S1 and S3 (Table 3). In general, quality defects in hazelnuts include changes in morphology and flavor, and their intensity depends mainly on seasonal weather conditions. The average percentage of hazelnuts without defect in this study was 75%, which is in the range between 64 and 88% reported by McCluskey et al. [36], McCluskey et al. [37], and Guerrero et al. [38].
Our results showed an interaction between TPC, and SxLxNM for both cultivars. Both cultivars grown in Cunco, had the highest TPC in the shell followed by the kernel in S1, under higher precipitations. However, in Perquenco in S2, characterized by lower temperatures, precipitations and radiation, opposite trends were observed in the cultivars compared to S1 (Figure 3). Overall, our results are concomitant with those reported by other authors, who determined TPC in 20 hazelnut cultivars, resulting in the highest concentration for the shell and the lowest for kernel, which ranged between 159–730 and 70–478 mg GAE 100 g−1 DW, respectively, whose values were statistically significant among cultivars Barcelona, TGDL and Lewis with high antioxidant and bioactive potential [39,40,41,42,43,44,45]. In addition, our study showed greater differences in TPC between tissues (shell and kernel) and cultivars for the evaluated seasons, probably in response to the structural diversity from which the extracts are derived and the weather conditions in cultivar plantings, which directly influenced the synthesis of these compounds.
Also, previous studies have observed that variations in temperature, light exposure, UV radiation, and high-altitude environments can influence the phenolic composition and antioxidant activity of hazelnuts [6,46]. However, altitudes alone did not directly characterize phenolic composition and antioxidant activity, suggesting that variations in the aforementioned variables observed in this study were mainly influenced by cultivars rather than altitudes.
Analysis of the antioxidant capacity of hazelnut is essential from a nutraceutical point of view, as the shell and kernel are valuable sources of bioactive compounds as antioxidants [5,47,48,49,50]. Regarding the RSA determined in shell and kernel samples, our results showed differences between the three seasons analyzed due to the interaction between SxLxNM (p < 0.001) determined for both cultivars. Barcelona and TDG grown in Cunco in S1, under higher precipitation, showed higher RSA in the shell followed by the kernel. However, in Perquenco in S1 and S2, characterized by lower temperatures, precipitations and radiation, higher RSA for the shell followed by the kernel was obtained compared to S3 (Figure 4). The differences are probably caused by environmental factors, including temperature, light exposure and UV radiation, in addition to cultivars, soil type and constant seasonal changes, which have a significant effect on the RSA composition of hazelnuts [6,46]. Furthermore, our results agree with those reported by Meriño-Gergichevich et al. [3], Manterola-Barroso et al. [5] who found that antioxidant activity measured by the DPPH method in shell and kernel ranged in values of 4290–8550 and 500–2520 mg TE 100 g−1 DW, respectively. However, for different cultivars of kernel samples [40,51] reported values of 146-1280 μmol TE 100 g−1 DW, higher than those reported in this study. Despite this, previous studies demonstrated a strong interaction between shell and kernel samples, seasons and localities on RSA, probably due to physiological responses to the prevailing abiotic conditions in the commercial orchards evaluated.
ORAC methodology was used to determine the AC which is characterized by hydrogen proton transfer (HAT) based on the stabilization of a free radical (AAPH or 2,2′azobis-(2-amidinopropane) by the action of an antioxidant and the complete transfer of a hydrogen proton measures the ability of an antioxidant system to inhibit oxidative damage of molecules susceptible to AAPH radicals [52]. Regarding the AC determined in shell and kernel samples, our results showed an interaction between SxLxNM for Barcelona and LxNM for TDG (Figure 5). Barcelona and TDG grown in Cunco for S1 and S3, under lower humidities, showed higher AC in shell followed by kernel compared to S2. However, TDG in Perquenco in S3, influenced by low temperatures and high precipitations and radiation showed the highest AC for shell followed by kernel. Currently there are scarce results under this methodology on hazelnut shell and kernel samples, therefore our results were compared with previous studies performed by our laboratory staff. Data obtained in related projects [53,54] showed an average shell AC value of 50 µmol TE g−1 DW, which was extremely low compared to the average shell values reported in this study. In addition, Manterola-Barroso et al. [5] showed that AC of shell and kernel extracts ranged between 1300–4500 and 304–965 μmol TE 100 g−1 DW, respectively, coincident with our study. In addition, they concluded that the planting location determined a relevant role in the antioxidant ORAC values in shell and kernel. Other factors that influenced seasonal AC corresponded to meteorological variability and agricultural practices, as reported for almonds (Prunus dulcis L.) by Garrido et al. [55] who reported differences of 36% for ORAC values between 2004 and 2006, that is, meteorological factors present in the evaluated seasons influence the synthesis pathway of polyphenols and their AC.
On the other hand, lipid peroxidation is the most common spoilage factor of oil-rich foods and occurs when oxygen reacts with lipids especially rich in unsaturated fatty acids and a series of free radical chain reactions result in complex chemical changes. Oxidation results in rancidity and an unpleasant taste, as well as nutritional loss and the formation of hazardous compounds toxic to human health of oil-rich foods such as hazelnuts [56,57]. Factors that enhance lipid oxidation correspond to agronomic management, crop location and seasonal weather differences [58]. In our study, an SxLxNM interaction was found for Barcelona, related to seasons with low humidity but not for TDG (Figure 6). Likewise, Meriño-Gergichevich et al. [3] performed a thiobarbituric acid reaction (TBARS) assay on hazelnut TDG samples, showing an average of 705–1573 and 2164–4130 nmol MDA 100 g−1 DW in shell and kernel respectively, showing lower results in shell, similarly as obtained in our study. These findings indicate that the antioxidants present in the shell prevent the oxidation process and that the shell, being the outermost part of the hazelnut, protects the other structures of the nut from unfavorable weather conditions, improving its tolerance to abiotic stress through the increased presence of antioxidants [41,59].
5. Conclusions
In this study, we show statistically significant effects of localities and seasons on two hazelnut cultivars, their morphological features and the antioxidant properties determined in kernel and shell. These properties vary due to the influence of the meteorological conditions of each season corresponding to the planting locations evaluated. Thus, Barcelona grown in Perquenco in S1 and S3, with less precipitation and higher solar radiation, performed better than in Cunco with respect to most morphological nut, kernel and shell features, and the same parameters generally had the same trends for TDG in S2 and S3. Among seasons at the same localities, the differences between the predominant condition defects for both cultivars were in the decreasing order wrinkled/deformed kernels, double kernels, blank and mold kernels, with about 75% of hazelnuts having no defect.
On the other hand, in both cultivars, the shell had higher antioxidant properties that increased in S1 and S3 at both localities and were consistently higher than in the kernels. While LP was higher in the kernel than shell in both cultivars and localities for S1 and S3. Which means that the antioxidants present in the shell prevent the oxidation process and that the shell, being the outermost part of the hazelnut, protects the other nut structures from unfavorable weather conditions, improving its tolerance to abiotic stress through the increased presence of antioxidants.
In summary, the knowledge obtained in this study on the morphological features and antioxidant properties of hazelnut structures and the variables evaluated are fundamental, as they are necessary to determine numerous parameters and decisions of utmost importance to guarantee the quality of the final product. Finally, further studies examining each of the abiotic factors affecting the features and properties of kernel and shell structures are needed to fully explain the causes of such variations.
Author Contributions
Conceptualization, G.G.-C., C.M.-G. and F.M.; methodology, G.G.-C. and C.M.-G.; formal analysis, G.G.-C. and C.M.-G.; investigation, G.G.-C. and C.M.-G.; writing—original draft preparation, G.G.-C., C.M.-G., C.M.-B., D.P.-C. and F.M.; visualization and project administration, G.G.-C. and C.M.-G.; funding acquisition, C.M.-G. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded (partially) by Universidad de La Frontera, thesis support DIUFRO TD23-0012, Dirección de Investigación DIUFRO DI22-2001 and DI22-0045, Fondecyt 11160762 from The National Research and Development Agency (ANID) and 16PTECFS-66647 from Corporación de Fomento de la Producción (CORFO).
Data Availability Statement
The data are available from the corresponding author upon request.
Acknowledgments
The authors thank to CORFO 16PTECFS-66647, Fondecyt 11160762 and DIUFRO TD23-0012 project, Dirección de Investigación DIUFRO (DI22-0045 and DI22-2001), UFRO Doctoral scholarship, Laboratory of Plant Physiology and Nutrition for Fruit Trees, Laboratory of Soil Fertility and BIOREN-UFRO. Authors thanks to IANCO and Santa Elena Orchards for providing tree and nut material. F.M. was supported by Ministry of Education, Youth and Sports of the Czech Republic with co-financing from the EU (KOROLID, CZ.02.1.01/0.0/0.0/15_003/0000336), Czech Academy of Sciences (RVO: 60077344).
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Nuts Dried Fruits Statistical Yearbook. International Nut and Dried Fruit Council. Available online: https://inc.nutfruit.org/technical-projects/ (accessed on 25 April 2023).
- Agrichile Ferrero Hazelnut Company. Available online: https://agrichile.cl/noticias/mas-de-550-productoresparticiparon-de-las-dos-jornadas-del-meeting-anual-del-avellano-2023/ (accessed on 25 April 2023).
- Meriño-Gergichevich, C.; Luengo-Escobar, A.; Alarcón, D.; Reyes-Díaz, M.; Ondrasek, G.; Morina, F.; Ogass, K. Combined spraying of boron and zinc during fruit set and premature stage improves yield and fruit quality of European hazelnut cv. Tonda di Giffoni. Front. Plant Sci. 2021, 12, 984. [Google Scholar] [CrossRef]
- Król, K.; Gantner, M.; Piotrowska, A. Morphological Traits, Kernel Composition and Sensory Evaluation of Hazelnut (Corylus avellana L.) Cultivars Grown in Poland. Agronomy 2019, 9, 703. [Google Scholar] [CrossRef]
- Manterola-Barroso, C.; Godoy, K.; Alarcón, D.; Padilla, D.; Meriño-Gergichevich, C. Antioxidants in Shell and Nut Yield Components after Ca, Mg and K Preharvest Spraying on Hazelnut Plantations in Southern Chile. Plants 2022, 11, 3536. [Google Scholar] [CrossRef]
- Gülsoy, E.; Kaya, E.D.; Türkhan, A.; Bulut, M.; Koyuncu, M.; Güler, E.; Sayın, F.; Muradoğlu, F. The Effect of Altitude on Phenolic, Antioxidant and Fatty Acid Compositions of Some Turkish Hazelnut (Coryllus avellana L.) Cultivars. Molecules 2023, 28, 5067. [Google Scholar] [CrossRef]
- Solar, A.; Medic, A.; Slatnar, A.; Mikulic-Petkovsek, M.; Botta, R.; Rovira, M.; Sarraquigne, J.P.; Silva, A.; Veberic, R.; Stampar, F.; et al. The Effects of the cultivar and environment on the phenolic contents of hazelnut kernels. Plants 2022, 11, 3051. [Google Scholar] [CrossRef]
- Agrometeorología Red Agrometeorológica INIA. Available online: https://agrometeorologia.cl/ (accessed on 7 May 2023).
- Ferrão, A.C.; Guiné, R.P.F.; Ramalhosa, E.; Lopes, A.; Rodrigues, C.; Martins, H.; Gonçalves, R.; Correia, P.M.R. Chemical and physical properties of some hazelnut varieties grown in Portugal. Agronomy 2021, 11, 1476. [Google Scholar] [CrossRef]
- Singleton, V.L.; Rossi, J.A. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar] [CrossRef]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C.L.W.T. Use of a free radical method to evaluate antioxidant activity. LWT Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Du, Z.; Bramlage, W.J. Modified thiobarbituric acid assay for measuring lipid oxidation in sugar-rich plant tissue extracts. J. Agric. Food Chem. 1992, 40, 1566–1570. [Google Scholar] [CrossRef]
- Simsek, A.; Aykut, O. Evaluation of the microelement profile of Turkish hazelnut (Corylus avellana L.) varieties for human nutrition and health. Int. J. Food Sci. Nutr. 2007, 58, 677–688. [Google Scholar] [CrossRef]
- Silvestri, C.; Bacchetta, L.; Bellincontro, A.; Cristofori, V. Advances in cultivar choice, hazelnut orchard management and nuts storage for enhancing product quality and safety: An overview. J. Sci. Food Agric. 2020, 101, 27–43. [Google Scholar] [CrossRef]
- Özdemir, F.; Akinci, I. Physical and nutritional properties of four major commercial Turkish hazelnut varieties. J. Food Eng. 2004, 63, 341–347. [Google Scholar] [CrossRef]
- Ercisli, S.; Ozturk, I.; Kara, M.; Kalkan, F.; Seker, H.; Duyar, O.; Erturk, Y. Physical properties of hazelnuts. Int. Agrophys. 2011, 25, 115–121. [Google Scholar]
- Petriccione, M.; Ciarmiello, L.F.; Boccacci, P.; De Luca, A.; Piccirillo, P. Evaluation of ‘Tonda di Giffoni’ hazelnut (Corylus avellana L.) clones. Sci. Hortic. 2010, 124, 153–158. [Google Scholar] [CrossRef]
- Delprete, C.; Sesana, R. Mechanical characterization of kernel and shell of hazelnuts: Proposal of an experimental procedure. J. Food Eng. 2014, 124, 28–34. [Google Scholar] [CrossRef]
- Milošević, T.; Milošević, N. Determination of size and shape features of hazelnuts using multivariate analysis. Acta Sci. Pol. Hortorum Cultus. 2017, 16, 49–61. [Google Scholar] [CrossRef]
- Mehlenbacher, S.A.; Smith, D.C.; McCluskey, R.L. ‘Sacajawea’ hazelnut. Hortic. Sci. 2008, 43, 255–257. [Google Scholar] [CrossRef]
- Solar, A.; Stampar, F. Characterisation of selected hazelnut cultivars: Phenology, growing and yielding capacity, market quality and nutraceutical value. J. Sci. Food Agric. 2011, 91, 1205–1212. [Google Scholar] [CrossRef]
- Mohammadzedeh, M.; Fattahi, R.; Zamani, Z.; Khadivi-Khub, A. Genetic Identity and Relationships of Hazelnut (Corylus avellana L.) Landraces as Revealed by Morphological Characteristics and Molecular Markers. Sci. Hortic. 2014, 167, 17–26. [Google Scholar] [CrossRef]
- Cristofori, V.; Bertazza, G.; Bignami, C. Changes in kernel chemical composition during nut development of three Italian hazelnut cultivars. Fruits 2015, 70, 311–322. [Google Scholar] [CrossRef]
- Ilić, P.; Mićić, N.; Đurić, G.; Tojnko, S.; Solares, A.; Bosančić, B. Pomological identification of hazelnut cultivars (Corylus avellana L.) in plantations Bosnia and Herzegovina. Agric. Conspec. Sci. 2017, 84, 389–395. [Google Scholar]
- Correia, P.; Rodrigues, C.; Filipe, A.; Guiné, R. Evaluation of the Biometric Characteristics of Hazelnuts. In Proceedings of the FABE 2019: Food and Biosystems Engineering Conference, Crete Island, Greece, 30 May–2 June 2019. [Google Scholar]
- Özdemir, M.; Açkurt, F.; Kaplan, M.; Yıldız, M.; Löker, M.; Gürcan, T.; Biringen, G.; Okay, A.; Seyhan, F.G. Evaluation of new Turkish hybrid hazelnut (Corylus avellana L.) varieties: Fatty acid composition, a-tocopherol content, mineral composition and stability. Food Chem. 2001, 73, 411–415. [Google Scholar] [CrossRef]
- Mehlenbacher, S.A. Hazelnuts (Corylus). Genet. Resour. Temp. Fruit Nut Crops. 1991, 290, 791–838. [Google Scholar] [CrossRef]
- Mehlenbacher, S.A.; Smith, D.C.; Brenner, L.K. Variance components and heritability of nut and kernel defects in hazelnut. Plant Breed. 1993, 110, 144–152. [Google Scholar] [CrossRef]
- Guerrero Contreras, J.; Galdames Gutierrez, R.; Ogass Contreras, K.; Pérez Fuentealba, S. First report of Diaporthe foeniculina causing black tip and necrotic spot on hazelnut kernel in Chile. Plant Dis. 2020, 104, 975. [Google Scholar] [CrossRef]
- Sezer, A.; Dolar, F.S. Hazelnut kernel defects and associated fungi in three provinces in Turkey. In Proceedings of the VII International Scientific Agriculture Symposium Agrosym 2016, Jahorina, Bosnia and Herzegovina, 6–9 October 2016; pp. 1312–1318. [Google Scholar]
- Battilani, P.; Chiusa, G.; Arciuolo, R.; Somenzi, M.; Fontana, M.; Castello, G.; Spigolon, N. Diaporthe as the main cause of hazelnut defects in the Caucasus region. Phytopathol. Mediterr. 2018, 57, 320–333. [Google Scholar]
- Pscheidt, J.W.; Heckert, S.; Wiseman, M.; Jones, L. Fungi associated with and influence of moisture on development of kernel mold of hazelnut. Plant Dis. 2019, 103, 922–928. [Google Scholar] [CrossRef]
- Wiman, N.G.; Webber III, J.B.; Wiseman, M.; Merlet, L. Identity and pathogenicity of some fungi associated with hazelnut (Corylus avellana L.) trunk cankers in Oregon. PLoS ONE 2019, 14, e0223500. [Google Scholar] [CrossRef]
- Beyhan, N.; Marangoz, D. An investigation of the relationship between reproductive growth and yield loss in hazelnut. Sci. Horticulturae. 2007, 113, 208–215. [Google Scholar] [CrossRef]
- Thompson, M.M.; Lagerstedt, H.B.; Mehlenbacher, S.A. Hazelnuts 125 184 Janick J. & Moore JN. Fruit Breed. 1996, 3, 1. [Google Scholar]
- McCluskey, R.L.; Mehlenbacher, S.A.; Azarenko, A.N.; Smith, D.C. “Santiam” Hazelnuts (OSU509.064); Extension Service of the Oregon State University: Corvallis, United States of America, 2005; 3p. [Google Scholar]
- McCluskey, R.; Mehlenbacher, S.; Smith, D. “Yamhill” Hazelnuts (OSU542.102); Extension Service of the Oregon State University: Corvallis, OR, USA, 2009; p. 3. [Google Scholar]
- Guerrero, C.J.; Merino-Gergichevich, C.; Ogass, C.K.; Alvarado, N.C.; Sobarzo, M.V. Quality and condition features of hazelnut (Corylus avellana L.) cv. Barcelona grown in South-Central of Chile. Rev. Fac. Cienc. Agrar. 2015, 47, 1–14. [Google Scholar]
- Cristofori, V.; Ferramondo, S.; Bertazza, G.; Bignami, C. Nut and kernel traits and chemical composition of hazelnut (Corylus avellana L.) cultivars. J. Sci. Food Agric. 2008, 88, 1091–1098. [Google Scholar] [CrossRef]
- Jakopic, J.; Petkovsek, M.M.; Likozar, A.; Solar, A.; Stampar, F.; Veberic, R. HPLC-MS identification of phenols in hazelnut (Corylus avellana L.) kernels. Food Chem. 2011, 124, 1100–1106. [Google Scholar] [CrossRef]
- Esposito, T.; Sansone, F.; Franceschelli, S.; Del Gaudio, P.; Picerno, P.; Aquino, R.P.; Mencherini, T. Hazelnut (Corylus avellana L.) shells extract: Phenolic composition, antioxidant effect and cytotoxic activity on human cancer cell lines. Int. J. Mol. Sci. 2017, 18, 392. [Google Scholar] [CrossRef]
- Sürek, E.; Büyükkileci, A.O. Extraction of antioxidant compounds from hazelnut wastes using subcritical water. GIDA J. Food 2018, 43, 211–221. [Google Scholar]
- Yilmaz, M.; Karakaya, O.; Balta, M.F.; Balta, F.; Yaman, İ. Changes in biochemical properties in Çakıldak hazelnut cultivar depending on the inner fruit size. Akademik. Ziraat. Dergisi. 2019, 8, 61–70. [Google Scholar] [CrossRef]
- Salem, M.A.; Aborehab, N.M.; Al-Karmalawy, A.A.; Fernie, A.R.; Alseekh, S.; Ezzat, S.M. Potential Valorization of Edible Nuts By-Products: Exploring the Immune-Modulatory and Antioxidants Effects of Selected Nut Shells Extracts in Relation to Their Metabolic Profiles. Antioxidants 2022, 11, 462. [Google Scholar] [CrossRef]
- Özcan, M.M.; Uslu, N. Investigation of changes in total phenol, flavonoid, antioxidant activity, fatty acids, polyphenol and mineral profiles of hazelnut kernels dried in air, oven and microwave. JSFA Reports 2023, 3, 72–81. [Google Scholar] [CrossRef]
- Koyuncu, M.A.; Islam, A.; Küçük, M. Fat and fatty acid composition of hazelnut kernels in vacuum packages during storage. Grasas Y Aceites 2005, 56, 263–266. [Google Scholar] [CrossRef]
- Mercanlıgil, S.M.; Arslan, P.; Alasalvar, C.; Okut, E.; Akgül, E.; Pınar, A.; Geyik, P.; Tokgözoğlu, L.; Shahidi, F. Effects of hazelnut-enriched diet on plasma cholesterol and lipoprotein profiles in hypercholesterolemic adult men. Eur. J. Clin. Nutr. 2007, 61, 212–220. [Google Scholar] [CrossRef]
- Alasalvar, C.; Pelvan, E.; Topal, B. Effects of roasting on oil and fatty acid composition of Turkish hazelnut varieties (Corylus avellana L.). Int. J. Food Sci. Nutr. 2010, 61, 630–642. [Google Scholar] [CrossRef]
- Mollica, A.; Zengin, G.; Stefanucci, A.; Ferrante, C.; Menghini, L.; Orlando, G.; Brunetti, L.; Locatelli, M.; Dimmito, M.; Novellinoc, E.; et al. Nutraceutical potential of Corylus avellana daily supplements for obesity and related dysmetabolism. J. Funct. Foods 2018, 47, 562–574. [Google Scholar] [CrossRef]
- Pelvan, E.; Öktem Olgun, E.; Karadağ, A.; Alasalvar, C. Phenolic profiles and antioxidant activity of Turkish Tombul hazelnut samples (natural, roasted, and roasted hazelnut skin). Food Chem. 2018, 244, 102–108. [Google Scholar] [CrossRef]
- Arcan, I.; Yemenicioğlu, A. Antioxidant activity and phenolic content of fresh and dry nuts with or without the seed coat. J. Food Compos. Anal. 2009, 22, 184–188. [Google Scholar] [CrossRef]
- Li, H.; Parry, J.W. Phytochemical compositions, antioxidant properties, and colon cancer antiproliferation effects of Turkish and Oregon hazelnut. Food Sci. Nutr. 2011, 2, 1142–1149. [Google Scholar] [CrossRef]
- 16PTECFS-66647; Sustainability and Efficient Use of Resources in the Production of European Hazelnut (Corylus avellana L.) in South-Central Chile. CORFO: Santiago, Chile, 2017.
- Ondrašek, G. Fondecyt 11160762; Quality Traits and Fruit Yield in Hazelnut (Corylus Avellana L.) Associated with Boron and Zinc Levels and Phenological Stage of Application in Plantations of Southern Chile. University of Zagreb Faculty of Agriculture: Zagreb, Croatia, 2016.
- Garrido, I.; Monagas, M.; Gómez-Cordovés, C.; Bartolomé, B. Polyphenols and antioxidant properties of almond skins: Influence of industrial processing. J. Food Sci. 2008, 73, C106–C115. [Google Scholar] [CrossRef]
- Alasalvar, C.; Shahidi, F.; Liyanapathirana, C.M.; Ohshima, T. Turkish tombul hazelnut (Corylus avellana L.). Compositional characteristics. J. Agric. Food Chem. 2003, 51, 3790–3796. [Google Scholar] [CrossRef]
- Pan, Y.; Zhu, J.; Wang, H.; Zhang, X.; Zhang, Y.; He, C.; Ji, X.; Li, H. Antioxidant activity of ethanolic extract of Cortex fraxini and use in peanut oil. Food Chem. 2007, 103, 913–918. [Google Scholar] [CrossRef]
- Siebeneichler, T.J.; Hoffmann, J.F.; Galli, V.; Zambiazi, R.C. Composition and impact of pre-and post-harvest treatments/factors in pecan nuts quality. Trends Food Sci Technol. 2023, 131, 46–60. [Google Scholar] [CrossRef]
- Masullo, M.; Cerulli, A.; Mari, A.; de Souza Santos, C.C.; Pizza, C.; Piacente, S. LC-MS Profiling Highlights Hazelnut (Nocciola Di Giffoni PGI) Shells as a Byproduct Rich in Antioxidant Phenolics. Food Res. Int. 2017, 101, 180–187. [Google Scholar] [CrossRef]
Figure 1.
Weather conditions during the productive seasons (October-March) for 2019/2020 (S1), 2020/2021 (S2) and 2021/2022 (S3). The figure shows the weather conditions in locality Cunco (A) temperature mean (T mean, °C), (B) relative humidity (RH, %), (C) precipitation (P, mm), (D) solar radiation (SR, MJ m2), and locality Perquenco (E) T mean (°C), (F) RH (%), (G) P (mm), (H) SR (MJ m2). Information extracted from Agromet INIA https://agrometeorologia.cl/ (accessed on 7 May 2023).
Figure 1.
Weather conditions during the productive seasons (October-March) for 2019/2020 (S1), 2020/2021 (S2) and 2021/2022 (S3). The figure shows the weather conditions in locality Cunco (A) temperature mean (T mean, °C), (B) relative humidity (RH, %), (C) precipitation (P, mm), (D) solar radiation (SR, MJ m2), and locality Perquenco (E) T mean (°C), (F) RH (%), (G) P (mm), (H) SR (MJ m2). Information extracted from Agromet INIA https://agrometeorologia.cl/ (accessed on 7 May 2023).
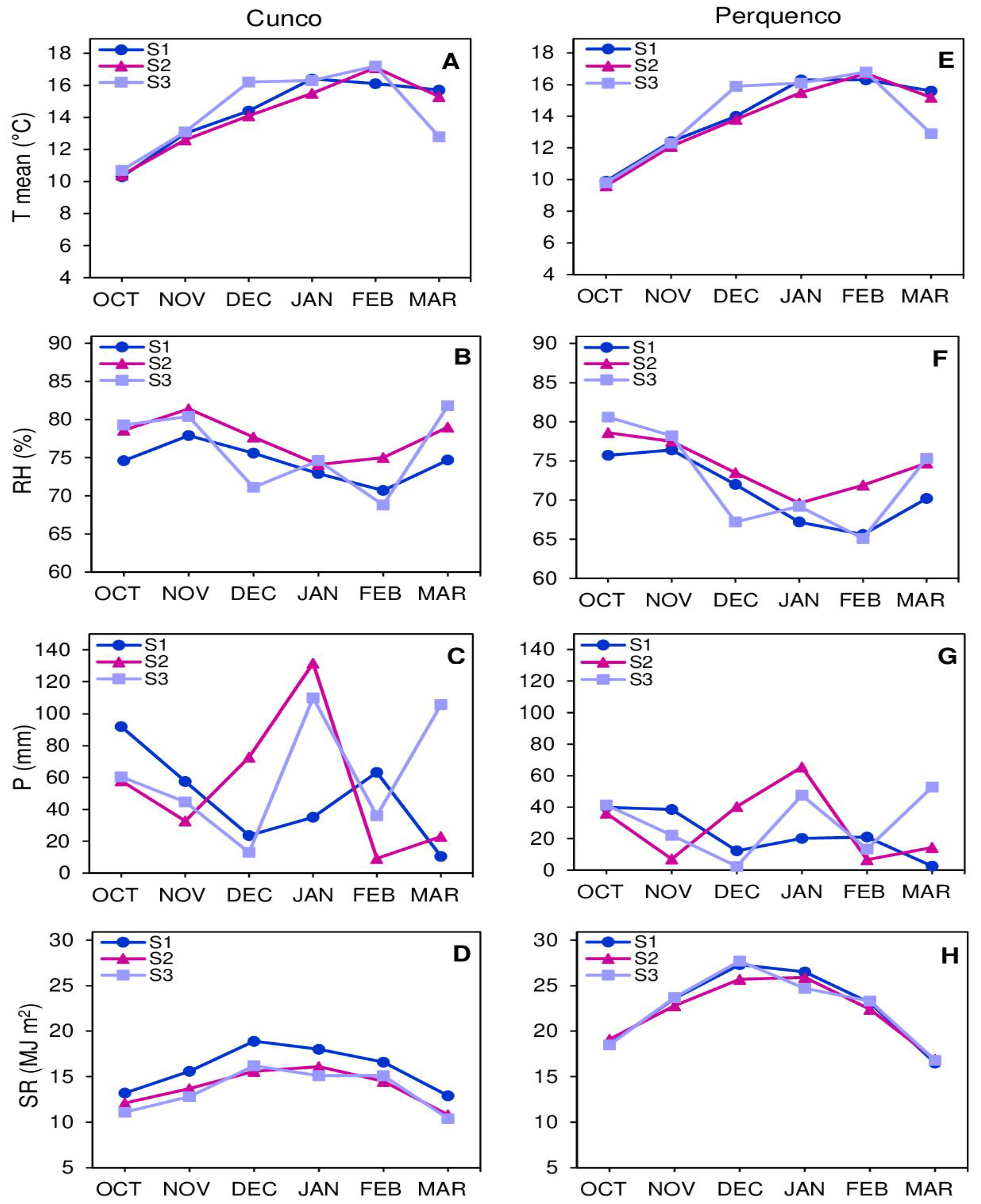
Figure 2.
Hazelnut defects: (A) blank, (B) wrinkled or misshapen kernel, (C) double kernel, and (D) molds.
Figure 2.
Hazelnut defects: (A) blank, (B) wrinkled or misshapen kernel, (C) double kernel, and (D) molds.

Figure 3.
Total phenolic compounds (mg GAE 100 g−1 DW) in kernel and shell of hazelnut Barcelona (A) and TDG (B) for three seasons 2019/2020 (S1), 2020/2021 (S2) and 2021/2022 (S3) in two localities in La Araucanía region. S-season, L-location, NM-nut material. Bars represent the average of three replicates ± S.E. Different lowercase letters indicate statistical differences (p < 0.05) between seasons for the same locality. NS, not significant; * p < 0.05; ** p < 0.01; *** p < 0.001.
Figure 3.
Total phenolic compounds (mg GAE 100 g−1 DW) in kernel and shell of hazelnut Barcelona (A) and TDG (B) for three seasons 2019/2020 (S1), 2020/2021 (S2) and 2021/2022 (S3) in two localities in La Araucanía region. S-season, L-location, NM-nut material. Bars represent the average of three replicates ± S.E. Different lowercase letters indicate statistical differences (p < 0.05) between seasons for the same locality. NS, not significant; * p < 0.05; ** p < 0.01; *** p < 0.001.
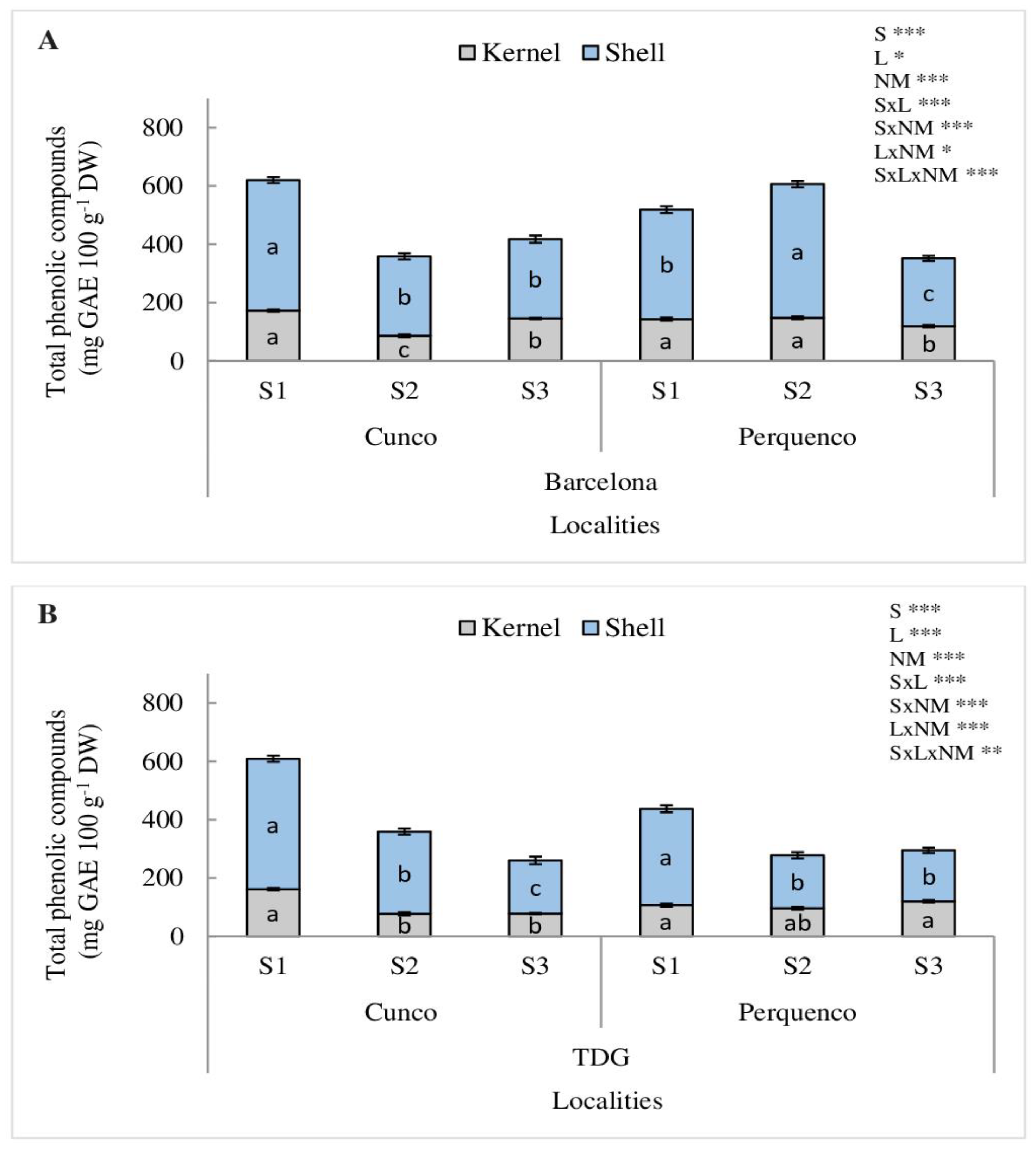
Figure 4.
Radical scavenging activity (mg TE 100 g−1 DW) in kernel and shell of hazelnut nuts Barcelona (A) and TDG (B) for three seasons 2019/2020 (S1), 2020/2021 (S2) and 2021/2022 (S3) in two localities in La Araucanía region. S-season, L-location, NM-nut material. Bars represent the average of three replicates ± S.E. Different lowercase letters indicate statistical differences (p < 0.05) between seasons for the same locality. NS, not significant; ** p < 0.01; *** p < 0.001.
Figure 4.
Radical scavenging activity (mg TE 100 g−1 DW) in kernel and shell of hazelnut nuts Barcelona (A) and TDG (B) for three seasons 2019/2020 (S1), 2020/2021 (S2) and 2021/2022 (S3) in two localities in La Araucanía region. S-season, L-location, NM-nut material. Bars represent the average of three replicates ± S.E. Different lowercase letters indicate statistical differences (p < 0.05) between seasons for the same locality. NS, not significant; ** p < 0.01; *** p < 0.001.
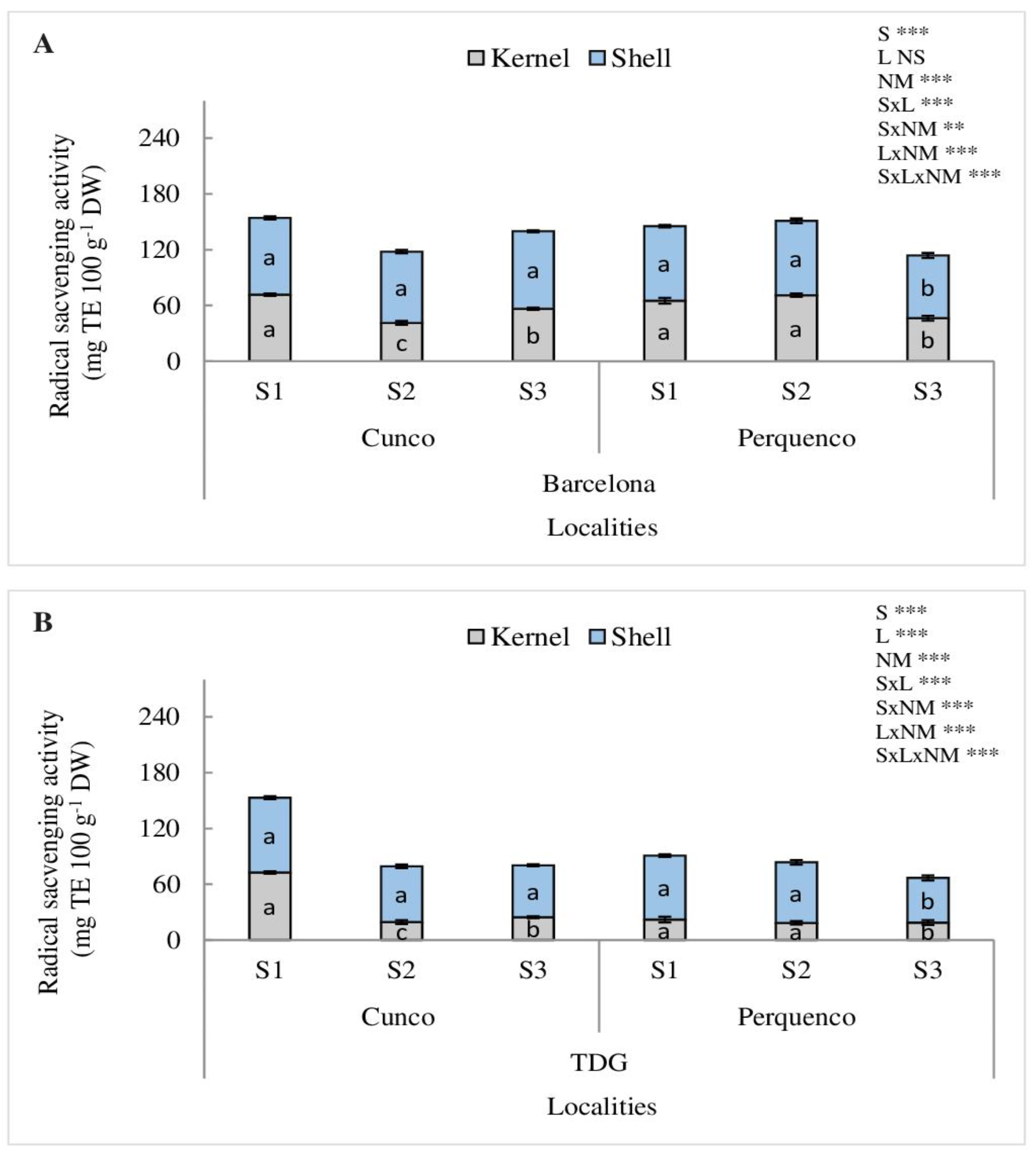
Figure 5.
Antioxidant capacity (µmol TE 100 g−1 DW) in kernel and shell of hazelnut nuts Barcelona (A) and TDG (B) for three seasons 2019/2020 (S1), 2020/2021 (S2) and 2021/2022 (S3) in two localities in La Araucanía region. S-season, L-location, NM-nut material. Bars represent the average of three replicates ± S.E. Different lowercase letters indicate statistical differences (p < 0.05) between seasons for the same locality. NS, not significant; *** p < 0.001.
Figure 5.
Antioxidant capacity (µmol TE 100 g−1 DW) in kernel and shell of hazelnut nuts Barcelona (A) and TDG (B) for three seasons 2019/2020 (S1), 2020/2021 (S2) and 2021/2022 (S3) in two localities in La Araucanía region. S-season, L-location, NM-nut material. Bars represent the average of three replicates ± S.E. Different lowercase letters indicate statistical differences (p < 0.05) between seasons for the same locality. NS, not significant; *** p < 0.001.
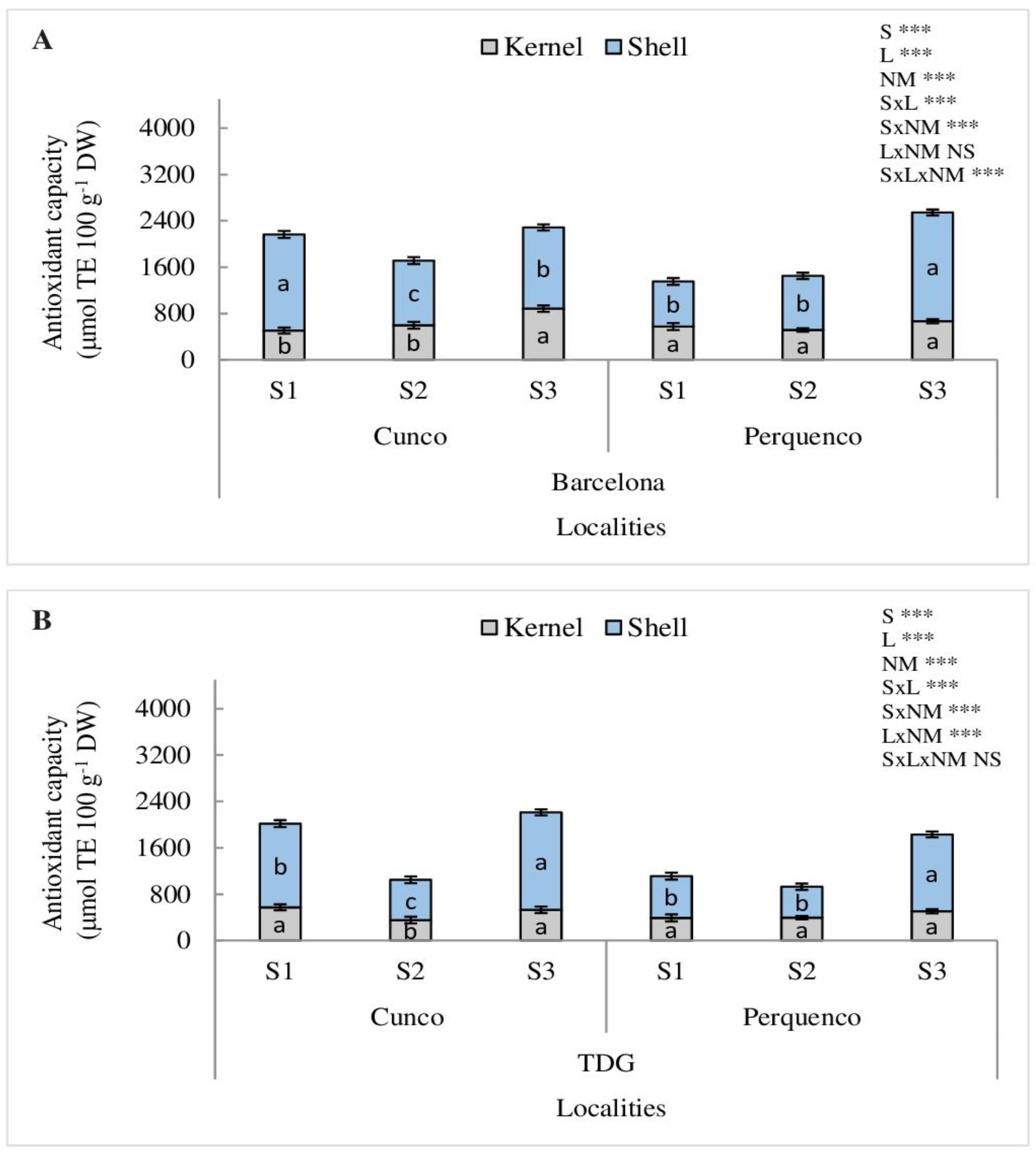
Figure 6.
Lipid peroxidation (nmol MDA 100 g−1 DW) in kernel and shell of hazelnut nuts Barcelona (A) and TDG (B) for three seasons 2019/2020 (S1), 2020/2021 (S2) and 2021/2022 (S3) in two localities in La Araucanía region. S-season, L-location, NM-nut material. Bars represent the average of three replicates ± S.E. Different lowercase letters indicate statistical differences (p < 0.05) between seasons for the same locality. NS, not significant; * p < 0.05; *** p < 0.001.
Figure 6.
Lipid peroxidation (nmol MDA 100 g−1 DW) in kernel and shell of hazelnut nuts Barcelona (A) and TDG (B) for three seasons 2019/2020 (S1), 2020/2021 (S2) and 2021/2022 (S3) in two localities in La Araucanía region. S-season, L-location, NM-nut material. Bars represent the average of three replicates ± S.E. Different lowercase letters indicate statistical differences (p < 0.05) between seasons for the same locality. NS, not significant; * p < 0.05; *** p < 0.001.
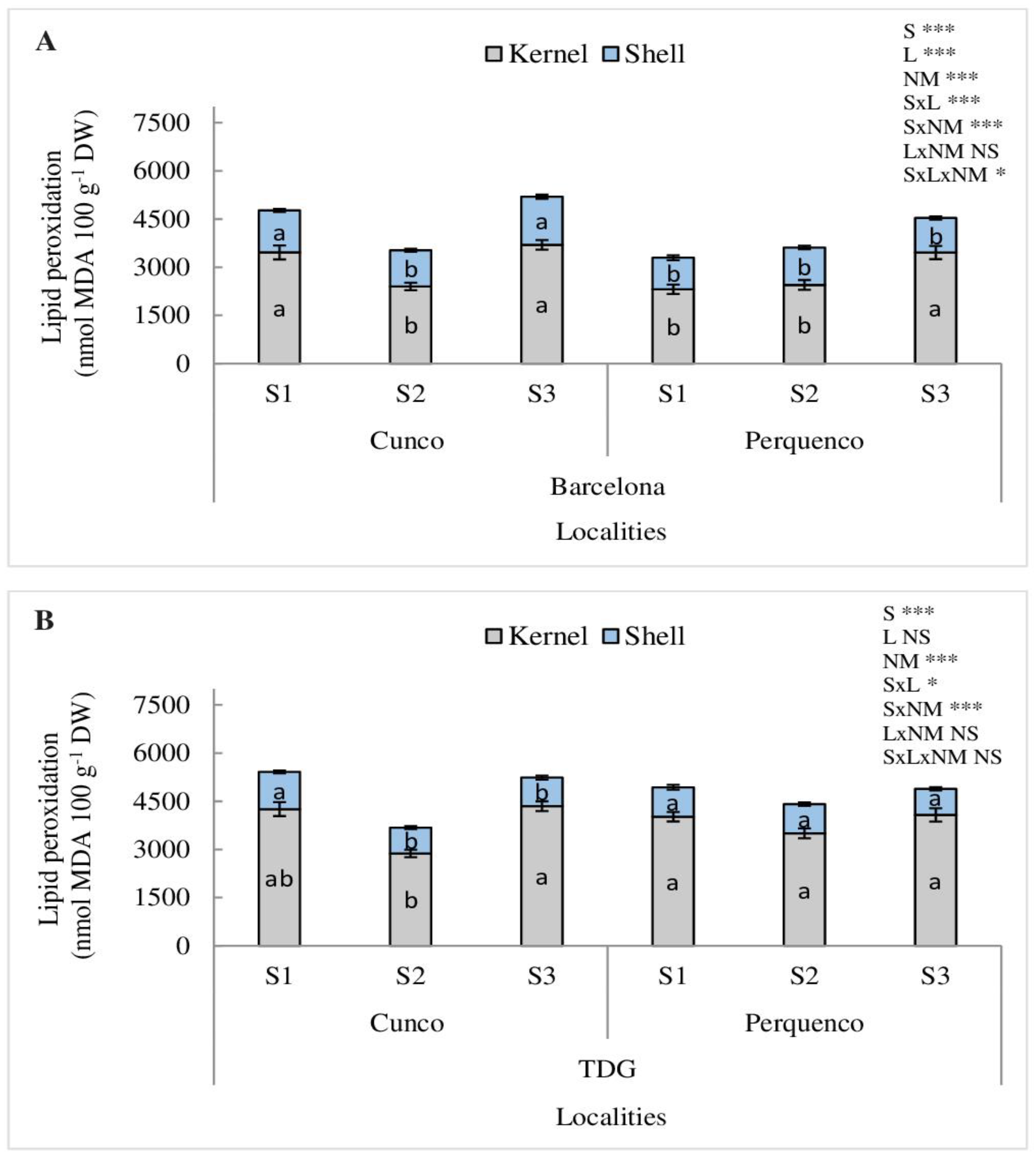

Table 1.
Nut and shell morphological parameters of hazelnut cultivars Barcelona and TDG harvested in seasons 2019/2020 (S1), 2020/2021 (S2) and 2021/2022 (S3) in Cunco and Perquenco localities, La Araucanía region. Different lowercase letters indicate statistical differences (p < 0.05) for season within locality and cultivar. NRI - nut roundness index. Values represent the average of three replicates ± S.E. NS, not significant; * p < 0.05; ** p < 0.01; *** p < 0.001.
Table 1.
Nut and shell morphological parameters of hazelnut cultivars Barcelona and TDG harvested in seasons 2019/2020 (S1), 2020/2021 (S2) and 2021/2022 (S3) in Cunco and Perquenco localities, La Araucanía region. Different lowercase letters indicate statistical differences (p < 0.05) for season within locality and cultivar. NRI - nut roundness index. Values represent the average of three replicates ± S.E. NS, not significant; * p < 0.05; ** p < 0.01; *** p < 0.001.
| Nut | Shell | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Length | Thickness | Width | Weight | NRI | Thickness | Weight | |||
| Season | Locality | Cultivar | (mm) | (g) | (mm) | (g) | |||
| S1 | Cunco | Barcelona | 19.38 ± 0.61 a | 15.95 ± 0.46 a | 18.74 ± 0.65 a | 2.49 ± 0.28 a | 0.90 ± 0.09 a | 1.48 ± 0.05 a | 1.38 ± 0.16 a |
| S2 | 20.20 ± 0.29 a | 17.11 ± 0.15 a | 19.89 ± 0.24 a | 2.77 ± 0.14 a | 0.92 ± 0.01 a | 1.42 ± 0.04 a | 1.55 ± 0.05 a | ||
| S3 | 19.79 ± 0.26 a | 16.30 ± 0.05 a | 19.64 ± 0.27 a | 2.81 ± 0.06 a | 0.91 ± 0.89 a | 1.48 ± 0.03 a | 1.68 ± 0.06 a | ||
| Mean | 19.79 ± 0.39 | 16.45 ± 0.22 | 19.42 ± 0.39 | 2.69 ± 0.16 | 0.91 ± 0.99 | 1.46 ± 0.04 | 1.54 ± 0.09 | ||
| S1 | Perquenco | 20.41 ± 0.32 ab | 18.16 ± 0.13 ab | 21.09 ± 0.29 a | 2.92 ± 0.21 a | 0.96 ± 0.01 a | 1.59 ± 0.02 a | 1.78 ± 0.07 a | |
| S2 | 19.10 ± 0.39 b | 17.72 ± 0.55 b | 20.03 ± 0.66 a | 2.90 ± 0.14 a | 0.99 ± 0.01 a | 1.43 ± 0.05 a | 1.66 ± 0.10 a | ||
| S3 | 22.80 ± 1.49 a | 19.84 ± 1.37 a | 22.82 ± 1.41 a | 3.36 ± 0.04 a | 0.96 ± 0.01 a | 1.59 ± 0.02 a | 1.94 ± 0.04 a | ||
| Mean | 20.77 ± 0.73 | 18.57 ± 0.68 | 21.31 ± 0.79 | 3.06 ± 0.39 | 0.97 ± 0.01 | 1.54 ± 0.03 | 1.79 ± 0.07 | ||
| S1 | Cunco | TDG | 20.23 ± 0.60 a | 17.01 ± 0.31 a | 20.06 ± 0.52 a | 2.80 ± 0.24 a | 0.92 ± 0.01 a | 1.57 ± 0.04 a | 1.64 ± 0.18 a |
| S2 | 18.85 ± 0.14 a | 16.20 ± 0.06 a | 18.86 ± 0.24 a | 2.61 ± 0.05 a | 0.93 ± 0.01 a | 1.49 ± 0.03 a | 1.38 ± 0.02 a | ||
| S3 | 18.54 ± 0.54 a | 15.42 ± 0.04 a | 18.31 ± 0.09 a | 2.29 ± 0.10 a | 0.92 ± 0.01 a | 1.57 ± 0.04 a | 1.31 ± 0.06 a | ||
| Mean | 19.20 ± 0.43 | 16.21 ± 0.14 | 19.08 ± 0.28 | 2.57 ± 0.39 | 0.92 ± 0.01 | 1.54 ± 0.04 | 1.44 ± 0.09 | ||
| S1 | Perquenco | 18.70 ± 0.16 a | 16.53 ± 0.31 a | 19.03 ± 0.24 a | 2.58 ± 0.17 a | 0.95 ± 0.01 a | 1.48 ± 0.02 a | 1.39 ± 0.07 a | |
| S2 | 19.63 ± 0.37 a | 17.69 ± 0.17 a | 20.01 ± 0.08 a | 2.96 ± 0.10 a | 0.96 ± 0.02 a | 1.56 ± 0.06 a | 1.58 ± 0.05 a | ||
| S3 | 19.22 ± 0.22 a | 16.01 ± 0.07 a | 18.32 ± 0.13 a | 2.61 ± 0.10 a | 0.95 ± 0.01 a | 1.48 ± 0.02 a | 1.41 ± 0.03 a | ||
| Mean | 19.18 ± 0.25 | 19.74 ± 0.18 | 19.12 ± 0.15 | 2.71 ± 0.12 | 0.95 ± 0.01 | 1.51 ± 0.03 | 1.46 ± 0.05 | ||
| Significance | S | NS | NS | NS | NS | *** | ** | NS | |
| L | NS | *** | ** | ** | *** | * | * | ||
| C | ** | *** | *** | * | NS | NS | *** | ||
| SxL | * | NS | NS | NS | * | NS | NS | ||
| SxC | * | ** | ** | * | NS | * | ** | ||
| LxC | NS | ** | ** | NS | * | NS | * | ||
| SxLxC | * | * | * | NS | NS | NS | * | ||
Table 2.
Kernel morphological characteristics in hazelnut Barcelona and TDG harvested in seasons 2019/2020 (S1), 2020/2021 (S2) and 2021/2022 (S3) in Cunco and Perquenco localities, La Araucanía region. Different lowercase letters indicate statistical differences (p < 0.05) for season within locality and cultivar. KRI-kernel roundness index, IC-internal cavity, Kernel (%)-kernel yield. Values represent the average of three replicates ± S.E. NS, not significant; * p < 0.05; ** p < 0.01; *** p < 0.001.
Table 2.
Kernel morphological characteristics in hazelnut Barcelona and TDG harvested in seasons 2019/2020 (S1), 2020/2021 (S2) and 2021/2022 (S3) in Cunco and Perquenco localities, La Araucanía region. Different lowercase letters indicate statistical differences (p < 0.05) for season within locality and cultivar. KRI-kernel roundness index, IC-internal cavity, Kernel (%)-kernel yield. Values represent the average of three replicates ± S.E. NS, not significant; * p < 0.05; ** p < 0.01; *** p < 0.001.
| Kernel | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Length | Thickness | Width | Weight | KRI | IC | Kernel | |||
| Season | Locality | Cultivar | (mm) | (g) | (mm) | (%) | |||
| S1 | Cunco | Barcelona | 14.19 ± 0.92 a | 10.51 ± 1.00 a | 13.59 ± 0.94 a | 1.16 ± 0.16 a | 0.82 ± 0.04 a | 2.40 ± 0.32 b | 45.25 ± 2.56 a |
| S2 | 14.36 ± 0.70 a | 10.81 ± 0.75 a | 13.70 ± 0.67 a | 1.21 ± 0.07 a | 0.83 ± 0.04 a | 4.08 ± 0.33 a | 43.47 ± 1.50 a | ||
| S3 | 14.45 ± 0.62 a | 10.31 ± 0.38 a | 12.85 ± 0.45 a | 1.13 ± 0.03 a | 0.75 ± 0.03 a | 2.40 ± 0.32 b | 39.19 ± 1.72 b | ||
| Mean | 14.33 ± 0.75 | 10.54 ± 0.71 | 13.38 ± 2.06 | 1.16 ± 0.09 | 0.80 ± 0.04 | 2.96 ± 0.32 | 42.64 ± 1.93 | ||
| S1 | Perquenco | 13.66 ± 0.65 a | 10.89 ± 1.37 a | 13.10 ± 1.63 a | 1.13 ± 0.17 a | 0.83 ± 0.13 a | 1.71 ± 0.41 b | 37.35 ± 4.36 b | |
| S2 | 13.73 ± 0.30 a | 11.94 ± 0.14 a | 13.70 ± 0.13 a | 1.24 ± 0.04 a | 0.93 ± 0.02 a | 2.73 ± 0.43 a | 42.31 ± 1.50 a | ||
| S3 | 17.06 ± 1.54 a | 14.70 ± 1.20 a | 16.85 ± 1.35 a | 1.43 ± 0.01 a | 0.83 ± 0.13 a | 1.71 ± 0.41 b | 37.35 ± 4.36 b | ||
| Mean | 14.82 ± 2.49 | 12.51 ± 0.90 | 14.55 ± 1.04 | 1.35 ± 0.07 | 0.86 ± 0.09 | 2.05 ± 0.42 | 39.00 ± 3.41 | ||
| S1 | Cunco | TDG | 14.13 ± 0.78 a | 10.95 ± 0.78 ab | 12.59 ± 0.84 ab | 1.23 ± 0.11 a | 0.78 ± 0.06 a | 2.50 ± 0.38 b | 42.83 ± 0.78 b |
| S2 | 14.16 ± 0.06 a | 11.45 ± 0.04 a | 13.39 ± 0.08 a | 1.22 ± 0.02 a | 0.86 ± 0.09 a | 3.67 ± 0.39 a | 46.46 ± 0.05 a | ||
| S3 | 11.83 ± 0.85 a | 8.23 ± 0.22 b | 11.07 ± 0.56 b | 1.03 ± 0.06 a | 0.78 ± 0.06 a | 2.50 ± 0.38 b | 41.62 ± 1.78 b | ||
| Mean | 13.37 ± 0.56 | 10.21 ± 0.35 | 12.35 ± 0.49 | 1.16 ± 0.06 | 0.81 ± 0.07 | 2.89 ± 0.38 | 43.64 ± 0.87 | ||
| S1 | Perquenco | 12.65 ± 0.60 a | 10.72 ± 0.63 a | 13.65 ± 0.75 ab | 1.19 ± 0.09 a | 0.90 ± 0.04 a | 2.91 ± 0.41 b | 44.26 ± 1.49 b | |
| S2 | 14.39 ± 0.38 a | 11.92 ± 0.20 a | 14.66 ± 0.07 a | 1.38 ± 0.04 a | 0.94 ± 0.03 a | 4.13 ± 0.33 a | 46.93 ± 0.24 a | ||
| S3 | 14.08 ± 0.73 a | 10.37 ± 0.46 a | 12.72 ± 0.45 b | 1.21 ± 0.06 a | 0.90 ± 0.04 a | 2.91 ± 0.41 b | 44.26 ± 1.49 b | ||
| Mean | 13.71 ± 0.57 | 11.00 ± 0.43 | 13.68 ± 0.42 | 1.26 ± 0.06 | 0.91 ± 0.04 | 3.32 ± 0.38 | 45.15 ± 1.07 | ||
| Significance | S | NS | NS | NS | NS | * | *** | NS | |
| L | NS | ** | * | NS | *** | NS | NS | ||
| C | * | * | NS | NS | NS | NS | ** | ||
| SxL | ** | * | NS | NS | NS | NS | * | ||
| SxC | * | ** | * | NS | NS | NS | NS | ||
| LxC | NS | NS | NS | NS | NS | NS | NS | ||
| SxLxC | NS | NS | NS | NS | NS | NS | NS | ||
Table 3.
The percentage of blank, wrinkled/misshapen kernel, double kernel and moldy kernel in two hazelnut cultivars Barcelona and TDG, harvested in seasons 2019/2020 (S1), 2020/2021 (S2) and 2021/2022 (S3) in Cunco and Perquenco localities, La Araucanía region (n = 30).
Table 3.
The percentage of blank, wrinkled/misshapen kernel, double kernel and moldy kernel in two hazelnut cultivars Barcelona and TDG, harvested in seasons 2019/2020 (S1), 2020/2021 (S2) and 2021/2022 (S3) in Cunco and Perquenco localities, La Araucanía region (n = 30).
| Nut | ||||||
|---|---|---|---|---|---|---|
| Blank | Wrinkled/Misshapen Kernel | Double Kernel | Mold | |||
| Season | Locality | Cultivar | (%) | |||
| S1 | Cunco | Barcelona | 3 | 10 | 0 | 0 |
| S2 | 3 | 7 | 3 | 0 | ||
| S3 | 3 | 13 | 10 | 7 | ||
| Mean | 3 | 10 | 4.3 | 2.3 | ||
| S1 | Perquenco | 7 | 3 | 13 | 3 | |
| S2 | 0 | 13 | 3 | 0 | ||
| S3 | 0 | 13 | 10 | 0 | ||
| Mean | 2.3 | 9.6 | 8.6 | 0.3 | ||
| S1 | Cunco | TDG | 7 | 10 | 3 | 7 |
| S2 | 0 | 0 | 0 | 3 | ||
| S3 | 13 | 10 | 3 | 7 | ||
| Mean | 6.6 | 6.6 | 2 | 5.6 | ||
| S1 | Perquenco | 7 | 10 | 3 | 0 | |
| S2 | 0 | 7 | 0 | 17 | ||
| S3 | 3 | 10 | 3 | 10 | ||
| Mean | 3.3 | 9 | 2 | 9 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



