Submitted:
13 June 2024
Posted:
14 June 2024
You are already at the latest version
Abstract
Abstract: Although the pathophysiology of fibromyalgia syndrome has been better understood in recent decades, a unified model of its pathogenesis and an effective therapeutic approach are still far from being realized. The main aim of this article will be to delve into the fundamental mechanisms of the pathophysiology of fibromyalgia conceptualized as stress intolerance syndrome. Using the biopsychosocial model of chronic pain syndromes, we will describe the potential role of the attachment system, C tactile fibers, and oxytocinergic system dysfunction in the pathophysiology of fibromyalgia syndrome and other central sensitivity syndromes. At the end of the article, the therapeutic implications of this new global and translational pathophysiological model will be briefly discussed.
Keywords:
fibromyalgia pathophysiology
; stress response system
; functional somatic disorders
; central sensitized syndromes
; central sensitization
; insecure attachment style
; oxytocinergic system
; small fiber neuropathy
; C tactile fibers
; C‐low‐threshold mechanoreceptors
; affective touch
1. Introduction
In clinical practice and also as clearly described by the American College of Rheumatology (ACR) diagnostic criteria [1], fibromyalgia syndrome (FMS) represents a chronic and heterogeneous primary multi-symptomatic disorder. Table 1 shows all the symptoms which, with different phenotypic expression, can be clinically reported by patients suffering from FMS.
Indeed, although widespread pain, stiffness, cognitive impairment, interrupted and non-restorative sleep, and fatigue have been considered the core symptoms of FMS, a constellation of other significant complaints, are often reported by FMS patients. In particular, the presence of anxiety and mood disorders are so important that for many years FMS was not considered a chronic pain syndrome per se, but somehow a sort of psychogenic pain which, at least in clinical psychiatric settings, it is still labeled as somatoform pain disorder or somatization disorder according to the International Classification of Diseases [2].
More recently the term Functional Somatic Disorders (FSDs) has been introduced as a consequence of recent discoveries in the field of brain-body interaction and to resolve historical controversies between the somatic and mental nature of central sensitivity syndromes (CSSs) such as FMS [3].
The complexity of FMS is even more evident if we consider the multiple pathophysiological mechanisms that have been hypothesized to be responsible for the wide spectrum of symptoms reported by fibromyalgia (FM) patients. From anomalies of the hypothalamic-pituitary-adrenal (HPA) axis and the autonomic nervous system (ANS), to dysregulation of the immune system; from the role of the central nervous system to that of the peripheral nervous system; from genetic factors to environmental components and epigenetic mechanisms.
Probably, each of these dysfunctions can have a role in the pathogenesis of FMS, but we believe it is important from both a clinical and therapeutic point of view to identify the “primum movens” capable of connecting all these potential pathophysiological mechanisms together.
In general, FMS is part of the spectrum of CSSs and chronic overlapping pain conditions (COPCs), all disorders that share central sensitization as an underlying pathophysiological mechanism [4,5].
The aim of this review is to report and analyze the main theories on the pathophysiology of FMS emerging from different fields of research focusing on the role of the bodily stress system (BSS).
In order to fill the lack of a unified theory, the main proposal of this article is to design a hypothetical and innovative pathophysiological model of FMS and more generally of CSSs using a biopsychological approach, capable of comprehensively integrating all mechanisms that until now have been considered in the pathophysiology of CSSs and COPCs.
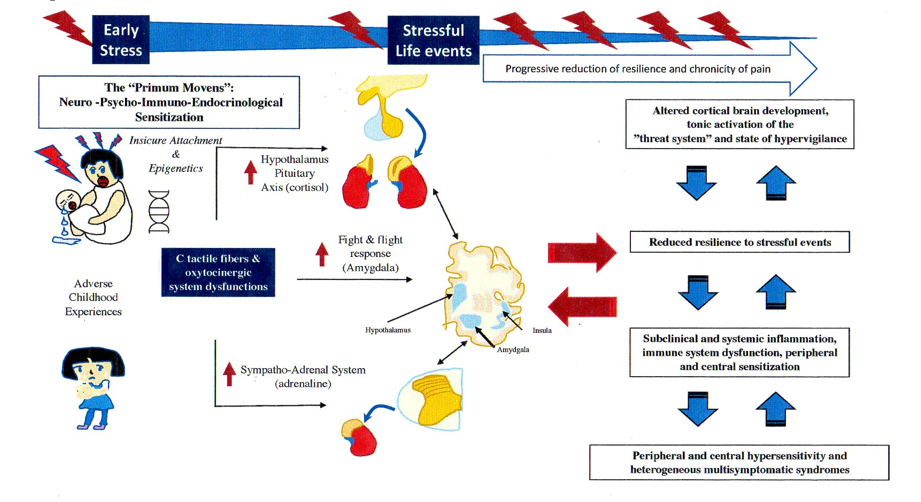
We hypothesize that insecure attachment (IA), C tactile (CT) fibers and oxytocinergic system (OS) dysfunction, together with epigenetic mechanisms, could underlie the different pathophysiological aspects of FMS and related multisensory hypersensitivity.
In this regard, we hope that our new pathophysiological model, common to different types of chronic pain syndromes and CSSs, can promote scientific research in this field and a more compassionate way of conducting the doctor-patient relationship, thus leading to a more effective clinical approach.
2. Pathophysiology of FMS
2.1. The Stress Response System in FMS and FSDs
Although clinical observations and scientific findings support the theory of FMS as a stress-related disorder due to dysregulation of the BSS, [6,7,8,9] the true origin of these pathophysiological mechanisms is still the subject of different interpretations [10,11].
From this perspective, many studies conducted on FMS have demonstrated that the two main players of the human stress system, the HPA axis and the sympathetic nervous system (SNS), present different types of dysfunctions, with some authors arguing for hypoactivity and others for hyperactivity of both systems [12].
The nature of these contradictory results probably depends on the complexity of human stress neural circuits which, to adapt the body to stress stimuli, work together in a mutual and synchronized way and influencing each other with different positive feedback loops [13]. Moreover, the heterogeneity of the study design with regards to the selection of the clinical sample (age of the patients and duration of the disease), the methods applied for the evaluation of the BSS, the different phases of the disease considered in the study, were considered the main factors responsible for these inconsistent findings observed in the pathophysiology of FMS [10,14].
Regardless, to explain this different pattern of BSS activation, it has been hypothesized that, over time, an increase in basal tone of both the HPA axis and SNS activity in the early stages of the disease (hyperactivity phase), could progressively predispose to a breakdown in the acute stress-related response (hypoactivity phase) during which the ability to cope with daily stressful stimuli could be compromised [6,7,10,14,15].
From this perspective and in line with previous results [14], more recently it has been demonstrated that in response to intense pain, the concentration of cortisol in the hair is higher in the early stages of FMS, while it tends to decrease afterwards, suggesting some sort of downregulation of cortisol during disease progression [16].
Certainly, these findings do not appear to be specific to FMS, but common to the spectrum of COPCs and FSDs [4]. Indeed, it has been theorized that the comorbidity between these overlapping clinical syndromes could depend on the same anomalies in the stress regulation system [6,7,17].
One of the most important methods used to evaluate ANS in patients with FM is cardiovascular regulation.
A large body of evidence suggests that in patients with FSDs, and in particular in patients with FM, basal heart rate variability (HRV) is lower than in healthy subjects, supporting the potential use of this cardiac parameter as a biological marker of chronic pain (CP) [18,19]. In fact, HRV and its sympathetic and parasympathetic components represent a reliable clinical tool for evaluating the body’s ability to cope with acute stress stimuli such as nociceptive stimulation [20] and for this reason it is also considered a biological indicator of human resilience [21].
Recently, the association between reduced HRV and FSDs was further found in a large-scale study suggesting a predominant role of the sympathetic arm of the ANS in the pathophysiology of CP [22].
The “chronic pain as ANS dysfunction” paradigm has been particularly demonstrated in FMS characterized by a general deterioration of the stress response evidenced by increased basal sympathetic activity, a reduction in sympathetic reactivity to acute stress, and a reduction in parasympathetic tone [23,24,25,26,27,28].
In a recent clinical study, lower HRV and a more impaired response to cognitive stress were found to be associated with mood and anxiety disorders in patients with FM, supporting a sharing of pathophysiological mechanisms between CP, psychiatric disorders and dysfunction of the autonomic nervous system [29].
Interestingly, the role of human stress dysfunction in the pathophysiology of FSDs was also confirmed in a general population study which led to a unifying diagnostic construct termed bodily distress syndrome (BDS) which may be common to several clinical syndromes [30,31]. The importance of this definition lies in its prognostic implications as highlighted by a prospective study which highlighted how in BDS, a deficit in response to psychological stress can predispose to the development of diseases over time [32].
On the other hand, as clearly reported by the International Association for the Study of Pain (IASP) [33], pain cannot be considered a pure sensation, but rather a stressful and unpleasant experience, composed of sensory, emotional and cognitive components linked to the perception of real damage or a threat of it. This definition highlights the cognitive aspect of pain perception and modulation and the significance of pain as a threat to survival.
With this in mind, a recent pathophysiological and integrative model called “Fibromyalgia: Imbalance of Threat and Soothing Systems” (FITSS) has been proposed [34]. According to this model, patients with FMS could have an excessive functionality of the threat system and the related defense cascade which over time could predispose and/or maintain the clinical phenotype of FMS [34].
2.2. The BDS and FMS: What Came First?
Although dysregulation of the bodily stress system (BSS) could explain the shared symptoms in FMS and other CSSs, it is still a matter of debate whether these neuroendocrine abnormalities represent the cause or consequence of CP and FMS [11,13,22,34,35].
A large body of scientific evidence supports the hypothesis of an association between trauma, both physical and psychological, and the onset of the first clinical manifestation of FMS, in particular when the trauma was emotional [36]. From this point of view, at least to a certain extent and in the initial phase of FMS, it is intuitive to consider the role of the BSS as primary and not secondary to the onset of pain.
Arguably, scientific discoveries emerging from studying the relationship between early life experiences and developmental psychobiology could be the key to answering what comes first between BSS abnormalities and CP [37,38]. In fact, numerous findings from both animal and human research have widely demonstrated that early stressful life events may be responsible for maladaptive neural plasticity and function and that these abnormalities may have a negative impact on children’s psychological and physical health throughout life [37].
From this perspective, it has been shown that subjects with a positive history for childhood trauma, are twice as likely to develop a CP adulthood (odds for fibromyalgia = 2·52; 95% CI = 1.92 – 3.31) [39]. Interestingly, the likelihood of developing FSDs is tripled when both childhood and adult trauma were included in the meta-analyses, supporting the key role of BSS in the pathophysiology of CSSs [40].
As previously mentioned, comorbidity between FMS and depressive and anxiety disorders is more the rule than the exception, and a positive history of childhood maltreatment has been considered to be a pathophysiological missing link of comorbidity between CP and psychiatric disorders. In particular, child maltreatment has been found to be highly associated with post-traumatic stress symptoms (PTSSs), a clinical spectrum that represents a frequent consequence of childhood maltreatment and which can also be considered a symptomatic phenotype predisposing to CP in childhood and adulthood [41,42]. Interestingly, adverse experiences in childhood have also been found to be associated with CP in youth, suggesting a clear cause-and-effect relationship between early stressful life events and the pathogenesis of CP [43]. From this point of view, it has been shown that early stress and in particular childhood maltreatment, is highly associated with HPA and ANS dysfunction, as well as alterations of other peripheral stress response systems such as the immune and inflammatory ones [37,44]. According to this pathophysiological “early trauma model” and considering the frequent clinical overlap between post-traumatic stress disorder (PTSD) and FMS (the prevalence varies between 56%), it has been suggested that all patients with FM should be screened for the presence of PTSD [45]. The importance of these epidemiological and clinical findings, is linked to the negative impact on therapy and prognosis that the presence of PTSD and also partial or subthreshold forms (PTSS) can have in patients suffering from FM [45,46]. In this regard, it has been shown that the level of central sensitization (CS), measured as widespread pain, pain intensity and multisensory complaints, could depend even more on the symptoms of PTSD than on the intensity of exposure to trauma itself, suggesting an important role of this psychiatric syndrome in the pathogenesis of CP [46]. Furthermore, as with other CSSs, FMS and PTSD may share some pathophysiological mechanisms such as HPA dysregulation (significantly low basal cortisol) [47].
2.3. The Relationship between BSS and the Insecure Attachment in CP and FMS
It has been shown that having a secure attachment (SA) predisposes one to having high-quality relationships and at the same time to positive health outcomes. John Bowlby (1969) [48], the father of attachment theory, defined attachment as a biobehavioral state composed of biological, physiological and behavioral systems, capable of maintaining internal homeostasis in the event of emotional arousal. Figure 1 schematizes the four different types of attachment model proposed by Bartholomew & Horowitz (1991) [49]. According to this model, several researchers consider the attachment system in itself to be a device responsible for regulating emotions [50]. From this perspective, the identikit of a person with SA is characterized by competence in social behavior, emotion regulation, executive cognitive functions, with a balance between exploration and attachment behavior and an ability to analyze one’s feelings and thoughts. On the contrary, while the ambivalent/preoccupied form of insecure attachment (IA) is characterized by feelings of anger and less autonomy, the typical aspect of the avoidant/dismissing form is to deactivate the attachment system to avoid discomfort; in both forms of IA, poor emotional regulation leads to an increase in the response of the stress system to stimuli related to the attachment system [51].
On the other hand, numerous evidence, supports the harmful role of IA on the biological, psychological and sociological aspects of adulthood. How can we explain these scientific findings?
Over the last three decades, numerous studies have been published on the neurobiological and physiological mechanisms of the attachment system, suggesting that SA can be considered an indicator of resilience [52]. In fact, it has been shown that the activation of the attachment system in adult subjects with SA increases HRV (one of the most important biomarkers of resilience) through the parasympathetic response to stress, explaining the psychophysiological benefits deriving from having positive social support [53]. These findings were recently also replicated in adolescents with SA who demonstrated higher HRV compared to the AI groups when subjected to the Adult Attachment Projective System interview, suggesting once again that secure adolescents are more efficient in dealing with stressors arising from attachment representations [51].
In fact, it has been demonstrated that attachment relationships have a regulatory role on the physiological and psychological response to stress through the modulation of BSS [54]. Consequently, compared to other attachment groups, the secure ones are able to respond with a lower increase in cortisol, lower skin conductivity and more flexible prefrontal functions during attachment-related stimuli [54]. On the contrary people with IA, both infants and adults, are characterized by an augmented activity of the stress response system supporting the paradigm of IA as a deficit of emotion regulation. In particular, a large body of evidence has demonstrated that early childhood adversity and/or poor parenting are associated with HPA axis hyper or hypo-responsiveness, sympathetic-adrenal-medullary (SAM) dysfunction, and immune system dysregulations in infants, children, and adults with IA [55]. The association between dysfunctional neuroendocrine and immunological responses observed in IA subjects is not surprising considering the reciprocal modulation that HPA, SAM axes and the immune system exert on each other [56]. On the other hand, the lower HRV observed in adolescents with insecure avoidant attachment and unresolved attachment compared to that of adolescents with SA, confirmed their reduced ability to regulate the ANS in response to stressors related to the representations of attachment [51].
For the data reported above, BSS dysregulation could represent the missing link between having an IA during childhood and negative health outcomes such as psychiatric, cardiovascular, metabolic and tumor diseases in adulthood [55,57,58].
Thus, considering the importance of the attachment system on the regulation of BSS and affective states, IA is considered a psychobiological indicator of vulnerability to stress and may represent an important risk factor for several diseases, including pain [59,60,61].
Several studies have shown that IA represents not only a predisposing factor for CP but also an important cause of altered pain perception, such as catastrophic pain, lack of ability to cope with pain and a significant predictive factor for anxiety and depression in CP disorders [59]. In reality, inconsistent results have been found on the relationship between IA and pain intensity, although numerous studies suggest that people with IA, in addition to being more prone to developing pain, perceive it as more pervasive, distressing and disabling [61,62].
In a cross-sectional study conducted on a large general population sample, subjects with chronic widespread pain (CWP) presented IA (preoccupied, dismissing, fearful) more frequently than subjects without pain. Interestingly, the number of pain sites was associated with IA along with more disabling pain symptoms [62]. Despite the cross-sectional nature of the study, also considering the results found by other researchers, the authors concluded that people with IA may be twice as likely to have CWP. In fact, it has been found that the origin of psychological trait “pain related to fear” is to be considered a consequence of IA, and in particular of its model of self-dimension (degree of anxiety regarding rejection based on beliefs of personal unworthiness) [63]. In this perspective, the old theory of CP as derived from a stress diathesis, has been transformed into attachment diathesis models of CP, suggesting a direct and causal pathophysiological link between IA, stress and CP disorders [59]. An important confirmation of IA as the main source of a more negative subjective perception of the painful experience came from studies conducted with experimentally induced pain [64].
In the context of FMS, it has been proposed that attachment style should be regularly explored in patients with FM, confirming the validity of the attachment-diathesis model of chronic pain [65]. In fact, as also found in other CP syndromes, IA is frequently observed in patients with FMS and appears not to be specific of this CP disorder [65,66]. Rather, it has been hypothesized that IA represents an important predisposing factor to alexithymia which, conversely, has been found to be a predictive personality trait of FMS [66,67]. Regardless, the presence of IA represents an important predictor of lower quality of life in patients with FMS and this effect appears to be mediated by low self-esteem [68] and depressive symtpoms [69]. In other words, the presence of IA (dismissive, preoccupied and fearful), could be responsible for a greater biopsychosocial impact and a higher disease burden in female patients with FMS [68,69].
As mentioned above, it has been shown that the negative effect of IA on the functioning of patients with CP, may not be mediated by an increase in pain intensity, suggesting that other variables like cognitive (catastrophizing, coping style) and affective components (anxiety and depression) of the pain experience and social status such as marital satisfaction and social support from significant others, may have a role [61,62].
2.4. Role of Oxytocin System in Attachment System and Pain Modulation
Oxytocin (OT) is a very ancient nonapeptide dating back almost 500 million years, initially conceived as a system for the regulation of basic functions, such as thermoregulation and energy balance (Feldman, 2020) [52]. Interestingly, over millions of years, OT analogue genes and related peptides have been highly conserved, but at the same time have undergone enormous evolutionary transformations structurally and functionally in order to adapt the organisms to rapidly changing of environments [70,71]. The importance of OT, throughout evolution, is suggested by its multiple social and non-social functions common to different species ranging from nematodes to humans [71]. In particular, in mammals, OT has been incorporated into labor and breastfeeding and then into the context of the mother-infant dyad, playing a critical role in the maturation process of the infant’s stress-reduction system [52,70,71]. Indeed, the OS in both zebrafish and mammals, is part of the hypothalamic paraventricular “stress response” network, which together with the locus coeruleus, the area postrema and the caudal hypothalamus, modulates the defense behavior in case of noxious and other threatening stimuli [72]. In other words, the OS represents the biological system that allows individuals to respond in an adaptive way to various life adversities, regulating the psycho-immuno-endocrine axis and promoting resilience [52]. According to this model, the OS, together with the affiliative brain and bio-behavioral synchrony represents a fundamental pillar of the neurobiology of affiliation and resilience [52]. From this perspective, it has been shown that the infant’s OS, during the sensitive period (the first nine months of life) and throughout the childhood, is hardwired and programmed through the parental behavior and care (both mother and father) in a sort of “rhythm of safety”, supporting the importance of the SA for social, emotional and cognitive development of the child’s brain [52]. Practically, the OS is the physiological system that regulates the bio-behavioral synchrony of the mother-child dyad. This external regulation of an immature brain from a mature brain is key to the programming of cognitive and emotional functions such as executive functions, empathy, stress management, behavioral adaptation, conflict dialogue and behavior health that are of crucial importance for social behavior and characterize the resilient brain [52,73].
It has been demonstrated that OT can exert an anti-nociceptive action together with anti-inflammatory and anxiolytic effects, thus improving perception and behavioral response to painful or aversive stimuli [74,75,76,77]. In particular, OT can exert its analgesic effect at both spinal cord level and above spinal cord level as well as in the peripheral nervous system. By interacting synergistically with the endogenous opioid system and activating GABA inhibitory interneurons, OT has been shown to inhibit nociceptive afferents (C & Aδ fibers) at the level of the most superficial layers of the dorsal horns of the spinal cord [74,77]. However, although the precise mechanism is not yet fully clarified, numerous data suggest that OT could be a modulator of pain perception by acting in several brain regions implicated in the cognitive-affective modulation of the pain experience [77]. In particular, the widespread presence of OT receptors (OTRs) in brain regions such as prefrontal cortex (PFC), Orbitofrontal Cortex (OFC), insular cortex (IC), amygdala (AMY), anterior cingulate cortex (ACC), supports the modulatory action of OS on the “medial pathways of suffering” of pain [77,78]. Furthermore, it has been hypothesized that OT could enhance the descending inhibitory system by stimulating the opioid system at the periaqueductal gray matter (PAG) and at the Rostral Ventromedial Reticular Structure (RVM), two other brain regions with extensive expression of OTRs (Yang et al., 2022) [77]. Nonetheless, OT, by strengthening the reward system on the ventral tegmental area (VTA) and the dopaminergic Nucleus Accumbens (NAc) and reducing fear behavior by inhibiting the central nucleus of AMY, could protect against the chronicity of the painful state.
Interestingly, good functionality of both the reward system and the opioid system provided by the OS, is the clear hallmark of an SA, further supporting the mutual connections between SA, OS and pain modulation [52,77].
A recent study conducted on a sample with CP suggested that the analgesic effect of mindfulness-based pain management could be linked to the action of OT by reducing pain sensitivity and improving mood symptoms. Interestingly, a trend towards reduction of inflammatory markers (IL-1b, IL-6, IL-8, TNF-a) and stress biomarkers (DHEA-S) was also observed in this clinical study, arguing that the anti-stress and anti-inflammatory properties of mindfulness-based pain management, could be mediated by OT [79].
On the other hand, during the different phases of life, particularly in the first ones (e.g., sensitive period of the first nine months) many variables including type of attachment, sex, social experience, genetic and epigenetic factors, could have a negative impact on the physiology of the OS which in turn, over the course of life, could be less efficient in regulating the stress response, allostatic load/overload and resilience, leading to different outcomes [80].
2.5. CT Fibers in FMS: from Small Fibers Pathology (SFP) to Oxytocin System
Although there is evidence of an important association between FMS and SFP, the pathophysiological mechanism underlying this comorbidity is still unclear. Indeed, SFP has been shown to be present in a proportion of nearly 50% of patients with FMS, supporting a role for peripheral mechanisms in the pathophysiology of FM at least in a broad range of patients [84]. On the other hand, with the increasing number of studies on intraepidermal nerve fiber density (IENFD) (65% in human studies and 440% in non-human studies from 2010 to 2020), a reduction in IENFD was surprisingly found in various human diseases [85]. In any case, on the one hand it is not yet clear whether these findings could be the cause or consequence of FMS, on the other hand there is an urgent need to discover the common mechanisms underlying the loss of IENFD observed in different types of diseases [84,85,86]. Interestingly, in an experimental model of pain in rats, it has been demonstrated that the bilateral increase in glutamatergic activity at the level of the IC is responsible for both the multimodal pain behavior and the loss of peripheral fibers. According to their results, the authors suggested that an overactivity of the IC could be the main cause of the pathogenesis of SFP in patients with FM [87]
In contrast to the theory of centralization of pain in FM, some authors have hypothesized that the dorsal roots of the spinal ganglia (DRG) could play a role in the genesis of hyperalgesia in FMS starting from various stressful stimuli, including the psychological ones. From this point of view, the alteration of small peripheral fibers has been considered the most important evidence of the link between FMS and DRG pathology [88]. In favor of a role of DRG pathology and peripheral sensitization as pathophysiological mechanisms of FMS, it has been demonstrated that IgG from FMS patients beyond the ability to bind to human DRG, inoculated into mice are able to produce painful sensory hypersensitivity by sensitizing peripheral nociceptive afferents and causing a reduction of IENFD [89]. According to other authors, a high sympathetic activity could be responsible for an increased response of the immune system which, together with the high level of muscular tension and the related myofascial-derived antigen anomalies, could lead to the formation of immune complexes capable of creating hyperexcitability in the DRG [90].
Moreover, it has been argued that, in the absence of severe depression and anxiety, the clinical phenotype of FMS and its associated disease burden correspond to that of neuropathic pain with dysautonomia. In this model, mood and anxiety disorders would not only not be a clinical indicator of pain centralization, but should also be considered confounders of the peripheral source of sensitization [91]. Indeed, the difficulty in interpreting the results stated above is even more pronounced considering a recent study which demonstrated the presence of a subgroup of patients with FMS who, without any alteration of nociceptive afferents, showed the clinical phenotype of small fibers neuropathy (SFN) [86]. On the other hand, in diabetic neuropathy, a positive correlation between the loss of IENFD and the presence of pain has not been found and for this reason the loss of fibers is not a reliable predictor of the development of neuropathic pain even in patients with diabetes [85].
Although the presence of SFP in FMS has recently been confirmed in a large percentage of patients, it has been clarified that SFP observed in FMS must be differentiated from SFN, being different from it not only from the neurophysiological point of view, but also from the point of view of the clinical phenotype [84,92,93].
How can we interpret such a large amount of SFP in FMS and what kind of role could it have from pathophysiological perspective?
Over the past three decades, a critical step in the study of the somatosensory system has been the discovery of a class of unmyelinated cutaneous mechanosensory fibers called C-low-threshold mechanoreceptors afferents (C-LTMRs) [94,95]. Clear evidence of the presence of this type of C fibers also in humans emerged in 1990s [96,97]. but it was only two decades later that “the affective touch hypothesis” was formulated [98]. In this model, “the essential role of the C tactile system is to convey pleasant aspects of light touch, especially skin-to-skin (SSC) contact with affiliated humans” and for this reason these C-LTMRs in humans are renamed C tactile (CT) afferents. It is interesting to note that, starting from this hypothesis, several research groups have stated that as in the pain system, also in the sensory system there are central tactile discriminative and affective representations that depend on the activity of the Aβ and CT afferents respectively [99].
It has been shown that CT afferents are abundant in hairy skin and that they can be activated when a tactile stimulus, such as a skin-stroking caress, is applied to the skin at a specific speed (3-6 cm/s), with a determined pressure (0,04 - 5 millinewton) and with an optimal temperature (skin temperature) [97,100].
Interestingly, the involvement of the opioid system both in the pathophysiology of FMS and in the physiology of the pleasant sensation due to the activation of CT afferents, has been found in patients suffering from FM, confirming an alteration of the opioid system in this CP condition and linking it, for the first time, to an abnormality of the function of CT afferents [101]. A confirmation of the involvement of a dysfunction of the CT afferents and the related affective tactile system was corroborated in a more recent neuroimaging study in FMS. In this study, the authors suggest that the reduction in the activity of the posterior part of the IC during the pleasant rating, and the activation of its anterior region during pain assessment, could be linked to normal sensory processing associated with anedhonia to pleasant touch and abnormal evaluative processing, confirming the potential role of CT afferents in the physiology and pathophysiology of both acute and CP modulation [102]. Interestingly, although to a lesser extent, an altered function of CT afferents has also been found in migraine which, like FMS, is part of COPCs [103]. In particular, during the interictal phase, migraine patients showed greater habituation to the stimulation of CT afferents on the trigeminal-innervated skin. The authors concluded that the reduction in pleasant tactile experience and increased lack of habituation to stimulation of CT afferents observed in migraine patients could be secondary to subclinical interictal mechanical allodynia [103]. A reduction in the pleasantness of touch was also found in patients with low back pain both to a lesser extent in the subacute and chronic phases. The authors suggested that the dysfunction of the brain pleasant representation of touch mediated by CT afferent abnormalities could be considered a marker of transition from the acute pain phase to a CP syndrome [104]. In line with this trend, a reduction in pleasantness through stimulation of CT afferents was also found in patients with post-herpetic neuralgia (PHN) and complex pain regional syndrome (CPRS) [105]. Interestingly, in this study, the typical inverted U curve of CT afferent stimulation was not found in patients with PHN, confirming the prevalent involvement of C-fiber damage over A-fiber damage in this CP condition [99,105]. Furthermore, in patients with unilateral CPRS, reduction in pleasantness due to stimulation of CT neurons was observed not only in the affected side, but also in the contralateral side, supporting a more systemic involvement of CT system dysfunction in CPRS [105]. On the other hand, as reported above, a reduction in IENFD has been found in several chronic disorders regardless of the presence of CP [85]. Therefore, the bilateral deficit in pleasantness from CT afferent stimulation observed on CPRS is not surprising since a bilateral reduction in IENFD was also observed in this CP disorder also in its unilateral form [105,106].
In any case, these data suggest that a dysfunction of the CT system may be common to several CP conditions probably characterized by a subclinical reduction in the perception of pleasantness. This could represent the missing link that can explain the pathophysiological mechanisms shared by COPCs [107].
However, it still remains to be clarified which peripheral and central mechanisms underlie the dysfunction of CT afferents and consequently the related lack of pleasant perception.
Furthermore, the enigmatic physiology of this C sensory system is based on its potential pro-nociceptive and non-nociceptive properties [108]. It has in fact been demonstrated that C-LTMRs stimulation is able to potentially produce not only a reduction in pain but also a type of allodynia which, compared to that mediated by Aß, is less topographically correlated to the side of the damage. From this point of view, C-LTMRs-mediated allodynia resembles that typically observed in patients with widespread pain such as FMS [108].
On the other hand, since the activation of CT afferents is associated with a subjective sensation of pleasantness, a series of studies have been conducted on acute pain induced in healthy controls, demonstrating the analgesic power of gentle touch [107,109].
The mechanism of action underlying the analgesic effect of stimulation of CT afferents has been studied in animal models and in human clinical studies. In particular, in animal models, CT afferents would act on the one hand on the substantia gelatinosa (SG) localized in the dorsal horn of the spinal cord by stimulating TAFA4+ C-LTMRs [108,110], on the other by increasing the secretion of oxytocin [111]. In humans, stimulation of CT afferents could be linked to increased tone of the parasympathetic system [112] and to a modulation of the activity of the endogenous μ-opioid receptor system [113]. More recently, a model of pain modulation has been hypothesized in which CT afferents would act by inhibiting ascending nociceptive pathways (bottom-up inhibition) and downregulating brain regions such as the ACC and the IC, two strategic cortices implicated in pain experience perception [114].
It has in fact been demonstrated that the activation of CT afferents is able to reduce the temporal summation of a second pain (TSSP), a neurophysiological marker of central sensitization and the pathophysiological basis of some CP syndromes like FMS [115]. The mechanism of action by which the stimulation of CT afferents would block the wind-up phenomenon in case of repetitive noxious heat stimulation would be based on the activation of inhibitory neurons which in turn would inhibit C nociceptors at the SG level of DRG [115]. Certainly, in healthy controls and acute pain studies, this model might have an own validity, but in CP patients where the maladaptive plasticity occurs, could be less applicable and suitable. In fact, during the process of pain becoming chronic, there is a progressive shift in activity from the somatosensory cortices that process nociceptive inputs to the brain regions that regulate the emotional-affective components of the painful experience [116]. At the same time, it has been highlighted that to obtain pain modulation from the stimulation of the “affective system C”, obviously, the CT afferents must be intact [117]. In this prospective functional magnetic resonance imaging (fMRI) and psychophysical study, the authors demonstrated that while in healthy subjects the activation of CT afferents is able to modulate heat pain regardless of its applied intensity, in patients with SFN this clinical response does not occur, indirectly confirming IENFD loss in this clinical sample. In addition to this, the authors concluded by arguing that CT fibers could exert their pain modulation at the DRG level in lamina I-II by activating inhibitory interneurons [117]. More recently, the same research group explored the analgesic properties of CT afferents in a sample of patients with chronic CRPS [118]. Although they observed a reduced threshold of heat sensation after repetitive stimulation of CT fibers and regardless of the presence of allodynia, the overall intensity of pain did not decrease, suggesting that the analgesic power of the “tactile affective system” is still present in patients with CP even if too weak to have a significant effect [118]. On the other hand, as reported above, in patients with CRPS, a widespread reduction in IENFD and perhaps CT fibers was found which explains to some extent the results of this study [105]. Unlike the results obtained in patients with chronic CRPS, a study conducted on a sample of different CP conditions found a significant and rapid improvement in pain intensity after an adequate CT fiber stimulation paradigm (23% of pain reduction after only 11 minutes of CT fibers stimulation at 3 cm/s with a force of 2.5 mN) [119]. Interestingly, in the same study the authors found a lower accuracy and confidence in the interoceptive processes of their own internal body sensation in patients with CP despite not being aware of it, supporting what is also reported in the scientific literature [120]. In this view, it has been highlighted that the relationship between pain modulation and body representation is bidirectional and reciprocal, with a dysfunction of one potentially predisposing to the pathogenesis of the other and vice versa. A potential therapeutic role of gentle, pleasant touch in modulating hypersensitivity of internal body sensation and associated elevated pain perception was demonstrated in an experimental study conducted on healthy controls [115].
Overall, the affective tactile system, represents an ancient phylogenetic system that not only has a fundamental function in the regulation of social behavior, but also has a peculiar role in the modulation of pain with both anti-nociceptive and pro-nociceptive properties [108]. Taken together, the above reported results are in line with the possibility that at least in CP conditions associated with C nociceptor dysfunction, stimulation of CT fibers may represent an alternative analgesic therapeutic approach [107,115,119]. From this perspective, an in-depth knowledge of the affective tactile system is crucial.
2.6. CT Fibers, Oxytocin System and Brain Development
Affective touch (AT) and the related CT fibers represent a fundamental functional system for the development of the newborn’s brain and, according to some evidence, also for that of the fetus [121,122,123]. It is known that the somatosensory system is almost completely mature by the 32nd week of pregnancy, with the exception of the insula and the superior temporal sulcus, two cortical regions involved in social cognition that become operational around 2-3 months and 12 months of age respectively [123]. It is interesting to note that it has been hypothesized that already in this gestational period the CT fibers may have a regulatory function on the maturation and differentiation of the “affective tactile system”. The rhythmic movement of the amniotic fluid in utero, would stimulate the CT fibers precisely through the delicate movements exerted on the “lanugo” of the fetus in a way similar to a physical massage [124]. Indeed, the effects of the therapeutic application of SSC contact and touch with both “kangaroo therapy” and pediatric massage have been studied in animal models and in premature human infants, demonstrating that CT fibers stimulation improve growth, regulate the stress response, reinforce and modulate immune system, and boost the development of cognition and sensory-motor integration in preterm human infant [125,126].
It is also important to highlight that the maturation of the CT afferents is already ready to carry out their function in the last gestational trimester, preparing the increase in the parasympathetic tone of newborns by reducing the “child’s stress for the birth” [127,128]. and preterm infants [129] and the analgesic power of the caresses provided by the parent-infant bond [130].
After birth, it was found that AT continues to exert a regulatory function on the development of the somatosensory system, BSS, ANS, as well as the child’s immune system [131]. Overall, it has been documented that AT by an attachment figure (parent or caregiver in general) is able to promote the global development of the child by stimulating five different but interconnected systems: the somatosensory system, the autonomic system and the immunity, the affiliative bond and social cognition [131].
From an evolutionary point of view, the CT fiber system of the higher primate may have modified its original functions, moving from a role as a simple regulator of the stress response to a sort of homeostatic system responsible for controlling of cognitive and emotional functions [121,122].
One of the most important mediators that parent-child bonding, affective touch and CT afferents stimulation use to promote social affiliative bonds and prosocial behavior is OT, a neuropeptide composed of just nine amino acids synthesized by the hypothalamic paraventricular nucleus (PVN) and supraoptic nucleus (SON). The OT receptors are widely express in the brain and in the entire organism underlying the importance of this neuropeptide in regulating several functions in the body. In rodents, the main brain source of OT has been shown to be magnocellular neurons and, to a lesser extent, parvocellular neurons of the PVN and SON. These neurons with their axons and dendrites reach different brain regions such as the frontal, cingulate cortex and IC, the basal ganglia, the limbic system (AMY, hippocampus and septum), the midbrain, the brainstem and the spinal cord [132]. Furthermore, some parvocellular neurons also project to the anterior pituitary where they regulate the release of adrenocorticotropic stress hormone [133]. Indeed, oxytocinergic projections from the PVN to the brainstem and HPA have been considered the anatomical and functional pathways responsible for the anti-stress and analgesic action of OT [80,131,133,134]. In fact, the increase in parasympathetic tone due to the activation of CT fibers recorded in the fetus during the last three months of pregnancy could be mediated by the stimulation of the nucleus of the solitary tract of the brainstem by the oxytocinergic pathways coming from the PVN [135].
Importantly, it has been suggested that the first activation of the OS depends on the projections of CT fibers that reach the PVN from the IC [136], supporting the role of AT in the development of CT fibers and the OT system and more generally for the regulation of pain modulation and the reward system.
In this line, it has been demonstrated that OT acts synergistically with the dopaminergic afferents of the mesocorticolimbic regions and with the opioidergic tachykinin + neurons of the reward system and more specifically of the lateral and ventrolateral PAG [123,131,137,138].
Taken together, the data reported above suggest that a normal development of CT fibers is fundamental for the physiological activation of the oxytocin system and the reward systems (dopamine and opioids) and at the same time for the pain modulation system (Figure 2). From this perspective, it has been found that, in newborns, the experience of pain is very similar to that of adults and that the caregiving environment is able to modify newborns’ sensitivity to pain [137]. In particular, SSC contact with caregivers has proven to be a valid non-pharmacological treatment for relieving pain in newborns in intensive care [139].
It is important to underline that it has been shown that from childhood to adolescence, pain represents a risk factor for the development of a dysfunctional attachment model while the caregiving environment can constitute a predisposing or protective factor [140].
3. Discussion
One of the most recent and interesting debates on the pathophysiology of FMS concerns the prevalent role of peripheral or central pathogenic mechanisms along nervous system pathways in combination with immune system dysregulation [34,141].
In their model called ‘Fibromyalgia: Imbalance of Threat and Soothing Systems (FITTS)’, Pinto et al. (2023) [34], define FMS as a disease linked to a dysfunction in emotional regulation that leads to an increase in the perception of threat and a reduction in the sense of safeness and protection regardless of the potential role of stress factors. Specifically, the hypothetical FITTS model would have been based on three main and interconnected pathophysiological factors: first, a high perception of threat; secondly, a reduced activity of the soothing affiliative system; and third, a persistent activation of the brain’s salience network [34]. This hypothesis found support in a recent study which highlighted the presence of emotional dysregulation with greater arousal towards unpleasant and socially unpleasant images in patients with FMS [142].
On the other hand, the FITTS model, has also been questioned on the one hand because it would not be a truly new theory but rather a different way of representing the model of ANS dysregulation, on the other hand because it does not mention the role of small fibers and the immune system activation on DRG which have recently been considered potentially involved in the pathophysiology of FMS [88,141].
In our opinion there is no real controversy between the two points of view, but rather a different perspective from which the authors see the phenomenon.
Certainly, it is still a matter of debate whether chronic painful states can be the result of “bottom-up” amplification mechanisms of nociceptive afferents or whether, on the contrary, a dysfunction of the descending nociceptive inhibitory system can lead the increase in pain perception towards a chronic state.
A different and more complete interpretation could be proposed by returning to clinical practice and the clinical phenotype of FMS and more generally of all CSSs. There is no doubt that FMS should not be simplistically considered a CP syndrome, both from a clinical and pathophysiological point of view. In clinical practice, in fact, CSSs are characterized not only by pain but above all by other conditions such as sleep disorders, cognitive problems, fatigue and psychiatric illnesses (depression, PTSD, panic disorder), which are often even more disabling than the pain itself and which are heterogeneously reported by patients.
As already widely reported above, despite the contradictory results probably linked to the complexity of human stress neural circuits and other factors already discussed, several scientific results and data support the theory of FMS as a stress-related disorder. In fact, a large body of evidence on HPA and ANS dysregulations has been found in FMS and more generally in other centralized pain syndromes [17]. In this vision, FMS and other CSSs can be considered a group of diseases characterized by hypersensitivity to the perception of pain and more specifically to the anomalous interpretation as a “threating” of innocuous sensory inputs both internal and external to the body [17,34].
It remains to be understood how this deficit in the interpretation of the salience of stimuli and in particular in the discrimination between threat and safety occurs in FMS and probably in other CSSs. According to extensive scientific evidence from both animal and human studies, we believe that, at least to a certain extent, the OS can represent a coherent answer to this question [143]. It has in fact been hypothesized that OT may play a crucial role in discriminating threat stimuli from safety stimuli, being a neuropeptide that is part of the hypothalamic paraventricular stress response network, widely represented in nature, from zebrafish to mammals [143,144]. In particular, it has been shown that OT is specifically involved in the recognition of the threat that could come from the environment or from fearful facial expressions, in order to identify a potential source of danger and consequently activate adequate adaptive responses [143]. The broad distribution of OT receptors within brain regions involved in stress regulation such as the prefrontal cortex, limbic area, hypothalamus, raphe, and medulla oblongata is in line with the OS functions mentioned above [145].
On the other hand, the analgesic power of OT has been widely attributed to its ability to improve the behavioral response to painful and aversive stimuli by reducing the perception of negative salience [77]. Indeed, it is widely demonstrated that OT receptors are widespread in different brain regions of the so-called “medial pain pathways” [77,78].
As previously described, it is important to underline that the analgesic action of OT is not limited to the modulation of the perception of the painful experience at the level of the higher brain centers, but extends to the spinal cord by activating the descending inhibitory and exerting a direct inhibitory effect on nociceptive afferents at the dorsal horn level [77,146]. It is also hypothesized that the physiology of OS, by strengthening the reward system (VTA and NAc) and inhibiting the central nucleus of the AMY [77], could represent a crucial mechanism of protection from the transformation of pain from acute to chronic.
Finally, in animal models, a large body of evidence supports the systemic anti-inflammatory properties of OT physiology, fundamental for preventing the pathogenesis of inflammatory pain, neuropsychiatric disorders and other chronic-degenerative diseases considered linked to systemic inflammation such as cardiovascular, gastrointestinal, diabetes and obesity [147]. On the other hand, it has been highlighted that OS might be susceptible to inflammation from early life and that neonatal inflammatory pain may lead to its dysregulation and neurodevelopmental diseases in later life [148].
In fact, it is important to underline that the OS is a very dynamic biological system and that it is not completely mature at birth, but on the contrary, is subjected to intense programming during the first nine months of age and throughout childhood through the maternal brain and caregiving behavior [52]. As already reported in the previous paragraphs, OS has a key role in the regulation of bio-behavioral synchrony of the mother-child dyad. For this reason, a mature maternal brain with a physiological OS, it has a key role in the physiological development of the immature infantile brain and consequently of executive cognitive functions, emotional regulation functions, empathy, stress regulation, behavioral adaptation and more generally the development of the resilient brain [52,73]. In fact, a resilient person is characterized by lower HPA reactivity and a higher parasympathetic tone in response to stressful conditions and compared to a subject without a resilient trait [80]. On the other hand, a large number of scientific findings support the detrimental effects of adverse events in early life on OS, with long-lasting negative consequences on both the physiological and behavioral development of the child [149]. From this perspective, we shed new light on the epidemiological data and the pathophysiological role of early adverse events and more generally of stressful events in the pathogenesis of FMS and comorbidity with PTSD [37,45,46,47,150].
As previously described, it has been reported that IA, the behavioral analogue of an OS dysfunction, predisposes in adolescence to emotional dysregulation and an increase in the response of the stress system secondary to stress factors related to attachment [51]. In adulthood, this behavioral pattern has been found associated with biological, psychological and sociological dysfunctions, highlighting the importance of functional programming of the OS and the consequent SA in the initial phase of brain development [55]. In other words, a dysfunction of OS and IA can be considered as two sides of the same coin, which, in a single construct, represent an important risk factor for several adult diseases including CP [55,57,58,59,60,61].
An important issue, and perhaps the most important if we look at the “primum movens” that could be responsible for the progressive dynamic process leading to FMS, is that of CT fibers and “affective tactile system”. It has already been extensively explained previously how important the role of CT fibers is in the regulation of pleasant sensations in a synergistic action with OS and the opioid system [77,113]. We have also described how CT fibers system is ready to act as a regulatory system for the maturation and differentiation of the “affective tactile system” already during the last trimester of pregnancy [121,122,123]. We have previously highlighted the importance of having a SA to achieve good functionality of the reward system and the opioid system, supporting the functional connections between SA, OS and pain modulation [52,77]. We also reported that a CT fibers dysfunction linked to opioid system abnormalities [101] together with a dysregulation of insular circuits [102], was observed in patients with FMS. We also underlined the possibility that a widespread subclinical dysfunction of the CT fiber system could be a common pathophysiological ground that predisposes to various CSS and COPCs, characterized by a subclinical reduction in the perception of pleasantness regardless the presence of pain [105,107]. This sheds light on another otherwise incomprehensible prevalence data, relating to the reduction of IENFD in various diseases and even in pain-free clinical samples [85].
From this perspective, it has been demonstrated in animal models that a sustained increase of bilateral glutamatergic activity of the IC, the first cortical station for CT afferents, is responsible not only for the pathogenesis of pain but also for the loss of peripheral fibers. According to their findings, the authors hypothesized that the high prevalence of SFP and IENFD observed in FMS might be mainly due to IC dysfunction [87]. Interestingly, psychophysiological studies have shown that skin stimulation of C-LTMRs with stroking at intermediate frequency speeds performed in healthy human controls subjected to tonic muscle pain has been shown to be capable of producing a type of allodynia very similar to that observed in patients with widespread pain such as FMS [108,151,152].
By putting together all the pieces of this scientific puzzle discussed along the different topics addressed in this overview, we can conclude with a pathophysiological model that can be applicable to FMS, CSS, COPCs and probably to all chronic-degenerative diseases.
In nature, there is an ancient phylogenetic and physiological system functional to the survival of life, common to many living species but species-specific, composed of at least 3 subsystems: the attachment system, the affective – CT fibers system and the OS. Over the course of millions of years, this multisystem physiological apparatus, particularly in the human race, has undergone a modification from its original functions as a simple stress regulator, towards a homeostatic system responsible for the emotional and cognitive regulation of social stimuli and in particular to differentiate what could be a real threat while maintaining the tendency to remain in a safe situation (e.g., affiliation system) [70,71,121,122]. This complex neurological, psychological, immunological and endocrine system begins to be programmed from the last three months of pregnancy of the fetus until childhood by a biological and physiological function called bio-behavioral synchrony and which is elaborated by the parent-child dyad [52,73]. Since this system is epigenetically determined, the type of attachment system of the parental figures, the early adversities and stressful life events particularly from the last period of pregnancy and the first nine months of life (e.g., sensitive period) until childhood and early adolescence, could represent important factors capable of modifying the neurobiological path and development of the infantile brain [148,149]. The final result of the consequent maladaptive brain, could lead progressively to a dysregulation of BSS together with cognitive-emotional dysfunction, systemic inflammation, central sensitization, and to an increasing sensitivity and perception of pain.
On the other hand, the clinical heterogeneity of syndromes such as FMS, CSS and COPCs, usually observed in daily clinical practice but also highlighted by the scientific literature, could also be linked to the type of attachment of the patients, to sex, to genetic and epigenetic factors and particularly to the life stage of the patients at the time of clinical observation [80,149]. In line with this, it has been found that in the general population a spectrum of fibromyalgia-like clinical symptoms exists within a continuum of biopsychological distress, which has been termed “fibromyalgia” [81,82,83].
Therapeutic Implications and Conclusive Remarks
As widely reported, a large amount of data would support the use of OT as a pain modulator in clinical practice, particularly in CP conditions with deep pressure pain such as FMS [153]. However, the scientific literature in this field is still in its infancy and the few available studies obtained on human samples have shown inconclusive results, although a recent narrative review has highlighted a significant dimension of the effectiveness of OT in reducing sensitivity to pain in patients with back pain, abdominal pain, and migraine [154]. Certainly, the scarcity of studies, the heterogeneity of clinical samples, the different design of the studies, the mode of administration and the dosage of OT are among the most important reasons for these conflicting results [154]. Nonetheless, OT does not cross the cerebrospinal barrier and this pharmacokinetic characteristic could represent an obstacle to its potential central analgesic activity.
To the best of our knowledge, the only published clinical study on the effectiveness of intranasal OT in reducing pain in patients with FM showed negative results, but as also stated by the authors themselves, this study presents many limitations that do not allow conclusions to be drawn definitive [155]. Indeed, better knowledge of the OTR system is needed which will help explain the mixed results of exogenous OT applications in humans [143]. At the same time, it is important to keep in mind that the effects of OT strongly depend on gender, social context, early life experiences and route of administration, epigenetic mechanisms and OTR system, all variables that will need to be taken into consideration for future studies [143,149]. As also reported in the body of text, depending on the different phases of life (e.g., the first nine months of life of sensitive period), various risk factors (type of attachment of parent in the parent-infant dyad, type of attachment of patient, social experience, genetic and epigenetic factors), could have a negative impact on the physiology of the OS [52,149] which in turn, could have a different type of response to exogeneous administration of OT.
The analgesic efficacy of gentle touch due to the activation of CT fibers has been explored both in healthy subjects to acutely induced pain and in patients with CP, showing a potential therapeutic role of this non-pharmacological treatment in CP conditions [107,109,115,118]. A study conducted on several CP syndromes deserves special mention especially for the rigorous CT fiber stimulation paradigm applied and the significant pain reduction achieved supporting the role of interoceptive tactile stimulation as a non-pharmacological and complementary approach for the treatment of CP [112]. However, in future studies, it will be important to explore the therapeutic potential of CT fibers stimulation also in chronic patients with SFP, that as we previously discussed represents a common condition in patients with FMS and many other diseases [84,85].
A significant clinical approach to understand for pain specialists is that of the neonatal intensive care unit. It has been shown that in the preterm newborn, a family education intervention, according to the calming cycle theory, is able to accelerate the maturation of the preterm newborn’s vagal control, highlighting the importance of the stimulation of the CT fibers and other sensory components of the bio-behavioral synchrony [156]. In this clinical setting, a nurture specialists, represents “a human tool” capable of promoting the system of social involvement between mother and infant, obtaining a positive outcome for both protagonists of the relationship [156]. From this perspective, in clinical practice, and for the scientific data reported above, it is important to explore the type of attachment of patients with CP together with that of the caregiver, in particular in patients with FMS who usually do not have good adherence to pharmacological therapy. In this way, it will be possible to collect a lot of information on the type of relationship between the caregiver-patient dyad, which could be useful for a global clinical approach composed a psychoeducational component, a compassionate physician-patient relationship, and for foster the engagement social system within the dyad physician-patient.
On the other hand, the biopsychosocial model for the clinical evaluation and treatment of patients with CP is based on a patient-centered approach in which individualized and multidisciplinary treatments must always be taken into consideration [157].
Therefore, the most effective treatment for CSSs should always be specifically tailored to each patient and set based on life history and clinical symptoms [17].
In conclusion, we strongly believe that the scientific community should consider extending research in the field of COPC, FSD, CSS, to evaluate the role of the AS, OS and CT fiber system in a multidisciplinary manner.
Improving knowledge of the pathophysiological mechanisms of these functionally integrated systems, probably crucial not only in CP conditions but also in the pathogenesis of chronic-degenerative diseases, could provide doctors and patients with valuable clinical tools to improve the psychological and social functioning of subjects affected by these chronic disorders.
Even more important could be the positive impact on prevention through scientific knowledge-based awareness campaigns for healthcare professionals, patients and the general population.
A reduction of the burden of the disease at an individual and social level, and above all a promotion of cognitive neuroscience and social sciences, should always be pursued in a civil society that looks to the future.
List of Abbreviations
- ACC: Anterior Cingulate Cortex
- ACR: American College of Rheumatology
- ACR: American College of Rheumatology
- AMY: Amygdala
- ANS: Autonomic Nervous System
- AT: Affective Touch
- BDS: Bodily Distress Syndrome
- BSS: bodily stress system
- C-LTMRs: C-low-threshold mechanoreceptors
- COPCs: Chronic Overlapping Pain Conditions
- CP: Chronic Pain
- CPRS: Chronic Pain Regional Syndrome
- CS: Central Sensitization
- CSSs: Central Sensitivity Syndromes
- CT: C tactile
- CWP: Chronic Widespread Pain
- DRG: Dorsal Roots of the Spinal Ganglion
- FITSS: Fibromyalgia: Imbalance of Threat and Soothing Systems
- FM: Fibromyalgia
- fMRI: functional Magnetic Resonance Imaging
- FMS: Fibromyalgia Syndrome
- FSDs: Functional Somatic Disorders
- HPA: Hypothalamus-Pituitary-Axis
- HRV: Heart Rate Variability
- IA: Insecure Attachment
- IA: Insecure Attachment
- IASP: International Association for the Study of Pain
- IC: Insular Cortex
- IENFD: Intraepidermal Nerve Fiber Density
- NAc: Nucleus Accumbens
- OFC: Orbitofrontal Cortex
- OS: Oxytocinergic System
- OS: Oxytocinergic System
- OT: Oxytocin
- OTRs: Oxytocinergic Receptors
- PAG: Periaqueductal Gray Matter
- PFC: Prefrontal Cortex
- PHN: Post-Herpetic Neuralgia
- PTSD: Post-Traumatic Stress Disorder
- PTSS: Post-Traumatic Stress Symptoms
- PVN: Paraventricular Nucleus
- RVM: Rostral Ventromedial Reticular Structure
- SA: Secure Attachment
- SAM: Sympathetic-Adrenal-Medullary
- SFN: Small Fiber Neuropathy
- SFP: Small Fiber Pathology
- SG: Substantia Gelatinosa
- SNS: Sympathetic Nervous System
- SON: Supraoptic Nucleus
- SSC: Skin-to-Skin Contact
- TSSP: Temporal Summation of a Second Pain
- VTA: Ventral Tegmental Area
References
- Wolfe, F.; Clauw, D.J.; Fitzcharles, M. A.; Goldenberg, D. L.; Katz, R. S.; Mease, P.; Russell, A. S.; Russell, I. J.; Winfield, J. B.; Yunus, M. B. The American College of Rheumatology preliminary diagnostic criteria for fibromyalgia and measurement of symptom severity. Arthritis Care Res. 2010, 62, 600–610. [Google Scholar] [CrossRef]
- World Health Organization. International Classification of Diseases (ICD). WHO http://www.who.int/ classifications/icd/en/ (2015). 2015. [Google Scholar]
- Burton, C.; Fink, P.; Henningsen, P.; Löwe, B.; Rief, W.; on behalf of the EURONET-SOMA, Group. Functional somatic disorders: discussion paper for a new common classification for research and clinical use. BMC Med. 2020, 18. [Google Scholar] [CrossRef]
- Yunus, M. B. Central sensitivity syndromes: a new paradigm and group nosology for fibromyalgia and overlapping conditions, and the related issue of disease versus illness. Semin. Arthritis Rheum. 2008, 37, 339–352. [Google Scholar] [CrossRef]
- Veasley, C. in Fibromyalgia Syndrome and Widespread Pain: From Construction to Relevant Recognition (eds Häuser, W. & Perrot, S.) 87–111 (Wolters Kluwer Health, 2018).
- Martinez-Lavin, M. Fibromyalgia as a sympathetically maintained pain syndrome. Curr. Pain. Headache Rep. 2004, 8, 385–389. [Google Scholar] [CrossRef]
- Martinez-Lavin, M. Biology and therapy of fibromyalgia. Stress, the stress response system, and fibromyalgia. Arthritis Res Ther. 2007, 9, 216. [Google Scholar] [CrossRef]
- Kosek, E.; Kadetoff, D. Evidence of reduced sympatho-adrenal and hypothalamic-pituitary activity during static muscular work in patients with fibromyalgia. J. Rehabil. Med, 2010, 42, 765–772. [Google Scholar] [CrossRef]
- Martínez-Martínez, L. A.; Mora, T.; Vargas, A.; Fuentes-Iniestra, M.; Martínez-Lavín, M. Sympathetic Nervous System Dysfunction in Fibromyalgia, Chronic Fatigue Syndrome, Irritable Bowel Syndrome, and Interstitial Cystitis. JCR J. Clin. Rheumatol. 2014, 20, 146–150. [Google Scholar] [CrossRef]
- Tanriverdi, F.; Karaca, Z.; Unluhizarci, K.; Kelestimur, F. “The hypothalamo-pituitary-adrenal axis in chronic fatigue syndrome and fibromyalgia syndrome”. Stress. 2007, 10, 13–25. [Google Scholar] [CrossRef]
- Becker, S.; Schweinhardt, P. Dysfunctional neurotransmitter systems in fibromyalgia, their role in central stress circuitry and pharmacological actions on these systems. Pain Res Treat. 2012, 2012. [Google Scholar] [CrossRef]
- Williams, D.A.; Clauw, D. J. “Understanding fibromyalgia: lessons from the broader pain research community”. Journal of Pain. 2009, 10, 777–791. [Google Scholar] [CrossRef] [PubMed]
- Wyns, A.; Hendrix, J.; Lahousse, A.; De Bruyne, E.; Nijs, J.; Godderis, L.; Polli, A. The Biology of Stress Intolerance in Patients with Chronic Pain—State of the Art and Future Directions. J. Clin. Med. 2023, 12, 2245. [Google Scholar] [CrossRef] [PubMed]
- Riva, R.; Mork, P.; Westgaard, R.; Rø, M.; Lundberg, U. Fibromyalgia syndrome is associated with hypocortisolism. Int. J. Behav. Med. 2010, 17, 223–233. [Google Scholar] [CrossRef] [PubMed]
- Fries, E.; Hesse, J.; Hellhammer, J. and Hellhammer D. H. “A new view on hypocortisolism,” Psychoneuroendocrinology. 2005, 30, 1010–1016. [Google Scholar]
- Reyes del Paso, G. A.; Garcia-Hernandez, A.; Contreras-Merino, A. M.; Galvez-Sánchez, C. M.; de la Coba, P.; Montoro, C. I. , & Davydov, D. M. A two-component model of hair cortisol concentration in fibromyalgia: Independent effects of pain chronicity and severity. European Journal of Pain. 2024, 28, 821–830. [Google Scholar]
- Eller-Smith, O. C.; Nicol, A. L. & Christianson, J. A. Potential mechanisms underlying centralized pain and emerging therapeutic interventions. Frontiers in Cellular Neuroscience. 2018, 12. [Google Scholar]
- Koenig, J.; Falvay, D.; Clamor, A.; Wagner, J.; Jarczok, M. N.; Ellis, R. J.; Weber, C.; Thayer, J. F. Pneumogastric (Vagus) Nerve Activity Indexed by Heart Rate Variability in Chronic Pain Patients Compared to Healthy Controls: A Systematic Review and Meta-Analysis. Pain. Physician. 2016, 19, E55–E78. [Google Scholar] [CrossRef]
- Ying-Chih, C; Yu-Chen, H; Wei-Lieh H. Heart rate variability in patients with somatic symptom disorders and functional somatic syndromes: a systematic review and meta-analysis. Neurosci Biobehav Rev. 2020, 112, 336–344. [Google Scholar] [CrossRef]
- Forte, G.; Troisi, G.; Pazzaglia, M.; Pascalis, V.; Casagrande, M. Heart Rate Variability and Pain: A Systematic Review. Brain. Sci. 2022, 12, 153. [Google Scholar] [CrossRef]
- An, E; Nolty, A. A. T.; Amano, S. S.; Rizzo, A. A.; Buckwalter, J. G.; Rensberger, J. Heart Rate Variability as an Index of Resilience. Mil Med. 2020, 2, 363–369. [Google Scholar]
- Jørgensen, T.; Dantoft, T. M.; Petersen, M.W.; Gormsen, L.; Winter-Jensen, M.; Fink, P; Linneberg, A.; Benros, M. E.; Eplov, L. F.; Bjerregaard, A. A.; Schovsbo, S. U.; Brinth, L. S. Is reduced heart rate variability associated with functional somatic disorders? A cross-sectional population-based study; DanFunD. BMJ Open. 2024, 14, e073909. [Google Scholar]
- Martínez-Martínez, L. A.; Mora, T.; Vargas, A.; Fuentes-Iniestra, M.; Martínez-Lavín, M. Sympathetic nervous system dysfunction in fibromyalgia, chronic fatigue syndrome, irritable bowel syndrome, and interstitial cystitis: a review of case-control studies. J Clin Rheumatol. 2014, 20, 146–150. [Google Scholar] [CrossRef]
- Reyes Del Paso, G. A.; Garrido, S.; Pulgar, A.; Martín-Vázquez, M.; & Duschek, S. Aberrances in autonomic cardiovascular regulation in fibromyalgia syndrome and their relevance for clinical pain reports. Psychosomatic medicine. 2010, 72, 462–470. [Google Scholar] [CrossRef]
- Meeus, M.; Goubert, D.; De Backer, F.; Struyf, F.; Hermans, L.; Coppieters, I.; De Wandele, I.; Da Silva, H.; Calders, P. Heart rate variability in patients with fibromyalgia and patients with chronic fatigue syndrome: a systematic review. Semin Arthritis Rheum. 2013, 43, 279–87. [Google Scholar] [CrossRef]
- Tracy, L. M.; Ioannou, L.; Baker, K. S.; Gibson, S. J.; Georgiou-Karistianis, N.; Giumarra, M. J. Meta-analytic evidence for decreased heart rate variability in chronic pain implicating parasympathetic nervous system dysregulation. Pain. 2016, 157, 7–29. [Google Scholar] [CrossRef]
- Reyes-Manzano, C. F.; Lerma, C.; Echeverría, J. C.; Martínez-Lavin, M.; Martínez-Martínez, L. A.; Infante, O. & Guzmán-Vargas, L. Multifractal Analysis Reveals Decreased Non-linearity and Stronger Anticorrelations in Heart Period Fluctuations of Fibromyalgia Patients. Frontiers in physiology. 2018, 9, 1118. [Google Scholar]
- Reyes Del Paso, G. A. & de la Coba, P. Reduced activity, reactivity and functionality of the sympathetic nervous system in fibromyalgia: an electrodermal study. PLoS One. 2020, 15. [Google Scholar]
- Zetterman, T. , Markkula, R., Miettinen, T. et al. Heart rate variability responses to cognitive stress in fibromyalgia are characterised by inadequate autonomous system stress responses: a clinical trial. Sci Rep. 2023, 13, 700. [Google Scholar] [CrossRef]
- Petersen, M. W.; Schröder, A.; Jørgensen, T.; Ørnbøl, E.; Dantoft, T. M.; Eliasen, M.; Thuesen, B. H.; Fink, P. The unifying diagnostic construct of bodily distress syndrome (BDS) was confirmed in the general population. Journal of psychosomatic research. 2020, 128. [Google Scholar] [CrossRef]
- Petersen, M. W.; Schröder, A.; Jørgensen, T.; Ørnbøl, E.; Meinertz Dantoft, T.; Eliasen, M.; Benros, M. E.; Fink, P. Irritable bowel, chronic widespread pain, chronic fatigue and related syndromes are prevalent and highly overlapping in the general population: DanFunD. Sci Rep. 2020, 24. [Google Scholar] [CrossRef]
- Turner, A. I.; Smyth, N.; Hall, S.J.; Torres, S. J.; Hussein, M.; Jayasinghe, S. U.; Ball, K.; Clow, A. J. Psychological stress reactivity and future health and disease outcomes: A systematic review of prospective evidence. Psychoneuroendocrinology 2020, 114, 104599. [Google Scholar] [CrossRef] [PubMed]
- Raja, S. N.; Carr, D. B.; Cohen, M.; Finnerup, N. B.; Flor, H.; Gibson, S.; Keefe, F. J.; Mogil, J. S.; Ringkamp, M.; Sluka, K. A.; Song, X. J.; Stevens, B.; Sullivan, M. D.; Tutelman, P. R.; Ushida, T.; Vader, K. The revised International Association for the Study of Pain definition of pain: concepts, challenges, and compromises. Pain. 2020, 161, 1976–1982. [Google Scholar] [CrossRef]
- Pinto, A. M.; Geenen, R.; Wager, T. D.; Lumley, M. A.; Häuser, W.; Kosek, E.; Ablin, J. N.; Amris, K.; Branco, J.; Buskila, D.; Castelhano, J.; Castelo-Branco, M.; Crofford, L. J.; Fitzcharles, M. A.; López-Solà, M.; Luís, M.; Marques, T. R.; Mease, P. J.; Palavra, F.; Rhudy, J. L.; Uddin, L.Q.; Castilho, P.; Jacobs, J. W. G.; da Silva, J. A. P. Emotion regulation and the salience network: a hypothetical integrative model of fibromyalgia. Nat Rev Rheumatol. 2023, 19, 44–60. [Google Scholar] [CrossRef]
- Generaal, E.; Vogelzangs, N.; Macfarlane, GJ.; Geenen, R.; Smit, J. H.; de Geus, E. J.; Penninx, B. W.; Dekker, J. Biological stress systems, adverse life events and the onset of chronic multisite musculoskeletal pain: a 6-year cohort study. Ann Rheum Dis. 2016, 75, 847–854. [Google Scholar] [CrossRef] [PubMed]
- Yavne, Y.; Amital, D.; Watad, A.; Tiosano, S.; Amital, H. A systematic review of precipitating physical and psychological traumatic events in the development of fibromyalgia. Semin Arthritis Rheum. 2018, 48, 121–133. [Google Scholar] [CrossRef]
- Smith, K. E.; Pollak, S. D. Early life stress and development: potential mechanisms for adverse outcomes. J Neurodev Disord. 2020, 16, 34. [Google Scholar] [CrossRef] [PubMed]
- Cay, M.; Gonzalez-Heydrich, J.; Teicher, M. H.; van der Heijden, H.; Ongur, D.; Shinn, A. K.; Upadhyay, J. Childhood maltreatment and its role in the development of pain and psychopathology. Lancet Child Adolesc Health. 2022, 6, 195–206. [Google Scholar] [CrossRef]
- Edwards, R. R.; Dworkin, R. H.; Sullivan, M. D.; Turk, D. C.; Wasan, A. D. The Role of Psychosocial Processes in the Development and Maintenance of Chronic Pain. J Pain. 2016, 17, T70–92. [Google Scholar] [CrossRef]
- Afari, N.; Ahumada, S. M.; Wright, L. J.; et al. Psychological trauma and functional somatic syndromes: a systematic review and meta-analysis. Psychosom Med. 2014, 76, 2–11. [Google Scholar] [CrossRef]
- Holley, A.; Wilson, A.; Noel, M.; Palermo, T. Post-traumatic stress symptoms in children and adolescents with chronic pain: A topical review of the literature and a proposed framework for future research. European journal of pain. 2016, 20, 1371–1383. [Google Scholar] [CrossRef]
- Beal, S. J.; Kashikar-Zuck, S.; King, C.; Black, W.; Barnes, J.; Noll, J. G. Heightened risk of pain in young adult women with a history of childhood maltreatment: a prospective longitudinal study. Pain. 2020, 161, 156–165. [Google Scholar] [CrossRef]
- Nelson, S. M.; Cunningham, N. R.; Kashikar-Zuck, S. A conceptual framework for understanding the role of adverse childhood experiences in pediatric chronic pain. The Clinical journal of pain. 2017, 33, 264–270. [Google Scholar] [CrossRef]
- McCrory, E; De Brito, S. A.; Viding, E. The impact of childhood maltreatment: a review of neurobiological and genetic factors. Front Psychiatry. 2011, 28. [Google Scholar]
- Nardi, A. E.; Karam, E. G.; Carta, M. G. Fibromyalgia patients should always be screened for post-traumatic stress disorder. Expert Rev Neurother. 2020, 20, 891–893. [Google Scholar] [CrossRef]
- McKernan, L. C.; Johnson, B. N.; Crofford, L. J.; Lumley, M. A.; Bruehl, S.; Cheavens, J. S. Posttraumatic Stress Symptoms Mediate the Effects of Trauma Exposure on Clinical Indicators of Central Sensitization in Patients With Chronic Pain. Clin J Pain. 2019, 35, 385–393. [Google Scholar] [CrossRef]
- González-Vives, S.; Díaz-Marsá, M.; De la Vega, I.; Palomares, N.; Vázquez, S.; López-Villatoro, J. M.; Palomo, T. & Carrasco, J. L. Hypothalamic-pituitary axis response to a 0.25-MG dexamethasone test in women with fibromyalgia. Stress (Amsterdam, Netherlands). 2020, 23, 284–289. [Google Scholar]
- Bowlby, J. Attachment and Loss: 1. Attachment. London: Hogarth Press. 1969.
- Bartholomew, K. and Horowitz L. M. “Attachment styles among young adults: a test of a four-category model”. Journal of Personality and Social Psychology. 1991, 61, 226–244. [Google Scholar] [CrossRef]
- Vrtička, P. , & Vuilleumier, P. Neuroscience of human social interactions and adult attachment style. Frontiers in human neuroscience. 2012, 6, 212. [Google Scholar]
- Gander, M.; Karabatsiakis, A.; Nuderscher, K.; Bernheim, D.; Doyen-Waldecker, C. , & Buchheim, A. Secure Attachment Representation in Adolescence Buffers Heart-Rate Reactivity in Response to Attachment-Related Stressors. Frontiers in human neuroscience. 2022, 16. [Google Scholar]
- Feldman, R. What is resilience: an affiliative neuroscience approach. World psychiatry : official journal of the World Psychiatric Association (WPA). 2020, 19, 132–150. [Google Scholar] [CrossRef]
- Bryant, R. A.; Hutanamon, T. Activating attachments enhances heart rate variability. PLoS One. 2018, 15. [Google Scholar] [CrossRef]
- Gander, M.; &, *!!! REPLACE !!!*; Buchheim, A. Attachment classification, psychophysiology and frontal EEG asymmetry across the lifespan: a review. Frontiers in human neuroscience. 2015, 9, 79. [Google Scholar] [CrossRef] [PubMed]
- Pietromonaco, P. R.; Powers, S. I. Attachment and Health-Related Physiological Stress Processes. Curr Opin Psychol. 2015, 1, 34–39. [Google Scholar] [CrossRef]
- Godoy, L.; Rossignoli, M.; Delfino-Pereira, P.; Garcia-Cairasco, N.; de Lima Umeoka, E. A. Comprehensive Overview on Stress Neurobiology: Basic Concepts and Clinical Implications. Front. Behav. Neurosci. 2018, 12. [Google Scholar] [CrossRef] [PubMed]
- Pietromonaco, P. R. , Uchino, B., Dunkel Schetter, C. Close relationship processes and health: implications of attachment theory for health and disease. Health Psychol. 2013, 32, 499–513. [Google Scholar] [CrossRef]
- Bosmans, G.; Borelli, J. L. Attachment and the Development of Psychopathology: Introduction to the Special Issue. Brain Sci. 2022, 12, 174. [Google Scholar] [CrossRef]
- Meredith, P.; Ownsworth, T.; Strong, J. A review of the evidence linking adult attachment theory and chronic pain: Presenting a conceptual model. Clin Psychol Rev. 2008, 28, 407–429. [Google Scholar] [CrossRef]
- McWilliams, L. A. Adult attachment insecurity is positively associated with medically unexplained chronic pain. Eur J Pain. 2017, 21, 1378–1383. [Google Scholar] [CrossRef]
- Romeo, A.; Tesio, V.; Castelnuovo, G.; Castelli, L. Attachment Style and Chronic Pain: Toward an Interpersonal Model of Pain. Front Psychol. 2017, 8, 284. [Google Scholar] [CrossRef]
- Davies, K. A.; Macfarlane, G. J.; McBeth, J.; Morriss, R.; Dickens, C. Insecure attachment style is associated with chronic widespread pain. Pain. 2009, 143, 200–205. [Google Scholar] [CrossRef]
- McWilliams, L. A. , & Asmundson, G. J. The relationship of adult attachment dimensions to pain-related fear, hypervigilance, and catastrophizing. Pain. 2007, 127, 27–34. [Google Scholar]
- Meredith, P. J. A review of the evidence regarding associations between attachment theory and experimentally induced pain. Current pain and headache reports. 2013, 17, 326. [Google Scholar] [CrossRef]
- Peñacoba, C.; Perez-Calvo, S.; Blanco, S. , & Sanroman, L. Attachment styles, pain intensity and emotional variables in women with fibromyalgia. Scandinavian journal of caring sciences. 2018, 32, 535–544. [Google Scholar]
- Romeo, A.; Di Tella, M.; Ghiggia, A.; Tesio, V.; Fusaro, E.; Geminiani, G. C.; Castelli, L. Attachment style and parental bonding: Relationships with fibromyalgia and alexithymia. PLoS One. 2020, 15. [Google Scholar] [CrossRef]
- Puşuroğlu, M.; Topaloğlu, M. S.; Hocaoğlu, Ç.; Yıldırım, M. Expressing emotions, rejection sensitivity, and attachment in patients with fibromyalgia. Turk J Phys Med Rehabil. 2023, 69. [Google Scholar] [CrossRef]
- Sechi, C.; Vismara, L.; Brennstuhl, M. J.; Tarquinio, C.; Lucarelli, L. Adult attachment styles, self-esteem, and quality of life in women with fibromyalgia. Health Psychol Open. 2020, 7. [Google Scholar] [CrossRef]
- Sechi, C.; Lucarelli, L.; Vismara, L. Depressive Symptoms and Quality of Life in a Sample of Italian Women with a Diagnosis of Fibromyalgia: The Role of Attachment Styles. Depress Res Treat. 2021, 16. [Google Scholar] [CrossRef]
- Knobloch, H. S.; Grinevich, V. Evolution of oxytocin pathways in the brain of vertebrates. Front Behav Neurosci. 2014, 14. [Google Scholar] [CrossRef]
- Feldman, R.; Monakhov, M.; Pratt, M. & Ebstein, R. P. Oxytocin Pathway Genes: Evolutionary Ancient System Impacting on Human Affiliation, Sociality, and Psychopathology. Biological psychiatry 2016, 79, 174–184. [Google Scholar] [PubMed]
- Wee, C. L.; Nikitchenko, M.; Wang, W. C.; Luks-Morgan, S. J.; Song, E.; Gagnon, J. A.; Randlett, O.; Bianco, I. H.; Lacoste, A. M. B.; Glushenkova, E.; Barrios, J. P.; Schier, A. F.; Kunes, S.; Engert, F.; Douglass, A. D. Zebrafish oxytocin neurons drive nocifensive behavior via brainstem premotor targets. Nat Neurosci. 2019, 22, 1477–1492. [Google Scholar] [CrossRef] [PubMed]
- Feldman, R. The adaptive human parental brain: implications for children’s social development. Trends Neurosci. 2015, 38, 387–399. [Google Scholar] [CrossRef] [PubMed]
- Eliava, M.; Melchior, M.; Knobloch-Bollmann, H. S.; Wahis, J.; da Silva Gouveia, M.; Tang, Y.; Ciobanu, A. C.; Triana Del Rio, R.; Roth, L. C.; Althammer, F.; Chavant, V.; Goumon, Y.; Gruber, T.; Petit-Demoulière, N.; Busnelli, M.; Chini, B.; Tan, L. L.; Mitre, M.; Froemke, R. C.; Chao, M. V.; Giese, G.; Sprengel, R.; Kuner, R.; Poisbeau, P.; Seeburg, P. H.; Stoop, R.; Charlet, A.; Grinevich, V. A New Population of Parvocellular Oxytocin Neurons Controlling Magnocellular Neuron Activity and Inflammatory Pain Processing. Neuron. 2016, 16, 1291–1304. [Google Scholar] [CrossRef] [PubMed]
- Knobloch, H. S.; Charlet, A.; Hoffmann, L. C.; Eliava, M.; Khrulev, S.; Cetin, A. H.; Osten, P.; Schwarz, M. K.; Seeburg, P. H.; Stoop, R. , & Grinevich, V. Evoked axonal oxytocin release in the central amygdala attenuates fear response. Neuron. 2012, 73, 553–566. [Google Scholar] [PubMed]
- Poisbeau, P.; Grinevich, V. & Charlet A. Oxytocin Signaling in Pain: Cellular, Circuit, System, and Behavioral Levels. Chapter in Current topics in behavioral neurosciences 2017. [Google Scholar]
- Yang, L. N.; Chen, K.; Yin, X. P.; Liu, D.; Zhu, L. Q. The Comprehensive Neural Mechanism of Oxytocin in Analgesia. Curr Neuropharmacol. 2022, 20, 147–157. [Google Scholar] [CrossRef] [PubMed]
- De Ridder, D.; Adhia, D. & Vanneste, S. The anatomy of pain and suffering in the brain and its clinical implications. Neuroscience and biobehavioral reviews. 2021, 130, 125–146. [Google Scholar] [PubMed]
- Aygün, O.; Mohr, E.; Duff, C.; Matthew, S.; Schoenberg, P. Oxytocin Modulation in Mindfulness-Based Pain Management for Chronic Pain. Life (Basel). 2024, 14, 253. [Google Scholar] [CrossRef] [PubMed]
- Takayanagi, Y.; Onaka, T. Roles of Oxytocin in Stress Responses, Allostasis and Resilience. Int J Mol Sci. 2021, 23, 150. [Google Scholar] [CrossRef] [PubMed]
- Wolfe, F. Fibromyalgianess. Arthritis Rheum. 2009, 61, 715–716. [Google Scholar] [CrossRef]
- Häuser, W.; Schmutzer, G.; Brähler, E. & Glaesmer, H. A cluster within the continuum of biopsychosocial distress can be labeled “fibromyalgia syndrome” — evidence from a representative German population survey. J. Rheumatol. 2009, 36, 2806–2812. [Google Scholar]
- Wolfe, F.; Brähler, E.; Hinz, A. & Häuser, W. Fibromyalgia prevalence, somatic symptom reporting, and the dimensionality of polysymptomatic distress: results from a survey of the general population. Arthritis Care Res. 2013, 65, 777–785. [Google Scholar]
- Grayston, R.; Czanner, G.; Elhadd, K.; Goebel, A.; Frank, B.; Üçeyler, N.; Malik, R. A.; Alam, U. A systematic review and meta-analysis of the prevalence of small fiber pathology in fibromyalgia: Implications for a new paradigm in fibromyalgia etiopathogenesis. Semin Arthritis Rheum. 2019, 48, 933–940. [Google Scholar] [CrossRef]
- Thomas, S. J.; Enders, J.; Kaiser, A.; Rovenstine, L.; Heslop, L.; Hauser, W.; Chadwick, A.; Wright, D. E. Abnormal Intraepidermal Nerve Fiber Density in Disease: A Scoping Review. Front Neurol. 2023, 20. [Google Scholar] [CrossRef]
- Marshall, A.; Rapteas, L.; Burgess, J.; Riley, D.; Anson, M.; Matsumoto, K.; Bennett, A.; Kaye, S.; Marshall, A.; Dunham, J.; Fallon, N.; Zhao, S. S.; Pritchard, A.; Goodson, N.; Malik, R. A.; Goebel, A.; Frank, B.; Alam, U. Small fibre pathology, small fibre symptoms and pain in fibromyalgia syndrome. Sci Rep. 2024, 14. [Google Scholar] [CrossRef]
- Harte, S. E.; Clauw, D. J.; Hayes, J. M.; Feldman, E. L.; St Charles, I. C.; Watson, C. J. Reduced intraepidermal nerve fiber density after a sustained increase in insular glutamate: a proof-of-concept study examining the pathogenesis of small fiber pathology in fibromyalgia. Pain Rep. 2017, 2. [Google Scholar] [CrossRef]
- Martínez-Lavín, M. Dorsal root ganglia: fibromyalgia pain factory? Clin Rheumatol. 2021, 40. [Google Scholar] [CrossRef]
- Goebel, A; Krock, E. ; Gentry, C.; Israel, M. R.; Jurczak, A.; Urbina, C. M.; Sandor, K.; Vastani, N.; Maurer, M.; Cuhadar, U.; Sensi, S.; Nomura, Y.; Menezes, J.; Baharpoor, A.; Brieskorn, L.; Sandström, A.; Tour, J.; Kadetoff, D., Haglund, L.; Kosek, E.; Bevan, S.; Svensson, C. I.; Andersson, D. A. Passive transfer of fibromyalgia symptoms from patients to mice. J Clin Invest. 2021, 131. [Google Scholar]
- Liptan, G. The widespread myofascial pain of fibromyalgia is sympathetically maintained and immune mediated. J Bodyw Mov Ther. 2023, 35, 394–399. [Google Scholar] [CrossRef]
- Ramírez, M.; Guerra-Juárez, A.; Miyake, D. Y.; Sebastian-Arellano, C.; Estrada-Mata, A. G.; González-Moyotl, N. J.; Rodríguez-Aguayo, A. M.; Martínez-Lavin, M.; Martínez-Martínez, L. A. Correlation Between Corneal Nerve Density and Symptoms of Small Fiber Neuropathy in Patients With Fibromyalgia: The Confounding Role of Severe Anxiety or Depression. J Clin Rheumatol. 2021, 27. [Google Scholar] [CrossRef]
- Jänsch, S.; Evdokimov, D.; Egenolf, N.; Meyer Zu Altenschildesche, C.; Kreß, L.; Üçeyler, N. Distinguishing fibromyalgia syndrome from small fiber neuropathy: a clinical guide. Pain Rep. 2024, 9. [Google Scholar] [CrossRef]
- Üçeyler, N.; Zeller, D.; Kahn, A. K.; Kewenig, S.; Kittel-Schneider, S.; Schmid, A.; Casanova-Molla, J.; Reiners, K.; Sommer, C. Small fibre pathology in patients with fibromyalgia syndrome. Brain. 2013, 136, 1857–1867. [Google Scholar] [CrossRef]
- Shea, V. K.; Perl, E. R. Sensory receptors with unmyelinated (C) fibers innervating the skin of the rabbit’s ear. J. Neurophysiol. 1985, 54, 491–501. [Google Scholar] [CrossRef]
- Lawson, S. N.; Crepps, B.A.; Perl, E. R. . Relationship of substance P to afferent characteristics of dorsal root ganglion neurones in guinea-pig. J. Physiol. 1997, 505, 177–191. [Google Scholar] [CrossRef]
- Nordin, M. Low-threshold mechanoreceptive and nociceptive units with unmyelinated (C) fibres in the human supraorbital nerve. J Physiol. 1990, 426, 229–240. [Google Scholar] [CrossRef]
- Vallbo, A. B.; Olausson, H.; and Wessberg, J. Unmyelinated afferents constitute a second system coding tactile stimuli of the human hairy skin. J. Neurophysiol. 1999, 81, 2753–2763. [Google Scholar] [CrossRef]
- Vallbo, Å. B.; Olausson, H.; Wessberg, J. Pleasant touch. In: Encyclopedia of neuroscience (Squire LR, ed). Amsterdam: Elsevier. 2009.
- Schirmer, A.; Croy, I.; Ackerley, R. What are C-tactile afferents and how do they relate to “affective touch”? Neurosci Biobehav Rev. 2023, 151. [Google Scholar] [CrossRef]
- Ackerley, R. C-tactile (CT) afferents: Evidence of their function from microneurography studies in humans. Curr. Opin. Behav. Sci. 2022, 43, 95–100. [Google Scholar] [CrossRef]
- Case, L. K.; Čeko, M.; Gracely, J. L.; Richards, E. A.; Olausson, H.; Bushnell, M. C. Touch Perception Altered by Chronic Pain and by Opioid Blockade. eNeuro. 2016, 3. [Google Scholar] [CrossRef]
- Boehme, R.; van Ettinger-Veenstra, H.; Olausson.; H, Gerdle, B.; Nagi, S. S. Anhedonia to Gentle Touch in Fibromyalgia: Normal Sensory Processing but Abnormal Evaluation. Brain Sci. 2020, 10. [Google Scholar] [CrossRef]
- Lapp, H. S.; Sabatowski, R.; Weidner, K.; Croy, I.; Gossrau, G. C-tactile touch perception in migraineurs - a case-control study. Cephalalgia. 2020, 40, 478–492. [Google Scholar] [CrossRef]
- Nees, F.; Usai, K.; Löffler, M.; Flor, H. The evaluation and brain representation of pleasant touch in chronic and subacute back pain. Neurobiol Pain. 2018, 5. [Google Scholar] [CrossRef] [PubMed]
- Gossrau, G.; Klimova, A.; Lapp, H. S.; Frost, M.; Peschel, E.; Weidner, K.; Koch, T.; Sabatowski, R.; Croy, I. C-tactile touch perception in patients with chronic pain disorders. Pain Rep. 2021, 6. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, V. F.; Karlsson, P.; Drummond, P. D.; Schaldemose, E. L.; Terkelsen, A. J.; Jensen, T. S.; Knudsen, L. F. Bilaterally Reduced Intraepidermal Nerve Fiber Density in Unilateral CRPS-I. Pain Med. 2018, 19. [Google Scholar] [CrossRef] [PubMed]
- Fusaro, M.; Bufacchi, R.J.; Nicolardi, V.; Provenzano, L. The analgesic power of pleasant touch in individuals with chronic pain: Recent findings and new insights. Front Integr Neurosci. 2022, 16. [Google Scholar] [CrossRef]
- Larsson, M. and Nagi S. S. Role of C-tactile fibers in pain modulation: animal and human perspectives. Current Opinion in Behavioral Sciences 2022, 43, 1–7. [Google Scholar]
- Liljencrantz, J.; Strigo, I.; Ellingsen, D. M.; Krämer, H. H.; Lundblad, L. C.; Nagi S., S.; Leknes, S.; Olausson, H. Slow brushing reduces heat pain in humans. Eur J Pain. 2017, 21. [Google Scholar] [CrossRef] [PubMed]
- Delfini, M. C.; Mantilleri, A.; Gaillard, S.; Hao, J.; Reynders, A.; Malapert, P.; Alonso, S.; François, A.; Barrere, C.; Seal, R.; Landry, M.; Eschallier, A.; Alloui, A.; Bourinet, E.; Delmas, P.; Le Feuvre, Y.; Moqrich, A. TAFA4, a chemokine-like protein, modulates injury-induced mechanical and chemical pain hypersensitivity in mice. Cell Rep. 2013, 5. [Google Scholar] [CrossRef] [PubMed]
- Walker, S. C.; Trotter, P. D.; Swaney, W. T.; Marshall, A. , and Mcglone, F. P. C-tactile afferents: Cutaneous mediators of oxytocin release during affiliative tactile interactions? Neuropeptides. 2017a, 64, 27–38. [Google Scholar] [CrossRef] [PubMed]
- Di Lernia, D.; Cipresso, P.; Pedroli, E. , and Riva, G. Toward an embodied medicine: A portable device with programmable interoceptive stimulation for heart rate variability enhancement. Sensors. 2018, 18. [Google Scholar] [CrossRef]
- Nummenmaa, L.; Tuominen, L.; Dunbar, R.; Hirvonen, J.; Manninen, S.; Arponen, E.; et al. Social touch modulates endogenous μ-opioid system activity in humans. Neuroimage. 2016, 138, 242–247. [Google Scholar] [CrossRef]
- Meijer, L. L.; Ruis, C.; van der Smagt, M. J.; Scherder, E. J. and Dijkerman, H. C. Neural basis of affective touch and pain: A novel model suggests possible targets for pain amelioration. J. Neuropsychol. 2022, 16, 38–53. [Google Scholar] [CrossRef] [PubMed]
- Fidanza, F.; Polimeni, E.; Pierangeli, V.; Martini, M. A Better Touch: C-tactile Fibers Related Activity is Associated to Pain Reduction During Temporal Summation of Second Pain. J Pain. 2021, 22, 567–576. [Google Scholar] [CrossRef] [PubMed]
- Hashmi, J. A.; Baliki, M. N.; Huang, L.; Baria, A. T.; Torbey, S.; Hermann, K. M.; Schnitzer, T. J.; Apkarian, A. V. Shape shifting pain: chronification of back pain shifts brain representation from nociceptive to emotional circuits. Brain. 2013, 136, 2751–2768. [Google Scholar] [CrossRef] [PubMed]
- Habig, K.; Schänzer, A.; Schirner, W.; Lautenschläger, G.; Dassinger, B. , Olausson, H.; Birklein, F.; Gizewski, E. R.; Krämer, H. H. Low threshold unmyelinated mechanoafferents can modulate pain. BMC Neurol. 2017, 17, 184. [Google Scholar] [CrossRef] [PubMed]
- Habig, K.; Lautenschläger, G.; Maxeiner, H.; Birklein, F.; Krämer, H. H.; Seddigh, S. Low mechano-afferent fibers reduce thermal pain but not pain intensity in CRPS. BMC Neurol. 2021, 21, 272. [Google Scholar] [CrossRef] [PubMed]
- Di Lernia, D.; Lacerenza, M.; Ainley, V.; Riva, G. Altered Interoceptive Perception and the Effects of Interoceptive Analgesia in Musculoskeletal, Primary, and Neuropathic Chronic Pain Conditions. J Pers Med. 2020, 10, 201. [Google Scholar] [CrossRef] [PubMed]
- Horsburgh, A.; Summers, S. J.; Lewis, A.; Keegan, R. J.; Flood, A. The Relationship Between Pain and Interoception: A Systematic Review and Meta-Analysis. J Pain. 2024, 18. [Google Scholar] [CrossRef] [PubMed]
- Croy, I.; Fairhurst, M. T. , and McGlone, F. The role of C-tactile nerve fibers in human social development. Curr. Op. Behav. Sci. 2022, 43, 20–26. [Google Scholar]
- Morrison, I. ALE meta-analysis reveals dissociable networks for affective and discriminative aspects of touch. Hum Brain Mapp. 2016, 37, 1308–1320. [Google Scholar] [CrossRef]
- Li, Q.; Zhao, W.; Kendrick, K. M. Affective touch in the context of development, oxytocin signaling, and autism. Front Psychol. 2022, 23, 13. [Google Scholar] [CrossRef]
- Bystrova, K. Novel mechanism of human fetal growth regulation: a potential role of lanugo, vernix caseosa and a second tactile system of unmyelinated low-threshold C-afferents. Med Hypotheses. 2009, 72, 143–146. [Google Scholar] [CrossRef] [PubMed]
- Feldman, R.; Eidelman, A. I.; Sirota, L., and Weller, A. Comparison of skin-to-skin and traditional care: parenting outcomes and preterm infant development. Pediatrics. 2001, 110, 16–26. [Google Scholar]
- Jefferies, A. L. Kangaroo care for the preterm infant and family. Paediatr. Child Health. 2012, 17, 141–143. [Google Scholar] [CrossRef] [PubMed]
- Bystrova, K.; Widström, A. M.; Matthiesen, A. S.; Ransjö-Arvidson AB, Welles-Nyström B, Wassberg C, Vorontsov I, Uvnäs-Moberg K. Skin-to-skin contact may reduce negative consequences of “the stress of being born”: a study on temperature in newborn infants, subjected to different ward routines in St. Petersburg. Acta Paediatr. 2003, 92, 320–326. [Google Scholar] [CrossRef]
- Van Puyvelde, M.; Collette, L.; Gorissen, A. S.; Pattyn, N.; McGlone, F. Infants Autonomic Cardio- Respiratory Responses to Nurturing Stroking Touch Delivered by the Mother or the Father. Front Physiol. 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Manzotti, A.; Cerritelli, F.; Esteves, J. E.; Lista, G.; Lombardi, E.; La Rocca, S.; Gallace, A.; McGlone, F. P.; Walker, S. C. Dynamic touch reduces physiological arousal in preterm infants: A role for c-tactile afferents? Dev Cogn Neurosci. 2019, 39. [Google Scholar] [CrossRef] [PubMed]
- Gursul, D.; Goksan, S.; Hartley, C.; Schmidt Mellado, G.; Moultrie, F.; Hoskin, A.; Adams, E.; Hathway, G.; Walker, S.; McGlone, F and Slater R. Stroking modulates noxious-evoked brain activity in human infants. Curr. Biol. 2018, 28, R1380–R1381. [Google Scholar] [CrossRef]
- Carozza, S.; Leong, V. The Role of Affectionate Caregiver Touch in Early Neurodevelopment and Parent-Infant Interactional Synchrony. Front Neurosci 2021, 14. [Google Scholar] [CrossRef] [PubMed]
- Liao, P. Y.; Chiu, Y. M.; Yu, J. H.; Chen, S. K. Mapping Central Projection of Oxytocin Neurons in Unmated Mice Using Cre and Alkaline Phosphatase Reporter. Front Neuroanat. 2020, 19. [Google Scholar] [CrossRef]
- Uvnäs-Moberg, K. , and Petersson, M. Physiological effects induced by stimulation of cutaneous sensory nerves, with a focus on oxytocin. Curr. Op. Behav. Sci. 2022, 43, 159–166. [Google Scholar]
- Bharadwaj, V. N.; Tzabazis, A. Z.; Klukinov, M.; Manering, N. A.; Yeomans, D. C. Intranasal Administration for Pain: Oxytocin and Other Polypeptides. Pharmaceutics. 2021, 13. [Google Scholar] [CrossRef]
- Uvnäs-Moberg, K.; Handlin, L. , and Petersson, M. Self-soothing behaviors with particular reference to oxytocin release induced by non-noxious sensory stimulation. Front. Psychol. 2015, 5. [Google Scholar] [CrossRef] [PubMed]
- McGlone, F.; Wessberg, J.; Olausson, H. Discriminative and affective touch: sensing and feeling. Neuron. 2014, 82, 737–755. [Google Scholar] [CrossRef]
- Yoshida, S.; Funato, H. Physical contact in parent-infant relationship and its effect on fostering a feeling of safety. iScience. 2021, 24. [Google Scholar] [CrossRef]
- Yu, H.; Miao, W.; Ji, E.; Huang, S.; Jin, S.; Zhu, X.; Liu, M. Z.; Sun, Y. G.; Xu, F.; Yu, X. Social touch-like tactile stimulation activates a tachykinin 1-oxytocin pathway to promote social interactions. Neuron. 2022, 110, 1051–1067. [Google Scholar] [CrossRef] [PubMed]
- Bucsea, O.; Pillai Riddell, R. Non-pharmacological pain management in the neonatal intensive care unit: Managing neonatal pain without drugs. Semin Fetal Neonatal Med. 2019, 24. [Google Scholar] [CrossRef]
- Failo, A.; Giannotti, M.; Venuti, P. Associations between attachment and pain: From infant to adolescent. SAGE Open Med. 2019, 7. [Google Scholar] [CrossRef]
- Martínez-Lavín, M. Imbalance of threat and soothing systems in fibromyalgia: rephrasing an established mechanistic model? Nat Rev Rheumatol. 2023, 19. [Google Scholar] [CrossRef] [PubMed]
- Ciuffini, R.; Cofini, V.; Muselli, M.; Necozione, S.; Piroli, A.; Marrelli, A. Emotional arousal and valence in patients with fibromyalgia: a pilot study. Front Pain Res (Lausanne). 2023, 4. [Google Scholar] [CrossRef]
- Janeček, M.; Dabrowska, J. Oxytocin facilitates adaptive fear and attenuates anxiety responses in animal models and human studies-potential interaction with the corticotropin-releasing factor (CRF) system in the bed nucleus of the stria terminalis (BNST). Cell Tissue Res. 2019, 375, 143–172. [Google Scholar] [CrossRef]
- Wee, C. L.; Nikitchenko, M.; Wang, W. C.; Luks-Morgan, S. J.; Song, E.; Gagnon, J. A.; Randlett, O.; Bianco, I. H.; Lacoste, A. M. B.; Glushenkova, E.; Barrios, J. P.; Schier, A. F.; Kunes, S.; Engert, F.; Douglass, A. D. Zebrafish oxytocin neurons drive nocifensive behavior via brainstem premotor targets. Nat Neurosci. 2019, 22, 1477–1492. [Google Scholar] [CrossRef] [PubMed]
- Grinevich, V.; Neumann, I. D. Brain oxytocin: how puzzle stones from animal studies translate into psychiatry. Mol Psychiatry. 2021, 26, 265–279. [Google Scholar] [CrossRef]
- Eliava, M.; Melchior, M.; Knobloch-Bollmann, H. S.; Wahis, J.; da Silva Gouveia, M.; Tang, Y.; Ciobanu, A. C.; Triana Del Rio, R.; Roth, L. C.; Althammer, F.; Chavant, V.; Goumon, Y.; Gruber, T.; Petit-Demoulière, N.; Busnelli, M.; Chini, B.; Tan, L. L.; Mitre, M.; Froemke, R. C.; Chao, M. V.; Giese, G.; Sprengel, R.; Kuner, R.; Poisbeau, P.; Seeburg, P. H.; Stoop, R.; Charlet, A.; Grinevich, V. A New Population of Parvocellular Oxytocin Neurons Controlling Magnocellular Neuron Activity and Inflammatory Pain Processing. Neuron. 2016, 89, 1291–1304. [Google Scholar] [CrossRef] [PubMed]
- Friuli, M.; Eramo, B.; Valenza, M.; Scuderi, C.; Provensi, G.; Romano, A. Targeting the Oxytocinergic System: A Possible Pharmacological Strategy for the Treatment of Inflammation Occurring in Different Chronic Diseases. Int J Mol Sci. 2021, 22. [Google Scholar] [CrossRef] [PubMed]
- Lee, J. H.; Espinera, A. R.; Chen, D.; Choi, K. E.; Caslin, A. Y.; Won, S.; Pecoraro, V.; Xu, G. Y.; Wei, L.; Yu, S. P. Neonatal inflammatory pain and systemic inflammatory responses as possible environmental factors in the development of autism spectrum disorder of juvenile rats. J Neuroinflammation. 2016, 13. [Google Scholar] [CrossRef] [PubMed]
- Onaka, T.; Takayanagi, Y. The oxytocin system and early-life experience-dependent plastic changes. J Neuroendocrinol. 2021, 33. [Google Scholar] [CrossRef] [PubMed]
- McCrory, E.; De Brito, S. A.; Viding, E. The impact of childhood maltreatment: a review of neurobiological and genetic factors. Front Psychiatry. 2011, 2. [Google Scholar] [CrossRef]
- Nagi, S. S.; Rubin, T. K.; Chelvanayagam, D. K.; Macefield, V. G.; Mahns, D. A. Allodynia mediated by C-tactile afferents in human hairy skin. J Physiol. 2011, 589, 4065–4075. [Google Scholar] [CrossRef] [PubMed]
- Nagi, S. S.; Mahns, D. A. Mechanical allodynia in human glabrous skin mediated by low-threshold cutaneous mechanoreceptors with unmyelinated fibres. Exp Brain Res. 2013, 231, 139–151. [Google Scholar] [CrossRef]
- Goodin, B. R.; Ness, T. J.; Robbins, M. T. Oxytocin - a multifunctional analgesic for chronic deep tissue pain. Curr Pharm Des. 2015, 21, 906–913. [Google Scholar] [CrossRef]
- Mekhael, A. A.; Bent, J. E. , Fawcett, J. M., Campbell, T. S., Aguirre-Camacho, A., Farrell, A., & Rash, J. A. Evaluating the efficacy of oxytocin for pain management: An updated systematic review and meta-analysis of randomized clinical trials and observational studies. Canadian journal of pain = Revue canadienne de la douleur 2023, 7. [Google Scholar]
- Mameli, S.; Pisanu, G. M.; Sardo, S.; Marchi, A.; Pili, A.; Carboni, M.; Minerba, L.; Trincas, G.; Carta, M. G.; Melis, M. R.; Agabio, R. Oxytocin nasal spray in fibromyalgic patients. Rheumatol Int. 2014, 34, 1047–1052. [Google Scholar] [CrossRef] [PubMed]
- Porges, S. W.; Davila, M. I.; Lewis, G. F.; et al. Autonomic regulation of preterm infants is enhanced by Family Nurture Intervention. Dev Psychobiol. 2019, 61, 942–952. [Google Scholar] [CrossRef] [PubMed]
- Chandler Iii, G.; Rojas, A. M.; Worts, P. R.; Flynn, H. A. Utilizing Multidisciplinary Medicine in Pain Management: A Narrative Review. Pain Physician. 2021, 24, 369–378. [Google Scholar]
Figure 1.
The four subtypes of adult attachment style according to Bartholomew and Horowitz’s model [49].
Figure 1.
The four subtypes of adult attachment style according to Bartholomew and Horowitz’s model [49].
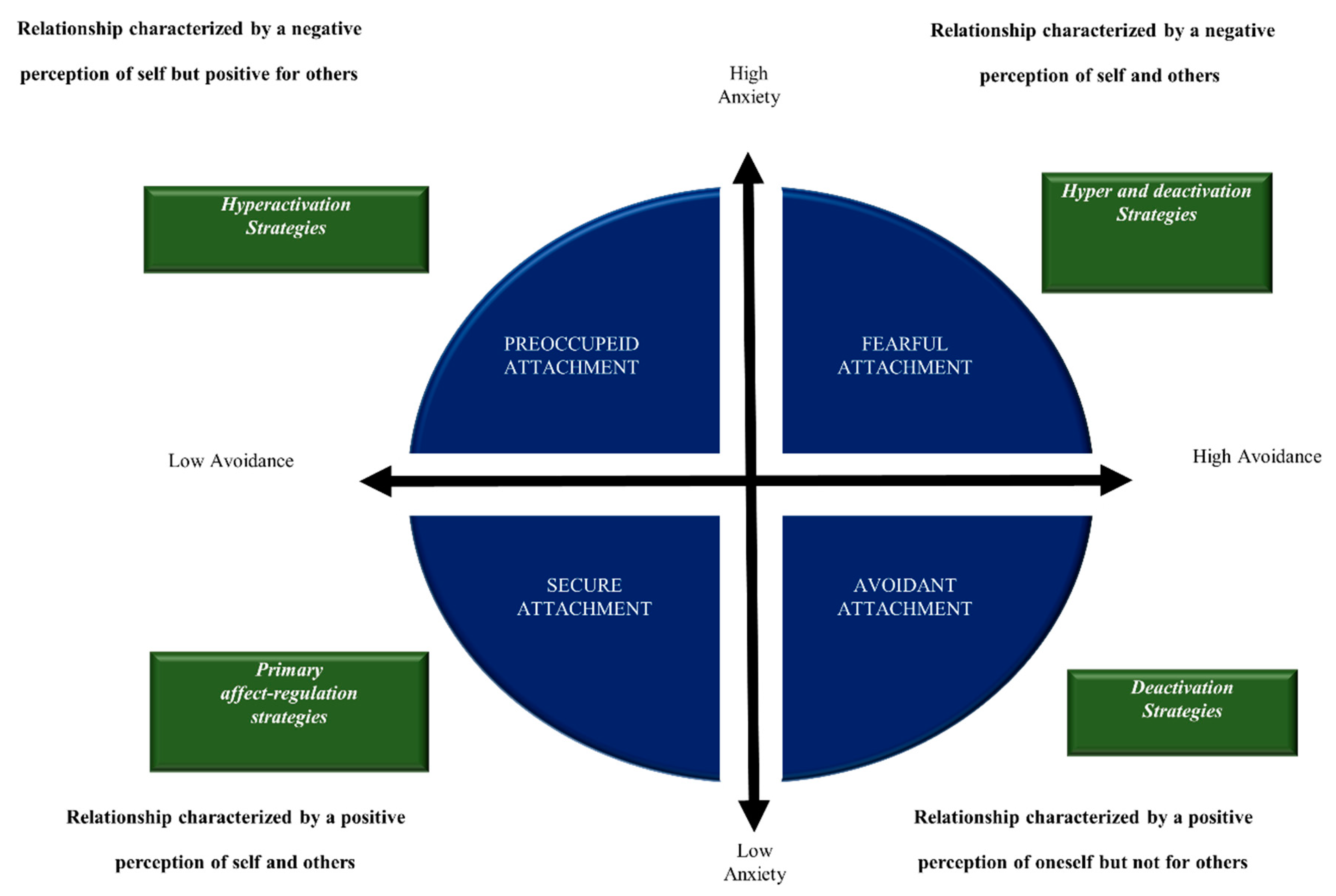
Figure 2.
Good resilience and pain modulation as a result of a physiological functional integration between the oxytocinergic system, CT fiber system and attachment system.
Figure 2.
Good resilience and pain modulation as a result of a physiological functional integration between the oxytocinergic system, CT fiber system and attachment system.
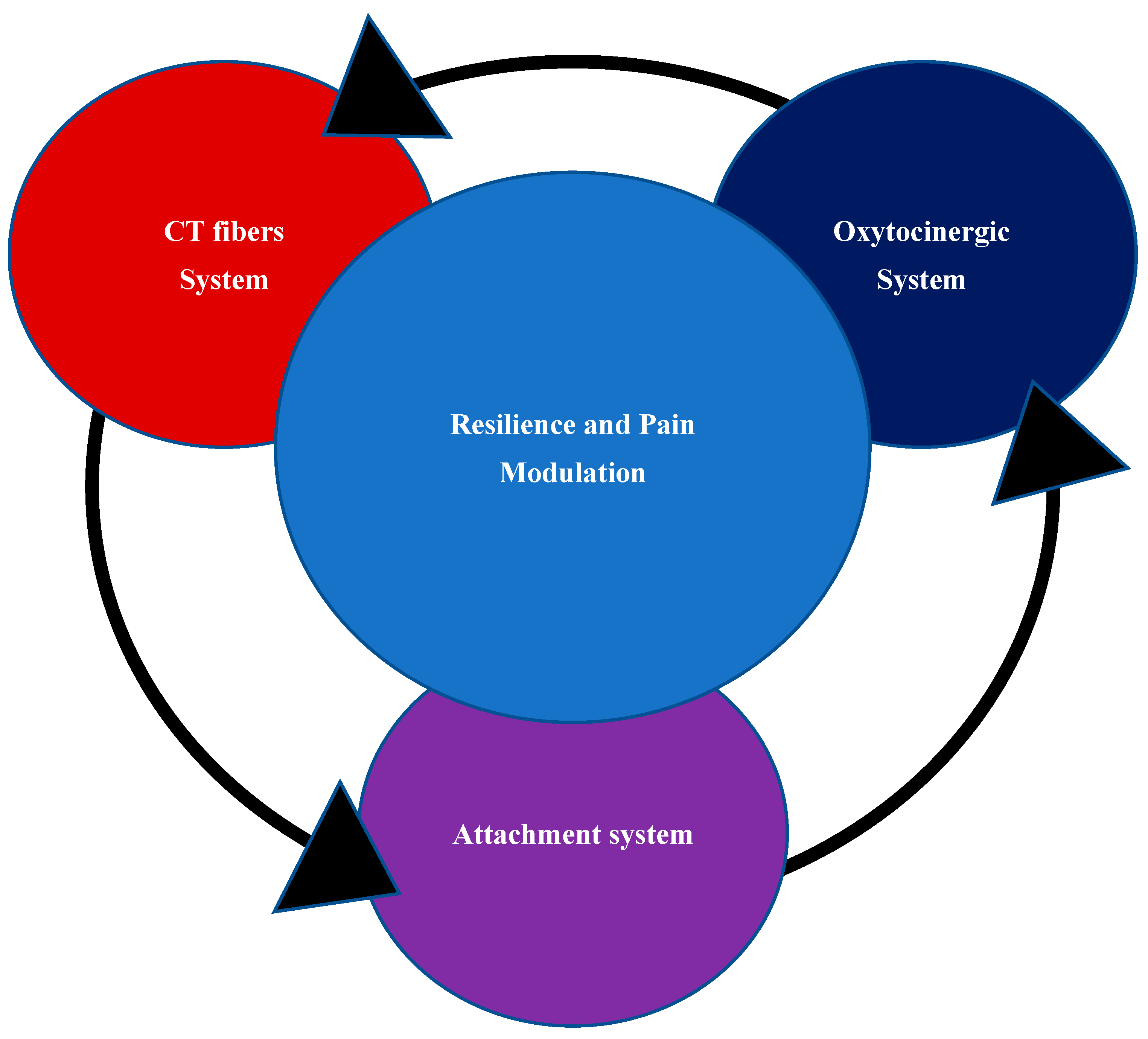

Table 1.
Cluster of symptoms and related organ system classification for the diagnosis of FMS according to ACR diagnostic criteria, Adapted from the reference [1].
Table 1.
Cluster of symptoms and related organ system classification for the diagnosis of FMS according to ACR diagnostic criteria, Adapted from the reference [1].
| Organ system | Symptoms |
|---|---|
| The nervous system | depression, thinking or remembering problems, numbness/tingling, nervousness, seizures, hearing difficulties, headache, dizziness, insomnia, blurred vision, ringing in ears |
| The gastrointestinal system | pain/cramps in the abdomen, constipation, pain in the upper abdomen, nausea, diarrhea, loss of appetite, loss of/change in taste, vomiting, oral ulcers, heartburn |
| The immune system | rash, sun sensitivity, easy bruising, dry eyes, fever, dry mouth, Raynaud’s phenomenon, itching, hives/welts |
| The musculoskeletal system | muscle pain, fatigue/tiredness, muscle weakness |
| The urinary system | bladder spasms, painful urination, frequent urination |
| The respiratory system | chest pain, shortness of breath, wheezing |
| The integumentary system | hair loss |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



