
Submitted:
13 June 2024
Posted:
14 June 2024
You are already at the latest version
Abstract
Pyrops candelaria is one of the common pests of fruit trees, but the research on the pathogenic microorganisms it may carry is very limited. Therefore, it is essential to reveal the pathogenic microbes it carries and their potential hazards. This study found a new virus from the transcriptome of P. candelaria, which was first reported in P. candelaria and named as PyCaV (Pyrops candelaria associated virus). RACE and bioinformatics assay revealed that the full-length of PyCaV is 10855 bp with the polyA tail, containing a single open reading frame (ORF) encoding a polyprotein consisting of 3171 amino acid (aa). The virus has a typical Iflavirus structure, including two rhv domains, a RNA helicase domain (HEL), a 3C cysteine protease domain (Pro), and a RNA-dependent RNA polymerase domain (RdRp). Further phylogenetic analysis revealed that this virus belongs to family Iflaviridae and sequence alignements analysis suggedted PyCaV is new member in unassigned genus of family Iflaviridae. Further in-depth analysis of virus-infection-part showed that PyCaV is distributing in the whole longan lanternfly, including its head, chest, and abdomen, but more PyCaV was identified in the chest. The distribution of PyCaV in different parts of P. candelaria was further explored, which showed that more PyCaV were detected in its piercing-sucking mouthparts and chest viscera. Statistical analysis showed that the PyCaV infection was affected by time and locations.
Keywords:
Iflavirus
; Pyrops candelaria
; virus
; insect
; distribution
1. Introduction
According to the different virus hosts, viruses can be roughly classified into plant viruses, animal viruses, and bacterial viruses. Iflavirus, belonging to small RNA viruses, is an animal virus composed of single-stranded, sense, non-segmented RNA sequence [1]. According to the Iflaviridae chapter of the ICTV online report, there are 16 recognized species within the genus Iflavirus. The virus particles of this genus are roughly spherical, with a diameter of about 22-30 nm. Iflavirus has the basic characteristics of small RNA viruses: the 5’ end of the RNA strand is connected to the viral protein genome (VPg), and the internal ribosome entry site (IRES)a is contained in the 5’ non-coding region, which mediates the initiation of RNA translation [2,3]. Therefore, Iflavirus can initiate RNA translation independent of the 5’ cap structure [4]. Iflavirus has a single open reading frame (ORF), which can be directly translated into a polyprotein, which then post-translationally hydrolyzed into several structural and nonstructural proteins (functional proteins), i.e., structural proteins and functional proteins are cleaved and separated after translation before implement their biological functions. The 3’ non-coding region of Iflavirus contains a typical poly-A (adenine) tail [2,5]. According to the related reports, a variety of Iflavirus viruses have been found, such as deformed wing virus [6], infectious flacheric virus [7], Perina nuda picornalike virus [8], Ectropis obliqua picornalike virus [9], Varrora destructor virus-1 virus [10] and sacbrood virus(SBV) [11].
At present, most known hosts of iflaviruses are arthropods, especially members of Hemiptera and Lepidoptera [12,13]. Infested hosts exhibit abnormal growth and development and have a high mortality rate [14]. However, not all iflaviruses cause observable pathology, and many appear to be transmitted vertically. The gregariousness of insects promotes the spread of viruses in their community [15]. Once the Iflavirus enters the host cell, the infection process is very rapid, and the offsprings of the virus replicate in large numbers within a few hours, and when Iflavirus infects the host, it will quickly infect the brain, intestine, and gland tissues of the host [16]. Iflaviruses pose a significant threat to certain essential environmental insects. It has been reported that Iflavirus infection of Spodoptera exigua (S. exigua) can increase host's susceptibility to nucleopolyhedrovirus infection, which could increase the larval mortality of S. exigua [17]. Infected by Iflavirus, the weight of S. exigua larvae is reduced and the sex ratio is unbalanced. Helicoverpa armigera is one of the most important agricultural pests of cotton and other important economic crops in the world. Iflavirus is mainly distributed in the fat bodies of larvae and adults of the infected H. armigera, which is not conducive to the growth and reproduction of this notorious pest [18]. In some cases, Iflavirus infection can be fatal to the host. sacbrood virus (SBV) mainly exists in the fat cells of the larvae of honeybees, impairing the metabolic function of honeybees and leading the failure of bee pupation, and ultimately causing the death of the hosts [19,20].
The Homoptera insect, Pyrops candelaria, is strange and novel for its long nose and brightly colorful wings. P. candelaria is also called Longan lanternfly, mainly damaging longan, litchi, olive, orange, mulberry, mango, and pomelo crops by sucking the juice from the trunks of these fruit trees [21,22]. Longan lanternflies prefer warm temperatures and high moisture, so they are distributed in the fruit-bearing forests in the low-altitude regions of the tropic and subtropic area, and they are found mainly in China (including Hainan, Fujian, Guangdong, Guangxi, and Hong Kong), Thailand, Vietnam, Malaysia, Cambodia, Sikkim, India, and Indonesia [21,22]. The development of longan lanternflies experiences three stages: egg, nymph, and adult, and their host preference are broad in the nymph stage, but become narrow to a few plant species in the adult stage, and the invasion of longan lanternflies serious endangers the culture and harvest of pomelo and longan trees [22]. The nymph of P.candelaria is good at bouncing, and the adult is good at bouncing and flying [23]. P.candelaria mainly endangers longan trees, when P. candelaria is prevalent, the dense population of the insects causes serious damage to longan trees. Adults and nymphs suck plant juice by inserting their needle-like mouthparts into the cortex of trunks and branches. After being sucked by longan lanternflies, small black spots would gradually appear in the cortex [24]. When the number of P. candelaria is too large the harm caused by them might become serious, which can weaken the longan trees, stunt their growth, make branches dry, and result in fruit drop or poor fruit quality, and the excreta of this insect can even cause soot disease of longan tree [24]. And it’s found that, in a forest mixed planting of longan, litchi, olive and mango, the population of this insect is usually especially large [25].
Longan lanternflies have co-evolved with various fruit trees, microbial pathogens, and other environmental insects through long-term interaction, and may play an important role in the transmission of some microbial pathogens of fruit trees or from other agricultural important insects. With the advancement of sequencing technology, microbial research has also entered the era of omics, in which to screen and identify possible microbial pathogens is one of the most important parts. The study found a new virus in the course of screening microbial pathogens carried by P. candelaria through high-throughput RNA sequencing, and the virus was further characterized in order to lay a solid foundation for revealing the possible interaction of the virus and its hosts, and providing new insights to reveal the role of P. candelaria as a potential vectot for microbial pathogens.
2. Materials and Methods
2.1. Insect Sample Collection
P. candelaria samples were collected from the longan forest in Fujian Academy of Agricultural Sciences (FRIFAAS) and Minhou County as insect samples in this study.
2.2. RNA Sequencing (RNA-seq) and Data Assembly
In this study, the collected P. candelaria samples were frozen and transported to BGI (Beijing Genomics Institute) for RNA extraction and RNA-seq. In this study, samples were collected for deep RNA-seq. After the possible contaminated DNA was degraded by DNaseI, ribosomal RNA was removed from the total RNA with the kit of Ribo-ZeroTM, and all mRNA was retained to the greatest extent. NEB Fragmentation Buffer was mixed with the greatest extent of retained mRNA to cut the RNA into 250 to 300 bp short fragments. Then, the first strand of cDNA was synthesized with random hexamers as primers, and the second strand of cDNA was further synthesized by adding buffer, dNTPs, RNase H, and DNA polymerase I at Thermomixer, and purified by AMPure XP beads. A-Tailing Mix was added to repair the end of double-stranded cDNA and add A base at the end of 3’end of it. Then, the linker mix was added to connect the linker with cDNA, the joint product was purified and recovered with magnetic beads. The library was established by amplifying recovered cDNA in a PCR mix, and the concentration of the library was quantified by Qubit2.0 Fluorometer, and the library was diluted to 1.5 ng/µL. At the same time, the quality of the library was assayed by Agilent 2100 Bioanalyzer. The qualified library is denatured into a single chain by adding NaOH, and the denatured and diluted library was added into FlowCell, hybridized with the linker in the FlowCell, completed bridge PCR amplification in cBot, The sequencing platform was illumina novaseq6000 to generate 15.48 GB 150 bp paired-end reads In quality control, the software fastqc (version 0.11.8) was chosen to check the raw clean reads from RNA-seq; and then the quality of clean reads was checked by software fastp (version 0.20.1) by which low-quality sequence reads were removed again. Following quality control, the clean reads were assembled de novo by Trinity (version 2.15.1) with default assembly parameters and blasted in the database of virus (https://ftp.ncbi.nlm.nih.gov/refseq/release/viral/).
2.3. Rapid Amplification of cDNA Ends
RNA was extracted from P. candelaria samples according to the methods described in Materials and Methods 2.2, and cDNA was obtained by reverse transcription. The specific primers were used to identify whether the sample carried PyCaV virus, and the cDNA carrying the virus was stored at −20°C. Race double-end specific primers and nested primers were designed according to the sequence obtained by the second RNA-seq carried out in this study. PCR amplification was performed using cDNA as a template using 5'-RACE end-specific primers and reverse transcription primers, and the PCR products were detected by agarose gel electrophoresis to ensure specific amplification. The PCR product was purified and the purified cDNA chain was used as a template for subsequent experiments. The adaptor was ligated to the 5' end of the cDNA chain. After the ligation reaction was completed, the adaptor-specific primers and internal primers were used for nested PCR amplification. The first round of PCR amplification was performed using internal primers and adapter-specific primers. The first round of PCR product was used as a template, and the second round of PCR amplification was performed using nested primers. PCR products were detected by agarose gel electrophoresis to ensure specific amplification. The PCR product was transformed into E. coli Competent Cell JM109 vector, and the positive clones were selected for sequencing to obtain the 5' and 3' end information of the cDNA sequence. The 3'RACE amplification primers of the target gene were designed, and the obtained cDNA was used as a template for 3'RACE PCR amplification. The PCR product was purified and recovered by DNA, and the 3-terminal full-length sequence was obtained by DNA sequencing.
2.4. Bioinformatics Analysis
The conserved domain of the amino acid sequence was predicted by were predicted with the NCBI Conserved Domain Database (CDD, https://www. ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi). Phylogenetic analysis was performed based on the aligned conserved RdRp domain sequence from PyCaV and other members in family Iflaviridae, two members in Picornavirales were selected as outgroup. MAFFT (v7.310) [26] was used to perform multiple sequence alignment with default parameters, while the best substitution model was estimated by Prottest3 (version 3.4.2) [27]. A maximum likelihood phylogenetic tree was constructed using PhyML (version 3.2.0) with 1000 bootstrap replicates with the best estimated model [28].
2.5. RNA Extraction
In this study, the collected P. candelaria samples were subjected to independent RNA extraction after alcohol disinfection.The preparation of RNA was following the protocol previously used by Toni et.al. with minor modifications [29]. The insect samples were first disinfected with sodium hypochlorite and ground into powder by liquid nitrogen. After 1 mL Trizol reagent mixed with the powder for 30 min at 4°C, 200 μL pre-cooling dichloromethane was added, and mixed thoroughly for 2 min, then centrifuge at 4°C, 12000 r/min for 20 min. A portion of 450 μL supernatant was transferred into the new sterile RNase EP tube and mixed with another 200 μL dichloromethane, mixed repeatedly for 2 min, then centrifuge at 4°C at 12000 r/min for 10 min. Following 350 μL supernatant transferred into a new sterile RNase EP tube, the same amount of pre-cooling isopropanol was added and mixed, and kept under -20°C for 20 min, then centrifuged under 4°C at 12000 r/min for 20 min. The pellets were washed with 1 mL of pre-cooled 75% ethanol (DEPC water) twice, air dried and finally dissolved in 20 μL DEPC water. The quality of prepared RNA samples was monitored by electrophoresis and with Nanodrop 2000, and the qualified RNA samples (the ratio of OD260/OD280 reaches about 2.0) were stored at -80°C for further use.
2.6.cDNA Preparation
The cDNA was obtained by reverse transcription of total RNA using Revert Aid First Strand cDNA Synthesis Kit (Quan, Beijing). 5 μg RNA, 1 μL Oligo [30] 18 Prime, 2 μL RNase-free water supplement were mixed, and put in a 65°C water bath for 5 min, then transferred immediately into an ice bath for 2 min. After the ice bath, 10 μL R-Mix, 1 μL E-Mix, and 1 μL gDNA-Remover were added to the system and kept at 42°C for 30 min, then heat-shocked at 85°C for 5 s to inactivate the activity of the reverse transcriptase. Finally, the prepared cDNA was stored at -20°C.
2.7. DNA Purification
The PyCaV virus fragment was amplified by PCR with specific primers, and the DNA was purified and recovered, and the recovered DNA fragment was sequenced. After electrophoresis, Gene Gel Extraction Kit (Omega, Shanghai) gel recovery kit was used for the recovery of DNA products. The amplified DNA fragment was mixed with a 10×DNA loading buffer, and analyzed by electrophoresis with a 1% agarose gel at 200 V (180 mA) in TBE buffer for 20 min. After electrophoresis, the target band was cut out and transferred into a sterile EP tube and mixed with the same volume of Binding buffer, then the gel was melted in a 65°C water bath. The melted mixture was transferred to an adsorption column from the gel recovery kit, centrifuged at 10000 r/min for 1 min, and the filtrate was discarded. Following 300 μL Binding buffer was added to the adsorption column, and centrifuged at 12000 r/min for 1 min, 700 μL SPW wash buffer was added to the adsorption column, centrifuged at 14000 r/min for 1 min. The adsorption column was centrifuged at 15000 r/min for 2 more min, followed by adding 30 μL Elution buffer to the adsorption column, standing at room temperature for 2 min, then centrifuged at 15000 r/min for 2 min, and the DNA recovery product was collected. Finally, the concentration of DNA recovery product was measured by Nanodrop 2000 and stored at -20°C for further use.
2.8. Virus Verification
In this study, the following methods were used to identify whether the P. candelaria samples carried PyCaV virus. After surface disinfected by sodium hypochlorite, the longan lanternfly samples were ground into a powder with liquid nitrogen and lysed at 4℃ for 30 min with 1 mL Trizol reagent. Followed by added with 500 µL chloroform, and standing for 3 min, then, the samples were centrifuged at 12000 rpm for 1 min. After the supernatant was mixed with the same volume of isopropanol and kept still at -20℃ for 30 min, the mixture was centrifuged at 12000 rpm for 1 min. The supernatant was discarded, and the pellet was thoroughly washed with 20% ethanol prepared with DEPC treated water. After further centrifuged at 12000 rpm for 1 min, let the pellet stand for 10 min to air dry. Finally, the prepared total RNA was dissolved in 20 µL DEPC pre-treated water, and kept at -80℃ for further use. The total RNA was reverse transcribed into cDNA with the kit of TransScript®One-Step gDNA Removal and cDNA Synthesis SuperMix (Transgen Biotech, Beijing). The cDNA was used as a template and amplified with primers designed according to the sequence information obtained from RNA-seq, and the amplification results were finally verified by agarose gel electrophoresis, and sequenced by Fuzhou Sunya Biotechnology Co., Ltd.
2.9. Statistics Analysis and Model Construction
The samples of longan lanternflies were collected and classified according to the location and time, and they were collected from two locations: Fruit Research Institute of Fujian Academy of Agricultural Science (FRIFAAS) and Minhou County of Fuzhou City. The total sample number and number of samples carrying viruses at each time were counted, and the positive rate was calculated (Number of samples carrying viruses/The total sample number). The histogram was established by importing statistical data into the software GraphPad Prism 9.0, and a T-test was selected for significance analysis (*P < 0.05). The simulated model was constructed with Origin 2019 and choosing the method of the scatter plot, and selecting related curve trends according to the distribution pattern of obtained point. If Nonlinear Curve was chosen, and the Logistic model was selected for further fitting.
3. Results
3.1. The Virus Screening from Pyrops Candelaria by RNA-seq
To screen the microbial pathogens carried by the fruit tree pest-longan lanternflies, the RNA-seq analysis was carried out, a total of 51.61 million paired-end reads were generated and were than assembled de novo by the software Trinity (version 2.15.1), generating 111,7086 candidate contigs for further analysis. To investigate the sequences originated from viruses, the 372,784 contigs with length no less than 500 nt were used to search against the viral refseq proteins from NCBI through BLASTX and 7,658 contigs showed best hits against viral proteins with longer than 200 aa. Among thes candidate viral-derived contigs, only 1932 no redundant contigs from 293 kinds of viruses, with phage removed (Supplementary Table S1). Based on the classification information, viral contig from 61 families were detected in P. candelaria (Figure 1). Among them, about 217 kinds of viruses detected in P. candelaria belonged to dsDNA, 18 kinds belonged to ssDNA(+), 10 kinds belong to ssRNA (+/), and 10 belonged to ssRNA-RT
The longest hit length blasted with known virus was found to be a 10916 nucleotide (nt) contig belonging to ssRNA (+) virus, shared 37.27% identified with polyprotein of Nilaparvata lugens honeydew virus-3, a member belonging to family Iflaviridae (Supplementary Table S1). Another contig of 6920 nt shared 23.18% identity with RdRp of Culex phasma-like virus, a member belonging to family Phasmaviridae. The limited sequence identity between viral contig and other known viruses suggested that the presence of several new viruses carried by P. candelaria. According to the above results and considering the length and the identity of these two contigs, the longest contig (10,916 nt) of the viral ssRNA (+) was selected for further analysis in this study, and the potential ssRNA (+) virus from P. candelaria were tentatively anamed as PyCaV (Pyrops candelaria associated virus) in the following sections of this study.
3.2. The Full-Length Sequencing and Related Bioinformatics Analysis Reflect PyCaV a New Virus
In order to obtain the full length of PyCaV sequence, Rapid Amplification cDNA End (RACE) experiments were carried out based on the assemblied contig of 10916 nt and the primers used in this study accessed in supplementary Table S2. The sequence of the PyCaV was validated by sanger sequencing and the final full sequence of the virus was submitted to NCBI GenBank (ID: ON382046). With a polyA tail, the full-genome sequence of PyCaV virus is 10855 bp. Similar to other members in family Iflaviridae, PyCaV contains only a single predicted open reading frame (ORF), and the ORF encodes a 3171 aa polyprotein (Figure 2A). The polyprotein encoded by PyCaV virus can be cut into structural proteins and non-structural proteins. The conserved protein domains of PyCaV virus were predicted using the NCBI conserved domain database, including two rhv domains, one RNA helicase domain (HEL), one 3C cysteine protease domain (Pro) and one RNA-dependent RNA polymerase domain (RdRp) (Figure 2A). The results showed that PyCaV virus had a typical Iflavirus structure. When mapping the RNA-Seq reads from P. candelaria samples to PyCaV, a total of 12506 reads were found mapped to the reference genome (Figure 2B), suggesting the high abundance of PyCaV virus in P. candelaria.
3.3. Construction of PyCaV Phylogenetic Tree
In order to explore the phylogenetic relationship between PyCaV and other viral members in Iflaviridae, two members of the small RNA virus Rabovirus D1 (RVD) and Aimelvirus 1 (AV1) were used as outgroups. The phylogenetic tree was constructed using the conserved RdRp domain sequence of PyCaV virus and the sequence of represented members from the family Iflaviridae (Figure 2B). Phylogenetic analysis showed that PyCaV was closely related to Nilaparvata lugens honeydew virus 3 (NLHV3). Further sequence alignments with the structure protein of PyCaV and other represented members suggested limited sequence identity between PyCaV and other known members of Iflaviridae, while the structure protein of PyCaV shared the highest sequence identity (67.67%) with that of Nilaparvata lugens honeydew virus-3. Based on the genus demarcation criteria of family Iflaviridae from ICTV, the sequence identity at the amino acid level between the capsid proteins of PyCaV and other reported member is lowere than 90%, suggesting that PyCaV is a new unclassified member of family Iflaviridae carried by P. candelaria.
3.4. P. Candelaria Is One of the Important Carriers of the Virus PyCaV
In order to facilitate the subsequent identification of PyCaV in P. candelaria, we designed a pair of specific primers from 4646 nt contig to identify PyCaV (The primers: LYJ-H-F: 5’GGTCTATGCTGTATCCAAA 3’; LYJ-H-R: 5’ATATTGTCAAGCTGGTGAG 3’), and the length of the amplified DNA fragment by the diagnostic PCR is 405 bp (Supplementary Figure S1). To explore the distribution of the virus PyCaV in time, space, and inside the insect, the total RNA was extracted from the P. candelaria samples, and reverse transcribed into cDNA. With the cDNA as templates, the results of following diagnostic PCR showed that the virus PyCaV was detected in the two samples (Figure 3A, Lane 3 and 4) from the 8 randomly collected P. candelaria samples from FRIFAAS (Fruit Research Institute of Fujian Academy of Agricultural Science, the green spot in the Figure 3B) and Minhou County (the red spot in the Figure 3B). The amplified DNA fragment was further collected, purified and confirmed by DNA sequencing by Fuzhou Sunya Biotechnology Co., Ltd. (Supplementary Figure S2). In order to detect the distribution of the virus PyCaV in various parts of the insect, the cDNA was prepared from three parts of the P. candelaria: head, chest, and abdomen (Figure 3C, the panel above), and the diagnostic PCR result showed that the virus PyCaV can infect the whole body of the insect, including its head, chest, and abdomen, and there is more virus found in its chest (Figure 3D, Group 1: Lane 1 to 3). In order to study the distribution of the PyCaV virus inside its body, the piercing-sucking mouthparts (PSM), chest viscera (CV), and abdomen viscera (AV) are separated under dissecting microscope (Figure 3C, the panel below), and the RT-PCR result showed that most PyCaV viruses are located in its PSM and CV (Figure 3D, Group 2: Lane 1 to 3), and what interesting is that the main tissue in CV is salivary glands.
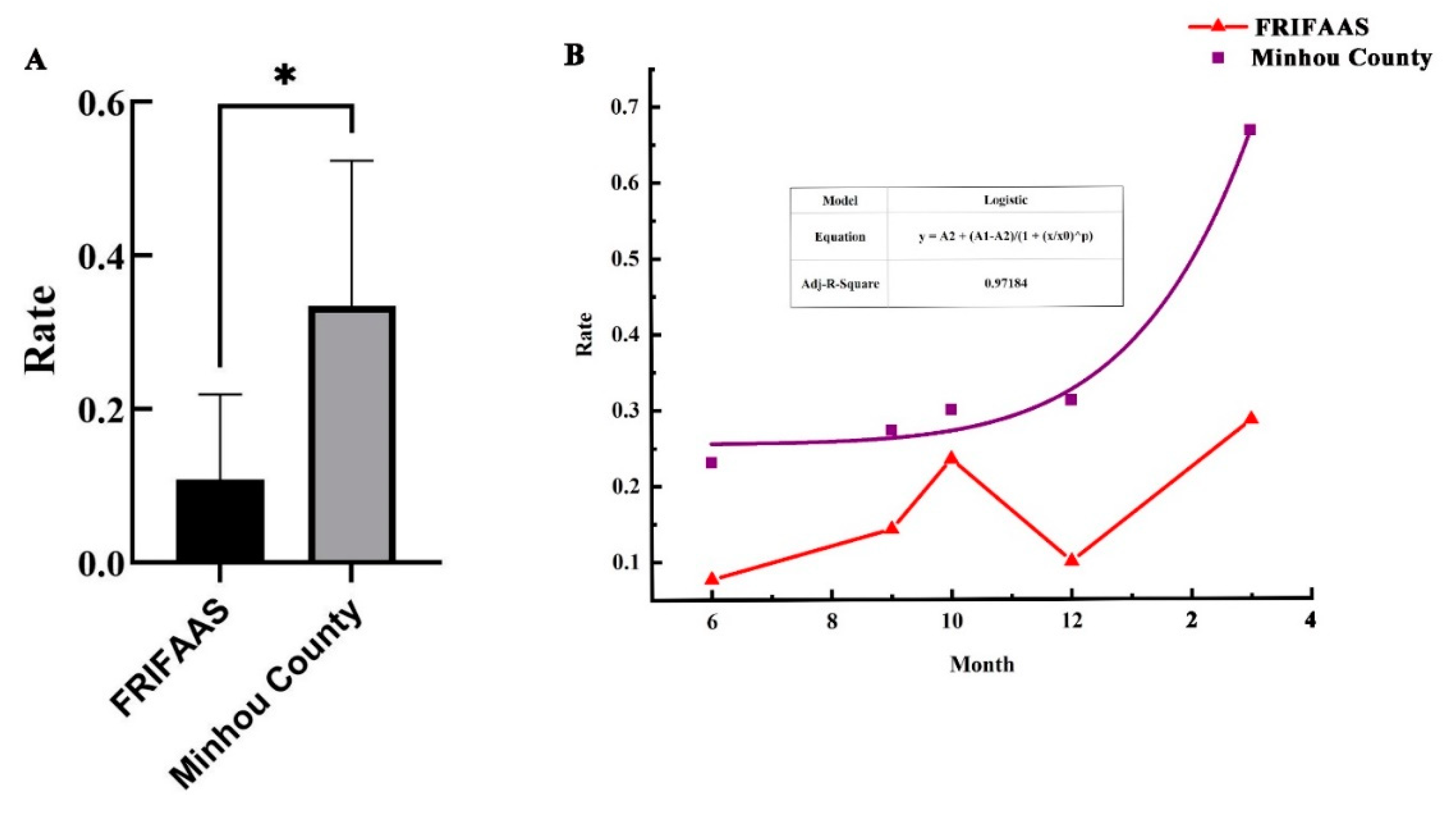
To explore if the location or distance could affect the distribution of the PyCaV, the PyCaV carrying rate between the P. candelaria samples from FRIFAAS and Minhou County was compared by One-way ANOVA, and the result reflects that there is a significant difference in the distribution of the virus between the two sampling sites (Figure 4A and Supplementary Table S4). To analyze the possible interaction of places and seasons of the PyCaV carrying rate, 123 longan lanternflies from these two places in different seasons were collected and it was found that 33 samples carried PyCaV. The changing trend of the PyCaV carrying rate was further simulated with Origin 2018, and the result showed the model of the PyCaV carrying rate of P. candelaria samples from Minhou County in different time point from spring to winter fitting to the Nonlinear Curve (Adj-r 2=0.97184), which reflected that from summer to winter and spring, the PyCaV carrying rate is stably increased (Figure 4B). But the carrying rate model of the samples from FRIFAAS are not fitting to the Nonlinear Curve or Generalized linear model, the fitted model showed that the changing trend of PyCaV is wavy, in which it reaches its peak in autumn and spring, and touches its valley at summer and winter (Figure 4B). These results reflected that the PyCaV carrying rates in these two sampling places are showing obviously different changing trends in the time dimension.
4. Discussion
According to the classification of ICTV, family Iflaviridae contains only one genus Iflavirus and several unassigned members [1]. It has been reported that most known Iflavirus viruses are isolated from arthropods, and the main infected host of the virus is insects. Iflavirus infection has different effects on hosts. Some hosts show no obvious symptoms after infection, but some hosts suffer abnormal development after the infection with Iflavirus, for example: silkworms, bees and many other invertebrates infected by corresponding Iflavirus virus can lead to fatal effect [1,31,32]. Honeybee (Apis mellifera), one of the common hosts of Iflavirus infection [33], are indispensable pollinators in ecosystems. [34]. However, in recent years, reports on the decline in honeybee pollination have been common, posing a serious threat to biodiversity and global food security [35,36,37]. McMahon D.P. et al. reported that deformed wing virus (DWV), one of the Iflavirus virus, can infect honeybees through Varroa and Tropilaelaps, resulting in a dramatic decrease in the number of honeybees [38]. When DWV infects honeybees, it can spread vertically within the bee population and increase the prevalence of the virus among honeybees [38]. In addition, Iflavirus can co-infect a host with multiple viruses and affect the growth and development of the host [39]. Two Iflavirus viruses were found in armyworm Spodoptera exigua and named as Spodoptera exigua iflavirus 1 (SeIV-1) [40] and Spodoptera exigua iflavirus 2 (SeIV-2) [41]. Experiments showed that S. exigua larvae infected with SeIV-1 and SeIV-2 increased the susceptibility of S. exigua to S. exigua multiple nucleopolyhedrovirus (SeMNPV) [17]. S. exigua infected with SeIV-1 and SeIV-2 viruses leads to a decrease in the weight gain of its larvae, and an imbalance in its sex ratio [42,43]. Thereby, Iflavirus has serious harm to some arthropods, especially insects. However, according to current reports, the species number of Iflavirus found is relatively small [43]. Therefore, finding potential members of Iflavirus and finding potential hosts of Iflavirus are of great research significance for biosafety.
Dimocarpus longan is an important subtropical fruit tree of family Sapindaceae, commonly known as longan in Asia [44]. According to published reports, longan fruits have a variety of functions, and their secondary metabolites have antioxidant, anti-cancer and other immunomodulatory activities [45,46,47]. In addition, the flowering period of longan is in spring and summer, which is an excellent nectar source plant. Pattamayutanon et al.found that honey derived from longan flowers contains more volatile organic compounds than all other flower sources [48].Therefore, longan is an important fruit crop. However, there are many pathogens that endanger the growth of longan, resulting in a serious reduction in its yield and quality. Trialeurodes vaporariorum, P. candelaria Deudorix dpijarbas Moore, and Cornegenapsylla sinica are the main pests that harm the growth and development of longan [49]. Tran et al. reported that the infection of C. sinica post a devastating effect on the growth of longan by spreading longan pathogen witches broom virus (LgWB) [50]. Therefore, it is very important to explore the potential microbial pathogens carry by main longan pests and their transmission mechanism. The P. candelaria is widely distributed in China, India, and many other countries in Southeast Asia and harms a variety of fruit trees [22]. It is also the main pest of longan. However, there are still few reports on the mechanism by which P. candelaria endanger longan as a microbial pathogen vehicle, herein, it is urgent to reveal the diversity of the microbial pathogens that this insect vector carries. In this study, a new virus PyCaV was identified in P. candelaria for the first time by RNA-seq. This is the first Iflavirus virus found in P. candelaria, which provided a new clue for the study of the pathogenicity of Iflavirus viral and cast new insight to the hazard of P. candelaria as a pathogenic vector of virus. However, the potential biological effect of PyCaV carrying by P. candelaria to longan or other insects remains to be further explored.
Through RNA-seq, based on the classification information, viral contig from 57 families and 4 unclassified groups were detected in P. candelaria (Figure 1A), which broadened our understanding of the virus that can be carried by P. candelaria. Apart from PyCaV, a near full length contig showed limited sequence identified with a member in Phasmaviridae, suggesting the high possibility of another virus carried by P. candelaria. Through RACE experiments and the construction of related phylogenetic trees, the similarity between PyCaV and Nilaparvata lugens honeydew virus 3 (NLHV3) was 67.67%, and it was distributed in the same cluster on the phylogenetic tree (Figure 2B). Based on the species demarcation criteria from ICTV, our results supported that PyCaV is a new member of family Iflaviridae carried by P. candelaria. Since Iflavirus only infects arthropods, especially insects, and as the largest group of animal, insect accounts for two-thirds of animal species [51]. In addition, insects occupy most of the ecological niches. More than 80% of flowering plants in the world are considered to be dependent on insect pollination, and about three-quarters of crops are also dependent on insect pollination [52,53]. Insects also become an important part of modern terrestrial ecosystems by regulating the carbon cycle in ecosystems [54,55]. Therefore, it is particularly important to further reveal the relationship between Iflavirus and insect hosts.
At present, new viruses account for almost 50% of emerging infectious diseases, which seriously threaten agricultural production, national biosafety, and even human survival [56]. Therefore, this study further explored the distribution of PyCaV virus in the insect. It is revealed that the PyCaV virus is distributed in every part of the P. candelaria, including the head, chest, and abdomen, which means that P. candelaria is, at least, one of the most important carriers of the PyCaV virus (Figure 3D, Group 1). The detection of the body section distributing of this virus reflected that more PyCaV virus accumulated in the chest of P. candelaria (Figure 3D, Group 1). The chest of the insect mainly including salivary glands and muscles, and when detecting the tissues inside the insect, it was found that the PyCaV virus mainly accumulated inside piercing-sucking mouthparts and viscera (mainly salivary glands) in the chest (Figure 3D, Group 2). The accumulation of the PyCaV virus in the piercing-sucking mouthparts and chest viscera inferred that the feeding system of the insect might play a critical role in the keeping and transmission of the virus. The salivary gland is one of the main tissues in psyllid (Diaphorina citri) that carries the DcFLV (Diaphorina citri flavi-like virus) [57]. Through the piercing-sucking mouthpart, the virions of PLRV (Potato leafroll virus) enter green peach aphid (Myzus persicae) and reach the salivary glands through the circulation in the hemolymph, and the virions are released into the saliva to contaminate other fruit plants [58]. And in the small brown planthopper (Laodelphax striatellus), a plasma membrane-associated protein, importin α, is utilized by the RSV (rice stripe virus) to enter the insect cell, especially the cells of salivary glands [59]. P. candelaria, psyllid, aphid, and the small brown planthopper all belong to Homoptera insects, so our results inferred that the PyCaV virus recovered in this study might follow the route of piercing-sucking mouthpart, the gut, the hemolymph, the salivary gland, and finally to be released into virus sensitive plants through saliva. But this speculation needs to be supported by further determining if it has any plant hosts.
In the study, in all 123 P. candelaria, 33 were found carrying the PyCaV virus, and the virus detection rate (VDR) is about 26%. Compared to the 6.93% detection rate of BtAstV (Bat-transmitted astroviruses), 1.79% detection rate of BtCoV (Coronaviruses), and 0.67% of BtCalV (novel caliciviruses) from bats [60], the VDR of the PyCaV virus is relatively high, especially in the autumn and spring in FRIFAAS, and the spring in Minhou County (Figure 4B), which means it is urgent to study its possible pathogenicity before it causes possible harm. To the different sampling regions, the VDRs varied significantly, too. The VDR is about 18% in FRIFAAS and 35% in Minhou County, which reflected the remarkable distribution difference of the PyCaV virus, which might reflect that the location or distance or orther factors which can hinder the migration and spread of the insect carrier might influence the transmission of this virus (Figures 3B and 4A, Supplementary Table S4). In spring, the VDR is about 46%; in summer, the VDR is about 15%; in autumn, it is about 24%; and in winter, it is about 23%. The VDR showed obvious seasonal variation, in which the VDR reaches its peak in the spring, touches its lowest valley in the summer, and rises in the autumn and winter (Figure 4B, Supplementary Table S4), and the VDR showed obvious regional variation (Figure 4A). Given more longan lanternflies carry the PyCaV virus in the spring, it is speculated that this season may be a high incidence period of PyCaV virus infection from this insects to other possible hosts.
In this study, the novel PyCaV virus carried by P. candelaria were found for the first time, its distribution in the pests and the changing trends of its carrying rates by P. candelaria in different time and space was revealed, which leads to an urgent need to explore whether this virus is harmful to longan tree or its pollinators. Totally, this study laid a new foundation for studying the role of P. candelaria as an insect vector in pathogen transmission, and expanded our understanding of Iflavirus virus.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Conceptualization, Z.Z. and J.X.; methodology, Z.Z.,H.L. and W.S..; software, J.X., R.M., and R.L.; validation,H.L, D.M.,C.Y.; formal analysis, H.L.,C.Y.; investigation, Z.Z., H.L., J.X., L.H., Y.Y., D.D and J.X.; resources, Z.Z., S.W., J.J. and R.L.; data curation, Z.Z.,Y.Y.; writing-original draft preparation, Z.Z., J.X., and H.L.; writing-review and editing, Z.Z.,J.X. and H.L.; visualization, Z.Z. and J.X.; supervision, Z.Z., and S.W.; project administration, Z.Z. and S.W.; funding acquisition, Z.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the National Key Research and Development plan project (2019YFC1200500, 2019YFC1200505).
Data Availability Statement
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding authors.
Acknowledgments
We thank Lifang Yu, Mengjuan Zhang, Xiaohua Pan, Wangzhuo Fu, and for their help in insect sampling, and thank Mengjuan Zhang for her help in experiments, and we are also very grateful to Dr. Shanshan Zhao for her professional advice on virus screening and sequencing.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Valles, S.M.; Chen, Y.; Firth, A.E.; Guérin, D.M.A.; Hashimoto, Y.; Herrero, S.; de Miranda, J.R.; Ryabov, E.; Consortium, I.R. ICTV Virus Taxonomy Profile: Iflaviridae. J. Gen. Virol. 2017, 98, 527–528. [Google Scholar] [CrossRef] [PubMed]
- Ongus, J.R.; Roode, E.C.; Pleij, C.W.A.; Vlak, J.M.; van Oers, M.M. The 5' non-translated region of Varroa destructor virus 1 (genus Iflavirus): Structure prediction and IRES activity in Lymantria dispar cells. J Gen Virol 2006, 87, 3397–3407. [Google Scholar] [CrossRef]
- Lee, Y.F.; Nomoto, A.; Detjen, B.M.; Wimmer, E. A protein covalently linked to poliovirus genome RNA. Proc Natl Acad Sci USA 1977, 74, 59–63. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.Y.; Wu, C.Y.; Chen, Y.J.; Chen, C.Y.; Wang, C.H. The 5' untranslated region of Perina nuda virus (PnV) possesses a strong internal translation activity in baculovirus-infected insect cells. Febs Lett 2007, 581, 3120–3126. [Google Scholar] [CrossRef] [PubMed]
- Roberts, L.O.; Groppelli, E. An atypical IRES within the 5' UTR of a dicistrovirus genome. Virus Res 2009, 139, 157–165. [Google Scholar] [CrossRef] [PubMed]
- Lanzi, G.; de Miranda, J.R.; Boniotti, M.B.; Cameron, C.E.; Lavazza, A.; Capucci, L.; Camazine, S.M.; Rossi, C. Molecular and biological characterization of deformed wing virus of honeybees (Apis mellifera L.). J Virol 2006, 80, 4998–5009. [Google Scholar] [CrossRef] [PubMed]
- Isawa, H.; Asano, S.; Sahara, K.; Iizuka, T.; Bando, H. Analysis of genetic information of an insect picorna-like virus, infectious flacherie virus of silkworm: Evidence for evolutionary relationships among insect, mammalian and plant picorna(-like) viruses. Arch Virol 1998, 143, 127–143. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.Y.; Lo, C.F.; Huang, C.J.; Yu, H.T.; Wang, C.H. The complete genome sequence of Perina nuda picorna-like virus, an insect-infecting RNA virus with a genome organization similar to that of the mammalian picornaviruses. Virology 2002, 294, 312–323. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhang, J.; Lu, J.; Yi, F.; Liu, C.; Hu, Y. Sequence analysis and genomic organization of a new insect picorna-like virus, Ectropis obliqua picorna-like virus, isolated from Ectropis obliqua. J Gen Virol 2004, 85, 1145–1151. [Google Scholar] [CrossRef]
- Ongus, J.R.; Peters, D.; Bonmatin, J.M.; Bengsch, E.; Vlak, J.M.; van Oers, M.M. Complete sequence of a picorna-like virus of the genus Iflavirus replicating in the mite Varroa destructor. J Gen Virol 2004, 85, 3747–3755. [Google Scholar] [CrossRef]
- Choe, S.E.; Nguyen, T.T.; Hyun, B.H.; Noh, J.H.; Lee, H.S.; Lee, C.H.; Kang, S.W. Genetic and phylogenetic analysis of South Korean sacbrood virus isolates from infected honey bees (Apis cerana). Vet Microbiol 2012, 157, 32–40. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Liu, S.; Bonning, B.C. Genome Sequence of a Novel Iflavirus from the Leafhopper Graminella nigrifrons. Genome Announc 2015, 3. [Google Scholar] [CrossRef]
- Wu, N.; Zhang, P.; Liu, W.; Cao, M.; Wang, X. Sequence analysis and genomic organization of a new insect iflavirus, Sogatella furcifera honeydew virus 1. Arch Virol 2018, 163, 2001–2003. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Liu, Y.; Liu, W.; Cao, M.; Wang, X. Full genome sequence of a novel iflavirus from the leafhopper Psammotettix alienus. Arch Virol 2019, 164, 309–311. [Google Scholar] [CrossRef] [PubMed]
- de Miranda, J.R.; Genersch, E. Deformed wing virus. J Invertebr Pathol 2010, 103 (Suppl. 1), S48–S61. [Google Scholar] [CrossRef] [PubMed]
- Ryabov, E.V.; Wood, G.R.; Fannon, J.M.; Moore, J.D.; Bull, J.C.; Chandler, D.; Mead, A.; Burroughs, N.; Evans, D.J. A virulent strain of deformed wing virus (DWV) of honeybees (Apis mellifera) prevails after Varroa destructor-mediated, or in vitro, transmission. PLoS Pathog 2014, 10, e1004230. [Google Scholar] [CrossRef] [PubMed]
- Carballo, A.; Williams, T.; Murillo, R.; Caballero, P. Iflavirus Covert Infection Increases Susceptibility to Nucleopolyhedrovirus Disease in Spodoptera exigua. Viruses 2020, 12, 509. [Google Scholar] [CrossRef] [PubMed]
- Yuan, H.; Xu, P.; Yang, X.; Graham, R.I.; Wilson, K.; Wu, K. Characterization of a novel member of genus Iflavirus in Helicoverpa armigera. J Invertebr Pathol 2017, 144, 65–73. [Google Scholar] [CrossRef]
- Bitondi, M.M.; Nascimento, A.M.; Cunha, A.D.; Guidugli, K.R.; Nunes, F.M.; Simoes, Z.L. Characterization and expression of the Hex 110 gene encoding a glutamine-rich hexamerin in the honey bee, Apis mellifera. Arch Insect Biochem Physiol 2006, 63, 57–72. [Google Scholar] [CrossRef]
- Park, C.; Kang, H.S.; Jeong, J.; Kang, I.; Choi, K.; Yoo, M.S.; Kim, Y.H.; Kang, S.W.; Lim, H.Y.; Yoon, B.S.; et al. In-situ Hybridization for the Detection of Sacbrood Virus in Infected Larvae of the Honey Bee (Apis cerana). J Comp Pathol 2016, 154, 258–262. [Google Scholar] [CrossRef]
- Lin, Y.S.; Liao, J.R.; Shiao, S.F.; Ko, C.C. Origin and Potential Expansion of the Invasive Longan Lanternfly, Pyrops candelaria (Hemiptera: Fulgoridae) in Taiwan. Biology 2021, 10, 678. [Google Scholar] [CrossRef]
- Hsu, M.H.; Yang, Y.L.; Wu, M.L.; Wang, L.J. Host Plants of the Immature Stages of the Invasive Longan Lanternfly, Pyrops candelaria (L.) (Hemiptera, Fulgoridae) in Taiwan. Insects 2021, 12, 1022. [Google Scholar] [CrossRef]
- Wu, Y.H.; Kamiyama, M.T.; Chung, C.C.; Tzeng, H.Y.; Hsieh, C.H.; Yang, C.S. Population Monitoring, Egg Parasitoids, and Genetic Structure of the Invasive Litchi Stink Bug, Tessaratoma papillosa in Taiwan. Insects 2020, 11, 690. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Huang, J.; Huang, B. Preliminary study on the biological characteristics of Fulgora candelaria (L.). J. East China Entomol. 2000, 61–65. [Google Scholar]
- Urban, J.M.; Leach, H. Biology and Management of the Spotted Lanternfly, Lycorma delicatula (Hemiptera: Fulgoridae), in the United States. Annu Rev Entomol 2023, 68, 151–167. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol Biol Evol 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. ProtTest 3: Fast selection of best-fit models of protein evolution. Bioinformatics 2011, 27, 1164–1165. [Google Scholar] [CrossRef]
- Guindon, S.; Dufayard, J.F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New algorithms and methods to estimate maximum-likelihood phylogenies: Assessing the performance of PhyML 3.0. Syst Biol 2010, 59, 307–321. [Google Scholar] [CrossRef]
- Toni, L.S.; Garcia, A.M.; Jeffrey, D.A.; Jiang, X.; Stauffer, B.L.; Miyamoto, S.D.; Sucharov, C.C. Optimization of phenol-chloroform RNA extraction. MethodsX 2018, 5, 599–608. [Google Scholar] [CrossRef] [PubMed]
- Affeldt, K.J.; Carrig, J.; Amare, M.; Keller, N.P. Global Survey of Canonical Aspergillus flavus G Protein-Coupled Receptors. Mbio 2014, 5. [Google Scholar] [CrossRef]
- Ryabov, E.V. Invertebrate RNA virus diversity from a taxonomic point of view. J Invertebr Pathol 2017, 147, 37–50. [Google Scholar] [CrossRef]
- Ribière, M.; Olivier, V.; Blanchard, P. Chronic bee paralysis: A disease and a virus like no other? J Invertebr Pathol 2010, 103 (Suppl. 1), S120–S131. [Google Scholar] [CrossRef]
- Jia, W.; Wang, F.; Li, J.; Chang, X.; Yang, Y.; Yao, H.; Bao, Y.; Song, Q.; Ye, G. A Novel Iflavirus Was Discovered in Green Rice Leafhopper Nephotettix cincticeps and Its Proliferation Was Inhibited by Infection of Rice Dwarf Virus. Front Microbiol 2020, 11, 621141. [Google Scholar] [CrossRef] [PubMed]
- Klein, A.M.; Vaissiere, B.E.; Cane, J.H.; Steffan-Dewenter, I.; Cunningham, S.A.; Kremen, C.; Tscharntke, T. Importance of pollinators in changing landscapes for world crops. Proc Biol Sci 2007, 274, 303–313. [Google Scholar] [CrossRef] [PubMed]
- Biesmeijer, J.C.; Roberts, S.P.; Reemer, M.; Ohlemuller, R.; Edwards, M.; Peeters, T.; Schaffers, A.P.; Potts, S.G.; Kleukers, R.; Thomas, C.D.; et al. Parallel declines in pollinators and insect-pollinated plants in Britain and the Netherlands. Science 2006, 313, 351–354. [Google Scholar] [CrossRef] [PubMed]
- Goulson, D.; Lye, G.C.; Darvill, B. Decline and conservation of bumble bees. Annu Rev Entomol 2008, 53, 191–208. [Google Scholar] [CrossRef] [PubMed]
- Potts, S.G.; Biesmeijer, J.C.; Kremen, C.; Neumann, P.; Schweiger, O.; Kunin, W.E. Global pollinator declines: Trends, impacts and drivers. Trends Ecol Evol 2010, 25, 345–353. [Google Scholar] [CrossRef] [PubMed]
- McMahon, D.P.; Furst, M.A.; Caspar, J.; Theodorou, P.; Brown, M.J.F.; Paxton, R.J. A sting in the spit: Widespread cross-infection of multiple RNA viruses across wild and managed bees. J Anim Ecol 2015, 84, 615–624. [Google Scholar] [CrossRef] [PubMed]
- Kemp, E.M.; Woodward, D.T.; Cory, J.S. Detection of single and mixed covert baculovirus infections in eastern spruce budworm, Choristoneura fumiferana populations. J Invertebr Pathol 2011, 107, 202–205. [Google Scholar] [CrossRef] [PubMed]
- Millan-Leiva, A.; Jakubowska, A.K.; Ferre, J.; Herrero, S. Genome sequence of SeIV-1, a novel virus from the Iflaviridae family infective to Spodoptera exigua. J Invertebr Pathol 2012, 109, 127–133. [Google Scholar] [CrossRef]
- Silva, L.A.; Ardisson-Araujo, D.M.; Tinoco, R.S.; Fernandes, O.A.; Melo, F.L.; Ribeiro, B.M. Complete genome sequence and structural characterization of a novel iflavirus isolated from Opsiphanes invirae (Lepidoptera: Nymphalidae). J Invertebr Pathol 2015, 130, 136–140. [Google Scholar] [CrossRef]
- Carballo, A.; Murillo, R.; Jakubowska, A.; Herrero, S.; Williams, T.; Caballero, P. Co-infection with iflaviruses influences the insecticidal properties of Spodoptera exigua multiple nucleopolyhedrovirus occlusion bodies: Implications for the production and biosecurity of baculovirus insecticides. PLoS ONE 2017, 12, e0177301. [Google Scholar] [CrossRef]
- Jakubowska, A.K.; Murillo, R.; Carballo, A.; Williams, T.; van Lent, J.W.; Caballero, P.; Herrero, S. Iflavirus increases its infectivity and physical stability in association with baculovirus. PeerJ 2016, 4, e1687. [Google Scholar] [CrossRef]
- Huang, S.; Han, D.; Wang, J.; Guo, D.; Li, J. Floral Induction of Longan (Dimocarpus longan) by Potassium Chlorate: Application, Mechanism, and Future Perspectives. Front Plant Sci 2021, 12, 670587. [Google Scholar] [CrossRef]
- Park, S.J.; Park, D.H.; Kim, D.H.; Lee, S.; Yoon, B.H.; Jung, W.Y.; Lee, K.T.; Cheong, J.H.; Ryu, J.H. The memory-enhancing effects of Euphoria longan fruit extract in mice. J Ethnopharmacol 2010, 128, 160–165. [Google Scholar] [CrossRef]
- Yang, L.; Fu, S.; Khan, M.A.; Zeng, W.; Fu, J. Molecular cloning and development of RAPD-SCAR markers for Dimocarpus longan variety authentication. Springerplus 2013, 2, 501. [Google Scholar] [CrossRef]
- Zhong, K.; Wang, Q.; He, Y.; He, X. Evaluation of radicals scavenging, immunity-modulatory and antitumor activities of longan polysaccharides with ultrasonic extraction on in S180 tumor mice models. Int J Biol Macromol 2010, 47, 356–360. [Google Scholar] [CrossRef] [PubMed]
- Pattamayutanon, P.; Angeli, S.; Thakeow, P.; Abraham, J.; Disayathanoowat, T.; Chantawannakul, P. Volatile organic compounds of Thai honeys produced from several floral sources by different honey bee species. PLoS ONE 2017, 12, e0172099. [Google Scholar] [CrossRef]
- Chen, Y.; Sun, J.; Lin, H.; Lin, M.; Lin, Y.; Wang, H.; Hung, Y.-C. Salicylic acid reduces the incidence of Phomopsis longanae Chi infection in harvested longan fruit by affecting the energy status and respiratory metabolism. Postharvest Biol. Technol. 2020, 160, 111035. [Google Scholar] [CrossRef]
- Seo, J.K.; Kim, M.K.; Kwak, H.R.; Kim, J.S.; Choi, H.S. Complete genome sequence of longan witches' broom-associated virus, a novel member of the family Potyviridae. Arch Virol 2017, 162, 2885–2889. [Google Scholar] [CrossRef]
- Behura, S.K. Insect phylogenomics. Insect Mol Biol 2015, 24, 403–411. [Google Scholar] [CrossRef]
- Klein, A.M.; Vaissière, B.E.; Cane, J.H.; Steffan-Dewenter, I.; Cunningham, S.A.; Kremen, C.; Tscharntke, T. Importance of pollinators in changing landscapes for world crops. Proc Biol Sci 2007, 274, 303–313. [Google Scholar] [CrossRef]
- Kremen, C. The value of pollinator species diversity. Science 2018, 359, 741–742. [Google Scholar] [CrossRef]
- Eggleton, P. The State of the World's Insects. Annu. Rev. Environ. Resour. 2020, 45, 61–82. [Google Scholar] [CrossRef]
- Crespo-Pérez, V.; Kazakou, E.; Roubik, D.W.; Cárdenas, R.E. The importance of insects on land and in water: A tropical view. Curr Opin Insect Sci 2020, 40, 31–38. [Google Scholar] [CrossRef]
- Anderson, P.K.; Cunningham, A.A.; Patel, N.G.; Morales, F.J.; Epstein, P.R.; Daszak, P. Emerging infectious diseases of plants: Pathogen pollution, climate change and agrotechnology drivers. Trends Ecol Evol 2004, 19, 535–544. [Google Scholar] [CrossRef]
- Rashidi, M.; Lin, C.Y.; Britt, K.; Batuman, O.; Al Rwahnih, M.; Achor, D.; Levy, A. Diaphorina citri flavi-like virus localization, transmission, and association with Candidatus Liberibacter asiaticus in its psyllid host. Virology 2022, 567, 47–56. [Google Scholar] [CrossRef]
- Patton, M.F.; Hansen, A.K.; Casteel, C.L. Potato leafroll virus reduces Buchnera aphidocola titer and alters vector transcriptome responses. Sci. Rep. 2021, 11, 23931. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Lu, H.; Wang, W.; Zhu, J.; Zhao, W.; Cui, F. Membrane association of importin α facilitates viral entry into salivary gland cells of vector insects. Proc. Natl. Acad. Sci. United States Am. 2021, 118. [Google Scholar] [CrossRef] [PubMed]
- Kemenesi, G.; Dallos, B.; Görföl, T.; Boldogh, S.; Estók, P.; Kurucz, K.; Kutas, A.; Földes, F.; Oldal, M.; Németh, V.; et al. Molecular survey of RNA viruses in Hungarian bats: Discovering novel astroviruses, coronaviruses, and caliciviruses. Vector Borne Zoonotic Dis 2014, 14, 846–855. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Taxonomic distribution of virus detected from P. candelaria. the total of 1932 viral contigs derived from 293 kinds of viruses were classified into 61 families based on ICTV and NCBI. The horizontal bar represents the number of viruses, while the color of bar representated the type of viruses in the correspoding family.
Figure 1.
Taxonomic distribution of virus detected from P. candelaria. the total of 1932 viral contigs derived from 293 kinds of viruses were classified into 61 families based on ICTV and NCBI. The horizontal bar represents the number of viruses, while the color of bar representated the type of viruses in the correspoding family.

Figure 2.
Schematic diagram of the genome structure of PyCaV (Pyrops candelaria associated virus). (A) The genome encodes a 3171 aa polyprotein with several conserved domains, including two rhv domains, a RNA helicase domain (HEL), a 3C cysteine protease (Pro) and a RNA-dependent RNA polymerase (RdRp). Numbers on the bottom indicate the start and end positions of the polyprotein coding region in the genome.(B) Maximum-likelihood phylogenetic tree based on RdRp of PyCaV and other Iflavirus viruses. Two members, Rabovirus D1 (RVD) and Aimelvirus 1 (AV1), belonging to family Picornaviridae were used as the out group. Bootstrap values (>60%) were shown at each node of the tree. Bar represents the genetic distance. Virus name abbreviations was listed in the Supplementary Table S3.
Figure 2.
Schematic diagram of the genome structure of PyCaV (Pyrops candelaria associated virus). (A) The genome encodes a 3171 aa polyprotein with several conserved domains, including two rhv domains, a RNA helicase domain (HEL), a 3C cysteine protease (Pro) and a RNA-dependent RNA polymerase (RdRp). Numbers on the bottom indicate the start and end positions of the polyprotein coding region in the genome.(B) Maximum-likelihood phylogenetic tree based on RdRp of PyCaV and other Iflavirus viruses. Two members, Rabovirus D1 (RVD) and Aimelvirus 1 (AV1), belonging to family Picornaviridae were used as the out group. Bootstrap values (>60%) were shown at each node of the tree. Bar represents the genetic distance. Virus name abbreviations was listed in the Supplementary Table S3.

Figure 3.
The detection of the virus in longan lanternflies. (A) The PyCaV was detected from the random P. candelaria samples. The lower panel was the amplification of actin used as the inner reference. (B)The map showing the sample collecting locations. The red spot is where the samples are collected in Minhou County and the green spot is the position of FRIFAAS. (C) The separated tissues. Above are the samples from the head, chest, and belly of longan lanternflies; below are corresponding piercing-sucking mouthparts from heads, viscera in the chest, and viscera in the abdomen. (D) The PyCaV was detected from different parts and tissues of the P. candelaria samples. Above panel, Group 1: The separated head (Lane 1), chest (Lane 2), and abdomen (Lane 3) samples of longan lanternflies; Group 2: piercing-sucking mouthparts (Lane 1), viscera in the chest including salivary glands (Lane 2), and viscera in the abdomen including gonad (Lane 3). The lower panel: amplification of actin as the inner reference.
Figure 3.
The detection of the virus in longan lanternflies. (A) The PyCaV was detected from the random P. candelaria samples. The lower panel was the amplification of actin used as the inner reference. (B)The map showing the sample collecting locations. The red spot is where the samples are collected in Minhou County and the green spot is the position of FRIFAAS. (C) The separated tissues. Above are the samples from the head, chest, and belly of longan lanternflies; below are corresponding piercing-sucking mouthparts from heads, viscera in the chest, and viscera in the abdomen. (D) The PyCaV was detected from different parts and tissues of the P. candelaria samples. Above panel, Group 1: The separated head (Lane 1), chest (Lane 2), and abdomen (Lane 3) samples of longan lanternflies; Group 2: piercing-sucking mouthparts (Lane 1), viscera in the chest including salivary glands (Lane 2), and viscera in the abdomen including gonad (Lane 3). The lower panel: amplification of actin as the inner reference.
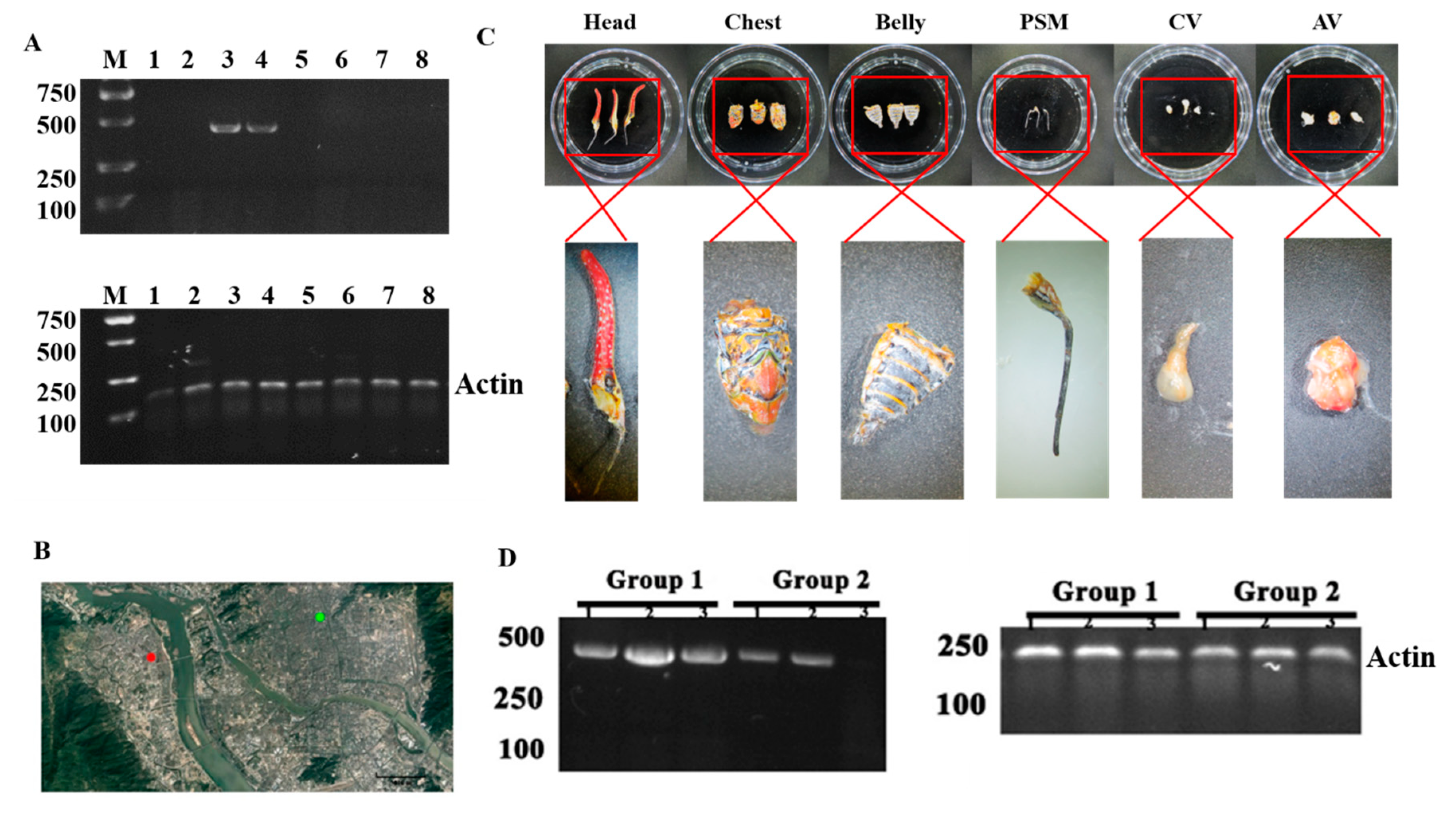
Figure 4.
The analysis of changing trend of the PyCaV carrying rate in different sampling sites and seasons. (A) The statistical analysis of PyCaV carrying rate of P. candelaria samples from FRIFAAS and Minhou County. (B) The simulation of the changing trend of the PyCaV carrying rate in both FRIFAAS and Minhou County with Origin 2018. Purple squares and lines are the simulated models for samples collected from Minhou County, and red triangles and lines are the models for the samples from FRIFAAS.
Figure 4.
The analysis of changing trend of the PyCaV carrying rate in different sampling sites and seasons. (A) The statistical analysis of PyCaV carrying rate of P. candelaria samples from FRIFAAS and Minhou County. (B) The simulation of the changing trend of the PyCaV carrying rate in both FRIFAAS and Minhou County with Origin 2018. Purple squares and lines are the simulated models for samples collected from Minhou County, and red triangles and lines are the models for the samples from FRIFAAS.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



