Submitted:
16 June 2024
Posted:
17 June 2024
You are already at the latest version
Abstract
In our previous papers, we demonstrated that inflammation and aging are the common roots of cancer and Alzheimer’s disease (AD). In both diseases, there is an inhibition of the mitochondria. In cancer, the alkaline intracellular pH (pHi) is associated with cell proliferation; acidic pHi causes apoptosis in AD. This difference is because cancer feeds on chemically neutral glucose, while neurons feed on acidic lactic acid. Increased uptake of lactic acid is responsible for acidic pH and cell death. Similarly, to AD, inflammation is responsible for metabolic shifts in age-related macular degeneration (AMD). In AD, there is hyperosmolarity responsible for increased lactic secretion by glial cells. Increased lactic secretion will cause neo-angiogenesis and neural cell death. Drugs increasing hyperosmolarity (Polyethylene Glycol) cause neo-angiogenesis and drusen-like structures in rodents. The link between AMD and inflammation is reinforced by the fact that treatments aiming at restoring mitochondrial activity, such as lipoic acid and/or methylene blue, have been experimentally shown to be effective in each set of diseases. The role of osmolarity has been demonstrated in the pathologies of the anterior segment of the eye, such as dry eye, corneal ulceration, cataracts, and glaucoma. The role of increased osmolarity in age-related macular degeneration (AMD) has been largely overlooked. We herein suggest that metabolic shift, inflammation, and hyperosmolarity are key players in the pathogenesis of AMD.
Keywords:
Macular degeneration
; osmotic pressure
; pHi
; apoptosis
; mitochondria
; Lipoic acid
; Methylene blue
1. Introduction
The past few decades have seen limited successes in the fight against age-related macular degeneration (AMD), cancer, and Alzheimer’s disease (AD). In each set of diseases, the paradigm has been centered on genomic alterations. Cancer and AD, or AMD, are widely considered to be different sets of diseases with different prognoses, sites of origin, patterns of spread, and treatments. Cancer, AD, or AMD can be seen at both opposite ends of a biological pattern. In cancer, there is unrelenting cell division. In AD or AMD, there is prominent apoptotic cell death. These diseases are different entities but share multiple common epidemiological and biological features [1,2,3]. Cancer and Alzheimer’s disease can be considered metabolic diseases [4,5]. Similarly, AMD can be seen as a disease with features common to cancer and AD [6,7].
Our work aims to highlight the underlying unity of these diseases and that metabolic shifts and increased osmotic pressure are also pivotal in the development of AMD. Increased osmotic pressure is well known to cause inflammation [8] and play a key role in cancer and Alzheimer’s disease [9].
We hypothesize that increased osmotic pressure because of inflammation of the posterior segment results in the increased secretion of lactic acid -by the Muller glial cells- which will feed the retinal cells. The retina cannot metabolize this lactic acid, resulting in acidic intracellular pH, synaptic dysfunction, extracellular deposition (Drusen), and ultimately apoptosis.
2. Overview
Age-related macular degeneration (AMD) is an eye condition strongly associated with aging. It can result in significant vision loss and blindness [10,11,12]. Age-related macular degeneration (AMD) is a chronic eye illness that mostly affects individuals over 50, while it can sometimes strike younger people in industrialized countries [13,14].
AMD is a retinal disease that typically affects the macula and gradually impairs vision in the central region of the field of vision. The primary clinical symptoms of early-stage AMD include drusen and changes to the retinal pigment epithelium [15].
Four stages can be distinguished in AMD [16,17,18]. The initial phase consists of typical aging changes with no aberrant pigmentation and just a little drusen. At this point, the drusen's diameter is less than 63 mm. While some intermediate drusen with diameters of 63 and 124 mm are in the second stage of AMD, no abnormalities are related to the RPE cells. The third stage is marked by RPE anomalies, at least one big drusen (diameter 125 mm; intermediate AMD), extensiveness, and moderateness. Advanced AMD, or stage 4, is characterized by geographic atrophy (GA) of the fovea or other age-related features of neovascular macular degeneration (nAMD) that lead to vision loss [15,19].
The non-dividing human retinal pigment epithelium (RPE) is a cell that performs several vital tasks for the survival of photoreceptor cells. As aging progresses, the RPE experiences several alterations that eventually result in the formation of drusen, a clinically noticeable localized yellow deposit of extracellular, polymorphous material at the interface between RPE and the inner collagenous zone of Bruch's membrane. The primary indicator of AMD is the presence of drusen inside the macula. In contrast, people with tiny drusen who do not have any other ocular abnormalities have a lower chance of developing the disease's more severe manifestations [11,17].
AMD patients exhibit a highly varied clinical appearance associated with drusen features and pigmentary abnormalities, including hypo- and hyperpigmentation. Numerous diseases can lead to geographic atrophy (GA) or choroidal neovascularization (CNV) AMD, often known as dry or wet AMD, respectively. These pathologies include focal detachment of the RPE, outer retinal atrophy, and new blood vessel growth between Bruch's membrane and the retina [11].
AMD is typically categorized based on the size of the drusen in clinically applied categorization methods that evaluate the severity of the non-sight-threatening early phases of this illness. Large drusen are classed as "intermediate" AMD and medium-sized drusen are regarded as "early" AMD. The likelihood of experiencing geographic atrophy (GA) and/or neovascular AMD, is highest when drusen are vast in area and accompanied by pigmentary changes [20].
3. Epidemiology
It has been estimated that 1.75 million Americans aged 40 and above have AMD, making the overall prevalence of neovascular AMD and geographic atrophy 1.47%. With more than 15% of White women over 80 years old having neovascular AMD and/or geographic atrophy, the prevalence of AMD rose with age. Over 7 million people were at significant risk of getting AMD because they had drusen that measured 125 microns or greater. The aging population is expected to lead to a 50% increase in AMD cases, reaching 2.95 million by 2020. White people had a considerably higher prevalence of age-related macular degeneration than did Black people [21].
The highest disease burden is seen in non-Hispanic White Europeans; however, the prevalence of AMD varies significantly by ethnicity. In a recent study, Wong et al. computed the pooled prevalence of population-based studies of AD across ethnically varied age groups (45–85 years) and found that the highest prevalence was found among people of European heritage, ranging from 12.3 to 30% as age increased [22]. The disease burden among Asians (7.4%), Africans (7.5%), and Hispanics (10.4%) is very high, despite a minor decrease (14). Nevertheless, the illness burden in the US has been estimated by some researchers to be lower, with non-Hispanic White Europeans having the highest prevalence at roughly 7.3% [15,23].
The incidence of age-adjusted dry AMD ranged from 0.3% to 0.4% in a population-based cohort research that integrated data from three population-based cohort studies (mean age: 60.1-65.7 years), with follow-up times ranging from 4.8 to 6.5 years. A higher incidence was associated with late-stage AMD disease. The incidence of dry AMD was also correlated with baseline drusen status in a North American study (N = 3,549). Patients with large bilateral drusen at baseline (n = 241) had a 10-year incidence risk of dry AMD diagnosis of 9.9%; patients with large bilateral drusen and RPE changes at baseline (n = 44.3%); and patients with late AMD (dry or wet) in one eye at baseline (n = 390) had a 10-year incidence risk of 53.9% [15,24].
Approximately 20 million Americans and 196 million people worldwide suffer from age-related macular degeneration. AMD is a major contributor to severe visual impairment in the elderly and is predicted to impact 288 million individuals globally by 2040. Furthermore, AMD affects 0.3 out of every 1000 persons in the 55 to 59 age group and 36.7 out of every 1000 people in the 90+ age group each year. It is estimated that 71% of late-stage AMD cases are heritable [25].
4. Risk Factors
Numerous factors, both modifiable and non-modifiable, have been associated with an elevated risk of AMD development. Aging, ethnicity, and genetics are some of the multiple risk factors contributing to advanced AMD development [11,26].
Extracellular debris deposits known as drusen are seen in the space between Bruch's membrane (BM) and the retinal pigment epithelium (RPE). The primary indicator of AMD is the presence of drusen inside the macula. The number of drusen overall, as well as the area or volume of drusen assessed, are risk factors for the development of GA and nAMD [27,28,29]. Additionally, the placement of the drusen has prognostic significance for the course of the disease; that is, eyes that have drusen close to the fovea are more likely to develop late AMD than eyes that have drusen farther from the fovea [30,31,32]. According to a recent study, the development of drusen may initiate an inflammatory cascade that contributes to the advancement of AMD [26,33].
It has been demonstrated that 34 genetic loci, comprising 52 gene variations, are connected to AMD [34]. Numerous combinations of gene variations have been found in many chromosomes, including chromosomes 1, 6, and 10 [35,36], using genome-wide screening techniques. These responsible genes regulate immune response, inflammatory processes, and retinal homeostasis. Variations in these loci are thought to be responsible for the degree of dysfunction of these responses in AMD patients. High-temperature requirement serine peptidase 1 (HTRA1; also known as age-related maculopathy [ARM] susceptibility 2 [ARMS2]) on chromosome 10 at 10q26, CFB/C2 on chromosome 6 at 6p21.3. and CFH on chromosome 1 at 1q31.3 are the most studied potential genes [11,37,38]. Genetic predisposition has a greater influence than just an individual's increased risk of getting AMD; it can also impact how well an individual respond to treatment. A combination of high-risk alleles in CFH, ARMS2, and VEGFA was shown by Smailhodzic et al.[39] to be linked to an earlier age of onset and a poor response to intravitreal anti-vascular endothelial growth factor (anti-VEGF) therapies in AMD patients. Additionally, variations of the CFH gene polymorphism T1277C were linked to a limited morphological response and a delayed functional response to the initial intravitreal injection of Avastin (bevacizumab) in patients with homozygous CC group, as reported by Medina et al.[26,40,41].
Age is by far the most significant demographic risk factor for AMD [26,30,31,32] According to a recent meta-analysis that combined data from 14 population-based studies, the prevalence of early AMD increased from 3.5% in individuals 55–59 years old to 17.6% in those 85 and older. These prevalence rates rise from 0.1% to 9.8%, respectively, for late AMD. [10,26]. Age is marginally more strongly related to the development of geographic atrophy (GA) compared to neovascular AMD (nAMD)[42,43,44]. Furthermore, While AMD is rare in individuals under 50, patients over 75 have a threefold higher risk of developing the condition than those between 65 and 74 years of age. In the United States, 30% of those over 85 have AMD [11].
The primary significant modifiable risk factor is smoking. AMD is two to four times more common in smokers over 40 than in nonsmokers of the same age [41]. Indeed, smoking has been the most reliable variable linked to neovascular AMD and geographic atrophy. A meta-analysis of five case-control studies, five cross-sectional studies, and six prospective cohort studies has also calculated the risk of smoking. Current smokers exhibited a significantly higher risk of AMD compared to never-smokers [45].
There are still contradictory results on the association of sex with predisposition and progression of AMD. While both men and women can get AMD, some research indicates that females have an increased likelihood of developing early AMD,[43,46] and late AMD [26,47] particularly nAMD [32,48]. Other research, however, indicates that there is no association between sex and the advancement of the illness [26,31,42,49].
Diet, lifestyle, and nutrition play important roles in AMD. The systolic or diastolic blood pressure or the use of antihypertensive drugs was not substantially connected with an increased risk of either early- or late-stage AMD. The French ALIENOR research found that high pulse pressure was linked to an increased risk of late-stage AMD [50]. While monounsaturated fats may be beneficial (30), a high diet of certain lipids, such as trans fats, saturated fats, and omega-6 fatty acids, has been linked to a twofold rise in the prevalence of AMD [26,51].
5. Inflammation as the Driving Force in Cancer, Alzheimer's, and Aging
In cancer, AD, and AMD, there is a small proportion of patients who have inheritable genetically transmissible risk factors [52,53]. Multiple genes are known to be implicated in cancer. Compared with the much more common sporadic cancer, these tumors arise most commonly before the age of fifty [54].
There are also cases of familial AD of autosomal-dominant inheritance, which usually has an onset at an early age [55]. Most autosomal-dominant familial AD can be attributed to mutations in one of three genes: those encoding amyloid precursor protein (APP) and presenilin 1 and 2 [56,57]. The vast majority of cases of AD do not exhibit autosomal-dominant inheritance and are thus termed sporadic AD. Similarly, a small proportion of AMD appears to have a genetic component [58]. These genetic features are rare in contrast to the most common sporadic ones. The vast majority of cases of AMD are sporadic. AMD, like cancer and Alzheimer’s disease, is strongly age-related [35].
In 1863, It was hypothesized that the origin of cancer occurs at sites of chronic inflammation [59]. A demonstration of the carcinogenicity of inflammation is foreign-body carcinogenesis [60]. The physical characteristics of the implant, rather than the chemical composition, are the critical determinants of tumor development. Abrasive, but not smooth, implants are both inflammatory and carcinogenic [60]. Inflammation is also a risk factor for AD [61]. Brain inflammation induced by repeated trauma increases brain levels of proteins associated with neurodegeneration such as amyloid β 1–42, observed in AD, total tau, and α-synuclein observed in Parkinson's disease (PD) [62].
Elderly people express low-level systemic inflammation [63]. Systemic inflammation plays a key role in the diseases of aging [63]. Inflammation, as seen in the process of aging, is a determinant in most of the diseases of the elderly [64]. The role of inflammation in AMD has been partially overlooked and is characterized by complement activation [65]. There are scanty reports about its importance but no clear explanation about its central role in this disease [24]. Another clue to the link between AD and AMD is the presence of beta-amyloid in the brain [66].
6. Inflammation and Increased Osmolarity
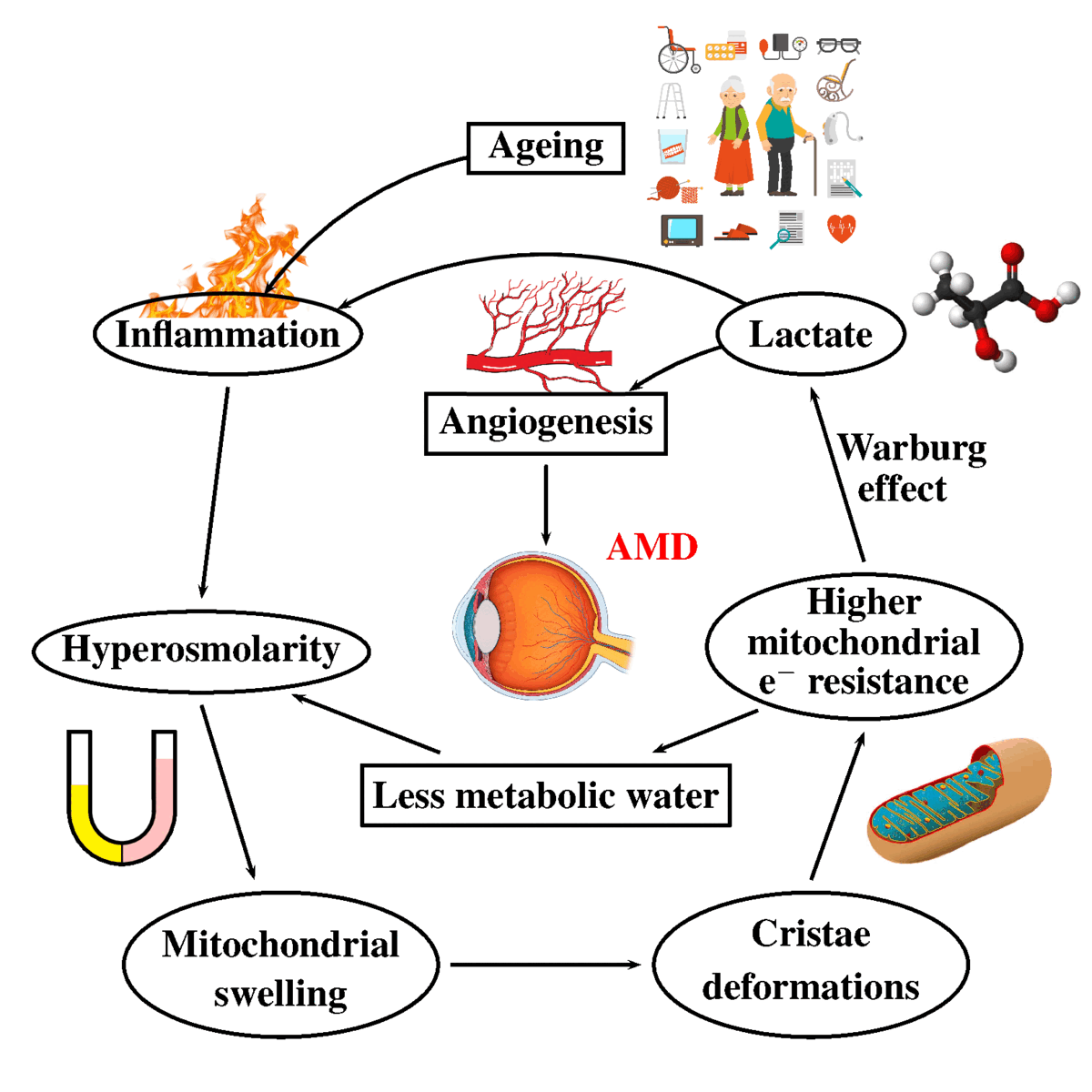
Inflammation is a clinical feature that can be caused by factors as diverse as heat, freezing temperature, trauma, or multiple chemicals resulting in vascular leakage [67,68]. Inflammation (a clinical feature) is closely related, if not synonymous, to hyperosmolarity (a physical feature) [8,69]. Animal models of inflammation demonstrate that in inflammatory fluids, whatever the cause, there is protein content, resulting in increased osmolarity. Increased osmolarity, whatever its cause, results in inflammation. Increased extracellular osmolarity stimulates cytokine synthesis and secretion and results in the proliferation and activation of immune cells [8].
7. Osmosis in the Posterior Segment
The posterior segment of the eye has one of the highest metabolic rates, requiring a rich supply of oxygen and other nutrients [74]. While the central retinal artery provides the inner retina's blood supply, most of its oxygen demand (seven times more important than that of the brain at constant weight) is supplied by diffusion from the underlying choroid, which is the sole supply of the avascular fovea. The choroid has the highest rate of blood flow per weight of any tissue [75]. The choroid, because of its intense blood flow and the permeability of the Bruch’s membrane, has a major role in the thermoregulation of the macula. The innermost layer of the choroid is the Bruch’s membrane, a 2-4μm thick elastic sheet. At a young age, it is permeable to liquids, ions, and small proteins [76].
There is evidence of a dialysis-like phenomenon in the eye. Like in an artificial kidney cartridge, where there is the concomitant circulation of arterial blood and dialysate, the choroidal and the retinal fluid circulation are separated by the Bruch’s membrane. The Bruch’s membrane is semi-permeable and this membrane is formed of glycated proteins such as heparin. The flow of the choroidal arteries is about 10 to 20 times more important than the flow of the retinal arteries [75,77], increasing the hydrostatic pressure in the retina. The two compartments of dialysis in the eye are the retinal pigment epithelium (RPE) and outer segments of the visual cells (OS) as the dialyzed sector animated by a centripetal flow, and secondary the choroid, which may be equated with a dialyzing bath driven by a centrifugal rather than counter flow. That is, to increase the exchange’s surface and to maintain a gradient, reinforce efficiency [78].
8. Aging of the Posterior Segment: Vascular Changes
During aging, there is a clear decrease in the perfusion of the choroid. The normal thickness of the choroid, measured in younger emmetropes (<27 years old) by EDI-OCT, varies from 264 to 436µ [79]. It decreases by 15.6µ per decade[80,81]. The Bruch membrane thickens, further altering exchanges [76]. The choroidal blood flow decreases, as does the porosity of the Bruch membrane. The decreased perfusion will result in ischemia and, therefore, the impairment of retinal pigment epithelium's (RPE) hemodynamic cellular antioxidant properties. The retinal pigment epithelium RPEis is deficient in managing such increased oxidative stress either by the apical synthesis of Reticular Pseudo Drusen (RPD), also named Subretinal Drusenoid Deposit (SDD) or by the more common basal exocytosis of multiple drusen composed of lipofuscin and bis retinoids [65].
The retinal pigment epithelium (RPE) separates the retinal (apical) and choroidal (basal) environments and contributes to a blood-retinal barrier (BRB), which provides a proper environment for photoreceptor cells. The osmolality on the choroidal side is higher than on the retinal side in physiological conditions [82]. Thus, the osmotic gradient from the apical to basal sides is thought to elevate transepithelial electric resistance (TER). TER reflects ion permeability across the epithelia [83]. So, gradients illustrate the direction of epithelia function or pathological changes according to apical and basal osmotic conditions through hydrostatic pressure (HP) [82]. The accumulation of these multiple deposits and wastes of various molecules will also result in a significant increase in osmolarity [8].
9. Increased Osmolarity and AMD
To the best of our knowledge, osmolarity has not been measured in any pathology of the posterior segment, but there is indirect and direct evidence of its importance in AMD. AMD is characterized by multiple features, such as infiltration of the retina by inflammatory cells, proliferation of fibroblasts, and new blood vessel formation. In the neovascular subfoveal membranes, the proliferation of fibroblasts goes along as a conjunctival tissue wearing the new vessel network [84].
The secretion of vascular growth factors that leads to wet macular degeneration by the retinal human pigment cells is increased by osmolarity [85,86]. Retinal detachment can be induced by the intravitreal injection of a hyperosmotic solution [87]. Fibroblast proliferation could be a consequence of increased osmolarity [88].
The best demonstration of the key role of osmolarity in AMD comes from animal models [89]. The retina can be damaged by either blue light or a laser. This results in vascular damage and, in turn, increased osmolarity because of an extravascular protein leak. Polyethylene Glycol (PEG) is a chemically inert but osmotically active chemical. PEG is not metabolized in vivo [90]. The subretinal injection of PEG results in choroidal neovascularization [90] and the formation of structures resembling drusen [91].
10. Metabolic Shift Because of Increased Pressure
To perform their normal physiological functions, cells must maintain the intracellular pH (pHi) within the physiological range. Intracellular enzyme activity, cytoskeleton component integration, and cellular growth and differentiation rates are all closely associated with pHi [92]. A fall in pHi decreases neuronal activity and is responsible for apoptosis [93].
In AD, there is a shift toward intracellular acidosis and apoptosis. It has been demonstrated that neurons feed on lactate secreted by glial cells [9]. In the case of AD, there is increased secretion of lactic acid as measured by spinal fluid [94].
he increased secretion of lactate by glial cells results in increased uptake by neurons and intracellular acidosis [9]. This is the inverse of Warburg’s effect, as first described by [95]. The fall in pHi will result in neuronal cell death.
A similar scenario is probably at stake in macular degeneration. Neurons feed on lactate released by glial cells, and similarly, retinal cells feed on lactate released by Muller glial cells [96]. Müller cells, akin to glial cells in the brain, metabolize glucose to lactate, which is preferentially taken up by photoreceptors as a fuel for their oxidative metabolism [97]. Even in the presence of glucose and oxygen, cultured human Müller cells obtain most of their ATP, principally from glycolysis, and display a low rate of oxygen consumption [98]. They feed lactate to the retinal cells. Indeed, the increased secretion of lactic acid will have multiple consequences: increased urinary secretion of lactate [99], increased secretion of markers of inflammation [100], and VEGF [101]. The acidic pH plays a crucial role in retinal cell death [102].
The increased secretion of lactic acid is a consequence of inflammation. In inflammation, whatever its cause, there is increased secretion of lactate, resulting in increased serum and urine levels of lactate [103,104]. It has been shown that in response to osmotic stress, cells display acute metabolic remodeling through the control of pyruvate dehydrogenase phosphorylation through direct osmosensing in mitochondria [105]. The correlations between inflammation, hyperosmolarity, and metabolic changes in AMD are shown in Figure 1.
The increased secretion of lactate is a consequence of hyperosmolarity. Exposure to increased osmotic pressure has major effects on the metabolic profile of the target cells. Hamraz exposed normal Chinese Ovarian cells (CHO cells) to increased osmotic pressure and showed that these cells expressed increased anaerobic glycolysis, decreased oxidative phosphorylation, and enhanced lactate secretion—the so-called Warburg effect [106]. Table 1 summarizes the metabolic similarities and differences between cancer, Alzheimer's disease, and AMD.
11. Shift to Alternative Energy Sources
There is very limited data on the metabolic pathways involved in macular degeneration. The crucial enzymatic activities of most enzymes in glycolysis and mitochondria have not been measured. It is probable that the metabolic patterns are the same in AMD as in Alzheimer’s disease or even cancer. However, it has been recently shown that homocysteine causes a metabolic shift from mitochondrial respiration to a high rate of glycolysis in RPE cells in AMD. A high rate of glycolysis was demonstrated by a high lactate/pyruvate ratio. This led to the activation of angiogenesis via increased levels of VEGF [107].
In cancer and Alzheimer's disease, there is a decrease in mitochondrial activity. There is a decrease in the activity of the pyruvate kinase, which conveys pyruvate into acetyl coA. Pyruvate dehydrogenase (PDH) is a complex set of subunits. It converts pyruvate into acetyl-CoA. This complex enzyme connects glycolysis, which takes place in the cytoplasm, and the mitochondria, where the Krebs cycle takes place. The activity of the PDH is decreased in both cancer and AD [7,8]. Lipoic acid is a cofactor of the second subunit of PDH.
In AMD, supplementation with lipoic acid improves long-term vision [108], suggesting the key role of pyruvate dehydrogenase. Methylene blue, a century-old drug, can receive two electrons from NADH in the presence of complex I and donate them to cytochrome C, providing an alternative electron transfer pathway in defective mitochondria [109]. The addition of Methylene Blue decreases Warburg’s effect induced by increased osmolarity and lowers the secretion of lactic acid [110].
Methylene blue provides a protective effect in neurons and astrocytes against various insults in vitro, like superoxide production. In humans, methylene blue appears effective in the treatment of early-stage AD and memory loss [109,110,111]. In animal models of AMD, methylene blue prevents neurodegeneration caused by rotenone in the retina [111].
12. Conclusions
This work strongly suggests that macular degeneration, like inflammation, is a straightforward consequence of increased osmotic pressure. In animals, drugs that increase osmotic pressure cause AMD. Increased pressure results in intracellular acidosis and apoptosis. Treatment should aim at normalizing the pressure and limiting the metabolic consequences of increased pressure. Drugs normalizing metabolic abnormalities such as those seen in AMD appear to be promising.
Author Contributions
LS drafted the manuscript; MH participated in the conceptualization of the paper; AB participated in writing and revising the manuscript. JS, critically read the manuscript.
Funding
The authors declare that no funds, grants, or other support were received during the preparation of this manuscript.
Acknowledgments
The authors want to acknowledge the help of Romain Attal, Dr Ricard Luscan and Professor Vincent Fleury.
Conflicts of Interest
The authors declare that the research was conducted lacking any commercial or financial relationships that could be construed as a potential conflict of interest.
References
- Reitz, C.; Brayne, C.; Mayeux, R. Epidemiology of Alzheimer Disease. Nature Reviews Neurology 2011, 7, 137–152. [Google Scholar] [CrossRef] [PubMed]
- Adami, H.-O.; Hunter, D.; Trichopoulos, D. Textbook of Cancer Epidemiology; Oxford University Press, 2008; ISBN 0-19-971863-6.
- Nogueira, M.L.; Moreira, J.d.V.; Baronzio, G.F.; Dubois, B.; Steyaert, J.-M.; Schwartz, L. Mechanical Stress as the Common Denominator between Chronic Inflammation, Cancer, and Alzheimer’s Disease. Front. Oncol. 2015, 5, 197. [Google Scholar] [CrossRef] [PubMed]
- Seyfried, T.N.; Shelton, L.M. Cancer as a metabolic disease. Nutr. Metab. 2010, 7, 7. [Google Scholar] [CrossRef] [PubMed]
- Craft, S. The Role of Metabolic Disorders in Alzheimer Disease and Vascular Dementia: Two Roads Converged. Archives of neurology 2009, 66, 300–305. [Google Scholar] [CrossRef] [PubMed]
- Vallée, A.; Lecarpentier, Y.; Guillevin, R.; Vallée, J.-N. Aerobic Glycolysis Hypothesis Through WNT/Beta-Catenin Pathway in Exudative Age-Related Macular Degeneration. J. Mol. Neurosci. 2017, 62, 368–379. [Google Scholar] [CrossRef] [PubMed]
- Léveillard, T.; Philp, N.J.; Sennlaub, F. Is Retinal Metabolic Dysfunction at the Center of the Pathogenesis of Age-Related Macular Degeneration? International journal of molecular sciences 2019, 20, 762. [Google Scholar] [CrossRef] [PubMed]
- Abolhassani, M.; Wertz, X.; Pooya, M.; Chaumet-Riffaud, P.; Guais, A.; Schwartz, L. Hyperosmolarity causes inflammation through the methylation of protein phosphatase 2A. Inflamm. Res. 2008, 57, 419–429. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, L.; Peres, S.; Jolicoeur, M.; da Veiga Moreira, J. Cancer and Alzheimer’s disease: intracellular pH scales the metabolic disorders. Biogerontology 2020, 21, 683–694. [Google Scholar] [CrossRef] [PubMed]
- Colijn, J.M.; Buitendijk, G.H.; Prokofyeva, E.; Alves, D.; Cachulo, M.L.; Khawaja, A.P.; Cougnard-Gregoire, A.; Merle, B.M.; Korb, C.; Erke, M.G. Prevalence of Age-Related Macular Degeneration in Europe: The Past and the Future. Ophthalmology 2017, 124, 1753–1763. [Google Scholar] [CrossRef]
- Al-Zamil, W.M.; Yassin, S.A. Recent developments in age-related macular degeneration: a review. Clin. Interv. Aging 2017, 12, 1313–1330. [Google Scholar] [CrossRef]
- Buitendijk, G.H.; Rochtchina, E.; Myers, C.; van Duijn, C.M.; Lee, K.E.; Klein, B.E.; Meuer, S.M.; de Jong, P.T.; Holliday, E.G.; Tan, A.G. Prediction of Age-Related Macular Degeneration in the General Population: The Three Continent AMD Consortium. Ophthalmology 2013, 120, 2644–2655. [Google Scholar] [CrossRef] [PubMed]
- Tan, W.; Zou, J.; Yoshida, S.; Jiang, B.; Zhou, Y. The Role of Inflammation in Age-Related Macular Degeneration. Int. J. Biol. Sci. 2020, 16, 2989–3001. [Google Scholar] [CrossRef]
- Brodzka, S.; Baszyński, J.; Rektor, K.; Hołderna-Bona, K.; Stanek, E.; Kurhaluk, N.; Tkaczenko, H.; Malukiewicz, G.; Woźniak, A.; Kamiński, P. The Role of Glutathione in Age-Related Macular Degeneration (AMD). Int. J. Mol. Sci. 2024, 25, 4158. [Google Scholar] [CrossRef] [PubMed]
- Hrishikesh, V.; Pranaykumar, S. Age-Related Macular Degeneration: Epidemiology, Pathophysiology, Diagnosis, and Treatment. Cureus 2022, 14. [Google Scholar]
- Davis, M.D.; Gangnon, R.E.; Lee, L.Y.; Hubbard, L.D.; Klein, B.; Klein, R.; Ferris, F.L.; Bressler, S.B.; Milton, R.C. The Age-Related Eye Disease Study Severity Scale for Age-Related Macular Degeneration: AREDS Report No. 17. Archives of ophthalmology (Chicago, Ill.: 1960) 2005, 123, 1484–1498. [Google Scholar] [PubMed]
- Ferris, F.L.; Davis, M.D.; Clemons, T.E.; Lee, L.-Y.; Chew, E.Y.; Lindblad, A.S.; Milton, R.C.; Bressler, S.B.; Klein, R. A Simplified Severity Scale for Age-Related Macular Degeneration: AREDS Report No. 18. Archives of ophthalmology (Chicago, Ill.: 1960) 2005, 123, 1570–1574. [Google Scholar] [PubMed]
- Lim, L.S.; Mitchell, P.; Seddon, J.M.; Holz, F.G.; Wong, T.Y. Age-Related Macular Degeneration. The Lancet 2012, 379, 1728–1738. [Google Scholar] [CrossRef] [PubMed]
- Liew, G.; Joachim, N.; Mitchell, P.; Burlutsky, G.; Wang, J.J. Validating the AREDS Simplified Severity Scale of Age-Related Macular Degeneration with 5- and 10-Year Incident Data in a Population-Based Sample. Ophthalmology 2016, 123, 1874–1878. [Google Scholar] [CrossRef]
- Garcia-Layana, A.; Cabrera-López, F.; García-Arumí, J.; Arias-Barquet, L.; Ruiz-Moreno, J.M. Early and intermediate age-related macular degeneration: update and clinical review. Clin. Interv. Aging 2017, 12, 1579–1587. [Google Scholar] [CrossRef]
- Friedman, D.S.; O'Colmain, B.J.; Muñoz, B.; Tomany, S.C.; McCarty, C.; De Jong, P.T.V.M.; Nemesure, B.; Mitchell, P.; Kempen, J.; Congdon, N. Prevalence of Age-Related Macular Degeneration in the United States. Arch. Ophthalmol. 2004, 122, 564–572. [Google Scholar] [CrossRef]
- von der Emde, L.; Pfau, M.; Holz, F.G.; Fleckenstein, M.; Kortuem, K.; Keane, P.A.; Rubin, D.L.; Schmitz-Valckenberg, S. AI-based structure-function correlation in age-related macular degeneration. Eye 2021, 35, 2110–2118. [Google Scholar] [CrossRef]
- Schultz, N.M.; Bhardwaj, S.; Barclay, C.; Gaspar, L.; Schwartz, J. Global Burden of Dry Age-Related Macular Degeneration: A Targeted Literature Review. Clin. Ther. 2021, 43, 1792–1818. [Google Scholar] [CrossRef] [PubMed]
- Telander, D.G. Inflammation and Age-Related Macular Degeneration (AMD). Semin. Ophthalmol. 2011, 26, 192–197. [Google Scholar] [CrossRef]
- Fleckenstein, M.; Schmitz-Valckenberg, S.; Chakravarthy, U. Age-Related Macular Degeneration: A Review. JAMA 2024, 331, 147–157. [Google Scholar] [CrossRef]
- Heesterbeek, T.J.; Lorés-Motta, L.; Hoyng, C.B.; Lechanteur, Y.T.; Hollander, A.I. Risk factors for progression of age-related macular degeneration. Ophthalmic Physiol. Opt. 2020, 40, 140–170. [Google Scholar] [CrossRef]
- Schlanitz, F.G.; Baumann, B.; Kundi, M.; Sacu, S.; Baratsits, M.; Scheschy, U.; Shahlaee, A.; Mittermüller, T.J.; Montuoro, A.; Roberts, P.; et al. Drusen volume development over time and its relevance to the course of age-related macular degeneration. Br. J. Ophthalmol. 2017, 101, 198–203. [Google Scholar] [CrossRef] [PubMed]
- Nathoo, N.A.; Or, C.; Young, M.; Chui, L.; Fallah, N.; Kirker, A.W.; Albiani, D.A.; Merkur, A.B.; Forooghian, F. Optical Coherence Tomography–Based Measurement of Drusen Load Predicts Development of Advanced Age-Related Macular Degeneration. Arch. Ophthalmol. 2014, 158, 757–761. [Google Scholar] [CrossRef] [PubMed]
- Folgar, F.A.; Yuan, E.L.; Sevilla, M.B.; Chiu, S.J.; Farsiu, S.; Chew, E.Y.; Toth, C.A. Drusen Volume and Retinal Pigment Epithelium Abnormal Thinning Volume Predict 2-Year Progression of Age-Related Macular Degeneration. Ophthalmology 2016, 123, 39–50. [Google Scholar] [CrossRef]
- Shim, S.H.; Kim, S.-G.; Bae, J.H.; Yu, H.G.; Song, S.J. Risk Factors for Progression of Early Age-Related Macular Degeneration in Koreans. Ophthalmic Epidemiology 2016, 23, 80–87. [Google Scholar] [CrossRef]
- Joachim, N.D.; Mitchell, P.; Kifley, A.; Wang, J.J. Incidence, Progression, and Associated Risk Factors of Medium Drusen in Age-Related Macular Degeneration: Findings from the 15-Year Follow-up of an Australian Cohort. JAMA ophthalmology 2015, 133, 698–705. [Google Scholar] [CrossRef]
- Hallak, J.A.; de Sisternes, L.; Osborne, A.; Yaspan, B.; Rubin, D.L.; Leng, T. Imaging, Genetic, and Demographic Factors Associated with Conversion to Neovascular Age-Related Macular Degeneration: Secondary Analysis of a Randomized Clinical Trial. JAMA ophthalmology 2019, 137, 738–744. [Google Scholar] [CrossRef] [PubMed]
- Johnson, L.V.; Leitner, W.P.; Staples, M.K.; Anderson, D.H. Complement Activation and Inflammatory Processes in Drusen Formation and Age Related Macular Degeneration. Exp. Eye Res. 2001, 73, 887–896. [Google Scholar] [CrossRef] [PubMed]
- Fritsche, L.G.; Igl, W.; Bailey, J.N.C.; Grassmann, F.; Sengupta, S.; Bragg-Gresham, J.L.; Burdon, K.P.; Hebbring, S.J.; Wen, C.; Gorski, M.; et al. A large genome-wide association study of age-related macular degeneration highlights contributions of rare and common variants. Nat. Genet. 2016, 48, 134–143. [Google Scholar] [CrossRef] [PubMed]
- Klein, R.; Peto, T.; Bird, A.; Vannewkirk, M.R. The epidemiology of age-related macular degeneration. Arch. Ophthalmol. 2004, 137, 486–495. [Google Scholar] [CrossRef] [PubMed]
- Majewski, J.; Schultz, D.W.; Weleber, R.G.; Schain, M.B.; Edwards, A.O.; Matise, T.C.; Acott, T.S.; Ott, J.; Klein, M.L. Age-Related Macular Degeneration—a Genome Scan in Extended Families. Am. J. Hum. Genet. 2003, 73, 540–550. [Google Scholar] [CrossRef] [PubMed]
- E Hughes, A.; Orr, N.; Esfandiary, H.; Diaz-Torres, M.; Goodship, T.; Chakravarthy, U. A common CFH haplotype, with deletion of CFHR1 and CFHR3, is associated with lower risk of age-related macular degeneration. Nat. Genet. 2006, 38, 1173–1177. [Google Scholar] [CrossRef]
- Cipriani, V.; Leung, H.-T.; Plagnol, V.; Bunce, C.; Khan, J.C.; Shahid, H.; Moore, A.T.; Harding, S.P.; Bishop, P.N.; Hayward, C.; et al. Genome-wide association study of age-related macular degeneration identifies associated variants in the TNXB–FKBPL–NOTCH4 region of chromosome 6p21.3. Hum. Mol. Genet. 2012, 21, 4138–4150. [Google Scholar] [CrossRef]
- Smailhodzic, D.; Muether, P.S.; Chen, J.; Kwestro, A.; Zhang, A.Y.; Omar, A.; Van de Ven, J.P.; Keunen, J.E.; Kirchhof, B.; Hoyng, C.B.; et al. Cumulative Effect of Risk Alleles in CFH, ARMS2, and VEGFA on the Response to Ranibizumab Treatment in Age-related Macular Degeneration. Ophthalmology 2012, 119, 2304–2311. [Google Scholar] [CrossRef]
- Medina, F.M.C.; da Motta, A.A.L.; Takahashi, W.Y.; Carricondo, P.C.; Motta, M.M.d.S.; Melo, M.B.; Vasconcellos, J.P.C. Pharmacogenetic Effect of Complement Factor H Gene Polymorphism in Response to the Initial Intravitreal Injection of Bevacizumab for Wet Age-Related Macular Degeneration. Ophthalmic Res. 2015, 54, 169–174. [Google Scholar] [CrossRef]
- McCarty, C.A.; Mukesh, B.N.; Fu, C.L.; Mitchell, P.; Wang, J.J.; Taylor, H.R. Risk Factors for Age-Related Maculopathy: The Visual Impairment Project. Archives of ophthalmology 2001, 119, 1455–1462. [Google Scholar] [CrossRef]
- Joachim, N.; Mitchell, P.; Kifley, A.; Rochtchina, E.; Hong, T.; Wang, J.J. Incidence and Progression of Geographic Atrophy: Observations from a Population-Based Cohort. Ophthalmology 2013, 120, 2042–2050. [Google Scholar] [CrossRef] [PubMed]
- Joachim, N.; Mitchell, P.; Rochtchina, E.; Tan, A.G.; Wang, J.J. Incidence and Progression of Reticular Drusen in Age-Related Macular Degeneration: Findings from an Older Australian Cohort. Ophthalmology 2014, 121, 917–925. [Google Scholar] [CrossRef] [PubMed]
- Jonasson, F.; Fisher, D.E.; Eiriksdottir, G.; Sigurdsson, S.; Klein, R.; Launer, L.J.; Harris, T.; Gudnason, V.; Cotch, M.F. Five-Year Incidence, Progression, and Risk Factors for Age-Related Macular Degeneration: The Age, Gene/Environment Susceptibility Study. Ophthalmology 2014, 121, 1766–1772. [Google Scholar] [CrossRef] [PubMed]
- Velilla, S.; García-Medina, J.J.; García-Layana, A.; Dolz-Marco, R.; Pons-Vázquez, S.; Pinazo-Durán, M.D.; Gómez-Ulla, F.; Arévalo, J.F.; Díaz-Llopis, M.; Gallego-Pinazo, R. Smoking and Age-Related Macular Degeneration: Review and Update. J. Ophthalmol. 2013, 2013, 895147. [Google Scholar] [CrossRef]
- Marin, A.I.; Poppelaars, F.; Wagner, B.D.; Palestine, A.G.; Patnaik, J.L.; Holers, V.M.; Frazer-Abel, A.A.; Mathias, M.T.; Manoharan, N.; Fonteh, C.N.; et al. Sex and Age-Related Differences in Complement Factors Among Patients With Intermediate Age-Related Macular Degeneration. Transl. Vis. Sci. Technol. 2022, 11, 22–22. [Google Scholar] [CrossRef] [PubMed]
- Smith, W.; Mitchell, P.; Wang, J. Gender, Oestrogen, Hormone Replacement and Age-related Macular Degeneration: Results from the Blue Mountains Eye Study. Australian and New Zealand journal of ophthalmology 1997, 25, 13–15. [Google Scholar] [CrossRef]
- Clemons, T.E.; Milton, R.C.; Klein, R.; Seddon, J.M.; Ferris 3rd, F.L. Risk Factors for the Incidence of Advanced Age-Related Macular Degeneration in the Age-Related Eye Disease Study (AREDS) AREDS Report No. 19. Ophthalmology 2005, 112, 533–539. [Google Scholar]
- Tikellis, G.; Robman, L.D.; Dimitrov, P.; Nicolas, C.; A McCarty, C.; Guymer, R.H. Characteristics of progression of early Age-related macular degeneration: the Cardiovascular Health and Age-related maculopathy Study. Eye 2006, 21, 169–176. [Google Scholar] [CrossRef] [PubMed]
- Cougnard-Grégoire, A.; Delyfer, M.-N.; Korobelnik, J.-F.; Rougier, M.-B.; Malet, F.; Le Goff, M.; Dartigues, J.-F.; Colin, J.; Barberger-Gateau, P.; Delcourt, C. Long-Term Blood Pressure and Age-Related Macular Degeneration: The ALIENOR Study. Investig. Opthalmology Vis. Sci. 2013, 54, 1905–1912. [Google Scholar] [CrossRef]
- Parekh, N.; Voland, R.P.; Moeller, S.M.; Blodi, B.A.; Ritenbaugh, C.; Chappell, R.J.; Wallace, R.B.; Mares, J.A. ; CAREDS Research Study Group Association between Dietary Fat Intake and Age-Related Macular Degeneration in the Carotenoids in Age-Related Eye Disease Study (CAREDS): An Ancillary Study of the Women’s Health Initiative. Archives of ophthalmology 2009, 127, 1483–1493. [Google Scholar] [CrossRef]
- Garber, J.E.; Offit, K. Hereditary Cancer Predisposition Syndromes. J. Clin. Oncol. 2005, 23, 276–292. [Google Scholar] [CrossRef] [PubMed]
- Rocchi, A.; Pellegrini, S.; Siciliano, G.; Murri, L. Causative and susceptibility genes for Alzheimer’s disease: a review. Brain Res. Bull. 2003, 61, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Brandt, A.; Bermejo, J.L.; Sundquist, J.; Hemminki, K. Age of onset in familial cancer. Ann. Oncol. 2008, 19, 2084–2088. [Google Scholar] [CrossRef] [PubMed]
- Blennow, K.; de Leon, M.J.; Zetterberg, H. Alzheimer’s Disease. The Lancet 2006, 368, 387–403. [Google Scholar] [CrossRef] [PubMed]
- Waring, S.C.; Rosenberg, R.N. Genome-Wide Association Studies in Alzheimer Disease. Arch. Neurol. 2008, 65, 329–334. [Google Scholar] [CrossRef] [PubMed]
- Selkoe, D.J. Translating cell biology into therapeutic advances in Alzheimer's disease. Nature 1999, 399, A23–A31. [Google Scholar] [CrossRef] [PubMed]
- Haddad, S.; Chen, C.A.; Santangelo, S.L.; Seddon, J.M. The Genetics of Age-Related Macular Degeneration: A Review of Progress to Date. Surv. Ophthalmol. 2006, 51, 316–363. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, A.; Weber, O. In Memoriam of Rudolf Virchow: A Historical Retrospective Including Aspects of Inflammation, Infection and Neoplasia. In Infection and inflammation: impacts on oncogenesis; Karger Publishers, 2006; Vol. 13, pp. 1–15.
- James, S.; Pogribna, M.; Miller, B.J.; Bolon, B.; Muskhelishvili, L. Characterization of cellular response to silicone implants in rats: implications for foreign-body carcinogenesis. Biomaterials 1997, 18, 667–675. [Google Scholar] [CrossRef] [PubMed]
- Tan, H.; Yang, W.; Wu, C.; Liu, B.; Lu, H.; Wang, H.; Yan, H. Assessment of the role of intracranial hypertension and stress on hippocampal cell apoptosis and hypothalamic-pituitary dysfunction after TBI. Sci. Rep. 2017, 7, 3805. [Google Scholar] [CrossRef]
- Nogueira, M.L.; Hamraz, M.; Abolhassani, M.; Bigan, E.; Lafitte, O.; Steyaert, J.-M.; Dubois, B.; Schwartz, L. Mechanical Stress Increases Brain Amyloid β, Tau, and α-Synuclein Concentrations in Wild-Type Mice. Alzheimer’s & Dementia 2018, 14, 444–453. [Google Scholar]
- Pawelec, G.; Goldeck, D.; Derhovanessian, E. Inflammation, Ageing and Chronic Disease. Current opinion in immunology 2014, 29, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Ginaldi, L.; Mengoli, L.P.; De Martinis, M. Osteoporosis, Inflammation and Ageing. Handbook on Immunosenescence: Basic Understanding and Clinical Applications 2009, 1329–1352.
- Sparrow, J.R. Bisretinoids of RPE Lipofuscin: Trigger for Complement Activation in Age-Related Macular Degeneration. Inflammation and Retinal Disease: Complement Biology and Pathology 2010, 63–74.
- Anderson, D.H.; Talaga, K.C.; Rivest, A.J.; Barron, E.; Hageman, G.S.; Johnson, L.V. Characterization of β amyloid assemblies in drusen: the deposits associated with aging and age-related macular degeneration. Exp. Eye Res. 2004, 78, 243–256. [Google Scholar] [CrossRef] [PubMed]
- Cotran, R.S.; Majno, G. The Delayed and Prolonged Vascular Leakage in Inflammation: I. Topography of the Leaking Vessels after Thermal Injury. The American Journal of Pathology 1964, 45, 261. [Google Scholar] [CrossRef] [PubMed]
- Roviezzo, F.; Tsigkos, S.; Kotanidou, A.; Bucci, M.; Brancaleone, V.; Cirino, G.; Papapetropoulos, A. Angiopoietin-2 Causes Inflammation in Vivo by Promoting Vascular Leakage. J. Pharmacol. Exp. Ther. 2005, 314, 738–744. [Google Scholar] [CrossRef]
- Schwartz, L.; Israël, M.; Philippe, I. Inflammation and Carcinogenesis: A Change in the Metabolic Process. Cancer Microenvironment and Therapeutic Implications: Tumor Pathophysiology Mechanisms and Therapeutic Strategies 2009, 3–18.
- Lemp, M.A.; Bron, A.J.; Baudouin, C.; del Castillo, J.M.B.; Geffen, D.; Tauber, J.; Foulks, G.N.; Pepose, J.S.; Sullivan, B.D. Tear Osmolarity in the Diagnosis and Management of Dry Eye Disease. Arch. Ophthalmol. 2011, 151, 792–798. [Google Scholar] [CrossRef] [PubMed]
- Gilbard, J.P.; Farris, R.L. OCULAR SURFACE DRYING AND TEAR FILM OSMOLARITY IN THYROID EYE DISEASE. Acta Ophthalmol. 1983, 61, 108–116. [Google Scholar] [CrossRef] [PubMed]
- Anwar, Z.; Wellik, S.R.; Galor, A. Glaucoma Therapy and Ocular Surface Disease: Current Literature and Recommendations. Current opinion in ophthalmology 2013, 24, 136–143. [Google Scholar] [CrossRef]
- Jacob, T.; Duncan, G. Osmotic influences on lens membrane characteristics. Exp. Eye Res. 1980, 31, 505–512. [Google Scholar] [CrossRef]
- Joyal, J.-S.; Gantner, M.L.; Smith, L.E. Retinal energy demands control vascular supply of the retina in development and disease: The role of neuronal lipid and glucose metabolism. Prog. Retin. Eye Res. 2017, 64, 131–156. [Google Scholar] [CrossRef] [PubMed]
- Alm, A.; Bill, A. Ocular and optic nerve blood flow at normal and increased intraocular pressures in monkeys (Macaca irus): a study with radioactively labelled microspheres including flow determinations in brain and some other tissues. Exp. Eye Res. 1973, 15, 15–29. [Google Scholar] [CrossRef]
- Moore, D.J.; Clover, G.M. The effect of age on the macromolecular permeability of human Bruch's membrane. . 2001, 42, 2970–5. [Google Scholar] [PubMed]
- Pournaras, C.J.; Rungger-Brändle, E.; Riva, C.E.; Hardarson, S.H.; Stefansson, E. Regulation of retinal blood flow in health and disease. Prog. Retin. Eye Res. 2008, 27, 284–330. [Google Scholar] [CrossRef] [PubMed]
- Boscher, C.; Erol, O.; Luscan, R. Remodeling of the choroidal vasculature and the role of choriocapillaris perfusion drop in pachychoroid diseases: a global rheological approach. Graefe's Arch. Clin. Exp. Ophthalmol. 2023, 261, 3045–3046. [Google Scholar] [CrossRef] [PubMed]
- Hoseini-Yazdi, H.; Vincent, S.J.; Collins, M.J.; Read, S.A.; Alonso-Caneiro, D. Wide-field choroidal thickness in myopes and emmetropes. Sci. Rep. 2019, 9, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Spaide, R.F.; Koizumi, H.; Pozonni, M.C. Enhanced Depth Imaging Spectral-Domain Optical Coherence Tomography. Am. J. Ophthalmol. 2008, 146, 496–500. [Google Scholar] [CrossRef] [PubMed]
- Margolis, R.; Spaide, R.F. A Pilot Study of Enhanced Depth Imaging Optical Coherence Tomography of the Choroid in Normal Eyes. Arch. Ophthalmol. 2009, 147, 811–815. [Google Scholar] [CrossRef] [PubMed]
- Tokuda, S.; Yu, A.S.L. Regulation of Epithelial Cell Functions by the Osmolality and Hydrostatic Pressure Gradients: A Possible Role of the Tight Junction as a Sensor. Int. J. Mol. Sci. 2019, 20, 3513. [Google Scholar] [CrossRef] [PubMed]
- Shirao, Y.; Steinberg, R.H. Mechanisms of effects of small hyperosmotic gradients on the chick RPE. . 1987, 28, 2015–25. [Google Scholar]
- Kvanta, A.; Algvere, P.V.; Berglin, L.; Seregard, S. Subfoveal fibrovascular membranes in age-related macular degeneration express vascular endothelial growth factor. . 1996, 37, 1929–34. [Google Scholar] [CrossRef]
- Hollborn, M.; Vogler, S.; Reichenbach, A.; Wiedemann, P.; Bringmann, A.; Kohen, L. Regulation of the hyperosmotic induction of aquaporin 5 and VEGF in retinal pigment epithelial cells: Involvement of NFAT5. 2015, 21, 360–377.
- Veltmann, M.; Hollborn, M.; Reichenbach, A.; Wiedemann, P.; Kohen, L.; Bringmann, A. Osmotic Induction of Angiogenic Growth Factor Expression in Human Retinal Pigment Epithelial Cells. PLOS ONE 2016, 11, e0147312. [Google Scholar] [CrossRef]
- Marmor, M. Retinal Detachment from Hyperosmotic Intravitreal Injection. Investigative Ophthalmology & Visual Science 1979, 18, 1237–1244.
- Turner, J.L.; Bierman, E.L. Effects of glucose and sorbitol on proliferation of cultured human skin fibroblasts and arterial smooth-muscle cells. Diabetes 1978, 27, 583–588. [Google Scholar] [CrossRef] [PubMed]
- Pennesi, M.E.; Neuringer, M.; Courtney, R.J. Animal models of age related macular degeneration. Mol. Asp. Med. 2012, 33, 487–509. [Google Scholar] [CrossRef] [PubMed]
- Lyzogubov, V.V.; Tytarenko, R.G.; Liu, J.; Bora, N.S.; Bora, P.S. Polyethylene Glycol (PEG)-induced Mouse Model of Choroidal Neovascularization. J. Biol. Chem. 2011, 286, 16229–16237. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, R.; Logan, C.; Lyzogubov, V.V.; Bora, N.S.; Bora, P.S. Wet and Dry Age-Related Macular Degeneration Induced by Polyethylene Glycol. 2023.
- Christen, R.; Schackmann, R.W.; Shapiro, B.M. Metabolism of sea urchin sperm. Interrelationships between intracellular pH, ATPase activity, and mitochondrial respiration. J. Biol. Chem. 1983, 258, 5392–5399. [Google Scholar] [CrossRef] [PubMed]
- Sinning, A.; Hübner, C.A. Minireview: pH and synaptic transmission. FEBS Lett. 2013, 587, 1923–1928. [Google Scholar] [CrossRef] [PubMed]
- Zebhauser, P.T.; Berthele, A.; Goldhardt, O.; Diehl-Schmid, J.; Priller, J.; Ortner, M.; Grimmer, T. Cerebrospinal Fluid Lactate Levels along the Alzheimer’s Disease Continuum and Associations with Blood-Brain Barrier Integrity, Age, Cognition, and Biomarkers. Alzheimer’s Research & Therapy 2022, 14, 1–8. [Google Scholar]
- Demetrius, L.A.; Simon, D.K. An inverse-Warburg effect and the origin of Alzheimer’s disease. Biogerontology 2012, 13, 583–594. [Google Scholar] [CrossRef]
- Poitry-Yamate, C.; Poitry, S.; Tsacopoulos, M. Lactate released by Muller glial cells is metabolized by photoreceptors from mammalian retina. J. Neurosci. 1995, 15, 5179–5191. [Google Scholar] [CrossRef]
- Ng, S.K.; Wood, J.P.; Chidlow, G.; Han, G.; Kittipassorn, T.; Peet, D.J.; Casson, R.J. Cancer-like metabolism of the mammalian retina. Clin. Exp. Ophthalmol. 2015, 43, 367–376. [Google Scholar] [CrossRef]
- Bringmann, A.; Reichenbach, A. Role of Muller cells in retinal degenerations. Front. Biosci. 2001, 6, e77–92. [Google Scholar] [CrossRef]
- Yokosako, K.; Mimura, T.; Funatsu, H.; Noma, H.; Goto, M.; Kamei, Y.; Kondo, A.; Matsubara, M. Glycolysis in Patients with Age-Related Macular Degeneration. Open Ophthalmol. J. 2014, 8, 39–47. [Google Scholar] [CrossRef]
- Dhup, S.; Dadhich, R.K.; Porporato, P.E.; Sonveaux, P. Multiple Biological Activities of Lactic Acid in Cancer: Influences on Tumor Growth,Angiogenesis and Metastasis. Curr. Pharm. Des. 2012, 18, 1319–1330. [Google Scholar] [CrossRef]
- Song, J.; Lee, K.; Park, S.W.; Chung, H.; Jung, D.; Na, Y.R.; Quan, H.; Cho, C.S.; Che, J.-H.; Kim, J.H.; et al. Lactic Acid Upregulates VEGF Expression in Macrophages and Facilitates Choroidal Neovascularization. Investig. Ophthalmol. Vis. Sci. 2018, 59, 3747–3754. [Google Scholar] [CrossRef] [PubMed]
- Reber, F.; Kasper, M.; Siegner, A.; Kniep, E.; Seigel, G.; Funk, R.H. Alteration of the intracellular pH and apoptosis induction in a retinal cell line by the AGE-inducing agent glyoxal. Graefe's Arch. Clin. Exp. Ophthalmol. 2002, 240, 1022–1032. [Google Scholar] [CrossRef]
- Kumar, A.; Haery, C.; Paladugu, B.; Kumar, A.; Symeoneides, S.; Taiberg, L.; Osman, J.; Trenholme, G.; Opal, S.M.; Goldfarb, R.; et al. The Duration of Hypotension before the Initiation of Antibiotic Treatment Is a Critical Determinant of Survival in a Murine Model ofEscherichia coliSeptic Shock: Association with Serum Lactate and Inflammatory Cytokine Levels. J. Infect. Dis. 2006, 193, 251–258. [Google Scholar] [CrossRef] [PubMed]
- Marcoux, J.; McArthur, D.A.; Miller, C.; Glenn, T.C.; Villablanca, P.; Martin, N.A.; Hovda, D.A.; Alger, J.R.; Vespa, P.M. Persistent metabolic crisis as measured by elevated cerebral microdialysis lactate-pyruvate ratio predicts chronic frontal lobe brain atrophy after traumatic brain injury*. Crit. Care Med. 2008, 36, 2871–2877. [Google Scholar] [CrossRef] [PubMed]
- Ikizawa, T.; Ikeda, K.; Arita, M.; Kitajima, S.; Soga, T.; Ichijo, H.; Naguro, I. Mitochondria directly sense osmotic stress to trigger rapid metabolic remodeling via regulation of pyruvate dehydrogenase phosphorylation. J. Biol. Chem. 2023, 299, 102837. [Google Scholar] [CrossRef]
- Hamraz, M.; Abolhassani, R.; Andriamihaja, M.; Ransy, C.; Lenoir, V.; Schwartz, L.; Bouillaud, F. Hypertonic external medium represses cellular respiration and promotes Warburg/Crabtree effect. FASEB J. 2019, 34, 222–236. [Google Scholar] [CrossRef]
- Samra, Y.A.; Zaidi, Y.; Rajpurohit, P.; Raghavan, R.; Cai, L.; Kaddour-Djebbar, I.; Tawfik, A. Warburg Effect as a Novel Mechanism for Homocysteine-Induced Features of Age-Related Macular Degeneration. Int. J. Mol. Sci. 2023, 24, 1071. [Google Scholar] [CrossRef]
- Tao, Y.; Jiang, P.; Wei, Y.; Wang, P.; Sun, X.; Wang, H. α-Lipoic Acid Treatment Improves Vision-Related Quality of Life in Patients with Dry Age-Related Macular Degeneration. Tohoku J. Exp. Med. 2016, 240, 209–214. [Google Scholar] [CrossRef]
- Yang, S.-H.; Li, W.; Sumien, N.; Forster, M.; Simpkins, J.W.; Liu, R. Alternative mitochondrial electron transfer for the treatment of neurodegenerative diseases and cancers: Methylene blue connects the dots. Prog. Neurobiol. 2017, 157, 273–291. [Google Scholar] [CrossRef] [PubMed]
- Montégut, L.; Martínez-Basilio, P.C.; Moreira, J.d.V.; Schwartz, L.; Jolicoeur, M. Combining lipoic acid to methylene blue reduces the Warburg effect in CHO cells: From TCA cycle activation to enhancing monoclonal antibody production. PLOS ONE 2020, 15, e0231770. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X. ; C. Rojas, J.; Gonzalez-Lima, F. Methylene Blue Prevents Neurodegeneration Caused by Rotenone in the Retina. Neurotoxicity research 2006, 9, 47–57. [Google Scholar] [PubMed]
Figure 1.
Correlations between inflammation, hyperosmolarity, and metabolic changes in AMD.
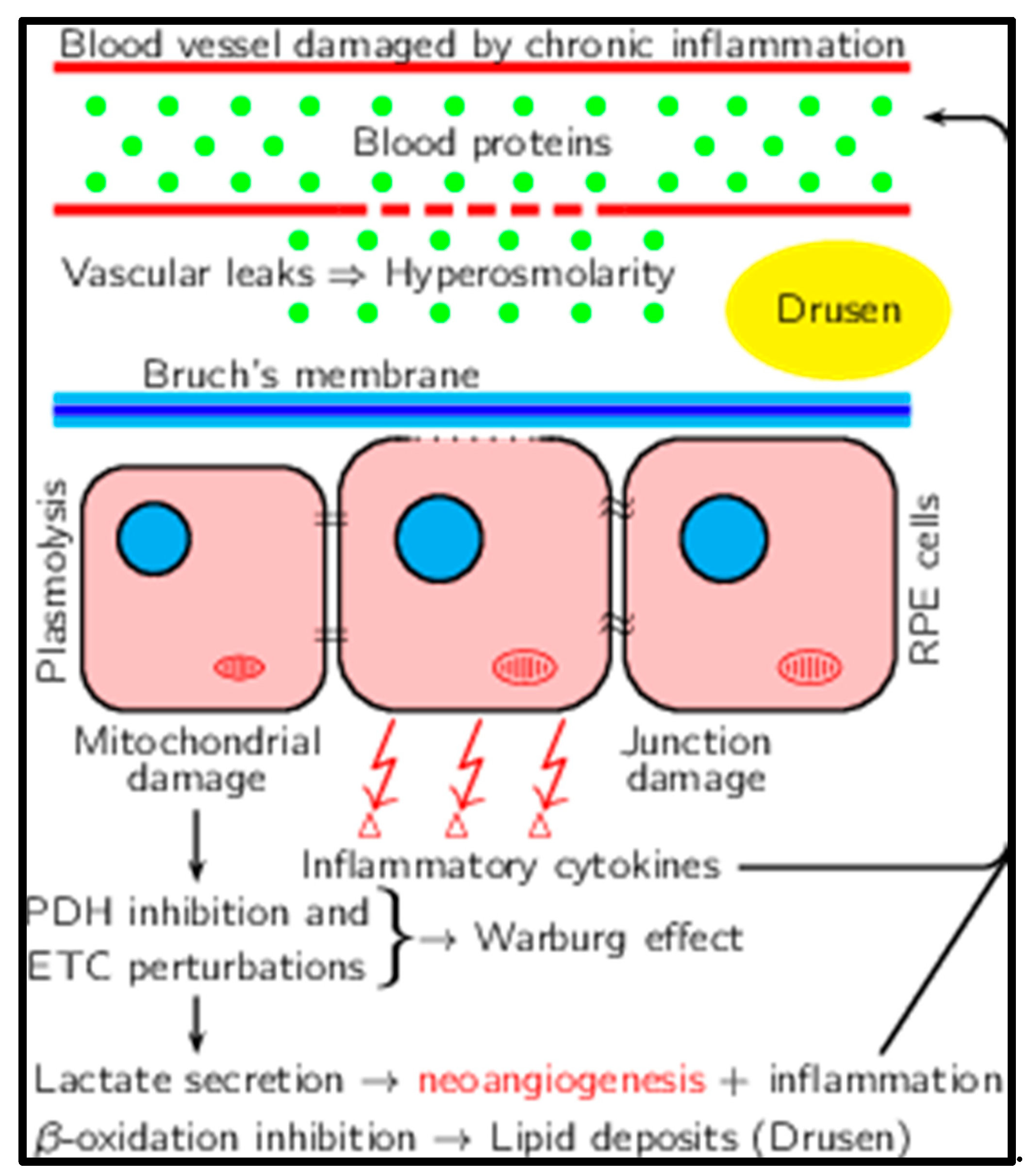

Table 1.
Comparison of metabolic characteristics of cancer, Alzheimer's disease and Macular degeneration.
Table 1.
Comparison of metabolic characteristics of cancer, Alzheimer's disease and Macular degeneration.
| Characteristic | Cancer | Alzheimer disease | Macular degeneration |
|---|---|---|---|
| Extracellular osmolarity | Increases | Increases | Increases |
| Oxidative phosphorylation | Decreases in cancer cells | Decreases in neurons | Decreases in photoreceptors |
| Extracellular lactate concentration | Increases | Increases | Increases |
| Intracellular pH | Increases in cancer cells | Decreases in neurons | Decreases in photoreceptors |
| Cell ecosystem | Cancer cells + Tumor Associated Macrophages + stroma | Neurons + glial cells | Photoreceptors + Müller cells |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



