
Submitted:
17 June 2024
Posted:
18 June 2024
You are already at the latest version
Abstract
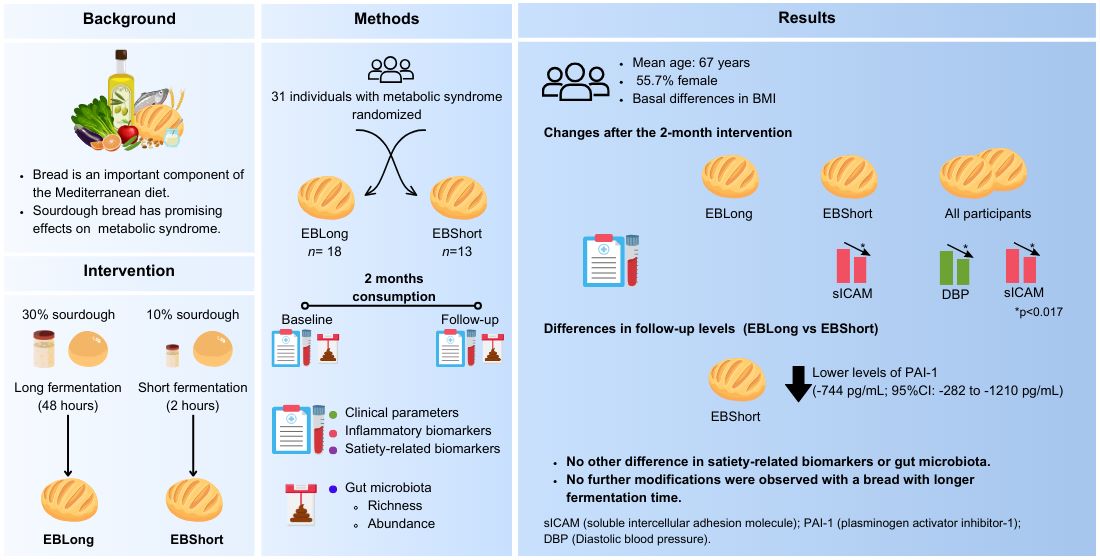
Mediterranean diet, featuring sourdough bread, shows promise in managing metabolic syndrome. This study explored the effects of two sourdough breads with differing fermentation times but similar nutritional profiles on inflammation, satiety, and gut microbiota composition in adults with metabolic syndrome. The double-blind clinical trial randomized participants to consume over two-months Elias Boulanger® long fermentation (48h) sourdough bread (EBLong) or Elias Boulanger® short fermentation (2h) sourdough bread (EBShort). We assessed clinical parameters, inflammatory- and satiety-related biomarkers; and richness and abundance of gut microbiota at baseline and follow-up. 31 individuals (mean age: 67, 55.7% female) participated. EBShort was associated with reduced soluble intercellular adhesion molecule (sICAM) levels, and irrespective of the adjudicated intervention, all participants had a decrease in sICAM and diastolic pressure relative to baseline (p<0.017). At follow-up, plasminogen activator inhibitor-1 (PAI-1) levels were lower in EBShort (-744 pg/mL; 95%CI: -282 to -1210 pg/mL) compared to EBLong. High-quality sourdough bread may offer mild benefits in blood pressure and inflammation for those with metabolic syndrome, however, only EBLong was associated to reduced Enterobacteriaceae abundance. Further research is needed to understand potential benefits and mechanisms of sourdough bread in managing metabolic syndrome and improving cardiovascular health.
Keywords:
sourdough bread
; metabolic syndrome
; sICAM
; PAI-1
; blood pressure
1. Introduction
Metabolic syndrome, a group of closely interrelated metabolic disorders, is strongly associated with cardiovascular disease incidence and the risk of developing type 2 diabetes mellitus [1,2]. The risk factors for metabolic syndrome include abdominal obesity, high levels of triglycerides, low concentrations of high-density lipoprotein cholesterol (HDLc), elevated blood pressure, insulin resistance and/or hyperglycemia [3]. The adoption of healthy lifestyle habits such as an adequate diet, weight management, regular physical activity, and avoiding smoking can reduce cardiometabolic risk [4]. However, a more holistic strategy is crucial to approach prevention; such strategies could include policies for easier access to healthy eating and physical activity and, innovations in the food industry.
In our context, the Mediterranean diet (MedDiet), which has proved to be effective in cardiovascular disease prevention [5], is characterized by the triad of bread, olives, and wine (but also their derived products) which have shown to improve metabolic syndrome risk factors [6]. Bread has been an important dietary component for Mediterranean populations since ancient times [7]. Sourdough bread, which dates back centuries, is considered a cultural heritage. It contains microorganisms such as yeast and lactic acid bacteria. The dough, made from wheat and/or rye flour and water, is left to naturally ferment at room temperature to increase its content of microorganisms [8]. These microorganisms provide the gas and acidity that give sourdough its aroma, texture, flavor, and keeping qualities [9]. Such characteristics are due to the longer fermentation time, as opposed to more commercial methods with very much shorter periods and higher temperatures [10]. It has yet to be established whether sourdough has a beneficial impact on health and whether a wholegrain sourdough, combining probiotics and prebiotics, could promote bacteria survival and microbiota equilibrium.
The present study aimed to evaluate the effects of two sourdough breads, with long (48 hours) and short (2 hours) fermentation in individuals with metabolic syndrome on clinical parameters, inflammatory biomarkers, and satiety hormones. Additionally, the study aimed to characterize the gut microbiota as a secondary objective. Our aim was to demonstrate a potentially yielding superior results with the long fermentation bread.
2. Materials and Methods
2.1. Study Design and Population
This is a randomized, controlled, double-blind clinical trial with two parallel groups.
Eligible participants were community-dwelling adults who met at least three components of metabolic syndrome according to the updated harmonized criteria of the joint statement from the International Diabetes Federation, the National Heart, Lung and Blood Institute, and the American Heart Association: hypertriglyceridemia [≥150 mg/dL (≥1.7 mmol/L)] or drug treatment for elevated triglycerides; low concentrations of HDLc [<50 mg/dL (<1.3 mmol/L) and <40 mg/dL (<1.03 mmol/L) in women and men, respectively] or drug treatment for low HDLc; elevated blood pressure (systolic ≥130 mmHg and/or diastolic ≥85 mmHg) or being treated for hypertension; high fasting plasma glucose [≥100 mg/dL (≥5.5 mmol/L)] or drug treatment; and elevated waist circumference for European individuals (≥88 cm in women and ≥102 cm in men) [3]. Exclusion criteria included: a) use of antibiotic, prebiotic and/or probiotic supplements in the 3 months prior to the start of the study; b) celiac disease; c) inflammatory bowel disease; d) history of bowel resection; e) alcoholism and/or any other drug dependence; f) use of non-steroidal anti-inflammatory drugs; g) use of immunosuppressants, antibiotics, and proton pump inhibitor; h) any disease or condition preventing compliance with the study protocol; and i) inability to provide informed consent.
Participants were assigned to one of the 2 sourdough bread (Elias Boulanger®) intervention arms: a) Elias Boulanger® long fermentation bread (EBLong) and, b) Elias Boulanger® short fermentation bread (EBShort). The volunteers were randomized between the 2 intervention arms using a random sequence generated by a computer program (Cardiovascular Epidemiology and Genetics Group, EGEC, Hospital del Mar Research Institute). The double-blind was maintained until the analysis of the results. No recommendations regarding diet, physical activity, or lifestyle were provided. Participants were instructed to replace their usual bread with the intervention bread while maintaining the same quantity consumed over a two-month period. Participants were instructed to collect the sliced bread from their chosen bakeries once a week or every two weeks (15 distributed throughout the province of Barcelona) and froze it. This trial was carried out at the Hospital del Mar Research Institute in Barcelona, Spain.
2.2. Ethical Aspects
The protocol of the study complied with the Declaration of Helsinki for Medical Research involving Human Subjects. The study protocol was reviewed and approved by the Clinical Research Ethics Committee of the Parc de Salut Mar Barcelona consortium (CEIC-Parc de Salut Mar) registry. Register number: ISRCTN89898870. All participants read and sign an informed consent before enrollment to the study.
2.3. Bread Composition and Fermentation Process
First, a starter was elaborated blending equal weight parts of whole-grain wheat flour (T110 flour, Triticum dicoccoides; Moulin de Colagne®, France) and water that rested for 24 hours at room temperature. At day 5 a mixture of infusions, dairy products and fruit (following the baker’s own receipt) was added and left at room temperature. The final starter had the following characteristics: a temperature of 34°C, a pH value of 6.68, and a total titratable acidity of 13.73 mL. Sourdough was created by combining the starter (30%), water (50%), and whole-grain wheat flour milled through a stone-mill (70%, T110 flour, T. dicoccoides; Moulin de Colagne®). Subsequently, a process known as feeding or back-slopping was carried out, involving the use of the initial mixture to ferment a new blend of water and flour at regular intervals of every 12–24 hours for 3 days and fermented for 4 days at 10-15 °C. This procedure aimed to stimulate the microbial fermentation and promote propagation of sourdough. Details of the high-quality sourdough preparation can be found elsewhere [11].
EBLong was formulated with sourdough (wt/wt; 30% flour basis), whole-grain flour (Triticum aestivum), water (wt/wt; 80% flour basis), dry baking yeast (Saccharomyces cerevisiae, wt/wt; <0.5% flour basis; Lesaffre (Hirondelle®)), and salt (wt/wt; 1.1% flour basis; Guerande®). Then, in a fermentation chamber (Eurifours®), bread was fermented for 48 hours: 46 hours of maturation at 4-6ºC and 2 hours of dough development at 28ºC.
EBShort was composed by sourdough (wt/wt; 10% flour basis), refined wheat flour (Farinera Corominas®) (being a blend of white bread since the starter contained whole grain flour), water (wt/wt 60% flour basis), yeast (S. cerevisiae, 0.010 g/kg of flour; Lesaffre (Hirondelle®)), salt (wt/wt 1.2% flour basis, Sal Costa®), enzymes(α-amylase, endoxylanasse, amyloglucosidanassa; Uniplus®), wheat gluten (0.010 g/kg of flour; Uniplus®), xanthan gum (0.005 g/kg of flour), and additive components: emulsifier E471, antioxidant E-300 (Uniplus®). It was fermented in a chamber (Eurifours®) for 2 hours at 28ºC.
Both breads were baked after the fermentation process in a Eurifours® oven at 200ºC for 90 minutes. Nutritional content of both breads can be found in Supplemental Table S1.
2.4. General and Life-Style Data
The following variables were recorded at baseline and follow-up: a) a questionnaire for adherence to the MedDiet [12]; b) a three-day food record collected during the three days prior to each visit (one of which at the weekend), which was further translated into nutrients using the Pro-PCN software (Barcelona, Spain) [13]; and c) an abbreviated questionnaire from the Minnesota Leisure Time Physical Activity Questionnaire [14].
2.5. Anthropometric and Exploration Data
The following variables were measured at baseline and follow-up: Participants’ weight was recorded in kilograms to one decimal point using a high-quality electronic scale, rounded to the nearest 100 grams, with individuals wearing light clothing and no shoes, jackets, or coats. Height was measured in centimeters with a stadiometer accurate to 1 centimeter. Waist circumference was calculated on expiration at the intermediate level between the last rib and the iliac crest, at the most prominent point of the trochanter. Body mass index (BMI) was derived by dividing weight (in kilograms) by height squared (in meters squared). Blood pressure was assessed while participants were seated with their backs and arms supported to ensure the cuff was at heart level, after refraining from smoking or consuming caffeine for 30 minutes. Measurements were taken on both arms using appropriate cuffs, with the arm displaying the higher mean diastolic blood pressure selected for subsequent measurements throughout the study. If the first two readings differed by more than 5 mmHg, additional readings were taken and averaged.
2.6. Laboratory Analysis
The following parameters were analyzed in fasting ethylenediaminetetraacetic acid (EDTA) plasma at baseline and after two-months follow-up: glucose (Glucose HK CP, Horiba ABX), triglycerides (Triglycerides CP, Horiba ABX), total cholesterol (Cholesterol CP, Horiba ABX) and, HDLc (HDLc Direct CP, Horiba ABX) were measured in an autoanalyzer ABX Pentra (Horiba ABX SAS, Spain). HOMA Index was calculated as (glucose x insulin)/405. We calculated LDLc with the Friedewald formula only when triglycerides were <300 mg/dL, higher values (≥300 mg) implied a missing value for LDLc. The following inflammation markers were analyzed: interleukin 6 and 8 (IL6 and IL8), tumor necrosis factor alpha (TNF-α) (Bio-Plex Cytokine 8-plex, Bio-Rad); soluble intercellular adhesion molecule (sICAM) (Bio-Plex Cytokine 2-plex, Bio-Rad), and Plasminogen activator inhibitor-1 (PAI-1) (Bio-Plex Pro Human Diabetes 10-plex, Bio-Rad). In addition, vascular-related hormones were also analyzed: insulin, C-peptide, ghrelin, leptin, glycoprotein 1 (GLP-1), glucagon, resistin, and vifastin (Bio-Plex Pro Human Diabetes 10-plex, Bio-Rad); a Luminex® xMAP® technology, in a BioPlex system was employed (Bio-Rad, Hercules, California, United States). The lipopolysaccharide binding protein (LBP) (Human LBP, Hycult Biotech) was measured with an Elisa Kit.
2.7. Intestinal Microbiota Analysis
Participants collected fecal samples at home in sterile containers provided for this purpose. They were instructed to keep their samples frozen until delivering them to the study staff the following day. The samples were sent to the Girona Biomedical Research Institute (IdIBGi) with dry ice to maintain the cold chain.
Genomic DNA was extracted from an approximately 0.25 g of faeces using commercial methods (NucleoSpin Soil kit, Macherey-Nagel®). The quality and quantity of the extracted nucleic acids were measured by Nanodrop ND 2000 UV-Vis spectrophotometer (Nanodrop, DE) and Qubit® (ThermoFisher Scientific) according to the manufacturers’ instructions. The region corresponding to the variable V3-V4 region of the 16S rRNA gene was determined with specific primers (341F/806R, [15]) and Illumina technology HiSeq 2000 using paired-end reads (generating 300 bp sequences). Briefly, amplification was performed after 25 PCR cycles. In this procedure, positive (CM) and negative (CN) controls were used to ensure quality control. The positive control is a Mock Community DNA (Zymobiomics Microbial Community DNA) control and it was processed the same way as the samples.
2.8. Bioinformatic Analysis
Raw demultiplexed forward reads were processed using the following methods and pipelines as implemented in QIIME2 version 2020.11 with default parameters unless stated [16]. DADA2 was used for quality filtering, denoising and amplicon sequence variant calling (ASV, i.e., phylotypes) using qiime dada2 denoise-sinle method [17]. Q16 was used as quality threshold to define read sizes for trimming (parameter: -- p-trunc-len). Reads were truncated at the position when the 75th percentile Phred score felt below Q16: 198 bp. After quality filtering steps, average sample size was 64,907 reads (min: 37,026 reads, max: 153,151 reads) and 1,896 phylotypes were detected. ASVs were aligned using the qiime alignment mafft method [18].The alignment was used to create a tree and to calculate phylogenetic relations between ASVs using qiime phylogeny fasttree method [19]. ASV tables were subsampled without replacement in order to even sample sizes for diversity analysis using qiime diversity core-metrics-phylogenetic pipeline. The smallest sample size was chosen for subsampling (i.e., 37,000 reads). Subsequently, reads were clustered into 1,896 operational taxonomic units (OTUs). The following alpha diversity metrics were calculated: observed ASVs number (i.e., richness) and Pielou’s evenness index. Weighted Unifrac distances were calculated to compare community structure [20].
2.9. Statistical Analysis
The study described categorical variables by proportions, normally distributed continuous variables by means and SDs, and non-normally distributed continuous variables by medians (1st– 3rd quartile). Changes relative to preintervention values were assessed in both groups separately and together by paired t-tests in normally distributed continuous variables and Wilcoxon signed-rank tests in non-normally distributed variables. Multivariable linear regressions were conducted to explore whether there were differences in the follow-up values in the EBLong group relative to the EBShort group. The models were adjusted for baseline levels of each parameter (continuous), age (continuous), sex, BMI (continuous), and MedDiet adherence(continuous).
We applied a Bonferroni correction to account for the multiple comparisons arising from the three sets of variables (clinical parameters, inflammatory-related biomarkers, and satiety-related biomarkers). Results were considered statistically significant if the p-value was less than 0.05/3 (0.017), ensuring a stringent threshold for significance. Analyses were performed using R Software version 4.3.1 [24].
Metagenome statistical analysis were done using Generalized Linear Mixed Models (GLMM). Alpha diversity comparisons were performed using R package NBZIMM version 1.0 [25] for richness and the R package betareg version 3.1-4 [26] for evenness. Beta diversity distance matrices were used to calculate principal coordinates analysis; the significance of groups in community structure was tested using Permanova. Differential abundance of taxa was tested using Negative Binomial Generalized Linear Mixed Models using the R package NBZIMM [25]. P-values were adjusted using false discovery rate (FDR). Significant threshold was set at 0.05.
3. Results
3.1. Study Population
Participants were recruited between July 2019 and February 2020. A total of 292 individuals were contacted and finally 61 were enrolled (55.7% female) (Figure 1). Of these, 31 were randomly assigned to the EBLong intervention and 30 to the EBShort intervention. Finally, due to the COVID pandemic only 31 participants finished the study resulting in 13 participants in EBShort and 18 in EBLong group. The participants were older adults, with a mean age of 66.7 years, and 51.6% were female. They exhibited a high prevalence of metabolic risk factors. The only variable in which intergroup differences were observed in baseline values was BMI. No difference in diet according to the Adherence to MedDiet questionnaire, kcal intake, and physical activity performance at baseline were found.

Table 1.
Basal characteristics of participants.
| All | EBLong | EBShort | p Value | |
|---|---|---|---|---|
| n | 31 | 18 | 13 | |
| Age, mean (SD) | 66.7 (5.94) | 66.6 (7.04) | 66.8 (4.36) | 0.954 |
| Sex, Female, n (%) | 16 (51.6%) | 8 (44.4%) | 8 (61.5%) | 0.565 |
| Diabetes, n (%) | 26 (83.9%) | 17 (94.4%) | 9 (69.2%) | 0.134 |
| Hypertension, n (%) | 30 (96.8%) | 18 (100%) | 12 (92.3%) | 0.419 |
| Triglycerides, mg/dL, median [1st-3rd quartile] | 142 [90.5;174] | 146 [90.0;168] | 139 [92.0;175] | 0.889 |
| HDLc, mg/dL, mean (SD) | 49.0 (11.7) | 50.3 (11.8) | 47.2 (11.7) | 0.475 |
| BMI, kg/m2, mean (SD) | 32.8 (3.26) | 34.1 (2.84) | 31.2 (3.13) | 0.015 |
| Scholarity | 0.895 | |||
| Elementary School | 13 (43.3%) | 7 (38.9%) | 6 (50.0%) | |
| Middle school | 9 (30.0%) | 6 (33.3%) | 3 (25.0%) | |
| Higher education | 8 (26.7%) | 5 (27.8%) | 3 (25.0%) | |
| Smoking habit | 0.634 | |||
| Non smoker | 13 (41.9%) | 9 (50.0%) | 4 (30.8%) | |
| Smoker | 5 (16.1%) | 2 (11.1%) | 3 (23.1%) | |
| Former smoker | 13 (41.9%) | 7 (38.9%) | 6 (46.2%) | |
| Adherence to MedDiet (14pt), points, mean (SD) | 9.71 (2.18) | 9.83 (2.07) | 9.54 (2.40) | 0.724 |
| Basal intake, kcal, mean (SD) | 1558 (345) | 1552 (387) | 1567 (291) | 0.900 |
| Physical activity, Mets/day, mean (SD) | 2502 (1885) | 2320 (1632) | 2753 (2234) | 0.560 |
EBLong, Elias Boulanger® long fermentation bread; EBShort, Elias Boulanger® long fermentation bread; BMI, body mass index; SD, standard deviation; HDLc, high density lipoprotein cholesterol; MedDiet, Mediterranean Diet.
3.1. Dietetic Assesment
Participants of both groups had similar diets before and during the last week of the study. No differences were found in the quantity of energy or macronutrients. The intake of some of the main nutrients recalled by the three-days food records can be found in Supplemental Table S2.
3.2. Sourdough Bread Intervention
Irrespective of the intervention, after two-months of intervention all participants had a decrease in sICAM and diastolic pressure (p<0.025) (Table 2). Whereas we observed a decrease in sICAM and PAI levels in the EBShort group (Table 2), a decrease of diastolic pressure was determined in EBLong group (p<0.025) (Table 2).
When the two groups were compared, no differences were seen in the follow-up values of these variables in non-adjusted model nor in the adjusted model, except for PAI -1 in EBLong (-744 pg/mL; 95%CI: -282 to -1210 pg/mL) (Table 3).
3.3. Microbiota Characterization
Alpha diversity analysis revealed no discernible differences in richness and evenness either within or between the intervention groups during the follow-up period (Figure 2). Beta diversity, evaluated using weighted UniFrac distance metrics, demonstrated no significant distinctions between the treatment groups throughout the study period (PERMANOVA R2= 0.0072, p = 0.389) (Figure 2).
Figure 3 illustrates the relative frequencies of the most abundant microbial phyla and family. At the phylum level, the relative abundance observed in both intervention groups was as expected for adult human fecal sample. No differences in baseline relative abundance of phyla were observed between the groups. After the intervention with EBLong bread there was a significant reduction in Synergistota abundance relative to baseline (p < 0.001). Additionally, when comparing the two groups at the follow-up assessment, the EBLong group exhibited lower abundance of Synergistota compared to the EBShort group, however its abundance did not exceed 1% (see Table 3). At family level, after the two-month intervention, the EBLong group showed a reduction in Enterobacteriaceae and Synergistaceae. In contrast, the EBShort group exhibited a decrease in Oscillospiraceae and an increase in Prevotellaceae. At follow-up, the abundance of Oscillospiraceae differed significantly between the groups; however, this difference was also observed at baseline.
4. Discussion
A two-months intervention with two sourdough bread, with similar nutrient composition but different fermentation time, decreased sICAM and diastolic pressure levels in all the participants. In the assessment of each bread, we observed a decrease in sICAM levels in the consumers of EBShort bread, relative to baseline values. When comparing the effects of each bread, PAI-1 levels were lower after the intervention with EBShort bread compared to EBLong bread. No changes were observed in any other clinical parameter or satiety-related biomarker. In the microbiota analysis, we found comparable composition between groups after the intervention, with no changes in the diversity and abundance, suggesting that gut microbial communities remain stable after both interventions. Nevertheless, at the phylum level, the abundance of Synergistota exhibited a reduction following the intervention with EBLong. At the family level, the intervention groups differed in the abundance of Oscillospiraceae at follow-up; however, these differences were already present at baseline.
Regarding inflammatory biomarkers, we have not found any difference in cytokines, similar to what has been reported in a previous article [27]. Nevertheless, after two-months of swapping their habitual bread for sourdough bread (either with short or long fermentation) participants had lower values of sICAM, an adhesion molecule that plays an important role in the development of atherosclerosis [28]. Comparable outcomes were found in a study with a 5-weeks intervention with a prebiotic antioxidant bread composed by wheat–rye bread, tomato paste, green tea powder and herbs [29]. In the previous study by Seidel et al., ICAM-1 significantly decreased after the intervention in non-smoker adults. Despite this bread not containing sourdough, it was enriched similarly to our starter, which included infusions, dairy products, and fruit resulting in a bread rich in flavonoids, pre and probiotics that could provide this effect. An in vitro study suggests that some flavonoids could inhibit ICAM-1 expression [30] and a clinical trial showed a decrease in ICAM-1 expression after the consumption of a probiotic sausage containing Lactobacillus paracasei [31]. Because of the limited number of participants involved in this study, besides the variability of flavonoids and probiotics in diet, more evidence is required to draw any conclusions regarding any mechanism associated with this observation. PAI-1 is a primary inhibitor of fibrinolysis, playing a critical role in the development of thrombosis, atherosclerosis, and cardiovascular risk [32]. Contrary to our hypothesis that bread with a higher sourdough proportion would reduce inflammation, PAI-1 levels were lower in the EBShort group compared to EBLong intervention at follow-up. Since our objective was to compare different sourdough breads, distinguishing between interventions is challenging without a poor-quality control bread for reference. Other authors, that compared the consumption of whole grain wheat sourdough bread to refined white bread in normoglycemic-normoinsulinemic and hyperglycemic-hyperinsulinemic patients observed no differences in PAI-1 levels after 6 weeks of consumption [33]. Different sample size of our intervention groups, the greater proportion of diabetics, differences in BMI (being greater in EBLong) and different breads may explain discrepancies in the findings presented here.
Regarding clinical parameters, after consuming for two-months sourdough bread there was a decrease in diastolic pressure. In contrast, another clinical trial comparing the effect of whole grain wheat sourdough to non-fermented white bread did not find differences in blood pressure after 6 weeks of treatment [34]. In vitro studies, have described an anti-hypertensive effect in spelt flours with 96 hours of fermentation [35], and quinoa and wheat flours with 46 hours of fermentation [36]. The authors reported that mechanism underlaying may be the release during fermentation of bioactive peptides involved in the inhibition of the Angiotensin-Converting Enzyme (ACE) activity, a molecule that plays a crucial role in the control of blood pressure with a vasoconstrictor effect [35,36]. Different conditions such as the types of flour, time, and microorganisms engaged in the fermentation process could have different impacts on the results. Future studies could assess polymorphisms in the ACE gene associated with hypertension to tailor recommendations for sourdough bread consumption. Despite minor variations in blood pressure, we did not observe any additional differences in lipid or glucose metabolism following the 2-month intervention. Similar outcomes were found in a previous clinical trial, which compared consumption of whole grain wheat sourdough bread and white bread over 6-weeks. Notably, no differences in serum lipids or glycemic parameters were observed in adults with normoglycemia nor hyperglycemia [34]. On the other hand, another clinical trial with healthy young adults found that after an intervention of 4 weeks with an ancient grain “Verna” bread (leavened with sourdough or yeast), LDL cholesterol decreased. Fasting glucose was higher in the group eating yeast-fermented bread, but when comparing both types of bread, there were no differences in lipids or glucose levels, and sourdough bread didn’t offer any extra benefits [27]. Although there’s no evidence that sourdough affects long-term glucose control, a meta-analysis suggested it might help lower post-prandial blood sugar compared to other breads [37].
As far as we know, no other study has examined long-term interventions of sourdough bread on satiety, although a few studies have looked for its acute effects. When appetite was assessed using subjective scales, the evidence was inconclusive [38,39,40]. However, when satiety was tested with an ad libitum meal after the consumption of different types of bread, sourdough breads did not seem to influence energy intake [38,39]. Some studies have specifically measured incretins. One study found that the postprandial concentrations of gastric inhibitory peptide (GIP) and GLP-1 were lower after consuming sourdough bread compared to whole wheat and whole wheat barley breads [41]. Another study described a lower ghrelin AUC after the consuming einkorn sourdough bread compared to commercial breads. However, there were no differences among the consumption of different sourdough breads [42]. The differences between these studies may be attributed to various factors, such as the use of different measurement scales, variations in control breads, and differences in the duration of the studies.
Regarding microbiota, the lack of changes in alpha diversity, which indicates species richness and diversity within individual samples, suggested consistent levels of microbial diversity regardless of sourdough bread consumption and its fermentation duration. Likewise, the consistency in beta diversity, which measures the differences in microbial community composition between different samples, suggests that there were no changes in microbial composition across the interventions. These results are consistent with a similar trial conducted in subjects diagnosed with ulcerative colitis, which examined the effects of a two-month intervention with two different sourdough breads (differing in percentage) [43]. Consistent with our results, the above-mentioned study with 23 subjects reported no differences microbiota diversity. Furthermore, although the bread treatment suggestively reduced the Firmicutes/Bacteroidetes ratio, both groups experienced some relief from symptoms resembling intestinal bowel disease [43]. Likewise, a one-week clinical trial involving 20 healthy subjects that compared sourdough bread consumption to white bread found similar results, with no noticeable differences in alpha and beta diversity or relative abundances at the phylum level [44]. Despite not finding differences in diversity, we have identified some differences in the abundances of the most prevalent phylum and families. We observed a reduction in the abundance of the Synergistota phylum following sourdough bread consumption, although its abundance did not surpass 1%. Although this phylum is poorly characterized, it is known to inhabit human soft tissues and the gut, and its main function is their role in amino acid degradation [45]. Interestingly, at family level, after the consumption of EBLong, the abundance of Enterobacteriaceae decrease. This family thrive in inflamed environments, commonly associated with conditions such as inflammatory bowel disease, obesity, colorectal cancer, celiac disease, and after antibiotic use [46]. EBLong may contribute to reducing the conditions for Enterobacteriaceae proliferation, consequently lowering their abundance. Further investigations could explore these alterations and their potential effects on gastrointestinal symptoms following sourdough consumption, as previously reported. For EBShort, we observed a decrease in the abundance of the Oscillospiraceae family. Within this family, the genus Oscillospira is the most studied and is associated with leanness, metabolic health, and reduced inflammation [47]. Additionally, we found an increase in the Prevotellaceae family, which has been described as more abundant in individuals with obesity [48]. It is important to note that our analysis did not extend to the genus level, making it difficult to draw specific conclusions about the implications of these changes. Furthermore, the weight-related effects described for these families are not reflected in the clinical data from our study. These collective findings suggest that the intestinal microbiota exhibits resilience to changes induced by bread consumption, particularly in the case of sourdough bread.
Our study has some limitations. First, due to the health contingency caused by the Covid-19 pandemic, the clinical trial was interrupted, affecting the disparity between the intervention groups in number and characteristics, especially in BMI. Nevertheless, we tried to minimize it by adjusting covariates (for age, sex, BMI, MedDiet adherence (14pt) and baseline values). Second, breads composition was very similar between them, both of them contained sourdough which could explain the findings, and the nutritional composition in terms of energy, carbohydrate, total fat, protein, fiber, and sodium was very similar. Third, intake of bread and nutritional assessment were auto-reported, which could imply some self-report bias. Fourth, due to the high number of determinations of the study, and even though we corrected our results for multiple testing, our results should be interpreted cautiously. Finally, we conducted the study in a very specific population, which could complicate the generalization of the results.
5. Conclusions
Sourdough bread may offer some mild benefits in blood pressure and inflammation markers in individuals with metabolic syndrome. Gut microbiota, however, did not exhibit differences when comparing both interventions, suggesting that it remains stable to changes in sourdough consumption. Apart from a higher abundance of Enterobacteriaceae in the group that consumed EBL, no further modifications were observed with a bread with longer fermentation time. More studies with larger samples comparing different fermented sourdough breads and control breads, are needed in order to verify our results and to fully understand the potential benefits and mechanisms of action of sourdough bread.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Supplemental Table S1: Nutritional composition of EBLong and EBshort breads; Supplemental Table S2: Difference in dietetic assessment of food records in EBShort and EBLong groups; Supplemental Table S3: Differential abundance at phylum taxonomic level.
Author Contributions
Conceptualization, María-Dolores Zomeño, Montserrat Fitó and Olga Castañer; Data curation, Karla Alejandra Pérez-Vega and Álvaro Hernáez; Formal analysis, Karla Alejandra Pérez-Vega, Nerea Carrón and Pedro González-Torres; Funding acquisition, Marc Llirós, Isidre Elias, Xavier Aldeguer and Montserrat Fitó; Investigation, Karla Alejandra Pérez-Vega, Albert Sanllorente, María-Dolores Zomeño, Ana Quindós, Júlia Muñoz-Martínez, Daniel Muñoz, Sònia Gaixas and Gemma Blanchart; Methodology, María-Dolores Zomeño, Daniel Muñoz, Montserrat Fitó and Olga Castañer; Project administration, Montserrat Fitó; Resources, Ana Aldea-Perona, Isidre Elias, Núria Elias-Masiques and Montserrat Fitó; Supervision, Álvaro Hernáez and Montserrat Fitó; Validation, Montserrat Fitó; Visualization, Montserrat Fitó and Olga Castañer; Writing—original draft, Karla Alejandra Pérez-Vega and Montserrat Fitó; Writing—review & editing, Albert Sanllorente, María-Dolores Zomeño, Ana Quindós, Júlia Muñoz-Martínez, Mireia Malcampo, Ana Aldea-Perona, Álvaro Hernáez, Aleix Lluansí, Marc Llirós, Isidre Elias, Núria Elias-Masiques, Xavier Aldeguer, Helmut Schröder, Javier Hernando-Redondo, Nerea Carrón, Pedro González-Torres, Valentini Konstantinidou and Olga Castañer. All authors have read and agreed to the published version of the manuscript.
Funding
Projects IFI20/00002, IFI16/00011, PI18/00020, PI20/00012, PI17/00214, CP21/00097 funded by Instituto de Salud Carlos III (ISCIII) and co-funded by the European Union; Ministry of Economy, Industry and Competitiveness (MINECO) RETOS program (RTC-2017-6467-2); and by Agència de Gestió d’Ajuts Universitaris i de Recerca (2021 SGR 00144).
Data Availability Statement
The generation and analysis of the data sets within this study are not projected to be open to access beyond the core research group. This is because the participants’ consent forms and ethical approval did not include provisions for public accessibility. However, we follow a controlled data-sharing collaboration model, as the informed consent documents signed by the participants allowed for regulated collaboration with other researchers for study-related research. The data described in the manuscript, alongside the codebook and analytic code, will be available upon request. Researchers interested in this study can reach out to corresponding author Montse Fito (mfito@researchmar.net).
Acknowledgments
The authors want to thank all the study participants. CIBER de Fisiopatología de la Obesidad y Nutrición (CIBEROBN) and CIBER de Epidemiología y Salud Publica; initiatives of Instituto de Salud Carlos III (Madrid, Spain), and financed by FEDER funds (CB06/03).
Conflicts of Interest
I.E. and N.EM. were employees of Elias–Boulanger who have received funding from RTC-2017-6467-2 program. They had no role in the collection, analyses, and interpretation of data; or in the decision to publish the results. The rest of the authors declare no conflict of interest.
References
- Shin, J.A.; Lee, J.H.; Lim, S.Y.; Ha, H.S.; Kwon, H.S.; Park, Y.M.; Lee, W.C.; Kang, M. Il; Yim, H.W.; Yoon, K.H.; et al. Metabolic Syndrome as a Predictor of Type 2 Diabetes, and Its Clinical Interpretations and Usefulness. J Diabetes Investig 2013, 4, 334–343. [CrossRef]
- Galassi, A.; Reynolds, K.; He, J. Metabolic Syndrome and Risk of Cardiovascular Disease: A Meta-Analysis. Am J Med 2006, 119, 812–819. [CrossRef]
- Alberti, K.G.M.M.; Eckel, R.H.; Grundy, S.M.; Zimmet, P.Z.; Cleeman, J.I.; Donato, K.A.; Fruchart, J.C.; James, W.P.T.; Loria, C.M.; Smith, S.C. Harmonizing the Metabolic Syndrome: A Joint Interim Statement of the International Diabetes Federation Task Force on Epidemiology and Prevention; National Heart, Lung, and Blood Institute; American Heart Association; World Heart Federation; International Atherosclerosis Society; And International Association for the Study of Obesity. Circulation 2009, 120, 1640–1645. [CrossRef]
- Rippe, J.M. Lifestyle Strategies for Risk Factor Reduction, Prevention, and Treatment of Cardiovascular Disease. Am J Lifestyle Med 2019, 13, 204–212. [CrossRef]
- Estruch, R.; Ros, E.; Salas-Salvadó, J.; Covas, M.-I.; Corella, D.; Arós, F.; Gómez-Gracia, E.; Ruiz-Gutiérrez, V.; Fiol, M.; Lapetra, J.; et al. Primary Prevention of Cardiovascular Disease with a Mediterranean Diet Supplemented with Extra-Virgin Olive Oil or Nuts. N Engl J Med. 2018, 378, e34. [CrossRef]
- Esposito, K.; Kastorini, C.M.; Panagiotakos, D.B.; Giugliano, D. Mediterranean Diet and Metabolic Syndrome: An Updated Systematic Review. Rev Endocr Metab Disord 2013, 14, 255–263. [CrossRef]
- Capurso, A.; Capurso, C. The Mediterranean Way: Why Elderly People Should Eat Wholewheat Sourdough Bread—a Little Known Component of the Mediterranean Diet and Healthy Food for Elderly Adults. Aging Clin Exp Res 2020, 32, 1–5. [CrossRef]
- Arendt, E.K.; Ryan, L.A.M.; Dal Bello, F. Impact of Sourdough on the Texture of Bread. Food Microbiol 2007, 24, 165–174. [CrossRef]
- Olagnero, G.; Abad, A.; Bendersky, S.; Genevois, C.; Granzella, L.; Montonati, M. Alimentos Funcionales: Fibra, Prebióticos, Probióticos y Simbióticos. Diaeta 2007, 25, 20–33.
- Akamine, I.T.; Mansoldo, F.R.P.; Vermelho, A.B. Probiotics in the Sourdough Bread Fermentation: Current Status. Fermentation 2023, 9, 90. [CrossRef]
- Lluansí, A.; Llirós, M.; Oliver, L.; Bahí, A.; Elias-Masiques, N.; Gonzalez, M.; Benejam, P.; Cueva, E.; Termes, M.; Ramió-Pujol, S.; et al. In Vitro Prebiotic Effect of Bread-Making Process in Inflammatory Bowel Disease Microbiome. Front Microbiol 2021, 12, 716307. [CrossRef]
- Schröder, H.; Fitó, M.; Estruch, R.; Martínez-González, M.A.; Corella, D.; Salas-Salvadó, J.; Lamuela-Raventós, R.; Ros, E.; Salaverría, I.; Fiol, M.; et al. A Short Screener Is Valid for Assessing Mediterranean Diet Adherence among Older Spanish Men and Women. J Nutr. 2011, 141, 1140–1145. [CrossRef]
- Cantós López, D.; Farran, A.; Palma Linares, I. Programa de Càlcul Nutricional Professional (PCN Pro) 2013.
- Molina, L.; Sarmiento, M.; Peñafiel, J.; Donaire, D.; Garcia-Aymerich, J.; Gomez, M.; Ble, M.; Ruiz, S.; Frances, A.; Schröder, H.; et al. Validation of the Regicor Short Physical Activity Questionnaire for the Adult Population. PLoS One 2017, 12, e0168148. [CrossRef]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glöckner, F.O. Evaluation of General 16S Ribosomal RNA Gene PCR Primers for Classical and Next-Generation Sequencing-Based Diversity Studies. Nucleic Acids Res 2013, 41, e1. [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, Interactive, Scalable and Extensible Microbiome Data Science Using QIIME 2. Nat Biotechnol 2019, 37, 852–857. [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-Resolution Sample Inference from Illumina Amplicon Data. Nat Methods 2016, 13, 581–583. [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol Biol Evol 2013, 30, 772–780. [CrossRef]
- Price, M.N.; Dehal, P.S.; Arkin, A.P. FastTree 2—Approximately Maximum-Likelihood Trees for Large Alignments. PLoS One 2010, 5. [CrossRef]
- Lozupone, C.; Lladser, M.E.; Knights, D.; Stombaugh, J.; Knight, R. UniFrac: An Effective Distance Metric for Microbial Community Comparison. ISME J 5 2011, 5, 169–172. [CrossRef]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naïve Bayesian Classifier for Rapid Assignment of RRNA Sequences into the New Bacterial Taxonomy. Appl Environ Microbiol 2007, 73, 5261–5267. [CrossRef]
- Pruesse, E.; Quast, C.; Knittel, K.; Fuchs, B.M.; Ludwig, W.; Peplies, J.; Glöckner, F.O. SILVA: A Comprehensive Online Resource for Quality Checked and Aligned Ribosomal RNA Sequence Data Compatible with ARB. Nucleic Acids Res 2007, 35, 7188–7196. [CrossRef]
- Pedregosa, F.; Varoquaux, G.; Gramfort, A.; Michel, V.; Thirion, B.; Grisel, O.; Blondel, M.; Prettenhofer, P.; Weiss, R.; Dubourg, V.; et al. Scikit-Learn: Machine Learning in Python. Journal of Machine Learning Research 2011, 12, 2825–2830.
- R Core Team R: A Language and Environment for Statistical Computing.
- Zhang, X.; Yi, N. NBZIMM: Negative Binomial and Zero-Inflated Mixed Models, with Application to Microbiome/Metagenomics Data Analysis. BMC Bioinformatics 2020, 21, 488. [CrossRef]
- Cribari-Neto, F.; Zeileis, A. Beta Regression in R. J Stat Softw 2010, 34, 1–24. [CrossRef]
- Pagliai, G.; Venturi, M.; Dinu, M.; Galli, V.; Colombini, B.; Giangrandi, I.; Maggini, N.; Sofi, F.; Granchi, L. Effect of Consumption of Ancient Grain Bread Leavened with Sourdough or with Baker’s Yeast on Cardio-Metabolic Risk Parameters: A Dietary Intervention Trial. Int J Food Sci Nutr 2021, 72, 367–374. [CrossRef]
- Singh, V.; Kaur, R.; Kumari, P.; Pasricha, C.; Singh, R. ICAM-1 and VCAM-1: Gatekeepers in Various Inflammatory and Cardiovascular Disorders. Clinica Chimica Acta 2023, 548, 117487. [CrossRef]
- Seidel, C.; Boehm, V.; Vogelsang, H.; Wagner, A.; Persin, C.; Glei, M.; Pool-Zobel, B.L.; Jahreis, G. Influence of Prebiotics and Antioxidants in Bread on the Immune System, Antioxidative Status and Antioxidative Capacity in Male Smokers and Non-Smokers. Br J Nutr 2007, 97, 349–356. [CrossRef]
- Chen, C.C.; Chow, M.P.; Huang, W.C.; Lin, Y.C.; Chang, Y.J. Flavonoids Inhibit Tumor Necrosis Factor-α-Induced up-Regulation of Intercellular Adhesion Molecule-1 (ICAM-1) in Respiratory Epithelial Cells through Activator Protein-1 and Nuclear Factor-ΚB: Structure-Activity Relationships. Mol Pharmacol 2004, 66, 683–693.
- Jahreis, G.; Vogelsang, H.; Kiessling, G.; Schubert, R.; Bunte, C.; Hammes, W.P. Influence of Probiotic Sausage (Lactobacillus Paracasei) on Blood Lipids and Immunological Parameters of Healthy Volunteers. Food Res Int 2002, 35, 133–138. [CrossRef]
- Tjärnlund-Wolf, A.; Brogren, H.; Lo, E.H.; Wang, X. Plasminogen Activator Inhibitor-1 and Thrombotic Cerebrovascular Diseases. Stroke 2012, 43, 2833–2839. [CrossRef]
- MacKay, K.A.; Tucker, A.J.; Duncan, A.M.; Graham, T.E.; Robinson, L.E. Whole Grain Wheat Sourdough Bread Does Not Affect Plasminogen Activator Inhibitor-1 in Adults with Normal or Impaired Carbohydrate Metabolism. Nutr Metab Cardiovasc Dis 2012, 22, 704–711. [CrossRef]
- Tucker, A.J.; MacKay, K.A.; Robinson, L.E.; Graham, T.E.; Bakovic, M.; Duncan, A.M. The Effect of Whole Grain Wheat Sourdough Bread Consumption on Serum Lipids in Healthy Normoglycemic/Normoinsulinemic and Hyperglycemic/Hyperinsulinemic Adults Depends on Presence of the APOE E3/E3 Genotype: A Randomized Controlled Trial. Nutr Metab 2010, 7, 37. [CrossRef]
- Gabriele, M.; Arouna, N.; Árvay, J.; Longo, V.; Pucci, L. Sourdough Fermentation Improves the Antioxidant, Antihypertensive, and Anti-Inflammatory Properties of Triticum Dicoccum. Int J Mol Sci 2023, 24, 6283. [CrossRef]
- Ayyash, M.; Johnson, S.K.; Liu, S.Q.; Mesmari, N.; Dahmani, S.; Al Dhaheri, A.S.; Kizhakkayil, J. In Vitro Investigation of Bioactivities of Solid-State Fermented Lupin, Quinoa and Wheat Using Lactobacillus Spp. Food Chem 2019, 275, 50–58. [CrossRef]
- Rolim, M.E.; Fortes, M.I.; Von Frankenberg, A.; Duarte, C.K. Consumption of Sourdough Bread and Changes in the Glycemic Control and Satiety: A Systematic Review. Crit Rev Food Sci Nutr 2022, 801–816. [CrossRef]
- Iversen, K.N.; Johansson, D.; Brunius, C.; Andlid, T.; Andersson, R.; Langton, M.; Landberg, R. Appetite and Subsequent Food Intake Were Unaffected by the Amount of Sourdough and Rye in Soft Bread—A Randomized Cross-over Breakfast Study. Nutrients 2018, 10, 1594. [CrossRef]
- Darzi, J.; Frost, G.S.; Robertson, M.D. Effects of a Novel Propionate-Rich Sourdough Bread on Appetite and Food Intake. Eur J Clin Nutr 2012, 66, 789–794. [CrossRef]
- Zamaratskaia, G.; Johansson, D.P.; Junqueira, M.A.; Deissler, L.; Langton, M.; Hellström, P.M.; Landberg, R. Impact of Sourdough Fermentation on Appetite and Postprandial Metabolic Responses-A Randomised Cross-over Trial with Whole Grain Rye Crispbread. Br J Nutr 2017, 118, 686–697. [CrossRef]
- Najjar, A.M.; Parsons, P.M.; Duncan, A.M.; Robinson, L.E.; Yada, R.Y.; Graham, T.E. The Acute Impact of Ingestion of Breads of Varying Composition on Blood Glucose, Insulin and Incretins Following First and Second Meals. Br J Nutr 2009, 101, 391–398. [CrossRef]
- Bo, S.; Seletto, M.; Choc, A.; Ponzo, V.; Lezo, A.; Demagistris, A.; Evangelista, A.; Ciccone, G.; Bertolino, M.; Cassader, M.; et al. The Acute Impact of the Intake of Four Types of Bread on Satiety and Blood Concentrations of Glucose, Insulin, Free Fatty Acids, Triglyceride and Acylated Ghrelin. A Randomized Controlled Cross-over Trial. Food Res Int 2017, 92, 40–47. [CrossRef]
- Lluansí, A.; Llirós, M.; Carreras-Torres, R.; Bahí, A.; Capdevila, M.; Feliu, A.; Vilà-Quintana, L.; Elias-Masiques, N.; Cueva, E.; Peries, L.; et al. Impact of Bread Diet on Intestinal Dysbiosis and Irritable Bowel Syndrome Symptoms in Quiescent Ulcerative Colitis: A Pilot Study. PLoS One 2024, 19, e0297836. [CrossRef]
- Korem, T.; Zeevi, D.; Zmora, N.; Weissbrod, O.; Bar, N.; Lotan-Pompan, M.; Avnit-Sagi, T.; Kosower, N.; Malka, G.; Rein, M.; et al. Bread Affects Clinical Parameters and Induces Gut Microbiome-Associated Personal Glycemic Responses. Cell Metab 2017, 25, 1243-1253.e5. [CrossRef]
- Marchandin, H.; Damay, A.; Roudière, L.; Teyssier, C.; Zorgniotti, I.; Dechaud, H.; Jean-Pierre, H.; Jumas-Bilak, E. Phylogeny, Diversity and Host Specialization in the Phylum Synergistetes with Emphasis on Strains and Clones of Human Origin. Res Microbiol 2010, 161. [CrossRef]
- Zeng, M.Y.; Inohara, N.; Nuñez, G. Mechanisms of Inflammation-Driven Bacterial Dysbiosis in the Gut. Mucosal Immunol 2017, 10. [CrossRef]
- Konikoff, T.; Gophna, U. Oscillospira: A Central, Enigmatic Component of the Human Gut Microbiota. Trends Microbiol 2016, 24. [CrossRef]
- Duan, M.; Wang, Y.; Zhang, Q.; Zou, R.; Guo, M.; Zheng, H. Characteristics of Gut Microbiota in People with Obesity. PLoS One 2021, 16. [CrossRef]
Figure 1.
Study flowchart.
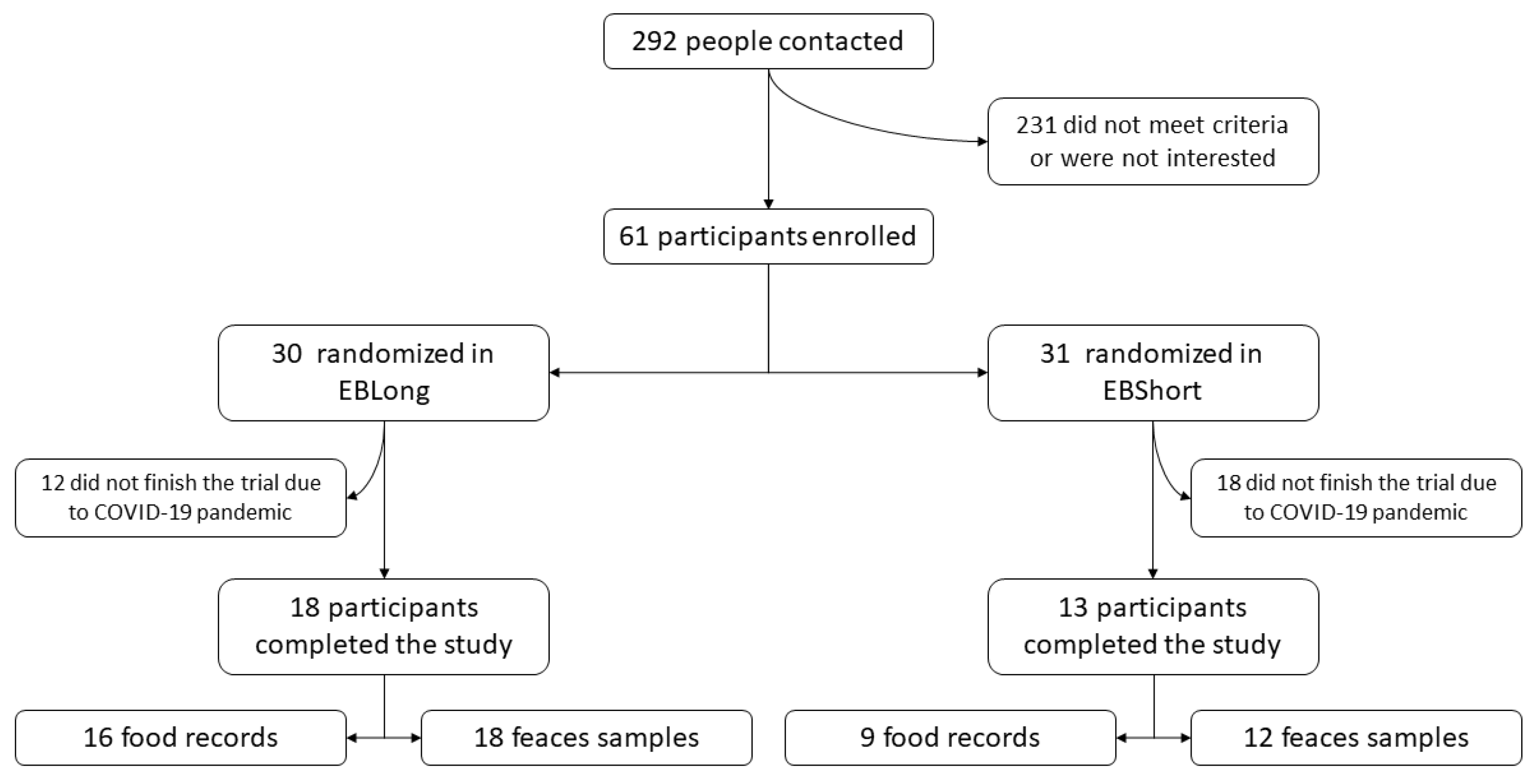
Figure 2.
Alpha and beta diversity of EBLong and EBShort intervention. (a) Diversity richness as the number of observed OTUs at baseline and at follow up in each intervention. (b) Pielou’s evenness index in each intervention groups at baseline and at follow up. (c) Principal coordinates analysis (PCoA) plot based on weighted UniFrac distances according to interventions and follow up.
Figure 2.
Alpha and beta diversity of EBLong and EBShort intervention. (a) Diversity richness as the number of observed OTUs at baseline and at follow up in each intervention. (b) Pielou’s evenness index in each intervention groups at baseline and at follow up. (c) Principal coordinates analysis (PCoA) plot based on weighted UniFrac distances according to interventions and follow up.
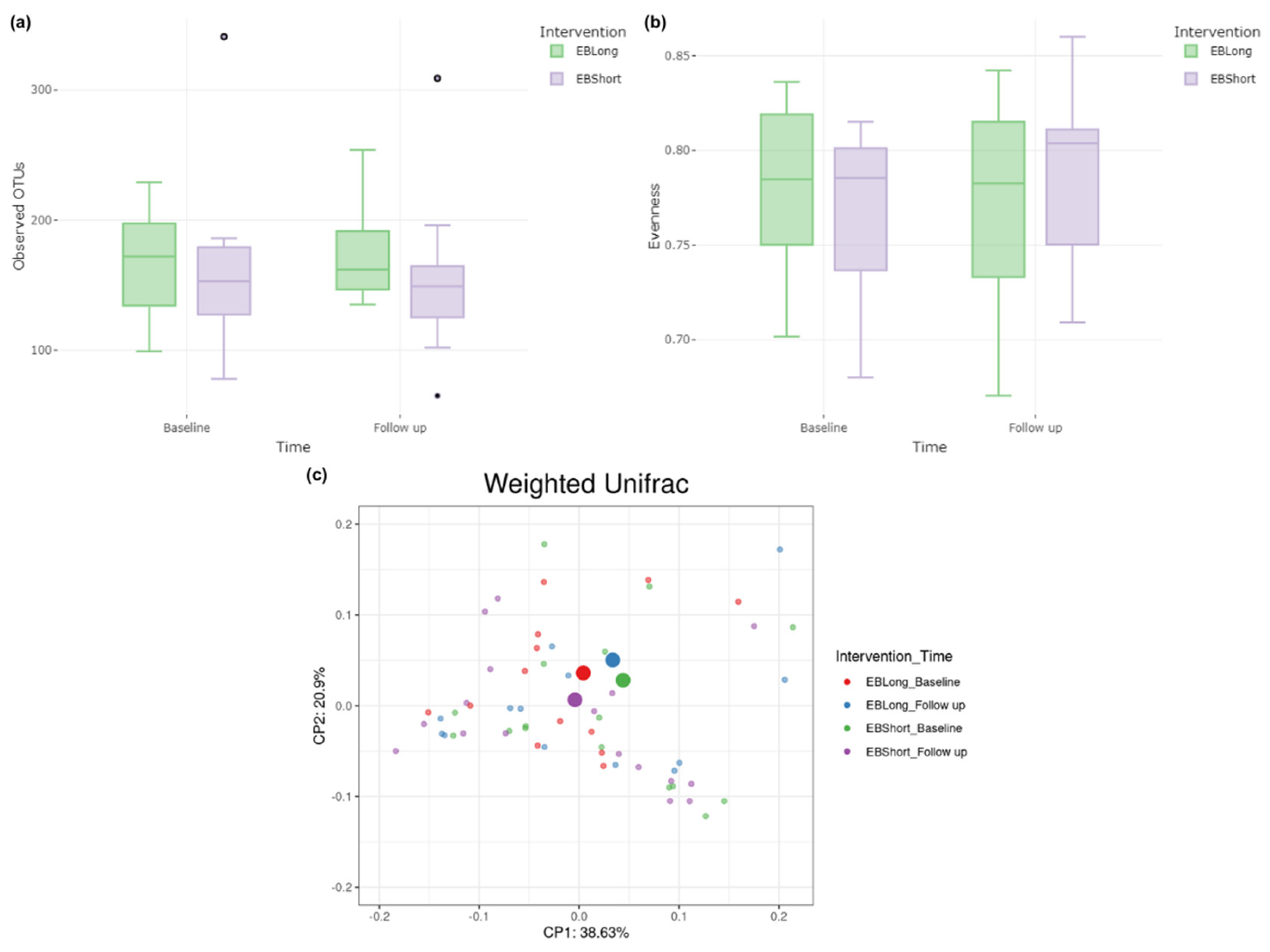
Figure 3.
Barplots for relative abundance. (a) Displays abundance at phylum level for each intervention. Phyla with a relative abundance lower than 1% grouped as “Others”. (b) Displays abundance at family level of the 10 most abundant taxa.
Figure 3.
Barplots for relative abundance. (a) Displays abundance at phylum level for each intervention. Phyla with a relative abundance lower than 1% grouped as “Others”. (b) Displays abundance at family level of the 10 most abundant taxa.
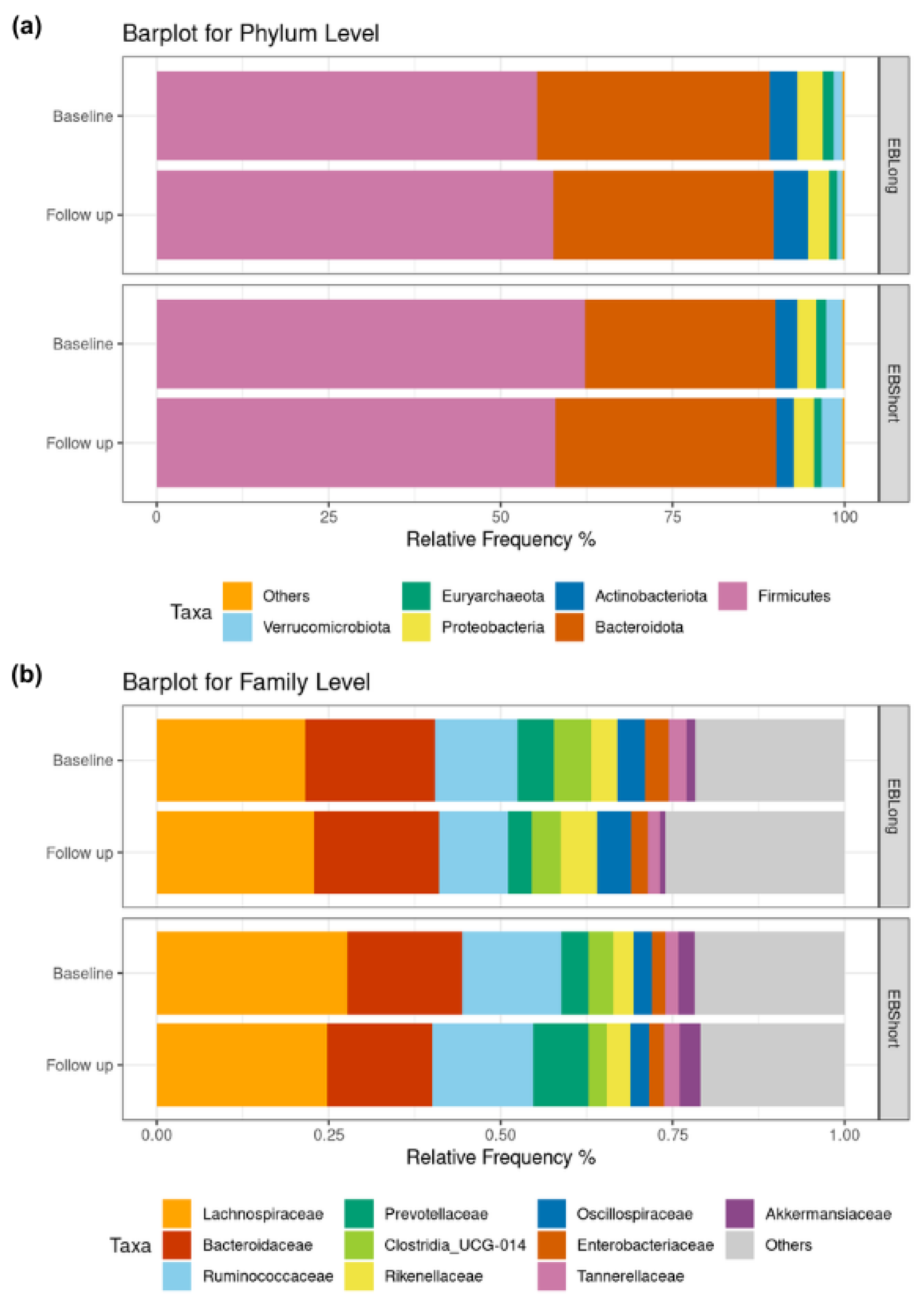
Table 2.
Intragroup differences between baseline and two-months follow-up.
| EBLong | EBShort | All | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Baseline | Follow up | p Value | Baseline | Follow up | p Value | Baseline | Follow up | p Value | |
| Weight, kg | 92.9 (14.7) | 93.4 (14.3) | 0.710 | 84 (10.2) | 83.5 (9.5) | 0.415 | 89.1 (13.5) | 89.1 (13.2) | 0.462 |
| Waist, cm | 119 (17.3) | 114 (12.5) | 0.237 | 111 (10.3) | 110 (9.18) | 0.796 | 115 (15.1) | 112 (11.2) | 0.223 |
| Systolic pressure, mmHg | 136 (11.3) | 132 (14.5) | 0.470 | 134 (10.1) | 135 (9.91) | 0.395 | 135 (10.7) | 134 (12.6) | 0.830 |
| Diastolic pressure, mmHg | 80.2 (12.2) | 72.5 (10.2) | 0.020 | 77.6 (12.7) | 72.7 (11.3) | 0.208 | 79.1 (12.2) | 72.6 (10.5) | 0.008 |
| Glucose, mg/dL | 125 (30.7) | 128 (33.2) | 0.162 | 117 (23.8) | 117 (21.9) | 0.967 | 122 (27.8) | 124 (29.2) | 0.318 |
| Insulin, pg/mL | 423 (201) | 388 (168) | 0.067 | 484 (282) | 490 (287) | 0.797 | 449 (236) | 431 (227) | 0.241 |
| Glucagon, pg/mL | 520 (188) | 493 (177) | 0.143 | 541 (117) | 539 (180) | 0.949 | 529 (160) | 512 (177) | 0.376 |
| Homa Index | 18.2 (7.97) | 17.3 (7.43) | 0.286 | 21.2 (15.6) | 21 (13.8) | 0.890 | 19.5 (11.7) | 18.8 (10.5) | 0.431 |
| Triglycerides, mg/dL | 146 [90; 168] | 130 [91; 158] | 0.862 | 139 [92; 175] | 124 [84; 179] | 0.839 | 139 [92; 175] | 124 [84; 179] | 0.814 |
| Total cholesterol, mg/dL | 199 (39.5) | 202 (38.6) | 0.378 | 189 (38.9) | 192 (56.3) | 0.707 | 195 (39) | 198 (46.3) | 0.444 |
| HDLc, mg/dL | 50.3 (11.8) | 50.6 (11.7) | 0.736 | 47.2 (11.7) | 48.7 (15.4) | 0.263 | 49 (11.7) | 49.8 (13.1) | 0.282 |
| LDLc, mg/dL | 120 (28.6) | 125 (35.1) | 0.123 | 115 (33.1) | 115 (44.2) | 0.918 | 118 (30.2) | 121 (38.8) | 0.314 |
| C-peptide, pg/mL | 1100 (423) | 1050 (358) | 0.389 | 1190 (511) | 1190 (613) | 0.940 | 1140 (457) | 1110 (481) | 0.618 |
| Ghrelin, pg/mL | 902 (297) | 904 (274) | 0.952 | 1180 (777) | 1130 (622) | 0.444 | 1020 (558) | 998 (458) | 0.531 |
| Leptin, pg/mL | 8920 (5110) | 8540 (5430) | 0.434 | 9170 (4680) | 9120 (4910) | 0.897 | 9020 (4850) | 8780 (5140) | 0.451 |
| GLP1, pg/mL | 164 (97.8) | 165 (111) | 0.960 | 187 (125) | 223 (122) | 0.191 | 174 (109) | 189 (118) | 0.304 |
| IL6, pg/mL | 2.4 (1.73) | 3.06 (1.93) | 0.106 | 2.5 (1.5) | 2.14 (1.08) | 0.350 | 2.44 (1.62) | 2.67 (1.67) | 0.426 |
| IL8, pg/mL | 4.49 (2.21) | 3.86 (2.02) | 0.116 | 4.63 (2.78) | 4.4 (2.04) | 0.563 | 4.55 (2.42) | 4.09 (2.02) | 0.099 |
| Resistin, pg/mL | 4320 (1720) | 4360 (1310) | 0.883 | 6260 (3020) | 5630 (2240) | 0.339 | 5130 (2510) | 4890 (1840) | 0.445 |
| TNF a, pg/mL | 29.6 (9.59) | 29.9 (11.8) | 0.898 | 40.7 (15.4) | 35.5 (12.4) | 0.032 | 34.2 (13.3) | 32.2 (12.2) | 0.246 |
| PAI-1, pg/mL | 2740 (1070) | 2840 (999) | 0.466 | 2750 (529) | 2330 (773) | 0.018 | 2740 (872) | 2630 (933) | 0.318 |
| Visfatin, pg/mL | 1910 (1310) | 1730 (1380) | 0.133 | 2030 (1440) | 1990 (1400) | 0.887 | 1960 (1340) | 1840 (1370) | 0.364 |
| sICAM, pg/mL | 179000 (67500) | 170000 (41800) | 0.325 | 192000 (59300) | 160000 (39200) | 0.013 | 184000 (63500) | 166000 (40300) | 0.014 |
| LBP, ng/mL | 15100 (2630) | 16500 (4370) | 0.095 | 14200 (3820) | 13900 (3690) | 0.761 | 14700 (3160) | 15400 (4230) | 0.259 |
Baseline and follow up values are presented as mean (SD) or median [1st-3rd quartile]. EBLong, Elias Boulanger® long fermentation bread; EBShort, Elias Boulanger® short fermentation bread; HDLc, high density lipoprotein cholesterol; LDLc, low density lipoprotein cholesterol; GLP-1, glycoprotein 1; IL6, interleukin 6; Il8, interleukin 8; TNF-a, tumor necrosis factor alpha; PAI-1, plasminogen activator inhibitor-1; sICAM, soluble intercellular adhesion molecule; LBP, lipopolysaccharide binding protein.
Table 3.
Intergroup differences between follow-up values.
| EBLong vs EBShort | ||||
|---|---|---|---|---|
| Non-Adjusted $(diff. [95% CI]) | p Value | Adjusted $(diff. [95% CI]) | p Value | |
| Weight, kg | 9.82 [0.47; 19.2] | 0.050 | -0.2 [-2.03; 1.62] | 0.829 |
| Waist, cm | 4.49 [-3.89; 12.9] | 0.303 | -4.46 [-9.22; 0.3] | 0.082 |
| Systolic pressure, mmHg | -3.16 [-12.7; 6.38] | 0.522 | -11.6 [-21.1; -2.12] | 0.026 |
| Diastolic pressure, mmHg | -0.18 [-8.17; 7.81] | 0.966 | -6.43 [-14.6; 1.76] | 0.140 |
| Glucose, mg/dL | 11.3 [-9.48; 32] | 0.296 | 5.71 [-4.76; 16.2] | 0.296 |
| Insulin, pg/mL | -102 [-262; 58.8] | 0.224 | -22 [-91.9; 47.9] | 0.543 |
| Glucagon, pg/mL | -46.3 [-173; 80.7] | 0.480 | -2.05 [-96.7; 92.6] | 0.966 |
| Homa Index | -3.64 [-11.2; 3.9] | 0.352 | 0.31 [-3.05; 3.67] | 0.858 |
| Triglycerides, mg/dL | -9.53 [-53.2; 34.1] | 0.672 | -33.3 [-66.6; -0.086] | 0.062 |
| Total cholesterol, mg/dL | 10.2 [-23.2; 43.5] | 0.554 | -4.44 [-25.3; 16.4] | 0.681 |
| HDLc Cholesterol | 1.87 [-7.65; 11.4] | 0.703 | -1.49 [-4.9; 1.91] | 0.399 |
| LDL cholesterol, mg/dL | 10.2 [-17.7; 38.1] | 0.479 | 2.03 [-16.1; 20.2] | 0.829 |
| C-peptide, pg/mL | -149 [-498; 200] | 0.411 | 50.4 [-205; 306] | 0.703 |
| Ghrelin, pg/mL | -225 [-548; 97.1] | 0.181 | -45.4 [-163; 72.1] | 0.457 |
| Leptin, pg/mL | -584 [-4310; 3140] | 0.761 | -276 [-1920; 1370] | 0.745 |
| GLP, pg/mL | -58.6 [-141; 24] | 0.175 | -16.9 [-88.1; 54.2] | 0.646 |
| IL6, pg/mL | 0.92 [-0.25; 2.08] | 0.134 | 1 [-0.17; 2.16] | 0.107 |
| IL8, pg/mL | -0.55 [-1.99; 0.9] | 0.466 | -0.62 [-1.57; 0.34] | 0.219 |
| Resistin, pg/mL | -1270 [-2530; -22.4] | 0.056 | -5.84 [-1190; 1180] | 0.992 |
| TNF a, pg/mL | -5.59 [-14.2; 3] | 0.212 | 3.81 [-3.15; 10.8] | 0.295 |
| PAI-1, pg/mL | 516 [-135; 1170] | 0.131 | 744 [282; 1210] | 0.004 |
| Visfatin, pg/mL | -258 [-1250; 731] | 0.613 | 19.9 [-626; 665] | 0.952 |
| sICAM, pg/mL | 9530 [-19500; 38600] | 0.525 | 22100 [2250; 42000] | 0.040 |
| LBP, ng/mL | 2520 [-411; 5450] | 0.103 | 1710 [-1210; 4630] | 0.263 |
Intergroup comparisons in follow-up values relative to control group were estimated by multivariable linear regression model 1 adjusted for age, sex, BMI, MedDiet adherence (14pt) and baseline values. EBLong, Elias Boulanger® long fermentation bread; EBShort, Elias Boulanger® short fermentation bread; HDLc, high density lipoprotein cholesterol; LDLc, low density lipoprotein cholesterol; GLP-1, glycoprotein 1; IL6, interleukin 6; Il8, interleukin 8; TNF-a, tumor necrosis factor alpha; PAI-1, plasminogen activator inhibitor-1; sICAM, soluble intercellular adhesion molecule; LBP, lipopolysaccharide binding protein.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



