
Submitted:
20 June 2024
Posted:
21 June 2024
You are already at the latest version
Abstract
High temperature stress affects tomato production both in tropical and sub-tropical environments worldwide. To explore genetic variation for heat tolerance in tomato, 329 transcontinental tomato varieties were evaluated at The Ministry of Municipality and Environment (MME) greenhouses near Doha, Qatar. Preliminary phenotypic analysis identified a diverse set of 71 hybrid and pure line tomato varieties for more detailed studies. The selected subset was examined in the greenhouse in a randomized complete block design under heat stress. The materials were phenotyped for fruit size, fruit weight, fruit hardness, fruit locules, fruit set (%), total soluble solids (TSS), and fruit yield. Significant phenotypic differences among genotypes were observed for all the traits assessed. To explore the genetic basis of variation among varieties the subset was genotyped using 104 SNP markers identified in previous heat tolerance genome wide association studies (GWAS). Nineteen QTL associated SNP markers could reliably select heat tolerant varieties in terms of better fruit yield, fruit set, and TSS. These markers are located on Chromosome 1, 5, 6, 8, 9, and 12. Interestingly, two clusters of markers on chromosome 6 were linked to significant effects on yield, fruit set, and TSS under high temperature. Some markers located within these QTL regions were also reported as associated with heat tolerance in other studies. Eighteen out of nineteen SNP markers were mapped within a gene body. Based on the phenotypic and the genotypic analysis, an elite set of five varieties was selected for approval for heat stress environments in Qatar. These findings have significance not only for Qatar, but they are also valuable for wider application at other locations worldwide. The 19 markers presented here could be adopted by breeders for marker-assisted selection (MAS) for heat tolerance.
Keywords:
high temperature stress
; QTL
; SNP marker
; genotypic selection
; tomato
1. Introduction
Global warming is increasing [1] and higher temperatures will progressively limit agricultural production, especially in tropical and subtropical regions [2,3,4,5]. A 28% reduction in tomato yield under high temperatures was reported in Australia [6]. There is an urgent need to improve adaptive management of crops and the selection of heat tolerant germplasm for current and future production environments [7,8]. Qatar has a desert climate, and crops can only grow in winter and spring. However, global warming is reducing the duration of both winter and spring with consequences for vegetable production [9]. Heat tolerant crops, therefore, will play an important role in future agriculture in Qatar.
Tomato (Solanum lycopersicum L.) is an important horticultural crop worldwide. It can be grown in both subtropical and tropical zones. The optimal daytime temperature for tomato production is 25 °C to 30 °C [10]. If temperature exceeds a critical point, productivity will fall significantly. For example, El Ahmadi and Stevens [11] reported that in several heat-tolerant tomato varieties, the number of flowers, pollen viability, fruit set, and yield were dramatically reduced under 38/27 °C day/night temperatures. Heat stress is defined as temperatures 10–15 °C higher than optimal [2]. High temperature stress can cause negative impact on plant development, including morphology, physiology, biochemistry, and molecular pathways at all vegetative and reproductive stages, which leads to loss of yield. During anthesis, tomato is very sensitive to temperature fluctuations, which impairs anther, pollen, and pistil development, leading to reduced fertilization, lower fruit set, and poorer quality fruit and yield [2,12,13,14]. At the physiological level, heat stress impacts photosynthesis, respiration, and membrane plasticity [15,16,17]. Damage to cell membranes results in electrolyte leakage [18,19]. Electrolyte leakage is commonly used to assess tolerance and sensitivity to heat stress [6,20]. Studies of the tomato transcriptome under normal and heat stressed conditions, identified hundreds of genes that changed expression, including heat shock proteins (HSPs) and their related transcription factors (HSFs) [21,22,23,24]. High temperature stress also causes biochemical changes, including changes to the levels of sugars, fatty acids, proline, salicylic acid, and abscisic acid. In addition, reactive oxygen species (ROS) accumulate and enzymes in chloroplasts and mitochondria are inactivated [17,25].
Heat tolerance is controlled by multiple genes which induce physiological and biochemical changes. Several studies have identified quantitative trait loci (QTL) linked to reproductive traits under heat stress using biparental QTL mapping, introgression lines, multiparent advanced generation intercross (MAGIC) populations [26,27,28,29] and genome-wide association studies (GWAS) [6]. Candidate genes linked to the heat stress response have also been identified [23,24,30]. Recently, genomic selection was applied to tomato [30,31,32] with some success. Heat tolerant genotypes were successfully predicted with good accuracy for yield (0.729) and total soluble solids (SCC, 0.715) [30]. Whole genome sequencing of a heat tolerant line revealed highly variable chromosome regions (QTL) compared to a reference genome and a high number of candidate genes [24]. While genomic selection and whole-genome sequencing may not be cost effective for a small breeding program, traditional marker-assisted selection (MAS) for key traits remains viable.
In this study, previously identified tomato QTL markers for agronomic traits under heat stress [6,26] were validated in a totally different set of 71 tomato varieties grown in Qatar. The aim was to identify and validate QTL for genotypic selection for heat tolerance. The identified markers would be useful to breeders for MAS.
2. Materials and Methods
2.1. Plant Materials and Growth Condition
Transcontinental tomato varieties were evaluated at the Ministry of Municipality and Environment (MME) net greenhouse (25.518031°N, 51.208001°E) at the Al-Utouriya research station, Doha, Qatar. In the first season 2019 – 2020, 329 varieties were grown in the greenhouse. Ten plants per variety were sown in October and seedlings were transplanted at the end of October following a completely randomized design in three replicates. Fruits were harvested from February to May. Plants were spaced 0.5 m between rows and 0.3 m within rows and watered via a fully automated drip irrigation system. In the second season 2020 - 2021, 71 transcontinental tomato varieties (Table 1) were grown in the same conditions in a completely randomized block design with three replicates. These varieties included different tomato types: globe, plum, cherry, oxheart, and beefsteak. Bumble bees were used in all seasons and greenhouses to help pollinate flowers. Mid-day temperature (11:00am) was recorded from December to May 2000 - 2021 (Figure 1). In the third season 2021 – 2022, 10 selected varieties were evaluated in a randomized complete block designs of three replicates.
2.2. Phenotyping and Statistical Analysis
Harvesting was conducted every 10 days in the first month, and twice per week afterwards. Yield was calculated as the weight of all harvested fruits per plant. Final yield was expressed as yield/3 plants (kg). Five fruits from each plot were taken at ripeness from the fourth harvest to determine fruit characteristics. Fruit length and diameter was determined using a digital caliper (cm). Fruit firmness (hardness) was measured on the two opposite sides of the fruit using a pressure test (lb/cm2). Total soluble solids Brix (TSS) in the fruit juice was determined using a handheld refractometer. Fruits were cut crosswise to determine the number of locules. Flowers/inflorescence were counted every 2 – 4 days. Fruits/inflorescence were also recorded. Fruit set (%) was then calculated as fruits/inflorescence divided by flowers/inflorescence.
Data were analysed for each year independently using a linear mixed model and the REML function of the statistical software package Genstat (64-bit release 22.1, VSN International Ltd). Genotype was considered a fixed effect and plot position within each replicate a random effect. The 10 selected genotypes were analysed across years. In this instance, year and genotype were considered fixed effects and plot position within replicates within years as random effects.
2.3. QTL Marker Selection and Genotyping
Previous GWAS of tomato identified more than 100 SNP markers [6] associated with various traits under heat stress. To validate these markers in a different set of tomato varieties, 96 markers were selected by filtering out those with low PIC, high percentage missing data and/or sequence uncertainty. Eight SNP markers from Xu et al. [26] were also included. All markers were projected on to the tomato genome SL4.0 (https://solgenomics.net/organism/Solanum_lycopersicum/genome). The names of 104 markers were designated as genome version (SL4-0) + chromosome number (ch01) + position, e.g., SL4-0ch01_1477682, to assist identification. All the marker sequences are listed in the supplementary document (Table S1). DNA extraction and genotyping (SeqSNP) of 71 tomato varieties was conducted by LGC Biosearch Technologies. The genotype and phenotype (fruit_set, yield and TSS) association study was done using Box plotting. A Student T-test was used to determine significance.
2.3. Comparison among Published QTL and QTL Identified in this Study
3. Results
3.1. Phenotypic Data Analysis
Phenotypic traits varied significantly among the 71 selected varieties. However, many of these varieties included different types and fruit shapes including globe, plum, cherry, oxheart, and beefsteak. This variation must be accounted for if the set was to be used in an association for heat tolerance. Therefore the differences among fruit types was assessed (Table 2). Results showed that TSS, fruit_set and yield (without beefsteak) were not significant among types. These three traits were subsequently used in the association study.
The 10 selected varieties were analysed across years to assess main effects and genotype x environment interaction (Table 3). Year and genotype effects were significant at P<0.05 for all traits. The genotype x environment interaction was also significan for all traits except locule number and fruit_set. When tomato types Globe and Plum were separated and analysed, difference in main and interaction effects were observed (Table 4). Globe genotype effects were all significant, as was the genotype x environment effect for yield and Frt_Width. However, the plum type genotype effects were only significant for Frt_Width, although a genotype x environment effect was observed for yield. The envionment main affects were all significant across types except for globle Locule_number and yield.
3.2. QTL Marker Genotyping and Association Study
A total of 104 QTL SNP markers selected from previous GWAS studies [6,26] were used to genotype the 71 varieties selected for evaluation in the second season. The marker profiles of the 71 varieties were listed in Table S3. Twenty-one markers had a minor allele frequency larger than 9% and were selected for genotypic and phenotypic association analysis. Among these 21 markers, 19 had a significant effect on tomato fruit_set, yield, and TSS under heat stress in Qatar. These markers are located on chromosomes 1, 5, 6, 8, 9, and 12 (Figure 2 and Figure 3). Two markers were located on each of chromosomes 1, 5, 9, and 12, one marker on chromosome 8, and 10 markers on chromosome 6. One marker located on chromosome 3 and another on chromosome 4 had no significant effect on the traits analysed.
Interestingly, clusters of markers were identified on chromosome 6, most of them on the short arm. Figure 4 shows the markers on chromosome 6 that associated with both fruit_set and yield, while those associated with yield only are shown in Figure 5.
The markers on chromosome 6 that associated with yield, fruit_set and TSS are shown in Figure 6. Another marker on the short arm of chromosome 9 was very significantly associated with TSS (Figure 7). Alleles that are favorable for yield and fruit_set often had the opposite effect on TSS. The 19 markers associated with traits and their favorable alleles are summarized in Table 5.
3.3. Genotypic and Phenotypic Selection
Favourable alleles for each marker were identified based on association with desirable phenotype. According to the genotype of each variety, favourable varieties for high fruit_set and yield were selected (Table 6). Based on phenotypic data, five varieties were selected for heat stress environments in Qatar (Table 7).
Five tomato cultivars selected phenotypically for growing under Qatari conditions had genotypes favorable for yield (#43, #44, #45, #58) and yield and quality (TSS, #47).
3.4. QTL Markers Comparsion
Tomato QTL linked to reproductive traits affected by high temperature, such as flower number (FLN) and fruit number (FRN) per truss and percentage of fruit set (FRS), pollen viability (PV), and yield, have been recently investigated [26,27,28]. The chromosome locations of the reported QTL were in the genome sequence of SL2.5. These reported QTL positions were projected into genome SL4.0, so that the QTL identified in this study can be compared with those reported. Some of the QTL identified in this study are in the same regions of reported QTL, while others are new (Figure 8).
Mapping the position of markers in the genome (SL4.0) showed that the majority of markers (18 out of 19 markers) were locate within a gene body (Table 8). SNPs locate either in the coding region or in the intron, 5’ untranslated region (UTR), 3’ UTR, and downstream (Table 8). Variation in the coding region could result in dysfunctional protein if it is a synonymous or missense mutation. Whereas sequence changes in the intron, 5’ UTR, 3’ UTR, and up- or downstream of a gene could cause transcriptional or translational change.
4. Discussion
Qatar has a desert climate. It has very low annual rainfall and a hot and long summer. The experiment was carried out in a net greenhouse to assess tomato production under high temperature. The temperature inside the net greenhouse was higher than the ambient temperature. Figure 1 demonstrated that day temperature during the tomato growth period was fluctuated severely. During the vegetative growth (December), the plants experienced a few days of over 40 °C. During the reproductive period (January and February), more days exceeded 40 °C. The effects of heat stress on vegetative development were evident at high temperature (i.e., 40 °C) [22], whereas reproductive traits are often affected by long-term mild heat stress (i.e., 31 °C) [26], or short periods of high heat stress (over 40 °C) [2]. Plant response to heat stress is complex and controlled by multiple genes. Phenotypic traits, such as flower number, fruit number, percentage of fruit set, stigma exsertion, pollen viability, electrolyte leakage, soluble solid content, were used for QTL analysis by others [6,26,27,28]. It was shown that fruit set is an important trait that directly affects yield. In this report, fruit set and yield were the focus of the genetic analysis. As TSS was an important trait for the fruit quality it was also included in the analysis.
Previous GWAS [6] used 144 tomato accessions and DArTseq (Diversity Arrays Technology by sequencing) for association study and identified 142 QTL markers (SNP) that had high log scores associated with heat tolerance. In the previous report, the arbitrary number from DArTseq were used as the marker name, which was not meaningful. In this study, 96 markers were selected, and the SNP position was converted from tomato genome SL2.4 to SL4.0. The name of the markers was converted to show genome sequence version, chromosome number, and position. This will allow the research community and breeders to use these markers easily. The QTL markers/positions identified in other studies [26,27,28] were also converted according to SL4.0. Thus, these QTL were comparable (Figure 8).
Comparison of markers identified in this study with the QTL reported by other researchers showed general agreement for several QTL. Markers SL4-0ch01_68237408 and SL4-0ch05_58243373 perfectly aligned with QTL associated with fruit number, flower number, and fruit set [27,28]. Other markers, SL4-0ch8_ 61071507, SL4-0ch09_59454323, SL4-0ch12_4886349, located within 4 Mbp of other QTL related to flower and fruit traits [27,28]. Marker SL4-0ch09_6794487 showed strong association with TSS, which is also located within 4 Mbp of a SSC QTL [27]. Three markers from Xu et al. (2017) showed correlation with yield and fruit set in this study. SL4-0ch01_1477682 (solcap_snp_sl_8704) was associated with style length [26]. SL4.0ch01_68237408 (solcap_snp_sl_13762) was associated with flower number per inflorescence [26]. These two traits were related to productivity and final yield. Another marker, SL4.0ch08_61071507 (solcap_snp_sl_15446), was associated with inflorescence number [26], which in this study was related to fruit set. Although 104 markers were used to genotype the 71 tomato varieties/accessions, only 21 markers showed enough polymorphism (minor allele frequency larger than 9%) for association analysis, and 19 markers showed significant association with yield, fruit set, and TSS. Two markers, SL4-0ch03_56340171 and SL4-0ch04_62857466, which mapped within the QTLs related to flower number, fruit number, and fruit set (Figure 8), were not significant for fruit set and yield in this study. This was probably due to the small population size and reduced power of the association study.
Interestingly 10 markers on chromosome 6 had a significant impact on yield, fruit set and TSS. Alsamir et al. (2017) [6] identified markers on chromosomes 1 and 6 that significantly impacted electrolyte leakage (EL). The EL trait was indicative of heat stress impact which was reflected in yield in this study. Cappetta et al. (2021) [30] found a high density of SNPs on chromosome 6 linked to heat tolerance. A major QTL was found on chromosome 6 (in a similar region to that reported here) which explained 86% of the phenotypic variance related to yield [30]. This QTL region contains Solyc06g006057, Solyc06g007310, Solyc06g007530, Solyc06g008720, Solyc06g009920, Solyc06g036260, Solyc06g036485, Solyc06g051190 variant genes, coding for Leucine-rich receptor-like protein kinase family protein, Deoxyribonuclease tatD, B3 domain-containing protein (Os05g0481400), Zinc ion binding protein, ATPase E1-E2 type family protein, Beta-carotene hydroxylase 1, Kinase family protein, and RNA-dependent RNA polymerase family protein. Using differential gene expression analysis of tolerant and sensitive accessions under high temperature, Gonzalo et al. (2021) [23] identified genes on chromosome 6 that upregulated during heat stress in tolerant accessions, including heat shock proteins, gibberellin-3-β-dioxygenase 1, and indeterminate-domain 16-like protein, which is plant specific transcription factor regulating in sugar homeostasis, leaf and root architecture, inflorescence and seed development.
The most interesting finding was that the markers identified in this study are all located in a gene body except one. Table 8 lists the SNP markers and their associated genes and gene functions. These genes may be important for conveying heat tolerance in tomato. For example, ABC transporter (SL4-0ch05_38882416) is a transmembrane protein; its function is to import essential nutrients to the cell and to export toxic molecules out. The role of ABC transporter in the defence of multiple plant pathogens has been demonstrated [33,34,35]. The role of ABC transporters in abiotic stress response, such as heat stress, could also be important, but have yet to be studied. Another protein, Cullin (SL4-0ch01_68237408) and its protein family is involved in protein degradation. Involvement of Cullin in the heat stress response is also possible. Multiple markers on chromosome 6 are located in different genes, including protein kinase (same finding as in [30]), Glyoxysomal fatty acid β-oxidation multifunctional protein (lipid metabolism), hydrolase, pleiotropic drug resistance protein, and phosphoinositide phospholipase C (signal transduction). Another chloroplastic Serine/threonine-protein kinase (SL4-0ch12_15246947) may play roles in photosynthesis during heat stress. Indeed, SNP location is important for genetic selection. Cappetta et al. (2021) [30] used a subset of 2,278 SNPs mapped in gene body regions to perform genomic selection (GS). They obtained similar accuracy to the full dataset of 10,648 SNPs. Overall, the markers/genes identified in this study are of importance for selection of heat tolerant tomato varieties/accessions.
5. Conclusions
Validation of previously identified QTL makers was successfully achieved by using different genetic resources. Nineteen SNP markers had significant effects on fruit set, yield, and total soluble solids of tomato under heat stress and are recommended for MAS in breeding programs. Mapping of SNPs identified eighteen candidate genes for heat tolerance. They are valuable for further study to explore the molecular mechanism of plant response to high temperature. Fundings in this study are of significance for the tomato industry and research community.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Table S1: The list of marker sequences; Table S2: Projection of published QTLs to genome SL4.0; Table S3: Marker profiles of 71 varieties used in this study.
Author Contributions
Conceptualization and funding acquisition, R.T., T.M., E.E.; methodology, E.E., T.M., C.D.; formal analysis, C.D., L.Z., R.T.; investigation, E.E., N.E., M.Q., N.S., A.K., M.M.; resources, E.E., T.M.; data curation, E.E., C.D.; writing—original draft preparation, C.D., E.E., T.M.; writing—review and editing, R.T., C.D., L.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Qatar National Research Fund, grant number NPRP11S-0129-180378.
Data Availability Statement
Data used in the article are available in all Figures and Tables.
Conflicts of Interest
The authors have no conflict of interest to disclose.
References
- Intergovernmental Panel on Climate Change (IPCC), Fifth Assessment Report: Synthesis, published online in 2014. https://www.ipcc.ch/report/ar5/syr/.
- Wahid, A.; Gelani, S.; Ashraf, M.; Foolad, M.R. Heat tolerance in plants: An overview. Environ. Exp. Bot. 2007, 61, 199–223. [Google Scholar] [CrossRef]
- Battisti, D.S.; Naylor, R.L. Historical warnings of future food insecurity with unprecedented seasonal heat. Science. 2009, 323, 240–244. [Google Scholar] [CrossRef] [PubMed]
- Schlenker, W.; Roberts, M.J. Nonlinear temperature effects indicate severe damages to US crop yields under climate change. Proc. Natl. Acad. Sci. USA. 2009, 106, 15594–15598. [Google Scholar] [CrossRef]
- Zhao, C.; Liu, B.; Piao, S.L.; Wang, X.H.; Lobell, D.B.; Huang, Y.; Huang, M.T.; Yao, Y.T.; Bassu, S.; Ciais, P. , et al. Temperature increase reduces global yields of major crops in four independent estimates. Proc. Natl. Acad. Sci. USA. 2017, 114, 9326–9331. [Google Scholar] [CrossRef] [PubMed]
- Alsamir, M.; Ahmand, N.; Ariel, V.; Mahmood, T.; Trethiwan, R. Phenotypic diversity and marker-trait association under heat stress in tomato (Solanum lycopersicum L.). Aust. J. Crop Sci. 2019, 13, 578–587. [Google Scholar] [CrossRef]
- Lohani, N.; Singh, M.B.; Bhalla, P.L. High temperature susceptibility of sexual reproduction in crop plants. J. Exp. Bot. 2020, 71, 555–568. [Google Scholar] [CrossRef] [PubMed]
- Janni, M.; Maestri, E.; Gulli, M.; Marmiroli, M.; Marmiroli, N. Plant responses to climate change, how global warming may impact on food security: A critical review. Front. Plant Sci. 2024, 14. [Google Scholar] [CrossRef]
- Francis, D.; Fonseca, R. Recent and projected changes in climate patterns in the middle east and north africa (MENA) region. Sci. Rep. 2024, 14. [Google Scholar] [CrossRef]
- Camejo, D.; Rodríguez, P.; Morales, A.; Dell'Amico, J.M.; Torrecillas, A.; Alarcón, J.J. High temperature effects on photosynthetic activity of two tomato cultivars with different heat susceptibility. J. Plant Physiol. 2005, 162, 281–289. [Google Scholar] [CrossRef]
- El Ahmadi, A. B., and Stevens, M. A. Reproductive responses of heat tolerant tomatoes to high temperatures. J. Am. Soc. Hort. Sci. 1979, 104, 686–691. [Google Scholar] [CrossRef]
- Xu, J.M.; Wolters-Arts, M.; Mariani, C.; Huber, H.; Rieu, I. Heat stress affects vegetative and reproductive performance and trait correlations in tomato (solanum lycopersicum). Euphytica. 2017, 213. [Google Scholar] [CrossRef]
- Gonzalo, M.J.; Li, Y.C.; Chen, K.Y.; Gil, D.; Montoro, T.; Nájera, I.; Baixauli, C.; Granell, A.; Monforte, A.J. Genetic control of reproductive traits in tomatoes under high temperature. Front. Plant Sci. 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.Y.; Impa, S.M.; Sunkar, R.; Jagadish, S.V.K. The neglected other half - role of the pistil in plant heat stress responses. Plant Cell Environ. 2021, 44, 2200–2210. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.S.; Ding, Y.F.; Zhu, C. Sensitivity and responses of chloroplasts to heat stress in plants. Front. Plant Sci. 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Haque, M.S.; Husna, M.T.; Uddin, M.N.; Hossain, M.A.; Sarwar, A.K.M.G.; Ali, O.M.; Abdel Latef, A.A.H.; Hossain, A. Heat stress at early reproductive stage differentially alters several physiological and biochemical traits of three tomato cultivars. Horticulturae 2021, 7, 330. [Google Scholar] [CrossRef]
- Moore, C.E.; Meacham-Hensold, K.; Lemonnier, P.; Slattery, R.A.; Benjamin, C.; Bernacchi, C.J.; Lawson, T.; Cavanagh, A.P. The effect of increasing temperature on crop photosynthesis: from enzymes to ecosystems. J. Exp. Bot. 2021, 72, 2822–2844. [Google Scholar] [CrossRef]
- Wahid, A.; Shabbir, A. Induction of heat stress tolerance in barley seedlings by pre-sowing seed treatment with glycinebetaine. Plant Growth Reg. 2005, 46, 133–141. [Google Scholar] [CrossRef]
- Biswas, P.; East, A.R.; Hewett, E.W.; Heyes, J.A. Increase in electrolyte leakage as a function of chilling stress and ripening of tomato. Acta Hortic. 2012, 945, 283–290. [Google Scholar] [CrossRef]
- Kumar, R.R., Goswami, S., Sharma, S.K., Singh, K., Gadpayle, K.A., Kumar, N., Rai, R.D. Protection against heat stress in wheat involves change in cell membrane stability, antioxidant enzymes, osmolyte, H2O2 and transcript of heat shock protein. Int. J. Plant Physiol. Biochem. 2012, 4 (4), 83–91. [CrossRef]
- Halter, G.; Simonetti, N.; Suguitan, C.; Helm, K.; Soroksky, J.; Waters, E.R. Patterns of thermotolerance, chlorophyll fluorescence, and heat shock gene expression vary among four boechera species and Arabidopsis thaliana. Botany. 2017, 95, 9–27. [Google Scholar] [CrossRef]
- Wen, J.; Jiang, F.; Weng, Y.; Sun, M.; Shi, X.; Zhou, Y.; Yu, L.; Wu, Z. Identification of heat-tolerance QTLs and high-temperature stress-responsive genes through conventional QTL mapping, QTL-seq and RNA-seq in tomato. BMC Plant Biol. 2019, 19. [Google Scholar] [CrossRef]
- Gonzalo, M.J.; Nájera, I.; Baixauli, C.; Gil, D.; Montoro, T.; Soriano, V.; Olivieri, F.; Rigano, M.M.; Ganeva, D.; Grozeva-Tileva, S., et al. Identification of tomato accessions as source of new genes for improving heat tolerance: From controlled experiments to field. BMC Plant Biol. 2021, 21. [CrossRef]
- Graci, S.; Ruggieri, V.; Francesca, S.; Rigano, M.M.; Barone, A. Genomic insights into the origin of a thermotolerant tomato line and identification of candidate genes for heat stress. Genes. 2023, 14. [Google Scholar] [CrossRef]
- Alsamir, M.; Trethowan, R.; Ahmad, N. An overview of heat stress in tomato (Solanum lycopersicum L.). Saudi J. Biol. Sci. 2021; 28, 1654–1663. [Google Scholar] [CrossRef]
- Xu, J.; Driedonks, N.; Rutten, M.J.M.; Vriezen, W.H.; de Boer, G.J.; Rieu, I. Mapping quantitative trait loci for heat tolerance of reproductive traits in tomato (solanum lycopersicum). Mol. Breed. 2017, 37. [Google Scholar] [CrossRef]
- Gonzalo, M.J.; Li, Y.C.; Chen, K.Y.; Gil, D.; Montoro, T.; Nájera, I.; Baixauli, C.; Granell, A.; Monforte, A.J. Genetic control of reproductive traits in tomatoes under high temperature. Front. Plant Sci. 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Bineau, E.; Diouf, I.; Carretero, Y.; Duboscq, R.; Bitton, F.; Djari, A.; Zouine, M.; Causse, M. Genetic diversity of tomato response to heat stress at the QTL and transcriptome levels. Plant J. 2021, 107, 1213–1227. [Google Scholar] [CrossRef] [PubMed]
- Gonzalo, M.J.; da Maia, L.C.; Nájera, I.; Baixauli, C.; Giuliano, G.; Ferrante, P.; Granell, A.; Asins, M.J.; Monforte, A.J. Genetic control of reproductive traits under different temperature regimes in inbred line populations derived from crosses between S. pimpinellifolium and S. lycopersicum accessions. Plants. 2022, 11. [Google Scholar] [CrossRef]
- Cappetta, E.; Andolfo, G.; Guadagno, A.; Di Matteo, A.; Barone, A.; Frusciante, L.; Ercolano, M.R. Tomato genomic prediction for good performance under high-temperature and identification of loci involved in thermotolerance response. Hortic. Res. 2021, 8. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, E.; Matsunaga, H.; Onogi, A.; Ohyama, A.; Miyatake, K.; Yamaguchi, H.; Nunome, T.; Iwata, H.; Fukuoka, H. Efficiency of genomic selection for breeding population design and phenotype prediction in tomato. Heredity. 2017, 118, 202–209. [Google Scholar] [CrossRef] [PubMed]
- Cappetta, E.; Andolfo, G.; Di Matteo, A.; Barone, A.; Frusciante, L.; Ercolano, M.R. Accelerating tomato breeding by exploiting genomic selection approaches. Plants. 2020, 9. [Google Scholar] [CrossRef] [PubMed]
- Krattinger, S.G. , Lagudah, E.S., Spielmeyer, W., Singh, R.P., Huerta-Espino, J., McFadden, H., Bossolini, E., Selter, L.L. and Keller, B. A putative ABC transporter confers durable resistance to multiple fungal pathogens in wheat. Science 2009, 323, 1360–1363. [Google Scholar] [CrossRef]
- Schnippenkoetter, W.; Lo, C.; Liu, G.Q.; Dibley, K.; Chan, W.L.; White, J.; Milne, R.; Zwart, A.; Kwong, E.; Keller, B. , et al. The wheat Lr34 multipathogen resistance gene confers resistance to anthracnose and rust in sorghum. Plant Biotechnol. J. 2017, 15, 1387–1396. [Google Scholar] [CrossRef]
- Sucher, J.; Boni, R.; Yang, P.; Rogowsky, P.; Büchner, H.; Kastner, C.; Kumlehn, J.; Krattinger, S.G. Keller, B. The durable wheat disease resistance gene Lr34 confers common rust and northern corn leaf blight resistance in maize. Plant Biotechnol. J. 2017, 15, 489–496. [Google Scholar] [CrossRef]
Figure 1.
Temperature at mid-day during the growth period of year 2.
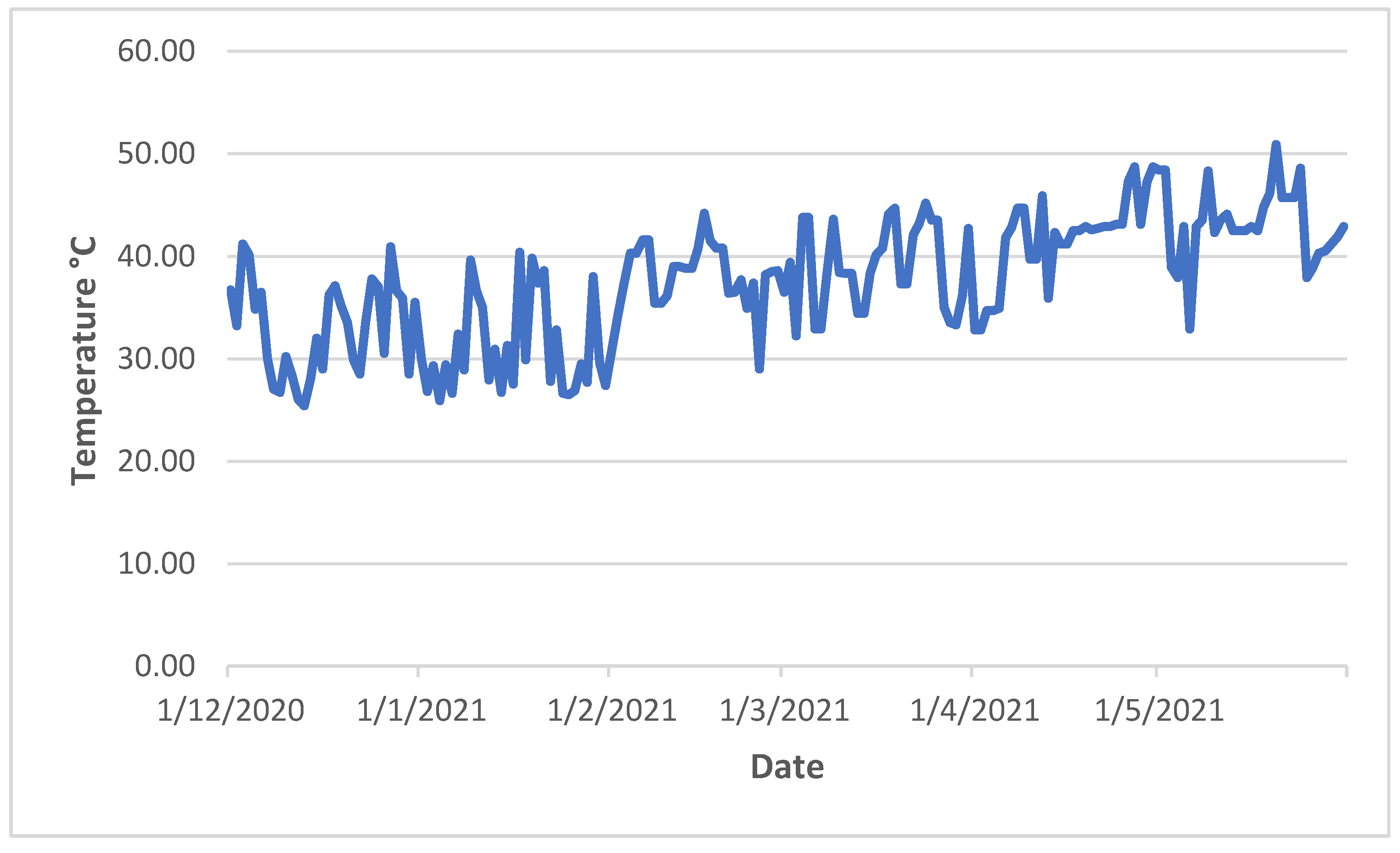
Figure 2.
Boxplots showing markers on chromosome 1, 5, 9 and 12 associated with yield under heat stress. The mean is represented by the x sign, while the median is represented by the horizontal line that divide the box. The lower and upper box boundaries represent 25th percentile and 75th percentile, respectively. ** and * indicate statistical significance p<0.01 and p<0.05, respectively.
Figure 2.
Boxplots showing markers on chromosome 1, 5, 9 and 12 associated with yield under heat stress. The mean is represented by the x sign, while the median is represented by the horizontal line that divide the box. The lower and upper box boundaries represent 25th percentile and 75th percentile, respectively. ** and * indicate statistical significance p<0.01 and p<0.05, respectively.
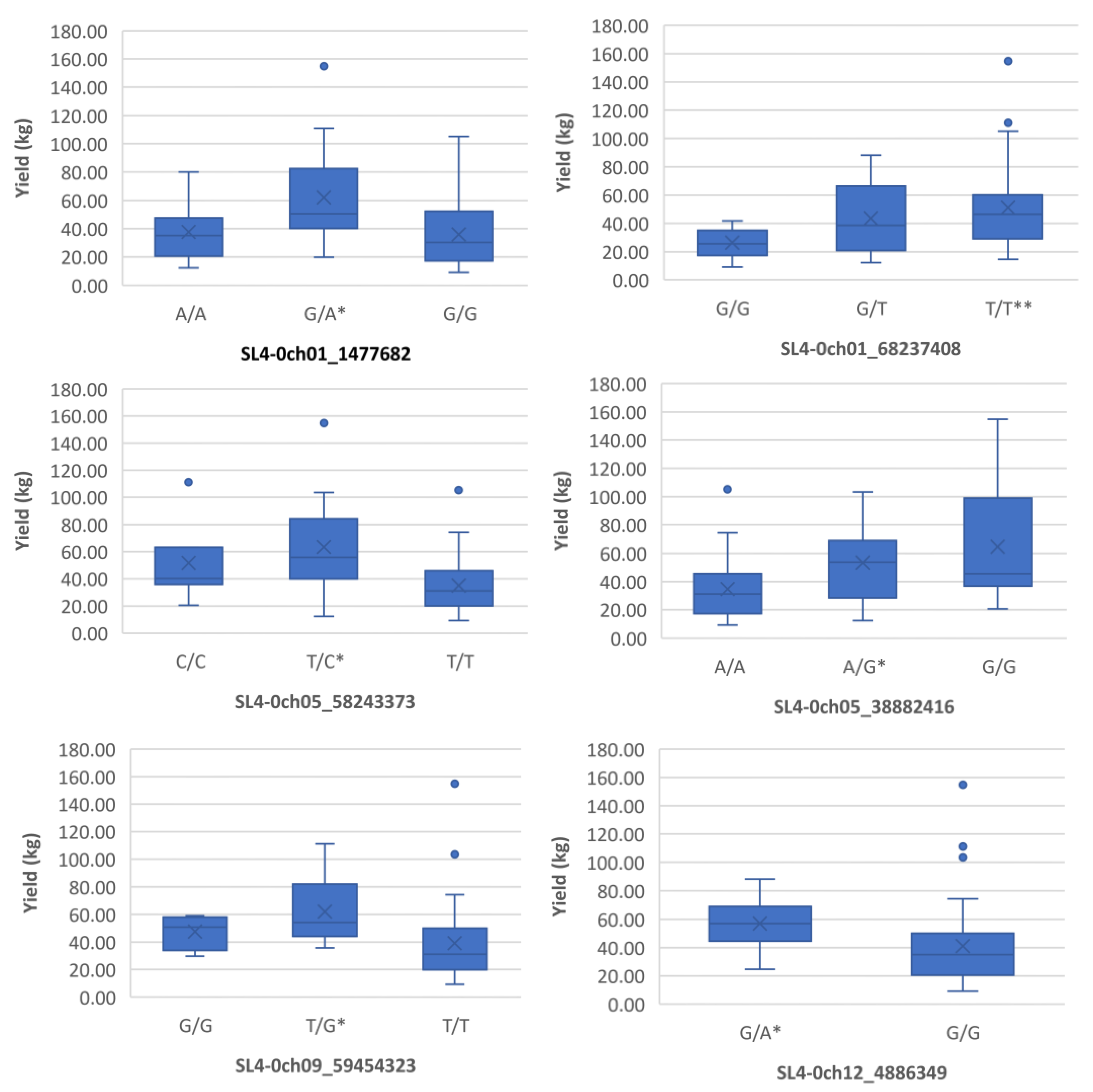
Figure 3.
Boxplots showing markers on chromosome 8 and 12 associated with fruit_set under heat stress. The mean is represented by the x sign, while the median is represented by the horizontal line that divide the box. The lower and upper box boundaries represent 25th percentile and 75th percentile, respectively. ** and * indicate statistical significance p<0.01 and p<0.05, respectively.
Figure 3.
Boxplots showing markers on chromosome 8 and 12 associated with fruit_set under heat stress. The mean is represented by the x sign, while the median is represented by the horizontal line that divide the box. The lower and upper box boundaries represent 25th percentile and 75th percentile, respectively. ** and * indicate statistical significance p<0.01 and p<0.05, respectively.
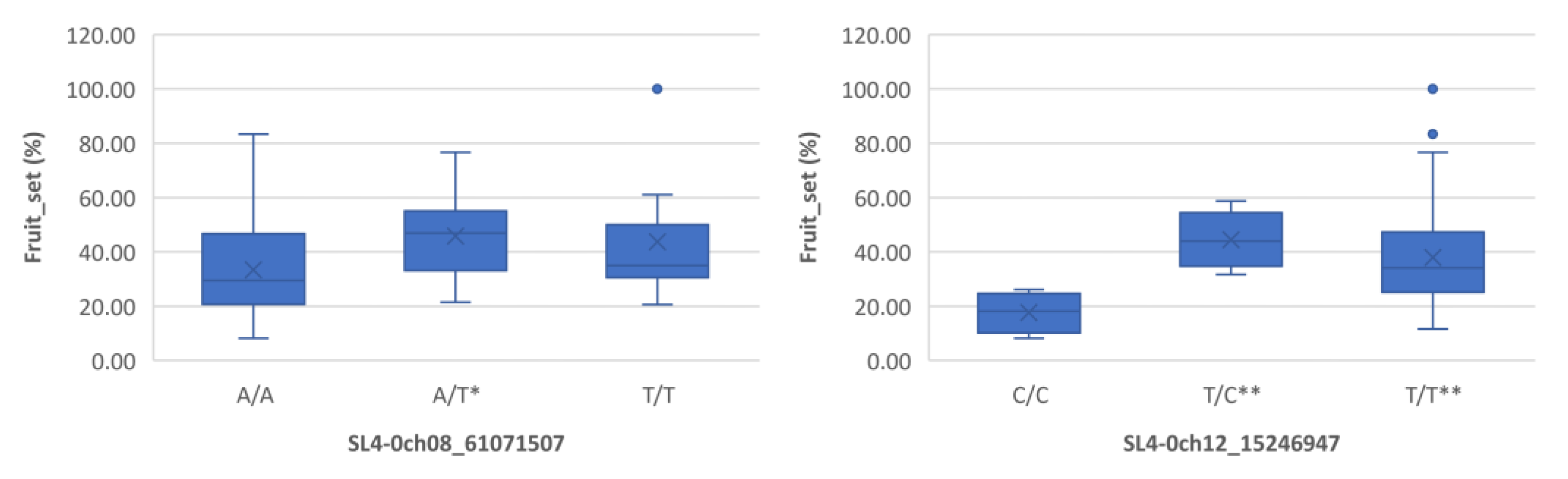
Figure 4.
Boxplots showing markers on chromosome 6 associated with both fruit_set and yield under heat stress. The mean is represented by the x sign, while the median is represented by the horizontal line that divide the box. The lower and upper box boundaries represent 25th percentile and 75th percentile, respectively. ** and * indicate statistical significance p<0.01 and p<0.05, respectively.
Figure 4.
Boxplots showing markers on chromosome 6 associated with both fruit_set and yield under heat stress. The mean is represented by the x sign, while the median is represented by the horizontal line that divide the box. The lower and upper box boundaries represent 25th percentile and 75th percentile, respectively. ** and * indicate statistical significance p<0.01 and p<0.05, respectively.
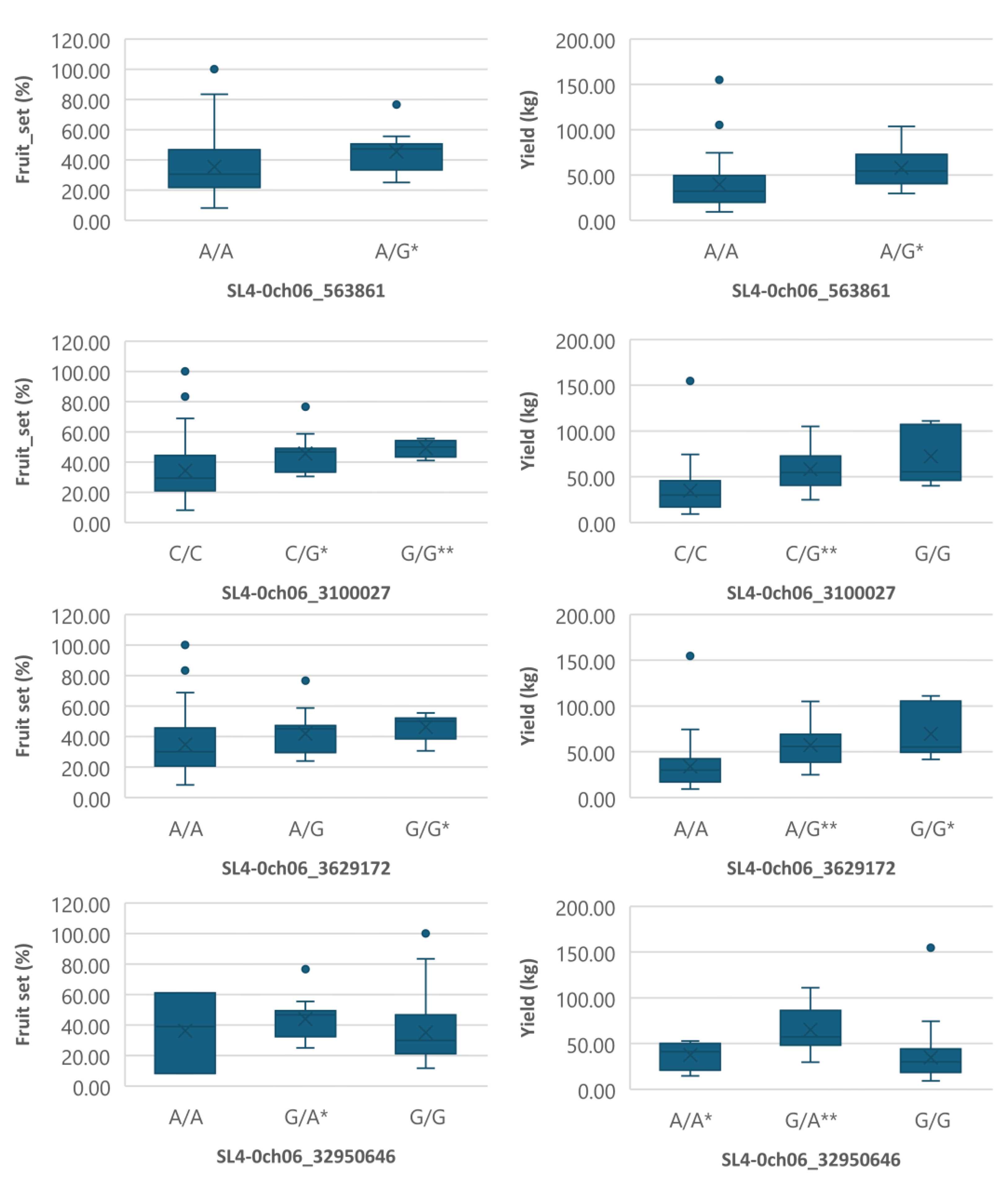
Figure 5.
Boxplots showing markers on chromosome 6 associated with yield under heat stress. The mean is represented by the x sign, while the median is represented by the horizontal line that divide the box. The lower and upper box boundaries represent 25th percentile and 75th percentile, respectively. ** and * indicate statistical significance p<0.01 and p<0.05, respectively.
Figure 5.
Boxplots showing markers on chromosome 6 associated with yield under heat stress. The mean is represented by the x sign, while the median is represented by the horizontal line that divide the box. The lower and upper box boundaries represent 25th percentile and 75th percentile, respectively. ** and * indicate statistical significance p<0.01 and p<0.05, respectively.
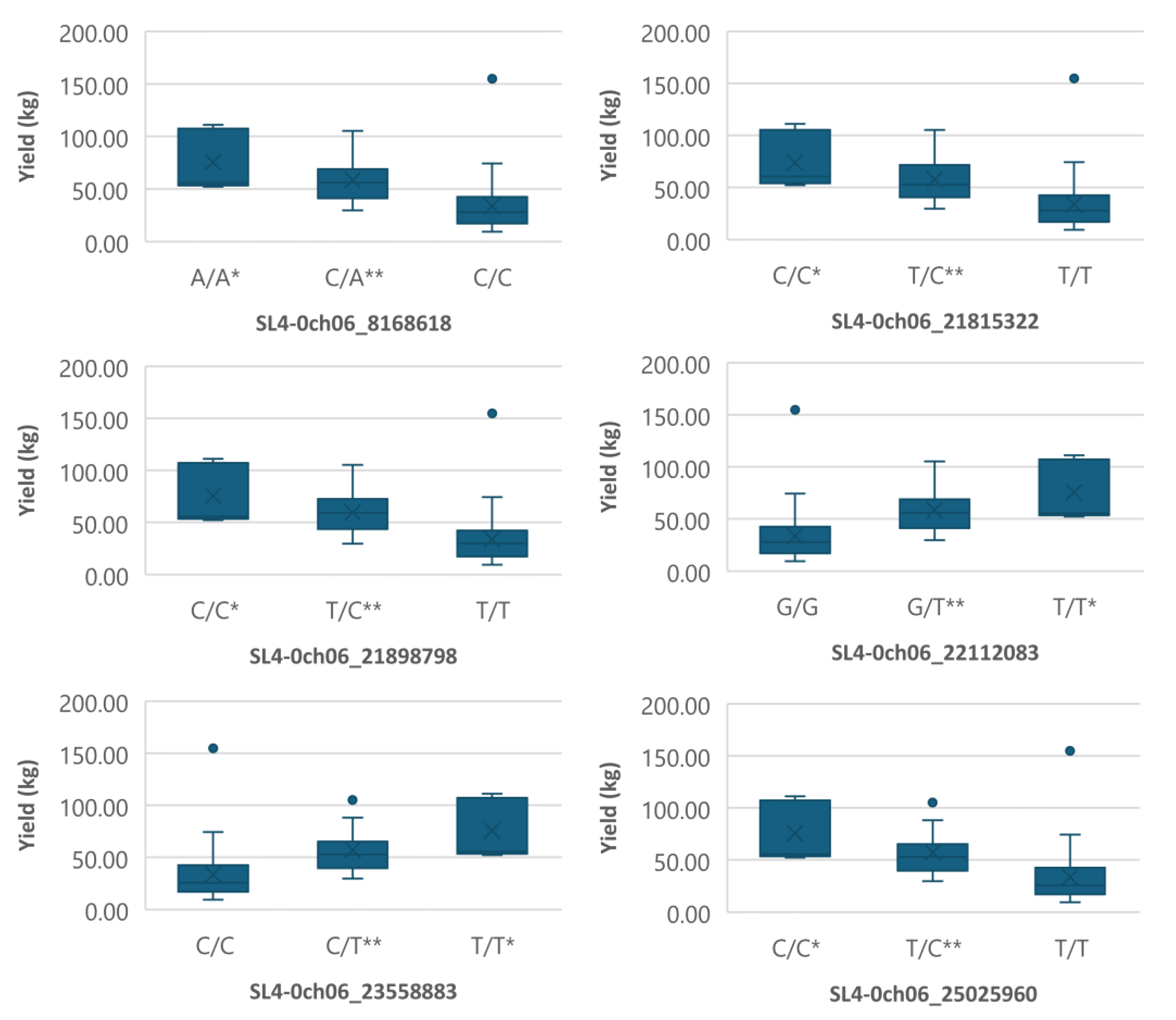
Figure 6.
Boxplots showing markers on chromosome 6 associated with TSS. The mean is represented by the x sign, while the median is represented by the horizontal line that divide the box. The lower and upper box boundaries represent 25th percentile and 75th percentile, respectively. ** and * indicate statistical significance p<0.01 and p<0.05, respectively.
Figure 6.
Boxplots showing markers on chromosome 6 associated with TSS. The mean is represented by the x sign, while the median is represented by the horizontal line that divide the box. The lower and upper box boundaries represent 25th percentile and 75th percentile, respectively. ** and * indicate statistical significance p<0.01 and p<0.05, respectively.
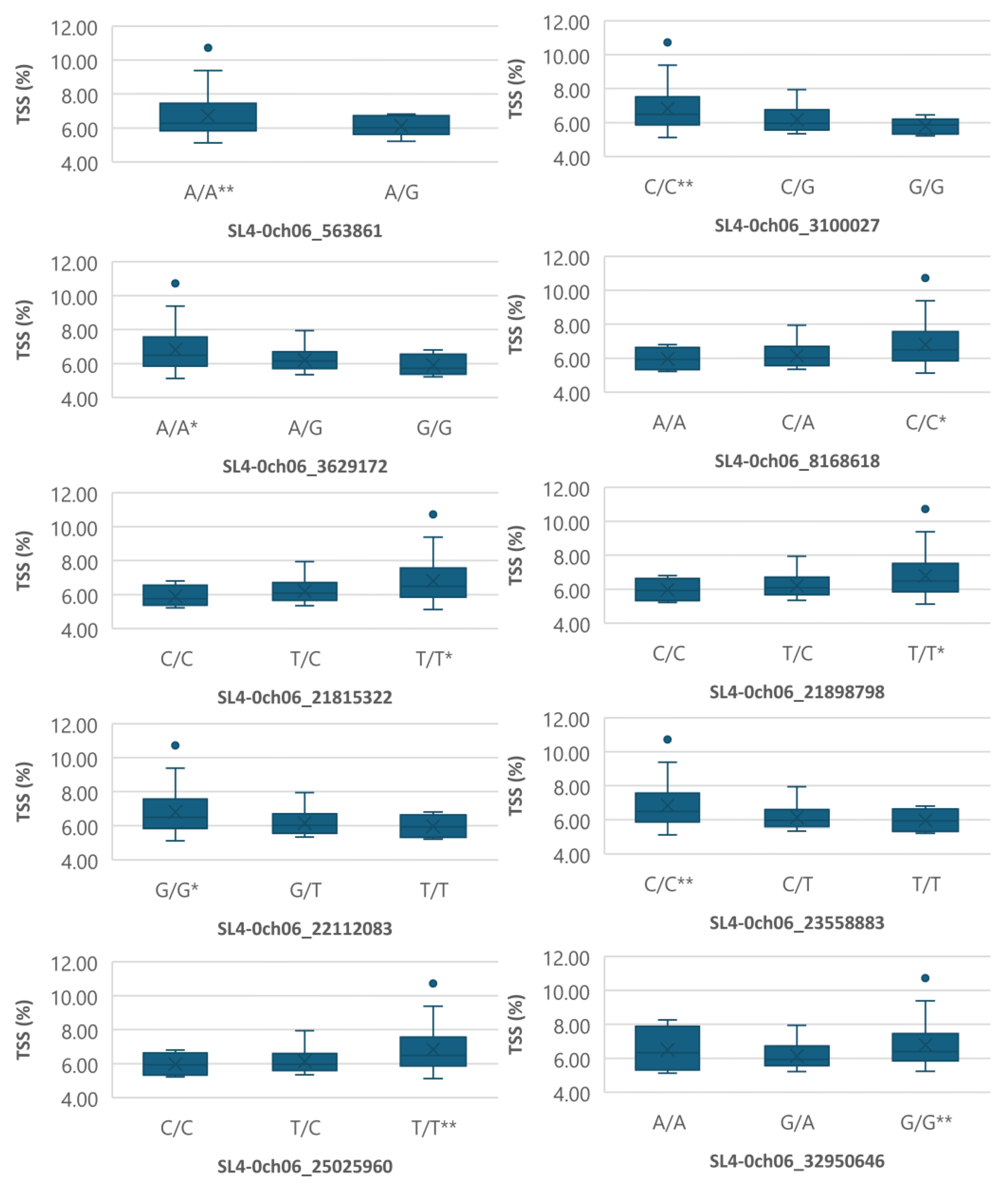
Figure 7.
Boxplots showing markers on chromosome 9 associated with TSS. The mean is represented by the x sign, while the median is represented by the horizontal line that divide the box. The lower and upper box boundaries represent 25th percentile and 75th percentile, respectively. ** and * indicate statistical significance p<0.01 and p<0.05, respectively.
Figure 7.
Boxplots showing markers on chromosome 9 associated with TSS. The mean is represented by the x sign, while the median is represented by the horizontal line that divide the box. The lower and upper box boundaries represent 25th percentile and 75th percentile, respectively. ** and * indicate statistical significance p<0.01 and p<0.05, respectively.
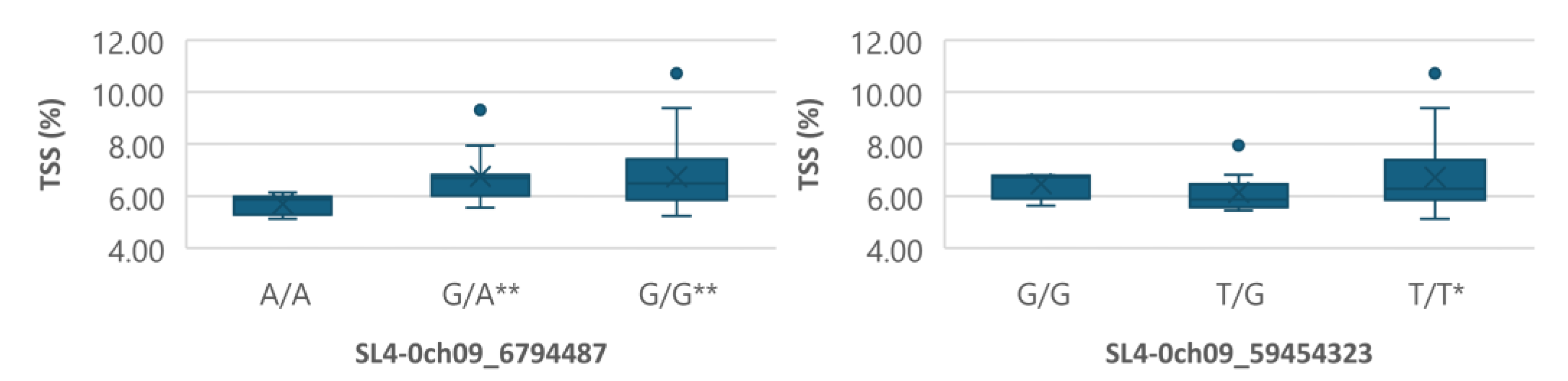
Figure 8.
comparison of QTL markers identified in this study (red) with QTL identified in Gonzalo et al. (2020, blue) and Bineau et al. (2021, green). Red rectangle shows the makers of interest are aligned in the same chromosome position of other QTL. Yellow rectangle shows the markers of interest are located within 4 Mbp of other QTL. Black rectangle shows clusters of markers identified in this study significantly associated with fruit_set, yield, and TSS under heat stress. (a) Chromosome 1 and 2; (b) Chromosome 3, 4, and 5; (c) Chromosome 6, 7, and 8; (d) Chromosome 9, 10, 11, and 12.
Figure 8.
comparison of QTL markers identified in this study (red) with QTL identified in Gonzalo et al. (2020, blue) and Bineau et al. (2021, green). Red rectangle shows the makers of interest are aligned in the same chromosome position of other QTL. Yellow rectangle shows the markers of interest are located within 4 Mbp of other QTL. Black rectangle shows clusters of markers identified in this study significantly associated with fruit_set, yield, and TSS under heat stress. (a) Chromosome 1 and 2; (b) Chromosome 3, 4, and 5; (c) Chromosome 6, 7, and 8; (d) Chromosome 9, 10, 11, and 12.
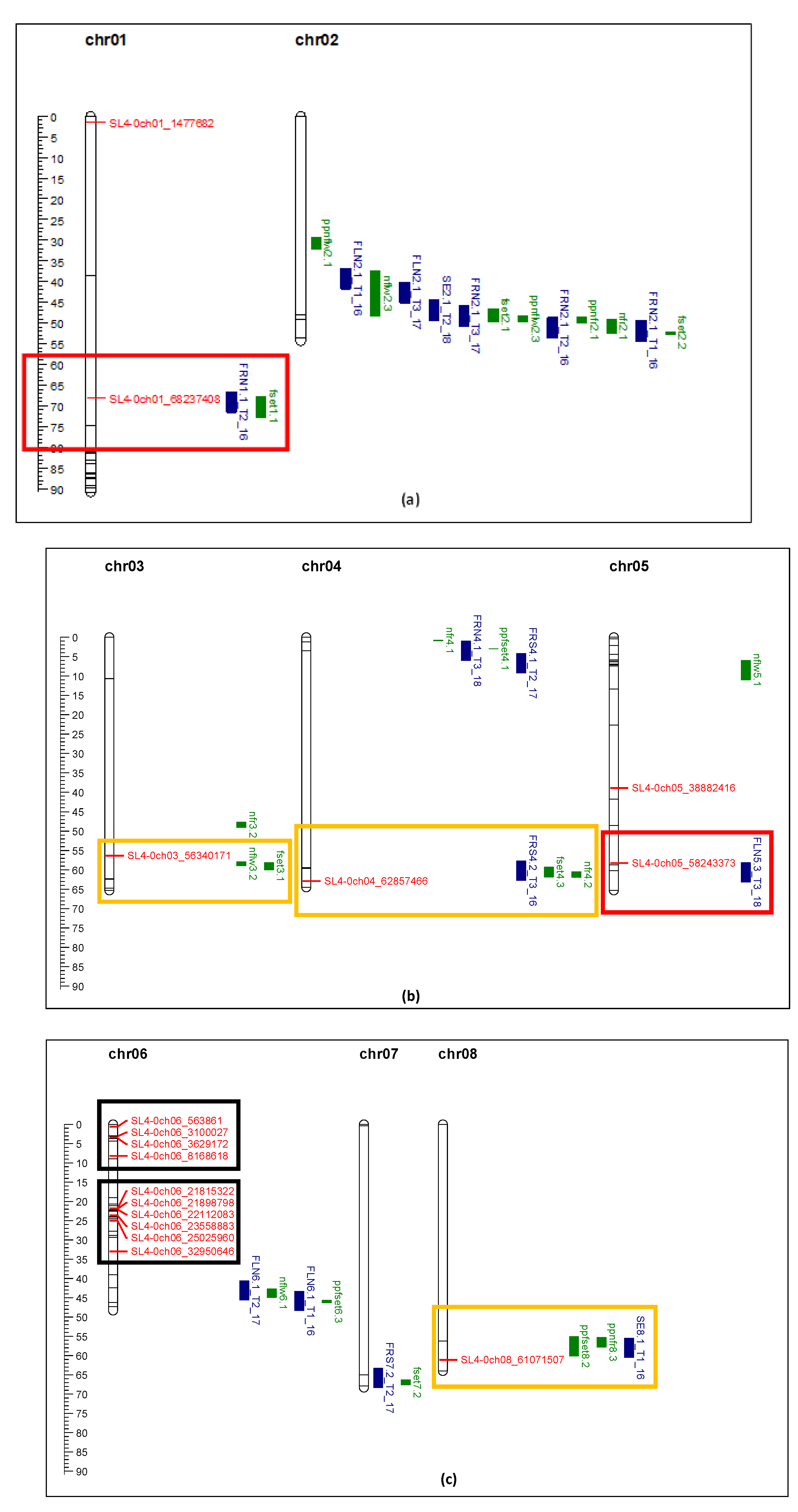
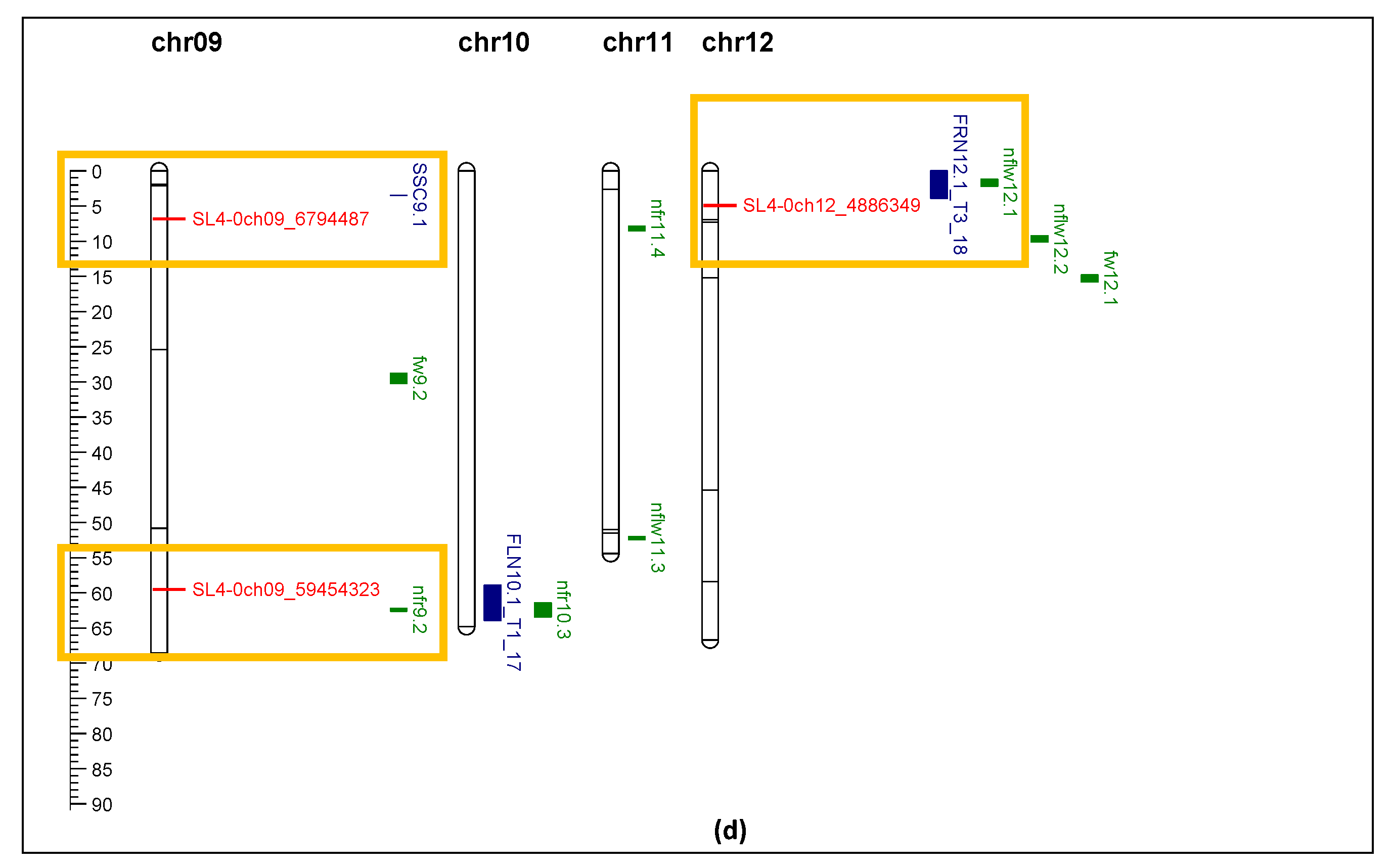

Table 1.
Tomato varieties selected from year 1 testing for evaluation in years 2 and 3.
| ID | Company code | Variety name | ID | Company code | Variety name | |
|---|---|---|---|---|---|---|
| 1 | TF-0014 | Amish Gold | 36 | TF-0141 | Ding Wall Scotty | |
| 2 | TF-0017S | Amy's Apricot | 37 | TF-0147 | Double Rich | |
| 3 | TF-0018 | Amy's Sugar Gem | 38 | TF-0176 | Florida Pink | |
| 4 | TF-0027 | Arkansas Marvel | 39 | Pakistan | Salar F1 | |
| 5 | TF-0032 | Aunt Lucy's Italian Paste | 40 | Pakistan | Surkhail F1 | |
| 6 | TF-0268 | Japanese Oxheart | 41 | Pakistan | Sundar F1 | |
| 7 | TF-0367 | Ozark Pink | 42 | Pakistan | Saandal F1 | |
| 8 | TF-0449 | Shenghaung Cherry | 43 | Pakistan | Tomato seed -2198 | |
| 9 | TF-0450 | Shirley Amish Red | 44 | Pakistan | Tomato seed -2199 | |
| 10 | TF-0173 | Fence Row Cherry | 45 | Pakistan | Tomato seed -2217 | |
| 11 | TF-0187 | German Johnson | 46 | Pakistan | Tomato seed -2218 | |
| 12 | TF-0197 | Giant Syrian | 47 | Pakistan | Tomato seed -2230 | |
| 13 | TF-0213F | Goose Creek | 48 | AVRDC | AVTO90304 | |
| 14 | TF-0227 | Gregori's Altai | 49 | AVRDC | AVTO9801 | |
| 15 | TF-0227G | Grightmire's Pride | 50 | AVRDC | AVTO1007 | |
| 16 | TF-0235 | Hazel Mae | 51 | AVRDC | AVTO1010 | |
| 17 | TF-0561 | Homer Fike's Yellow Oxheart | 52 | AVRDC | AVTO9001 | |
| 18 | TF-0035A | Austin's Red Pear | 53 | DRW7806 | hybrid tomato | |
| 19 | TF-0036 | Australia | 54 | Bright Star F1 | Bright Star F1 | |
| 20 | TF-0070A | Bloody Butcher | 55 | Roenza | Roenza | |
| 21 | TF-0077M | Brandy Sweet Plum | 56 | 619 | 619 | |
| 22 | TF-0078 | Brandywine | 57 | SV7846TH | SV7846TH | |
| 23 | TF-0106 | Chadwick Cherry | 58 | syngenta | Tomato: 413485 | |
| 24 | TF-0109 | Chapman | 59 | syngenta | Jarawa Ind tomato | |
| 25 | TF-0129 | Creole | 60 | syngenta | Tomato -Beef Vikllio | |
| 26 | TF-0135 | Dad's Sunset | 61 | syngenta | Pilavy Ind tomato | |
| 27 | TF-0137 | Debarao | 62 | syngenta | T415271 Ind tomato | |
| 28 | TF-0142 | Dinner Plate | 63 | syngenta | Tomato Dafnis | |
| 29 | TF-0146 | Dona | 64 | syngenta | Tomato Commondo | |
| 30 | TF-0149 | Dr. Lyle | 65 | syngenta | Tomato Izmono | |
| 31 | TF-0150 | Dr. Neal | 66 | TF-0004 | Ace 55 | |
| 32 | TF-0167 | Ethiopia Roi Humbert | 67 | TF-0021 | Anahu | |
| 33 | TF-0024 | Angora Super Sweet | 68 | TF-0330 | Mrs. Houseworth | |
| 34 | TF-0093 | Bulgarian Triumph | 69 | TF-0404J | Punta Banda | |
| 35 | TF-0121 | Clint Eastwood's Rowdy Red | 70 | TF-0486 | Sweet Organic tomato | |
| 71 | Banana Legs | Banana Legs |
Table 2.
The probability values of Wald statistics for different tomato types for various traits assessed on 71 tomato varieties.
Table 2.
The probability values of Wald statistics for different tomato types for various traits assessed on 71 tomato varieties.
| Frt_Width (cm) | Frt_Length (cm) | TSS (%) | Flowers /inflo | Fruit_set (%) | yield (kg) | yield* (kg) | |
|---|---|---|---|---|---|---|---|
| Tomato_type | <0.001 | <0.001 | 0.058 | <0.001 | 0.237 | 0.002 | 0.056 |
| *yield excluding Beefsteak. | |||||||
Table 3.
The probability values of Wald statistics from the 2 year analysis of 10 varieties.
| Frt_Width (cm) | Frt_Length (cm) | Hardness (lb/cm2) | TSS (%) | Locule_ number | Flowrs /inflo | Fruits /inflo | Fruit_set (%) | yield (kg) | |
|---|---|---|---|---|---|---|---|---|---|
| Year | 0.005 | 0.043 | <0.001 | <0.001 | 0.01 | 0.466 | <0.001 | <0.001 | 0.005 |
| Genotype | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | 0.001 | <0.001 |
| Year x Genotype | 0.029 | <0.001 | <0.001 | <0.001 | 0.354 | 0.016 | 0.013 | 0.132 | <0.001 |
Table 4.
The probability values of Wald statistics from the 2 year analysis of Globe only and Plum only.
Table 4.
The probability values of Wald statistics from the 2 year analysis of Globe only and Plum only.
| Globe | Plum | |||||||
|---|---|---|---|---|---|---|---|---|
| Frt_Width (cm) | Frt_Length (cm) | Locule_ number | yield (kg) | Frt_Width (cm) | Frt_Length (cm) | Locule_ number | yield (kg) | |
| Year | 0.066 | 0.008 | 0.154 | 0.467 | 0.027 | 0.026 | 0.015 | 0.001 |
| Genotype | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | 0.369 | 0.215 | 0.316 |
| Year x Genotype | 0.031 | 0.433 | 0.49 | <0.001 | 0.116 | 0.252 | 0.32 | 0.015 |
Table 5.
List of favorable alleles of SNP markers for yield, fruit_set, and TSS.
| Marker | Traits | Favorable alleles |
|---|---|---|
| SL4-0ch01_1477682 | Yield | GA* > GG, AA |
| SL4-0ch01_68237408 | Yield | TT**, GT > GG |
| SL4-0ch05_38882416 | Yield | GA*, GG > AA |
| SL4-0ch05_58243373 | Yield | CT*, CC > TT |
| SL4-0ch06_563861 | Fruit set | GA* > AA |
| SL4-0ch06_563861 | Yield | GA* > AA |
| SL4-0ch06_563861 | TSS | AA** > GA |
| SL4-0ch06_3100027 | Fruit set | GG**, GC* > CC |
| SL4-0ch06_3100027 | Yield | GC**, GG > CC |
| SL4-0ch06_3100027 | TSS | CC** > GC, GG |
| SL4-0ch06_3629172 | Fruit set | GG*, GA > AA |
| SL4-0ch06_3629172 | Yield | GA**, GG* > AA |
| SL4-0ch06_3629172 | TSS | AA* > GA, GG |
| SL4-0ch06_8168618 | Yield | CA**, AA* > CC |
| SL4-0ch06_8168618 | TSS | CC* > CA, AA |
| SL4-0ch06_21815322 | Yield | CC**, CT* > TT |
| SL4-0ch06_21815322 | TSS | TT* > CT, CC |
| SL4-0ch06_21898798 | Yield | CT**, CC* > TT |
| SL4-0ch06_21898798 | TSS | TT* > CT, CC |
| SL4-0ch06_22112083 | Yield | GT**, TT* > GG |
| SL4-0ch06_22112083 | TSS | GG* > GT, TT |
| SL4-0ch06_23558883 | Yield | CT**, TT* > CC |
| SL4-0ch06_23558883 | TSS | CC** > CT, TT |
| SL4-0ch06_25025960 | Yield | CT**, CC* > TT |
| SL4-0ch06_25025960 | TSS | TT** > CT, CC |
| SL4-0ch06_32950646 | Fruit set | GA* > GG, (AA) |
| SL4-0ch06_32950646 | Yield | GA** > AA* > GG |
| SL4-0ch06_32950646 | TSS | GG** > GA, (AA) |
| SL4-0ch08_61071507 | Fruit set | AT*, TT > AA |
| SL4-0ch09_6794487 | TSS | GG**, GA* > AA |
| SL4-0ch09_59454323 | Yield | GT*, (GG) > TT |
| SL4-0ch09_59454323 | TSS | TT*, (GG) > GT |
| SL4-0ch12_4886349 | Yield | GA* > GG |
| SL4-0ch12_15246947 | Fruit set | CT**, TT** > CC |
**p<0.01, *p<0.05, () indicates the number of the individuals in the population is too small. TSS is total soluble solids.
Table 6.
The fifteen varieties selected for high yield based on genotype (favourable alleles > 15 out of total 19 markers).
Table 6.
The fifteen varieties selected for high yield based on genotype (favourable alleles > 15 out of total 19 markers).
| ID | Company code | Variety name | Favorable allele % |
|---|---|---|---|
| 43 | Pakistan | Tomato seed -2198 | 89.5 |
| 44 | Pakistan | Tomato seed -2199 | 100 |
| 45 | Pakistan | Tomato seed -2217 | 78.9 |
| 46 | Pakistan | Tomato seed -2218 | 89.5 |
| 53 | DRW7806 | hybrid tomato | 100 |
| 55 | Roenza | Roenza | 89.5 |
| 56 | 619 | 619 | 84.2 |
| 57 | SV7846TH | SV7846TH | 89.5 |
| 59 | syngenta | Jarawa Ind tomato | 94.7 |
| 60 | syngenta | Tomato -Beef Vikllio | 84.2 |
| 61 | syngenta | Pilavy Ind tomato | 100 |
| 62 | syngenta | T415271 Ind tomato | 84.2 |
| 63 | syngenta | Tomato Dafnis | 94.7 |
| 64 | syngenta | Tomato Commondo | 100 |
| 65 | syngenta | Tomato Izmono | 89.5 |
Table 7.
Genotypes of heat tolerant lines selected by phenotyping based on yield and quality.
| #43 | #44 | #45 | #47 | #58 | |
|---|---|---|---|---|---|
| SL4-0ch01_1477682 | GA | GA | GA | AA | GG |
| SL4-0ch01_68237408 | TT | TT | TT | TT | TT |
| SL4-0ch05_38882416 | GG | GG | GG | GA | GA |
| SL4-0ch05_58243373 | CC | C/C | C/C | T/T | CT |
| SL4-0ch06_563861 | GA | GA | AA | AA | AA |
| SL4-0ch06_3100027 | GC | GC | GC | GC | GG |
| SL4-0ch06_3629172 | GA | GA | GA | GA | GG |
| SL4-0ch06_8168618 | CA | CA | CA | CC | AA |
| SL4-0ch06_21815322 | CT | CT | CT | TT | CC |
| SL4-0ch06_21898798 | CT | CT | CT | TT | CC |
| SL4-0ch06_22112083 | GT | GT | GT | GG | TT |
| SL4-0ch06_23558883 | CT | CT | CT | CC | TT |
| SL4-0ch06_25025960 | CT | CT | CT | T/T | CC |
| SL4-0ch06_32950646 | GA | GA | GA | GG | GA |
| SL4-0ch08_61071507 | AA | AT | AA | AT | AT |
| SL4-0ch09_6794487 | GA | GG | AA | GA | AA |
| SL4-0ch09_59454323 | TT | GT | TT | TT | TT |
| SL4-0ch12_4886349 | GA | GA | GA | GA | GG |
| SL4-0ch12_15246947 | TT | TT | TT | CT | TT |
| Allele for yield/total markers | 89.5% | 100% | 78.9% | 42.1% | 73.7% |
Yellow highlighted is the less favorable allele for yield; green highlight is favorable for TSS.
Table 8.
Locations of markers in gene body.
| Marker | Genes SNP located | SNP position | gene function |
|---|---|---|---|
| SL4-0ch01_1477682 | Solyc01g006890 | coding | EEIG1/EHBP1 N-terminal domain (C2 domain superfamily) |
| SL4-0ch01_68237408 | Solyc01g067100 | coding | Cullin |
| SL4-0ch05_38882416 | Solyc05g025955 | intron | ABC transporter B family member 11 |
| SL4-0ch05_58243373 | Solyc05g047450 | intron | Methyl-CpG-binding domain-containing protein 2 |
| SL4-0ch06_563861 | Solyc06g005520 | coding | Protein kinase superfamily |
| SL4-0ch06_3100027 | Solyc06g009160 | 3’ UTR | Glyoxysomal fatty acid β-oxidation multifunctional protein MFP-a |
| SL4-0ch06_3629172 | Solyc06g009680 | coding | BRCT domain-containing protein |
| SL4-0ch06_8168618 | Solyc06g011570 | coding | Haloacid dehalogenase-like hydrolase family protein |
| SL4-0ch06_21815322 | Solyc06g034330 | 3’ UTR | unknown |
| SL4-0ch06_21898798 | intergenic region | ||
| SL4-0ch06_22112083 | Solyc06g035450 | 5’ UTR | DEAD-box ATP-dependent RNA helicase |
| SL4-0ch06_23558883 | Solyc06g036240 | 3’ UTR | Pleiotropic drug resistance protein |
| SL4-0ch06_25025960 | Solyc06g036690 | intron | unknown |
| SL4-0ch06_32950646 | Solyc06g051630 | intron | Phosphoinositide phospholipase C |
| SL4-0ch08_61071507 | Solyc08g079440 | coding | UDP-glucuronate 4-epimerase 4 |
| SL4-0ch09_6794487 | Solyc09g014720 | coding | Protein kinase domain |
| SL4-0ch09_59454323 | Solyc09g065300 | coding | spindle pole body-associated protein |
| SL4-0ch12_4886349 | Solyc12g014010 | coding | Glycosyltransferase |
| SL4-0ch12_15246947 | Solyc12g021280 | downstream | Serine/threonine-protein kinase STN7, chloroplastic |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



