
Submitted:
30 August 2024
Posted:
03 September 2024
You are already at the latest version
Abstract
The present work offers a contemporary synthesis of the application of the Oxford Nanopore® MinION™ within the context of cultural heritage biodeterioration investigations, while also providing a comprehensive discussion of possible future perspectives for their utilization in this research field. Owing to the distinctive characteristics of this device, the past few years have witnessed the consistent increase on the application of this system in a variety of cultural heritage materials. In fact, the technique has been useful to understand microbial biodeteriogens diversity and some of their metabolic and biodeteriogenic features. Considering the immense potential for application of this system, this manuscript explores and discusses further possibilities of the technique, focusing on helping understand critical questions on the cultural heritage biodeterioration area. Its application in various differential contexts, has opened doors for prospective applications on other interesting sub-areas of research meriting future investigations, including: biodeteriogens genome and transcriptome analysis, metatranscriptomics, biodeteriorative metabolism studies, inter and intra kingdom interactions analysis, resistome profiling, object history and context studies, in situ applications, bioprospecting and biotechnology.
Keywords:
microbiome
; MinION™
; biodeterioration
; -omics
; next-generation-sequencing
; cultural heritage
1. Introduction
Humanity’s cultural heritage artifacts, relics, and sites are susceptible to undesirable alterations and significant damage due to the growth and metabolic activities of living organisms [1,2,3]. These biodeterioration phenomena can manifest in both indoor settings (e.g., museums) and outdoor environments (e.g., monuments), posing a serious threat to historical sites, properties and objects on a global scale. Various materials, including paper, ceramics, textiles, glass and stone, as well as items such as parchment, books, paintings, frescoes, stained glass, photographs, sculptures and funerary accessories, can be colonized and deteriorated through microorganisms’ aesthetic, mechanical, acidic and enzymatic actions [4]. Consequently, biodeterioration may arise from the influence of multiple organisms (including bacteria, cyanobacteria, microalgae, archaea, fungi and lichens [4]), requiring that protective measures are constantly considered, developed and implemented. Reliant on specific conditions and the substrate type, some of these microorganisms can also contribute to the protection of the materials, thus displaying a biodeteriorative/bioprotective dualistic effect/nature [5,6]. For these reasons, molecular techniques, such as DNA sequencing, have been extensively applied for to investigate, understand and monitor biological colonization on art objects and cultural heritage monuments for more than two decades [7,8,9]. Accordingly, the current focus resides on the application of Next-Generation-Sequencing (NGS) methodologies, since these techniques have powerfully expanded the possibility to characterize microbial communities in a cost-effective, rapid and more comprehensive manner [10,11,12,13,14,15,16]. Owing to the peculiar characteristics of this device, the last few years have seen the steady rise of the popular Oxford Nanopore® MinION™ sequencer in this research area, which has been applied in a variety of cultural heritage materials [8,9,17]. The MinION™ device is a compact (Figure 1), portable, relatively affordable, long-read DNA/RNA sequencer, offering an extensive range of applications. This technology operates on the principle of nanopores (protein-based pores), which function as biosensors to detect negatively charged single-stranded DNA or RNA molecules as they pass through a nanopore driven by an ionic current, with subsequent decoding of electrical variations via computational algorithms [16]. Compared with other NGS platforms, MinION™ offers the advantages of a low startup cost, real-time sequencing, portability and the ability to generate ultra-long reads useful for resolving complex genomic regions. Nonetheless, classical disadvantages associated with the technique have been pointed as lower accuracy and the need for robust bioinformatics tools. In this review article, we aim to provide a brief updated summary of previous studies using the MinION™ sequencer in the area of cultural heritage biodeterioration (CHB), while simultaneously providing a holistic discussion of possible future directions, additional applications and associated impacts of their utilization in this research field.
2. Application of Oxford Nanopore® in Cultural Heritage Biodeterioration Studies
Constrains of conventional cultivation approaches (they primarily detect active forms while neglecting to identify inactive (viable non-culturable) and non-viable states; in vitro culture conditions may preferentially support the proliferation of rapidly growing opportunistic species, thereby hindering the isolation of fastidious species; a substantial proportion of microorganisms exist in symbiotic relationships; and isolated microorganisms may not exhibit the characteristics observed in the original complex communities from which they were derived [4,7]), have hampered an holistic analysis of microorganisms in cultural heritage materials. To address the limitations associated with cultivation, researchers adeptly employed culture-independent molecular biology techniques, such as fingerprinting via denaturing gradient gel electrophoresis (DGGE) or temperature gradient gel electrophoresis (TGGE) in conjunction with clone libraries, or whole-cell hybridization (FISH), across a diverse array of cultural heritage materials [4]. While these methodologies yielded valuable insights and facilitated critical conservation decision-making processes for monuments worldwide, they also encountered the limitation of offering (to varying extents) limited data. In fact, both cultivations paired with molecular analysis, DNA-fingerprinting and the creation of clone libraries, also faced additional constraints, such as the often labor-intensive and time-consuming nature of these experimental protocols and the potential for inherent errors associated with several of these techniques [4,7]. For these reasons, researchers rapidly transitioned to the emerging paradigms of modern high-throughput next-generation sequencing (NGS) technologies that were being developed, introduced and rendered commercially available in the 2000s. Later on, with the development of long read sequencing devices and considering their associated advantages, a strong focus was given to these methodologies. Beginning in 2020, a notable trend observed, with the implementation, assessment and evaluation of the Oxford Nanopore® MinION™ sequencer within the CHB field. In fact, to date the Oxford Nanopore® MinION™ sequencer has been employed across an array of cultural heritage materials, encompassing stone monuments, granite chapels, salt-weathered edifices, petroglyph sites, oil paintings, drawings, textiles, waxes, bronze sculptures, waterlogged archaeological wooden pirogues, iron nails from a whale skeleton, documents and within museum settings (Table 1) [18,19,20,21,22,23,24,25,26,27,28,29,30,31,32,33,34,35,36,37,38,39]. Moreover, the sequencer has progressively evolved into an essential element within the domain of microbial cultural heritage biodeterioration and conservation research [8,9,17,40] and certain patterns regarding the utilization of this technology are already observable. For instance, the prevalence of studies employing a metabarcoding methodology is notable, with the most frequently sequenced genetic regions being the ITS, 16S and 28S rDNA. Concurrently, the predominant subjects of study are related to stone materials and substrates associated with museum environments (Figure 2).
In addition, the most common keywords found in these works include: biodeterioration, metagenomics, nanopore sequencing technology, building materials, stone, microbial community, 16s RNA ribosomal gene, microbiome, bacteria, nanopore sequencing technology, MinION™, microbiota and conservation (Figure 3). Moreover, through an examination of the most frequently occurring terms in the abstracts of these publications (Figure 4), additional insights into the emerging trends associated with the Oxford Nanopore® MinION™ can be further elucidated. Indeed, a significant emphasis in these studies is placed on the capacity of microorganisms to proliferate within museum environments and artworks, the biodeterioration of stone substrates influenced by saline conditions and the resistance mechanisms exhibited by biodeteriogens.
Common workflows for the employment of the MinION™ sequencer within CHB area typically encompass: (1) sampling, DNA extraction and quantification; (2) library preparation according to the selected protocol (including shotgun metagenomics, specific genes PCR or whole genome amplification); and (3) sequencing followed by bioinformatic analysis (Figure 5). Additionally, the library preparation process can incorporate supplementary steps such as barcoding, pooling, DNA fragmentation, DNA end repair, transposase-based adapter ligation, or multiple displacement amplification (MDA) to achieve an adequate yield of DNA [18,19,20,21,22,23,24,25,26,27,28,29,30,31,32,33,34,35,36,37,38,39]. The vast majority of the extant literature employed an amplification step, either via PCR gene amplification or through whole genome amplification (Table 1). However, the studies conducted by Li and colleagues [26] and Nir and colleagues [27] adopted an amplification-free strategy, integrating a MinION™ shotgun metagenomics approach with Illumina sequencing. While shotgun metagenomics methodologies can effectively mitigate PCR bias, further analysis of their inherent characteristics for this sequencer, remains essential. In terms of bioinformatic analysis and the software utilized in CHB investigations, there appears to be a discernible preference for the company’s EPI2ME / “What is in my pot” (WIMP) workflow, which is the most frequently employed approach (Table 2).
From the available literature, several studies utilized gene-targeted sequencing approaches to analyze specific genomic regions such as ITS, 16S and 28S rDNA to study microbial communities associated with cultural heritage artifacts and sites. Grottoli et al. [23] employed a targeted approach using the ITS, 16S, and 18S rDNA regions with the Oxford Nanopore® MinION™ sequencer to study the microbiomes in the Basilica di San Nicola’s hypogeum. They developed the “AmpLIcon SequencIng Analysis” (ALISIA) bioinformatics tool, discovering limited overlap between their sequencing data and culture-dependent methods. Kisová et al. [25] used metabarcoding of ITS, 16S and 28S rDNA to characterize microorganisms on funeral accessories, finding bacteria responsible for metal corrosion and fungi that could degrade textiles. Šoltys et al. [37] sequenced the same regions to study the biodeteriorative microbiomes on an 18th-century wax seal, revealing a diverse community of fungi contributing to lipid alterations and lead soap formation. Other studies focused on the biodeterioration of various cultural materials using similar gene-targeted approaches. For example, Planý et al. [33] analyzed the 16S and 28S rDNA regions to understand microbial communities associated with corroded iron nails from a whale skeleton exhibit. Brimblecombe et al. [20] and Derksen et al. [22] sequenced the ITS region to study fungal contamination in monastic libraries, while Delegou et al. [21] used 16S rDNA sequencing to investigate the microbiota on the Aedicule sepulcher’s building materials. Pavlović et al. [28,29] conducted multiple studies targeting various genes (ITS, 16S rDNA, nirK, dsr, soxB and/or amoA) to analyze microbial influences on salt-contaminated chapels and wax drippings on manuscripts, revealing complex relationships between microbial communities and biodeterioration processes. Additionally, Haedar et al. [24] and Tichy et al. [38] utilized 16S rDNA analysis to explore the microbial influences on patinas and rock paintings, respectively, while Bastholm et al. [36] targeted the calmodulin gene to investigate Aspergillus growth in museum repositories.
On the other hand, Whole-genome amplification (WGA) approaches were employed in several studies to gain a more comprehensive understanding of microbial communities associated with cultural heritage objects. Piñar and associates [31,32] were pioneers in employing a whole genome amplification (WGA) methodology utilizing this sequencer within the context of CHB. This advancement understandably marked a significant progression in comprehending both the microbiome and their potential metabolic characteristics that contribute to the biodeterioration of materials. They applied WGA to study microbiomes on various oil paintings and Leonardo da Vinci’s drawings, while also developing a rapid WGA protocol, revealing significant microbiome differences related to the artworks’ composition and conservation status and also highlighting the impact of geographical factors on microbiome composition (providing a valuable reference for future monitoring). Rabbachin et al. [34,35] also used WGA sequencing to study microbial communities on petroglyphs in the Negev Desert and the Austrian Alps, identifying relationships between stone microbiomes and biodeterioration processes and noting that biofilm removal could enhance microbial recolonization cycles. In another study, Li et al. [26] applied a WGA approach using both Illumina and Nanopore® sequencing to investigate biofilms on stone cultural heritage sites in China. Their findings elucidated the biofilms’ taxonomic profiles and the roles of microbial groups in nitrogen and sulfur metabolism, which contribute to stone deterioration. Nir et al. [27] used the sequencer to characterize a cyanobacterial metagenome from the Negev petroglyphs, while also identifying genes crucial for survival in extreme environments and their contributions to biodeterioration through elemental exchanges. Overall, these whole-genome studies provided a more holistic understanding of the microbial communities and their roles in biodeterioration, offering insights into developing more effective preservation strategies for cultural heritage conservation.
In light of the research that has been conducted so far, some consequently current gaps of the application of the technique in this area can be identified and include the need: (1) to expand the number of studies in supports less studied or not yet explored; (2) to conduct further discussions and comparisons regarding bioinformatic pipelines and analysis; and (3) to proceed with the expansion of application of WGA tailored studies, in order to deepen the knowledge of metabolic and resistance characteristics of microorganisms. Moreover, other additional topics could also be further investigated, such as the case of biodeteriogens metatranscriptomics, microbial ecology, on-site real-time application and molecular monitoring; biotechnology focusing in preservation/restoration efforts; and the monitoring of treatment efficiency through time.
3. Future Directions for Oxford Nanopore® Applications in Biodeterioration Studies
Pavlović and colleagues [17] conducted an initial examination of the first five scholarly articles wherein the aforementioned technique was applied within this particular domain. The authors further postulated that forthcoming investigations could emphasize in the exploration of metabolic and degradative characteristics of cultural heritage microbiomes, the continued establishment of biobanks, the consequences of climate change on microbial populations, the challenges associated with bioinformatics and existing analytical pipelines, as well as the advancements being achieved in sequencing capabilities of the devices. Moreover, over the past five years, the application of this technique across a multitude of differential contexts has also facilitated potential further explorations into other compelling sub-disciplines of research. These encompass, for example: genomic and transcriptomic investigations of biodeteriogens, metatranscriptomic analyses, studies on biodeteriorative metabolic processes, assessments of inter and intra-kingdom interactions, comprehension of resistomes, biocodicological inquiries, analyses of object history and contextual factors, in situ applications, as well as bioprospecting and biotechnological endeavors (Figure 6).
3.1. Biodeteriogens Genomics and Transcriptomics
Genome-wide and transcriptomic investigations can yield significant insights regarding the genetic foundations and microbial mechanisms implicated in the process of biodeterioration. However, a predominant contemporary challenge hindering progress in this field is the scarcity of sequenced microbial genomes coupled with the scarcity of experimental transcriptomic data. For instance, concerning black fungi (which represent a major problematic group of microorganisms affecting stone), only a minimal number of genomes sourced from isolates derived from stone monuments, currently exist [41,42]. Furthermore, the quantity of accessible transcriptomic studies within this domain is considerably much lower.
In a recent investigation, Quach and colleagues [43] illustrated the efficiency of genome sequencing in elucidating lifestyle adaptations and mechanisms of glass biodeterioration exhibited by Curvularia eragrostidis through the Illumina sequencing technique. The authors successfully characterized the metabolic pathways integral to the species’ adaptations to its glassy environment, as well as the genomic underpinnings associated with the biosynthesis of organic acids, exopolysaccharides and enzymatic products that facilitate biodeterioration. Paiva and colleagues [44] similarly employed the Illumina platform to sequence the genome of the microcolonial black fungus Saxispiralis lemnorum and executed an extensive genomic analysis of the Aeminiaceae family. The authors discovered not only various distinctive traits associated with extremotolerance, but also metabolic characteristics that may hypothetically contribute to biodeterioration, including enzymatic properties and capabilities for nitrate assimilation and sulfate reduction. Pei and colleagues [45] investigated the genomic and transcriptomic attributes of Naumannella cuiyingiana AFT2T, which was implicated in the lead-containing pigment discoloration of a 1500-year-old tomb wall painting in China. The authors identified that the presence of Pb ions in the culture medium prompted alterations in gene expression pertaining to metal ion transport and metabolism, as well as to TlpA family protein disulfide reductases. Enzymes exhibiting disulfide-bond reducing capabilities and genes encoding the transporter for divalent Pb ions were also underscored as critical components in the discoloration process of this tomb wall. Wang and colleagues [46] examined the biodegradation mechanisms employed by Fusarium solani NK-NH1 on the hull wood of the Nanhai No. 1 shipwreck. The authors conducted whole genome sequencing of this isolate and successfully characterized the key genes responsible for the degradation of cellulose and lignin.
Currently, the Oxford Nanopore® methodology has demonstrated its capability to sequence entire bacterial and plasmid genomes without necessitating short-read sequencing [47]. In a complementary manner, this technique has also been validated for its ability to procure near-complete, telomere-to-telomere fungal genomes [48,49,50] and with the ongoing advancements in flow cell technology, it is anticipated to yield enhanced outcomes in the imminent future. However, there has been a notable absence of genomic or transcriptomic analyses pertaining to biodeteriogenic microorganisms utilizing this system in CHB studies. Such an approach could represent a compelling and advantageous application of the technique in forthcoming investigations, particularly in light of the long-read capabilities of the device. Their implementation could potentially enhance (alongside metatranscriptomics) the understanding of one of the predominant inquiries in the area: “What do organisms do on and with the object?” [8].
3.2. Metatranscriptomics
While a substantial volume of research employing metagenomics or metabarcoding methodologies exists, there remains a deficiency in metatranscriptomic investigations concentrating on the biodeterioration of cultural heritage materials. As pointed by Sterflinger and Piñar [8], this methodological framework possesses the potential to significantly advance the field, as it would facilitate an understanding of the biodeteriogenic and metabolic repercussions exerted by microorganisms on an object under specified conditions. Zhang and associates [51] have examined the active RNA-level community located at the Beishiku Temple in China utilizing Illumina sequencing data. The researchers identified a concordance between the findings derived from metagenomic and metaproteomic methodologies, thereby affirming the influence of cyanobacteria in the formation of biofilms on stone monuments at this site. While these are rather critical findings, the Oxford Nanopore® sequencer has yet to be employed for metatranscriptomic investigations within this domain, a subject that merits rigorous exploration, as the sequencer has demonstrated its capability to conduct such analyses even under atypical conditions, such as those present at the International Space Station (ISS) [52].
3.3. Biodeteriorative Metabolic Insights
Although a significant number of studies utilizing the sequencer have been executed with a WGA approach (Table 1), the investigations conducted by Li and colleagues [26] and Nir and colleagues [27] have delved more deeply into the biodeteriorative metabolic capabilities inferred from metagenomic analysis data, through a shotgun metagenomics approach. In the first study, this methodology facilitated an in-depth comprehension of the contributions of metabolic pathways to biogeochemical nitrogen and sulfur cycling, which are known to substantially exacerbate stone biodeterioration [53]. The latter investigation concentrated on the acquisition and characterization of the metagenome of a cyanobacteria, yielding insights into genes implicated in element exchanges, specifically ATPases and membrane transporters, which may play a pivotal role in weathering processes. As such, it is evident that, notwithstanding the application of alternative NGS techniques [54,55], such studies are indispensable for elucidating not only the microbial agents accountable for biodeterioration but also their biodeteriorative mechanisms. Consequently, forthcoming studies utilizing this sequencer within these domains may present an extensive array of opportunities and contribute to addressing essential inquiries in the field, specifically regarding the mechanisms underlying biodeterioration in distinct contextual settings.
3.4. Multi-Kingdom Interactions and Microbial Ecology
The biodeterioration of materials constituting cultural heritage may sporadically arise from the influence of a single or a limited ensemble of microorganisms. However, the predominant mechanisms underlying these processes are attributable to a multifaceted microbial consortium that develops and exhibits intra- and inter-species interactions [56,57]. Indeed, the microbiome of cultural heritage materials can be shaped by multi-kingdom mutualistic, competitive, and neutral relationships [56,57], which in turn affect the biodeterioration phenomena that impact these materials. Therefore, an enhanced comprehension of ecological networks can yield essential insights regarding biodeteriorative microbes, thereby aiding in the prediction of their assemblages and assisting in their management, protection and conservation [57].
Liu and colleagues [56] ascertained that interactions across multiple kingdoms dictated the microbiome present within the subterranean Chinese Dahuting Han Dynasty Tomb. The researchers discovered that Actinobacteria and Pseudonocardiaceae, through the emission of volatile geosmin, had the capacity to attract Collembola, which facilitate their dispersal into the tomb’s interior from the surrounding milieu (inter-kingdom mutualism). Additionally, the findings also underscored the occurrence of intra-kingdom competition, as Pseudonocardiaceae flourish due to their production of cellulases and potentially antimicrobial compounds. Yu and associates [57] performed a metabarcoding meta-analysis encompassing approximately 1000 microbiomes sourced from cultural heritage sites situated in diverse environmental contexts. The authors concluded that, on a global scale, bacterial communities are predominantly shaped by overarching climatic factors, whereas the predominance of fungal communities is primarily elucidated by localized habitat conditions. In one of our recent investigations concerning a marble statue [58], we observed some unusual findings, as multiple negative correlations were identified between Bacteria and Fungi at this monument. Given that the majority of the Bacteria identified were Cyanobacteria, one would anticipate the presence of more positive correlations, given their role in fostering the development of heterotrophs.
Acknowledging the significance and necessity of comprehending climatic, and intra- and inter-species ecological interactions to enhance the long-term preservation of cultural heritage materials, prospective studies utilizing the Nanopore® sequencer (employing metagenomics and metatranscriptomics in tandem, for instance) could provide additional pivotal insights into both the microbiome and the metabolic characteristics that facilitate these interactions.
3.5. Cultural Heritage Resistome
The cultural heritage resistome, defined as the antimicrobial resistance genes (AMR or ARGs) present within a community, has recently emerged as a significant area of scholarly interest in this research field [26,59,60]. Understandably, investigations into the AMR genes associated with cultural heritage monuments and artifacts are of considerable importance, as such studies facilitate the comprehension of anthropogenic influences, the evaluation of resistance to preservation and conservation practices, the tracing of contamination sources and even allow the enhancement of public health management initiatives [26,59,60].
Li et al. [26] employed both the Illumina and Oxford Nanopore® sequencing technologies to elucidate the microbiome and pivotal genetic frameworks within the Leshan and Feilaifeng stone heritage sites. The authors innovatively utilized the Nanopore® sequencer within this context for the first time, revealing significant abundances and diversity of genes associated with antibacterial biocides and resistance to metals (specifically copper and quaternary ammonium compounds). Furthermore, they verified that a variety of antimicrobial resistance genes and mobile genetic elements are capable of being disseminated through horizontal gene transfer, ostensibly complicating interventions on these relics. Interestingly, the authors also conducted a comparative analysis of both methodologies, confirming the Nanopore® sequencer capacity to: (1) achieve average read lengths that are considerably longer (8510 bp versus 304,680 bp from Illumina); (2) enhance contig results (9390 versus 542,843 bp from Illumina); and (3) attain an overall superior average contig N50 value in contrast to the exclusive application of Illumina reads [26]. In parallel, He et al. [59] investigated the environmental resistome and mobilome at Feilaifeng stone heritage sites located in Hangzhou, Zhejiang Province, China, utilizing the Illumina sequencing technique. The authors identified a substantial and diverse array of AMR genes that confer resistance to pharmacological agents (antibiotics), biocides, and metals within the studied substrates, inferring that anthropogenic activities likely exert an influence on stone resistomes across various heritage stone areas. Moreover, the authors noted that the stone resistome and mobilome possess the capacity to enhance their adaptability and confer resistance to antimicrobials employed against biodeterioration. Ding et al. [60] examined the pathogenicity characteristics and resistome profiles of the microbiome inhabiting the Angkor sandstone monuments in Cambodia through the Illumina technique. The authors identified a distinct assemblage of ARGs exhibiting cross-niche affiliations between the environmental context and warm-blooded fauna. Additionally, they elucidated the mechanisms of resistance present on these substrates and deliberated on their implications for public health in relation to tourism at cultural heritage sites.
Given the critical nature and necessity of comprehending the resistance mechanisms associated with treatments utilized in the conservation of materials of cultural heritage, as well as the AMR threat to public health, further investigations employing this sequencer could yield profound insights into visitor safety, thereby contributing to a one-health perspective and enhancing conservation endeavors. Recently, this technique has been validated as a rapid, straightforward, and dependable method for detecting such resistances [61] and its application in this field is poised to significantly deepen our understanding of microbiome dynamics and resistance through the monitoring of treatment efficacy.
3.6. Cultural Heritage Objects History
Molecular investigations have been documented to yield significant insights pertaining to the historical context, manufacturing constituents, preservation conditions, geographic provenance and potential adulteration of artistic artifacts [31,32,62,63]. Furthermore, such studies may offer critical evidence regarding the interactions between these relics and human engagement [62]. Consequently, they hold substantial relevance in disciplines such as biocodicology, archaeology, criminology, as well as in assessing human contamination resulting from physical handling [31,32,64,65]. Although this remains an emergent field, efforts have been made to utilize the Oxford Nanopore® sequencer for the identification of the microbiome and ancient animal DNA (aDNA) derived from a 15th-century parchment housed within the Graphic Collection (Kupferstichkabinett) of the Academy of Fine Arts in Vienna [63]. Conversely, the application of these methodologies has the potential to enrich our comprehension of object context and to elucidate the dualistic influences of the microbiome, encompassing both biodeteriorative and bioprotective aspects [65]. It is reasonable to assert that this sub-discipline may pave the way for numerous applications of the sequencer that merit further investigation.
3.7. On-Site Application
Owing to the portability, operational features, versatility and analytical velocity of the sequencer, the platform facilitates on-site analytical procedures. Indeed, currently a wide range of reports documenting such applications across diverse environmental contexts, including Ebola surveillance in West Africa, water quality assessment, environmental microbiota analysis, SARS-CoV-2 diagnostics and even aboard the International Space Station (ISS) [66,67,68,69,70,71,72,73,74], are available. The methodology has further proven to be dependable in resource-constrained environments [72], a condition that is likewise applicable to various cultural heritage sites globally. Moreover, WGS analysis is poised to enhance the study of microbial threats to historical properties within developing nations. Nonetheless, the potential of on-site capabilities within this specific research domain remains to be thoroughly investigated. Recently, Tamames and collaborators [74] designed and evaluated an in-situ protocol for Oxford Nanopore® metagenomic analysis in volcanic rocks (La Palma, Canary Islands) and marine environments, achieving results in less than one day. Given such advancements, the deployment of this technique for on-site analysis can present an extensive array of opportunities within the realm of cultural heritage biodeterioration, including the surveillance of microbial outbreaks and the implementation of rapid control measures [75,76]. In addition, while culture-dependent approaches encounter limitations in delivering a comprehensive understanding of the microbiome responsible for the biodeterioration of artifacts, they nonetheless provide valuable insights as they facilitate the isolation of microbes for differential analysis, discovery of novel microorganisms and enhancement of biological databases [77,78]. The sequencer has also proven effective for in situ bioprospecting initiatives and microbial characterization [79], which, considering the frequently uncharacterized elements of the microbiome associated with heritage materials [78], could present valuable avenues for future alternative research paths.
3.8. Biotechnology and Restoration Efforts
From a biotechnological standpoint, green intervention strategies such as bioconsolidation and biocleaning [78] may significantly benefit from the integration of this sequencer. Bioconsolidation is defined as the array of methodologies that involve the utilization of bacteria to remediate construction materials, emulating the natural bacterial precipitation of carbonates found in carbonate rock formations [80,81,82]. Conversely, biocleaning is characterized by the investigation of microbial metabolic processes that facilitate the removal of sulfates, nitrates and organic matter, aiming to restore or enhance compromised stone surfaces [83]. In this context, the Oxford Nanopore® sequencer could serve as a valuable tool for investigating and gaining deeper insights into the mechanisms underlying bioconsolidation and biocleaning through genomic and transcriptomic analyses, which could subsequently pave the way for genomic manipulation and enhancement of these beneficial properties. Furthermore, contributions towards the potential implementation and enhancement of alternative microbial-derived methodologies, including the utilization of antagonistic microorganisms or bacteria capable of facilitating bioremoval, may also derive benefits from this system [84,85,86,87]. Finally, the application of this methodology to evaluate the efficacy of biocide treatments also presents a promising perspective, as its utility has already been established with the Illumina system [88].
4. Challenges in the Oxford Nanopore® Application in Biodeterioration Studies
While the application of sequencers offers a multitude of benefits for the investigation of microbial communities present within cultural heritage materials, it is imperative to recognize several inherent limitations. Notably, it is essential to consider that the comparatively high costs associated with this technology may pose significant challenges for certain researchers, alongside the difficulties encountered in securing appropriate reagent storage in resource-limited or remote environments, as well as the requisite for robust internet connectivity to facilitate data analysis, which may impede the broader implementation of this methodology [72]. Conversely, the occurance of sequencing inaccuracies and the intricate nature of bioinformatics pipelines necessary for data analysis, can pose additional obstacles for researchers employing this technique. At present, despite not being devoid of errors, a substantial portion of the challenges associated with long-read sequencing has been to some degree, effectively mitigated through the advent of various error correction software developed in recent years [89], as well as the innovation of new flow cells. Indeed, the Oxford Nanopore® systems rank among the most rapidly evolving NGS platforms currently available, with their limitations being systematically enhanced. For instance, the newly developed flow cell R10.4.1 facilitates quality scores of Q20+, achieves exceptionally high accuracy levels (frequently exceeding 99%), and even accommodates Short Fragmented Mode (SFM) applications [64,90,91,92,93], which, in turn, can provide an expanded array of analytical possibilities. However, there exists a lesser degree of consensus regarding the bioinformatic analysis of the obtained data. Existing constraints and critiques encompass: the necessity for user-friendly bioinformatics platforms; the substantial volume of data generated, which can impose significant demands on IT resources [73]; and the accessibility of methods and comprehensive databases pertinent to these analyses [88]. Furthermore, the standard bioinformatics platforms, such as Epi2me, relies on the availability of an internet connection, although offline applications have also been introduced [94]. Nevertheless, the ongoing evaluation of alternative bioinformatic pipelines and databases both within and beyond the cultural heritage biodegradation domain [19,23,95,96,97,98,99,100,101,102] represents a continual endeavor that will undoubtedly require further analytical scrutiny and standardization in the foreseeable future. Moreover, while MinION™ provides advantages in terms of portability and capabilities for in-field analysis, the protocols for sample preparation may necessitate optimization to guarantee reproducibility and dependable outcomes in non-laboratory environments. Conversely, and to a lesser extent, the challenges associated with executing routine procedures in situ, such as suboptimal DNA recovery from environmental samples, warrant acknowledgment [74]. Although protocols aimed at enhancing DNA extraction efficiency (e.g., [103]) have been formulated, they require further validation within contexts of CHB. Addressing these limitations is undeniably critical to effectively ensure the applicability of nanopore sequencing [89] in studies related to cultural heritage biodeterioration, as well as to advance our comprehension of microbial interactions and degradation processes that affect cultural heritage artifacts.
5. Conclusions
The utilization of Oxford Nanopore® MinION™ sequencing technology in the field of cultural heritage biodeterioration research has thus far revealed significant potential and adaptability. This technology has been proficiently employed across a diverse array of materials, facilitating the examination of various microbial communities and certain metabolic pathways implicated in their biodeterioration processes. Current trends within this domain indicate a growing predisposition towards studies that focus on gene amplification and an increased emphasis on materials associated with stone. Prospective applications of this technique appear promising, as the characteristics of the sequencer enable its use for in situ analyses and bioprospecting; for comprehensive genomic, transcriptomic and environmental metatranscriptomic investigations of microorganisms to elucidate active metabolic processes; to explore inter- and intra-kingdom microbial interactions; and to reconstruct the historical context of objects. Moreover, resistome analysis of cultural heritage materials can yield critical insights into antimicrobial resistance, thereby informing the development and implementation of enhanced conservation strategies. In addition, the application of this technology for biotechnology, holds the potential to refine innovative conservation methodologies. In summary, this sequencer has established itself as an indispensable instrument in the domain of cultural heritage conservation and the exploration of novel applications within this area can significantly aid in the safeguarding of cultural heritage artifacts and sites for forthcoming generations.
Author Contributions
Conceptualization, J.T.; formal analysis, J.T. and F.S.; investigation, J.T. and F.S.; writing—original draft preparation, J.T.; writing—review and editing, F.S.; project administration, J.T. and F.S.; funding acquisition, J.T.; All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by Santander/University of Coimbra in the framework of the SeedProjects@UC 2024 with the reference PT0054.A.01.C. This work was carried out at the R&D Unit Centre for Functional Ecology—Science for People & the Planet (CFE) and Associate Laboratory TERRA. The Centre for Functional Ecology—Science for People and the Planet (CFE) was supported by FCT—Fundação para a Ciência e Tecnologia, I.P. by project reference UIDB/04004/2020 and the DOI identifier 10.54499/UIDB/04004/2020 (https://doi.org/10.54499/UIDB/04004/2020). The Associate Laboratory TERRA was supported by FCT—Fundação para a Ciência e Tecnologia, I.P. by project reference LA/P/0092/2020 and the DOI identifier 10.54499/LA/P/0092/2020 (https://doi.org/10.54499/LA/P/0092/2020). The authors also thank the funding of PRR—Recovery and Resilience Plan and by the NextGeneration EU European Funds.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Hueck, H. J., The biodeterioration of materials as part of hylobiology. Mater Org, 1965, 1, 1. 5– 34.
- Hueck, H.J., The biodeterioration of materials—an appraisal. In Biodeterioration of Materials. Walters, A.H., Elphick, J.S., Eds, Elsevier, London, UK, 1968, 6–12.
- Urzì, C., Krumbein. W.E., Microbiological impacts on the cultural heritage. In Durability and change: the science, responsability and cost of sustaining cultural heritage. Krumbein, W.E., Brimblecombe, P., Cosgrove D.E., Stainforth, S., Eds, John Wiley & Sons Ltd., London, UK, 1994, 107–135.
- Sterflinger, K.; Piñar, G. Microbial Deterioration of Cultural Heritage and Works of Art--Tilting at Windmills? Appl Microbiol Biotechnol, 2013, 97, 9637–9646. [CrossRef]
- Favero-Longo, S.E.; Viles, H.A. A Review of the Nature, Role and Control of Lithobionts on Stone Cultural Heritage: Weighing-up and Managing Biodeterioration and Bioprotection. World J. Microbiol. Biotechnol. 2020, 36, 100. [CrossRef]
- Liu, X.; Qian, Y.; Wu, F.; Wang, Y.; Wang, W.; Gu, J.-D. Biofilms on Stone Monuments: Biodeterioration or Bioprotection? Trends Microbiol, 2022, 30, 816–819. [CrossRef]
- Beata, G. The Use of -Omics Tools for Assessing Biodeterioration of Cultural Heritage: A Review. J Cult Herit, 2020, 45, 351–361. [CrossRef]
- Sterflinger, K.; Piñar, G. Molecular-Based Techniques for the Study of Microbial Communities in Artworks. In Microorganisms in the Deterioration and Preservation of Cultural Heritage; Joseph, E., Ed.; Springer International Publishing: Cham, 2021, 59–77 ISBN 978-3-030-69411-1.
- Piñar, G.; Sterflinger, K. Natural Sciences at the Service of Art and Cultural Heritage: An Interdisciplinary Area in Development and Important Challenges. Microb Biotechnol, 2021, 14, 806–809. [CrossRef]
- Quail, M.A.; Smith, M.; Coupland, P.; Otto, T.D.; Harris, S.R.; Connor, T.R.; Bertoni, A.; Swerdlow, H.P.; Gu, Y. A tale of three next generation sequencing platforms: Comparison of Ion Torrent, Pacific Biosciences and Illumina MiSeq sequencers. BMC Genom, 2012, 13, 341. [CrossRef]
- Shokralla, S.; Spall, J.L.; Gibson, J.F.; Hajibabaei, M. Next-generation sequencing technologies for environmental DNA research. Mol Ecol, 2012, 21, 1794–1805. [CrossRef]
- Buermans, H.P.J.; den Dunnen, J.T. Next generation sequencing technology: Advances and applications. Biochim Biophys Acta, 2014, 1842, 1932–1941. [CrossRef]
- Goodwin, S.; McPherson, J.D.; McCombie, W.R. Coming of age: Ten years of next-generation sequencing technologies. Nat Rev Genet, 2016, 17, 333–351. [CrossRef]
- Amarasinghe, S.L.; Su, S.; Dong, X.; Zappia, L.; Ritchie, M.E.; Gouil, Q. Opportunities and challenges in long-read sequencing data analysis. Genome Biol, 2020, 21, 30. [CrossRef]
- Mohammed, A.A.; Senbeta, B.; Worku, T.; Mohammed, A.A.; Senbeta, B.; Worku, T. Pacific bioscience sequence technology: Review. Int J Vet Sci Res, 2022, 8, 27–33. [CrossRef]
- Wang, Y.; Zhao, Y.; Bollas, A.; Wang, Y.; Au, K.F. Nanopore sequencing technology, bioinformatics and applications. Nat Biotechnol, 2021, 39, 1348–1365. [CrossRef]
- Pavlovic, J.; Cavalieri, D.; Mastromei, G.; Pangallo, D.; Perito, B.; Marvasi, M. MinION Technology for Microbiome Sequencing Applications for the Conservation of Cultural Heritage. Microbiol Res, 2021, 247, 126727. [CrossRef]
- Bastholm, C.J.; Andersen, B.; Frisvad, J.C.; Oestergaard, S.K.; Nielsen, J.L.; Madsen, A.M.; Richter, J. A Novel Contaminant in Museums? A Cross-Sectional Study on Xerophilic Aspergillus Growth in Climate-Controlled Repositories. Sci Total Environ, 2024, 173880. [CrossRef]
- Beccaccioli, M.; Moricca, C.; Faino, L.; Reale, R.; Mineo, M.; Reverberi, M. The Neolithic Site “La Marmotta”: DNA Metabarcoding to Identify the Microbial Deterioration of Waterlogged Archeological Wood. Front Microbiol, 2023, 14. [CrossRef]
- Brimblecombe, P.; Sterflinger, K.; Derksen, K.; Haltrich, M.; Querner, P. Thermohygrometric Climate, Insects and Fungi in the Klosterneuburg Monastic Library. Heritage, 2022, 5, 4228–4244. [CrossRef]
- Delegou, E.; Karapiperis, C.; Hilioti, Z.; Chasapi, A.; Valasiadis, D.; Alexandridou, A.; Rihani, V.; Kroustalaki, M.; Bris, T.; Ouzounis, C.; et al. Metagenomics of the built cultural heritage: microbiota characterization of the building materials of the holy aedicule of the holy sepulchre in Jerusalem. CIENTIFIC CULTURE, 2022, 8(2), 59-83. [CrossRef]
- Derksen, K.; Brimblecombe, P.; Piñar, G.; Waldherr, M.; Graf, A.; Haltrich, M.; Querner, P.; Sterflinger, K. Fungal Biodeterioration Risk in Monastic Libraries without Climate Control. Microorganisms, 2024, 12, 1450. [CrossRef]
- Grottoli, A.; Beccaccioli, M.; Zoppis, E.; Fratini, R.S.; Schifano, E.; Santarelli, M.L.; Uccelletti, D.; Reverberi, M. Nanopore Sequencing and Bioinformatics for Rapidly Identifying Cultural Heritage Spoilage Microorganisms. Front Mater, 2020, 7. [CrossRef]
- Haedar, N.; Iqram, M.; Priosambodo, D.; Lebe, R. Bacterial Communities on Degraded Prehistoric Rock Paintings in Maros-Pangkep Global Geopark. Philippine Journal of Science, 2024, 153(1): 391-402.
- Kisová, Z.; Planý, M.; Pavlović, J.; Bučková, M.; Puškárová, A.; Kraková, L.; Kapustová, M.; Pangallo, D.; Šoltys, K. Biodeteriogens Characterization and Molecular Analyses of Diverse Funeral Accessories from XVII Century. Appl Sci, 2020, 10, 5451. [CrossRef]
- Li, Q.; Wu, C.; He, J.; Zhang, B. Unraveling the Microbiotas and Key Genetic Contexts Identified on Stone Heritage Using Illumina and Nanopore Sequencing Platforms. Int Biodeterior Biodegradation, 2023, 185. [CrossRef]
- Nir, I.; Barak, H.; Rabbachin, L.; Arielle, K.; Pavan, M.; Winter, E.; Pinar, G.; Sterflinger, K.; Ariel, K. Trichocoleus Desertorum Isolated from Negev Desert Petroglyphs: Characterization, Adaptation and Bioerosion Potential. The Sci Total Environ, 2023, 904, 166739. [CrossRef]
- Pavlović, J.; Bosch-Roig, P.; Rusková, M.; Planý, M.; Pangallo, D.; Sanmartín, P. Long-Amplicon MinION-Based Sequencing Study in a Salt-Contaminated Twelfth Century Granite-Built Chapel. Appl Microbiol Biotechnol, 2022, 106, 4297–4314. [CrossRef]
- Pavlović, J.; Sclocchi, M.C.; Planý, M.; Ruggiero, D.; Puškárová, A.; Bučková, M.; Šoltys, K.; Colaizzi, P.; Riccardi, M.L.; Pangallo, D.; et al. The Microbiome of Candle Beeswax Drops on Ancient Manuscripts. Int Biodeterior Biodegradation, 2022, 174, 105482. [CrossRef]
- Pavlović, J.; Puškárová, A.; Planý, M.; Farkas, Z.; Rusková, M.; Kvalová, K.; Kraková, L.; Bučková, M.; Pangallo, D. Colored Stains: Microbial Survey of Cellulose-Based and Lignin Rich Papers. Int J Biol Macromol, 2023, 241, 124456. [CrossRef]
- Pinar, G.; Poyntner, C.; Lopandic, K.; Tafer, H.; Sterflinger, K. Rapid Diagnosis of Biological Colonization in Cultural Artefacts Using the MinION Nanopore Sequencing Technology. Int Biodeterior Biodegradation, 2020, 148. [CrossRef]
- Piñar, G.; Sclocchi, M.C.; Pinzari, F.; Colaizzi, P.; Graf, A.; Sebastiani, M.L.; Sterflinger, K. The Microbiome of Leonardo Da Vinci’s Drawings: A Bio-Archive of Their History. Front Microbiol, 2020, 11. [CrossRef]
- Planý, M.; Pinzari, F.; Šoltys, K.; Kraková, L.; Cornish, L.; Pangallo, D.; Jungblut, A.D.; Little, B. Fungal-Induced Atmospheric Iron Corrosion in an Indoor Environment. Int Biodeterior Biodegradation, 2021, 159, 105204. [CrossRef]
- Rabbachin, L.; Piñar, G.; Nir, I.; Kushmaro, A.; Pavan, M.J.; Eitenberger, E.; Waldherr, M.; Graf, A.; Sterflinger, K. A Multi-Analytical Approach to Infer Mineral–Microbial Interactions Applied to Petroglyph Sites in the Negev Desert of Israel. Appl Sci, 2022, 12, 6936. [CrossRef]
- Rabbachin, L.; Pinar, G.; Nir, I.; Kushmaro, A.; Eitenberger, E.; Waldherr, M.; Graf, A.; Sterflinger, K. Natural Biopatina on Historical Petroglyphs in the Austrian Alps: To Clean or Not to Clean? Int Biodeterior Biodegradation, 2023, 183, 105632. [CrossRef]
- Rabbachin, L.; Nir, I.; Waldherr, M.; Vassallo, Y.; Piñar, G.; Graf, A.; Kushmaro, A.; Sterflinger, K. Diversity of Fungi Associated with Petroglyph Sites in the Negev Desert, Israel, and Their Potential Role in Bioweathering. Front Fungal Biol, 2024, 5. [CrossRef]
- Šoltys, K.; Planý, M.; Biocca, P.; Vianello, V.; Bučková, M.; Puškárová, A.; Sclocchi, M.C.; Colaizzi, P.; Bicchieri, M.; Pangallo, D.; et al. Lead Soaps Formation and Biodiversity in a XVIII Century Wax Seal Coloured with Minium. Environ Microbiol, 2020, 22, 1517–1534. [CrossRef]
- Tichy, J.; Waldherr, M.; Ortbauer, M.; Graf, A.; Sipek, B.; Jembrih-Simbuerger, D.; Sterflinger, K.; Piñar, G. Pretty in Pink? Complementary Strategies for Analysing Pink Biofilms on Historical Buildings. Sci Total Environ, 2023, 904, 166737. [CrossRef]
- Timoncini, A.; Costantini, F.; Bernardi, E.; Martini, C.; Mugnai, F.; Mancuso, F.; Sassoni, E.; Ospitali, F.; Chiavari, C. Insight on Bacteria Communities in Outdoor Bronze and Marble Artefacts in a Changing Environment. Sci Total Environ, 2022, 850, 157804. [CrossRef]
- Marvasi, M.; Pangallo, D.; Cavalieri, D.; Poyatos-Jiménez, F. Editorial: Multi-Omics Revolution in Microbial Cultural Heritage Conservation. Front Microbiol, 2021, 12. [CrossRef]
- De Leo, F.; Marchetta, A.; Urzì, C. Black Fungi on Stone-Built Heritage: Current Knowledge and Future Outlook. Appl Sci, 2022, 12, 3969. [CrossRef]
- Trovão, J.; Tiago, I.; Soares, F.; Paiva, D.S.; Mesquita, N.; Coelho, C.; Catarino, L.; Gil, F.; Portugal, A. High-Quality Draft Genome Sequence of the Microcolonial Black Fungus Aeminium Ludgeri DSM 106916. Microbiol Resour Announc, 2019, 8, e00202-19. [CrossRef]
- Quach, N.T.; Ngo, C.C.; Nguyen, T.H.; Nguyen, P.L.; Vu, T.H.N.; Phan, T.H.T.; Nguyen, Q.H.; Le, T.T.M.; Chu, H.H.; Phi, Q.-T. Genome-Wide Comparison Deciphers Lifestyle Adaptation and Glass Biodeterioration Property of Curvularia Eragrostidis C52. Sci Rep, 2022, 12, 11411. [CrossRef]
- Paiva, D.S.; Fernandes, L.; Portugal, A.; Trovão, J. First Genome Sequence of the Microcolonial Black Fungus Saxispiralis Lemnorum MUM 23.14: Insights into the Unique Genomic Traits of the Aeminiaceae Family. Microorganisms, 2024, 12, 104. [CrossRef]
- Pei, S.; Wu, F.; Chen, Y.; Ma, W.; He, D.; Zhang, Q.; Gu, J.-D.; Wang, W.; Tian, T.; Feng, H. Mechanisms of Lead-Containing Pigment Discoloration Caused by Naumannella Cuiyingiana AFT2T Isolated from 1500 Years Tomb Wall Painting of China. Int Biodeterior Biodegradation, 2023, 185, 105689. [CrossRef]
- Wang, Y.; Han, Y.; Li, N.; Wang, C.; Ma, K.; Huang, X.; Du, J.; Guo, H.; Pan, J. Study on Biodegradation Mechanism of Fusarium Solani NK-NH1 on the Hull Wood of the Nanhai No. 1 Shipwreck. Front Microbiol, 2024, 15, 1382653. [CrossRef]
- Zhao, W.; Zeng, W.; Pang, B.; Luo, M.; Peng, Y.; Xu, J.; Kan, B.; Li, Z.; Lu, X. Oxford Nanopore Long-Read Sequencing Enables the Generation of Complete Bacterial and Plasmid Genomes without Short-Read Sequencing. Front Microbiol, 2023, 14. [CrossRef]
- Salazar, A.N.; Gorter de Vries, A.R.; van den Broek, M.; Wijsman, M.; de la Torre Cortés, P.; Brickwedde, A.; Brouwers, N.; Daran, J.-M.G.; Abeel, T. Nanopore Sequencing Enables Near-Complete de Novo Assembly of Saccharomyces Cerevisiae Reference Strain CEN.PK113-7D. FEMS Yeast Res, 2017, 17, fox074. [CrossRef]
- McGinnis, J.L.; Giguere, D.J. High-Quality Genome Assembly of a Pestalotiopsis Fungus Using DIY-Friendly Methods 2022. [version 1; peer review: 3 approved with reservations]. F1000Research, 2022, 11:442 . [CrossRef]
- Witte, T.E.; Hicks, C.; Shoukouhi, P.; Dadej, K.; Findlay, W.; Liu, M.; Overy, D.P. Chromosome-Level Draft Genome Sequences of Three Isolates of the Toxigenic Fungus Claviceps Purpurea Showing Structural Rearrangements. Microbiol Resour Announc, 2023, 12, e00234-23. [CrossRef]
- Zhang, Y.; Wu, F.; Gu, J.-D.; He, K.; Fang, Z.; Liu, X.; He, D.; Ding, X.; Li, J.; Han, Z.; et al. Dominance by Cyanobacteria in the Newly Formed Biofilms on Stone Monuments under a Protective Shade at the Beishiku Temple in China. Env Res, 2024, 251, 118576. [CrossRef]
- Haveman, N.J.; Khodadad, C.L.M.; Dixit, A.R.; Louyakis, A.S.; Massa, G.D.; Venkateswaran, K.; Foster, J.S. Evaluating the Lettuce Metatranscriptome with MinION Sequencing for Future Spaceflight Food Production Applications. npj Microgravity, 2021, 7, 1–11. [CrossRef]
- Liu, X.; Koestler, R.J.; Warscheid, T.; Katayama, Y.; Gu, J.-D. Microbial Deterioration and Sustainable Conservation of Stone Monuments and Buildings. Nat Sustain, 2020, 3, 991–1004. [CrossRef]
- Wu, F.; Ding, X.; Zhang, Y.; Gu, J.-D.; Liu, X.; Guo, Q.; Li, J.; Feng, H. Metagenomic and Metaproteomic Insights into the Microbiome and the Key Geobiochemical Potentials on the Sandstone of Rock-Hewn Beishiku Temple in Northwest China. Sci Total Environ, 2023, 893, 164616. [CrossRef]
- Qian, Z.; Li, Y.; Pratush, A.; Kan, J.; Gu, J.-D.; Peng, T.; Huang, T.; Hu, Z. A Comparative Analysis of the Microbial Communities and Functional Genes of the Nitrogen Cycling in Mangroves of China, Indian and Malaysia. Int Biodeterior Biodegradation, 2024, 190, 105767. [CrossRef]
- Liu, W.; Zhou, X.; Jin, T.; Li, Y.; Wu, B.; Yu, D.; Yu, Z.; Su, B.; Chen, R.; Feng, Y.; et al. Multikingdom Interactions Govern the Microbiome in Subterranean Cultural Heritage Sites. PNAS, 2022, 119, e2121141119. [CrossRef]
- Yu, Y.; Zhang, J.; Chen, R.; Coleine, C.; Liu, W.; Delgado-Baquerizo, M.; Feng, Y. Unearthing the Global Patterns of Cultural Heritage Microbiome for Conservation. Int Biodeterior Biodegradation, 2024, 190, 105784. [CrossRef]
- Trovão, J.; Portugal, A. The Impact of Stone Position and Location on the Microbiome of a Marble Statue. The Microbe, 2024, 2, 100040. [CrossRef]
- He, J.; Zhang, N.; Shen, X.; Muhammad, A.; Shao, Y. Deciphering Environmental Resistome and Mobilome Risks on the Stone Monument: A Reservoir of Antimicrobial Resistance Genes. Sci Total Environ, 2022, 838, 156443. [CrossRef]
- Ding, X.; Lan, W.; Li, J.; Deng, M.; Li, Y.; Katayama, Y.; Gu, J.-D. Metagenomic Insight into the Pathogenic-Related Characteristics and Resistome Profiles within Microbiome Residing on the Angkor Sandstone Monuments in Cambodia. Sci Total Environ, 2024, 918, 170402. [CrossRef]
- Solcova, M.; Demnerova, K.; Purkrtova, S. Application of Nanopore Sequencing (MinION) for the Analysis of Bacteriome and Resistome of Bean Sprouts. Microorganisms, 2021, 9, 937. [CrossRef]
- Piñar, G.; Poyntner, C.; Tafer, H.; Sterflinger, K. A Time Travel Story: Metagenomic Analyses Decipher the Unknown Geographical Shift and the Storage History of Possibly Smuggled Antique Marble Statues. Ann Microbio,l 2019, 69, 1001–1021. [CrossRef]
- Vassallo, Y.; Waldherr, M.; Lehner, E.; Graf, A.; Cappa, F.; Hartl, A.; Schober, R.; Beccaccioli, M.; Sterflinger, K.; Piñar, G.; et al. Oxford Nanopore Technologies for Biocodicology: A Case Study on a 15th-Century Parchment. In DTC Lazio: Tecnologie e patrimonio culturale: nuove competenze e professioni. 2023. https://iris.uniroma1.it/handle/11573/1695400?mode=complete.
- Simon, L.M.; Flocco, C.; Burkart, F.; Methner, A.; Henke, D.; Rauer, L.; Müller, C.L.; Vogel, J.; Quaisser, C.; Overmann, J.; et al. Microbial Fingerprints Reveal Interaction between Museum Objects, Curators, and Visitors. iScience, 2023, 26, 107578. [CrossRef]
- Cao, Y.; Bowker, M.A.; Delgado-Baquerizo, M.; Xiao, B. Biocrusts Protect the Great Wall of China from Erosion. Sci Adv, 2023, 9, eadk5892. [CrossRef]
- Castro-Wallace, S.L.; Chiu, C.Y.; John, K.K.; Stahl, S.E.; Rubins, K.H.; McIntyre, A.B.R.; Dworkin, J.P.; Lupisella, M.L.; Smith, D.J.; Botkin, D.J.; et al. Nanopore DNA Sequencing and Genome Assembly on the International Space Station. Sci Rep, 2017, 7, 18022. [CrossRef]
- Goordial, J.; Altshuler, I.; Hindson, K.; Chan-Yam, K.; Marcolefas, E.; Whyte, L.G. In Situ Field Sequencing and Life Detection in Remote (79°26′N) Canadian High Arctic Permafrost Ice Wedge Microbial Communities. Front Microbiol, 2017, 8. [CrossRef]
- Harcourt, J.; Tamin, A.; Lu, X.; Kamili, S.; Sakthivel, S.K.; Murray, J.; Queen, K.; Tao, Y.; Paden, C.R.; Zhang, J.; et al. Isolation and Characterization of SARS-CoV-2 from the First US COVID-19 Patient. bioRxiv, 2020, 2020.03.02.972935.
- Latorre-Pérez, A.; Pascual, J.; Porcar, M.; Vilanova, C. A Lab in the Field: Applications of Real-Time, in Situ Metagenomic Sequencing. Biology Methods and Protocols, 2020, 5, bpaa016. [CrossRef]
- Moore, S.C.; Penrice-Randal, R.; Alruwaili, M.; Dong, X.; Pullan, S.T.; Carter, D.P.; Bewley, K.; Zhao, Q.; Sun, Y.; Hartley, C.; et al. Amplicon Based MinION Sequencing of SARS-CoV-2 and Metagenomic Characterisation of Nasopharyngeal Swabs from Patients with COVID-19 bioRxiv, 2020, 2020.03.05.20032011.
- Quick, J.; Loman, N.J.; Duraffour, S.; Simpson, J.T.; Severi, E.; Cowley, L.; Bore, J.A.; Koundouno, R.; Dudas, G.; Mikhail, A.; et al. Real-Time, Portable Genome Sequencing for Ebola Surveillance. Nature, 2016, 530, 228–232. [CrossRef]
- Wasswa, F.B.; Kassaza, K.; Nielsen, K.; Bazira, J. MinION Whole-Genome Sequencing in Resource-Limited Settings: Challenges and Opportunities. Curr Clin Micro Rpt, 2022, 9, 52–59. [CrossRef]
- Werner, D.; Acharya, K.; Blackburn, A.; Zan, R.; Plaimart, J.; Allen, B.; Mgana, S.M.; Sabai, S.M.; Halla, F.F.; Massawa, S.M.; et al. MinION Nanopore Sequencing Accelerates Progress towards Ubiquitous Genetics in Water Research. Water, 2022, 14, 2491. [CrossRef]
- Tamames, J.; Jiménez-Lalana, D.; Redondo, Á.; Martínez-García, S.; De Los Rios, A. In Situ Metagenomics: A Platform for Rapid Sequencing and Analysis of Metagenomes in Less than One Day. Mol Ecol Resour, 2024, 24, e13909. [CrossRef]
- Bastholm, C.J.; Madsen, A.M.; Andersen, B.; Frisvad, J.C.; Richter, J. The Mysterious Mould Outbreak—A Comprehensive Fungal Colonisation in a Climate-Controlled Museum Repository Challenges the Environmental Guidelines for Heritage Collections. J Cult Herit, 2022, 55, 78–87. [CrossRef]
- Martin-Pozas, T.; Nováková, A.; Jurado, V.; Cuezva, S.; Fernandez-Cortes, A.; Saiz-Jimenez, C.; Sanchez-Moral, S. A Second Fungal Outbreak in Castañar Cave, Spain, Discloses the Fragility of Subsurface Ecosystems. Microb Ecol, 2024, 87, 53. [CrossRef]
- Trovão, J.; Portugal, A. Current Knowledge on the Fungal Degradation Abilities Profiled through Biodeteriorative Plate Essays. Appl Sci, 2021, 11, 4196. [CrossRef]
- Pyzik, A.; Ciuchcinski, K.; Dziurzynski, M.; Dziewit, L. The Bad and the Good—Microorganisms in Cultural Heritage Environments—An Update on Biodeterioration and Biotreatment Approaches. Materials, 2021, 14, 177. [CrossRef]
- Latorre-Pérez, A.; Gimeno-Valero, H.; Tanner, K.; Pascual, J.; Vilanova, C.; Porcar, M. A Round Trip to the Desert: In Situ Nanopore Sequencing Informs Targeted Bioprospecting. Front Microbiol, 2021, 12. [CrossRef]
- Andreolli, M.; Lampis, S.; Bernardi, P.; Calò, S.; Vallini, G. Bacteria from Black Crusts on Stone Monuments Can Precipitate CaCO3 Allowing the Development of a New Bio-Consolidation Protocol for Ornamental Stone. Int Biodeterior Biodegradation, 2020, 153, 105031. [CrossRef]
- Dhami, N.K.; Reddy, M.S.; Mukherjee, A. Application of Calcifying Bacteria for Remediation of Stones and Cultural Heritages. Front Microbiol, 2014, 5. [CrossRef]
- Reddy, M.S. Biomineralization of Calcium Carbonates and Their Engineered Applications: A Review. Front Microbiol, 2013, 4. [CrossRef]
- Cappitelli, F. Biocleaning of Cultural Heritage Surfaces. Open Conf Proc J, 2016, 7. [CrossRef]
- Ranalli, G.; Zanardini, E. Biocleaning on Cultural Heritage: New Frontiers of Microbial Biotechnologies. Journal of Applied Microbiology, 2021, 131, 583–603. [CrossRef]
- Bosch-Roig, P.; Sanmartín, P. Bioremoval of Graffiti in the Context of Current Biocleaning Research. In Microorganisms in the Deterioration and Preservation of Cultural Heritage; Joseph, E., Ed.; Springer International Publishing: Cham, 2021, 175–197 ISBN 978-3-030-69411-1.
- Cattò, C.; Sanmartín, P.; Gulotta, D.; Troiano, F.; Cappitelli, F. Bioremoval of Graffiti Using Novel Commercial Strains of Bacteria. Sci Total Environ, 2021, 756, 144075. [CrossRef]
- Sanmartín, P.; Bosch-Roig, P.; Pangallo, D.; Kraková, L.; Serrano, M. Unraveling Disparate Roles of Organisms, from Plants to Bacteria, and Viruses on Built Cultural Heritage. Appl Microbiol Biotechnol, 2023, 107, 2027–2037. [CrossRef]
- Villar-dePablo, M.; Ascaso, C.; Rodríguez-Pérez, E.; Urizal, M.; Wierzchos, J.; Pérez-Ortega, S.; de los Ríos, A. Innovative Approaches to Accurately Assess the Effectiveness of Biocide-Based Treatments to Fight Biodeterioration of Cultural Heritage Monuments. Sci. Total Environ. 2023, 897, 165318. [CrossRef]
- Ciuffreda, L.; Rodríguez-Pérez, H.; Flores, C. Nanopore Sequencing and Its Application to the Study of Microbial Communities. CSBJ 2021, 19, 1497–1511. [CrossRef]
- Kerkhof, L.J. Is Oxford Nanopore Sequencing Ready for Analyzing Complex Microbiomes? FEMS Microbiol. Ecol., 2021, 97, fiab001. [CrossRef]
- Ni, Y.; Liu, X.; Simeneh, Z.M.; Yang, M.; Li, R. Benchmarking of Nanopore R10.4 and R9.4.1 Flow Cells in Single-Cell Whole-Genome Amplification and Whole-Genome Shotgun Sequencing. Comput Struct Biotechnol J, 2023, 21, 2352–2364. [CrossRef]
- Zhang, T.; Li, H.; Ma, S.; Cao, J.; Liao, H.; Huang, Q.; Chen, W. The Newest Oxford Nanopore R10.4.1 Full-Length 16S rRNA Sequencing Enables the Accurate Resolution of Species-Level Microbial Community Profiling. Appl Environ Microbiol, 2023, 89, e0060523. [CrossRef]
- Sereika, M.; Kirkegaard, R.H.; Karst, S.M.; Michaelsen, T.Y.; Sørensen, E.A.; Wollenberg, R.D.; Albertsen, M. Oxford Nanopore R10.4 Long-Read Sequencing Enables the Generation of near-Finished Bacterial Genomes from Pure Cultures and Metagenomes without Short-Read or Reference Polishing. Nat Methods, 2022, 19, 823–826. [CrossRef]
- Zorz, J.; Li, C.; Chakraborty, A.; Gittins, D.A.; Surcon, T.; Morrison, N.; Bennett, R.; MacDonald, A.; Hubert, C.R.J. SituSeq: An Offline Protocol for Rapid and Remote Nanopore 16S rRNA Amplicon Sequence Analysis. ISME Commun, 2023, 3, 1–11. [CrossRef]
- Chandrakumar, I.; Gauthier, N.P.G.; Nelson, C.; Bonsall, M.B.; Locher, K.; Charles, M.; MacDonald, C.; Krajden, M.; Manges, A.R.; Chorlton, S.D. BugSplit Enables Genome-Resolved Metagenomics through Highly Accurate Taxonomic Binning of Metagenomic Assemblies. Commun Biol 2022, 5, 1–10. [CrossRef]
- Curry, K.D.; Wang, Q.; Nute, M.G.; Tyshaieva, A.; Reeves, E.; Soriano, S.; Wu, Q.; Graeber, E.; Finzer, P.; Mendling, W.; et al. Emu: Species-Level Microbial Community Profiling of Full-Length 16S rRNA Oxford Nanopore Sequencing Data. Nat Methods 2022, 19, 845–853. [CrossRef]
- Fan, J.; Huang, S.; Chorlton, S.D. BugSeq: A Highly Accurate Cloud Platform for Long-Read Metagenomic Analyses. BMC Bioinformatics 2021, 22, 160. [CrossRef]
- Jung, A.; Chorlton, S.D. BugSeq 16S: NanoCLUST with Improved Consensus Sequence Classification bioRxiv. 2021. [CrossRef]
- Petrone, J.R.; Rios Glusberger, P.; George, C.D.; Milletich, P.L.; Ahrens, A.P.; Roesch, L.F.W.; Triplett, E.W. RESCUE: A Validated Nanopore Pipeline to Classify Bacteria through Long-Read, 16S-ITS-23S rRNA Sequencing. Front. Microbiol. 2023, 14. [CrossRef]
- Planý, M.; Sitarčík, J.; Pavlović, J.; Budiš, J.; Koreňová, J.; Kuchta, T.; Pangallo, D. Evaluation of Bacterial Consortia Associated with Dairy Fermentation by Ribosomal RNA (Rrn) Operon Metabarcoding Strategy Using MinION Device. Food Bioscience 2023, 51, 102308. [CrossRef]
- Rodríguez-Pérez, H.; Ciuffreda, L.; Flores, C. NanoCLUST: A Species-Level Analysis of 16S rRNA Nanopore Sequencing Data. Bioinformatics 2021, 37, 1600–1601. [CrossRef]
- Rodríguez-Pérez, H.; Ciuffreda, L.; Flores, C. NanoRTax, a Real-Time Pipeline for Taxonomic and Diversity Analysis of Nanopore 16S rRNA Amplicon Sequencing Data. CSBJ 2022, 20, 5350–5354. [CrossRef]
- Maghini, D.G.; Moss, E.L.; Vance, S.E.; Bhatt, A.S. Improved High-Molecular-Weight DNA Extraction, Nanopore Sequencing and Metagenomic Assembly from the Human Gut Microbiome. Nat Protoc, 2021, 16, 458–471. [CrossRef]
Figure 1.
Example of an Oxford Nanopore® MinION™ sequencer device.

Figure 2.
Alluvial plot displaying a summary of works were Oxford Nanopore® MinION™ sequencer was applied in the cultural heritage biodeterioration field (SM: Shotgun metagenomics; PCR: Polymerase chain reaction; WGA: Whole genome amplification; WG: Whole genome).
Figure 2.
Alluvial plot displaying a summary of works were Oxford Nanopore® MinION™ sequencer was applied in the cultural heritage biodeterioration field (SM: Shotgun metagenomics; PCR: Polymerase chain reaction; WGA: Whole genome amplification; WG: Whole genome).
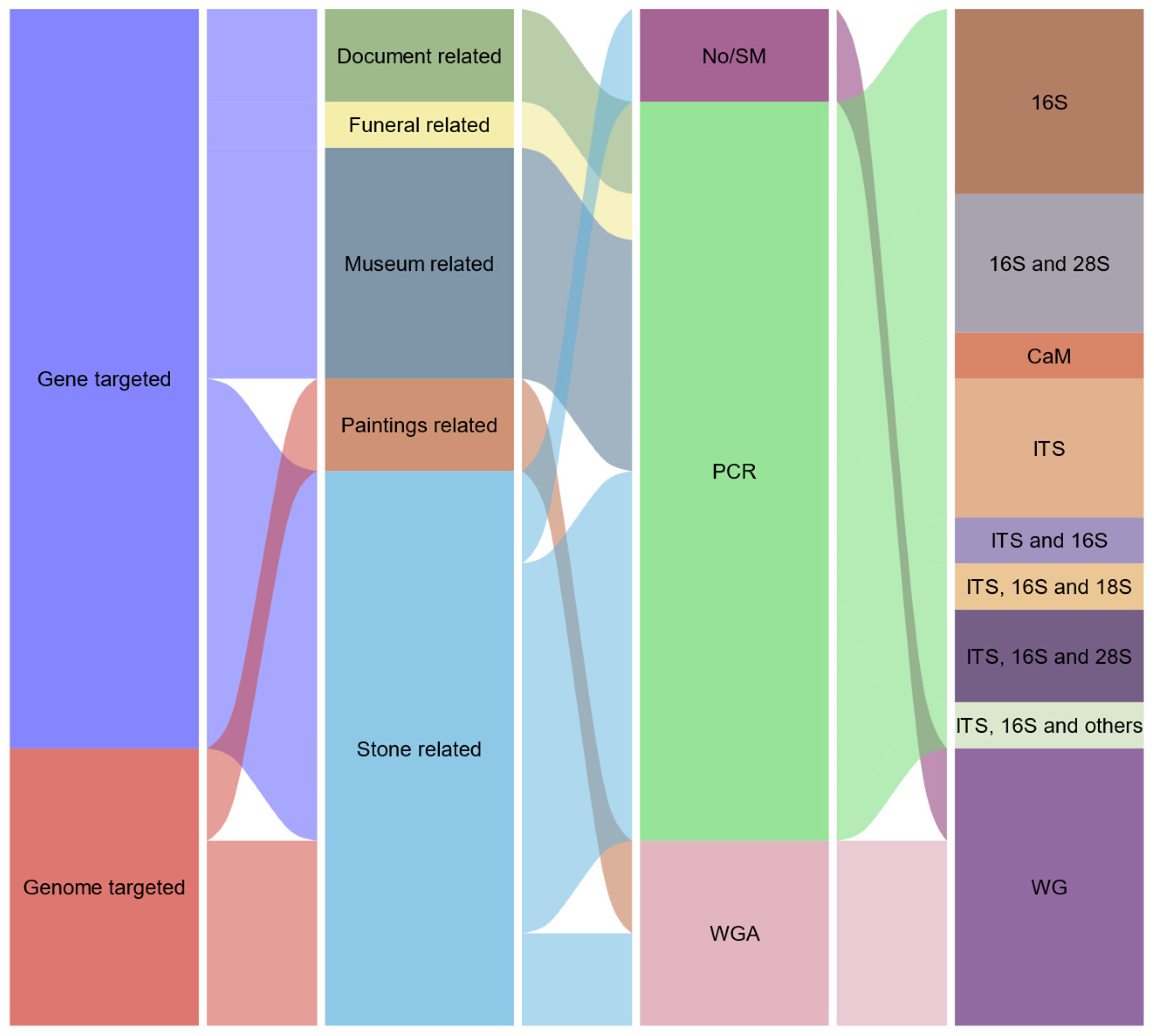
Figure 3.
VOSviewer (https://www.vosviewer.com/) co-occurrence network displaying the main keywords (appearing at least two times) from works regarding the application of the Oxford Nanopore® sequencer in the cultural heritage biodeterioration area.
Figure 3.
VOSviewer (https://www.vosviewer.com/) co-occurrence network displaying the main keywords (appearing at least two times) from works regarding the application of the Oxford Nanopore® sequencer in the cultural heritage biodeterioration area.
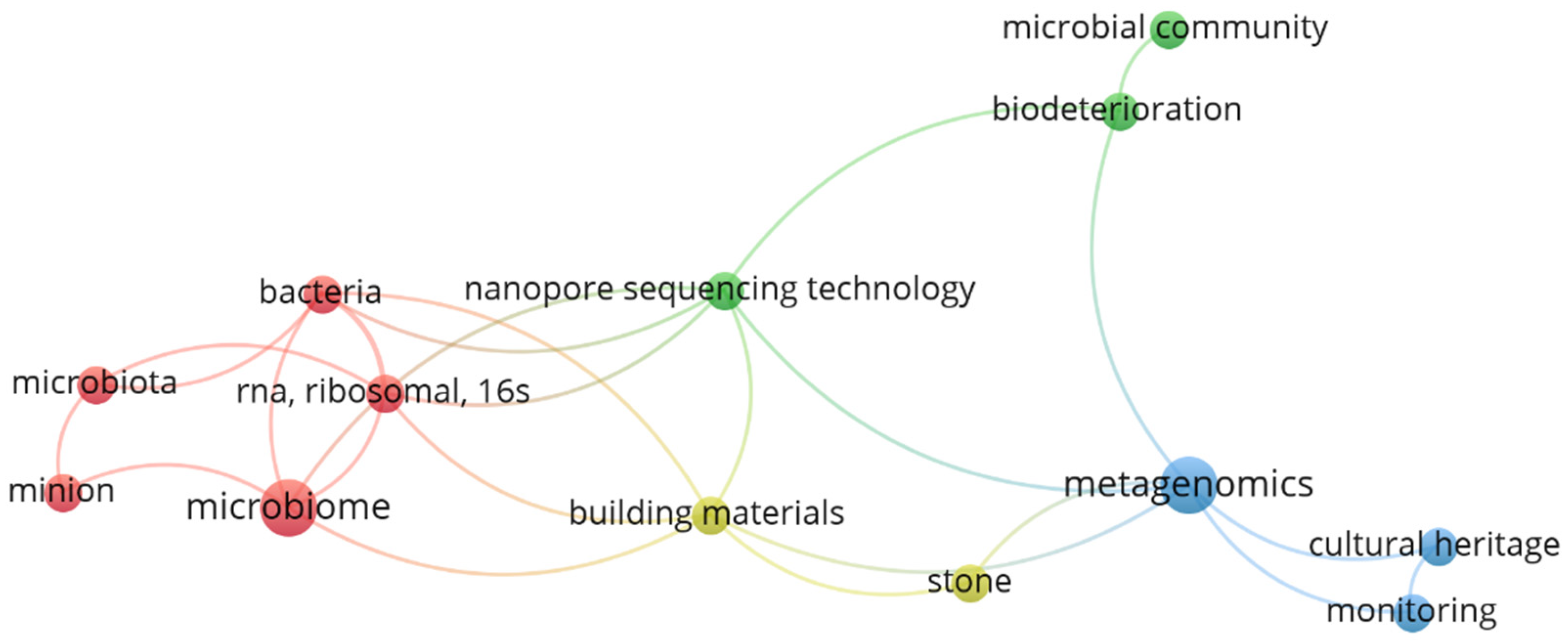
Figure 4.
VOSviewer (https://www.vosviewer.com/) co-occurrence network displaying the main words (appearing at least two times in the abstract section) from works regarding the application of the Oxford Nanopore® sequencer in the cultural heritage biodeterioration area.
Figure 4.
VOSviewer (https://www.vosviewer.com/) co-occurrence network displaying the main words (appearing at least two times in the abstract section) from works regarding the application of the Oxford Nanopore® sequencer in the cultural heritage biodeterioration area.
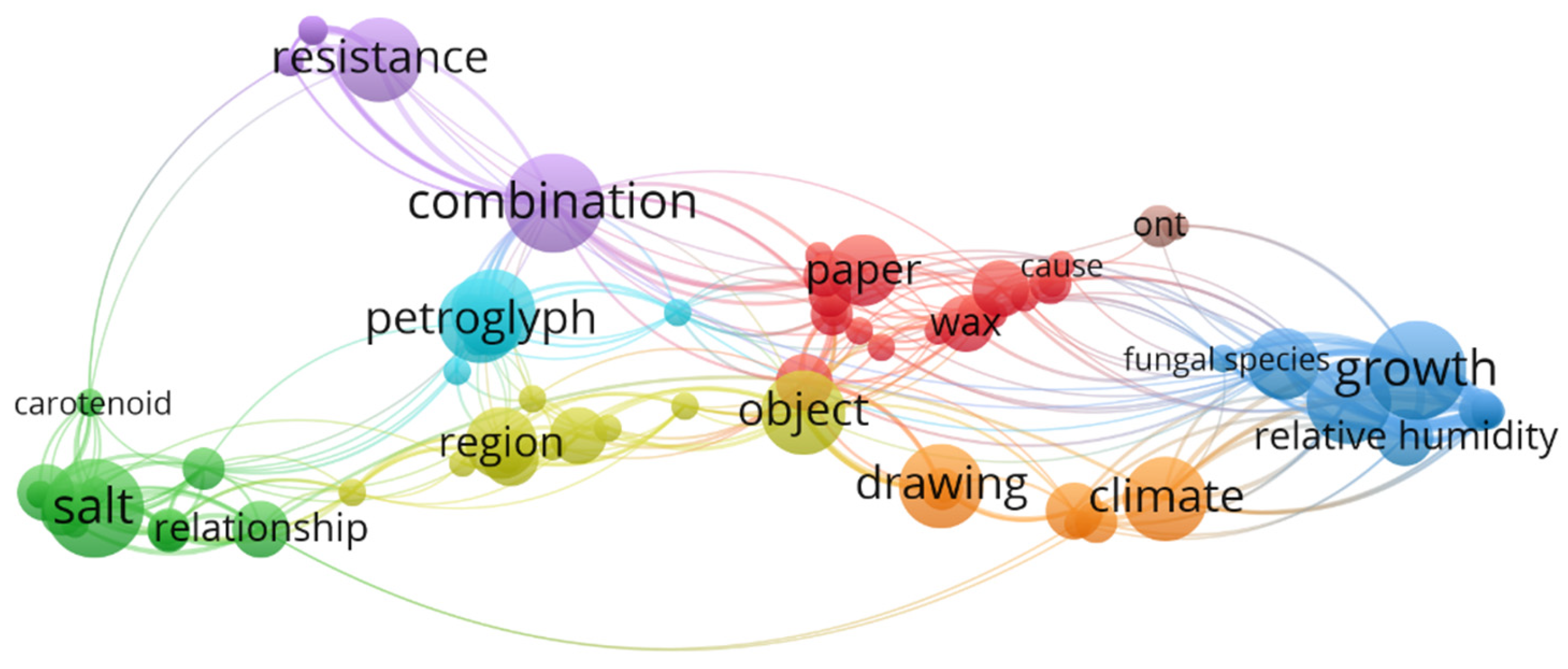
Figure 5.
Common workflows for the application of MinION™ sequencer in the CHB area include.
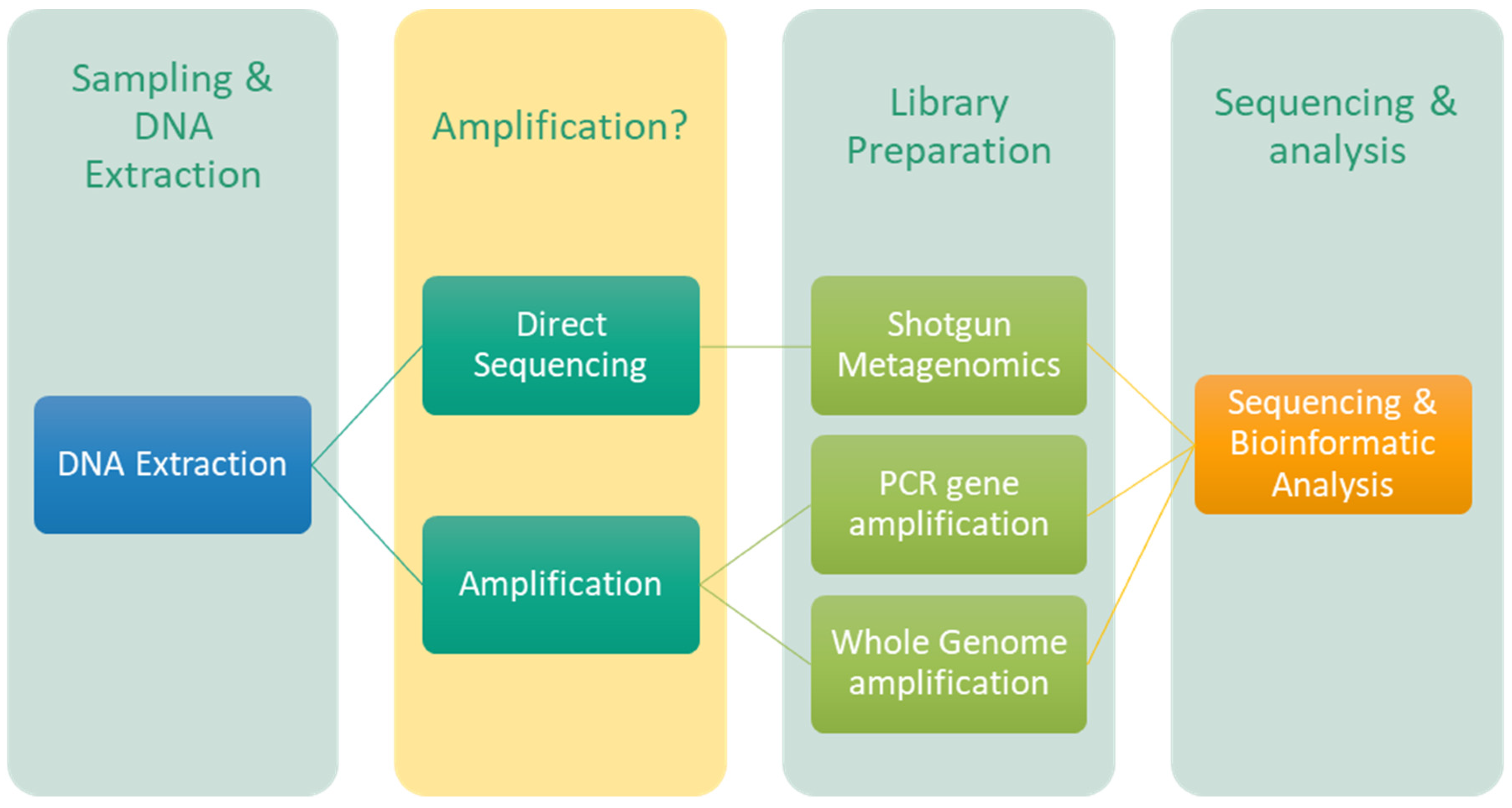
Figure 6.
Possible future directions for the Oxford Nanopore® application in biodeterioration studies.
Figure 6.
Possible future directions for the Oxford Nanopore® application in biodeterioration studies.
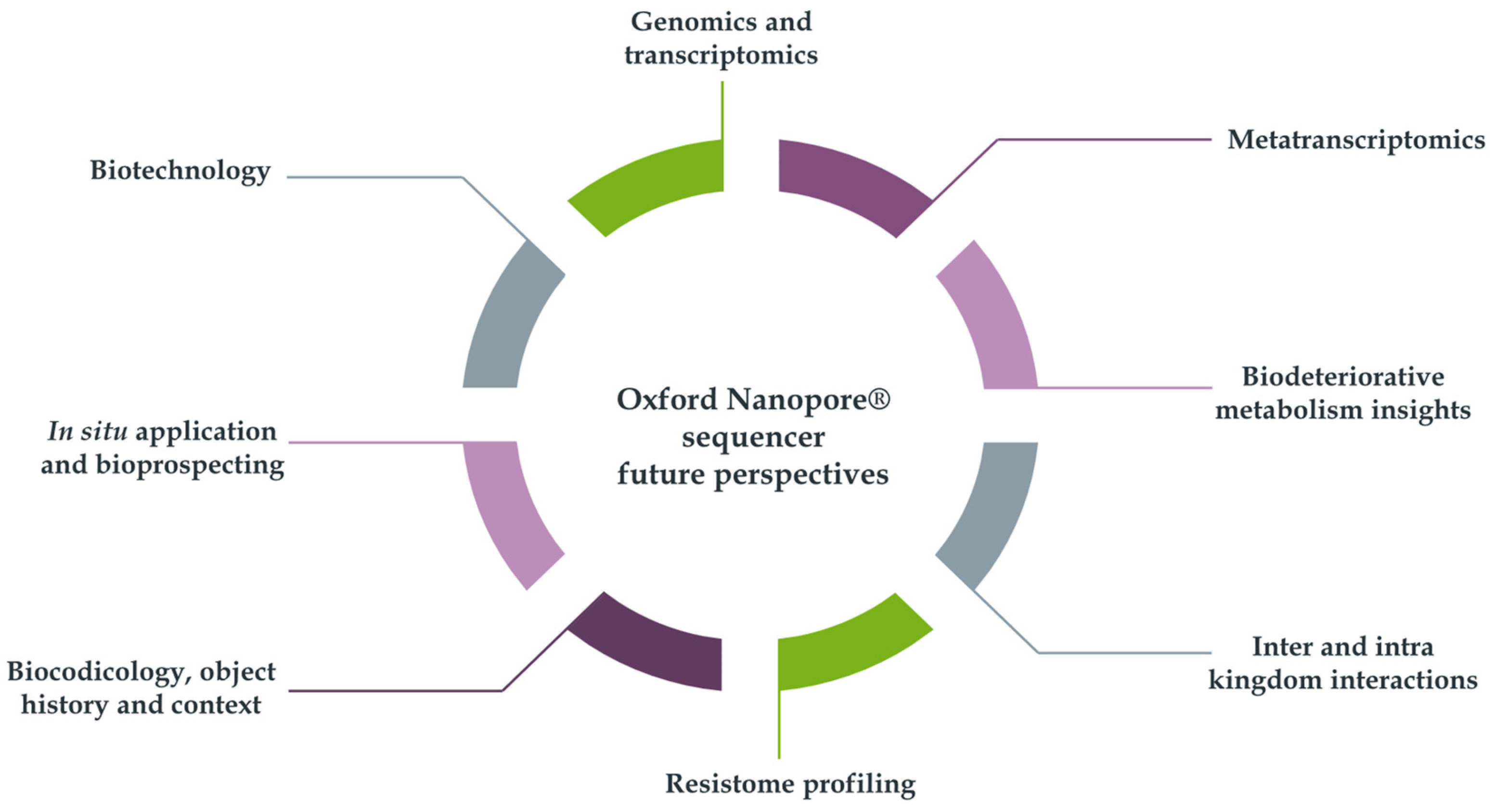

Table 1.
Cultural heritage biodeterioration studies conducted with the MinION™ sequencer.
| Study | Material sampled | DNA Extraction Method | Approach | Amplification Step | Genes | WGA Protocol Followed | Reference |
|---|---|---|---|---|---|---|---|
| Bastholm and colleagues (2024) | Air and museum surfaces | FastDNA Spin kit for soil | Gene targeted | Yes (PCR amplification) | CaM | NA | [18] |
| Beccaccioli and colleagues (2023) | Waterlogged wood | CTAB method | Gene targeted | Yes (PCR amplification) | ITS and 16S | NA | [19] |
| Brimblecombe and colleagues (2022) | Museum surfaces | FastDNA Spin kit for soil | Gene targeted | Yes (PCR amplification) | ITS | NA | [20] |
| Delegou and colleagues (2022) | Stone monument | zymoBIOMICS DNA miniprep kit | Gene targeted | Yes (PCR amplification) | 16S | NA | [21] |
| Derksen and colleagues (2024) | Museum surfaces | FastDNA Spin kit for soil | Gene targeted | Yes (PCR amplification) | ITS | NA | [22] |
| Grottoli and colleagues (2020) | Stone monument | CTAB method | Gene targeted | Yes (PCR amplification) | ITS, 16S and 18S | NA | [23] |
| Haedar and colleagues (2024) | Rock Paintings | zymoBIOMICS DNA miniprep kit | Gene targeted | Yes (PCR amplification) | 16S | NA | [24] |
| Kisová and colleagues (2020) | Funeral accessories textiles | DNeasy PowerSoil extraction kit | Gene targeted | Yes (PCR amplification) | ITS, 16S and 28S | NA | [25] |
| Li and colleagues (2023) | Stone monument | DNeasy PowerSoilPro Kit | Genome targeted | No | NA (WG) | Shotgun metagenomics paired with Illumina Sequencing | [26] |
| Nir and colleagues (2023) | Petroglyph sites | MoBio Power-Soil DNA isolation kit | Genome targeted | No | NA (WG) | Shotgun metagenomics paired with Illumina Sequencing | [27] |
| Pavlović and colleagues (2022a) | Granite chapel | DNeasy PowerSoil extraction kit | Gene targeted | Yes (PCR amplification) | ITS, 16S and others | NA | [28] |
| Pavlović and colleagues (2022b) | Beeswax drops | DNeasy PowerSoil extraction kit | Gene targeted | Yes (PCR amplification) | 16S and 28S | NA | [29] |
| Pavlović and colleagues (2023) | Documents | DNeasy PowerSoil extraction kit | Gene targeted | Yes (PCR amplification) | 16S and 28S | NA | [30] |
| Piñar and colleagues (2020a) | Oil painting | FastDNA Spin kit for soil | Genome targeted | Yes (Whole Genome (WG) amplification) | NA (WG) | Whole Genome Amplification (WGA) using multiple displacement amplification (MDA) | [31] |
| Piñar and colleagues (2020b) | Drawings | FastDNA Spin kit for soil | Genome targeted | Yes (Whole Genome (WG) amplification) | NA (WG) | Whole Genome Amplification (WGA) using multiple displacement amplification (MDA) | [32] |
| Planý and colleagues (2021) | Iron nails from whale skeleton | DNeasy PowerSoil extraction kit | Gene targeted | Yes (PCR amplification) | 16S and 28S | NA | [33] |
| Rabbachin and colleagues (2022) | Petroglyph sites | FastDNA Spin kit for soil | Genome targeted | Yes (Whole Genome (WG) amplification) | NA (WG) | Whole Genome Amplification (WGA) using multiple displacement amplification (MDA) | [34] |
| Rabbachin and colleagues (2023) | Petroglyph sites | FastDNA Spin kit for soil | Genome targeted | Yes (Whole Genome (WG) amplification) | NA (WG) | Whole Genome Amplification (WGA) using multiple displacement amplification (MDA) | [35] |
| Rabbachin and colleagues (2024) | Petroglyph sites | FastDNA Spin kit for soil | Gene targeted | Yes (PCR amplification) | ITS | NA | [36] |
| Šoltys and colleagues (2020) | Wax seal | DNeasy PowerSoil extraction kit | Gene targeted | Yes (PCR amplification) | ITS, 16S and 28S | NA | [37] |
| Tichy and colleagues (2023) | Salt-weathered buildings | FastDNA Spin kit for soil | Gene targeted | Yes (PCR amplification) | 16S | NA | [38] |
| Timoncini and colleagues (2022) | Bronze and marble statues | E.Z.N.A. Soil DNA Kit | Gene targeted | Yes (PCR amplification) | 16S | NA | [39] |
Table 2.
Bioinformatic approaches applied to the MinION™ sequencer data in the CHB area.
| Study | Bioinformatic approach | Reference |
|---|---|---|
| Bastholm and colleagues (2024) | Guppy; Porechop; Nanoplot; Nanofilt; minimap2; Racon; VSEARCH; Blastn | [18] |
| Beccaccioli and colleagues (2023) | Guppy; Porechop; BLASTN; KronaTools | [19] |
| Brimblecombe and colleagues (2022) | EPI2ME / “What is in my pot” (WIMP) | [20] |
| Delegou and colleagues (2022) | EPI2ME | [21] |
| Derksen and colleagues (2024) | Guppy; Porechop; NanoFilt; Emu with UNITE database | [22] |
| Grottoli and colleagues (2020) | “AmpLIcon SequencIng Analysis” (ALISIA) | [23] |
| Haedar and colleagues (2024) | Guppy; NanoPlot; Nanofilt; Centrifuge classifier with NCBI 16S RefSeq database; Pavian; KronaTools | [24] |
| Kisová and colleagues (2020) | Albacore; EPI2ME / “What is in my pot” (WIMP) | [25] |
| Li and colleagues (2023) | Guppy; NanoStat; Flye; Pilon; QUAST; cd-hit | [26] |
| Nir and colleagues (2023) | metaWRAP; Hybrid Spades; PROKKA | [27] |
| Pavlović and colleagues (2022a) | EPI2ME / “What is in my pot” (WIMP); | [28] |
| Pavlović and colleagues (2022b) | Albacore; EPI2ME / “What is in my pot” (WIMP) | [29] |
| Pavlović and colleagues (2023) | EPI2ME / “What is in my pot” (WIMP); MetONTIIME | [30] |
| Piñar and colleagues (2020a) | Guppy; EPI2ME / “What is in my pot” (WIMP) | [31] |
| Piñar and colleagues (2020b) | Guppy; EPI2ME / “What is in my pot” (WIMP) | [32] |
| Planý and colleagues (2021) | Albacore; EPI2ME / “What is in my pot” (WIMP) | [33] |
| Rabbachin and colleagues (2022) | Guppy; Centrifuge with custom database | [34] |
| Rabbachin and colleagues (2023) | Guppy; Centrifuge with custom database | [35] |
| Rabbachin and colleagues (2024) | Guppy; NanoFilt; Emu; UNITE | [36] |
| Šoltys and colleagues (2020) | Albacore; EPI2ME / “What is in my pot” (WIMP) | [37] |
| Tichy and colleagues (2023) | Guppy; Porechop; NanoStat; NanoPlot; Emu with NCBI 16S RefSeq | [38] |
| Timoncini and colleagues (2022) | EPI2ME / “What is in my pot” (WIMP) | [39] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



