
Submitted:
20 June 2024
Posted:
21 June 2024
You are already at the latest version
Abstract
Autism spectrum disorder (ASD) is a neurodevelopmental disorder characterized by impairments in social interaction and communication, anxiety, hyperactivity, and restricted interest for specific subjects. In addition to the genetic factors, multiple environmental factors have been related to the development of ASD. Animal models can serve as crucial tools for understanding the complexity of ASD and identifying potential therapeutic strategies. In this study, a chemical model of ASD has been developed in zebrafish by exposing embryos to valproic acid (VPA) from 4 to 48 hours post-fertilization. After the exposure, embryos were transferred to fish water and the effects on behavior and neurotransmitters profile were analyzed at both larval and adult stages. Larvae from VPA-treated embryos showed hyperactivity and decreased visual and vibrational escape responses, as well as an altered neurotransmitters profile, with increased glutamate and decreased acetylcholine and norepinephrine levels. Adults from VPA-treated embryos exhibited impaired social behavior characterized by larger shoal sizes and a decreased interest for their conspecifics. Neurotransmitter analysis revealed a significant decrease in dopamine and GABA levels in the brain. These results support the interest of this model in the study of ASD.
Keywords:
valproic acid
; animal model
; zebrafish
; autism spectrum disorder
; social behavior
; neurotransmitter
1. Introduction
Autism spectrum disorder (ASD) is a neurodevelopmental disorder characterized by impairments in social interaction and communication, anxiety, hyperactivity, and restricted interest for specific subjects [1,2,3]. It is believed that emergence of autism involves a combination of anatomical brain abnormalities, genetic anomalies, and neurochemical imbalances [2,4,5]. Epidemiological studies emphasize the significance of genetic factors, revealing that autistic disorder ranks among the most genetically influenced neuropsychiatric conditions [6]. While candidate genes implicated in autism primarily involve proteins that regulate neuronal network patterning and the balance between excitatory and inhibitory signaling, ASD is highly polygenic, with no specific gene or locus associated with a large group of patients [6,7,8]. Approximately 20% of patients exhibit genetic mutations affecting known major genes, contributing to the complexity of clinical manifestations and etiology, without a specific biological hallmark identified for this disorder [2]. In addition to the genetic factors, multiple environmental factors have been related to the development of ASD [1,3,6,7].
Animal models in ASD research can serve as crucial tools for understanding the complexity of the disorder, as they allow testing specific hypotheses of its causes and identifying potential therapeutic strategies [9,10]. However, challenges persist in creating models that faithfully represent the complexity of ASD due to its unclear etiology and heterogeneous symptomatology [9]. Replicating ASD symptoms through animal models is crucial for the identification of the neurobiological basis associated with it [2].
Molecular genetic models allow to simulate core features of human autism, including the disruption of glutamatergic synaptic transmission, loss of inhibitory GABAergic interneurons, and impairments in synaptic plasticity [11]. However, while mammalian models expressing autism-linked genetic mutations have been developed, their utility is constrained by the associated costs. Thus, simpler models such as Danio rerio, are proposed for initial mechanistic and screening studies [7]. Zebrafish, a widely used model in biomedical research, emerges as an ideal candidate for behavioral screening in ASD research, as its predominant social behavior along with other advantages make it a valuable tool for investigating the neurobiological basis of the disorder [12,13,14].
Environmental factors, such as drug and toxin exposure, viral infections, and immune dysfunctions during pre- and post-natal stages can influence ASD risks [6,7,8]. For instance, prenatal exposure to valproic acid (VPA) has been associated with an increased incidence of autism [7,15]. Because of the association observed in humans between maternal treatment with VPA and ASD, the use of prenatal VPA exposure has been used to build chemical models in different animal models, including zebrafish, to study ASD [9,16,17]. VPA dysregulates key transcription factors, signal transduction pathways, inositol metabolism, and direct modulation of epigenetic regulators, specifically histone deacetylases (HDACs), resulting in more compact DNA-histone packaging and reduced gene transcription [8,9]. Moreover, VPA exposure has been reported to lead to changes in gamma-aminobutyric acid (GABA) levels in the central nervous system [18,19,20].
Studies indicate that chemical models built using VPA exhibit construct, face and predictive validity as an ASD model. In this study, 4 hours-post fertilization (hpf) zebrafish embryos have been exposed to 48 mM VPA for 48 h, and then, transferred to clean fish water. The effect of the early-life exposure to VPA on the basal locomotor activity and the escape response evoked by visual and vibrational stimuli were determined at 8 days post-fertilization. At adult stage (3 months), social behavior (shoaling and social preference tests) was analyzed, and transcriptomic and metabolomic changes in the brain were determined.
2. Results
2.1. Systemic and Developmental Effects in Zebrafish larvae Exposed to VPA during Early Development
Embryos exposed to VPA exhibited a significant delay in hatching time compared to controls. At 48 hpf, around 40% of the control embryos, but none of those exposed to VPA, had hatched (Supplementary Figure S1A; z = -8.33, p < 0.0001). At 72 hpf, the 95% of the control and 80% of the VPA-exposed embryos had hatched (Supplementary Figure S1B; z = -5.75, p < 0.0001).
When the toxicity of the VPA-treatment was determined in 7 dpf larvae, the VPA-exposed group exhibited a similar cumulative mortality to the control group (Supplementary Figure S1C). However, VPA exposure led to a mild but significant increase in phenotypic abnormalities, such as pericardial and yolk sac edemas (Supplementary Figure S1D; 0.83% and 4.65%, median values for control and VPA, respectively; z = -3.52, p < 0.0001).
VPA exposure during early development (4-48 hpf) also led to a significant increase in heart rate when larvae reached 8 dpf, compared to the corresponding controls (Supplementary Figure S2; t (31) = 3.504, p = 0.0014).
2.2. Neurobehavioral Effects in Zebrafish larvae Exposed to VPA during Early Development
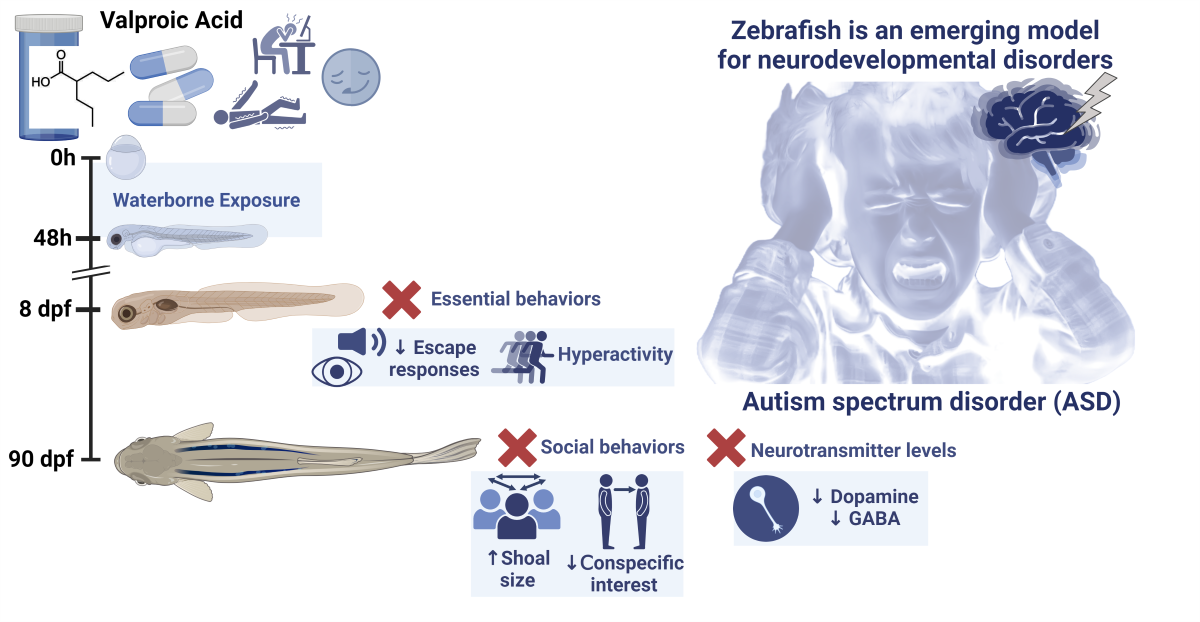
For neurobehavioral assessment, 8 dpf zebrafish larvae without any evidence of systemic toxicity were selected. Moreover, no differences in the standard length were found between the control and the VPA treated groups [t (18) = 1.852, p = 0.08]. As shown in Figure 1, the exposure of embryos to VPA from 4 to 48 hpf led to significant changes in all three analyzed behaviors. While BLA increased in the treated larvae (z = -2.92, p = 0.0034), the escape response evoked by a visual and a vibrational stimulus decreased (z = -2.35, p = .0185 for VMR; z = -3.86, p < .0001 for VSR).
The following step was to determine if the observed changes in the different behaviors might be related with changes in the neurotransmitter profile of larvae exposed to VPA during the early development. As shown in Figure 2 and Supplementary Table S1, larval heads exposed to VPA during early development presented a significant reduction on the levels of acetylcholine (U(Ncontrol = 7, NVPA = 7) = 6.00, p = 0.017), and norepinephrine(U(Ncontrol = 7, NVPA = 7) = 4.00, p = 0.007), whereas the levels of glutamate increased in this group (U(Ncontrol = 7, NVPA = 7) = 48.00, p = 0.001). VPA-exposed larvae also showed a significant decrease in epinephrine levels (U(Ncontrol = 7, NVPA = 7) = 2.00, p = 0.002; Supplementary ST1).
2.3. Neurobehavioral Effects in Adult Zebrafish Exposed to VPA during Early Development
Adult zebrafish selected for behavioral analysis had a similar body length (19.97 ± 0.28 mm for the control and 18.95 ± 0.34 for the VPA-treated group) and no differences in the total distance moved in 6 min were found between VPA-exposed and control fish (Supplementary Figure S2).
The effects of early life exposure to VPA on adult social behavior were explored using two experimental paradigms, the shoaling test and the social preference test. The shoaling test (Figure 3A) showed significantly higher average interfish distance (U(Ncontrol = 18, NVPA = 16) = 261.00, p = 1.28 × 10−5) and farthest interfish distance (t (32) = -8.667, p = 6.65 × 10−10), a behavioral phenotype consistent with social isolation, in the adults exposed to VPA during early development. The social preference test (Figure 3B) showed a significant decrease in both the time spent and distance moved by treated fish in the zone closest to conspecifics (time: t (31) = 2.350, p = 0.025; distance: t (31) = 2.348, p = 0.025), and a concomitant increase in the time spent and the distance moved by these fish in the empty virtual zone (time: t (31) = -2.112, p = 0.043; distance: t (31) = -2.125, p = 0.042). These results are consistent with the social isolation phenotype suggested by the shoaling test results.
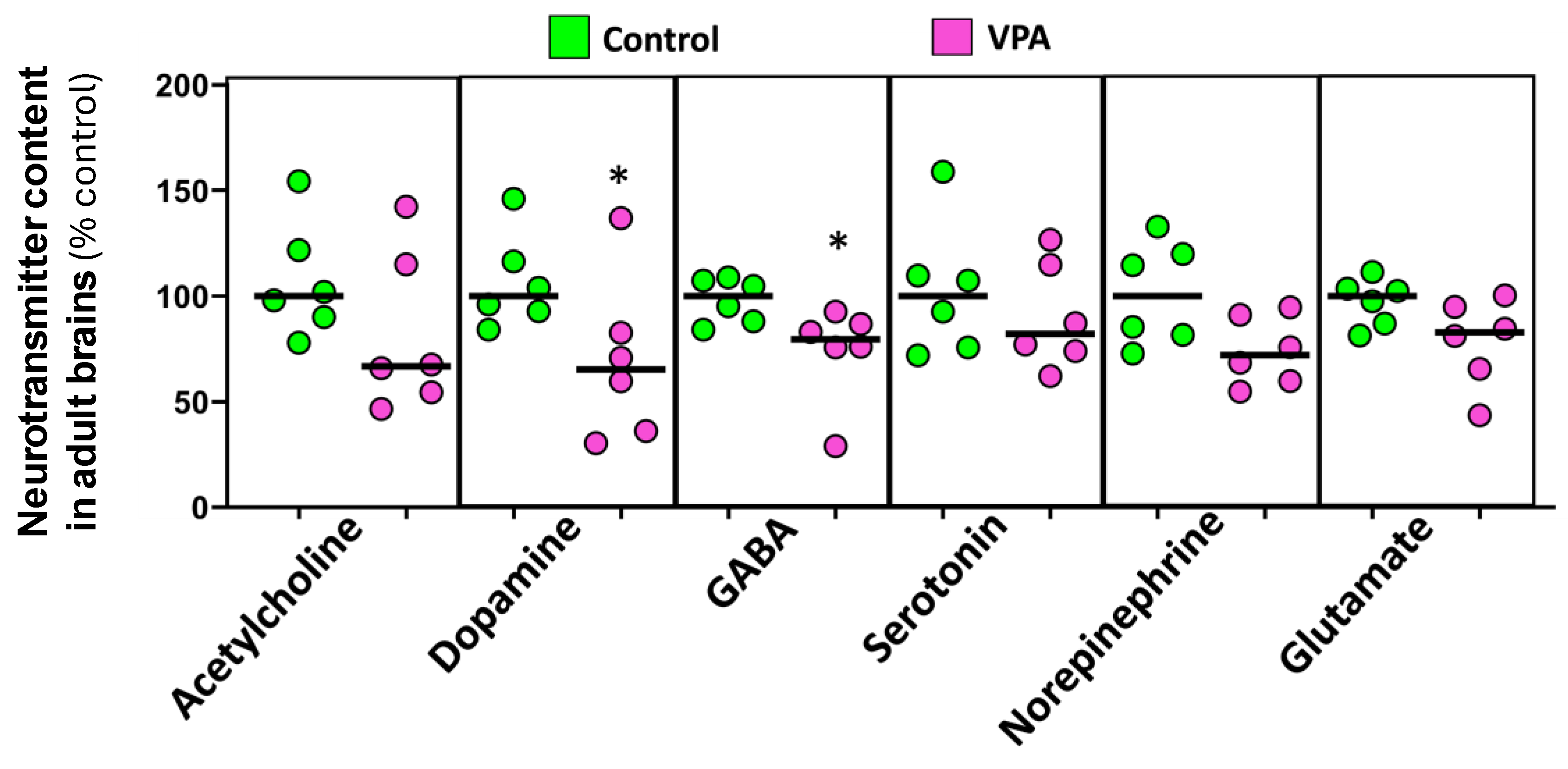
As shown in Figure 4, when the levels of different neurotransmitters in the brain were analyzed, a significant decrease in the levels of GABA (U(Ncontrol = 6, NVPA = 6) = 3.00, p 0.015) and dopamine (U(Ncontrol = 6, NVPA = 6) = 5.00, p 0.041) was found in VPA-exposed fish compared to controls. A trend to a reduction in glutamate levels was also found in the brain of the exposed fish (U(Ncontrol = 6, NVPA = 6) = 6.00, p 0.065).
3. Discussion
VPA is a medication widely used to treat epilepsy, bipolar disorder and migraines [21]. Despite its therapeutic effect, exposure to VPA during the first three months of pregnancy can result in a rare condition known as fetal valproate syndrome (FVS) [21]. Children with FVS have also an increased risk of ASD [22,23]. In fact, VPA is commonly used to build chemical models of ASD in different animal species, including zebrafish [12,24]. In this study we present the effects on social behavior and neurotransmitter profile found in zebrafish larvae and adult exposed to VPA during the early development. First of all, we have found that the exposure of 4 hpf zebrafish embryos to 48 μM VPA for 48 h did not result in mortality when this endpoint was evaluated at 7 dpf. Consistently, no mortality in zebrafish embryos has been reported by other authors using the same [12] or similar exposure conditions [25] to VPA. However, the effect of VPA exposure on zebrafish embryos mortality is, anyway, controversial. Whereas in one report embryos exposed to 40 μM VPA from 8 to 120 hpf showed a 96% mortality at 7 dpf [26], in another report the exposure of 8 hpf zebrafish embryos for 100 h to 1500 μM VPA did not result in mortality [16].
In our study we have found that exposure of zebrafish embryos during the first 48 h of development to VPA leaded to a mild (4.65%) but significant increase in phenotypic abnormalities [21]. In contrast, no phenotypic effects have been reported in exposed embryos by other authors using similar exposure conditions [12,25]. While morphological alteration were also not reported after exposure of embryos to 75 μM VPA from 4 to 120 hpf [17] or to 50 μM VPA from 8 to 108 hpf [16], exposure to 30 μM VPA from 8 to 120 hpf showed 30.5% malformations.
The observed variability in mortality and malformation rate among these studies based on the waterborne exposure to VPA could be related to the differences in the final pH of the experimental solutions. VPA is a weak acid with a pKa of 4.86, and depending on the pH of the experimental solution, the chemical will be ionized (low bioavailability) or non-ionized (high bioavailability). The percentage of non-ionized VPA molecules is higher at low pH (39% of VPA is in its non-ionized form at pH 5.0), than at the physiological pH 6.3 (3.1% non-ionized) [27]. Therefore, VPA uptake should be significantly higher in studies using experimental solutions with low pH than in those using experimental solutions with a final pH above 7. In fact, when we performed the first experiments with VPA, using our standard fish water to prepare the experimental solution of VPA (pH 6.5), we found a huge mortality of exposed embryos. At pH 6.5, about 1.96% of the VPA is expected to be in the form of valproate (non-ionized form) and, therefore, bioavailable. The pH of the fish water was then raised to 7.5 in both fish water and experimental solutions by adding bicarbonate and, at this pH, the non-ionized fraction should be only 0.23%. When this water was used to prepare the experimental solutions, we found no mortality and a very limited percentage of malformations. In addition to the potential differences in VPA bioavailability, the sensitivity of the zebrafish strain and batch could have also played a role, as some malformations were observed on the controls too.
The hyperactivity found in VPA-exposed larvae in this study aligns with the results obtained by most of the previous studies using this chemical [12,16,26,28], and may be related to the dysregulation of dopaminergic and glutamatergic neurotransmission in certain brain regions, as suggested for ADHD (attention-deficit hyperactivity disorder), a disorder related to and co-occurring with ASD [29,30,31,32]. However, no effects on BLA have been reported in shank3b-/- zebrafish larvae, a result suggesting that hyperactivity is a VPA effect unrelated with ASD [14].
In this study we have also found a decrease in the light-off VMR in larvae exposed to VPA during early development, a result consistent with the reported by Bailey et al. [33] in 6 dpf larvae exposed to 30-50 μM VPA from 4 to 120 hpf, but not with the significant increase reported by Joseph et al [26] in 5 and 7 dpf larvae exposed from 8 to 108 hpf to 5-10 μM VPA and by Baronio et al. [34] in 5 dpf larvae exposed to 25 μM VPA from 10 to 24 hpf. A similar decrease in light-off VMR was also reported in shank3b-/- zebrafish larvae, suggesting that this effect of VPA could be related with ASD. The decrease in the VMR could be related to an excitatory-inhibitory imbalance in the visual cortex suggested to happen in ASD patients [34]. We also observed a significant decrease in the vibrational-evoked startle response in those larvae exposed to VPA during the early embryonic development. Interestingly, Gupta et al. [25], in a seminal article on neuroanatomical mapping of the zebrafish brain, found that 6 dpf zebrafish larvae exposed to 50 μM VPA during early development presented a loss of glutamatergic signal in part of the statoacoustic ganglion leading to a decreased acoustic startle responsiveness. Glutamate dysregulation is thought to be implicated in the changes of the startle response through NMDA and mGlu5 receptors [34,35,36,37].
It is difficult to explain the behavioral changes found in the VPA-exposed larvae by the observed changes in the neurotransmitter profiles found in their heads. For instance, hyperactivity has been linked to the increase in the dopamine, acetylcholine [38,39] and/or norepinephrine [40,41] levels in the zebrafish larvae. However, levels of acetylcholine and norepinephrine in the heads of the exposed larvae decreased, and no changes were found in dopamine levels. The observed hyperactivity is also not explained by increased glutamate levels in the head of VPA-exposed larvae, as glutamate has been reported to reduce the motor activity of zebrafish larvae [42]. These results emphasize the difficult to correlate behavioral changes with changes in the neurotransmitter profile observed in larval heads, since (1) the analysis includes the whole set of neurotransmitters, not just those released into the synaptic space, and (2) the analysis is performed on the entire head, not on discrete nuclei. This is a clear limitation of the zebrafish model, in which, due to its small size, it is not possible to use microdialysis [44] to selectively determine neurotransmitters in the synaptic clefts of specific CNS nuclei, as for example is done in rats.
The decrease in the social behavior found in this study in adult zebrafish (90 dpf) exposed to VPA during early embryonic development is consistent with previous studies reporting increased size of the shoals [33] and decreased preference for being near conspecifics [12] in VPA-treated zebrafish. Similar disorders of social behavior have been described in genetic models of ASD constructed in zebrafish [13,14]. Interestingly, the decrease found in the dopamine levels in the brain of the VPA-treated fish has also been reported in different mammalian models [45,46]. Since the dopaminergic system is involved in social behavior, the observed decrease in dopamine levels could be behind the impaired social behavior found in ASD [45,47,48]. Moreover, the significant decrease in GABA levels found in the brain of adult fish exposed to VPA during early development is consistent with the decrease in this neurotransmitter reported in ASD patients [49,50].
In summary, the neurobehavioral short- and long-term effects of the developmental exposure to valproic acid in zebrafish have been analyzed, with special emphasis on determining the relationship between behavioral and neurochemical changes. Our results suggest the importance of the pH of the experimental solution on the bioavailability of the VPA and therefore, in the severity of the observed effects. Moreover, our results show that the developed ASD model exhibited an impaired social behavior, characterized by an increase in the average inter-fish distance, in the shoaling test, and a decrease in the time spent and distance moved in the closest zone to conspecifics, in the social preference test. The impairment of social behavior combined with the significant decrease found in brain levels of dopamine and GABA in adult zebrafish treated with VPA during early development strongly suggests that this model is of great interest for both the study of FVS and ASD.
4. Materials and Methods
4.1. Zebrafish Housing and Husbandry
Adult zebrafish were supplied by Pisciber BSF (Terrassa, Barcelona) and maintained at the Research and Development Center of the Spanish Research Council (CID-CSIC) facilities in fish water [FW: reverse-osmosis purified water containing 90 μg/mL of Instant Ocean® (Aquarium Systems, Sarrebourg, France), 0.58 mM CaSO4·2H2O, pH 6.5] at 28 ± 1 °C. A 12:12 light:dark photoperiod was used. Fish were fed twice a day with flake food (TetraMin, Tetra, Germany).
For breeding, a series of polycarbonate crossing tanks were prepared with a total of five adult zebrafish in each one, the ratio of female:male being 3:2. After a treatment with 0.1% methylene blue, embryos were maintained in crystallizing dishes with fish water inside the incubator (at 28°C, 12:12 light:dark photoperiod). At 4 hpf, the embryos at sphere and dome stages [51] were selected and transferred to 6-well cell culture plates (20 embryos/well) with the exposure medium.
All procedures were approved by the Institutional Animal Care and Use Committees at the CID-CSIC and conducted in accordance with the institutional guidelines under a license from the local government (agreement number 11336).
4.2. Experimental Procedure
Sodium valproate (VPA; CAS: 1069-66-5) was purchased from Sigma-Aldrich (St Louis, Missouri, U.S.). For treatment, an exposure protocol similar to that described by Zimmermann et al. (2015) was used [12]. Basically, the day of the experiment a fresh 48 µM VPA solution was prepared in FW and then, pH was adjusted to 7.5 with a 1 M NaHCO3 solution. At 4 hpf, embryos at sphere-dome stages were exposed to the VPA solution (treated group), or to FW (also adjusted to pH 7.5, control group), for 48h without medium renewal. Zimmermann et al. (2015) reported that this concentration of VPA is stable in fish water for at least 48 h [12]. At 52 hpf, embryos were removed from the experimental solutions, washed and transferred to pH 7.5 FW (28°C and 12L:12D photoperiod) until 8 dpf, when the behavioral effects of the early exposure to VPA were initially tested. Afterwards, larvae were placed in a 2L tank with fish water and fed twice a day. After three months, when zebrafish reached the adult state, social behavior was determined. Finally, fish were euthanized by inducing hypothermic shock in ice-chilled water (2–4°C), and the brains were immediately dissected and individually stored at −80°C for neurotransmitter and transcriptional analysis.
4.3. Phenotypic Analysis
Live embryos and larvae were examined with a Nikon SMZ 1500 stereomicroscope every 24 h to observe the phenotype, recording the lethality, malformations and hatching time in both control and VPA-exposed groups. Standard length of the animals from both experimental groups was determined at 8, 30, 60 and 90 dpf from pictures taken with a GigE camera mounted on the stereomicroscope using the GIMP software (version 2.10.32).
4.4. Neurobehavioral Assay in Larvae
Behavioral assays including basal locomotor activity (BLA), light-off visual motor response (VMR), and vibrational startle response (VSR) assay, were conducted and analyzed in a DanioVision platform driven by EthoVision XT 13 software (Noldus, Wageningen, The Netherlands). The behavioral assays in larvae were conducted as previously described [52,53].
At day 7, those larvae with no signs of systemic toxicity were placed in 48-well microplates, with 1 larva per well in 1 ml FW. For the behavioral analysis, larvae were first acclimated to the new environment inside the observation chamber for 10 minutes in darkness, after which the plate received one vibrational stimulus by means of a solenoid (tapping stimulus). The distance (cm) moved in response to the tap of the routine corresponds to the vibrational startle response (VSR). After delivering the tapping, the larvae were maintained in the dark without any stimulus for 15 minutes. The last 10 minutes in the dark were used to calculate their basal locomotor activity (BLA), corresponding to the total distance moved during that time. The routine continues by switching a light-on for 10 minutes and then turning it off for 15 more minutes. The light-off visual motor response (VMR) is calculated as the distance moved in the first two minutes of darkness minus the distance moved in the last two minutes of light. After the behavioral battery, larvae were euthanized by inducing hypothermic shock in ice-chilled water (2–4°C) and then the heads were cut. Eight pools of fifteen heads were prepared for each experimental condition and stored at -80 °C.
4.5. Neurobehavioral Assays in Adults
All tests were conducted in an isolated behavioral room at 27–28 °C. The assessing effects on social behavior, shoaling and social preference test (SPT) were performed following the protocol described by Bedrossiantz et al. [53]. Ethovision XT 13.0 (Noldus, Wageningen, Netherlands) was used for the video-tracking analysis. After these tests, the fish were euthanized by inducing hypothermic shock in ice-chilled water (2–4°C) and the brain of each individual was dissected and transferred into an Eppendorf tube. Eight brains of each concentration were used for the analysis. During all the process, the samples were maintained on ice. When all the samples were prepared, they were kept in a freezer at -80°C until the day of the extraction.
4.6. Neurochemical Analysis by UHPLC-MS/MS
For the neurochemical extraction of larval heads and adult brains and UHPLC-MS/MS, a protocol similar to that described by Ricarte et al. [52] was used. Basically, samples were homogenized by means of a bead mill (TissueLyser LT, Quiagen, Hilden, Germany) and centrifuged. 0.22 μm nylon filters were used to filter the supernatant into chromatographic vials that were kept at -20 ºC until the analysis. To extract and conduct the analysis of neurotransmitters, acetonitrile (ACN) HPLC-MS grade was supplied from VWR chemicals Prolabo (Leuven, Belgium), formic acid (FA) from Fisher Scientific (Loughborough, UK), and ammonium formate from Sigma-Aldrich (St. Louis, MO, USA). Ultra-pure water was obtained through Millipore Milli-Q purification system (Millipore, Bedford, MA, USA).
The content in neurochemicals of the extract were determined by UHPLC–MS/MS, using conditions described elsewhere [55,56]. BEH Amide column was used for separation and elution, and the detection was performed in MRM mode with ESI+, ensuring specificity during detection and quantification. For the calibration curve, pure reference standards were used: serotonin hydrochloride (5-HT), dopamine hydrochloride (DA), γ-aminobutyric acid (GABA), epinephrine (Epi), and acetylcholine (ACh), were supplied by Sigma-Aldrich (St. Louis, MO, USA). Glutamic acid (Glu) was supplied by BLD Pharmatech (Shanghai, China), and norepinephrine (NE) was obtained from Tocris Bioscience (Ellisville, USA). A mixture of isotopically labelled standards was used as internal standard. 5-HIAA-d5, 5-HTP-d4, 5HT-d4, NE-d6, DA-1,1,2,2-d4 and 3-MT-d4 were purchased from Toronto Research Chemicals (TRC, Toronto, Canada).
4.7. Statistical Analysis
Data were analyzed with IBM SPSS v29 (Statistical Package 2010, Chicago, IL) and GraphPad Prism 9 for Windows (GraphPad software Inc, La Jolla, CA)and plotted with GraphPad Prism 9 for Windows (GraphPad software Inc, La Jolla, CA).
In order to determine if the samples followed a normal distribution, Shapiro-Wilk test was used. For normally distributed groups, Unpaired t-test was used for determining statistical significance and one-way ANOVA followed by Dunnett’s as multiple comparison test. When parametric assumptions could not be made, statistical significance was determined by Mann-Whitney U test and the Kruskal-Wallis test followed by Dunn-Bonferroni’s test to see if there were any differences between more than two groups. Significance was set at P < 0.05.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Supplementary Figures: Figure S1. Systemic and developmental effects in zebrafish larvae; Figure S2. Total distance moved in 6 minutes (cm); Supplementary Table 1: Statistical results from Mann-Whitney U test in neurotransmitter levels of larval heads.
Author Contributions
Conceptualization: D.R., N.G.R.; Methodology: M.B., J.B.; Formal Analysis: D.R., M.R. M.B.; Investigation: M.R., N.T., E.P.; Resources, D.R., C.G.C.; Writing – Original Draft Preparation: D.R., M.R., N.T., M.B., J.B., E.P., C.G.C., N.G.R; Writing – Review & Editing: D.R., M.R., N.T., M.B., J.B., E.P., C.G.C., N.G.R; Visualization, D.R., M.J.B., J.B.; Supervision, D.R., C.G.C.; Project Administration, D.R., C.G.C; Funding Acquisition, D.R., C.G.C. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by “Agencia Estatal de Investigación” from the Spanish Ministry of Science and Innovation (projects PID2020-113371RB-C21 and TED2021-130845B-C3), and IDAEA-CSIC, Severo Ochoa Centre of Excellence (CEX2018-000794-S). Juliette Bedrossiantz was supported by a PhD grant (PRE2018-083513) co-financed by the Spanish Government and the European Social Fund (ESF).
Institutional Review Board Statement
All procedures were approved by the Institutional Animal Care and Use Committees at the CID-CSIC and conducted in accordance with the institutional guidelines under a license from the local government (agreement number 11336).
Informed Consent Statement
Not applicable.
Data Availability Statement
Data supporting the findings of this study are available within the manuscript, and its Supplementary Material file, or will be available from the corresponding author upon request.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Bölte, S.; Girdler, S.; Marschik, P.B. The contribution of environmental exposure to the etiology of autism spectrum disorder. Cell. Mol. Life Sci. 2019, 76, 1275–1297. [Google Scholar] [CrossRef]
- Fernández, M.; Mollinedo-Gajate, I.; Peñagarikano, O. Neural Circuits for Social Cognition: Implications for Autism. Neuroscience 2018, 370, 148–162. [Google Scholar] [CrossRef] [PubMed]
- Polšek, D.; Jagatic, T.; Cepanec, M.; Hof, P.R.; Šimić, G. RECENT DEVELOPMENTS IN NEUROPATHOLOGY OF AUTISM SPECTRUM DISORDERS. Transl. Neurosci. 2011, 2, 256–264. [Google Scholar] [CrossRef] [PubMed]
- Hettinger, J.A.; Liu, X.; Schwartz, C.E.; Michaelis, R.C.; Holden, J.J.A. A DRD1 haplotype is associated with risk for autism spectrum disorders in male-only affected sib-pair families. Am. J. Med. Genet. B. Neuropsychiatr. Genet. 2008, 147B, 628–636. [Google Scholar] [CrossRef]
- Marotta, R.; Risoleo, M.C.; Messina, G.; Parisi, L.; Carotenuto, M.; Vetri, L.; Roccella, M. The Neurochemistry of Autism. Brain Sci. 2020, 10. [Google Scholar] [CrossRef]
- Andres, C. Molecular genetics and animal models in autistic disorder. Brain Res. Bull. 2002, 57, 109–119. [Google Scholar] [CrossRef]
- Keil-Stietz, K.; Lein, P.J. Gene×environment interactions in autism spectrum disorders. Curr. Top. Dev. Biol. 2023, 152, 221–284. [Google Scholar] [CrossRef] [PubMed]
- Meshalkina, D.A.; N. Kizlyk, M.; V. Kysil, E.; Collier, A.D.; Echevarria, D.J.; Abreu, M.S.; Barcellos, L.J.G.; Song, C.; Warnick, J.E.; Kyzar, E.J.; et al. Zebrafish models of autism spectrum disorder. Exp. Neurol. 2018, 299, 207–216. [Google Scholar] [CrossRef]
- Ranger, P.; Ellenbroek, B.A. Perinatal Influences of Valproate on Brain and Behaviour: An Animal Model for Autism. Curr. Top. Behav. Neurosci. 2016, 29, 363–386. [Google Scholar] [CrossRef]
- Roullet, F.I.; Crawley, J.N. Mouse models of autism: testing hypotheses about molecular mechanisms. Curr. Top. Behav. Neurosci. 2011, 7, 187–212. [Google Scholar] [CrossRef]
- Topál, J.; Román, V.; Turcsán, B. The dog (Canis familiaris) as a translational model of autism: It is high time we move from promise to reality. Wiley Interdiscip. Rev. Cogn. Sci. 2019, 10. [Google Scholar] [CrossRef]
- Zimmermann, F.F.; Gaspary, K.V.; Leite, C.E.; De Paula Cognato, G.; Bonan, C.D. Embryological exposure to valproic acid induces social interaction deficits in zebrafish (Danio rerio): A developmental behavior analysis. Neurotoxicol. Teratol. 2015, 52, 36–41. [Google Scholar] [CrossRef] [PubMed]
- Kim, O.H.; Cho, H.J.; Han, E.; Hong, T.I.; Ariyasiri, K.; Choi, J.H.; Hwang, K.S.; Jeong, Y.M.; Yang, S.Y.; Yu, K.; et al. Zebrafish knockout of Down syndrome gene, DYRK1A, shows social impairments relevant to autism. Mol. Autism 2017, 8, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.X.; Li, C.Y.; Hu, C.C.; Wang, Y.; Lin, J.; Jiang, Y.H.; Li, Q.; Xu, X. CRISPR/Cas9-induced shank3b mutant zebrafish display autism-like behaviors. Mol. Autism 2018, 9, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Dufour-Rainfray, D.; Vourc’h, P.; Tourlet, S.; Guilloteau, D.; Chalon, S.; Andres, C.R. Fetal exposure to teratogens: evidence of genes involved in autism. Neurosci. Biobehav. Rev. 2011, 35, 1254–1265. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Lei, L.; Tian, L.; Hou, F.; Roper, C.; Ge, X.; Zhao, Y.; Chen, Y.; Dong, Q.; Tanguay, R.L.; et al. Developmental and behavioral alterations in zebrafish embryonically exposed to valproic acid (VPA): An aquatic model for autism. Neurotoxicol. Teratol. 2018, 66, 8–16. [Google Scholar] [CrossRef] [PubMed]
- Dwivedi, S.; Medishetti, R.; Rani, R.; Sevilimedu, A.; Kulkarni, P.; Yogeeswari, P. Larval zebrafish model for studying the effects of valproic acid on neurodevelopment: An approach towards modeling autism. J. Pharmacol. Toxicol. Methods 2019, 95, 56–65. [Google Scholar] [CrossRef] [PubMed]
- Chomiak, T.; Turner, N.; Hu, B. What We Have Learned about Autism Spectrum Disorder from Valproic Acid. Patholog. Res. Int. 2013, 2013. [Google Scholar] [CrossRef]
- Ghodke-Puranik, Y.; Thorn, C.F.; Lamba, J.K.; Leeder, J.S.; Song, W.; Birnbaum, A.K.; Altman, R.B.; Klein, T.E. Valproic acid pathway: pharmacokinetics and pharmacodynamics. Pharmacogenet. Genomics 2013, 23, 236–241. [Google Scholar] [CrossRef]
- Terbach, N.; Williams, R.S.B. Structure-function studies for the panacea, valproic acid. Biochem. Soc. Trans. 2009, 37, 1126–1132. [Google Scholar] [CrossRef]
- Bialer, M.; Yagen, B. Valproic acid: Second generation. Neurotherapeutics 2007, 4, 130. [Google Scholar] [CrossRef] [PubMed]
- Williams, G.; King, J.; Cunningham, M.; Stephan, M.; Kerr, B.; Hersh, J.H. Fetal valproate syndrome and autism: additional evidence of an association. Dev. Med. Child Neurol. 2001, 43, 202–206. [Google Scholar] [CrossRef] [PubMed]
- Tayanloo-Beik, A.; Hamidpour, S.K.; Abedi, M.; Shojaei, H.; Tavirani, M.R.; Namazi, N.; Larijani, B.; Arjmand, B. Zebrafish Modeling of Autism Spectrum Disorders, Current Status and Future Prospective. Front. psychiatry 2022, 13. [Google Scholar] [CrossRef] [PubMed]
- Mabunga, D.F.N.; Gonzales, E.L.T.; Kim, J.; Kim, K.C.; Shin, C.Y. Exploring the Validity of Valproic Acid Animal Model of Autism. Exp. Neurobiol. 2015, 24, 285–300. [Google Scholar] [CrossRef] [PubMed]
- Gupta, T.; Marquart, G.D.; Horstick, E.J.; Tabor, K.M.; Pajevic, S.; Burgess, H.A. Morphometric analysis and neuroanatomical mapping of the zebrafish brain. Methods 2018, 150, 49–62. [Google Scholar] [CrossRef] [PubMed]
- Joseph, T.P.; Zhou, F.; Sai, L.Y.; Chen, H.; Lin, S.L.; Schachner, M. Duloxetine ameliorates valproic acid-induced hyperactivity, anxiety-like behavior, and social interaction deficits in zebrafish. Autism Res. 2022, 15, 27–41. [Google Scholar] [CrossRef]
- Terbach, N.; Shah, R.; Kelemen, R.; Klein, P.S.; Gordienko, D.; Brown, N.A.; Wilkinson, C.J.; Williams, R.S.B. Identifying an uptake mechanism for the antiepileptic and bipolar disorder treatment valproic acid using the simple biomedical model Dictyostelium. J. Cell Sci. 2011, 124, 2267–2276. [Google Scholar] [CrossRef]
- Zellner, D.; Padnos, B.; Hunter, D.L.; MacPhail, R.C.; Padilla, S. Rearing conditions differentially affect the locomotor behavior of larval zebrafish, but not their response to valproate-induced developmental neurotoxicity. Neurotoxicol. Teratol. 2011, 33, 674–679. [Google Scholar] [CrossRef]
- Bai, W.J.; Luo, X.G.; Jin, B.H.; Zhu, K.S.; Guo, W.Y.; Zhu, X.Q.; Qin, X.; Yang, Z.X.; Zhao, J.J.; Chen, S.R.; et al. Deficiency of transmembrane AMPA receptor regulatory protein γ-8 leads to attention-deficit hyperactivity disorder-like behavior in mice. Zool. Res. 2022, 43, 851. [Google Scholar] [CrossRef]
- Maltezos, S.; Horder, J.; Coghlan, S.; Skirrow, C.; O’Gorman, R.; Lavender, T.J.; Mendez, M.A.; Mehta, M.; Daly, E.; Xenitidis, K.; et al. Glutamate/glutamine and neuronal integrity in adults with ADHD: a proton MRS study. Transl. Psychiatry 2014 43 2014, 4, e373–e373. [Google Scholar] [CrossRef]
- Naaijen, J.; Bralten, J.; Poelmans, G.; Glennon, J.C.; Franke, B.; Buitelaar, J.K.; Faraone, S.; Asherson, P.; Banaschewski, T.; P Ebstein, R.; et al. Glutamatergic and GABAergic gene sets in attention-deficit/hyperactivity disorder: association to overlapping traits in ADHD and autism. Transl. Psychiatry 2017 71 2017, 7, e999–e999. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.; Liu, A.; Shi, M.Y.; Yan, Z. Disrupted Glutamatergic Transmission in Prefrontal Cortex Contributes to Behavioral Abnormality in an Animal Model of ADHD. Neuropsychopharmacol. 2017 4210 2017, 42, 2096–2104. [Google Scholar] [CrossRef]
- Bailey, J.M.; Oliveri, A.N.; Karbhari, N.; Brooks, R.A.J.; De La Rocha, A.J.; Janardhan, S.; Levin, E.D. Persistent behavioral effects following early life exposure to retinoic acid or valproic acid in zebrafish. Neurotoxicology 2016, 52, 23–33. [Google Scholar] [CrossRef]
- Montanari, M.; Martella, G.; Bonsi, P.; Meringolo, M. Autism Spectrum Disorder: Focus on Glutamatergic Neurotransmission. Int. J. Mol. Sci. 2022, Vol. 23, Page 3861 2022, 23, 3861. [Google Scholar] [CrossRef] [PubMed]
- Rojas, D.C. The role of glutamate and its receptors in autism and the use of glutamate receptor antagonists in treatment. J. Neural Transm. 2014, 121, 891–905. [Google Scholar] [CrossRef] [PubMed]
- Wright, J.W.; Harding, J.W. Contributions of Matrix Metalloproteinases to Neural Plasticity, Habituation, Associative Learning and Drug Addiction. Neural Plast. 2009, 2009, 579382. [Google Scholar] [CrossRef] [PubMed]
- Faria, M.; Bedrossiantz, J.; Prats, E.; Garcia, X.R.; Gómez-Canela, C.; Piña, B.; Raldúa, D. Deciphering the mode of action of pollutants impairing the fish larvae escape response with the vibrational startle response assay. Sci. Total Environ. 2019, 672, 121–128. [Google Scholar] [CrossRef]
- Mora-Zamorano, F.X.; Svoboda, K.R.; Carvan, M.J. The nicotine-evoked locomotor response: A behavioral paradigm for toxicity screening in zebrafish (Danio rerio) embryos and eleutheroembryos exposed to methylmercury. PLoS One 2016, 11, 1–15. [Google Scholar] [CrossRef]
- Kalueff, A. V. The rights and wrongs of zebrafish: Behavioral phenotyping of zebrafish; 2017; ISBN 9783319337746.
- Abbas, K.; Saputra, F.; Suryanto, M.E.; Lai, Y.H.; Huang, J.C.; Yu, W.H.; Chen, K.H.C.; Lin, Y.T.; Hsiao, C. Der Evaluation of effects of ractopamine on cardiovascular, respiratory, and locomotory physiology in animal model zebrafish larvae. Cells 2021, 10. [Google Scholar] [CrossRef]
- Basnet, R.M.; Zizioli, D.; Taweedet, S.; Finazzi, D.; Memo, M. Zebrafish larvae as a behavioral model in neuropharmacology. Biomedicines 2019, 7. [Google Scholar] [CrossRef]
- McCutcheon, V.; Park, E.; Liu, E.; Wang, Y.; Wen, X.Y.; Baker, A.J. A Model of Excitotoxic Brain Injury in Larval Zebrafish: Potential Application for High-Throughput Drug Evaluation to Treat Traumatic Brain Injury. Zebrafish 2016, 13, 161–169. [Google Scholar] [CrossRef] [PubMed]
- Horzmann, K.; Freeman, J. Zebrafish Get Connected: Investigating Neurotransmission Targets and Alterations in Chemical Toxicity. Toxics 2016, 4, 19. [Google Scholar] [CrossRef] [PubMed]
- Chefer, V.I.; Thompson, A.C.; Zapata, A.; Shippenberg, T.S. Overview of Brain Microdialysis. Curr. Protoc. Neurosci. 2009, 47, 7.1.1–7.1.28. [Google Scholar] [CrossRef] [PubMed]
- Kuo, H.Y.; Liu, F.C. Pathophysiological Studies of Monoaminergic Neurotransmission Systems in Valproic Acid-Induced Model of Autism Spectrum Disorder. Biomedicines 2022, 10. [Google Scholar] [CrossRef]
- Ali, E.H.A.; Elgoly, A.H.M. Combined prenatal and postnatal butyl paraben exposure produces autism-like symptoms in offspring: Comparison with valproic acid autistic model. Pharmacol. Biochem. Behav. 2013, 111, 102–110. [Google Scholar] [CrossRef] [PubMed]
- Pavǎl, D. A Dopamine Hypothesis of Autism Spectrum Disorder. Dev. Neurosci. 2017, 39, 355–360. [Google Scholar] [CrossRef]
- Nguyen, M.; Roth, A.; Kyzar, E.J.; Poudel, M.K.; Wong, K.; Stewart, A.M.; Kalueff, A. V. Decoding the contribution of dopaminergic genes and pathways to autism spectrum disorder (ASD). Neurochem. Int. 2014, 66, 15–26. [Google Scholar] [CrossRef]
- Schür, R.R.; Draisma, L.W.R.; Wijnen, J.P.; Boks, M.P.; Koevoets, M.G.J.C.; Joëls, M.; Klomp, D.W.; Kahn, R.S.; Vinkers, C.H. Brain GABA levels across psychiatric disorders: A systematic literature review and meta-analysis of 1H-MRS studies. Hum. Brain Mapp. 2016, 37, 3337–3352. [Google Scholar] [CrossRef]
- Purkayastha, P.; Malapati, A.; Yogeeswari, P.; Sriram, D. A Review on GABA/Glutamate Pathway for Therapeutic Intervention of ASD and ADHD. Curr. Med. Chem. 2015, 22, 1850–1859. [Google Scholar] [CrossRef]
- Kimmel, C.B.; Ballard, W.W.; Kimmel, S.R.; Ullmann, B.; Schilling, T.F. Stages of embryonic development of the zebrafish. Dev. Dyn. 1995, 203, 253–310. [Google Scholar] [CrossRef]
- Ricarte, M.; Prats, E.; Montemurro, N.; Bedrossiantz, J.; Bellot, M.; Gómez-Canela, C.; Raldúa, D. Environmental concentrations of tire rubber-derived 6PPD-quinone alter CNS function in zebrafish larvae. Sci. Total Environ. 2023, 896, 165240. [Google Scholar] [CrossRef] [PubMed]
- Bellot, M.; Manen, L.; Prats, E.; Bedrossiantz, J.; Barata, C.; Gómez-Canela, C.; Antolin, A.A.; Raldúa, D. Short-term exposure to environmental levels of nicotine and cotinine impairs visual motor response in zebrafish larvae through a similar mode of action: Exploring the potential role of zebrafish α7 nAChR. Sci. Total Environ. 2024, 912, 169301. [Google Scholar] [CrossRef] [PubMed]
- Bedrossiantz, J.; Prats, E.; Raldúa, D. Neurotoxicity Assessment in Adult Danio rerio using a Battery of Behavioral Tests in a Single Tank. J. Vis. Exp. 2023, 2023. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Canela, C.; Tornero-Cañadas, D.; Prats, E.; Piña, B.; Tauler, R.; Raldúa, D. Comprehensive characterization of neurochemicals in three zebrafish chemical models of human acute organophosphorus poisoning using liquid chromatography-tandem mass spectrometry. Anal. Bioanal. Chem. 2018, 410, 1735–1748. [Google Scholar] [CrossRef]
- Prats, E.; Gómez-Canela, C.; Ben-Lulu, S.; Ziv, T.; Padrós, F.; Tornero, D.; Garcia-Reyero, N.; Tauler, R.; Admon, A.; Raldúa, D. Modelling acrylamide acute neurotoxicity in zebrafish larvae. Sci. Rep. 2017, 7, 1–12. [Google Scholar] [CrossRef]
Figure 1.
Effects of embryonic exposure to VPA on the behavior of zebrafish larvae. (A) Percentage of basal locomotor activity relative to control; (B) Percentage of light-off visual motor response relative to control; (C) Percentage of vibrational startle response relative to control. Data reported as scatter plot with the median (n = 160-209). *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001; Mann-Whitney U test. Data from 3 independent experiments.
Figure 1.
Effects of embryonic exposure to VPA on the behavior of zebrafish larvae. (A) Percentage of basal locomotor activity relative to control; (B) Percentage of light-off visual motor response relative to control; (C) Percentage of vibrational startle response relative to control. Data reported as scatter plot with the median (n = 160-209). *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001; Mann-Whitney U test. Data from 3 independent experiments.
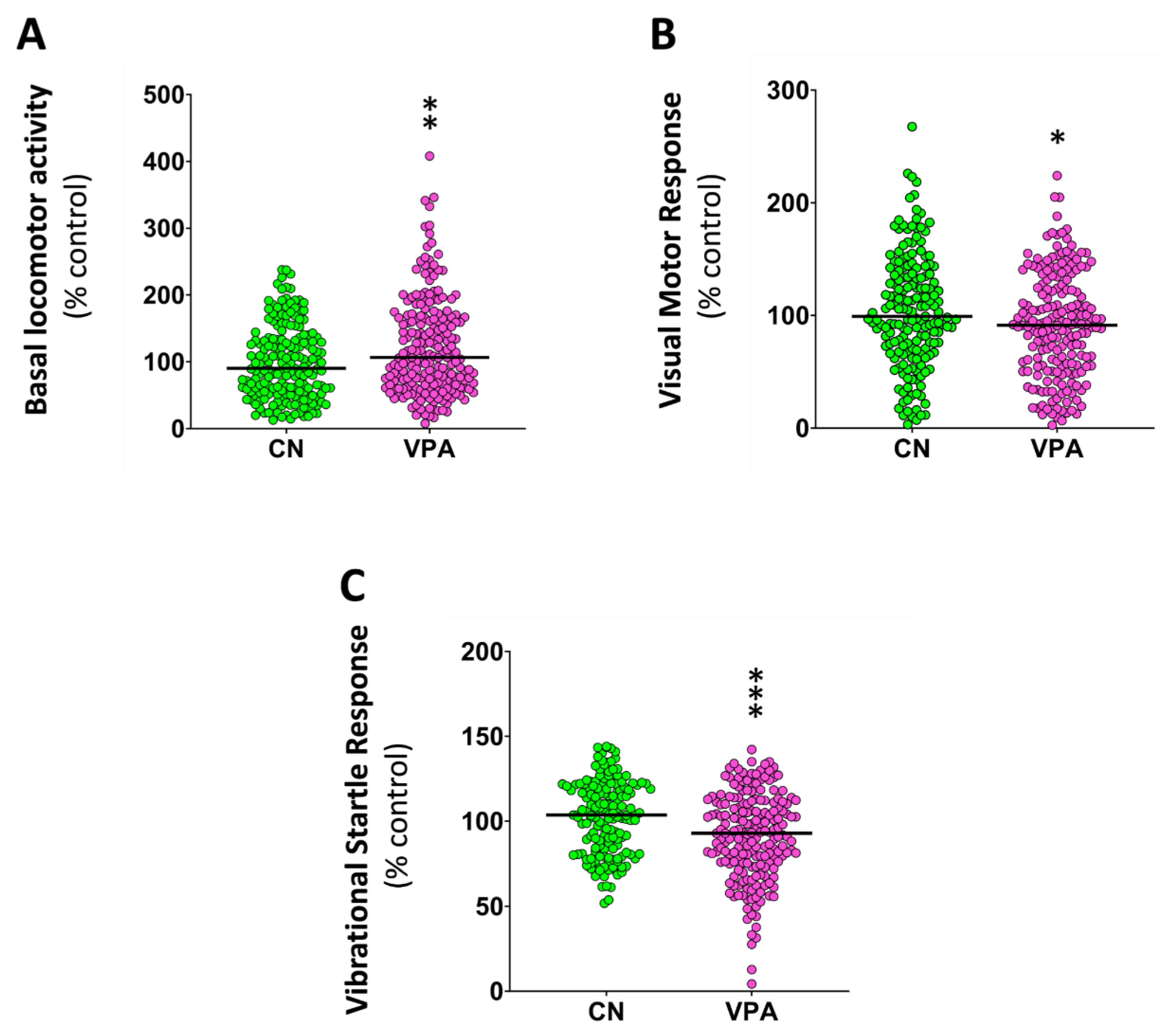
Figure 2.
Effects of embryonic exposure to VPA on the neurotransmitter profile of zebrafish larvae. The neurotransmitters analyzed in the larval heads include catecholamines (dopamine, norepinephrine, epinephrine), serotonin, acetylcholine, GABA, aspartic acid, and glutamate. Data reported as scatter plot with median (n = 7) *p < 0.05, **p < 0.01; Mann-Whitney U test. Data from two independent experiments.
Figure 2.
Effects of embryonic exposure to VPA on the neurotransmitter profile of zebrafish larvae. The neurotransmitters analyzed in the larval heads include catecholamines (dopamine, norepinephrine, epinephrine), serotonin, acetylcholine, GABA, aspartic acid, and glutamate. Data reported as scatter plot with median (n = 7) *p < 0.05, **p < 0.01; Mann-Whitney U test. Data from two independent experiments.
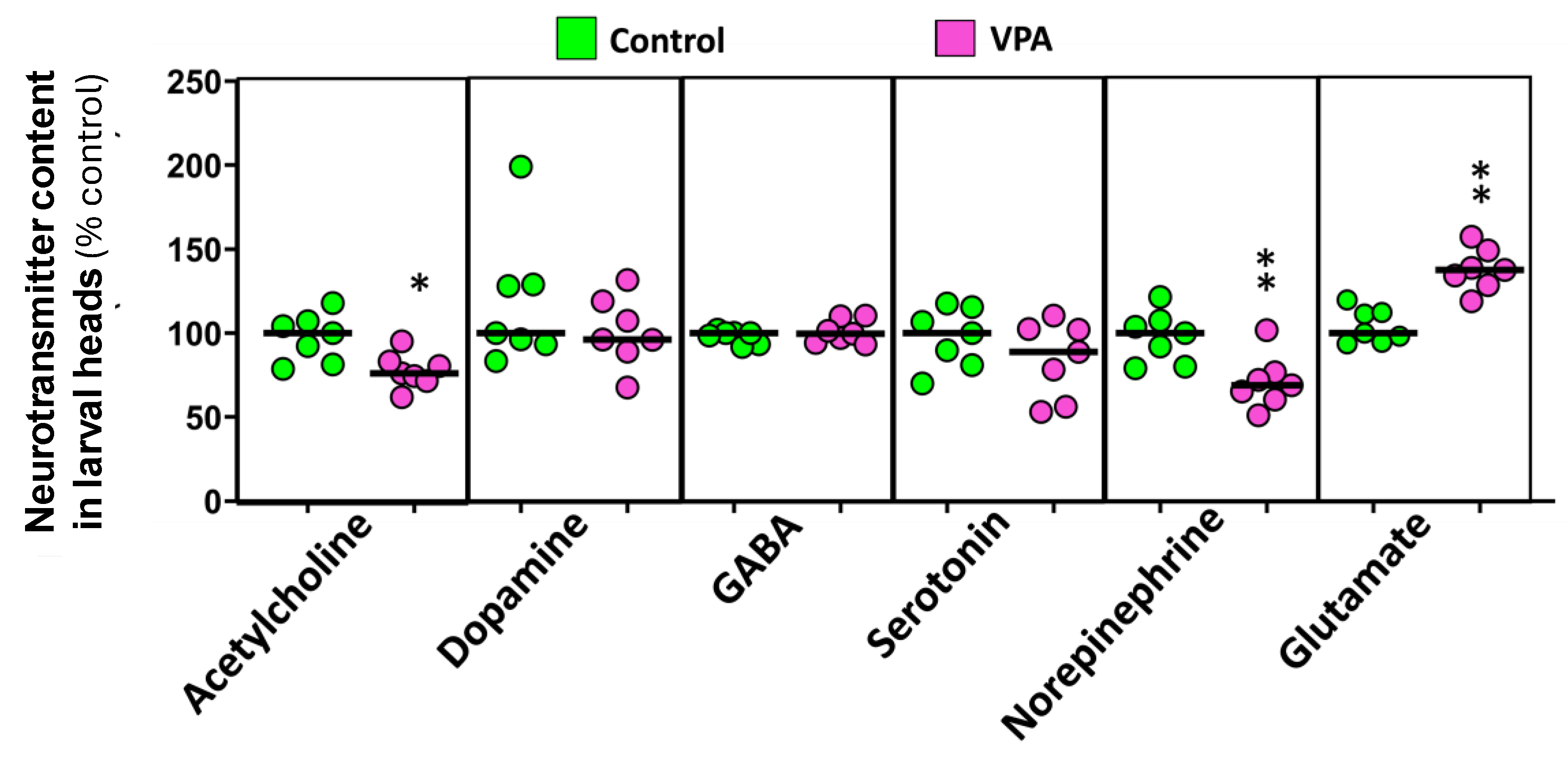
Figure 3.
Effect of early-life VPA exposure on social behavior. (A) VPA-exposure during the early embryonic development increases the average interfish distance and the farthest interfish distance in the shoaling test. (B) VPA-exposure during early embryonic development decreases the time fish spend in the zone closest to conspecifics, increasing, however, the time spent in the empty zone in the social preference test (SPT). In the SPT, the time spent in each virtual zone for each fish was normalized to the total time of the assay. Data reported as scatter plot with median (n = 16-18) *p < 0.05, **p < 0.01, ***p < 0.001; t-test or Mann-Whitney U test. Data from two independent experiments.
Figure 3.
Effect of early-life VPA exposure on social behavior. (A) VPA-exposure during the early embryonic development increases the average interfish distance and the farthest interfish distance in the shoaling test. (B) VPA-exposure during early embryonic development decreases the time fish spend in the zone closest to conspecifics, increasing, however, the time spent in the empty zone in the social preference test (SPT). In the SPT, the time spent in each virtual zone for each fish was normalized to the total time of the assay. Data reported as scatter plot with median (n = 16-18) *p < 0.05, **p < 0.01, ***p < 0.001; t-test or Mann-Whitney U test. Data from two independent experiments.
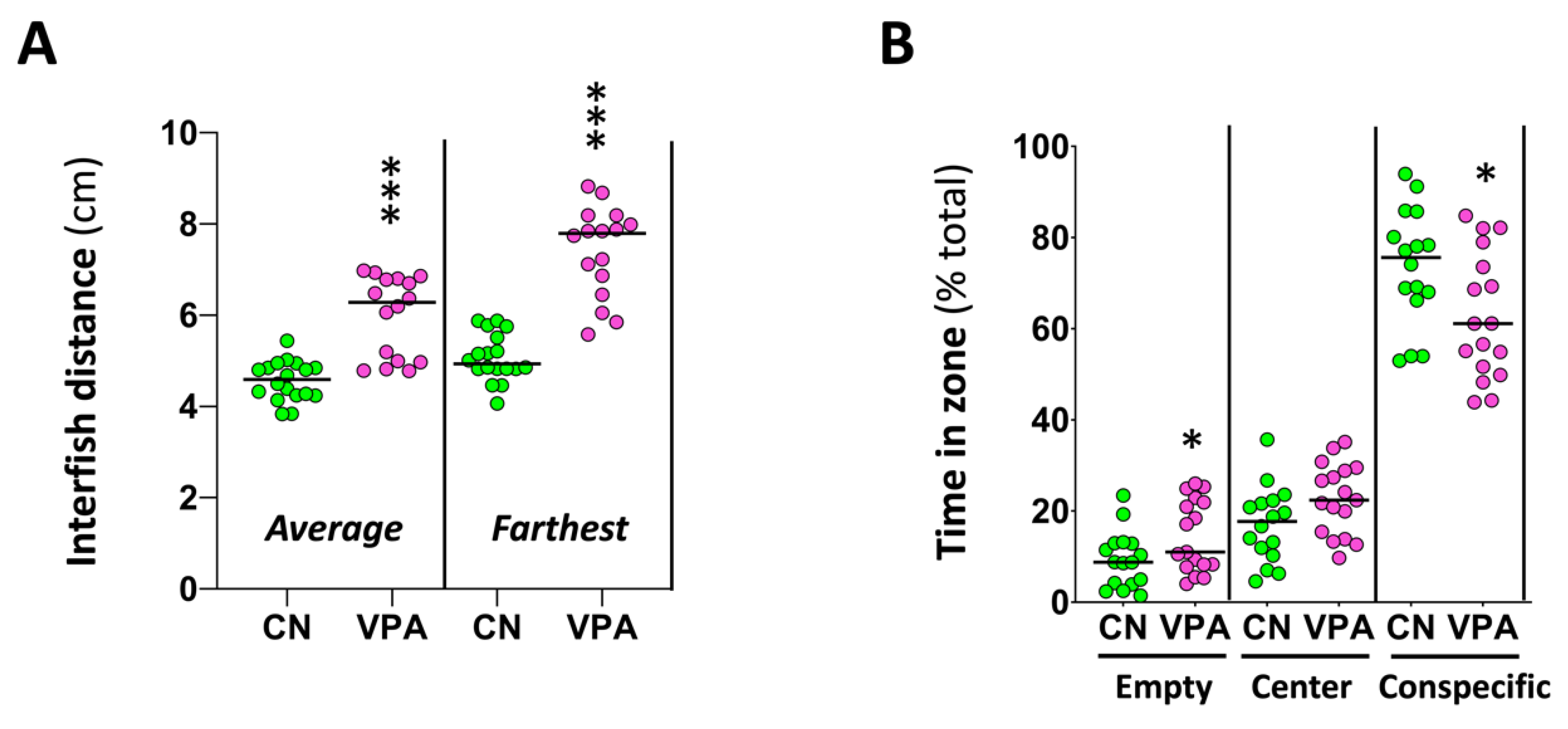
Figure 4.
Effects of embryonic exposure to VPA on the neurotransmitter profile in the brain of adult zebrafish. The neurotransmitters analyzed include catecholamines (dopamine and norepinephrine), serotonin, acetylcholine, GABA, and glutamate. Data reported as scatter plot with median (n = 6) *p < 0.05; Mann-Whitney U test. Data from two independent experiments.
Figure 4.
Effects of embryonic exposure to VPA on the neurotransmitter profile in the brain of adult zebrafish. The neurotransmitters analyzed include catecholamines (dopamine and norepinephrine), serotonin, acetylcholine, GABA, and glutamate. Data reported as scatter plot with median (n = 6) *p < 0.05; Mann-Whitney U test. Data from two independent experiments.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



