
Submitted:
21 June 2024
Posted:
24 June 2024
You are already at the latest version
Abstract
Salivary gland tumors are highly variable in clinical presentation and histology. The World Health Organization (WHO) classifies 22 types of malignant and 11 types of benign tumors of the salivary glands. Diagnosis of salivary gland tumors is based on imaging (ultrasound, magnetic resonance imaging) and fine-needle aspiration biopsy, but the final diagnosis is based on histopathological examination of the removed tumor. In this pilot study, we are testing a new approach to identifying peptide biomarkers in saliva that can be used to diagnose salivary gland tumors. The research material for the peptidomic studies was extracts from washings of neoplastic tissues and healthy tissues (control samples). At the same time, saliva samples from patients and healthy individuals were analysed. The comparison of the peptidome composition of tissue extracts and saliva samples may allow the identification of potential peptide markers of salivary gland tumors in patients' saliva. The peptidome composition extracted from 18 tumor and 18 healthy tissue samples, patient (11 samples) and healthy saliva samples (8 samples) was analysed by LC-MS tandem mass spectrometry. A group of 109 peptides were identified that were present only in the tumor tissue extracts and in the patient saliva samples. Some of the identified peptides were derived from proteins previously suggested as potential biomarkers of salivary gland tumors (ANXA1, BPIFA2, FGB, GAPDH, HSPB1, IGHG1, VIM) or tumors of other tissues or organs (SERPINA1, APOA2, CSTB, GSTP1, S100A8, S100A9, TPI1). Unfortunately, none of the identified peptides were present in all samples analysed. This may be due to the high heterogeneity of this type of cancer. The surprising result was that extracts from tumor tissue did not contain peptides derived from salivary gland-specific proteins (STATH, SMR3B, HTN1, HTN3). These results could suggest that the developing tumor suppresses the production of proteins that are essential components of saliva.
Keywords:
salivary glands
; saliva
; cancer
; biomarkers
; peptidome
1. Introduction
Neoplasms of the major salivary glands account for approximately 3% of all tumors in the head and neck region. Most tumors develop in the parotid gland (80%), of which 4/5 are benign lesions [1,2]. These tumors are very different regarding their histology and clinical course. The World Health Organization (WHO) classification distinguishes 22 types of malignant and 11 benign tumors originating from the salivary glands [3]. Diagnostics of salivary gland tumors are based on imaging (ultrasound, magnetic resonance imaging) and fine-needle aspiration biopsy; however, the final diagnosis is based on the histopathological examination of the removed tumor [2,4]. The most common treatment for salivary glands tumors is surgical removal and complementary radiotherapy in the case of cancer detection. Removal of the parotid gland is always connected with a risk of damaging the facial nerve, which causes significant functional disorders (disorders of the mouth, eyes closing) and cosmetic defects. In the case of benign tumors, the priority is to maintain the anatomical continuity of the facial nerve. In the case of a malignant tumor, its radical removal is the most important, even with the sacrifice of the facial nerve [2,5,6]. Therefore, preoperative diagnostics differentiating malignant from benign changes are crucial. The prognosis for benign neoplasms is excellent, as the recurrence rate is about 1-2%. The disease-free 5-year survival is observed in about 60% of malignant parotid tumors cases, which depends on their stage and grade [2,7]. The signs and symptoms indicating malignancy of salivary gland tumors (e.g. facial nerve palsy, skin infiltration) are related to the highest stage of disease and poor prognosis. Clinical picture, modern imaging techniques, and fine needle aspiration biopsy (FNA) do not allow distinguishing between benign and malignant tumors in every case; thus, other diagnostic methods (i.a., based on saliva testing) are developed [8].
Due to their rarity and high heterogeneity, salivary gland tumors remain relatively poorly characterized in terms of molecular analysis; most of the studies on this topic have focused on the genetic aspect of tumorigenesis [9], demonstrating, among other things, the presence of characteristic fusion oncogenes in salivary gland tumors [10]. Recently, there has been a gradual increase in interest in proteomic studies using mass spectrometric methods as a source of new tumor biomarkers [11]. Although various proteins [12,13,14,15] have been proposed as potential biomarkers for different salivary gland tumors, their diagnostic efficacy has not been confirmed [11]. No study has been published to date analyzing salivary gland tumors at the peptidomic level. The peptidome – consisting of thousands of peptides: remnants of protein degradation; specific precursor-derived peptides such as hormones, cytokines, or growth factors; and translation products of small open reading frames (smORFs) [16] – responds dynamically to changes in the body, providing a rich source of potential biomarkers. This results in the rapid expansion of peptidomics research observed in recent years [17,18], demonstrating characteristic peptidomic patterns of many diseases, including various types of cancer [19,20,21]. Furthermore, it has been shown that peptidomic analyses can detect statistically significant differences observed in bottom-up proteomics [22].
Proteomic studies of salivary tumor tissues known from the literature used either homogenized frozen [15] or FFPE tissues [12,23]. Homogenized tissues are a commonly used material in proteomic research. However, its major disadvantage is the presence of many proteins (some of these are in high concentrations) from the examined tissue, making it challenging to identify potential biomarkers. An interesting solution that allows the analysis of tissues derived from tumor tissues of the salivary glands without the need to use homogenates is the method of analyzing the composition of the FNA fluid (a saline wash of needle after fine needle aspiration of tumor) [12,13]. Using the FNA fluid as a source of proteins/peptides deserves attention, given the widespread use of FNA in diagnosing salivary gland tumors. The problems related to the FNA fluid analysis are the minimal amount of the sample and the problem related to the FNA procedure itself, i.e., the high probability of collecting the sample from the tissue not covered by the tumor.
In this pilot study, we propose a new approach for identification of peptide biomarkers that can be used in diagnosing tumors of the salivary glands. As a research material for peptidomic studies, we use extracts from washing neoplastic tissues and healthy tissues (control samples). At the same time, samples of saliva from patients, as well as from healthy individuals, will be analyzed. Comparison of the peptidome composition of tissue extracts and saliva samples may allow the identification of potential peptide markers of a salivary gland tumor in patients' saliva.
2. Results
In our research we use fresh or frozen (samples are referred to as fresh or frozen) salivary gland tissue samples, healthy or tumor. Peptides are extracted from the tissue samples using phosphate buffer (PBS extracts) or 0.1% trifluoroacetic acid solution in water (TFA extracts). A more detailed description of sample preparation is provided in the Methods section.
We analysed tissue samples from 14 patients. In addition, we analysed saliva samples from 8 healthy donors (control) and 11 patients. Information on the samples analysed, as well as the handling of the tissue samples (frozen or fresh) and the peptide extraction methods are summarised in Table 1. Patient information is presented in Supplementary Table 1. Detailed information on each of the extracts used in the study is provided in Supplementary Table 2.
2.1. Comparison of Peptide Extraction Methods
The first step of our research was to compare the peptidome composition extracted from tissue samples obtained by two different extraction methods: (i) extraction with PBS alone (referred to in the text as the PBS method); (ii) extraction with 0.1% TFA solution following PBS extraction (referred to in the text as the TFA method). 20 PBS extracts and 16 TFA extracts from different samples were used in the analysis. A total of 7771 peptides were identified in all extracts obtained with PBS extracts, more than twice as many as in the TFA extracts, where only 3540 peptides were identified (Supplementary Figure S1). The two types of extracts also differed significantly in composition, with only about 9% (931 out of 10380) of all unique peptides identified in both PBS and TFA extracts. The PBS extracts (bearing in mind that the PBS wash was performed first) were richer in peptides, but the extraction with TFA allowed the isolation of a different fraction of peptides. On average, about 388 unique peptides were identified in the PBS extract samples and about 221 unique peptides were identified in the TFA extract samples. On the other hand, we tested the effect of sample handling on peptide extraction by comparing extracts obtained from 18 frozen and 18 fresh salivary gland samples (Supplementary Figure S1). A total of 10,380 unique peptides were identified in all samples [Supplementary Table S3], with an average of approximately 288 unique peptides per sample. Similar numbers of peptides were identified in both extracts (6751 in fresh and 6913 in frozen). Almost one third of all identified peptides (3284 out of 10,380) were common to both fresh and frozen tissue extracts. The remaining peptides were clustered into two almost equal sets (3467 and 3629 peptides), also close to 1/3 of the total pool, containing peptides unique to either fresh or frozen tissue extracts. Thus, although fresh and frozen salivary gland tissue extracts are similar in terms of the number of peptides identified, they differ significantly in terms of peptidome composition. The results of the experiments shown above indicate that there is no best sample preparation or extraction method in terms of the number of unique peptides identified. Each of the analysis options shown above yields a significant number of unique peptides specific to the type of sample preparation. Therefore, the optimal method for sample preparation and peptide extraction could not be identified. The following sections analyse the results of all the measurements performed.
2.2. Comparison Peptidome Composition of Salivary Glands Extracts and Saliva
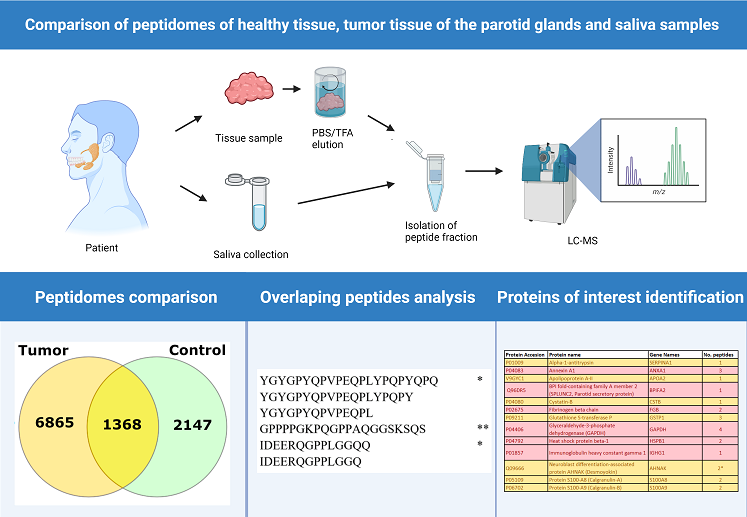
To investigate differences in the peptidome of salivary gland tumors and healthy tissue, we compared 18 extracts (both PBS and TFA) obtained from tumor tissue and 18 extracts from healthy control tissue [Figure 1A]. The extracts from tumor tissue were significantly richer in peptides, with 8233 unique peptides identified, compared to only 3515 peptides identified in the control samples. Surprisingly only a small number of peptides (1368) were shared between the tumor and healthy tissue (see Figure 1A). In similar manner we analyzed saliva samples. The study used saliva samples from 8 healthy subjects (control) and 11 patients. 3272 peptides were identified in patient saliva, compared to 2461 peptides identified in control samples. 1474 peptides were standard for both sample types, while 1798 peptides were found only in patient saliva (see Figure 1B). Two methods were used to isolate the peptide fraction: Direct method and SPE extraction (see Materials and methods). SPE extraction resulted in a significantly higher number of identified peptides, so all patient samples underwent this procedure.
2.3. Bioinformatic Analysis
In searching for potential biomarkers of salivary gland cancer, peptides with overlapping sequences are of little interest. The peptides identified in each of the analyses (see Section 2.2) were compared with each other for overlapping sequences, and then only those not contained in the sequence of other peptides were selected as unique peptides. In addition, stand-alone peptides were also identified that were not included in the sequence of others and did not contain any identified peptides themselves (see Figure 2). As it is shown in Figure 2 peptides do not mark * or ** are not consider in further analysis.
Table 2 provides a number of all peptides identified in each analysis, categorized into unique and stand-alone statuses. Less than half of all identified peptides could be classified as unique peptides, and 10-20% as stand-alone peptides. There are no significant differences between the content of unique and stand-alone peptides between tumor samples and controls and between different extraction methods. However, we noted noticeable differences between the peptide composition of samples from salivary glands and saliva. Unique peptides accounted for 46.9% of all peptides identified in salivary gland samples (4869 of 10380) and stand-alone 20% (2077 of 10380). For saliva samples, the values are much lower, at 35% for unique peptides (1490 of 4259) and 10% for stand-alone peptides (420 of 4259). Lower number of unique and stand-alone peptides in saliva samples probably are due higher proteolytic activity in saliva caused by presence of large number of microorganisms in oral cavity.
The next step was to look at peptides identified in saliva and compare them with peptide sequences from salivary gland extracts. We assumed that intense proteolytic enzyme activity occurs in human saliva, so we focused on looking for saliva peptides whose sequences are identical with peptides identified in salivary glands extracts (common peptides) or entirely contained within the sequences of longer peptides of salivary glands origin (overlapping sequences). The results of this analysis are presented in Table 3.
We analyzed the peptide data sets obtained in the study (e.g., saliva patient) and the sets created by excluding peptides in common with the control (e.g., saliva patient only). The comparison of peptides specific to patients' saliva (Saliva patient only) with peptides specific to salivary gland tumors (SG tumor only) is particularly noteworthy. The peptides identified in this analysis may be potential biomarkers to characterize salivary gland tumors. We found 16 peptides that are common to both data sets compared (Saliva patient only and SG tumor only), but also 109 peptides identified in saliva whose sequences were contained within the sequences of longer peptides identified in salivary gland tumor extracts. Sequences of peptides identified in the group Saliva patient only - SG tumor only (see Table 3) are shown in Supplementary Table S4.
2.4. Characterization of Peptide Biomarkers of Salivary Gland Tumors
Our research focuses on peptidomic characterization of salivary gland tumors, but it is challenging to analyze peptides in isolation from the parent protein. Therefore, the next step was to look at the proteins from which the peptides were identified as observed only in patient saliva samples and extracts from tumor tissue (Saliva patient only - SG tumor only in Table 3). All 109 peptides were assigned to 24 protein groups (see Table 4). Typical saliva-specific proteins were the most heavily represented, accounting for nearly half of all identified peptides. Essential belongs to basic salivary proline-rich proteins (32 peptides) (marked white in Table 4) and salivary acidic proline-rich phosphoprotein (17 peptides). In addition, one peptide belonging to the parotid secretory SPLUNC2 (short palate, lung and nasal epithelium clone 2) protein was identified. Among other proteins, relatively many peptides were identified for actin (9 peptides), histones (9 peptides), hemoglobin (5 peptides), and keratin (4 peptides) (marked white in Table 4). These proteins, and consequently the peptides derived from them, are commonly found in many tissues, so, like saliva-specific proteins, they do not appear to be particularly interesting from a diagnostic point of view.
The situation is different for the remaining proteins: 17 of them, identified based on 30 peptides, have been linked in the literature to various malignancies. The studies that report those findings concern proteomics, not peptidomics, but they remain an exciting benchmark for considering the biological significance of the peptides found in this study.
At least 7 (marked in red in Table 4) of these 15 proteins have previously been shown to have differential expression in salivary gland tumors: annexin A1 (ANXA1), glyceraldehyde-3-phosphate dehydrogenase (GAPDH), fibrinogen beta chain (FBG), heat shock protein beta-1 (HSPB1), immunoglobulin gamma-1 chain C region (IGHG1), BPI fold-containing family A member 2 (BPIFA2), and vimentin (VIM). Donadio et al. [13] observed that ANXA1 and GAPDH are up-regulated in pleomorphic adenomas; and FBG, HSPB1, and IGHG1 are up-regulated in Warthin's tumors. Also, Seccia et al. [12] observed up-regulation of ANXA1 in pleomorphic adenomas and IGHG1 in Warthin's tumors, while Mutlu et al. [15] observed up-regulation of VIM and down-regulation of FGB in pleomorphic adenomas patients. Pereira et al. [24] suggested BPIFA2 as a potential biomarker for salivary mucoepidermoid carcinomas. González-Arriagada et al. [25] showed that BPIFA2 level in head and neck cancer decreases after radiotherapy.
To our knowledge, the remaining proteins (marked yellow in Table 4) have not been associated with salivary gland tumors but are associated with other types of cancer. Overexpression of alpha-1-antitrypsin (SERPINA1) promotes tumor progression in colorectal [26] and gastric [27] cancer, and its high level in plasma was observed in lung and prostate cancer patients [28]. Apolipoprotein A-II (APOA2) was proposed as a potential marker for urinary bladder [29] and pancreatic cancer [30]. Cystatin B was proposed as a biomarker in various tumors like bladder and ovarian cancer [31,32]. Glutathione S-transferase P (GSTP1) is downregulated in prostate cancer and up-regulated in many cancer types, e.g., colorectal, thyroid, or breast cancer [33]. Also, a high level of glyceraldehyde-3-phosphate dehydrogenase (GAPDH) was found in various cancer types [34]. AHNAK was proposed as a biomarker for bladder urothelial carcinoma [35] and is strictly related to cell migration in mesothelioma [36]. Calprotectin, a heterodimer formed from the combination of S100A8 and S100A9 proteins, is commonly up-regulated in many tumors and likely plays a critical (essential) role in inflammation-associated cancers [37]. Protein-glutamine gamma-glutamyltransferase E (TGM3) was proposed as a marker for some head and neck cancer types [38,39]. Thymosin beta-4 (TMSB4X) is overexpressed in pancreatic cancer [40] and was proposed as a biomarker for colorectal cancer [41]. Triosephosphate isomerase (TPI1) is up-regulated in neoplasms like gastric or breast cancer and is related to cell migration and invasion [42,43].
Analyzing the peptidome of tumor tissue extracts, a complete absence of peptides derived from saliva-specific proteins was observed. As it is shown in Figure 3 and Supplementary Figure S2 extracts derived from tumor tissue do not contain peptides derived from statherin (STATH), submaxillary gland androgen-regulated protein 3b (SMR3B), histatin 1 (HTN1) and histatin 3 (HTN3) proteins. Peptides derived from these four proteins are abundantly represented in healthy tissue extracts and saliva samples (both patient and control). To ensure quality of identification we compare our results with reference data form Peptide Atlas database [44]. As it is shown Supplementary Table S5, for all mentioned proteins we detect larger number of peptides than is deposited in Peptide Atlas database [44]. In the case of histatin 1 and histatin 3 we can identify in one extract sample from tumor tissue only one peptide belonging to those proteins, we suspect that such identification in caused by not perfect separation of healthy tissue form tumor (see panel B and C in Supplementary Figure S2). It is very interesting that all four proteins mentioned above are specifically produced in salivary glands [45] and their genes are located on chromosome 4 in very close proximity one to another (www.ncbi.nlm.nih.gov/gene/3347;www.ncbi.nlm.nih.gov/gene/3346;www.ncbi.nlm.nih.gov/gene/6779; www.ncbi.nlm.nih.gov/gene/10879). The lack of identification of peptides from proteins STATH, SMR3B, HTN1, HTN3 in extracts from tumor tissues may suggest that there is a complete suppression of the production of these proteins in tumor tissues. It is of great interest that salivary gland tumors are extremely diverse [3] and in our study samples from different types of tumors were used (see Table S1). However, regardless of tumor type, the effect is the same lack of occurrence of peptides derived from proteins STATH, SMR3B, HTN1, HTN3. The fact that peptides from these proteins are present in the saliva of patients with detected cancer is related to the fact that not all salivary glands are occupied by cancer, so that the remaining healthy glands are able to produce the proteins in question.
3. Conclusions
The most interesting results for diagnostic reasons are the peptides identified only in the tissues of tumor tissue extracts and found in the patient's saliva (see Table 4 and Table S4). Two buffers (PBS and TFA) were used to extract peptides from tissues, and analyses were performed on fresh and frozen samples. It shows that out of 109 selected peptides (Table 4), 104 are identified in the frozen samples, while only 88 peptides are identified in the fresh samples. Based on these preliminary results, it seems more promising to analyze the frozen samples. This observation has a purely practical dimension. The use of frozen samples allows for an effective organization of analyses, which is extremely difficult when analyzing fresh samples. As for the buffers' effectiveness, 104 out of 109 selected peptides were identified in PBS extracts, and only 79 in TFA extracts. However, it should be noted that various types of acidic buffers have long been used to extract potentially immunogenic peptides that may be used in anti-cancer therapies [46,47]. Thus, even though the fractions from the acid extraction seem poorer, they are perhaps more interesting from a diagnostic point of view. In the planned research, we intend to use buffers and extraction protocols to isolate immunogenic peptides [46,47] to possibly expand the area of biomarker searches. Isolation of immunogenic peptides may be the beginning of the development of immunotherapy in this type of disease. Unfortunately, each of the peptides identified exclusively in tumor tissue extracts was only found in a maximum of 2-3 samples analyzed. This state of affairs may be related to the remarkable heterogeneity of salivary gland tumors [3]. Most of the samples examined in this work were from so-called mixed tumors (see Table S1). The results were obtained on a relatively small number of samples (18 tissue samples and 11 patient saliva samples) (see Table 1). The tumor tissue samples used for the study were derived from different types of tumors (see Supplementary Table S1). In addition, all data analyses were performed collectively for all available data and not for individual samples. The obtained results (see Table 4), despite the limitations presented above, are very promising and allow planning more targeted studies on a larger group of patients divided into different types of cancer.
Using the OpenProt library [48] to identify proteins/peptides gives exciting and promising results. However, as mentioned in the Introduction, this is a pilot study to show that the proposed new methodological solutions (peptide extraction, use of the OpenProt library) are applicable to the planned research. Using the OpenProt library does not generate additional costs and allows to increase the number of identified peptides by a few percent. Moreover, in the case of the data in Table 4, 3 out of 30 (10%) of the identified potential biomarkers were identified thanks to the OpenProt database [48]. These peptides would be omitted if classical UniProt or NCBI databases were used in searches. It should be noted however, that recent editions of the UniProt database [49] go in the direction of including not only the so-called canonical sequences, but also protein isoforms, which means that the content of the UniProt database also begins to include some of the information available only in the OpenProt database [48,50]. Reason for that could be that non-canonical products of eukaryotic genes (isoforms, pseudogenes) are of increasing interest in studies of tumorigenesis [51,52].
A surprising result of the studies presented here is the absence of peptide traces (in all samples tested) of four proteins (statherin (STATH), submaxillary gland androgen-regulated protein 3b (SMR3B), histatin 1 (HTN1) and histatin 3 (HTN3)) in tumor tissues. The aforementioned proteins are specifically expressed in salivary glands [45]. As shown in this study, no peptide traces of the proteins were found in the mixture of peptides extracted from tumor tissues of all analyzed samples (Figure 3): STATH, SMR3B, HTN1, HTN3. However, a significant number of peptides that are fragments of the proteins in question are present in healthy tissue samples obtained from the same patients. The simplest interpretation of these results seems to be the hypothesis that the onset of tumorigenesis in salivary gland tissue either completely or significantly stops the expression of these proteins. As described in the Results section, all four genes encoding these proteins are located in their immediate vicinity on chromosome 4, which may support the thesis that expression is blocked within a selected portion of the genetic material. Peptides derived from other proteins produced in salivary glands, e.g. basic salivary proline-rich protein 2, are observed in all types of samples (healthy tissue, cancerous tissue), but the gene encoding this protein (PRB2) is located on chromosome 12. The result obtained is extremely interesting for several reasons. Firstly, it may have enormous diagnostic significance even with current methods based on fine-needle biopsies after using appropriate antibodies to pick out healthy tissue (presence of proteins) and no staining with tumor tissue (absence of proteins). With the heterogeneity of salivary gland tumors, a method capable of distinguishing healthy tissue from neoplastic tissue would be extremely valuable. Secondly, if the expression of a specific number of genes is stopped during the tumorigenesis process, then perhaps further research would lead to the description of the mechanism of this process, which in the long run would open the way to new diagnostic tests and/or therapeutic methods.
4. Materials and Methods
4.1. Salivary Glands Tissue Collection
A fragment of the tumor and salivary gland tissue was collected after parotidectomy. Some samples were stored at 4oC after the surgery procedure, and peptide extraction was processed on the same day (those samples are called Fresh). The remaining samples were placed in a freezer at a temperature of -20oC within 4 hours of collection and processed later (those samples are called Frozen). A portion of each specimen was fixed in buffered formalin and sent to the Department of Pathomorphology of the University Clinical Center in Gdańsk (Medical University of Gdańsk) for further research (histological examination with immunohistochemical staining of the material). It is a generally accepted procedure during the diagnosis of neoplasms provided by the diagnostic protocol for patients with suspected neoplasms in the head and neck area.
4.2. Saliva Samples Collection
The collection of biological material (saliva) in the control group and the research group was performed by collecting the saliva collected in the oral cavity into a sterile tube by spitting the saliva directly into the tube. The above-described method of collecting biological material is a non-invasive method, the execution of which is painless, quick, and does not cause discomfort or other damage to the patient's health. The collected biological material does not require fixation. The samples were frozen at -20oC immediately after collection and stored until peptide extraction.
4.3. Isolation of Peptide Fractions from Salivary Gland Tissue
The general workflow of tissue sample handling is shown in Figure 4. Every tissue sample was processed with two steps of peptide elution. At first, samples were sliced into 5-10 small pieces to increase the elution surface. Then, the tissue was transferred to glass bottles and washed with 5 ml of ice-cold PBS. After 3 minutes of mix gently rinsing, PBS was collected and stored at -20oC as PBS Fraction 1. The whole procedure was repeated up to 8 times until all traces of blood were removed. The next step after PBS washing was peptide elution. Samples were eluted with 0.1% TFA in MS-grade water (called TFA). 5 ml of the appropriate, ice-cold elution solutions were added to the tissue and incubated with mixing for 3 minutes on ice. After that, the elution solution was removed and centrifuged for 5 minutes at 500xG, 4oC. The supernatant was collected and stored at -20oC as TFA extract. A total of 36 extracts were obtained from clinical samples taken from 14 different patients (Table 1). Frozen elution samples were lyophilized using CentriVap Cold Trap (Labconco, Kansas City, MO, USA). After that, lyophilisates were dissolved in 1 mL of MS-grade water and filtrated on Amicon filters with 10kDa NMWCO (Merck Millipore, Burlington, MA, USA). The flow-through fractions containing peptides were collected. The concentration of peptides was measured with Multiskan SkyHigh Microplate Spectrophotometer (Thermo Scientific, Waltham, MA, USA), and 10µg of every sample was desalted in STAGE (Stop And Go Extraction) TIPS procedure [53] using Empore C18 extraction disks (3M) with elution by 60% acetonitrile/1% acetic acid solution. Eluates were concentrated in SpeedVac to 25 µl volume and stored at -20oC for further LC-MS analysis.
4.4. Peptide Isolation from Saliva
Peptides were isolated from saliva by two methods. In the first (Direct method) method, 500 µl of saliva was filtrated on Amicon filters with 10kDa NMWCO. The concentration of peptides in the flow-through fraction was measured, and 10µg of every sample was desalted in STAGE TIPS procedure using Empore C18 extraction disks (3M) with elution by 60% acetonitrile/1% acetic acid solution. Eluates were concentrated in SpeedVac to 25 µl volume and stored at -20oC for further LC-MS analysis.
The second method was peptide isolation by solid phase extraction (SPE). 100 µl of saliva was diluted in 900 µl of TFA (0.1 or 0.5%) and incubated on ice for 5 minutes. After incubation, all samples were centrifuged for 5 minutes at 4oC and 5000xG. Samples were transferred to new tubes, diluted with 0.2% formic acid in a 1:1 ratio, mixed intensively for 20 seconds, and filtrated on Amicon filters with 10kDa NMWCO. The flow-through fraction proceeded on SupelcleanTM SPE tubes (LC-18 or HLB) (Supelco, Bellefonte, PA, USA) as previously described [54] and eluted with 500 µl 50% ACN/0.2% FA. The solvent was evaporated in SpeedVac to complete dryness, resuspended in 30 µl of 0.1% FA, and stored at -20oC for further LC-MS analysis.
4.5. LC-MS/MS
Liquid chromatography tandem mass spectrometry measurements were performed using the Triple TOF 5600+ mass spectrometer with DuoSpray Ion Source (SCIEX, Framingham, MA, USA) coupled with Ekspert MicroLC 200 Plus (Eksigent, Dublin, CA, USA) similar to the previously described [55]. The column used for chromatographic separation was ChromXP C18CL (3 μm, 120 Å, 150 × 0.3 mm; Eksigent, Dublin, CA, USA), and the sample injection volume was 5µL. Mobile phase A used in chromatography was 0.1% formic acid in water, and mobile phase B was 99.9% acetonitrile/0.1% formic acid. The gradient was 11% B for 2 minutes, then 48 minutes gradient to 43.5% B, 2 minutes gradient to 98% B, 98% B for 2 minutes, drop to 11% B and 11% B for 4 minutes in total flowrate 5 µL/min. The column temperature was stabilized at 35oC. The mass spectrometer operated in data-dependent acquisition (DDA) mode. MS1 scan was acquired in mass range 400-1200Da with an accumulation time of 100ms and followed by MS2 scan in range 100-1800Da with an accumulation time of 50ms. The raw data from the mass spectrometer was converted to a .mzML file with MSConvert software (ProteoWizard), then processed in PEAKS Studio software (Bioinformatics Solutions Inc.) [56] and searched against Homo sapiens OpenProt 1.6 database (May 2021) [48]. The mass spectrometry proteomics data have been deposited to the ProteomeXchange Consortium via the PRIDE [57] partner repository with the dataset identifier PXD038985.
4.6. Exploratory Data Analysis
Exploratory Data Analysis (EDA) of peptide sequences were performed by using custom scripts written in Python programming language (ver. 3.8.8) [58]. Loading, filtering, and cleaning of data was achieved by using NumPy (ver. 1.19.5) [59] and pandas (ver. 1.1.1) [60] libraries. To compare sequences (of peptides) with each other, built-in functions/methods and data structures from the standard library were applied. Jupyter-notebook (ver. 6.1.3) was used as an environment for computations and result validation.
4.7. Ethics Committee Approval and Personal Data Handling
The Regional Bioethics Committee approved the research protocol (NKBBN/308/2021). Written informed consent was obtained from each participant. Each participant could opt out of the study at any stage without any consequences. The obtained biological material was placed in sterile containers with a tight lid and marked with a number corresponding to the order of obtaining during the study, which allowed for segregation of the material and conducts of analysis and correlation with clinical data. Personal data has been classified and not placed on the containers used.
Supplementary Materials
Author Contributions
Conceptualization, D.T. and S.O.; methodology, M.P., D.T., P.C. and S.O.; investigation, M.P., A.S., D.S., K.S. B.M., A.O., N.M., M.T., P.C.; resources, S.O., P.C.; writing—original draft preparation, M.P., D.T., S.O.; writing—review and editing, M.P., D.T. and S.O.; supervision, S.O. and D.T.; funding acquisition, M.P. and S.O. All authors have read and agreed to the published version of the manuscript.
Funding
The research was funded by the research project "Young Creator of Science" of the Medical University of Gdansk, project number 71-01418, task number K15 0004729.
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki, and approved by the Institutional Bioethics Committee of Medical University of Gdańsk (NKBBN/308/2021, 2021.05.20).
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
The mass spectrometry proteomics data have been deposited to the ProteomeXchange Consortium via the PRIDE partner repository with the dataset identifier PXD038985.
Acknowledgments
Computational resources used in this project were provided by the Informatics Center of the Metropolitan Academic Network (IC MAN-TASK) in Gdańsk. Authors thank Dr Aleksandra Bogucka for proofreading the manuscript and valuable comments.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Sood, S.; McGurk, M.; Vaz, F. Management of Salivary Gland Tumors: United Kingdom National Multidisciplinary Guidelines. J. Laryngol. Otol. 2016, 130, S142–S149. [Google Scholar] [CrossRef] [PubMed]
- Jegadeesh, N.; Liu, Y.; Prabhu, R.S.; Magliocca, K.R.; Marcus, D.M.; Higgins, K.A.; Vainshtein, J.M.; Trad Wadsworth, J.; Beitler, J.J. Outcomes and prognostic factors in modern era management of major salivary gland cancer. Oral Oncol. 2015, 51, 770–777. [Google Scholar] [CrossRef] [PubMed]
- Mikaszewski, B.; Markiet, K.; Smugała, A.; Stodulski, D.; Szurowska, E.; Stankiewicz, C. Parotid malignancies are not homogenous in terms of their presentation on dynamic MRI - a preliminary report. Acta Otolaryngol. 2017, 137, 215–220. [Google Scholar] [CrossRef] [PubMed]
- Tretiakow, D.; Mikaszewski, B.; Skorek, A. The role of fine-needle aspiration biopsy (FNAB) in the diagnostic management of parotid gland masses with emphasis on potential pitfalls. Eur. Arch. Otorhinolaryngol. 2020, 277, 2939–2940. [Google Scholar] [CrossRef] [PubMed]
- Lammek, K.; Tretiakow, D.; Skorek, A. The first bite syndrome after parotidectomy. Oral Oncol. 2021, 113, 105028. [Google Scholar] [CrossRef] [PubMed]
- Tretiakow, D.; Skorek, A. Regarding to functional outcomes after extracapsular dissection with partial facial nerve dissection for small and large parotid neoplasms. Am. J. Otolaryngol. 2021, 42, 102965. [Google Scholar] [CrossRef]
- Lombardi, D.; McGurk, M.; Vander Poorten, V.; Guzzo, M.; Accorona, R.; Rampinelli, V.; Nicolai, P. Surgical treatment of salivary malignant tumors. Oral Oncol. 2017, 65, 102–113. [Google Scholar] [CrossRef] [PubMed]
- Stodulski, D.; Mikaszewski, B.; Stankiewicz, C. Signs and symptoms of parotid gland carcinoma and their prognostic value. Int. J. Oral Maxillofac. Surg. 2012, 41, 801–806. [Google Scholar] [CrossRef]
- Fonseca, F.P.; Sena Filho, M.; Altemani, A.; Speight, P.M.; Vargas, P.A. Molecular signature of salivary gland tumors: potential use as diagnostic and prognostic marker. J. Oral Pathol. Med. 2016, 45, 101–110. [Google Scholar] [CrossRef]
- Stenman, G. Fusion oncogenes in salivary gland tumors: molecular and clinical consequences. Head Neck Pathol. 2013, 7 (Suppl. S1), S12–9. [Google Scholar] [CrossRef] [PubMed]
- Lima-Souza, R.A. de; Scarini, J.F.; Lavareze, L.; Emerick, C.; Dos Santos, E.S.; Paes Leme, AF; Egal, ESA; Altemani, A. ; Mariano, F.V. Protein markers of primary salivary gland tumors: A systematic review of proteomic profiling studies. Arch. Oral Biol. 2022, 136, 105373. [Google Scholar] [CrossRef]
- Seccia, V.; Navari, E.; Donadio, E.; Boldrini, C.; Ciregia, F.; Ronci, M.; Aceto, A.; Dallan, I.; Lucacchini, A.; Casani, A.P.; et al. Proteomic Investigation of Malignant Major Salivary Gland Tumors. Head Neck Pathol. 2020, 14, 362–373. [Google Scholar] [CrossRef] [PubMed]
- Donadio, E.; Giusti, L.; Seccia, V.; Ciregia, F.; Da Valle, Y.; Dallan, I.; Ventroni, T.; Giannaccini, G.; Sellari-Franceschini, S.; Lucacchini, A. New insight into benign tumors of major salivary glands by proteomic approach. PLoS One 2013, 8, e71874. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, C.M.; Jesus, S.F. de; Souza, M.G. de; Queiroz, L.D.R.P.; Santos, E.M.; Dos Santos, E.P.; Oliveira, L.P.; Cordeiro Santos, C.K.; Santos, S.H.S.; Paula, A.M.B. de; et al. High levels of ANXA2 are characteristic of malignant salivary gland tumors. J. Oral Pathol. Med. 2019, 48, 929–934. [Google Scholar] [CrossRef]
- Mutlu, A.; Ozturk, M.; Akpinar, G.; Kasap, M.; Kanli, A. Proteomics analysis of pleomorphic adenoma of the human parotid gland. Eur. Arch. Otorhinolaryngol. 2017, 274, 3183–3195. [Google Scholar] [CrossRef] [PubMed]
- Lyapina, I.; Ivanov, V.; Fesenko, I. Peptidome: Chaos or Inevitability. Int. J. Mol. Sci. 2021, 22. [Google Scholar] [CrossRef] [PubMed]
- Hellinger, R.; Sigurdsson, A.; Wu, W.; Romanova, E. V.; Li, L.; Sweedler, J. V.; Süssmuth, R. D.; Gruber, C. W. Peptidomics. Nat Rev Methods Primers 2023, 3, 25. [Google Scholar] [CrossRef]
- Latosinska, A.; Frantzi, M.; Siwy, J. Peptides as "better biomarkers"? Value, challenges, and potential solutions to facilitate implementation. Mass Spec Rev. 2023, 1–42. [Google Scholar] [CrossRef]
- Villanueva, J.; Shaffer, D.R.; Philip, J.; Chaparro, C.A.; Erdjument-Bromage, H.; Olshen, A.B.; Fleisher, M.; Lilja, H.; Brogi, E.; Boyd, J.; et al. Differential exoprotease activities confer tumor-specific serum peptidome patterns. J. Clin. Invest. 2006, 116, 271–284. [Google Scholar] [CrossRef]
- Tanuwidjaya, E.; Schittenhelm, R.B.; Faridi, P. Soluble HLA peptidome: A new resource for cancer biomarkers. Front. Oncol. 2022, 12, 1069635. [Google Scholar] [CrossRef]
- Han, T.; Cong, H.; Yu, B.; Shen, Y. Application of peptide biomarkers in life analysis based on liquid chromatography–mass spectrometry technology. BioFactors. 2022, 48, 725–743. [Google Scholar] [CrossRef] [PubMed]
- Neves, L.X.; Granato, D.C.; Busso-Lopes, A.F.; Carnielli, C.M.; Patroni, F.M.d.S.; Rossi, T. de; Oliveira, A.K.; Ribeiro, A.C.P.; Brandão, T.B.; Rodrigues, A.N.; et al. Peptidomics-Driven Strategy Reveals Peptides and Predicted Proteases Associated With Oral Cancer Prognosis. Mol. Cell. Proteomics 2021, 20, 100004. [Google Scholar] [CrossRef] [PubMed]
- Fonseca, F.P.; Macedo, C.C.S.; Dos Santos Costa, S.F.; Leme, AFP; Rodrigues, R. R.; Pontes, H.A.R.; Altemani, A.; van Heerden, WFP; Martins, M.D.; Almeida, O.P. de; et al. Mass spectrometry-based proteome profile may be useful to differentiate adenoid cystic carcinoma from polymorphous adenocarcinoma of salivary glands. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. 2019, 128, 639–650. [Google Scholar] [CrossRef] [PubMed]
- Pereira, D.L.; Bingle, L.; Bingle, C.D.; Lopes, M.A.; Vargas, P.A. Expression of BPIF/PLUNC Protein Family and MECT1-MAML2 Fusion in Salivary Mucoepidermoid Carcinomas: a Preliminary Study. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. 2018, 126, e158. [Google Scholar] [CrossRef]
- González-Arriagada, W.A.; Ramos, L.M.A.; Silva, A.A.; Vargas, P.A.; Della Coletta, R.; Bingle, L.; Lopes, M.A. Salivary BPIFA1 (SPLUNC1) and BPIFA2 (SPLUNC2 A) are modified by head and neck cancer radiotherapy. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. 2015, 119, 48–58. [Google Scholar] [CrossRef] [PubMed]
- Kwon, C.H.; Park, H.J.; Choi, J.H.; Lee, J.R.; Kim, H.K.; Jo, H.-J.; Kim, H.S.; Oh, N.; Song, G. am; Park, D.Y. Snail and serpinA1 promote tumor progression and predict prognosis in colorectal cancer. Oncotarget 2015, 6, 20312–20326. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Hu, L.G. Serpin peptidase inhibitor clade A member 1-overexpression in gastric cancer promotes tumor progression in vitro and is associated with poor prognosis. Oncol. Lett. 2020, 20, 278. [Google Scholar] [CrossRef] [PubMed]
- El-Akawi, Z.J.; Al-Hindawi, F.K.; Bashir, N.A. Alpha-1 antitrypsin (alpha1-AT) plasma levels in lung, prostate and breast cancer patients. Neuro Endocrinol. Lett. 2008, 29, 482–484. [Google Scholar]
- Salem, H.; Ellakwa, D.E.-S.; Fouad, H.; Hamid, M.A. APOA1 AND APOA2 proteins as prognostic markers for early detection of urinary bladder cancer. Gene Reports 2019, 16, 100463. [Google Scholar] [CrossRef]
- Honda, K.; Srivastava, S. Potential usefulness of apolipoprotein A2 isoforms for screening and risk stratification of pancreatic cancer. Biomark. Med. 2016, 10, 1197–1207. [Google Scholar] [CrossRef]
- Feldman, A.S.; Banyard, J.; Wu, C.-L.; McDougal, W.S. ; Zetter, BR Cystatin B as a tissue and urinary biomarker of bladder cancer recurrence and disease progression. Clin. Cancer Res. 2009, 15, 1024–1031. [Google Scholar] [CrossRef]
- Wang, X.; Gui, L.; Zhang, Y.; Zhang, J.; Shi, J.; Xu, G. Cystatin B is a progression marker of human epithelial ovarian tumors mediated by the TGF-β signaling pathway. Int. J. Oncol. 2014, 44, 1099–1106. [Google Scholar] [CrossRef] [PubMed]
- Cui, J.; Li, G.; Yin, J.; Li, L.; Tan, Y.; Wei, H.; Liu, B.; Deng, L.; Tang, J.; Chen, Y.; et al. GSTP1 and cancer: Expression, methylation, polymorphisms and signaling (Review). Int. J. Oncol. 2020, 56, 867–878. [Google Scholar] [CrossRef]
- Altenberg, B.; Greulich, K.O. Genes of glycolysis are ubiquitously overexpressed in 24 cancer classes. Genomics 2004, 84, 1014–1020. [Google Scholar] [CrossRef]
- Lee, H.; Kim, K.; Woo, J.; Park, J.; Kim, H.; Lee, K.E.; Kim, H.; Kim, Y.; Moon, K.C.; Kim, J.Y.; et al. Quantitative Proteomic Analysis Identifies AHNAK (Neuroblast Differentiation-associated Protein AHNAK) as a Novel Candidate Biomarker for Bladder Urothelial Carcinoma Diagnosis by Liquid-based Cytology. Mol. Cell. Proteomics 2018, 17, 1788–1802. [Google Scholar] [CrossRef] [PubMed]
- Sudo, H.; Tsuji, A.B.; Sugyo, A.; Abe, M.; Hino, O.; Saga, T. AHNAK is highly expressed and plays a key role in cell migration and invasion in mesothelioma. Int. J. Oncol. 2014, 44, 530–538. [Google Scholar] [CrossRef] [PubMed]
- Gebhardt, C.; Németh, J.; Angel, P.; Hess, J. S100A8 and S100A9 in inflammation and cancer. Biochem. Pharmacol. 2006, 72, 1622–1631. [Google Scholar] [CrossRef]
- Nayak, S.; Bhatt, M.L.B.; Goel, MM; Gupta, S. ; Mahdi, A.A.; Mishra, A.; Mehrotra, D. Tissue and serum expression of TGM-3 may be prognostic marker in patients of oral squamous cell carcinoma undergoing chemo-radiotherapy. PLoS One 2018, 13, e0199665. [Google Scholar] [CrossRef]
- Uemura, N.; Nakanishi, Y.; Kato, H.; Saito, S.; Nagino, M.; Hirohashi, S.; Kondo, T. Transglutaminase 3 as a prognostic biomarker in esophageal cancer revealed by proteomics. Int. J. Cancer 2009, 124, 2106–2115. [Google Scholar] [CrossRef]
- Zhang, Y.; Feurino, L.W.; Zhai, Q.; Wang, H.; Fisher, W.E.; Chen, C.; Yao, Q.; Li, M. Thymosin Beta 4 is overexpressed in human pancreatic cancer cells and stimulates proinflammatory cytokine secretion and JNK activation. Cancer Biol. Ther. 2008, 7, 419–423. [Google Scholar] [CrossRef]
- Gemoll, T.; Strohkamp, S.; Schillo, K.; Thorns, C.; Habermann, J.K. MALDI-imaging reveals thymosin beta-4 as an independent prognostic marker for colorectal cancer. Oncotarget 2015, 6, 43869–43880. [Google Scholar] [CrossRef] [PubMed]
- Tamesa, M.S.; Kuramitsu, Y.; Fujimoto, M.; Maeda, N.; Nagashima, Y.; Tanaka, T.; Yamamoto, S.; Oka, M.; Nakamura, K. Detection of autoantibodies against cyclophilin A and triosephosphate isomerase in sera from breast cancer patients by proteomic analysis. Electrophoresis 2009, 30, 2168–2181. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Huang, Z.; Tian, Y.; Lin, B.; He, R.; Wang, H.; Ouyang, P.; Chen, H.; Wu, L. Clinical significance and prognostic value of Triosephosphate isomerase expression in gastric cancer. Medicine (Baltimore) 2017, 96, e6865. [Google Scholar] [CrossRef] [PubMed]
- Deutsch, E.W. The PeptideAtlas Project. In: Hubbard, S.; Jones, A. (eds) Proteome Bioinformatics. Methods in Molecular Biology™, 2010, vol 604, Humana Press. [CrossRef]
- Fagerberg, L.; Hallström, B.M.; Oksvold, P.; Kampf, C.; Djureinovic, D.; Odeberg, J.; Habuka, M.; Tahmasebpoor, S.; Danielsson, A.; Edlund, K.; Asplund, A.; Sjöstedt, E.; Lundberg, E.; Szigyarto, C. A.; Skogs, M.; Takanen, J. O.; Berling, H.; Tegel, H.; Mulder, J.; Nilsson, P.; Schwenk, J. M.; Lindskog, C.; Danielsson, F.; Mardinoglu, A.; Sivertsson, A.; von Feilitzen, K.; Forsberg, M.; Zwahlen, M.; Olsson, I.; Navani, S.; Huss, M.; Nielsen, J.; Ponten, F.; Uhlén, M. Analysis of the human tissue-specific expression by genome-wide integration of transcriptomics and antibody-based proteomics. Mol Cell Proteomics. 2014, 13, 397–406. [Google Scholar] [CrossRef] [PubMed]
- Storkus, W.J. , Zeh H.J., Salter R.D., Lotze M.T. Identification of T-Cell epitopes: Rapid isolation of class i-presented peptides from viable cells by mild acid elution. J. Immunother. 1993, 14, 94–103. [Google Scholar] [CrossRef]
- Purcell, A.W.; Ramarathinam, S.H.; Ternette, N. Mass spectrometry based identification of MHC-bound peptides for immunopeptidomics. Nature Protocols, 2019; 14, 1687–170. [Google Scholar] [CrossRef]
- Brunet, M.A.; Lucier, J.-F.; Levesque, M.; Leblanc, S.; Jacques, J.-F.; Al-Saedi, H.R.H.; Guilloy, N.; Grenier, F.; Avino, M.; Fournier, I.; et al. OpenProt 2021: deeper functional annotation of the coding potential of eukaryotic genomes. Nucleic Acids Res. 2021, 49, D380–D388. [Google Scholar] [CrossRef] [PubMed]
- The UniProt Consortium. UniProt: the Universal Protein Knowledgebase in 2023, Nucleic Acids Research, Volume 51, Issue D1, 6 January 2023, Pages D523–D531. [CrossRef]
- Newfound Coding Potential of Transcripts Unveils Missing Members of Human Protein Communities. Leblanc S, Brunet MA, Jacques JF, Lekehal AM, Duclos A, Tremblay A, Bruggeman-Gascon A, Samandi S, Brunelle M, Cohen AA, Scott MS, Roucou X. Genomics Proteomics Bioinformatics. 2022 Sep 29:S1672-0229(22)00124-3. [CrossRef]
- Wang, ET; Sandberg, R; Luo, S; Khrebtukova, I; Zhang, L; Mayr, C; Kingsmore, SF;, Schroth GP; Burge CB Alternative Isoform Regulation in Human Tissue Transcriptomes Nature. 2008 November 27; 456(7221): 470–476. [CrossRef]
- The Landscape of Isoform Switches in Human Cancers Kristoffer Vitting-Seerup and Albin Sandelin. Mol Cancer Res; 15(9) September 2017.
- Rappsilber, J.; Ishihama, Y.; Mann, M. Stop and go extraction tips for matrix-assisted laser desorption/ionization, nanoelectrospray, and LC/MS sample pretreatment in proteomics. Anal. Chem. 2003, 75, 663–670. [Google Scholar] [CrossRef] [PubMed]
- Nobilis, M.; Pour, M.; Kuneš, J.; Kopecký, J.; Květina, J.; Svoboda, Z.; Sládková, K.; Vortel, J. High-performance liquid chromatographic determination of ursodeoxycholic acid after solid phase extraction of blood serum and detection-oriented derivatization. Journal of Pharmaceutical and Biomedical Analysis 2001, 24, 937–946. [Google Scholar] [CrossRef] [PubMed]
- Lewandowska, A.E.; Macur, K.; Czaplewska, P.; Liss, J.; Łukaszuk, K.; Ołdziej, S. Qualitative and Quantitative Analysis of Proteome and Peptidome of Human Follicular Fluid Using Multiple Samples from Single Donor with LC-MS and SWATH Methodology. J. Proteome Res. 2017, 16, 3053–3067. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Xin, L.; Shan, B.; Chen, W.; Xie, M.; Yuen, D.; Zhang, W.; Zhang, Z.; Lajoie, G.A.; Ma, B. PEAKS DB: de novo sequencing assisted database search for sensitive and accurate peptide identification. Mol. Cell. Proteomics 2012, 11, M111–010587. [Google Scholar] [CrossRef]
- Perez-Riverol Y, Bai J, Bandla C, García-Seisdedos, D. ,.; Hewapathirana, S.,.; Kamatchinathan S, Kundu DJ, Prakash A, Frericks-Zipper A, Eisenacher M, Walzer M, Wang S, Brazma A, Vizcaíno, J.A. The PRIDE database resources in 2022: a hub for mass spectrometry-based proteomics evidences. Nucleic Acids Res. 2022, 50, D543–D552. [Google Scholar] [CrossRef] [PubMed]
- van Rossum, G.; Drake, F.L. Python 3 Reference Manual; CreateSpace: Scotts Valley, CA, 2009; ISBN 1441412697. [Google Scholar]
- Harris, C.R.; Millman, K.J.; van der Walt, S.J.; Gommers, R.; Virtanen, P.; Cournapeau, D.; Wieser, E.; Taylor, J.; Berg, S.; Smith, N.J.; et al. Array programming with NumPy. Nature 2020, 585, 357–362. [Google Scholar] [CrossRef] [PubMed]
- McKinney, W. Data Structures for Statistical Computing in Python. In Proceedings of the 9th Python in Science Conference. Python in Science Conference, Austin, Texas, 2010, 28 June–3 July 2010; SciPy; pp. 56–61. [Google Scholar]
Figure 1.
Venn diagrams of showing number of identified peptides: A - peptides identified in extracts from tumor tissue and healthy salivary glands samples, B - peptides identified in extracts from patients’ and healthy subjects’ saliva. Venn diagrams show combined results from analyzing all samples.
Figure 1.
Venn diagrams of showing number of identified peptides: A - peptides identified in extracts from tumor tissue and healthy salivary glands samples, B - peptides identified in extracts from patients’ and healthy subjects’ saliva. Venn diagrams show combined results from analyzing all samples.
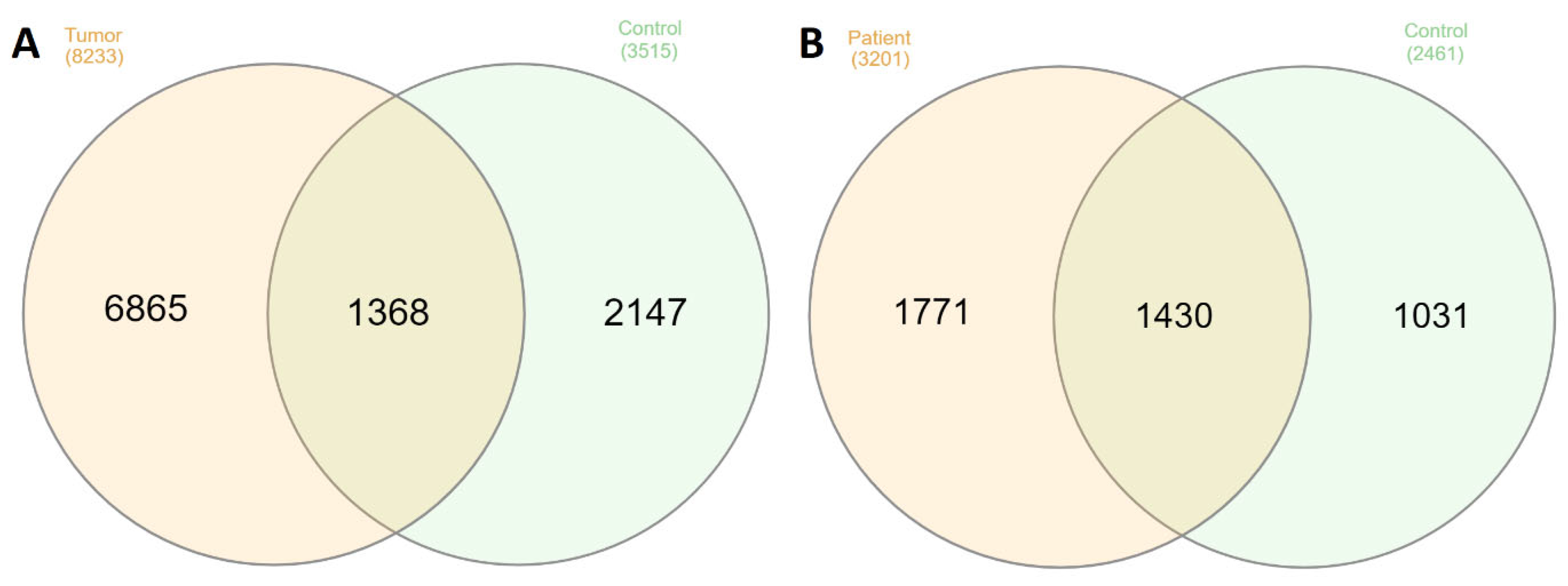
Figure 2.
An example of the classification of identified peptides. * - unique peptide , ** - stand-alone peptide.
Figure 2.
An example of the classification of identified peptides. * - unique peptide , ** - stand-alone peptide.
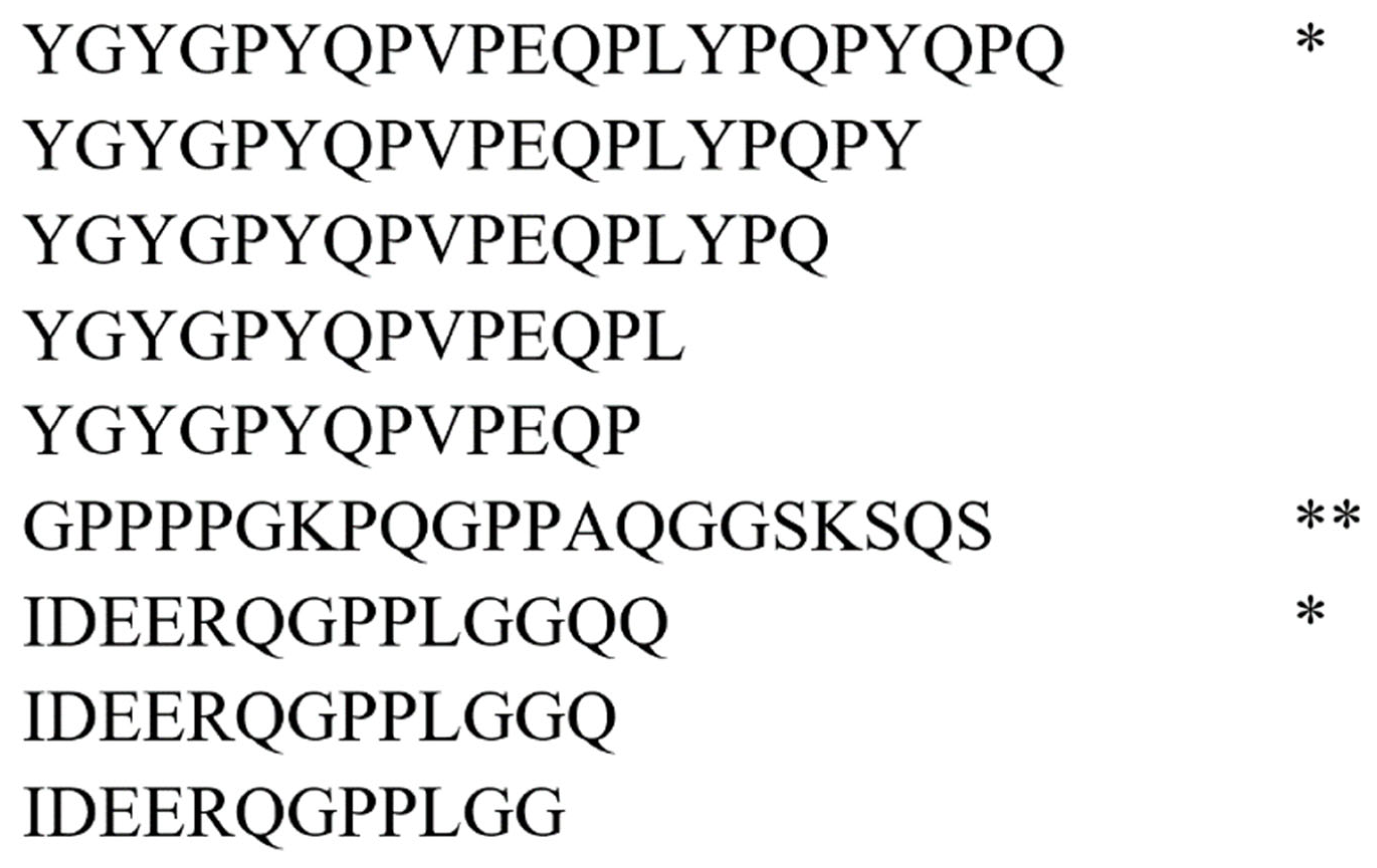
Figure 3.
Coverage of the amino acid sequence of statherin (UniProt ID P02808) by peptides identified in saliva (left) and tissue extracts (right). The green line schematically shows the amino acid sequence of statherin, the x-axis shows the length of the protein. In both panels, red and blue lines represent peptides identified as fragments of the parent protein. Blue colors represent peptides identified in saliva from healthy individuals and peptides extracted from healthy tissue samples. Peptides identified in patient saliva or extracted from tumor tissue are shown in red.
Figure 3.
Coverage of the amino acid sequence of statherin (UniProt ID P02808) by peptides identified in saliva (left) and tissue extracts (right). The green line schematically shows the amino acid sequence of statherin, the x-axis shows the length of the protein. In both panels, red and blue lines represent peptides identified as fragments of the parent protein. Blue colors represent peptides identified in saliva from healthy individuals and peptides extracted from healthy tissue samples. Peptides identified in patient saliva or extracted from tumor tissue are shown in red.
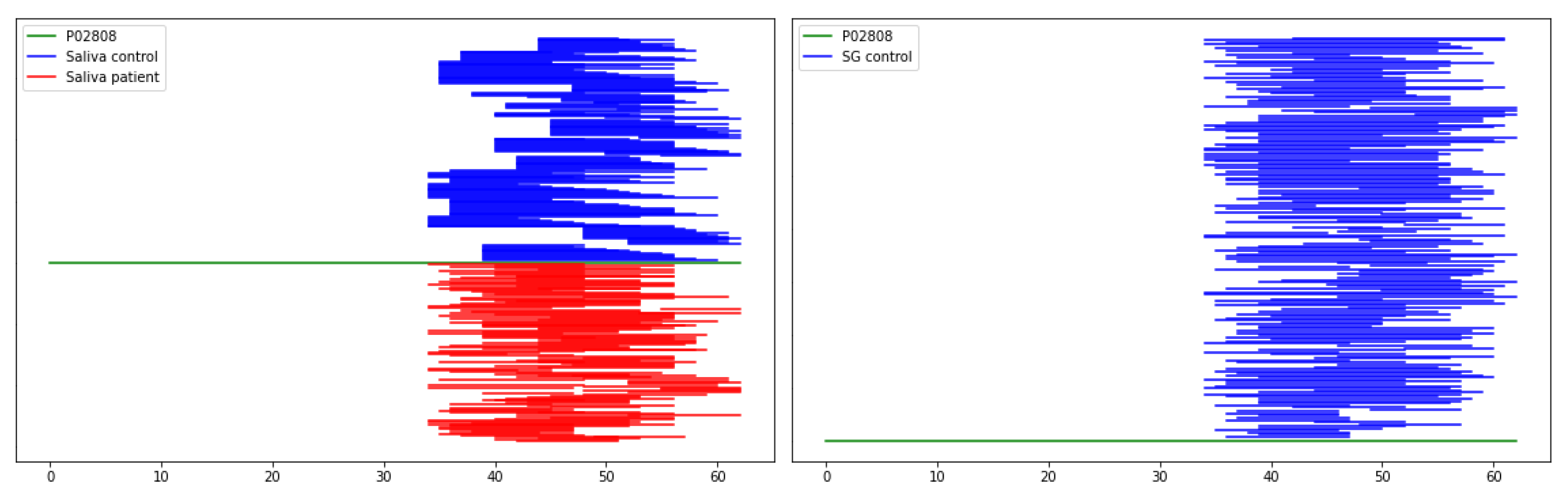
Figure 4.
Workflow of peptide isolation from salivary gland tissue.
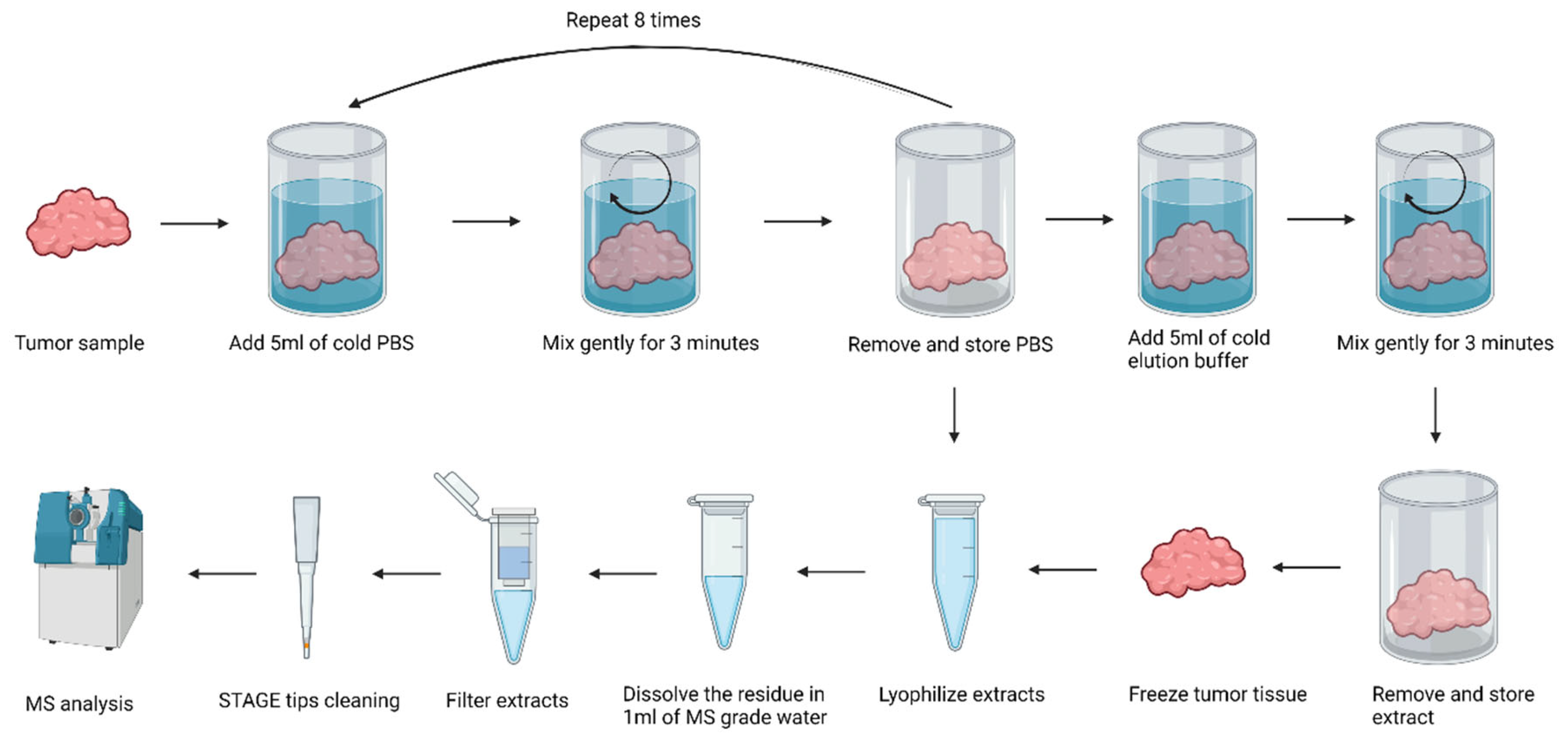

Table 1.
Number of samples analyzed in this work with information about sample handling (fresh or frozen) or peptide methods extraction used (PBS or TFA). For details see Method section.
Table 1.
Number of samples analyzed in this work with information about sample handling (fresh or frozen) or peptide methods extraction used (PBS or TFA). For details see Method section.
| Salivary glands tissue samples | |||||
| Fresh | Frozen | Total | |||
| PBS | TFA | PBS | TFA | ||
| Tumor | 5 | 4 | 5 | 4 | 18 |
| Control | 5 | 4 | 5 | 4 | 18 |
| Total | 10 | 8 | 10 | 8 | 36 |
| Saliva samples | |||||
| Patient | Control | Total | |||
| 11 | 8 | 19 | |||
Table 2.
Number of peptides identified in all analyses with the distinction of unique and stand-alone peptides (see text for details).
Table 2.
Number of peptides identified in all analyses with the distinction of unique and stand-alone peptides (see text for details).
| Data sets | Peptides | Unique peptides | Stand-alone peptides |
|---|---|---|---|
| SG tumor | 8233 | 3916 | 1733 |
| SG control | 3515 | 1586 | 589 |
| SG fresh | 6752 | 3205 | 1383 |
| SG frozen | 6913 | 3212 | 1496 |
| SG PBS | 7771 | 3649 | 1667 |
| SG TFA | 3540 | 1770 | 702 |
| SG All | 10380 | 4869 | 2077 |
| Saliva patient | 3201 | 1166 | 325 |
| Saliva control | 2461 | 836 | 258 |
| Saliva all | 4189 | 1490 | 420 |
SG – salivary glands; SG tumor – peptides identified in extracts from SG tumor tissue; SG control – peptides identified in extracts from SG healthy tissue; SG fresh – peptides identified in extracts from fresh samples of SG tissue (tumor and control); SG frozen – peptides identified in extracts from frozen samples of SG tissue (tumor and control); SG PBS – peptides identified in extracts from samples of SG tissue (tumor and control, fresh and frozen) using PBS; SG TFA - peptides identified in extracts from samples of SG tissue (tumor and control, fresh and frozen) using water with TFA; SG all – all peptides isolated form SG all samples. Saliva patient – peptides identified in patient saliva; Saliva control – peptides identified in control group saliva; Saliva all - peptides identified in all saliva samples (patient and control).
Table 3.
Comparison of the numbers of peptides unique to a given group of samples.
| Datasets | Overlapping sequences | Common peptides |
|---|---|---|
| Saliva patient - SG tumor | 377 | 86 |
| Saliva patient only - SG tumor only | 109 | 16 |
| Saliva patient - SG control | 960 | 182 |
| Saliva patient only - SG control only | 362 | 42 |
| Saliva control - SG tumor | 425 | 121 |
| Saliva control only - SG tumor only | 154 | 39 |
| Saliva control - SG control | 950 | 209 |
| Saliva control only - SG control only | 351 | 58 |
See the description of data sets in Table 2. Word “only” added to data sets means that the given data set contains peptides unique for a given group. For example, the “Saliva patient only” data set was built by removing from the data set "Saliva patient" (see Table 2) all peptides present in the data set “Saliva control” (see Table 2).
Table 4.
A list of proteins which are the source of peptides observed only in patient saliva samples and extracts from tumor tissue samples (see Table 3).
Table 4.
A list of proteins which are the source of peptides observed only in patient saliva samples and extracts from tumor tissue samples (see Table 3).
| Protein Accession | Protein name | Gene Names | No. peptides |
|---|---|---|---|
| P63261, P60709 | Actin, cytoplasmic | ACTB, ACTG | 9 |
| P01009 | Alpha-1-antitrypsin | SERPINA1 | 1 |
| P04083 | Annexin A1 | ANXA1 | 3 |
| V9GYC1 | Apolipoprotein A-II | APOA2 | 1 |
| P04280, P02812, Q04118, P10163 | Basic salivary proline-rich proteins | PRB1/4 | 32 |
| Q96DR5 | BPI fold-containing family A member 2 (SPLUNC2, Parotid secretory protein) | BPIFA2 | 1 |
| P04080 | Cystatin-B | CSTB | 1 |
| P02671 | Fibrinogen alpha chain | FGA | 2 |
| P02675 | Fibrinogen beta chain | FGB | 2 |
| P09211 | Glutathione S-transferase P | GSTP1 | 3 |
| P04406 | Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) | GAPDH | 4 |
| P04792 | Heat shock protein beta-1 | HSPB1 | 2 |
| P69905, P68871 | Haemoglobin | HBA1/HBA2, HBB | 5 |
| P16402, P10412, P16401, P20671 | Histones | H1-3, H1-4, H1-5, H2AC7 | 9 |
| P01857 | Immunoglobulin heavy constant gamma 1 | IGHG1 | 1 |
| P04264, P13647 | Keratin | KRT1 , KRT5 | 4 |
| Q09666 | Neuroblast differentiation-associated protein AHNAK (Desmoyokin) | AHNAK | 2* |
| P05109 | Protein S100-A8 (Calgranulin-A) | S100A8 | 2 |
| P06702 | Protein S100-A9 (Calgranulin-B) | S100A9 | 2 |
| Q08188 | Protein-glutamine gamma-glutamyltransferase E | TGM3 | 1 |
| P02810 | Salivary acidic proline-rich phosphoprotein 1/2 | PRH1; PRH2 | 17 |
| P62328 | Thymosin beta-4 | TMSB4X | 2(1**) |
| P60174 | Triosephosphate isomerase | TPI1 | 2 |
| P08670 | Vimentin | VIM | 1 |
In red – are proteins previously linked with salivary gland tumors; in yellow – are proteins previously linked with other cancer types; * - are peptides of isoform origin (OpenProt), ** - are peptides of pseudogene origin (OpenProt).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



