
Submitted:
24 June 2024
Posted:
25 June 2024
You are already at the latest version
Abstract
Creatine has been linked to the development of kidney damage since 1998. However, uncertainties persist regarding the actual impact of creatine supplementation on individuals with kidney disease before supplementation or with pathological conditions that have an increased risk of developing kidney dysfunction. This study performed a comprehensive review of the literature and utilized bioinformatics tools to investigate the effects of creatine on renal tissue and function. Moreover, we identified 44 genes modulated explicitly in response to creatine exposure using the Comparative Toxicogenomics Database, including IGF1, SLC2A4, and various creatine kinase genes. The analysis revealed associations with metabolic processes such as amino acid metabolism, indicating a connection between creatine and tissue physiology. Gene expression analysis using the Genotype-Tissue Expression Portal highlighted tissue-specific expression patterns in kidney and pancreas tissues. Furthermore, the study explored creatine metabolism pathways and kinase regulation through biological databases and Gene Expression Omnibus data, shedding light on the potential regulatory roles of creatine in cellular processes during kidney diseases. Understanding the impact of the balance between the endogenous creatine synthesis and the creatine uptake, particularly the genes GATM, GAMT, SLC6A8 and IGF1, insights into molecular mechanisms underlying creatine's physiological effects on renal tissues. Overall, this work provides new insights into the molecular interactions and pathways influenced by creatine in renal function.
Keywords:
creatine monohydrate
; kidney
; renal injury
Introduction
Creatine (α-methylguanidinoacetic acid) (Figure 1) is an amino acid endogenously synthesized in the kidneys and liver of animals from reactions involving arginine, glycine, and methionine. It is also obtained exogenously from consuming meat or as a dietary supplement [1].
Its primary role, especially within muscle tissue, is to participate in the resynthesis of ATP (adenosine triphosphate) in the immediate bioenergetic system. In the cytosol, creatine phosphate loses its phosphate group through the enzyme creatine kinase (CK), transferring this group to ADP (adenosine diphosphate) to produce ATP, which fuels the cell's energy needs. Within mitochondria, creatine can also be recycled by mitochondrial creatine kinase (MtCK), generating ADP and phosphocreatine, thereby completing the Cr/ATP/CK system cycle [2].
Originally discovered by Michel Chevreul in 1832, it took over a century and a half for the first evidence to suggest that some of the consumed creatine could be retained in the body [3]. Subsequently, Harris et al. conclusively demonstrated that creatine retention primarily occurs within skeletal muscle, raising significant interest in sports [4,5]. Indeed, creatine is currently regarded as one of the most effective ergogenic supplements in sports practice [6], given its potential to enhance muscle performance, particularly during short-duration, high-intensity resistance exercises [7]. Due to its anabolic potential, creatine supplementation may benefit a range of acute and chronic clinical conditions [8]. It is currently associated with positive effects on heart disease, muscle dystrophies [5], and brain health, such as cognitive processing, brain function, memory, and trauma recovery [9,10,11].
The continuous, low-grade, and non-enzymatic degradation of creatine produces creatinine as the resultant end product [12]. After filtration through the renal glomeruli, creatinine and plasma creatine are excreted. To counterbalance the daily depletion, the classic understanding is that our body synthesizes creatine through two reactions: the first occurs in the kidneys, where the enzyme arginine amidinotransferase (GATM/AGAT) converts arginine and glycine into guanidinoacetate, whereas the second reaction involves guanidinoacetate N-methyltransferase (GAMT), which catalyzes the conversion of guanidinoacetate to creatine through methylation in the liver [13]. However, this notion had been intensely debated since several gene expression studies showed the expression of both GATM and GAMT in other tissues [14] and significant contributions from other tissues may be common [15].
On the other hand, it is well-known that serum creatinine concentration is a crude index of renal function [12]. Nevertheless, exogenous creatine intake markedly increases urine creatinine concentration and excretion, potentially leading to minor changes in markers of renal and hepatic function [16], which led to the understanding that creatine supplementation could pose risks to healthy individuals, despite the lack of concrete evidence. The first concern about adverse effects from creatine supplementation emerged after two works in the late 1990s [17,18]. For instance, Pritchard and Kalra reported a case where a patient with previously controlled renal lesions experienced changes in kidney function after creatine supplementation. Notably, these alterations were ameliorated upon cessation of the supplementation regimen [18]. Since then, concerns have arisen about creatine's potential impact on renal function, despite increasing scientific interest.
The association between creatine and renal injuries primarily originates from its tendency to elevate serum creatinine levels [5]. However, the interpretation of serum creatinine levels in the context of creatine supplementation must be cautious, as creatine can lead to false-positive results for renal injury due to non-enzymatic hydrolysis generating creatinine.
To date, a significant misconception exists regarding whether creatine supplementation is associated with kidney damage or renal dysfunction in humans. Therefore, this work aims to summarize the current evidence concerning the potential association between creatine supplementation and nephropathies, as well as provides new insights into the molecular interactions, pathways modulated by creatine-related gene expression in renal issues through a systems biology approach. Through this in silico exploration, we aimed to uncover potential therapeutic strategies and enhance our comprehension of how creatine impacts renal function in various physiological states.
Methods
Narrative Review
We performed a comprehensive search for articles using PubMed, Science Direct and Web of Science with no data or language restrictions. We searched Abstracts and full texts for the terms “creatine” OR “creatine supplementation” AND/OR “kidney disease”, “kidney injury” and “nephropathy”. Inclusion criteria were: (i) case reports and clinical trials with creatine supplementation conducted in humans and murine models and (ii) the presence of nephropathies and alterations in the kidney’s function and morphology. The exclusion criteria were studies with creatine administration that did not assess renal morphology or function in murine models and humans. The search retrieved 108 articles, of which 23 were included in this review. The present study also considered relevant reviews on the subject matter.
Bioinformatics analyses
We initially used the term "creatine" in a search limited to the organism Homo sapiens, using the Stitch Database [19], which lists the known and predicted interactions between chemicals and proteins. Then, using the advanced settings, we restricted the search for integration of RNA-Seq expression data from kidney tissues using the Human Protein Atlas database. In the updated gene list, the proteins AKT1, AKT2, and AKT3 appeared as tissue-specific interactions. We also used the restriction from the Tissues Database, using the kidney as a term.
To widen our searches for the creatine-related genes, we also used the Comparative Toxicogenomics Database (CTD) [20], using "creatine" as a search term. After obtaining a list of genes from CTD, both gene lists, from CTD and Stitch databases, were submitted to a batch query in the CTD database to retrieve the genes-disease interactions. Then, we merged all the gene terms into one list. From this merged list, we performed a gene enrichment analysis (GEA), using enrichR [21] and enrich-Kg [22] to explore the biological functions and metabolic pathways of the identified genes, using Gene Ontology (GO) and the Kyoto Encyclopedia of Genes and Genomes (KEGG), respectively. We also performed GEA against the ReactomeDB. We retrieved the general expression panel of these genes for kidney tissues from the GTEx Portal (https://gtexportal.org). Additionally, we conducted GEA against the kinase regulation datasets ARCHS4_Kinases [23] and the Kinase Library 2023 [24], to assess the overall regulation of proteins encoded by the creatine-related genes in our list. The inclusion criteria were consistent across all analyses: selecting the first ten results based on a P-value cutoff of ≤ 0.05.
We also evaluated the expression of creatine-related genes using datasets and gene expression series from the Gene Expression Omnibus (GEO) database [25]. We used each gene symbol as the primary search term, applying the following criteria: (i) Organism, Homo sapiens; (ii) Differential expression, Up/down genes; (iii) Dataset keyword, Kidney and Renal tissues. The exclusion criteria were an undetected expression of the genes, experimental designs that did not compare disease/metabolic/chemical treatment conditions, and mutant cell lines. The gene expression series were analyzed using the R packages GEO2R (https://www.ncbi.nlm.nih.gov/geo/geo2r/), GEOquery [26], limma [27], and DESeq2 [28], employing a case-control approach. We considered significant differential expression (DE) expression values with Log2 Fold Change >0.5 and P-Value>0.05. Afterward, we generated specific plots for the evaluated genes using ggplot2 [29] and Python programming language modules numpy [30] and matplotlib [31]. The scripts used are available on GitHub.
Results and Discussion
Association of Creatine Supplementation and renal injury in Humans
As previously mentioned, the initial connection between renal injuries and creatine supplementation in sports was briefly described in the study by Pritchard and Kalra [18]. At that time, three Olympic wrestlers reportedly had died after using the amino acid, which garnered significant attention. However, as afterward found, only one of the three athletes was supplementing with creatine. The probable cause of death for the athletes was hyperthermia resulting from severe dehydration during competition preparation [32].
Pritchard and Kalra also discussed the correlation between creatine supplementation and renal injury in their case study [18]. However, establishing a cause-effect relationship was challenging due to several factors, including the low dosage of creatine supplementation during maintenance. Subsequent case studies and trials have highlighted that creatine supplementation was associated with certain adverse [18,33,34,35,36,37,38] or showed the absence of evident effects [16,39,40,41,42,43] on the renal function (Table 1). Notably, most cases already demonstrated a pre-existing condition that could affect renal function, inefficient diagnostic tools in the face of creatine supplementation, and occasional conflicting results, including a minor beneficial outcome in one instance (Table 1).
With the progress of prospective studies and improved methods for evaluating renal function, the idea that creatine exerts a negative impact on kidney balance began to be refuted. Since then, more methodologically robust studies have been conducted, with similar results [44]. The work of Poortmans and colleagues [16] was the first to report the effects of creatine supplementation in a group of five young males. The study observed that creatine did not impair renal function in healthy individuals, as indicated by creatinine and glomerular filtration rate analysis. However, it had significant limitations, including a small sample size (n = 5), a short duration of creatine supplementation (five days), and the absence of an active control group.
Subsequent studies aimed to expand the understanding of the effects of creatine supplementation on renal function. For instance, Gualano and colleagues [39] evaluated creatine administration in individuals undergoing moderate-intensity aerobic training (three times a week for three months). It demonstrated a decrease in Cystatin C levels over time, indicating an increased glomerular filtration rate. Serum creatinine decreased in the placebo group but remained unchanged in the creatine group. Another study examined the effects of two doses of creatine supplementation (0.03g/kg and 5g/day) on renal and hepatic function in 35 healthy adults during eight weeks of resistance exercise training [41]. Biochemical tests showed that creatinine levels increased slightly in both creatine groups but remained within normal ranges. Liver function tests showed no significant changes across all groups. These observations concluded that creatine supplementation at the given doses for eight weeks did not adversely affect renal or hepatic function in healthy individuals [41].
The impact of creatine supplementation was also assessed in type 2 diabetic patients (DMT2) [42]. This study included 25 participants randomly allocated to receive either creatine (5 g/day) or placebo for 12 weeks, and they underwent a protocol involving training sessions consisting of a 5-minute treadmill warm-up followed by 25 minutes of resistance training, 30 minutes of treadmill aerobic training, and 5 minutes of stretching exercises. No changes were found in creatinine clearance, serum and urinary urea, electrolytes, proteinuria, and albuminuria, indicating that creatine supplementation does not affect kidney function in DMT2 [42].
The most recent study examining the safety of creatine supplementation was conducted by Almeida and colleagues [43]. They collected a panel of blood and urine health indicators of eighteen resistance training practitioners before and after creatine supplementation (0.3 g/kg per day for seven days) to evaluate several biochemical parameters and renal function. Compared with the control group, no changes were observed in the supplemented group in the red blood cell parameters, white blood cell profile, blood lipid profile, metabolic and urine markers, or hepatic and renal function.
In general, these studies suggest that high-dose creatine (>3 g/day) may not be recommended for individuals with existing kidney issues or those at risk for kidney problems such as diabetes or hypertension. However, there are no restrictions for healthy individuals [44]. However, extensive cohort studies assessing long-term effects are imperative to understand better the impact of creatine supplementation on kidney health and function.

Table 1.
Studies with creatine supplementation in humans.
| Study | Object of study | Creatine intake protocol | Observations | |
|---|---|---|---|---|
| [16] | Five young, healthy males | 20 g of creatine monohydrate per day for 5 consecutive days | There was no detrimental effect on renal function; low sample size. | |
| [17] | Eight young men and one women | 1 to 80 g·day−1 for 10 months to 5 years | There was no detrimental effect on renal function; low sample size; long-term supplementation (years). | |
| [18] | Man, 25 years old, with a history of renal injury 8 years ago. | L: 15 g·day−1 for 1 week M: 2 g·day−1 for 7 weeks |
Previous renal injuries; low dosage in the maintenance phase (similar to amounts obtained dietetically and endogenously). | |
| [33] | 19 years old Man, soccer athlete. | 10 g·day−1 for 3 months | Renal insufficiency induced by creatine supplementation; serum creatinine was the sole marker of renal injury. | |
| [34] | Man, 20 years old. | 20 g·day−1 for 4 weeks | Severe interstitial nephritis four weeks after taking high dose creatine; insufficient past data on the patient. | |
| [35] | Male, 18 years old, renal insufficiency secondary to mitochondrial encephalopathy. | L: 20 g·day−1 for 12 days M: 5 g·day−1 for 28 months |
Previous renal injuries; creatinine levels were determined using a less accurate method; neuroprotective effect observed. | |
| [36] | Man, 22 years old, athlete | 200 g·day−1 continuously | Concomitant use of anabolic steroid; creatine overdose (200 g/day); renal injury markers were not reported. | |
| [37] | Man, 24 years old, athlete | 15 g·day−1 for 6 months | Acute renal failure; in addition to (low dose) creatine, other supplements were consumed for the purpose of bodybuilding. | |
| [38] | Man, 18 years old | L: 20 g·day−1 for 5 days M: 1 g·day−1 for 6 weeks |
Patient had acute renal failure while taking creatine; the study lacked a description of the participant's diet, whether he used anabolic steroids and a discussion of the possible contamination of the supplement | |
| [39] | Eighteen sedentary males performing resistance training | 10 g·day−1 for 3 months | Increased performance and body weight; no changes in metabolic and urine markers, or hepatic and renal function | |
| [40] | Man, 20 years old, single kidney | L: 20 g·day−1 for 5 days M: 5 g·day−1 for 30 days |
There were no changes in renal function; there was a slight improvement in glomerular filtration rate. | |
| [41] | 35 male individuals, 18 and 42 years, with a minimum of two consecutive months of training with resistance exercises | L: 20 g·day−1 for 7 days M: 0.03 g·kg-1day−1 for 7 weeks |
There were no changes in hepatic and renal function; relative small sample size and absence of long-term effects of creatine supplementation | |
| [42] | 25 men and women, > 45 years, with type 2 diabetes, physically inactive for at least 1 year, and with BMI ≥ 30 kg/m2 | 5 g·day−1 for 12 weeks | Creatine supplementation does not impair kidney function; study with short duration without long-term follow-up | |
| [43] | 18 healthy males | L: 0,3 g·kg-1day−1 for 7 days | Creatine supplementation did not alter metabolic and urine markers, hepatic, and renal function vs control group; short duration of supplementation | |
L: loading phase; M: maintenance phase; BMI: Body mass index.
Association of creatine supplementation and renal injury in experimental models
In addition to clinical studies, several studies with experimental models have been conducted to investigate the toxicity of creatine doses, as well as its effect on renal function and morphology (Table 2). However, the findings from these studies could have been more consistent, underscoring the necessity for tailored investigation into specific nephropathies in the context of creatine supplementation [45,46,47,48,49,50,51,52,53,54].
To the best of our knowledge, the study by Baracho et al. [45] exclusively examined the potential toxicity of creatine, assessing renal and hepatic function after 14 days of supplementation in physically inactive rats. Biochemical analysis revealed no significant changes in various plasma markers, such as creatinine, urea, and creatinine clearance, as well as in water and food intake and urinary output compared to the control group. The authors concluded that oral creatine supplementation at the tested doses did not induce renal or hepatic toxicity [45].
While creatine supplementation may not impact renal function in healthy individuals, the question remains regarding its effects on renal function in pre-existing pathological conditions [46]. Studies addressing this concern have predominantly utilized animal models, yielding varied outcomes in each model. Edmunds et al. [47] investigated the effects of creatine supplementation in Han:SPRD-cy rats, a genetically modified strain of rats commonly used in research on kidney diseases, such as polycystic kidney disease. After evaluating the kidney size and fluid content and determining cyst scores, as well as assessing renal function by measuring serum urea and creatinine concentrations and creatinine clearance, the authors concluded that creatine administration (2.0 g/kg for one week, followed by 0.4 g/kg for the next five weeks) accelerated the progression of renal disease in these animals [47].
Taes et al. [48] did not observe negative impacts of creatine supplementation on renal function in an animal model with pre-existing renal insufficiency. Moreover, Genc et al. [49] found that free creatine therapy did not protect rat kidneys from cisplatin-induced damage, suggesting no clear benefit of free creatine in preventing or reducing cisplatin-induced kidney damage.
Another research group investigated the long-term effects of oral creatine supplementation on renal function and body composition. In this experimental model involving creatine supplementation and exercise, the authors found that the group receiving creatine supplementation alone experienced a significant reduction in both glomerular filtration rate and renal plasma flow [50].
Souza et al. [51] evaluated the effects of short-term (1 week) and long-term (4-8 weeks) high-dose creatine supplementation on kidney and liver structure and function in sedentary and exercised Wistar rats. While results showed no differences between sedentary and exercised groups, the long-term creatine group exhibited higher creatinine and urea levels, along with increased levels of liver enzymes at four and eight weeks and structural alterations indicating renal and hepatic damage. Their findings suggested that prolonged creatine supplementation had a detrimental effect on kidney and liver function in sedentary rats but not in exercised rats [51].
Furthermore, another study indicated that creatine supplementation in exercised young rats led to increased levels of hepatic biomarkers (aspartate transaminase and gamma-glutamyltranspeptidase) and renal biomarkers (urea and creatinine), suggesting potential metabolic or functional changes in the liver and kidneys [52]. However, a study by Ramos Fernandes et al. [53] did not identify significant morphological damage to the kidneys or liver in older animals receiving creatine supplementation.
Recently, a study conducted by our research group investigated the effects of creatine supplementation in streptozotocin-induced diabetic rats [54]. We observed that although creatine administration attenuated some biochemical and morphological parameters of pancreatic and renal tissues in diabetic animals, normoglycemic animals supplemented with creatine were diagnosed with pancreatitis and presented renal tubular necrosis. These findings suggest that despite the absence of clinical symptoms and unaltered biochemical parameters, creatine supplementation as adjuvant therapy for DM requires careful evaluation due to potential adverse effects on tissue morphology [54]. These results contrast with the study by Gualano and colleagues in patients with DMT2 [42], where no changes in renal function were observed after creatine supplementation. However, it is essential to note that one of the exclusion criteria in their study was a low glomerular filtration rate, suggesting that patients were at risk of kidney injury due to diabetes but did not exhibit alterations in renal function.
Table 2.
Experimental studies with creatine supplementation.
| Study | Object of study | Creatine intake protocol | Conclusions | |
|---|---|---|---|---|
| [45] | 24 male Wistar rats; three dosages of creatine for 2 weeks | 0.5 g·kg-1day−1 1 g·kg-1day−1 2 g·kg-1day−1 |
Creatine supplementation did not result in renal and/or hepatic toxicity; short experimental period | |
| [47] | 23 male and 24 female Han:SPRD-cy rats (cystic kidney disease) | L: 2 g·kg-1day−1 for 1 week M: 0,4 g·kg-1day−1 for 35 days |
Creatine supplementation exacerbated pre-existing polycystic kidney disease; creatine was combined with glutamine | |
| [48] | 43 male Wistar rats; 23 with moderate renal failure | Creatine monohydrate (2% w/w) was added to this diet in the creatine-supplemented groups for 4 weeks | Creatine supplementation does not impair kidney function in animals with pre-existing renal failure or in control animals | |
| [49] | 60 male Sprague–Dawley rats; 40 with cisplatin-induced nephrotoxicity | 300 mg·kg-1day−1 for 30 days | Creatine administration was considered a promising adjuvant protective drug for reducing nephrotoxic effect of cisplatin | |
| [50] | 36 male Wistar rats | 2 g·kg-1day−1 for 10 weeks | The use of creatine alone induced an important and significant reduction of both renal plasma flow and glomerular filtration rate | |
| [51] | 72 male Wistar rats; swimming training | Short-term: 5 g·kg-1day−1 for 1 week Long-term: 1 g·kg-1day−1 for 4-8 weeks |
Long-term creatine supplementation impacted kidney and liver structure and function of sedentary but not of exercised rats | |
| [52] | 35 male young Wistar rats; swimming training | L: 5 g·kg-1day−1 for 1 week M: 1 g·kg-1day−1 for 40 days |
Increased hepatic and renal (urea and creatinine) biomarkers levels were observed in the groups supplemented with creatine | |
| [53] | 12 old Wistar rats | 0.3 mg/kg for 8 weeks. |
The supplemented group showed no significant organ damage, such as reductions in glomerular size or hepatic degeneration; however; low sample size | |
| [54] | 32 male Wistar rats, 16 with streptozotocin-induced type 1 DM | L: 13% (g·kg-1 of feed) for 5 days M: 2% (g·kg-1 of feed) for 35 days |
Creatine supplementation as adjuvant therapy for DM should be carefully evaluated |
DM: Diabetes mellitus; L: loading phase; M: maintenance phase; w/w: creatine weight / food weight.
Is there a therapeutic potential for creatine supplementation?
Research into its therapeutic potential in pathological conditions commenced following the demonstration of creatine supplementation safety in healthy individuals. Indeed, the extensive therapeutic potential of creatine supplementation has been explored in various conditions [55], including neuromuscular disorders, muscle injuries, neurodegenerative diseases, and even certain metabolic conditions (Table 3). As previously commented, the theoretical basis for this lies in the fact that phosphorylated creatine plays a role in the resynthesis of ATP, serving as an essential energy reserve for cellular work. In this context, some studies have shown promising effects of creatine supplementation in specific health conditions, such as Huntington's disease [56], cachexia in rats with cancer [57], sarcopenia, and osteoporosis [58].
Op 't Eijnde et al. [59] observed that creatine may increase the expression of GLUT-4 receptors, and subsequently, Gualano et al. [39] reported greater glucose uptake with creatine supplementation possible through this pathway. The increase in intramuscular glycogen concentration after creatine supplementation supports increased glucose uptake [60]. Due to these effects of creatine, supplementation could be beneficial in various pathological conditions. Thus, we utilized bioinformatics analyses to explore how the differential expression of creatine-related genes is disturbed in several kidney pathological conditions and hypothesize the potential outcomes of creatine supplementation.
Comparative Toxicogenomics Database
We utilized the Comparative Toxicogenomics Database to analyze the effects of creatine on renal and pancreatic tissues. We explored the data to identify genes and pathways affected by creatine exposure and potential associations with specific physiological and metabolic processes. We observed that creatine exposure is associated with changes in the expression of 44 specific genes. After filtering the repetitive entries and excluding the pseudogenes, we obtained the following gene list from CTD: CKB, CKM, CKMT1A, CKMT1B, CKMT2, SLC16A12, and SLC6A8.
Using these two lists, we filtered the inferred gene/diseases associations since the curated ones are enriched to genetic diseases (inborn metabolic errors), and filtered the results using the presence of the terms kidney and renal, obtaining 2596 and 818 rows, for Stitch and CTD gene lists, respectively. The higher number of entries in the Stitch list is probably associated with the initial filtering process for expression in kidney tissues. Figure 3 describes the DiseaseIDs (MESH) and counts from association with both gene lists. This analysis revealed that several diseases with similar names or related conditions, such as "Kidney Failure/ Chronic", "Kidney Diseases", and "Kidney Neoplasms", have identical counts, which may suggest potential clustering or co-occurrence of related diseases
We found at least five disease associations for each gene, demonstrating that these genes presented altered expression or direct associations to kidney-related disturbs, including chronic kidney diseases (CKD), cancer, and kidney failure. Detailed plots from each gene-disease association are available in Supplementary File 1.
Gene Expression Analysis and Enrichment of Creatine-Related Genes
We merged the two above gene lists into one list (Table 4). Then, we obtained a comprehensive expression profile of these genes across all tissues using the Multi-Gene Query tool available on the GTEx Portal. Specifically, we highlighted the expression patterns in kidney (Figure 3A) and pancreas tissues (Figure 3B).
Figure 4.
A) Kidney and B) pancreas tissue expression levels of the 17 creatine-related genes used in the present study (TPM - Transcripts per million bases).
Figure 4.
A) Kidney and B) pancreas tissue expression levels of the 17 creatine-related genes used in the present study (TPM - Transcripts per million bases).
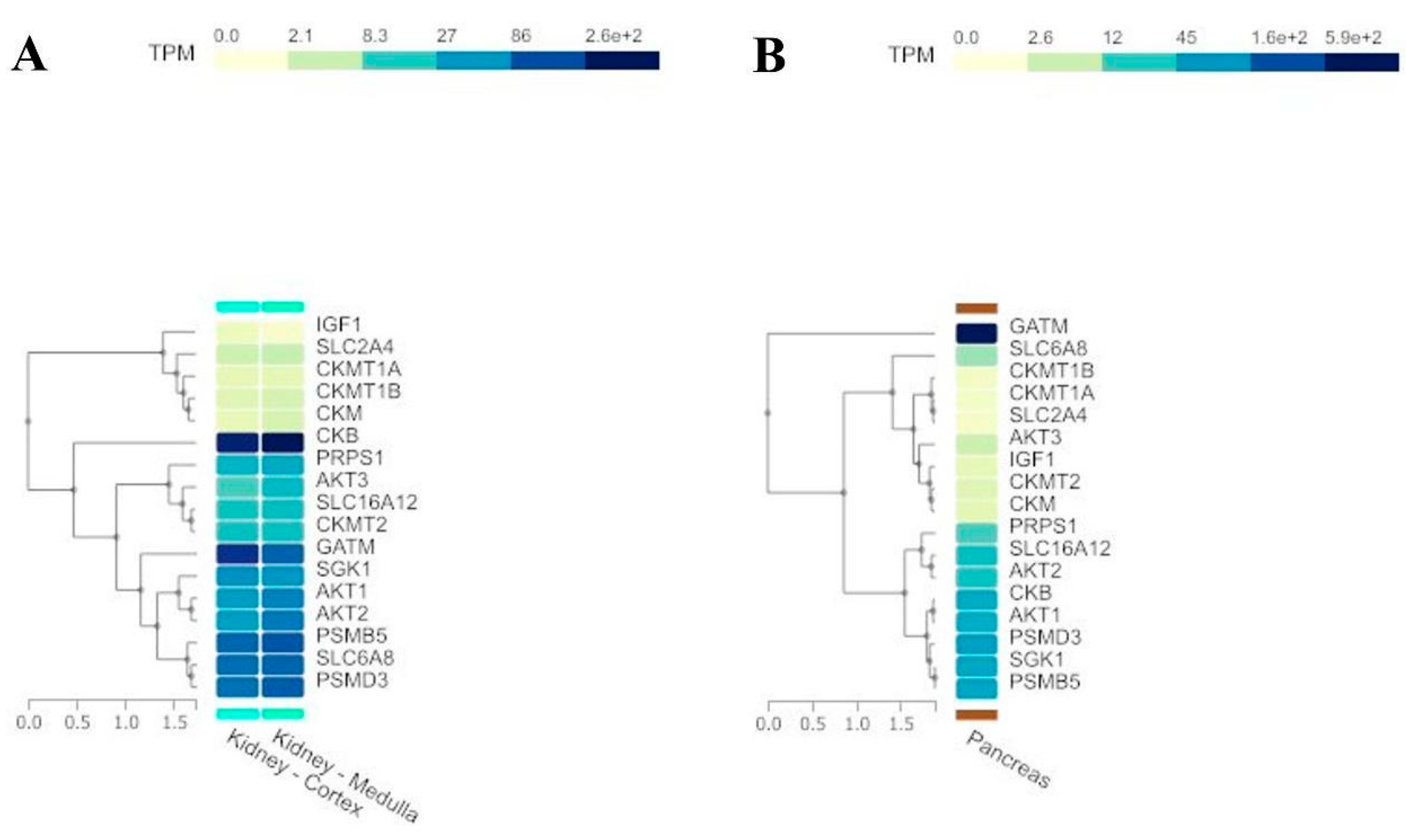
We can observe similar expression patterns for several creatine-related genes, especially GATM, SLC6A12, and SLC6A8. These genes are responsible for cell creatine homeostasis, balancing the production and uptake of this compound [65]. GATM encodes the enzyme Glycine Amidinotransferase, the primary regulatory step in creatine biosynthesis, and SLC6A8 and SLC6A12 genes encode the specific creatine transporter and the monocarboxylate transporter 12 (MCT12), respectively. Creatine concentration controls its synthesis by a feedback loop, regulating GATM expression [65].
As anticipated, the first two terms from the gene enrichment analysis against the ReactomeDB were directly related to creatine and amino acid metabolism (Figure 5A). Interestingly, the remaining eight terms are associated with processes related to cell cycle regulation, including the degradation and stability of well-known tumor suppressor genes TP53 and PTEN. We also noted that several terms overlapped with the genes PSMB5, AKT2, AKT1, AKT3, PSMD3, and SGK1. This integration of amino acid metabolism with specific glycolysis reactions may explain their association with cell cycle control. For instance, the glycolytic enzyme PKM2 phosphorylates and activates ERK1/2, crucial for cell proliferation. Another glycolytic enzyme, PFKFB3, promotes cell proliferation by enhancing glycolytic ATP production and modulating the expression of cell cycle regulators [67]. The involvement of creatine kinase (CK) in mitosis regulation is well-documented in scientific literature. Moreover, previous studies have reported conflicting roles of creatine metabolism in tumor cells, with both upregulation and downregulation of CK potentially affecting cell viability and inducing cell death, depending on the tumor's nature [68]. Through the GO analysis (Figure 5B), the study was able to categorize the genes in terms of biological processes, cellular components, and molecular functions, providing a broader view of these genes' capabilities and actions, and how their expression may be influenced by the presence of creatine in renal tissues. Our first analysis was against the GO dataset to verify the terms associated with these 17 genes. The resulting network from the top ten P-value-rated results is described in Figure 5B. The only direct integration of creatine metabolism to all other processes is the gene IGF1, which also participates in several carbohydrate-metabolism-related terms. Increasing the number of results to the top 30 rated results did not retrieve any other gene integration between creatine metabolism and other terms.
The pathway analysis using KEGG (Figure 5C) enabled the mapping of genes into specific metabolic pathway contexts, such as those involved in amino acid metabolism, elucidating how the convergence of metabolic activities and creatine supplementation may interact to impact renal function and homeostasis. Essentially, these analyses provided insights into how creatine can precisely alter renal metabolic pathways and which cellular processes may be particularly involved or affected.Additional findings are associated with insulin resistance and signaling metabolism, renal cell carcinoma, type 2 DM, central carbon metabolism in cancer, and others. The significant presence of AKT genes in various metabolic processes accounts for the numerous associated terms.
Moreover, this phase of analysis facilitated the identification of changes in the activity of specific genes that may be associated with processes such as cellular hypertrophy, apoptosis, and cellular repair. These findings are crucial for building a more comprehensive understanding of the implications of creatine intake, as alterations in cellular activity may have direct implications on renal function.
Pathway Analysis and Regulation by Kinases and Phosphatases
To investigate how kinase regulation processes influence creatine metabolism pathways, we perform GEA against two kinases-specific databases: ARCHS4 and The Kinase Library 2023. After merging the results, we found 24 kinases that are directly or indirectly related to the creatine-related genes (Table 5). We also joined this list with the creatine genes to evaluate their normal expression in kidney and pancreas tissues from GTEx (Figure 6). All the kinases presented high basal expression in the two tested tissues. A noteworthy observation is that both tissues present a very similar expression of these genes, and are consequently grouped in the same cluster when we analyze them together with the expression of all other human tissues (Supplementary Figure 1).
The study of metabolic pathways and enzymatic regulation helps elucidate not only how creatine is metabolized by the kidneys, but also how this metabolism may affect other cellular functions through kinase modulation. For instance, the impact of creatine on cellular energy levels and renal tissue homeostasis may be mediated by pathways regulated by specific kinases, providing a direct link between creatine supplementation and renal health status [68]. Additionally, exploring kinase regulation opens pathways to understanding broader cellular events, such as signal transduction and cell cycle regulation, which may have significant implications in renal disease contexts or under conditions of oxidative stress.
We focused on the AKT gene family due to their significant role in intracellular signaling and the regulation of processes such as cell growth, proliferation, and survival [69]. AKT proteins are involved in several important metabolic pathways and are known to be sensitive to variations in nutrient levels, including creatine [70]. Therefore, understanding how creatine influences AKT activity can reveal significant insights into the molecular mechanisms that result in the physiological effects of creatine in the kidneys. For instance, changes in AKT activity may impact cellular energy balance and protection against oxidative stress. These findings are significant as they offer potential for therapeutic manipulation in conditions where renal function is compromised or where regulation of energy metabolism is crucial.
Furthermore, by studying the expression of these kinases and their potential regulation in different kidney conditions/diseases, we can highlight a new layer of complexity in understanding the effects of creatine. It is not just a supplement that can affect energy capacity and muscle performance, but also an agent that interacts with signaling mechanisms and cellular regulation that are pivotal for normal renal health and function.
GEO datasets analyses
We did not find specific datasets from human organisms or cell models that evaluated the creatine treatment or supplementation on renal tissues. Therefore, we restricted our search to datasets that evaluated specific kidney diseases to analyze the expression of the genes in our list. The first part was to search the list's gene symbols against the GEO profiles database, which uses curated datasets to evaluate individual gene expression profiles. Within our search criteria, the gene GATM presented the highest number of profiles, with 16 distinct ones, most of them with significant differential expressions between the conditions tested. Then, we chose four distinct expression datasets from kidney conditions to widen the creatine-related genes assessment from array (three) and RNA-Seq platforms (one).
The first dataset evaluated was from the study of Neusser et al. [71], which analyzed genome-wide array expression of renal biopsy specimens from 14 patients with nephrosclerosis (NSC) against four tumor-free kidney specimens from patients undergoing tumor nephrectomy (TN) (GDS3712, GSE20602). The authors revealed a significant regulation of hypoxia-associated biological processes in NSC samples, including angiogenesis, fibrosis, and inflammation. The glomerular expression levels of most genes regulated by the hypoxia-inducible factors (HIFs) were also significantly altered in these samples [71]. We re-analyzed the dataset, focusing on the expression values from the creatine-related genes (Figure 7). Ten of the initial 17 genes of the list had significative differential expression in the above dataset: GATM, SLC6A8, IGF1, AKT1, AKT3, PRPS1, CKB, SGK1, PSMD3, and PSMB5.
Notably, the expression levels of these genes can retrieve the exact clusterization of the experiment samples (TN and NSC samples) (Figure 7). Compared to NSC samples, TN samples have an upregulated expression of transcript isoforms of GATM and SLC6A8 genes. In addition, the average expression of IGF1 transcripts is also higher in TN. As previously described, GATM encodes the enzyme Glycine Amidinotransferase, which plays a crucial role in creatine biosynthesis, and IGF1 is related to creatine and amino acid metabolism. Furthermore, the SLC6A8 codes the sodium- and chloride-dependent creatine transporter 1, responsible for the creatine transport into cells, against a concentration gradient. Recent studies reported the transcriptional upregulation of this gene in tumoral cells under hypoxic conditions [72,73]. The authors of these studies concluded that increased creatine production or uptake promotes survival by maintaining redox homeostasis in hypoxic cells.
Moreover, reduced active oxygen species (AOS) accumulation could activate the AKT-ERK signaling, which protects the viability of hypoxic tumoral cells and explains the relationship between creatine accumulation and members of the AKT gene family. Nephrosclerosis patients may have reduced creatine production and accumulation in the cell. Indeed, this compound is crucial to kidney functioning [14] and even more critical during gluconeogenesis cell metabolism to maintain energy levels when kidney cells rely on amino acids, fatty acids oxidation, and non-lactic pathways. Notably, the gradual reduction of tubular gluconeogenesis is a crucial feature of chronic kidney disease [74]. An uninterrupted and basal creatine supply could be why TN samples had regular expression of these genes, which may reflect normal kidney functioning. However, further studies are necessary to elucidate if creatine supplementation would be an efficient adjuvant protocol to improve renal function under these conditions.
We also analyzed the dataset from Flechner et al. [75] which evaluated kidney biopsy expression profiles after transplant, unique to acute rejection (AR samples), dysfunction without rejection (NR samples), and well-functioning transplants (TX Samples) (GDS724/GSE1563). For our analyses, we excluded the samples extracted from lymphocytes. Then, we evaluate the expression of the creatine-related genes of TX, AR, and NR samples versus Control kidney biopsies. Not all the genes were differentially expressed under our criteria (P<0.05). This dataset is particularly interesting due to a recent study by Post et al. [14], who conducted a clinical trial on kidney transplant recipients, evaluating creatine intake and endogenous production levels.
Our first comparison was AR against control samples, where 3607 transcripts were differentially expressed. Among them, we observed differential expression for the creatine-related genes AKT1, CKB, GATM, and SGK1. Although the DE of these genes was significant (P<0.05), only the GATM expression surpassed the threshold of Log2 Fold Change±0.5, which was intensely downregulated (Figure 8A). On the other hand, compared to the Controls, the expression of TX samples presented DE of the genes AKT1, AKT2, IGF1, PSMB5, and SLC6A8, but only IGF1 and AKT2 expressions were above the Log2FC threshold (Figure 8B). We can observe that GATM has the most significant downregulation in AR versus Control (p=0.00e+00) but no significant change in TX vs. Control. In addition, IGF1 and AKT1 show substantial changes in both comparisons, with smaller p-values in TX vs. Control (p=1.00e-03 and p=1.00e-04, respectively). On the other hand, AKT2 and SGK1 were significantly downregulated in AR samples (p=1.00e-04 and p=1.00e-03, respectively) but were not different in TX samples. We found no differential expression of the creatine-related genes in NR samples compared to controls.
We also compared the DE between conditions. In the TX versus AR comparison, GATM shows the most significant upregulation with a Log2 fold change around 2.4. PRPS1, CKMT2, PSMB5, SGK1, and PSMD3 are also upregulated to a lesser extent (log2 fold change between 0.3 and 0.7), and the genes AKT2, SLC6A8, and AKT3 were slightly downregulated (Figure 8C). Compared to NR samples, TX biopsies had increased expression of the genes GATM and PRPS1 and decreased expression of the AKT2, IGF1, and SLC6A8 (Figure 8D). Only IGF1 was upregulated in the comparison of AR versus NR samples. These results demonstrate that TX samples have induced expression of endogenous creatine production, marked by the increase in GATM compared to acute rejected transplant biopsies (AR) and dysfunctional but non-rejected samples (NR). It is also important to highlight that GATM was not differentially expressed in well-functioning transplants, whereas SLC6A8 was slightly downregulated when compared to control samples, which might indicate that the endogenous creatine production, or at least the activity of the GATM/AGAT enzyme, is the preferred pathway instead of the creatine uptake. Post et al. [14] stated that kidney transplant recipients may be at risk of impaired creatine synthesis, and the results of the present analysis partially corroborate theirs.
Since there is no consensus on the role of creatine homeostasis in cancer cells (47), we decided to analyze the expression of the creatine-related genes in the dataset GDS2880 (series GSE6344), which evaluated the transcriptional profile of two stages of Clear-Cell Renal cell Carcinomas (cRCC), paired with controls [76,77]. The first analysis was the comparison of cRCC versus Control samples, both in Stage I, where we observed the differential expression of 4694 genes. Still, only three are creatine-related: CKMT2, GATM, and SLC6A8 (Figure 9A). The genes CKMT2 and GATM show the largest negative log2 fold changes, indicating significant downregulation in the tumor cells compared to Normal ones. On the other hand, the gene SLC6A8 exhibits substantial upregulation in cRCC tissues. We also compared Stage II cRCC samples versus their paired Controls. In this analysis, we also found a strong downregulation of the genes GATM and CKMT2 in cRCC samples. These results indicate that cRCCs tissues have a severely affected endogenous creatine production. The higher expression of SLC6A8, especially during Stage I, might be a mechanism to increase the intracellular levels of this compound and maintain tumor viability [78] or to compensate for lower endogenous creatine synthesis to support kidney function.
It is interesting to note that cRCC samples have a downregulated GATM expression in both stages tested. The enzyme (AGAT) coded by this gene is the rate-limiting step of endogenous creatine biosynthesis [79]. Several previous studies discuss renal GATM expression and AGAT activity in the context of the creatine kidney contribution to the total body guanidinoacetate (GAA) synthesis (approximately 20%). However, recent studies have been relating kidney GATM expression to renal function and homeostasis maintenance, where the capacity of GAA production decreases with increased states of CKD, becoming virtually nonexistent in dialysis patients [13]. On the other side of the spectrum, several mutations in GATM result in known genetic inborn diseases linked to creatine synthesis deficiency, like Cerebral Creatine Synthesis Deficiency 3 (CCDS3), and to altered phenotypes, like renal Fanconi syndrome and progressive kidney failure [80], and in dialysis-independent CKD patients [81]. Our analysis also reveals that a downregulated GATM expression also occurs in cRCC samples, implying that increased creatine production, or at least GAA, may not be a strategy for this tumor-type survival, especially during Stage II of the disease. It could also imply that regular GATM expression is essential for human kidney function.
To better understand and confirm these observations, we analyzed the data of the RNA-Seq transcriptome profiling of chronic kidney disease from the series GSE137570 (BioProject: PRJNA565950; SRA: SRP222033). This data series provides clinical/morphological parameters for CKD samples, such as estimated Glomerular Filtration Rate (eGFR) and the tubulointerstitial fibrosis percentual of kidney biopsies. Using these parameters, we performed six different analyses. The first one was the general comparison of all Fibrosis samples versus Normal tissue biopsies, where three genes presented Log2 Fold Changes higher than the threshold used (Log2 Fold Change±0.5): IGF1 and SLC6A8 were upregulated. At the same time, SLC6A12 was downregulated (Figure 10A). However, fibrosis samples had different values of % Tubulointerstitial Fibrosis, which ranged from 0 to 77.5%. So, we separated the samples into two additional groups with tubulointerstitial fibrosis above and below 50% (50%<Fibrosis and 50%>Fibrosis, respectively). Two creatine-related genes presented downregulation, PSMB5 and SLC6A12, while IGF-1 was upregulated when comparing 50%<Fibrosis versus Normal samples (Figure 10B). We also observed similar IGF-1 expression values when comparing 50%>Fibrosis versus Normal samples (Figure 10C). The genes GAMT, SLC6A8, and PSMD3 showed significant upregulation in the 50%>Fibrosis (Figure 10C).
Fibrotic renal tissues from this cohort could have increased IGF1 due to insulin resistance or in the first stages of diabetic kidney disease (DKD), or its upregulation could be linked to kidney hypertrophy, a known indicative of DKD, as previously demonstrated in animal models [82]. The action of this hormone-like molecule is to induce glucose consumption, inducing Glycolytic enzymes, which in turn could feed the creatine synthesis. Renal tissues require high energy levels, and during CKD, this tissue has a partial loss of fatty acid oxidation and an impaired capacity to produce glucose due to loss of gluconeogenesis and an increase in glycolysis [74]. However, we did not observe differential expression of the GATM gene, which is the committing step for the pathway of this compound synthesis. Perhaps GATM expression levels were close to the ones observed in Normal samples, maintaining GAA intracellular levels. GAA is toxic to the cell at high concentrations. Therefore, renal samples with lower than 50% tubulointerstitial fibrosis could upregulate the GAMT gene to metabolize GAA excess, leading to creatine synthesis. The significant upregulation of the gene PSMD3, a proteasome protease, might provide the amino acids as substrate for this pathway and not carbon skeletons to gluconeogenesis, which would be reduced under the action of IGF-1 [82]. Since we also observed an upregulation of the creatine transporter gene (SLC6A8) in these samples, creatine must be related to renal function maintenance or energy supply. As fibrosis increases, the decreased expression of the monocarboxylic acid transporter, SLC6A12, may be linked to renal medulla osmoregulation. This transporter is crucial to the control of betaine accumulation [83].
We also categorized the samples from this study by the eGFR values, using a threshold of 50. We know this value is not a dividing point between the presence and absence of renal disease. The eGFR is indicative, calculated from the plasma creatinine levels, and, though related, is not equal to the actual glomerular filtration rate (GFR) [14]. The usual range used to indicate a normal kidney function is above 90; values between 60-89 may indicate early-stage kidney disease; values between 15-59 may mean kidney disease; and values below 15 indicate kidney failure (National Institute of Diabetes and Digestive and Kidney diseases and National Kidney Foundation guidelines). However, we chose a lower threshold because gene expression alterations commonly occur before observable changes in clinical and morphological parameters. So, eGFR values indicative of early-stage renal disease may not imply an intense remodeling of gene expression, especially the ones related to energy production shifts.
Interestingly, the kidney biopsy samples with eGFR values above 50 (eGFR>50) had a similar differential expression of the creatine-related genes observed in the samples with lower Fibrosis percentages (50%>Fibrosis) (Figure 10D and 10C), respectively, including the same DE genes. In addition, samples with lower eGFR (eGFR<50) presented altered expressed levels of the genes GATM and SLC6A12 (downregulated) and the genes IGF1 and SLC6A8 (upregulated) (Figure 10E). These results demonstrate that eGFR<50 samples have impaired endogenous creatine synthesis, marginally compensated by the increase of this compound absorption by its specific transporter, SLC6A8, compared to the Normal samples. Also, a previous study associated increased serum IGF1 levels with low eGFR rates [84]. From our gene list, only the GATM and GAMT genes were significantly downregulated when we contrasted the gene expression of eGFR<50 versus eGFR>50 samples (Figure 10F). These observations mean that there might be a relationship between the eGFR values and endogenous creatine production. These findings also corroborate previous suggestions of [13,14] and [85] that impaired endogenous creatine production in chronically and dialysis-dependent kidney disease patients creatine becomes an essential nutrient, and they may require creatine supplementation. However, carefully evaluating the patient's diet and creatine ingestion/supplementation is imperative to maintain kidney function and homeostasis.
Our creatine-related gene expression analyses provide additional support for previous studies, but they are not without limitations. The case-control comparisons could be more complex because gene expression overlaps between different tested case conditions could occur. In addition, no information on the creatine content or consumption of the samples' donors is available, which could directly impact our evaluations. Finally, the studies used experimental designs, which may need to be more suited to our questions. Therefore, we emphasize the need for specific studies with human or animal models that could address the questions raised here and confirm the central role of creatine homeostasis in kidney function.
Future Directions
Creatine supplementation still warrants a deeper understanding of its mechanisms of action, potential therapeutic applications in diseases, and its association with nephropathies. For instance, the potential elevation in serum creatinine levels, resulting from the spontaneous conversion of creatine to creatinine, was among the factors that led to scrutiny and questioning of the sale of creatine supplements in certain countries in the past. This episode appears to continue influencing new studies, as there is still hesitation in exploring the effects of creatine supplementation on nephropathies. Due to the spontaneous conversion of creatine to creatinine, serum creatinine should not be solely relied upon as a biomarker of renal function post-creatine supplementation. In such instances, elevated creatinine levels stem from increased creatine levels rather than renal dysfunction, resulting in a false positive indication of kidney injury. Other markers such as serum/urinary urea, cystatin-C, imaging, and molecular tests [86] should be considered in future studies to enhance the accuracy of renal function evaluation, whether in experimental or clinical settings.
In the case of diabetic nephropathy (DN), there are currently no studies confirming the effects of creatine supplementation in patients with this condition, only animal model studies [54]. However, like CKD and RT, some hypotheses suggest a potential positive outcome with creatine. Creatine supplementation might enhance renal function in DN patients by increasing glucose uptake through GLUT-4 translocation to the sarcolemma, as observed in type 2 diabetes patients [42]. However, the mechanism by which creatine supplementation increases GLUT-4 translocation has not been fully elucidated. The findings of Alves et al. [87] suggested that the increased expression of AMPK-α could explain the increased translocation of GLUT-4 to the cell membrane, but the increased expression of AMPK-α did not show statistical significance between the groups in this study. Therefore, this mechanism requires further investigation.
In a previous study, we observed that creatine supplementation did not induce significant changes in the renal morphology of diabetic animals [54]. Additionally, we found a hypoglycemic effect of creatine supplementation in this model. Based on the findings of Gualano et al. [42], the hypoglycemic effect we observed may have occurred due to increased expression of GLUT-4 and unconventional myosin-Ic, proteins that could enhance glucose uptake.
In addition, several recent studies evidence that the balance between creatine synthesis and uptake in renal tissues is directly linked to kidney function. Several abnormal kidney conditions induce the differential expression of creatine-related genes, especially GATM, GAMT, SLC6A8, AKTs, and IGF1. Further studies with specific experimental designs need to address whether the expression levels of these genes can be molecular markers of creatine homeostasis and kidney function.
Conclusions
There is compelling evidence that creatine supplementation is safe and does not alter function in healthy individuals without prior renal impairment. However, in individuals with kidney diseases, clinical studies are scarce, and experimental data suggest that, due to the particularities of each condition, the effects of creatine supplementation should be considered on an individual basis.
The bioinformatic approach of this study sheds light on the intricate relationship between creatine and renal tissues, offering valuable insights into the potential effects of creatine supplementation on renal function. The findings highlight the importance of understanding the impact of creatine on renal health and underscore the need for further research in this area. Future studies could explore the expression of kinases in different renal conditions, elucidating a new layer of complexity in understanding the effects of creatine.
Furthermore, experimental studies in human or animal models could confirm the central role of creatine homeostasis in renal function. Additional research could advance our understanding of the influence of creatine on renal tissues and contribute to the development of personalized approaches to support renal function. Based on current information, hypotheses suggest that creatine may benefit patients with kidney diseases directly or indirectly, or at least not present adverse effects.
Author Contributions
Conceptualization: JPMSL and MAM; writing—original draft preparation: JPMSL, MAM, and BJA.; writing-review and editing: JPMSL and BJA. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
AGAT - Arginine
ADP - adenosine diphosphate Glycine Amidinotransferase deficiency syndrome
AMPK-α - AMP-activated protein kinase alpha
AOS - Active oxygen species
ATK1 - V-akt murine thymoma viral oncogene homolog 1
ATK2 - V-akt murine thymoma viral oncogene homolog 2
ATK3 - V-akt murine thymoma viral oncogene homolog 3
ATP - Adenosine triphosphate
CK - Creatine kinase
CKB - Creatine Kinase B
CKD - Chronic kidney disease
CKM - Creatine kinase, muscle
CKMT1A - Creatine kinase, mitochondrial 1A
CKMT1B - Creatine kinase, mitochondrial 1B
DKD – Diabetic kidney disease
DN - Diabetic nephropathy
ERK1/2 - Extracellular signal-regulated kinase ½
GAA - Guanidinoacetate
GAMT - Guanidinoacetate methyltransferase
GEO - Gene Expression Omnibus
GLUT4 - Glucose transporter type 4
GO - Gene Ontologies
GTEx - Genotype-Tissue Expression
HIV - Human Immunodeficiency Virus
IGF1 - Insulin-like growth factor 1
KEGG - Kyoto Encyclopedia of Genes and Genomes
MtCK - Mitochondrial creatine kinase
MYO1C - Myosin IC
NSC - Nephrosclerosis
PFKFB3 - 6-phosphofructo-2-kinase/fructose-2,6-biphosphatase 3
PKM2 - Pyruvate kinase isozymes M2
PPI - Protein-protein interaction
PSMB5 - Proteasome subunit beta type-5
PSMD3 - 26S proteasome non-ATPase regulatory subunit 3
PTEN - Phosphatase and tensin homologue
RT - Renal transplant
SGK1 - Serine/threonine-protein kinase 1
SLC2A4 - Solute Carrier Family 2 Member 4
TN - Tumor nephrectomy
TP53 - Tumor protein p53
TPM - Transcripts per million
References
- Antonio, J.; Candow, D.G.; Forbes, S.C.; Gualano, B.; Jagim, A.R.; Kreider, R.B.; Rawson, E.S.; Smith-Ryan, A.E.; VanDusseldorp, T.A.; Willoughby, D.S.; et al. Common questions and misconceptions about creatine supplementation: what does the scientific evidence really show? J. Int. Soc. Sports Nutr. 2021, 18, 13. [Google Scholar] [CrossRef]
- Gastin, P.B. Energy System Interaction and Relative Contribution During Maximal Exercise. Sports Med. 2001, 31, 725–741. [Google Scholar] [CrossRef]
- Chanutin, A.B.; Guy, P. The fate of creatine when administered to man. J Biol. Chem. 1926, 67, 29–37. [Google Scholar] [CrossRef]
- Harris, R.C.; Söderlund, K.; Hultman, E. Elevation of creatine in resting and exercised muscle of normal subjects by creatine supplementation. Clin. Sci. 1992, 83, 367–374. [Google Scholar] [CrossRef]
- Poortmans, J.R.; Francaux, M. Adverse effects of creatine supplementation: fact or fiction? Sports Med. 2000, 30, 155–170. [Google Scholar] [CrossRef]
- Kreider, R.B.; Kalman, D.S.; Antonio, J.; Ziegenfuss, T.N.; Wildman, R.; Collins, R.; Candow, D.G.; Kleiner, S.M.; Almada, A.L.; Lopez, H.L. International Society of Sports Nutrition position stand: safety and efficacy of creatine supplementation in exercise, sport, and medicine. J. Int. Soc. Sports Nutr. 2017, 14, 1–18. [Google Scholar] [CrossRef]
- Hall, M.; Trojian, T.H. Creatine supplementation. Curr. Sports Med. Rep. 2013, 12, 240–244. [Google Scholar] [CrossRef]
- Harmon, K.K.; Stout, J.R.; Fukuda, D.H.; Pabian, P.S.; Rawson, E.S.; Stock, M.S. The Application of Creatine Supplementation in Medical Rehabilitation. Nutrients 2021, 13, 1825. [Google Scholar] [CrossRef]
- Roschel, H.; Gualano, B.; Ostojic, S.M.; Rawson, E.S. Creatine Supplementation and Brain Health. Nutrients 2021, 13, 586. [Google Scholar] [CrossRef] [PubMed]
- Forbes, S.C.; Cordingley, D.M.; Cornish, S.M.; Gualano, B.; Roschel, H.; Ostojic, S.M.; Rawson, E.S.; Roy, B.D.; Prokopidis, K.; Giannos, P.; et al. Effects of Creatine Supplementation on Brain Function and Health. Nutrients 2022, 14, 921. [Google Scholar] [CrossRef] [PubMed]
- Prokopidis, K.; Giannos, P.; Triantafyllidis, K.K.; Kechagias, K.S.; Forbes, S.C.; Candow, D.G. Effects of creatine supplementation on memory in healthy individuals: a systematic review and meta-analysis of randomized controlled trials. Nutr. Rev. 2023, 81, 416–427. [Google Scholar] [CrossRef]
- Farquhar, W.B.; Zambraski, E.J. Effects of creatine use on the athlete's kidney. Curr. Sports Med. Rep. 2002, 1, 103–106. [Google Scholar] [CrossRef]
- Post, A.; Tsikas, D.; Bakker, S.J. Creatine is a Conditionally Essential Nutrient in Chronic Kidney Disease: A Hypothesis and Narrative Literature Review. Nutrients 2019, 11, 1044. [Google Scholar] [CrossRef]
- Post, A.; Groothof, D.; Kremer, D.; Knobbe, T.J.; Abma, W.; Koops, C.A.; Tsikas, D.; Wallimann, T.; Dullaart, R.P.; Franssen, C.F.; et al. Creatine homeostasis and the kidney: comparison between kidney transplant recipients and healthy controls. Amino Acids 2024, 56, 42. [Google Scholar] [CrossRef]
- Baker, S.A.; Gajera, C.R.; Wawro, A.M.; Corces, M.R.; Montine, T.J. GATM and GAMT synthesize creatine locally throughout the mammalian body and within oligodendrocytes of the brain. Brain Res. 2021, 1770, 147627. [Google Scholar] [CrossRef]
- Poortmans, J.R.; Auquier, H.; Renaut, V.; Durussel, A.; Saugy, M.; Brisson, G.R. Effect of short-term creatine supplementation on renal responses in men. Eur. J. Appl. Physiol. Occup. Physiol. 1997, 76, 566–567. [Google Scholar] [CrossRef]
- Poortmans, J.R.; Francaux, M. Long-term oral creatine supplementation does not impair renal function in healthy athletes. Med. Sci. Sports Exerc. 1999, 31, 1108–1110. [Google Scholar] [CrossRef]
- Pritchard, N.; Kalra, P. Renal dysfunction accompanying oral creatine supplements. Lancet 1998, 351, 1252–1253. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Santos, A.; von Mering, C.; Jensen, L.J.; Bork, P.; Kuhn, M. STITCH 5: Augmenting Protein–Chemical Interaction Networks with Tissue and Affinity Data. Nucleic Acids Res. 2016, 44, D380–D384. [Google Scholar] [CrossRef]
- Davis, A.P.; Wiegers, T.C.; Johnson, R.J.; Sciaky, D.; Wiegers, J.; Mattingly, C.J. Comparative Toxicogenomics Database (CTD): update 2023. Nucleic Acids Res. 2022, 51, D1257–D1262. [Google Scholar]
- Xie, Z.; Bailey, A.; Kuleshov, M.V.; Clarke, D.J.B.; Evangelista, J.E.; Jenkins, S.L.; Lachmann, A.; Wojciechowicz, M.L.; Kropiwnicki, E.; Jagodnik, K.M.; et al. Gene Set Knowledge Discovery with Enrichr. Curr. Protoc. 2021, 1, e90. [Google Scholar] [CrossRef]
- Evangelista, J.E.; Xie, Z.; Marino, G.B.; Nguyen, N.; Clarke, D.J.B.; Ma’ayan, A. Enrichr-KG: bridging enrichment analysis across multiple libraries. Nucleic Acids Res. 2023, 51, W168–W179. [Google Scholar] [CrossRef]
- Lachmann, A.; Torre, D.; Keenan, A.B.; Jagodnik, K.M.; Lee, H.J.; Wang, L.; Silverstein, M.C.; Ma’ayan, A. Massive mining of publicly available RNA-seq data from human and mouse. Nat. Commun. 2018, 9, 1366. [Google Scholar] [CrossRef]
- Johnson, J.L.; Yaron, T.M.; Huntsman, E.M.; Kerelsky, A.; Song, J.; Regev, A.; Lin, T.-Y.; Liberatore, K.; Cizin, D.M.; Cohen, B.M.; et al. An atlas of substrate specificities for the human serine/threonine kinome. Nature 2023, 613, 759–766. [Google Scholar] [CrossRef]
- Barrett, T.; Wilhite, S.E.; Ledoux, P.; Evangelista, C.; Kim, I.F.; Tomashevsky, M.; Marshall, K.A.; Phillippy, K.H.; Sherman, P.M.; Holko, M.; et al. NCBI GEO: archive for functional genomics data sets—update. Nucleic Acids Res. 2013, 41, D991–D995. [Google Scholar] [CrossRef]
- Davis, S.; Meltzer, P.S. GEOquery: a bridge between the Gene Expression Omnibus (GEO) and BioConductor. Bioinformatics 2007, 23, 1846–1847. [Google Scholar] [CrossRef]
- Smyth, G.K. Limma: linear models for microarray data. In: Bioinformatics and Computational Biology Solutions using R and Bioconductor, R. Gentleman, V. Carey, S. Dudoit, R. Irizarry, W. Huber (eds.), Springer, New York, 2005, 397-420.
- Love, M.I.; Huber, W.; Anders, S. “Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. ” Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer-Verlag: New York, NY, USA, 2016; ISBN 978-3-319-24277-4. https://ggplot2.tidyverse.org. [Google Scholar]
- Harris, C.R.; Millman, K.J.; van der Walt, S.J.; Gommers, R.; Virtanen, P.; Cournapeau, D.; Wieser, E.; Taylor, J.; Berg, S.; Smith, N.J.; et al. Array programming with NumPy. Nature 2020, 585, 357–362. [Google Scholar] [CrossRef]
- Hunter, J.D. Matplotlib: A 2D graphics environment. Comput. Sci. Eng. 2007, 9, 90–95. [Google Scholar] [CrossRef]
- Centers for disease control and prevention (CDC). Hyperthermia and dehydration-related deaths associated with intentional rapid weight loss in three collegiate wrestlers--North Carolina, Wisconsin, and Michigan, November-December 1997. MMWR. Morb. Mortal. Wkly. Rep. 1998, 47, 105–108. [Google Scholar]
- Kuehl, K.M.; Goldberg, L.M.; Elliot, D.M. Renal insufficiency after creatine supplementation in a college football athlete. Med. Sci. Sports Exerc. 1998, 30, 235. [Google Scholar] [CrossRef]
- Koshy, K.M.; Griswold, E.; E Schneeberger, E. Interstitial Nephritis in a Patient Taking Creatine. New Engl. J. Med. 1999, 340, 814–815. [Google Scholar] [CrossRef]
- Barisic, N.; Bernert, G.; Ipsiroglu, O.; Stromberger, C.; Müller, T.; Gruber, S.; Prayer, D.; Moser, E.; Bittner, R.E.; Stöckler-Ipsiroglu, S. Effects of Oral Creatine Supplementation in a Patient with MELAS Phenotype and Associated Nephropathy. Neuropediatrics 2002, 33, 157–161. [Google Scholar] [CrossRef] [PubMed]
- Révai, T.; Sápi, Z.; Benedek, S.; Kovács, A.; Kaszás, I.; Virányi, M.; Winkler, G. Severe nephrotic syndrome in a young man taking anabolic steroid and creatine long term. Orvosi hetilap. 2003, 144, 2425–2427. [Google Scholar] [PubMed]
- Thorsteinsdottir, B.; Grande, J.P.; Garovic, V.D. Acute Renal Failure in a Young Weight Lifter Taking Multiple Food Supplements, Including Creatine Monohydrate. J. Ren. Nutr. 2006, 16, 341–345. [Google Scholar] [CrossRef]
- Taner, B.; Aysim, O.; Abdulkadir, U. The effects of the recommended dose of creatine monohydrate on kidney function. NDT Plus 2011, 4, 23–24. [Google Scholar] [CrossRef]
- Gualano, B.; Ugrinowitsch, C.; Novaes, R.B.; Artioli, G.G.; Shimizu, M.H.; Seguro, A.C.; Harris, R.C.; Lancha, A.H., Jr. Effects of creatine supplementation on renal function: a randomized, double-blind, placebo-controlled clinical trial. Eur. J. Appl. Physiol. 2008, 103, 33–40. [Google Scholar] [CrossRef]
- Gualano, B.; Ferreira, D.C.; Sapienza, M.T.; Seguro, A.C.; Lancha, A.H., Jr. Effect of Short-term High-Dose Creatine Supplementation on Measured GFR in a Young Man With a Single Kidney. Am. J. Kidney Dis. 2010, 55, e7–e9. [Google Scholar] [CrossRef]
- Carvalho, A.P.P.F.; Molina, G.E.; Fontana, K.E. Creatine supplementation associated with resistance training does not alter renal and hepatic functions. Rev. Bras. Med. Esporte. 2011, 17, 1–5. [Google Scholar]
- Gualano, B.; de Salles Painelli, V.; Roschel, H.; Lugaresi, R.; Dorea, E.; Artioli, G.G.; Lima, F.R.; da Silva, M.E.; Cunha, M.R.; Seguro, A.C.; et al. Creatine supplementation does not impair kidney function in type 2 diabetic patients: A randomized, double-blind, placebo-controlled, clinical trial. Eur. J. Appl. Physiol. 2011, 111, 749–756. [Google Scholar] [CrossRef]
- Almeida, D.; Colombini, A.; Machado, M. Creatine supplementation improves performance, but is it safe? Double-blind placebo-controlled study. J. Sports Med. Phys. Fit. 2020, 60, 1034–1039. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Kim, C.K.; Carpentier, A.; Poortmans, J.R. Studies on the safety of creatine supplementation. Amino Acids 2011, 40, 1409–1418. [Google Scholar] [CrossRef] [PubMed]
- Baracho, N.C.; Castro, L.P.; Borges, N.C.; Laira, P.B. Study of renal and hepatic toxicity in rats supplemented with creatine. Acta Cir. Bras. 2015, 30, 313–318. [Google Scholar] [CrossRef] [PubMed]
- Davani-Davari, D.; Karimzadeh, I.; Ezzatzadegan-Jahromi, S.; Sagheb, M.M. Potential Adverse Effects of Creatine Supplement on the Kidney in Athletes and Bodybuilders. Iran. J. Kidney Dis. 2018, 12, 253–260. [Google Scholar] [PubMed]
- Edmunds, J.W.; Jayapalan, S.; DiMarco, N.M.; Saboorian, H.; Aukema, H.M. Creatine Supplementation Increases Renal Disease Progression in Han:SPRD-cy Rats. Am. J. Kidney Dis. 2001, 37, 73–78. [Google Scholar] [CrossRef] [PubMed]
- Taes, Y.E.C.; Delanghe, J.R.; Wuyts, B.; van de Voorde, J.; Lameire, N.H. Creatine supplementation does not affect kidney function in an animal model with pre-existing renal failure. Nephrol. Dial. Transplant. 2003, 18, 258–264. [Google Scholar] [CrossRef]
- Genc, G.; Okuyucu, A.; Meydan, B.C.; Yavuz, O.; Nisbet, O.; Hokelek, M.; Bedir, A.; Ozkaya, O. Effect of free creatine therapy on cisplatin-induced renal damage. Ren. Fail. 2014, 36, 1108–1113. [Google Scholar] [CrossRef]
- Ferreira, L.G.; Bergamaschi, C.D.T.; Lazaretti-Castro, M.; Heilberg, I.P. Effects of Creatine Supplementation on Body Composition and Renal Function in Rats. Med. Sci. Sports Exerc. 2005, 37, 1525–1529. [Google Scholar] [CrossRef]
- A Souza, R.; Miranda, H.; Xavier, M.; A Lazo-Osorio, R.; A Gouvea, H.; Cogo, J.C.; Vieira, R.P.; Ribeiro, W. Effects of high-dose creatine supplementation on kidney and liver responses in sedentary and exercised rats. J Sports Sci Med 2009, 8, 672–681. [Google Scholar]
- Souza, W.M.; Heck, T.G.; Wronski, E.C.; Ulbrich, A.Z.; Boff, E. Effects of creatine supplementation on biomarkers of hepatic and renal function in young trained rats. Toxicol. Mech. Methods 2013, 23, 697–701. [Google Scholar] [CrossRef]
- Fernandes, V.A.R.; Delforno, M.C.; Banov, G.C.; Shmayev, M.; Leandro, J.V.A.; Teixeira, K.F.G.; Iatecola, A.; Cardozo, M.F.I.; Caldeira, E.J.; da Cunha, M.R. Renal, hepatic and muscle effects of creatine supplementation in an older adults experimental model. Clin. Nutr. ESPEN 2022, 48, 464–471. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, M.G.; Medeiros, M.A.; de Lemos, L.I.C.; Pedrosa, L.d.F.C.; Santos, P.P.d.A.; Abreu, B.J.; Lima, J.P.M.S. Effects of Creatine Supplementation on Histopathological and Biochemical Parameters in the Kidney and Pancreas of Streptozotocin-Induced Diabetic Rats. Nutrients 2022, 14, 431. [Google Scholar] [CrossRef]
- Gualano, B.; Roschel, H.; Lancha, A.H., Jr.; Brightbill, C.E.; Rawson, E.S. In sickness and in health: the widespread application of creatine supplementation. Amino Acids. 2012, 43, 519–529. [Google Scholar] [CrossRef] [PubMed]
- Ferrante, R.J.; Andreassen, O.A.; Jenkins, B.G.; Dedeoglu, A.; Kuemmerle, S.; Kubilus, J.K.; Kaddurah-Daouk, R.; Hersch, S.M.; Beal, M.F. Neuroprotective effects of creatine in a transgenic mouse model of Huntington's disease. J Neurosci. 2000, 20, 4389–4397. [Google Scholar] [CrossRef]
- Deminice, R.; Cella, P.S.; Padilha, C.S.; Borges, F.H.; da Silva, L.E.C.M.; Campos-Ferraz, P.L.; Jordao, A.A.; Robinson, J.L.; Bertolo, R.F.; Cecchini, R.; et al. Creatine supplementation prevents hyperhomocysteinemia, oxidative stress and cancer-induced cachexia progression in Walker-256 tumor-bearing rats. Amino Acids 2016, 48, 2015–2024. [Google Scholar] [CrossRef]
- Candow, D.G.; Chilibeck, P.D.; Forbes, S.C.; Fairman, C.M.; Gualano, B.; Roschel, H. Creatine supplementation for older adults: Focus on sarcopenia, osteoporosis, frailty and Cachexia. Bone 2022, 162, 116467. [Google Scholar] [CrossRef]
- Op 't Eijnde, B.; Ursø, B.; Richter, E.A.; Greenhaff, P.L.; Hespel, P. Effect of oral creatine supplementation on human muscle GLUT4 protein content after immobilization. Diabetes 2001, 50, 18–23. [Google Scholar] [CrossRef]
- Young, J.C.; E Young, R. The effect of creatine supplementation on glucose uptake in rat skeletal muscle. Life Sci. 2002, 71, 1731–1737. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, S.S.; Ahmad, K.; Lee, E.J.; Lee, Y.-H.; Choi, I. Implications of Insulin-Like Growth Factor-1 in Skeletal Muscle and Various Diseases. Cells 2020, 9, 1773. [Google Scholar] [CrossRef]
- Kineman, R. D.; Rio-Moreno, M.; del Sarmento-Cabral, A. 40 YEARS of IGF1: Understanding the tissue-specific roles of IGF1/IGF1R in regulating metabolism using the Cre/loxP system. J. Mol. Endocrinol. 2018, 61, T187–T198. [Google Scholar] [CrossRef]
- Bach, L.A.; Hale, L.J. Insulin-like Growth Factors and Kidney Disease. Am. J. Kidney Dis. 2015, 65, 327–336. [Google Scholar] [CrossRef] [PubMed]
- Mohebi, R.; Liu, Y.; Hansen, M.K.; Yavin, Y.; Sattar, N.; Pollock, C.A.; Butler, J.; Jardine, M.; Masson, S.; Heerspink, H.J.L.; et al. Insulin growth factor axis and cardio-renal risk in diabetic kidney disease: an analysis from the CREDENCE trial. Cardiovasc. Diabetol. 2023, 22, 176. [Google Scholar] [CrossRef] [PubMed]
- Stelzer, G.; Rosen, N.; Plaschkes, I.; Zimmerman, S.; Twik, M.; Fishilevich, S.; Stein, T.I.; Nudel, R.; Lieder, I.; Mazor, Y.; et al. The GeneCards Suite: From Gene Data Mining to Disease Genome Sequence Analyses. Curr. Protoc. Bioinform. 2016, 54, 1–30. [Google Scholar] [CrossRef] [PubMed]
- Tropak, M.B.; Tkachyova, I.; Gu, R.; Lee, A.; Schulze, A. Evidence of an intracellular creatine-sensing mechanism that modulates creatine biosynthesis via AGAT expression in human HAP1 cells. Sci. Rep. 2023, 13, 22392. [Google Scholar] [CrossRef]
- Kalucka, J.; Missiaen, R.; Georgiadou, M.; Schoors, S.; Lange, C.; De Bock, K.; Dewerchin, M.; Carmeliet, P. Metabolic control of the cell cycle. Cell Cycle 2015, 14, 3379–3388. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.-B. Creatine kinase in cell cycle regulation and cancer. Amino Acids 2016, 48, 1775–1784. [Google Scholar] [CrossRef] [PubMed]
- Tang, C.; Livingston, M.J.; Liu, Z.; Dong, Z. Autophagy in kidney homeostasis and disease. Nat. Rev. Nephrol. 2020, 16, 489–508. [Google Scholar] [CrossRef]
- Yuan, Q.; Tang, B.; Zhang, C. Signaling pathways of chronic kidney diseases, implications for therapeutics. Signal Transduct. Target. Ther. 2022, 7, 182. [Google Scholar] [CrossRef] [PubMed]
- Neusser, M.A.; Lindenmeyer, M.T.; Moll, A.G.; Segerer, S.; Edenhofer, I.; Sen, K.; Stiehl, D.P.; Kretzler, M.; Gröne, H.-J.; Schlöndorff, D.; et al. Human Nephrosclerosis Triggers a Hypoxia-Related Glomerulopathy. Am. J. Pathol. 2010, 176, 594–607. [Google Scholar] [CrossRef]
- Li, Q.; Liu, M.; Sun, Y.; Jin, T.; Zhu, P.; Wan, X.; Hou, Y.; Tu, G. SLC6A8-mediated intracellular creatine accumulation enhances hypoxic breast cancer cell survival via ameliorating oxidative stress. J. Exp. Clin. Cancer Res. 2021, 40, 168. [Google Scholar] [CrossRef]
- Rashidi, A.; Billingham, L.K.; Zolp, A.; Chia, T.-Y.; Silvers, C.; Katz, J.L.; Park, C.H.; Delay, S.; Boland, L.; Geng, Y.; et al. Myeloid cell-derived creatine in the hypoxic niche promotes glioblastoma growth. Cell Metab. 2024, 36, 62–77.e8. [Google Scholar] [CrossRef] [PubMed]
- Dalga, D.; Verissimo, T.; Seigneux, S. de Gluconeogenesis in the Kidney: In Health and in Chronic Kidney Disease. Clin. Kidney J. 2023, 16, 1249–1257. [Google Scholar] [CrossRef] [PubMed]
- Flechner, S.M.; Kurian, S.M.; Head, S.R.; Sharp, S.M.; Whisenant, T.C.; Zhang, J.; Chismar, J.D.; Horvath, S.; Mondala, T.; Gilmartin, T.; et al. Kidney Transplant Rejection and Tissue Injury by Gene Profiling of Biopsies and Peripheral Blood Lymphocytes. Am. J. Transplant. 2004, 4, 1475–1489. [Google Scholar] [CrossRef] [PubMed]
- Gumz, M.L.; Zou, H.; Kreinest, P.A.; Childs, A.C.; Belmonte, L.S.; LeGrand, S.N.; Wu, K.J.; Luxon, B.A.; Sinha, M.; Parker, A.S.; et al. Secreted Frizzled-Related Protein 1 Loss Contributes to Tumor Phenotype of Clear Cell Renal Cell Carcinoma. Clin. Cancer Res. 2007, 13, 4740–4749. [Google Scholar] [CrossRef] [PubMed]
- Tun, H.W.; Marlow, L.A.; von Roemeling, C.A.; Cooper, S.J.; Kreinest, P.; Wu, K.; Luxon, B.A.; Sinha, M.; Anastasiadis, P.Z.; Copland, J.A. Pathway Signature and Cellular Differentiation in Clear Cell Renal Cell Carcinoma. PLOS ONE 2010, 5, e10696. [Google Scholar] [CrossRef]
- Rashidi, A.; Billingham, L.K.; Zolp, A.; Chia, T.-Y.; Silvers, C.; Katz, J.L.; Park, C.H.; Delay, S.; Boland, L.; Geng, Y.; et al. Myeloid cell-derived creatine in the hypoxic niche promotes glioblastoma growth. Cell Metab. 2024, 36, 62–77. [Google Scholar] [CrossRef]
- Yu, L.; Wang, L.; Hu, G.; Ren, L.; Qiu, C.; Li, S.; Zhou, X.; Chen, S.; Chen, R. Reprogramming alternative macrophage polarization by GATM-mediated endogenous creatine synthesis: A potential target for HDM-induced asthma treatment. Front. Immunol. 2022, 13, 937331. [Google Scholar] [CrossRef]
- Reichold, M.; Klootwijk, E.D.; Reinders, J.; Otto, E.A.; Milani, M.; Broeker, C.; Laing, C.; Wiesner, J.; Devi, S.; Zhou, W.; et al. Glycine Amidinotransferase (GATM), Renal Fanconi Syndrome, and Kidney Failure. J. Am. Soc. Nephrol. 2018, 29, 1849–1858. [Google Scholar] [CrossRef] [PubMed]
- Šalamon, Š.; Bevc, S.; Ekart, R.; Hojs, R.; Potočnik, U. Polymorphism in the GATM Locus Associated with Dialysis-Independent Chronic Kidney Disease but Not Dialysis-Dependent Kidney Failure. Genes 2021, 12, 834. [Google Scholar] [CrossRef]
- Mohebi, R.; Liu, Y.; Hansen, M.K.; Yavin, Y.; Sattar, N.; Pollock, C.A.; Butler, J.; Jardine, M.; Masson, S.; Heerspink, H.J.L.; et al. Insulin growth factor axis and cardio-renal risk in diabetic kidney disease: an analysis from the CREDENCE trial. Cardiovasc. Diabetol. 2023, 22, 176. [Google Scholar] [CrossRef]
- Kempson, S.A.; Zhou, Y.; Danbolt, N.C. The betaine/GABA transporter and betaine: roles in brain, kidney, and liver. Front. Physiol. 2014, 5, 159. [Google Scholar] [CrossRef]
- Dittmann, K.; Wallaschofski, H.; Rettig, R.; Stracke, S.; Endlich, K.; Völzke, H.; Nauck, M.; Friedrich, N. Association between serum insulin-like growth factor I or IGF-binding protein 3 and estimated glomerular filtration rate: results of a population-based sample. BMC Nephrol. 2012, 13, 169. [Google Scholar] [CrossRef]
- van der Veen, Y.; Post, A.; Kremer, D.; Koops, C.A.; Marsman, E.; Appeldoorn, T.Y.J.; Touw, D.J.; Westerhuis, R.; Heiner-Fokkema, M.R.; Franssen, C.F.M.; et al. Chronic Dialysis Patients Are Depleted of Creatine: Review and Rationale for Intradialytic Creatine Supplementation. Nutrients 2021, 13, 2709. [Google Scholar] [CrossRef] [PubMed]
- Pasala, S.; Carmody, J.B. How to use… serum creatinine, cystatin C and GFR. Arch. Dis. Child. Educ. Pract. Ed. 2017, 102, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Alves, C.R.; Ferreira, J.C.; de Siqueira-Filho, M.A.; Carvalho, C.R.; Lancha, A.H., Jr.; Gualano, B. Creatine-induced glucose uptake in type 2 diabetes: a role for AMPK-α? Amino Acids. 2012, 43, 1803–1807. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Chemical structure of creatine. Source: PubChem (CID: 586).
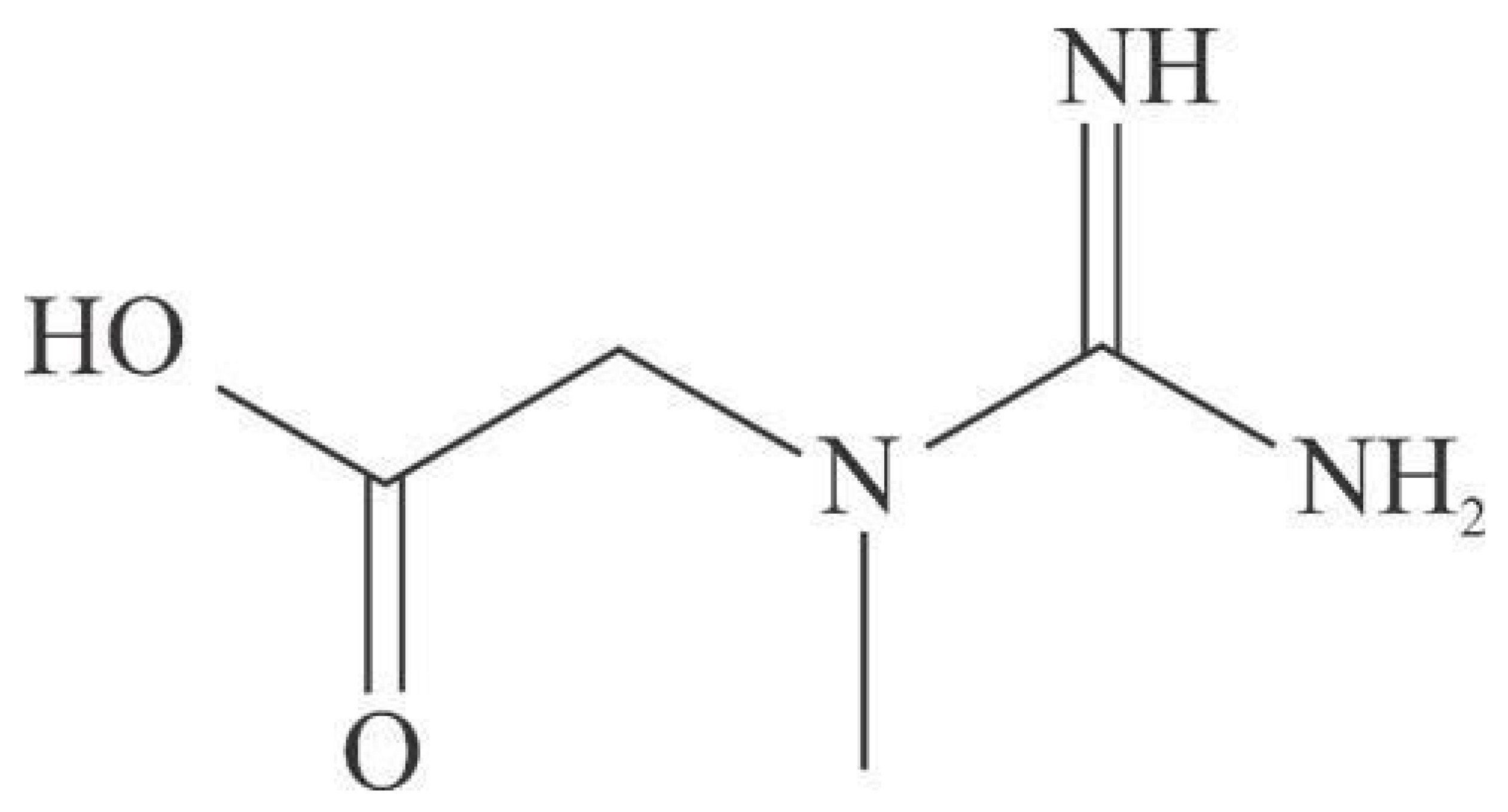
Figure 2.
Chemical-Protein Interaction Network (Green) and Protein-Protein Interaction (Gray) Network of Creatine Obtained by STITCH.
Figure 2.
Chemical-Protein Interaction Network (Green) and Protein-Protein Interaction (Gray) Network of Creatine Obtained by STITCH.
Figure 3.
Number of associations Gene-DiseaseID (MESH - Medical Subject Headings) from the creatine-related genes from Stitch and CTD databases. Names on bars indicate the Medical Subject Headings associated with each DiseaseID.
Figure 3.
Number of associations Gene-DiseaseID (MESH - Medical Subject Headings) from the creatine-related genes from Stitch and CTD databases. Names on bars indicate the Medical Subject Headings associated with each DiseaseID.
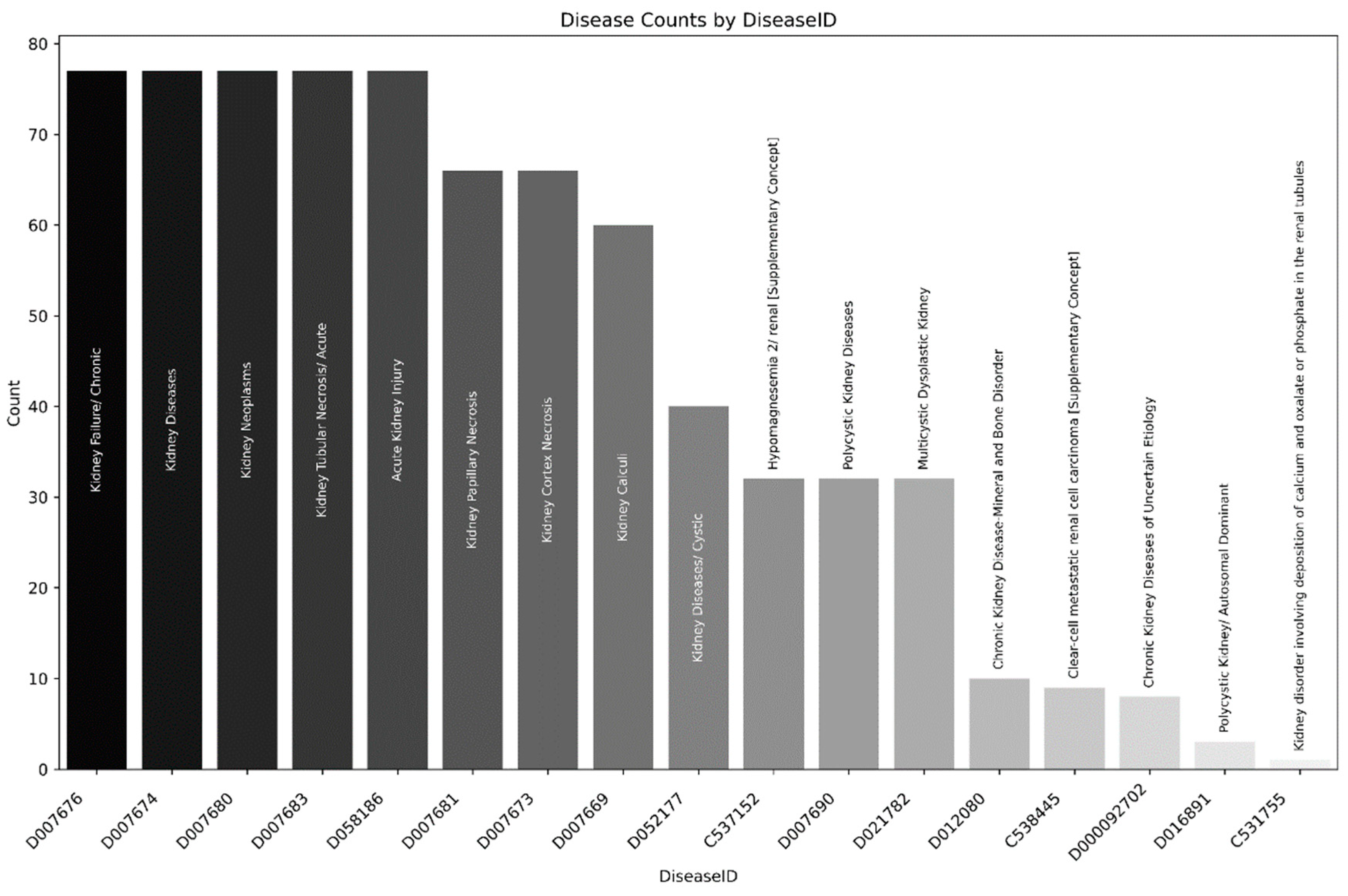
Figure 5.
Gene Enrichment Analysis from EnrichR-Kg using the creatine-related gene list against the following databases: (A) ReactomeDB; (B) Gene Ontology; and (C) KEGG terms. Green circles represent the genes. The first ten results are represented, using p-values ≤ 0.05 as the threshold.
Figure 5.
Gene Enrichment Analysis from EnrichR-Kg using the creatine-related gene list against the following databases: (A) ReactomeDB; (B) Gene Ontology; and (C) KEGG terms. Green circles represent the genes. The first ten results are represented, using p-values ≤ 0.05 as the threshold.
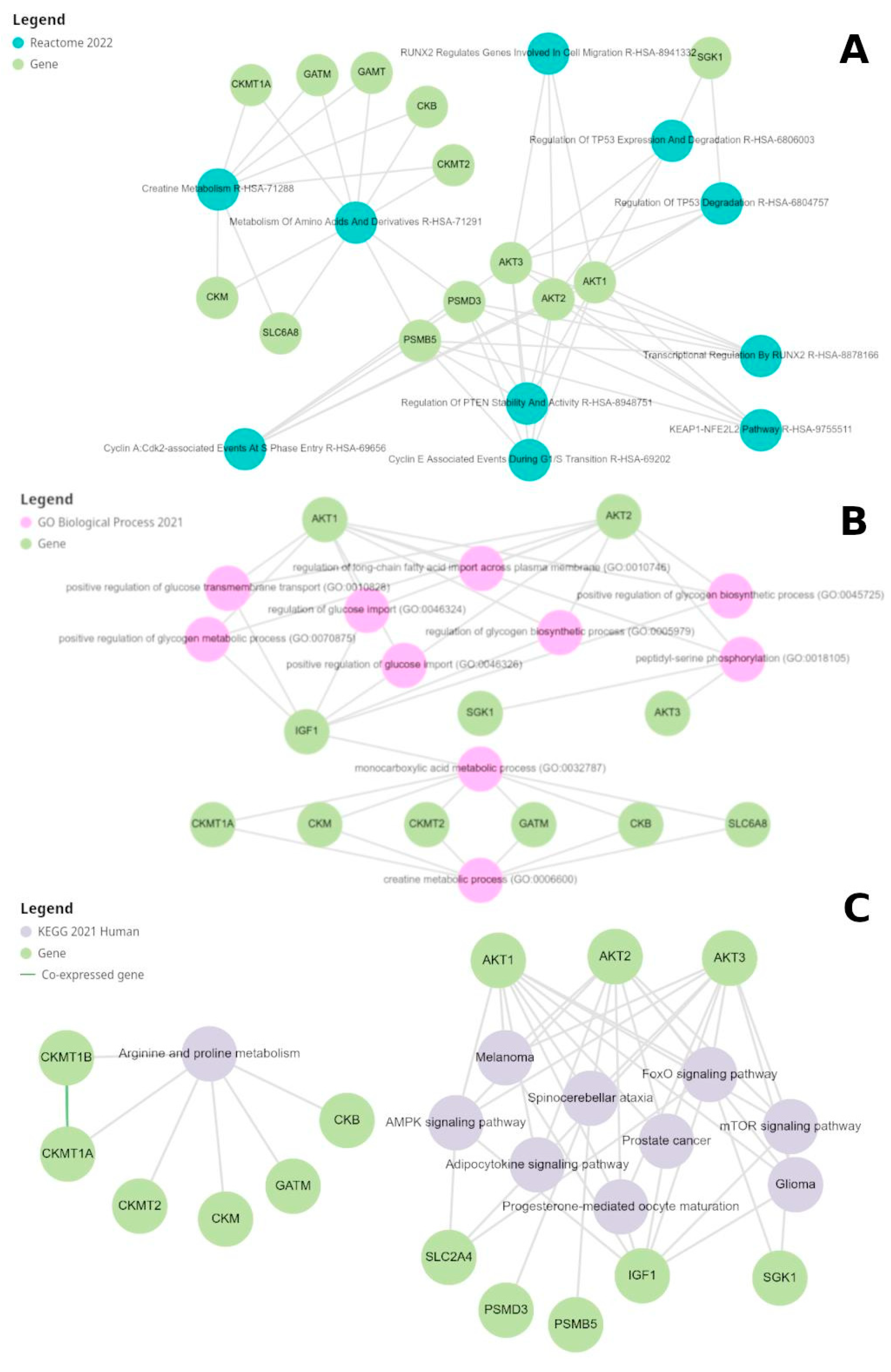
Figure 6.
Expression of creatine-related genes and the kinases retrieved from gene enrichment analysis against kinase-specific databases. (TPM - Transcripts per million bases).
Figure 6.
Expression of creatine-related genes and the kinases retrieved from gene enrichment analysis against kinase-specific databases. (TPM - Transcripts per million bases).
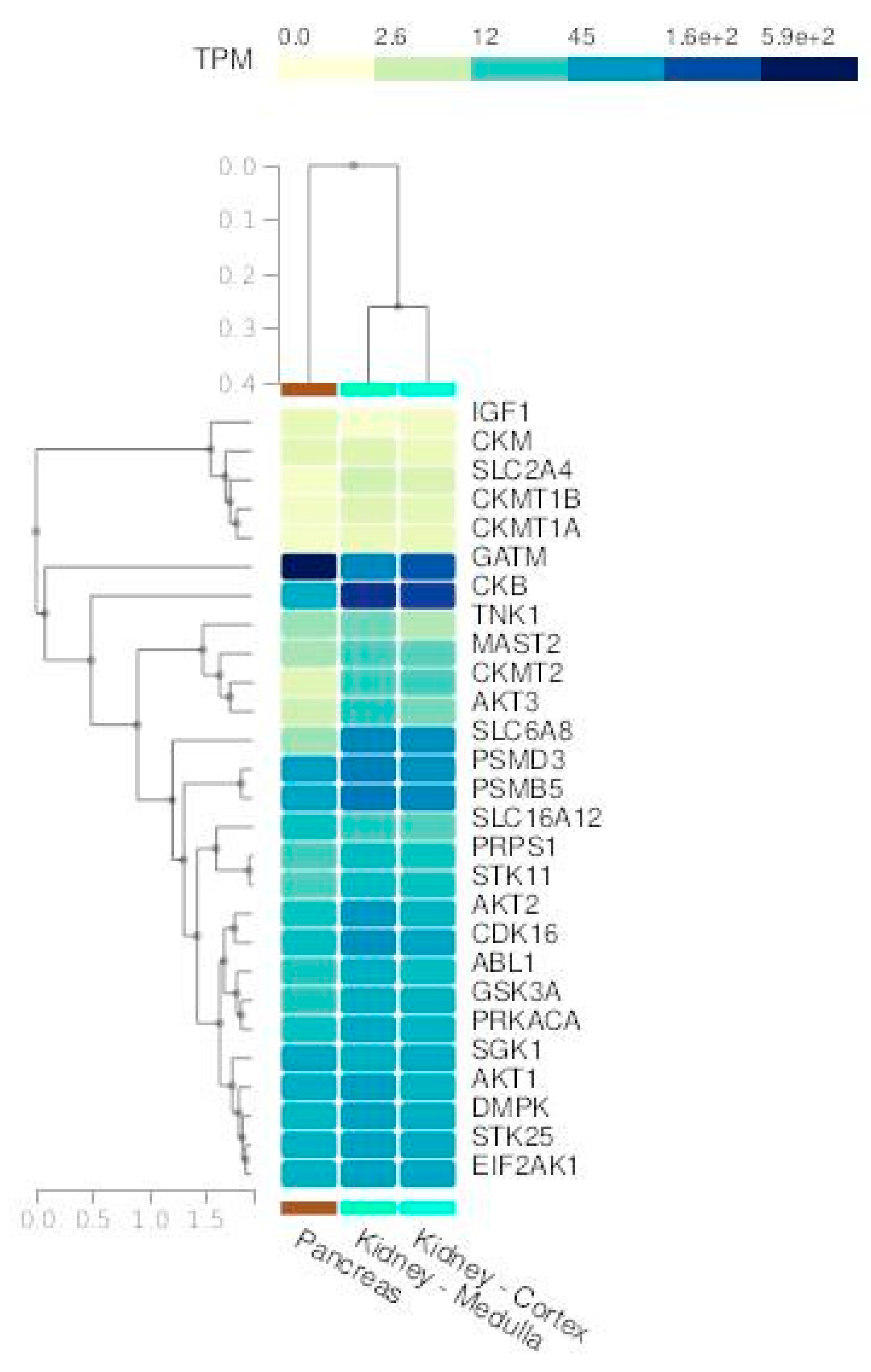
Figure 7.
Heatmap with the expression of creatine-related genes, from the study of Neusser et al. [71]. (GEO Accession number: GDS3712). The samples codes and gene symbols are represented horizontally and vertically, respectively. Samples states (TN - Control; NSC - Nephrosclerosis) are represented in the first colored line. The following cutoffs were used P-values < 0.05 and Log2Fold Change±1.
Figure 7.
Heatmap with the expression of creatine-related genes, from the study of Neusser et al. [71]. (GEO Accession number: GDS3712). The samples codes and gene symbols are represented horizontally and vertically, respectively. Samples states (TN - Control; NSC - Nephrosclerosis) are represented in the first colored line. The following cutoffs were used P-values < 0.05 and Log2Fold Change±1.
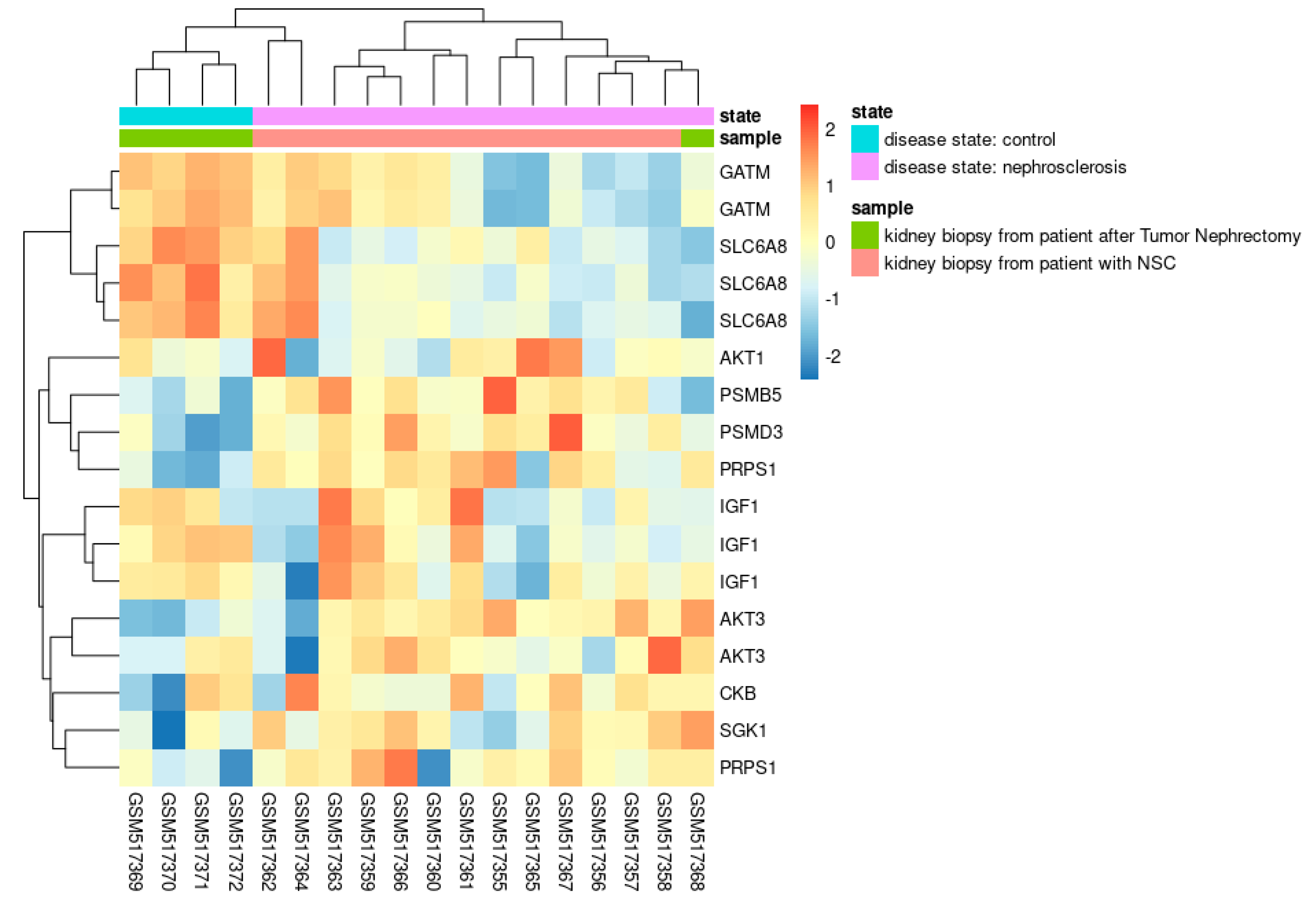
Figure 8.
Expression analyses of the creatine-related genes from transplanted patients kidney biopsies (dataset GDS724, Flechner et al. 2004). (A) Acute-rejection transplant (AR) versus Control samples. (B) Well-functioning transplant (TX) versus Control samples. (C) Well-functioning (TX) versus acute-rejection transplant samples (AR). (D) Well-functioning (TX) versus dysfunctional non-rejected transplant samples (NR). The plot shows the log2 fold change for the genes that met the significance threshold in each comparison (P<0.05). The error bars represent the standard deviation of log2 fold change for genes with multiple measurements (distinct transcripts or probes for the same gene). This provides a measure of variability in the fold change estimate. Printed numbers in each bar describe the p-values.
Figure 8.
Expression analyses of the creatine-related genes from transplanted patients kidney biopsies (dataset GDS724, Flechner et al. 2004). (A) Acute-rejection transplant (AR) versus Control samples. (B) Well-functioning transplant (TX) versus Control samples. (C) Well-functioning (TX) versus acute-rejection transplant samples (AR). (D) Well-functioning (TX) versus dysfunctional non-rejected transplant samples (NR). The plot shows the log2 fold change for the genes that met the significance threshold in each comparison (P<0.05). The error bars represent the standard deviation of log2 fold change for genes with multiple measurements (distinct transcripts or probes for the same gene). This provides a measure of variability in the fold change estimate. Printed numbers in each bar describe the p-values.
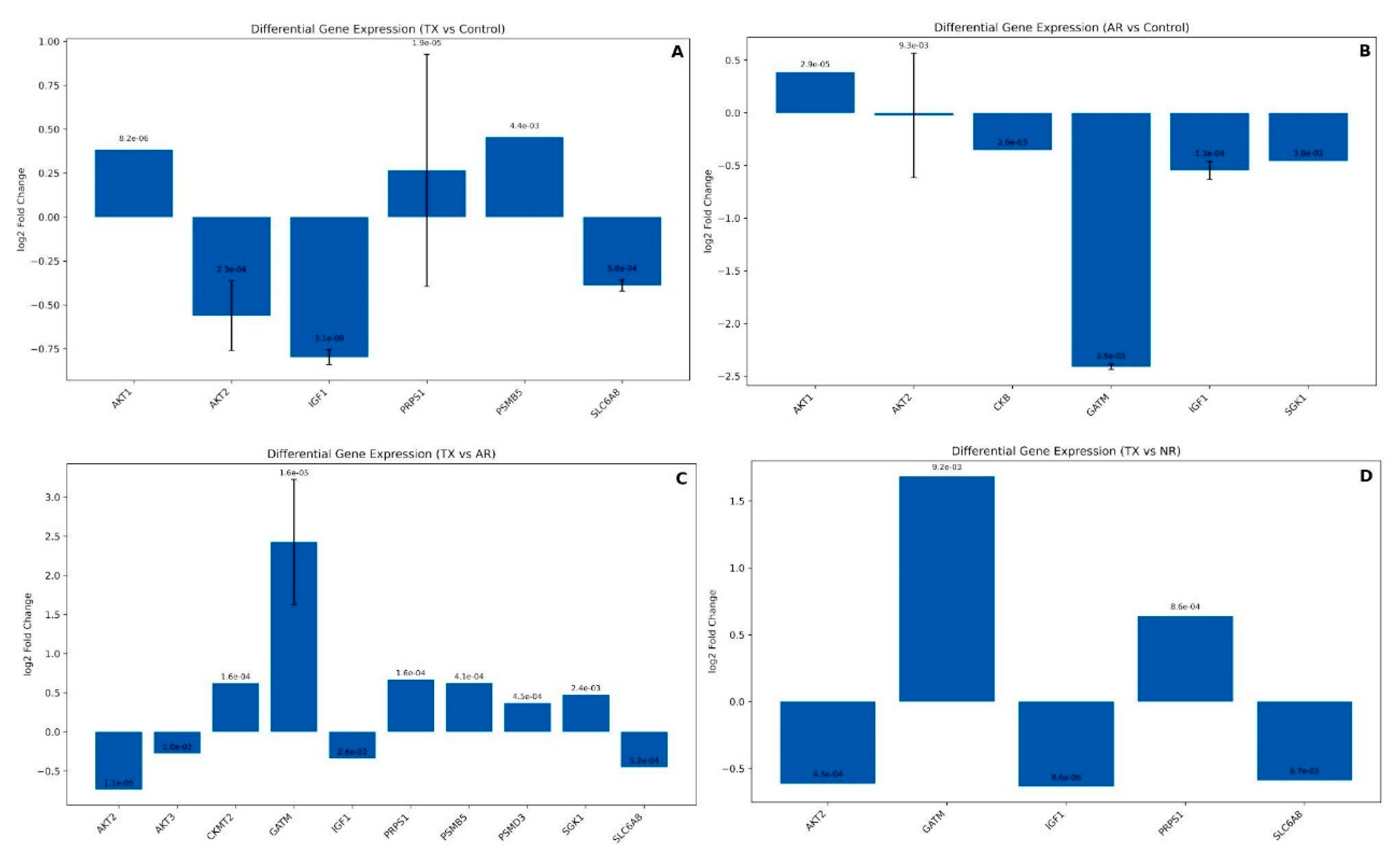
Figure 9.
Expression analyses of the creatine-related genes transcriptional profile of two stages of Clear-Cell Renal cell Carcinomas (cRCC), paired with controls [76,77] (GEO Accession Number: GDS2880) (A) Stage I cRCC versus Stage I Control samples. (B) Stage II cRCC versus Stage II Control samples. The comparisons of Stage I versus Stage II, within normal and tumor samples, did not retrieve differential expression of the creatine-related genes. The plot shows the log2 fold change for the genes that met the significance threshold in each comparison (P<0.05). The error bars represent the standard deviation of log2 fold change for genes with multiple measurements (distinct transcripts or probes for the same gene). This provides a measure of variability in the fold change estimate. Printed numbers in each bar describe the p-values.
Figure 9.
Expression analyses of the creatine-related genes transcriptional profile of two stages of Clear-Cell Renal cell Carcinomas (cRCC), paired with controls [76,77] (GEO Accession Number: GDS2880) (A) Stage I cRCC versus Stage I Control samples. (B) Stage II cRCC versus Stage II Control samples. The comparisons of Stage I versus Stage II, within normal and tumor samples, did not retrieve differential expression of the creatine-related genes. The plot shows the log2 fold change for the genes that met the significance threshold in each comparison (P<0.05). The error bars represent the standard deviation of log2 fold change for genes with multiple measurements (distinct transcripts or probes for the same gene). This provides a measure of variability in the fold change estimate. Printed numbers in each bar describe the p-values.
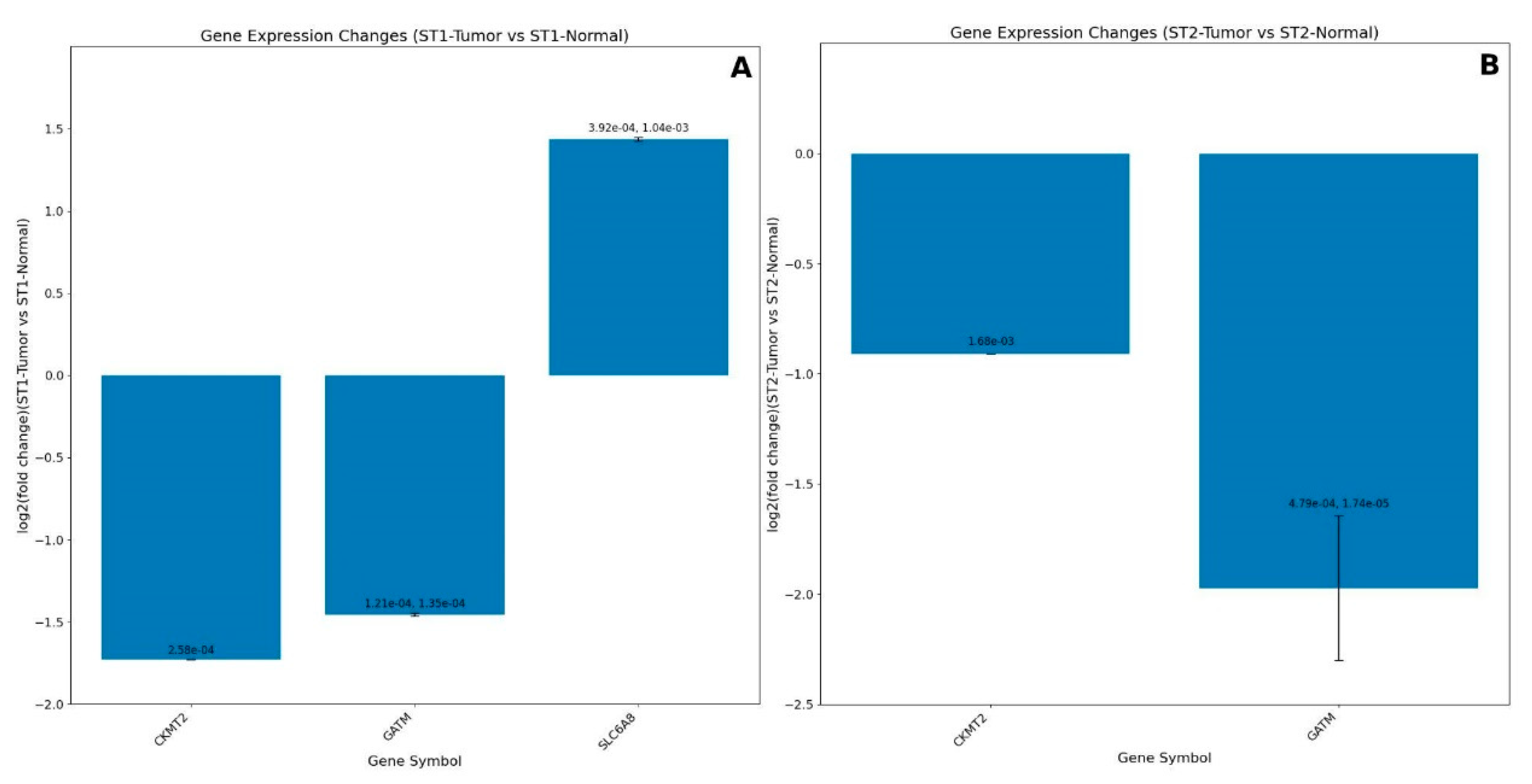
Figure 10.
Expression analyses of the creatine-related genes transcriptional profile from kidney fibrosis biopsies (GEO Accession Number: GSE137570) (A) All Fibrosis biopsy versus Normal samples. (B) Samples with Tubulointerstitial Fibrosis above 50% (Fibrosis>50%) versus Normal ones. (C) Tubulointerstitial Fibrosis below 50% (Fibrosis<50%) versus Normal samples. (D) Samples with eGFR higher than 50 (eGFR>50) versus Normal ones. (E) Samples with eGFR lower than 50 (eGFR<50) versus Normal ones. (F) Comparisons between eGFR<50 and eGFR>50 samples. The plot shows the Log2 fold change for the genes that met the significance threshold in each comparison (P<0.05). Printed numbers in each bar describe the p-values.
Figure 10.
Expression analyses of the creatine-related genes transcriptional profile from kidney fibrosis biopsies (GEO Accession Number: GSE137570) (A) All Fibrosis biopsy versus Normal samples. (B) Samples with Tubulointerstitial Fibrosis above 50% (Fibrosis>50%) versus Normal ones. (C) Tubulointerstitial Fibrosis below 50% (Fibrosis<50%) versus Normal samples. (D) Samples with eGFR higher than 50 (eGFR>50) versus Normal ones. (E) Samples with eGFR lower than 50 (eGFR<50) versus Normal ones. (F) Comparisons between eGFR<50 and eGFR>50 samples. The plot shows the Log2 fold change for the genes that met the significance threshold in each comparison (P<0.05). Printed numbers in each bar describe the p-values.
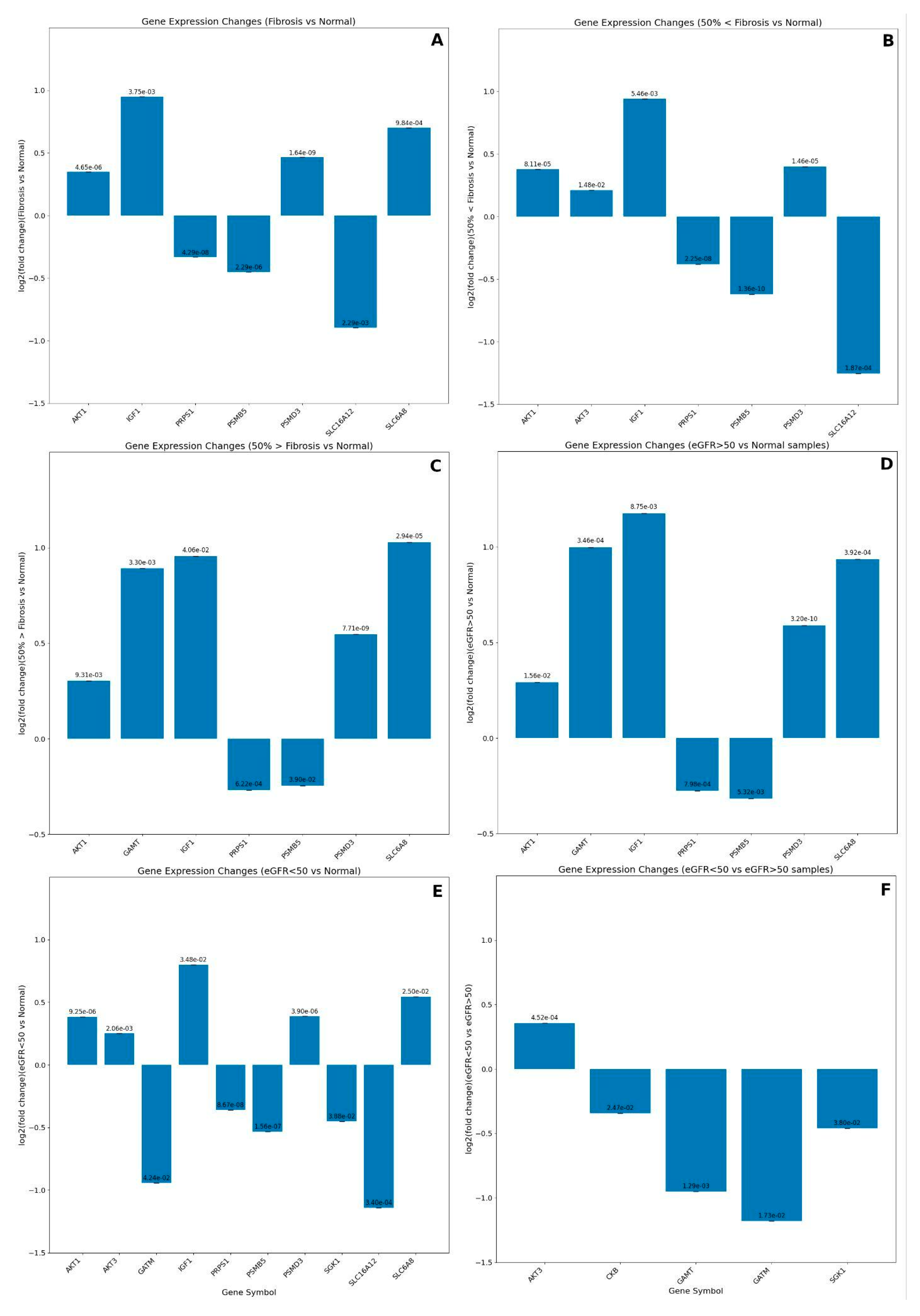
Table 3.
Compiled studies on pathological conditions and potential benefits of creatine. Adapted from [40].
Table 3.
Compiled studies on pathological conditions and potential benefits of creatine. Adapted from [40].
| Clinical conditions | Effect of creatine supplementation |
|---|---|
| GAMT deficiency; AGAT deficiency; Chronic Heart Failure, Myotonic dystrophy type II, Huntington’s disease, Dystrophinopathies, Sarcopenia | Most likely beneficial |
| Cancer, Mitochondrial cytopathies, Type II diabetes, Depression, Osteoarthritis, Osteoporosis, post-traumatic stress disorder, Alzheimer’s disease, Dyslipidemia, Traumatic brain injury, Charcot Marie-Tooth disease, Fibromyalgia, Myositis, Drug addiction, Parkinson’s disease | Possibly beneficial |
| HIV infection, Myotonic dystrophy type I, Creatine transporter deficiency, Chronic obstructive pulmonary disease, Facioscapulohumeral dystrophy, Amyotrophic lateral sclerosis, Schizophrenia | Unlikely beneficial |
Table 4.
Creatine-related gene list according to our search criteria. Gene short descriptions were retrieved from GeneCards database [65].
Table 4.
Creatine-related gene list according to our search criteria. Gene short descriptions were retrieved from GeneCards database [65].
| Gene Symbol (Homo sapiens, txid:9606) | Short description |
|---|---|
| SLC2A4 | Insulin-regulated facilitative glucose transporter GLUT4. |
| IGF1 | Insulin-like growth factor 1, a hormone involved in growth and development. |
| GATM | Glycine amidinotransferase, key in creatine biosynthesis. |
| GATM | Guanidinoacetate N-Methyltransferase, converts GAA in creatine. |
| SLC6A8 | Creatine transporter involved in transporting creatine into cells. |
| AKT1 | AKT serine-threonine protein kinase 1, involved in signaling pathways regulating cell growth and survival. |
| AKT2 | AKT serine-threonine protein kinase 2, involved in metabolism and insulin signaling. |
| AKT3 | AKT serine-threonine protein kinase 3, involved in brain development and function. |
| PSMB5 | A component of the 20S core proteasome complex involved in protein degradation. |
| PSMD3 | A non-ATPase subunit of the 26S proteasome involved in protein degradation. |
| SGK1 | Serum/glucocorticoid regulated kinase 1, involved in cellular stress response. |
| PRPS1 | Phosphoribosyl pyrophosphate synthetase 1, involved in purine metabolism and nucleotide biosynthesis. |
| CKB | Creatine kinase B, which plays a role in energy homeostasis by transferring phosphate between ATP and various phosphogens. |
| CKM | Creatine kinase M, involved in energy homeostasis and is a marker for myocardial infarction. |
| CKMT1A | Ubiquitous mitochondrial creatine kinase, involved in energy transduction and impaired in various diseases. |
| CKMT1B | Identical protein to CKMT1A and is also involved in energy transduction in tissues with large energy demands. |
| CKMT2 | Sarcomeric mitochondrial creatine kinase, involved in energy transduction in muscle tissues. |
| SLC16A12 | Transporter likely involved in monocarboxylic acid transport and associated with juvenile cataracts and renal glucosuria. |
Table 5.
Protein kinases directly and indirectly related to the creatine-related genes according to our search criteria. Gene short descriptions were retrieved from GeneCards database [65].
Table 5.
Protein kinases directly and indirectly related to the creatine-related genes according to our search criteria. Gene short descriptions were retrieved from GeneCards database [65].
| Kinase Gene Symbol | Short description (Source: GeneCards) |
|---|---|
| CAMKK2 | Calcium/calmodulin-dependent protein kinase kinase 2, involved in various signaling pathways. |
| MAP2K2 | Mitogen-activated protein kinase kinase 2, a key component in the MAPK/ERK pathway. |
| VRK1 | Serine/threonine-protein kinase VRK1, involved in cell cycle and nuclear envelope formation. |
| PDK4 | Pyruvate dehydrogenase kinase isozyme 4, regulating glucose and fatty acid metabolism. |
| PLK4 | Polo-like kinase 4, a regulator of centriole duplication in cell cycle progression. |
| PDK1 | Pyruvate dehydrogenase kinase 1, regulating glucose metabolism by phosphorylating pyruvate dehydrogenase. |
| DMPK | Myotonic dystrophy protein kinase, involved in muscle, heart, and brain function. |
| AAK1 | AP2 associated kinase 1, involved in clathrin-mediated endocytosis. |
| TBK1 | TANK-binding kinase 1, central in innate immune response and inflammation. |
| GAK | Cyclin G-associated kinase, involved in cell cycle regulation and clathrin-mediated endocytosis |
| NEK7 | Serine/threonine-protein kinase involved in mitotic cell cycle progression and cytokinesis. |
| PBK | Serine/threonine-protein kinase involved in mitotic cell cycle and DNA damage response. |
| BMP2K | Kinase implicated in bone morphogenic protein signaling and osteoblast differentiation. |
| MASTL | Kinase that regulates mitosis entry and maintenance which is involved in cell cycle checkpoint recovery. |
| VRK2 | Kinase involved in cell cycle regulation and neuronal apoptosis. |
| STK25 | Kinase that regulates stress response and cell migration. |
| CDK16 | Kinase involved in vesicle-mediated transport and exocytosis. |
| MAST2 | Kinase that regulates spermatid differentiation and interleukin-12 production. |
| PRKACA | Catalytic subunit of protein kinase A, involved in various cellular processes including metabolism and memory. |
| ABL1 | Tyrosine-protein kinase involved in cell differentiation, division, adhesion, and stress response. |
| STK11 | Serine/threonine kinase 11, also known as liver kinase B1 (LKB1), which regulates cell polarity and functions as a tumor suppressor. |
| TNK1 | Tyrosine kinase non-receptor 1, involved in negative regulation of cell growth and acting as a tumor suppressor. |
| EIF2AK1 | Eukaryotic translation initiation factor 2-alpha kinase 1, which plays a role in the cellular response to stress by phosphorylating the alpha subunit of the eukaryotic translation initiation factor 2 (eIF2). |
| GSK3A | Glycogen synthase kinase 3 alpha, involved in energy metabolism, neuronal cell development, and body pattern formation. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



