Submitted:
24 June 2024
Posted:
25 June 2024
You are already at the latest version
Abstract
Animals use dietary lipids to sustain their growth and survival. Insects can synthesize fatty acids (FAs) and are autotroph for a number of lipids, but auxotroph for specific lipids classes (e.g. sterols, polyunsaturated FAs). Once ingested, lipids are hydrolyzed in the intestinal lumen and taken up into intestinal cells within specific regions of the insect digestive tract. These lipids can be either stored in the intestinal cells or exported through the haemolymph circulation to specific organs. In this chapter, we describe the various lipids provided by insect diets, their extracellular hydrolysis in the gut lumen and their intake and metabolic fate in the intestinal cells. This review emphasizes the critical role of the digestive tract and its regionalization in processing dietary lipids prior to their transfer to the requiring tissues.
Keywords:
Digestion
; Enterocyte
; Microbiota
; Lipase
; Hydrolysis
; Emulsifier
; Lipid transporter
; Membrane.
Insects colonize a great variety of ecological niches on Earth and, thus, their survival relies on diverse feeding diets. Dietary macromolecules provided by the environment are used to produce energy and building blocks required to sustain developmental growth and reproduction, and to maintain homeostasis. The digestive tract is the first organ exposed to these external macromolecules. It must take in charge several essential functions, including protection against pathogens, neutralization of toxic compounds, extra-cellular digestion of macromolecules into assimilable nutrients, and absorption and efflux of these nutrients into the haemolymph (the insect open circulatory system). In addition, intestinal cells can fulfil storage and further enzymatic modifications prior to the distribution of nutrients throughout the overall body to the needing organs. Therefore, the digestive tract plays a major role in coordinating local and systemic homeostasis (Zhao and Karpac 2020). Mechanistically, the food bolus progresses through specialized compartments that process specific nutrients. The diversity of insect species and their broad diet spectrum is associated with a large variety of digestive tract anatomical structures favoring nutrient breakdown and assimilation (Terra and Ferreira 2020). Nonetheless, despite this huge diversity, the nutrient requirements and the digestive tracts of insects exhibit conserved functional and structural properties.
Enzymatic digestions of carbohydrates, proteins and lipids have been investigated in several insect species for more than a century (Plateau 1876) to understand how the basic nutrients are produced and assimilated. Food provides essential and dispensable lipids, which are enzymatically processed in the gut lumen and taken up by the enterocytes. In this chapter, we will first provide an overview of the insect digestive tract and discuss essential dietary lipids. Next, we will describe the lipid digestive processes in the gut lumen and the uptake of the resulting products by enterocytes. Finally, we will describe the lipid metabolic processes that take place in midgut cells. Digestive processes are tightly regulated by systemic controls, including endocrine signals from the brain and the fat body, which will not be addressed here. In this chapter, we will focus on lipid digestion, the dynamics of this process along the whole midgut and its essential role prior to the transfer of basic dietary lipids into the body internal milieu.
1. The Insect Digestive Tract
1.1. Structure of the Digestive Tract
The diversity in the morphology of the alimentary canal strongly depends on the taxon rather than on the insect feeding regimen (Santos et al. 2017; Terra and Ferreira 2020). Nonetheless, irrespective of the phylogeny, the insect midgut always comprises three major regions: the foregut, the midgut and the hindgut (Chapman et al. 2013) (Figure 1A). The foregut and the hindgut are of ectodermal origin and are covered by a cuticle facing the intestinal lumen. The midgut is of endodermal origin and is protected by a thin mucus layer and, for most species, by another structure called the peritrophic matrix (PM, also called peritrophic membrane) (Terra 2001). From the anterior to the posterior ends, the foregut comprises the mouth, the pharynx connected to the salivary glands, the esophagus, the crop in which the food bolus is transiently stored and the proventriculus that controls the entry of food into the midgut. The midgut is subdivided in an anterior, a median and a posterior region. Genomic and morphological analyses revealed that the midgut of Drosophila melanogaster contains five regions, which can be further divided into up to 14 sub-regions (Buchon et al. 2013; Dutta et al. 2015; Marianes and Spradling 2013). In most species, the anterior midgut is connected to gastric caeca (Terra and Ferreira 2020). The hindgut contributes to water reabsorption and feces formation. It comprises the ileum connected to the Malpighian tubules, which fulfil excretory functions, the colon, the rectum and the anus.
The midgut is the main organ of food digestion and nutrient absorption, which are facilitated by the presence of a PM and the absence of an external cuticle. The PM is a chitin-based lattice of proteoglycans and proteins. Depending on the insect species, the PM is produced either by the proventriculus or by delamination from the intestinal cells along the whole midgut (Hegedus et al. 2009; Lehane 1997). Hemipterans and thysanopterans do not have a PM but a lipoproteic perimicrovillar membrane that fulfils a similar function (Ribeiro et al. 2014). The PM compartmentalizes the midgut lumen into an endoperitrophic space, the PM itself and an ectoperitrophic space (Figure 1A). Depending on their physicochemical properties, the digestive enzymes are either attached to the epithelium membrane, free in the ectoperitrophic or endoperitrophic spaces, or framed within the PM (Hegedus et al. 2009; Hu et al. 2012). Analysis of the digestive enzymes that cross the larval PM of Spodoptera frugiperda suggested that the PM pore diameter is about 7.5-8 nm (Ferreira et al. 1994). As a result, macromolecules and undigested materials remain in the endoperitrophic space, whereas small molecules and nutrients can pass through, for eventual digestion and absorption by the intestinal cells (Bolognesi et al. 2008). The PM allows the establishment of bidirectional fluxes along the midgut: contractions of the proventriculus pushes the food bolus into the midgut generating an anterior/posterior flux inside the endoperitrophic space (Peller et al. 2009; Woodring and Lorenz 2007), while fluid secretions by the Malpighian tubules exerts a counter pressure within the ectoperitrophic space (Biagio et al. 2009) (Figure 1A). These opposing fluxes favor the mixing of foods to be digested and the recycling of the digestive enzymes.
1.2. Cell Types and Their Functions
The midgut is a monolayer epithelium supported by a basal lamina with the apical cell side facing the intestinal lumen (Caccia et al. 2019). This epithelium is surrounded by striated muscles and connected to fine-branched tracheoles and nerves (Miguel-Aliaga et al. 2018) (Figure 1B). Pluripotent intestinal stem cells (ISCs), located at the basis of the epithelium, ensure a constant homeostatic renewal of intestinal cells (Hakim et al. 2010; Zwick et al. 2019). ISCs give rise to either enteroblasts or enteroendocrine progenitors that differentiate into enterocytes or enteroendocrine cells, respectively (Zeng and Hou 2015).
The functions of enteroendocrine cells have been studied in several insect species (Reiher et al. 2011; Wegener and Veenstra 2015), but elucidation of their activity mostly relies on D. melanogaster studies (Issaq et al. 2009). These cells secrete various peptidic hormones that systemically control appetite and homeostasis (Veenstra 2009; Veenstra et al. 2008). They also act locally to control midgut peristalsis (LaJeunesse et al. 2010) and lipid metabolism (Song et al. 2014).
The enterocytes, which should formally be called columnar cells (Caccia et al. 2019), are the most abundant cell type in the midgut and display a characteristic organization that accounts for their secretive and/or absorptive functions (Azevedo et al. 2009; Fialho et al. 2013; Lehane 1977; Rost-Roszkowska et al. 2007; Santos et al. 2017; Teixeira et al. 2013). Columnar cells mainly specialized in enzyme secretion contain in their apical cytosol rough endoplasmic reticulum (ER) and granules, and display apical microvilli (de Sousa et al. 2009). Release of vesicles containing digestive enzymes may occur through various types of secretion systems, namely holocrine, apocrine and merocrine (Fialho et al. 2013; Fuzita et al. 2019; Silva et al. 2013). During holocrine secretion, secretory vesicles and the cytoplasm are released into the gut lumen by rupture of the plasma membrane, whereas in apocrine secretion, a variable part of the apical cytoplasm containing vesicles is released by membrane budding. During merocrine secretion, vesicles fuse with the apical membrane and release their content into the gut lumen without membrane and cytoplasm loss. Conversely, columnar cells mainly specialized in absorption exhibit a dense apical microvillar structure, basal membrane infolds and accumulate lipid droplets (LDs) depending on diet, gut region and insect species. These subcellular structures as well as the membrane composition greatly vary along the anterior-posterior axis of the midgut, suggesting different nutrient absorption and enzyme secretion abilities, although the existence of populations of columnar cells strictly specialized in either function has not been formally demonstrated (Caccia et al. 2019; Terra et al. 2006).
The pH greatly varies along the intestinal tract depending on gut regions and insect species (Harrison 2001; Terra and Ferreira 2020). In various species, restricted regions display extreme pH values; for instance, in the median region of the midgut of some Scarabaeidae, alkaline pH can reach 12, which favors lipid saponification (Lehane 1977) and lipase activity (Canavoso et al. 2001). The precise regulation of the luminal pH depends on active acid-base secretion processes across epithelia, including an H+-V-ATPase potassium/proton exchanger (Harrison 2001). Extreme pH values appear to rely on specialized cells that are specific to certain insect species (Caccia et al. 2019). Goblet cells mostly studied in lepidopteran larvae, but also reported in ephemeropterans, plecopterans and trichopterans, display an invagination of their apical membrane that forms a cavity (Cioffi 1984). Active ion exchanges in this cavity generate an alkaline pH in the gut lumen, which favors the neutralization of toxic compounds present in the plant material eaten by phytophagous lepidopteran larvae. Copper cells found in brachycerous dipterans generate an acidic pH in a restricted region of the median midgut, thereby favoring the killing of luminal bacteria (Dubreuil 2004; Storelli et al. 2018). In sum, the whole lumen environment depends on exogenous and endogenous parameters, which determines the final digestive abilities of each insect species.
2. Essential and Non-Essential Dietary Lipids
2.1. Dietary Lipids
Insects find in their food a great variety of lipids, including those essential for survival and development that they are unable to synthesize. The main dietary lipids are triacylglycerols (TAGs), phospholipids, galactolipids, sulpholipids and sterols. TAGs and phospholipids are present in several types of foods. Galactolipids (mono- and di-galactosyl diacylglycerols, MGDG and DGDG respectively) (Figure 2A) and sulpholipids (sulfated glycolipids), the most abundant membrane lipids of chloroplasts, represent up to 80% of total lipids of leaf cells consumed by folivorous insects (Sahaka et al. 2020). Depending on their diet, insects ingest sterols as phytosterol from plants, cholesterol from animals or ergosterol from fungi (Jing and Behmer 2020).
Only a few studies investigated the need for fat-soluble vitamins; they are not metabolized in the midgut but directly transferred to the requiring organs. Experimental strategies to determine the essential need of a given lipid are based on either controlling the food composition or feeding radiolabeled precursors to monitor their incorporation in final products (Canavoso et al. 2001). In this way, it has been shown that insects are unable to synthesize sterols, most of them being also disabled for polyunsaturated fatty acids (PUFAs) synthesis.
Only a few studies investigated the need for fat-soluble vitamins; they are not metabolized in the midgut but directly transferred to the requiring organs. Experimental strategies to determine the essential need of a given lipid are based on either controlling the food composition or feeding radiolabeled precursors to monitor their incorporation in final products (Canavoso et al. 2001). In this way, it has been shown that insects are unable to synthesize sterols, most of them being also disabled for polyunsaturated fatty acids (PUFAs) synthesis.
2.2. Essential Fatty Acids
Although most insect diets contain TAGs, insects are able to synthesize long chain fatty acids (LCFAs), the monounsaturated palmitoleic (C16:1) and oleic (C18:1) acids, and TAGs (Garrido et al. 2015). In contrast, they are classically considered to be auxotrophs for the polyunsaturated linoleic (C18:2) and α-linolenic (C18:3) acids (Figure 2B). Nevertheless, Blomquist and colleagues have shown that a few insect species, including the cockroach Periplaneta americana and the house cricket Acheta domesticus, are autotrophs for linoleic but not α-linolenic acid (Blomquist et al. 1991). Linoleic acid is the precursor of arachidonic acid (AA, C20:4), whereas α-linolenic acid is the precursor of docosahexanoic (DHA, C22:6) and eicosapentanoic (EPA, C20:5) acids (Figure 2B), both pathways requiring the same metabolic enzymes (Stark et al. 2008). Surprisingly, D. melanogaster is disabled for AA, EPA and DHA synthesis (Shen et al. 2010), a default likely shared by mosquitoes that require dietary AA or EPA to sustain their larval development (Dadd and Kleinjan 1979; Merritt et al. 1992). Oxidation of AA directs the synthesis of eicosanoids (e.g., prostaglandins), appearing to be important for juvenile development, immune response and reproductive functions in insects (Kim and Stanley 2021). The specific requirements of EPA and DHA have been investigated in D. melanogaster, and revealed that feeding an EPA/DHA mixture is neuroprotective against intoxication with the Paraquat insecticide (de Oliveira Souza et al. 2019).
The phenotypes induced by PUFA deficiency differ between insect species. In the locusts Schistocerca gregaria and Locusta migratoria, this deficiency results in impaired adult emergence (Dadd 1961), whereas in other species it may result in milder phenotypes, including wing expansion defect and reduced reproductive capacity (Beenakkers et al. 1985). In some coleopteran, dipteran and lepidopteran species, phenotypes may appear only after two generations of deprivation, suggesting that the very low amounts provided by the oocyte stores are sufficient to sustain their critical functions (Canavoso et al. 2001; Stanley-Samuelson 1994).
2.3. Sterol Requirement
Insects are unable to synthesize sterols (Clark and Block 1959; Svoboda 1999), although for a few species, sterols are provided by intestinal symbionts (Clayton 1964; Jurzitza 1974). Cholesterol is essential i) as a membrane component present at high levels in lipid rafts (Carvalho et al. 2010; Rietveld et al. 1999), ii) as a precursor of the molting ecdysteroid hormones (Rewitz et al. 2006; Yamanaka et al. 2013), and iii) as a Hedgehog conjugate required for signaling efficiency (Cooper et al. 2003). The need for dietary sterols for insect development was first discovered in the blow-fly Lucilia sericata (Hobson 1935); this requirement was next extended to all investigated insect species (Canavoso et al. 2001; Jing and Behmer 2020). Deficiency in dietary sterols results in defects in developmental growth, molting transitions and reproduction, although maternally provided sterol stores may be sufficient to support early developmental processes, as reported for the first and second larval stages of D. melanogaster (Fluegel et al. 2006).
Sterol synthesis requires the condensation of two farnesylpyrophsophate units and its reduction to squalene (Figure 2C); next, cyclization and oxidation of squalene leads to the formation of lanosterol, which after further oxidation steps results in cholesterol. Insects can synthesize the sesquiterpene intermediate farnesylpyrophsophate in the biosynthetic pathway of the juvenile hormone (Noriega 2014). In contrast, insects lack the enzymes required for squalene, lanosterol and cholesterol synthesis (Zhang et al. 2019). These final metabolic steps demand a high number of oxygen molecules. Given their blind tracheal system limiting oxygen delivery to the needing tissues, it has been proposed that the loss of such a high oxygen-consuming pathway may provide an evolutionary advantage to insects (Harrison et al. 2010; Zhang et al. 2019).
2.4. Carotenoids and Fat-Soluble Vitamins
Carotenoids are lipophilic isoprenoids divided into carotenes and xanthophylls, the latter result from the oxidation of the former. They constitute pigments that color the tegument and the eye photoreceptors of several insect species (Canavoso et al. 2001). Carotenoids were thought to be synthesized only by plants, fungi and microorganisms, but recent studies revealed that aphid and cecidomyiid genomes encode the enzymes needed for their synthesis (Cobbs et al. 2013; Moran and Jarvik 2010).
A few studies investigated the role of the fat-soluble vitamins A, D, E and K in insects. Vitamin A derives from β-carotene; its deprivation in D. melanogaster does not affect essential functions but results in blindness (Dewett et al. 2021). Three forms of vitamin K exist VK1, VK2 and VK3. It has been shown that a VK1 diet supplementation improves the developmental growth of the house cricket A. domesticus (McFarlane 1976). Studies in D. melanogaster revealed that VK2 acts as a mitochondrial electron carrier (Vos et al. 2012) and that feeding VK2 protects against mitochondrial dysfunctions in lymphoma and Alzheimer models (Dragh et al. 2017; Lin et al. 2021). The need for vitamin E (α-tocopherol) has been studied in a few insect species and shown to be essential for spermatogenesis in the house cricket A. domesticus and for female reproduction in Agria affinis (Fraenkel and Blewett 1946; House 1966; Meikle and McFarlane 1965). Vitamin D3 is a steroid whose deficiency increases DNA damage. In D. melanogaster, vitamin D3 supplementation reduces the formation of experimentally induced tumors (Vasconcelos et al. 2020). These requirements open the door to the search for strategies aimed at controlling insect pest propagation (Moadeli et al. 2020).
2.5. Microbiota and Dietary Lipids
The gut microbiota comprises all the microorganisms that colonize transiently or permanently the digestive tract. The gut microbiota has been shown to contribute to insect digestion, homeostasis, growth, detoxification and resistance to pathogens (Engel and Moran 2013; Grenier and Leulier 2020; Lee and Hase 2014; Schmidt and Engel 2021). The microbiota is essential for xylophagous insects, as described in termites, which ─when deprived of their anaerobic protists─ continue to feed on wood but die of starvation (Cleveland 1923). Lower termites are exclusive wood-feeders and their hindgut paunch hosts flagellates, bacteria and archaea, whereas higher termites have a more diverse diet but are devoid of flagellates (Brune and Dietrich 2015). Digestion of wood polysaccharides by these symbionts yields short-chain FAs (SCFAs), namely acetate, propionate and butyrate, which are taken up by the hindgut epithelium. Therefore, the hindgut paunch is referred to as a micro-bioreactor mostly providing acetate, which constitutes the major source of energy and anabolic metabolism of xylophagous termites (Brune 2014).
The digestive physiology of xylophagous but also of polyphagous cockroaches shares characteristic features with that of termites: i) their hindgut contains the highest bacterial density and is critical for metabolizing indigestible dietary compounds, and ii) these biochemical activities result in the production SCFAs, which are taken up by intestinal epithelial cells (Cruden and Markovetz 1987). Studies investigating the effect of diet in shaping the microbiota composition found variable results. One of those showed no significant change in the core microbial populations after shifting P. americana on a butter-based diet, suggesting that a selection pressure acts to maintain the specificity of the symbiosis between the insect and its microbiota (Tinker and Ottesen 2016). Volatile acids, namely acetate, propionate, butyrate, isobutyrate, valerate and isovalerate, are produced by microbiota-mediated digestion of cellulose; these compounds have been proposed to mediate social aggregation in several insect species (McFarlane and Alli 1985). In Blattella germanica, volatile FAs produced by the hindgut microbiota are excreted in the frass and are the main olfactory molecules mediating social aggregation (Wada-Katsumata et al. 2015).
In the honeybee Apis mellifera, social interactions elicit the colonization of the digestive tract by a conserved gut microbial community that comprises five core bacterial strains (Kwong et al. 2017). Pollen wall components resistant to midgut enzymes are metabolized to organic acids in the hindgut, where the bacterial density is the highest (Kesnerova et al. 2017). Honeybees devoid of microbiota exhibit reduced body and midgut weight through an insulin-dependent process (Zheng et al. 2017). These defects result from the lack of SCFAs produced from sugar fermentation by specialized bacteria in the hindgut.
The burying beetle Nicrophorus vespilloides (Coleoptera), which feeds its offspring on vertebrate carrions, excretes with their frasses bacteria and yeasts that preserve carcasses from putrefaction, but also secrete lipases to operate extracellular lipid digestion (Shukla et al. 2018; Vogel et al. 2017a). Lipase activities have also been reported in the bacteria of the anal droplets from another coleopteran, the weevil Cryptorhynchus lapathi (Jing et al. 2020).
In the velvetbean caterpillar (Lepidoptera) Anticarsia gemmatalis, antibiotic (tetracycline) treatment to dampen the gut microbiota slightly affected survival and development, indicating that gut bacteria are not essential for this species survival in laboratory setting. On the other hand, this antibiotic treatment strongly suppressed intestinal lipase activity, suggesting that the microbiota contributes to lipid digestion in velvetbean caterpillars (Visotto et al. 2009). Analysis of the gut bacteria of three other lepidopteran species, Antheraea assamensis, Helicoverpa armigera and Plutella xylostella, revealed that among 71 isolates, 46 exhibited lipolytic activity, suggesting that their microbiota likely play a role in lipid digestion (Gandotra et al. 2016).
The microbiota has been reported to modulate midgut lipid metabolism in several dipteran species. The Tsetse fly (Glossina pallidipes) and the sand fly (Lutzomyia longipalpis) are the vectors of Trypanosoma brucei and Leishmania infantum, respectively. In the Tsetse fly, midgut infection provokes a severe immune response to eliminate Trypanosoma brucei, while Leishmania infantum establishes a commensal association in the midgut sand fly. However, in both species, midgut infection induces lipid remodeling (Bateta et al. 2017; Coutinho-Abreu et al. 2020). Axenic mosquito Aedes aegypti larvae, lacking bacteria, cannot reach the critical size required for molting; this phenotype is associated with several metabolic perturbations, including changes in lipid metabolism (Valzania et al. 2018; Vogel et al. 2017b).
During the past two decades, the D. melanogaster model has been extensively used to investigate the physiological impact of gut microbiota, which, in the fruitfly, comprises only a few bacterial species. The two major strains, Acetobacter pomorum and Lactobacillus plantarum cooperate in a metabolic dialogue that sustains larval growth under nutritional stress (Consuegra et al. 2020a; Consuegra et al. 2020b). Axenic D. melanogaster larvae are viable and develop to adulthood, but are sensitive to food scarcity (Shin et al. 2011; Storelli et al. 2011). This phenotype may in part rely on digestion efficiency, as colonization of the midgut by commensal bacteria enhances, through the IMD/Relish pathway, the expression of digestive enzymes, including lipases (Erkosar and Leulier 2014). The mechanism of this regulation has been in part decrypted and initially involves the production of acetate by the commensal bacteria in the anterior midgut, which is the intestinal segment exhibiting the highest bacterial density (Jugder et al. 2021; Shin et al. 2011; Storelli et al. 2018). Acetate activates the IMD/Relish pathway in enteroendocrine cells, which in turn enhances the expression of tachykinin (Jugder et al. 2021; Kamareddine et al. 2018). Tachykinin is a secreted peptide that binds its cognate receptor at the enterocyte membrane to regulate the intracellular lipid storage/catabolism rate (Kamareddine et al. 2018; Song et al. 2014).
In summary, the insect microbiota strongly affects host homeostasis by providing additional nutrients and optimizing the digestion process. As to lipid metabolism, the microbiota stimulates the expression of host lipases and produces energy metabolites, in particular SCFAs. Acetate appears to play a central role, both as a carbon source for energy metabolism and as a signaling molecule that triggers a lipid homeostasis response. In human, acetate produced by the colon microbiota provides the most abundant energy source for enterocytes and protects against infections (Fukuda et al. 2011; Lee and Hase 2014), a topic that deserves further investigations in insects.
3. Lipid Hydrolysis in the Gut Lumen
3.1. Lipase Characterization
Early studies on lipid digestion were based on the usage of radiolabeled molecules and the analyses of the end-products after feeding or incubation with midgut extracts. Further experimentations were based on the separation of midgut sections and/or lumen gut fractions, and on the purification of enzymes. More recently, genomic and transcriptomic strategies allowed the identification of all the potential digestive enzymes and the analysis of their tissue-specific expression. Digestive enzymes have to hydrolyze dietary lipids, i.e. TAGs, phospholipids, galactolipids and sterols, the latter being possibly ingested as sterol conjugates. TAG hydrolases/lipases, commonly called lipases, cleave carboxylester bonds in TAGs, diacylglycerols (DAGs) and monoacylglycerols (MAGs), but may also be active on steryl-esters and phospholipids (Horne et al. 2009).
Lipases typically contain a Gly-X-Ser-X-Gly consensus sequence, where the Ser together with a His and an Asp/Glu residue form the tridimensional catalytic site (Derewenda 1994). When covered by a peptidic domain ─called the lid─ the catalytic site is not accessible to the substrate. Lipases are water-soluble but poorly active in aqueous solution, whereas at the interface of lipid micelles, a conformational rearrangement opens the lid so that the catalytic site becomes accessible to the substrate (Holmquist 2000). Depending on their primary structures, lipases are classified as neutral, acid, lipase2 (not reported in insects), lipase3, GDSL, hormone sensitive lipase (HSL) and adipocyte triglyceride lipase (ATGL) (Horne et al. 2009). In mammals, digestive lipases correspond to the families of gastric/acid lipases active at low pH, and of neutral/pancreatic lipases active at neutral pH, the activity of the latter depending on colipase specific binding at the C-terminal domain of the lipase (Miled et al. 2000). Congruent with the absence of colipase in insects, their neutral-lipases lack such a C-terminal domain (Sahaka et al. 2020).
TAG Lipases (EC 3.1.1.3) cleave ester bounds between FA carboxylic and glycerol-alcohol residues, although several of these lipases preferentially remove FA in position sn-1 and sn-3 of the glycerol, leading to sn-2 MAGs (Figure 3A). Phospholipases are defined in respect of their specific cleavage site. Phospholipases A1 (PLA1, EC 3.1.1.32) and A2 (PLA2, EC 3.1.1.4) catalyze the formation of lysophospholipids by removing FA at the sn-1 and sn-2 positions of glycerol, respectively (Figure 3B). Phospholipases B (PLB) cleave both sn-1 and sn-2 sites. Finally, phospholipases that hydrolyze the phosphate bonds located proximally or distally relative to the glycerol backbone are referred to as PLC (EC 3.1.4.3) and PLD (EC 3.1.4.4), respectively (Figure 3B). The lid domain and a β-loop located near the catalytic site are involved in substrate recognition; both long domains are characteristic of TAG lipases, whereas shorter domains are characteristic of phospholipases (Horne et al. 2009).
3.2. Insect Digestive Lipases
Insect genomes encode a high number of lipases, most of them belonging to the acid and neutral lipase families (Table 1). Interestingly, these lipases are more abundant in insects than in mammals (Horne et al. 2009), which may be the consequence of a high level of redundancy and/or of a broader diversity of dietary lipids. The high number of lipases together with their substrate flexibility renders difficult the precise physiological assignment of a specific substrate to a given lipase. Consequently, investigations aimed at deciphering the lipid digestion processes hardly rely on genetic approaches but rather on monitoring enzymatic activities of midgut extracts or of purified enzymes, and on analyzing gene expression in enterocytes.
In the insect midgut, a few studies suggest that TAG hydrolysis preferentially removes FA from the sn-1 and sn-3 positions of the glycerol, leading to sn-2 MAGs (Bollade et al. 1970; Hoffman and Downer 1979; Treherne 1958), while others rather suggest complete digestion to glycerol and free FAs (FFAs) (Table 2) (Canavoso et al. 2004; Tsuchida and Wells 1988; Weintraub and Tietz 1973). Given that alkaline pH favors the complete hydrolysis to glycerol and FFAs (Canavoso et al. 2001), these discrepancies may be the consequence of species variability in intestinal luminal pH (Terra and Ferreira 2020). It may also depend on experimental conditions and/or lipase affinity for specific lipid classes. For instance, midgut lipases of L. migratoria hydrolyze triolein more efficiently than tripalmitin (Weintraub and Tietz 1973), suggesting a higher affinity for unsaturated FAs, as reported for mammalian pancreatic lipases.
Phospholipase activity was first described in the midgut of several species of coleopterans, lepidopterans and dipterans (Somerville and Pockett 1976; Turunen 1988a; Turunen and Chippendale 1989; Turunen and Kastari 1979). To date, PLA2 are the most studied phospholipases (Nor Aliza et al. 2018; Nor Aliza and Stanley 1998; Rana et al. 1997; Sajjadian et al. 2019; Stanley et al. 1998; Uscian et al. 1995; Weiher and Komnick 1997), while only a few studies focused on digestive phospholipases A1, C and D (references in Table 2). Further, some of the enzymes referred to as PLA1 are actually galactolipases that hydrolyze preferentially the acyl residue at the sn-1 position of the glycerol backbone of galactolipids (Christeller et al. 2011; Sahaka et al. 2020).
Removal of phosphate groups is catalyzed by non-specific alkaline (EC 3.1.3.1) or acid (EC 3.1.3.2) phosphatases, defined by the optimal pH range of their activity. These phosphatases are active on a great variety of phosphomonoester compounds. Both acid and alkaline phosphatases have been described as soluble or membrane-bound forms in the midgut of several insect species (da Silva et al. 2019; Eguchi 1995; Matic et al. 2021; Terra et al. 1988).
Waxes are LCFA/long-chain-alcohol esters, which are structural components of most insect cuticles and, thus, are present in the insect-based diet of carnivorous insects (Table 2). It has been suggested that wax hydrolysis might be accomplished by intestinal symbionts, but a recent study revealed that the honeycomb moth Galleria mellonella exhibits a microbiota-independent digestive enzyme activity that hydrolyzes the wax ester bonds (Kong et al. 2019).
Dietary sterols are present in a free form or as steryl-ester or steryl-glycoside (Li and Jing 2020). Cholesterol ester hydrolase (EC 3.1.1.13) was first characterized in the beetle Trogoderma granarium (Agarwal and Nair 1976). In D. melanogaster, the lipase Magro acts as a sterol esterase in the midgut (Sieber and Thummel 2012). Phytosterols that contain an alkyl branching are not readily usable by phytophagous insects (Svoboda 1999). The efficiency of dealkylation varies, depending on the species (Jing and Behmer 2020). A few enzymes potentially required for phytosterol dealkylation have been identified in Helicoverpa zea (Jing et al. 2012), while a desmosterol reductase that catalyzes the final step of this process has been characterized in Bombyx mori (Ciufo et al. 2011).
Database mining revealed that the genome of D. melanogaster encodes at least 90 putative lipases (Flybase 2003), 44 of which are significantly expressed in the midgut (Chintapalli et al. 2007). Among those, a few are expected to be inactive, such as the yolk proteins of higher dipterans, and therefore unlikely to participate in lipid digestion (Bownes 1992). Furthermore, given that lipid digestion mostly takes place in the gut lumen, the presence of a signal peptide as prerequisite for protein secretion identified 29 of them as putative functional lipases. These enzymes are predicted (Flybase 2003) to fulfil most lipid-related enzymatic functions (Table 3), although specific activity has been formally demonstrated for only a few. Due to a high level of redundancy, the critical requirement of a given enzyme for a specific dietary lipid substrate cannot be easily addressed by a systematic gene mutant approach.
3.3. Lipase Activity and Expression
In the mammalian pancreas, signal peptide cleavage converts several proenzymes into active secreted enzymes. This maturation process might be conserved among insect digestive lipases, as reported for a secreted PLA2 of Spodoptera exigua (Sajjadian et al. 2019; Vatanparast et al. 2018). Lipase activity mainly relies on the penetration of the enzyme at the interface of lipid micelles. In contrast to mammals, insects do not produce bile acids to emulsify dietary lipids. This process likely involves lysophospholipids, which derive from phospholipid cleavage by either PLA1 or PLA2 (Figure 3B) (Ian De Veau and Schultz 1992; Sajjadian et al. 2019; Stanley 2006). In addition, the penetration of lipases into micelles is influenced by the acyl chain length, the pH and the concentration of several ions, in particular Ca2+ (Verger et al. 1973). For instance, the midgut lipase activity of the bloodsucking bug, Rhodnius prolixus, is optimal at neutral pH (Grillo et al. 2007), whereas that of PLA2 in Cicindella circumpicta (Uscian et al. 1995) and in Nicrophorus marginatus (Rana et al. 1997) is optimal at alkaline pH; all of them depending on Ca2+ concentration. However, a digestive PLA2 of A. aegypti has been shown to be Ca2+-independent (Nor Aliza and Stanley 1998). In most insect species, the midgut lumen exhibits an alkaline pH (Harrison 2001), which favors the activity of neutral lipases. Conversely, a few species exhibit an acidic midgut region (Barroso et al. 2021; Bonelli et al. 2019; Dubreuil 2004), which is optimal to sustain the activity of acid lipases. Nonetheless, genes encoding acid lipases can be found in most insect species (Table 1). In Manduca sexta, acid lipases are expressed in the midgut, although devoid of acidic region. Given that three of these acid lipases are not detected in the lumen juice, it has been proposed that they act in acidic lysosomes to hydrolyze dietary lipids taken up by the enterocytes (Miao et al. 2020).
Early studies focusing on monitoring lipase activity of the midgut juice revealed an effect of the nutritional status. In the hematophagous species A. aegypti, Panstrongylus megistus and R. prolixus, lipase activity is induced by the blood meal and reaches a maximum a few days later (Canavoso et al. 2004; Grillo et al. 2007; Rudin and Hecker 1979). In M. sexta (Stanley et al. 1998) and Gryllus bimaculatus (Weidlich et al. 2015) the activity of midgut lipases is higher in fed than in starved animals. Furthermore, a positive correlation between midgut lipase activity and dietary lipid content has been observed in Oedaleus asiaticus (Huang et al. 2017).
Conversely, FA supplementation in the diet of Epiphyas pastvittana represses lipase gene expression (Christeller et al. 2010), suggesting the existence of a FA sensing mechanism that adjusts lipase levels to FFA amounts. These studies revealed that the expression and/or the secretion of lipases is controlled by the diet, a vision generally recognized for discontinuous insect feeders but not for continuous ones. However, transcriptomic analyses in D. melanogaster larvae ⸺a continuous feeder⸺ counteract this vision, in that starvation but also high dietary glucose levels repress the expression of digestive lipases (Chng et al. 2014; Mattila et al. 2015; Zinke et al. 2002). At the molecular scale, the nuclear receptor DHR96 controls midgut TAG and cholesterol metabolism through the expression of lipases (Sieber and Thummel 2009; 2012). Finally, the insect gut microbiota, which contributes to homeostasis by providing essential nutrients (Grenier and Leulier 2020), has also been shown to modulate enzyme expression. In D. melanogaster, midgut colonization by Lactobacillus plantarum enhances the expression of digestive lipases (Erkosar and Leulier 2014). In summary, these studies support the notion that the activity and the production of lipases are tightly regulated by the midgut lumen content to optimize the digestion process.
4. Lipid Uptake into Enterocytes
4.1. Lipid Emulsifiers
The molecular mechanism of lipid crossing cell membranes remains puzzling, albeit extensively studied. The idea that lipids ─due to their biochemical nature─ could easily cross cell membranes suggests that the lipid bilayer would fail to strictly maintain compartment integrity. Single lipid molecules are poorly soluble, if not insoluble, in aqueous solution. Lipids integrating a membrane bilayer tend to remain within the lipid leaflets, rather than to exit at the other membrane side in the cytosolic aqueous phase. Within a membrane leaflet, lipids are free in their lateral movements, while, unless catalyzed by flippases (Montigny et al. 2016), spontaneous lipid flip-flops are rather rare events (Perez-Salas et al. 2021). Nonetheless, spontaneous flip-flops are favored by an extreme membrane curvature, which may happen in the enterocyte microvilli (Niot et al. 2009). Thus, a critical issue is lipid desorption, which likely requires specific cytosolic proteins. These proteins may also act as lipid sequesters, thereby preventing lipid transfer back into the gut lumen upon fasting.
Emulsification of lipids facilitates the access to water-soluble lipases (§ 6.3.1.) and the tight interaction of lipid molecules with the hydrophilic side of the microvillar membrane prior to their absorption. Insects do not produce bile acid and the intestinal absorption of lipids is proposed to proceed through the formation of glycolipids and amino-lipid complexes (Chapman et al. 2013; Miguel-Aliaga et al. 2018). In lepidopterans, the alkaline midgut favors the formation of FA micelles (Canavoso et al. 2001), questioning whether the molecular nature of the lipids emulsifiers varies among insect species. Analysis of surface-active substances in G. bimaculatus, revealed that FA-amino acid complexes (FACs) were potential emulsifiers (Collatz and Mommsen 1974). In phytophagous lepidopterans, FACs synthesis depends on the gut bacteria (Spiteller et al. 2000). FACs are regurgitated in oral secretions and act as potential emulsifiers to favor external lipolysis, yielding volatile compounds (Alborn et al. 1997; Halitschke et al. 2001; Spiteller and Boland 2003). These volatiles act at long distance to elicit a protective plant response (Dicke et al. 1990). Despite this adversative effect, the conservation of FAC synthesis and secretion suggests a benefit for the insect. Indeed, in Spodoptera litura and Heliothis virescens, these compounds are proposed to constitute nitrogen stores in the gut lumen, in the form of FA-glutamine complexes (Kuhns et al. 2012; Yoshinaga et al. 2008). As mentioned above (§ 6.3.3.), lysophospholipids resulting from phospholipid hydrolysis are likely playing a critical digestive role as lipid emulsifiers (Ian De Veau and Schultz 1992; Sajjadian et al. 2019; Turunen and Kastari 1979). Finally, the MGDG galactolipid, highly abundant in chloroplast thylakoids (§ 6.2.1.), has been shown to possess emulsifier properties (Chu et al. 2009). As for lysophospholipids, lysogalactolipid resulting from partial hydrolysis might act as intestinal emulsifiers in the midgut of phytophagous species. However, in spite of the characterization of all these putative lipid emulsifiers, their effective contribution to lipid uptake is not formally demonstrated and remains to be experimentally addressed in various insect species.
4.2. Lipid Transporters
The molecular mechanism of lipid uptake into the enterocytes has been poorly studied in insects, with most data relying on mammalian investigations. In mammals, dietary lipids are emulsified by bile acids, which facilitate lipolysis and lipid absorption, mostly MAGs, FFAs, lysophospholipids and cholesterol (Ko et al. 2020). An early study in the rat intestine proposed that linoleic acid uptake proceeds through two distinct mechanisms. At low concentrations, the uptake is saturable, thus, potentially energy- and/or transporter-dependent. In contrast, at higher concentrations, linoleic acid intake follows a linear plot, suggesting a concentration-dependent passive diffusion (Chow and Hollander 1979). This passive diffusion mechanism may result from a vectorial fusion/fission between micelles and membranes from the gut lumen towards the enterocyte cytoplasm, although thermodynamic considerations postulate that micelle-membrane fusion proceeds through unstable states of high-energy intermediates (Markvoort and Marrink 2011). Alternatively, it is tempting to speculate that high amounts of FAs might react with monovalent cations to form soaps, which could act as detergents, destabilizing cell membranes. Congruently, it has been shown that perfusion of the rat jejunum with emulsified lipids provokes membrane injuries in the brush border of the enterocytes, allowing lipid micelles intake (Kvietys et al. 1991). In sum, the molecular mechanism of this so-called passive diffusion remains largely unclear. The uptake of oleic acid has been shown to be more active than the one of palmitic or stearic acid in the midgut of the dragonfly Aeshna cyanea (Kirfel and Komnick 1999), suggesting either the existence of selective transporters or that passive diffusion is favored for unsaturated FAs. In addition, the observation that the uptake of lipids in not uniform along the entire midgut but takes place within specific sub-regions (see below § 6.6.) further supports the notion that the principle of passive lipid diffusion must be considered with caution. Of note, while intestinal absorption of cholesterol and FAs was initially believed to proceed merely in this way, transporter-dependent intake is now characterized for either.
In mammals, several proteins have been proposed to contribute to FA absorption into the enterocytes, including FATP4 (Fatty Acid transporter protein 4), FABPs (Fatty Acid binding proteins), FAT/CD36 and caveolin. FATP4 knockdown by antisense oligonucleotides reduces FA uptake by 50% in murine primary enterocytes (Stahl et al. 1999). However, a recent study indicated that FATP4 localizes at the ER to contribute to FA esterification, subsequently driving FA intake (Milger et al. 2006). Two other FA binding proteins, FABP1 and FABP2, despite their high intestinal expression, are also improbable FA transporter, rather acting to sequester FAs, thereby preventing lipotoxicity (Cifarelli and Abumrad 2018). The scavenger receptor CD36 is another candidate for mediating FA uptake, although such involvement in mammalian enterocytes remains controversial. In the mouse, one study reports a role of CD36 in FA absorption in the proximal, but not in the distal region of the small intestine (Nassir et al. 2007). However, two other studies rather suggest that CD36 plays a role in intracellular lipid trafficking and chylomicron formation (Goudriaan et al. 2002; Nauli et al. 2006). SR-B1, another scavenger receptor of the CD36 family highly expressed in the mouse intestine, is required for intestinal absorption of vitamin E (Reboul et al. 2006), but neither for FA nor for cholesterol intake (Cifarelli and Abumrad 2018). Caveolin1-3 are proteins associated with lipid rafts, which cover 50% of the membrane microvilli of the enterocytes (Kunding et al. 2010). Caveolins are proteins that channel lipid raft-derived endocytic vesicles to the ER. Caveolin-1 has been identified as a FA-binding protein (Trigatti et al. 1999) and its knockout in mice virtually abolishes the uptake of albumin-bound oleic acid (Siddiqi et al. 2013).
Most of these potential lipid transporters are conserved in insects (Table 4), although their roles in midgut lipid metabolism remain elusive. Two FABP homologues, MFB1 and MFB2 have been isolated from the midgut cytosol of M. sexta larvae (Smith et al. 1992). The D. melanogaster FATP4 homologue, Fatp1, has been the focus of interest in several tissues, although not in the midgut (Parvy et al. 2012). As reported for its mammalian counterpart, a study on D. melanogaster oenocytes suggests that it controls FA-CoA synthesis rather than FA uptake (Wicker-Thomas et al. 2015). A great number of scavenger receptors are encoded by insect genomes (Table 4), one of them, NinaD, has been shown to mediate carotenoid uptake in the midgut of D. melanogaster (Kiefer et al. 2002). To date, no caveolin genes have been found in insects. Nevertheless, two flotillin genes, encoding caveolae-associated proteins, have been described in D. melanogaster (Galbiati et al. 1998). Interestingly, ectopic expression of the murine flotillin-1 in Sf21 insect cells drives the formation of caveolae-like vesicles (Volonte et al. 1999).
The mechanism of cholesterol uptake into the enterocytes has been elucidated thanks to mammalian and insect studies. The Niemann-Pick type C genes (NPC1 or NPC2), which cause abnormal cholesterol accumulation, are conserved in insects (Table 4) (Zheng et al. 2018). The D. melanogaster Npc1a and Npc1b genes are the homologues of the mammalian NPC1 and NPC1-like 1 (NPC1L1) genes, whose products fulfil intracellular sterol trafficking and intestinal sterol absorption, respectively (Huang et al. 2005; Voght et al. 2007). The expression of either NPC1L1 in mammals or Npc1b in D. melanogaster is restricted to the apical membrane of enterocytes (Jing and Behmer 2020). In a rat hepatic cell line, flotillins have been shown to associate with NPC1L1, thereby mediating micelle sterol uptake (Ge et al. 2011). Further, in D. melanogaster, Npc1b mutation results in a defect of intestinal sterol absorption (Voght et al. 2007). In vertebrates, the sterol carrier protein-2 (SCP-2) is a cytoplasmic protein involved in the ER to plasma transfer of cholesterol (Jing and Behmer 2020). SCP-2 homologues have been identified and shown to be highly expressed in the midgut of lepidopteran and dipteran species (Gong et al. 2006; Kim and Lan 2010; Lan and Wessely 2004). SCP-2 knockdown in A. aegypti and in S. litura decreases overall cholesterol levels, whereas its over-expression in cell lines improves cholesterol uptake (Blitzer et al. 2005; Guo et al. 2009; Lan and Massey 2004). These studies identified bona fide cholesterol transporters, acting on cholesterol uptake and transfer between subcellular compartments.
In summary, the mechanism of intestinal sterol uptake has been elucidated and depends on evolutionary conserved transporters. In contrast, the intestinal uptake of FA and phospholipids is lesser elucidated but likely involves specific transporters, which remain to be clearly identified in insects.
5. Enterocyte Lipid Metabolism
5.1. Lipid Usage to Sustain Enterocyte Activity
Once uptaken, digested lipid products can be directly exported through the haemolymph, stored within specialized organelles (LDs, lysosomes) or used to sustain the energy and structural needs of the enterocytes. The diet likely influences lipid metabolism in the enterocytes. For instance, in D. melanogaster, the FA composition of the diet significantly modifies the phospholipid composition of the different tissues, including the midgut (Carvalho et al. 2012). In addition, this lipidome profiling revealed that, compared to other tissues, the midgut contains a higher fraction of sphingolipids with two double bonds in the long-chain base and in the esterified fatty acyl chain.
Enterocytes are highly active cells that require ATP as an energy source. The major ATP pool is produced via the respiratory chains coupled to the tricarboxylic acid cycle that relies on glycolysis, FA β-oxidation or amino acid catabolism. However, it has not been formally demonstrated which of these metabolic pathways is preferential in the enterocytes; most likely it depends on the diet composition and thus, on the insect species. For instance, in the stablefly Stomoxys calcitrans, a midgut posterior region (called lipoid zone) incorporates TAGs and phospholipids in the form of LDs often associated with mitochondria, suggesting that these lipid stores could be used for β-oxidation (Lehane 1977). In A. cyanea, peroxisome content increases in the apical region of the enterocytes of larvae fed on either natural lipid-rich food or high amounts of monounsaturated LCFA (Faber and Komnick 1989). Given that peroxisomes act to shorten and process FA not readily usable by mitochondria (Renne and Hariri 2021), these observations suggest that excess dietary LCFAs are first metabolized in peroxisomes to potentially sustain mitochondrial ATP production. In R. prolixus, high expression levels of β-oxidation enzymes have been detected in the midgut, suggesting the use of FA as a major source of energy in the enterocytes of this species (Ribeiro et al. 2014). Further, the microbiota may affect the midgut energy metabolism, through the production of short-chain FAs that may be consumable by the enterocytes, as described for the digestion of wood by termites and of plant polymers from pollen by honeybees (Brune 2014; Zheng et al. 2017).
The secretion of enzymes and the absorption of nutrients by enterocytes suggest a high rate of microvillar membrane replacement. In L. migratoria, it has been shown that glycerol and FFAs hydrolyzed from dietary triolein are found in the phospholipids of the intestinal wall (Weintraub and Tietz 1973). In Pieris brassicae larvae, the kinetics of phosphatidylcholine (PC) and phosphatidylethanolamine (PE) biosynthesis in the enterocyte, compared to that of DAG, suggests that these phospholipids are used for enterocyte membrane regeneration (Turunen 1993). In R. prolixus, the microvillar membrane contains high cholesterol levels (Albuquerque-Cunha et al. 2009) and the dynamics of cholesterol accumulation in the enterocytes, further suggests utilization for membrane regeneration (Entringer et al. 2021).
5.2. Lipid Metabolic Pathways in Enterocytes
In the enterocytes, dietary lipids can be converted to DAGs, TAGs or phospholipids. In S. calcitrans, it has been estimated that 60% of dietary FAs are converted to TAGs and 20% to phospholipids (Lehane 1977). In M. sexta fed radioactive triolein, labelling is eventually found in phospholipids and TAGs of the enterocytes and in the circulating DAGs (Tsuchida and Wells 1988). DAGs are the prevalent form of lipid transport through the haemolymph and are also the precursors for phospholipid biogenesis (Vance 2015).
FFAs, in particular the saturated ones, are highly toxic (Listenberger et al. 2003). To prevent this deleterious effect, FAs can be either degraded through mitochondrial β-oxidation or esterified to TAGs and stored in LDs. The biosynthesis of TAGs may proceed through three distinct pathways, which begin by either acylation of glycerol-phosphate in the mitochondria or the ER, acylation of dihydroxyacetone in the peroxisomes, or acylation of MAGs in the ER; the subsequent steps taking place in the ER (Coleman and Mashek 2011; Ko et al. 2020). MAG acylation gives rise to DAG, whereas the glycerol-phosphate and dihydroxyacetone pathways result in phosphatidic acid (PA), which after dephosphorylation by Lipin leads to DAG (Figure 4). The glycerol-phosphate pathway also called the Kennedy pathway is the most universal. In mammals, it takes place in the liver and adipose tissue, while the conversion of MAGs ─resulting from luminal TAG hydrolysis─ into DAGs is prevalent in the small intestine. To date, besides the characterization of a putative MAG acyltransferase activity in the midgut of P. americana (Hoffman and Downer 1979), no evidence suggests that the MAG pathway operates in the insect midgut. In contrast, the PA pathway has been reported in the midgut of several insect species, including P. brassicae (Turunen 1993), M. sexta (Canavoso and Wells 2000), P. megistus (Canavoso et al. 2004) and D. melanogaster (Kuhnlein 2012).
DAGs are the precursors for biogenesis of the most abundant phospholipids, PC and PE (Figure 4) (Vance 2015). First, phosphocholine and phosphoethanolamine are converted to CDP-choline and CDP-ethanolamine at the nuclear and ER membranes, respectively. The final step to PC and PE synthesis takes place at the ER membrane. Synthesis of phosphatidylserine may proceed by the exchange of the head group of PC or PE to a serine residue at the ER membrane. Synthesis of other phospholipids utilizes CTP and PA to form CDP-DAG; replacement of the CDP group by other functional groups results in the formation of phosphatidylinositol (PI), phosphatidylglycerol (PG) or cardiolipin (Figure 4). The synthesis of sphingomyelin, the most abundant sphingolipid, occurs at the Golgi apparatus from ceramide and PC. Phospholipid biosynthesis and expression of the potentially required enzymes have been reported in the midgut of several insect species, including S. calcitrans, M. sexta, R. prolixus and D. melanogaster (Buchon et al. 2013; Grillo et al. 2007; Lehane 1977; Ribeiro et al. 2014; Tsuchida and Wells 1988). However, the functional characterization of the underlying enzymes in the enterocytes remains poorly investigated.
Excesses of dietary sterols can be temporarily stored as free or steryl-ester forms in lysosomes or LDs, or efflux back to the gut lumen likely via ABC transporters, as reported in mammal studies (Entringer et al. 2021; Jing et al. 2012; Sieber and Thummel 2012). In insects, Npc2 are soluble proteins encoded by multiple genes (Table 4), which are not equally expressed across different organs (Huang et al. 2007; Jing and Behmer 2020; Jupatanakul et al. 2014). Two of the D. melanogaster homologues, Npc2a and Npc2b, have a redundant function in controlling intracellular sterol distribution (Huang et al. 2007). Based on mammalian and insect studies, Npc2 homologues are required for intralysosomal sterol transfer to Npc1a (Huang et al. 2007; Jing and Behmer 2020). Loss of Npc1a in D. melanogaster results in abnormal cholesterol retention in lysosome-like organelles in different tissues, including the midgut (Huang et al. 2005). Importantly, Npc1a and Npc1b are two large transmembrane proteins, which fulfil cholesterol intracellular trafficking and its uptake into enterocytes (§ 6.4.2.), respectively. Although they likely evolved from a single ancestor gene, Npc1a and Npc1b are not interchangeable transporters.
5.3. Lipid Efflux to Haemolymph
The distribution of lipids to the different tissues mostly proceeds via lipoprotein transfer through the haemolymph. In most insect species, lipoproteins typically carry sterols, phospholipids and DAGs (Canavoso et al. 2001), although DAGs are replaced by TAGs in some dipteran species (Ford and Van Heusden 1994; Pennington and Wells 2002). The transfer of lipids from the enterocytes to lipoproteins requires first a remobilization from the storage organelles. Given that intracellular LDs store TAGs in higher proportions than DAGs, the former must be hydrolyzed to DAGs prior to their haemolymph transfer. In D. melanogaster, the adipocyte triglyceride lipase Brummer, which converts TAG to DAG, is expressed in the midgut and, thus, potentially works in the enterocytes (Gronke et al. 2005). Free sterols rather than steryl-esters are loaded on lipoproteins (Carvalho et al. 2012; Entringer et al. 2021; Jouni et al. 2002; Yun et al. 2002), suggesting that a steryl-esterase must be active in the enterocytes prior to haemolymph efflux (Sieber and Thummel 2012).
The lipid transfer from the enterocytes to the haemolymph requires a lipophorin receptor, a lipid transfer particle (LTP) and a circulating lipophorin (Lpp). The lipophorin receptor(s) acting to recruit both lipophorin and LTP at the basal membrane of the enterocytes have not been formally identified (Arrese et al. 2001; Rodriguez-Vazquez et al. 2015). A number of studies in several insect species have reported the purification of lipophorins, characterized their binding to the enterocyte membrane and analyzed their lipid content (Atella et al. 1995; Canavoso and Wells 2001; Ford and Van Heusden 1994; Fruttero et al. 2009; Gondim and Wells 2000; Grillo et al. 2003). LTP that acts to transfer lipids between intracellular and extracellular compartments in a bidirectional manner (Arrese et al. 2001), has been shown to interact with the midgut of M. sexta (Canavoso and Wells 2001) and of D. melanogaster (Palm et al. 2012; Rodriguez-Vazquez et al. 2015). An in-depth study of the D. melanogaster scaffolding apolipoproteins demonstrated that both apoLTP and apoLpp are synthesized in the fat body, matured by lipid loading and transferred through the haemolymph to reach the midgut, where LTP mediates the transfer of lipids from the enterocytes to Lpp (Palm et al. 2012).
6. The Dynamics of Lipid Digestion in Insects
6.1. An Attempt to Schematize the Lipid Digestive Process
Lipid digestion mostly proceeds in the midgut lumen (Chapman et al. 2013; Gazara et al. 2017; Toprak et al. 2020), although it can occur externally in a few species that regurgitate salivary gland enzymes to liquefy their food (Canton and Bonning 2020; Holtof et al. 2019; Rana et al. 1997). Lipid digestion may also start within the crop in species for which the countercurrent flux of the ectoperitrophic space dumps enzymes secreted from the anterior midgut into the foregut lumen (Holtof et al. 2019; Weidlich et al. 2015). For instance, in P. americana, the crop is the largest digestive organ, which contains high amounts of lipases (Tamaki et al. 2014) and can uptake FFAs (Hoffman and Downer 1976). Conversely, in xylophagous and phytophagous insects, the digestion of plant-derived macromolecules by symbionts and the uptake of the resulting SCFAs take place in the hindgut (Breznak 1982; Brune and Dietrich 2015).
Figure 5 summarizes the various processes of lipid digestion that occur in insect midguts. Most studies suggest that TAGs are completely digested to glycerol and FFAs, although MAGs may also be produced in some species (Hoffman and Downer 1979; Turunen 1975), but likely excreted within the frass. Phospholipids may be partially or completely hydrolyzed; the resulting FAs, glycerol and choline, and the hydrolate intermediates, such as lysophosphatidylcholine, are absorbed by enterocytes (Turunen and Kastari 1979; Weiher and Komnick 1997). Galactolipids are hydrolyzed to lysogalactolipids (Christeller et al. 2011), and likely through complete hydrolysis to galactose, glycerol and FAs, as described in mammals (Sahaka et al. 2020). Lipid uptake might proceed through passive diffusion, although not in all midgut sub-regions, suggesting that it depends on the molecular composition of the plasma membrane or on putative transporters (§ 6.4.2.). Indeed, transporters for cholesterol have been identified in the midgut, whereas those for FAs and lysophospholipids have not been formally identified to date. To renew their plasma membrane, enterocytes can incorporate dietary sterol and phospholipids derived from lipid digestive products.
TAGs, steryl-esters and phospholipids can be stored in LDs, whereas lysosomes store free sterols. In most species, lipids are exported to the organs via the haemolymph in the form of DAGs, sterols and phospholipids loaded on lipophorins. This synthetic picture partly reflects the process of lipid digestion, which depends on specific activities in specialized midgut sub-regions and on the diet type of each insect species.
6.2. Midgut Regionalization of Lipid Digestion
The anterior midgut and the associated caeca play a critical role in lipid digestion (Toprak et al. 2020). For instance, the caeca of G. bimaculatus (Orthoptera) fulfil most of the total lipase activity (Weidlich et al. 2015). Studies in three lepidopteran species, Diatraea grandiosella (Turunen and Chippendale 1977), P. brassicae (Turunen 1990) and M. sexta (Smith et al. 1992), suggest that lipid absorption takes place in the anterior midgut, although secretion of PLA2 in the midgut of M. sexta is low in the anterior region and highest in the middle region (Rana and Stanley 1999).
In three species of hemipterans, R. prolixus (Billingsley 1990), P. megistus (Canavoso et al. 2004) and Cimex hemipterus (Azevedo et al. 2009), lipid inclusions have been observed in the anterior midgut after the blood meal. However, in R. prolixus, it has not been demonstrated whether these inclusions directly result from lipid uptake in the anterior midgut cells (Billingsley 1990), and recent studies revealed that breakdown of complex lipids and the uptake of the resulting metabolites mostly take place in the posterior midgut (Grillo et al. 2007; Ribeiro et al. 2014). Furthermore, lipase activity has been observed in all midgut regions of Brontocoris tabidus, another hemipteran species (Guedes et al. 2007).
Studies on dipterans suggest that the posterior midgut is a critical region for lipid digestion. In S. calcitrans, enzymatic digestion and lipid absorption have been reported in the posterior midgut (Lehane 1977). In the midgut juice of Hermatia illucens, lipase activity is high in the anterior and posterior, but not in the middle region (Bonelli et al. 2019). Gene expression analyses from midgut sections provide considerable information about functional midgut regionalization. In D. melanogaster, lipid-related digestive enzymes are expressed in specific midgut regions, suggesting that each region fulfils distinct digestive functions (Buchon et al. 2013; Zinke et al. 2002). In Anopheles gambiae, several genes encoding lipids transporters are enriched in the gastric caeca and the anterior midgut, although a few appear to be also enriched in the posterior midgut, the Malpighian tubules and the hindgut (Neira Oviedo et al. 2008). In Musca domestica, the anterior midgut expresses PLA1 at high levels, whereas the middle midgut expresses high levels of acid lipases and the posterior midgut acid lipase as well as potential lipid transporters (Barroso et al. 2021). Based on these results, the authors propose that lysophospholipids resulting from PLA1 activity in the anterior midgut favor micelle formation in the middle midgut and the digestion of TAGs to FFAs, which are eventually absorbed in the posterior midgut (Barroso et al. 2021).
In summary, lipid digestion is a multiple-step process recruiting specific enzymes active in distinct specialized midgut sections, following a functional sequence of events during which the digestion of certain classes of lipids potentiates the digestion of other lipids and their subsequent absorption. The functional regionalization of the midgut is likely necessary for efficient lipid digestion and has been observed in all insects studied to date but does not exhibit an identical architecture among species.
7. Conclusions
This chapter aimed to provide an overview of the studies that reported on lipid digestion in the insect midgut. Accordingly, we apologize to those that could not be cited. In spite of a great number of publications, data related to lipid digestion processes remain sparse and many questions are still opened. Not surprisingly, it is not possible to draw a uniform picture of insect digestion, because of the tremendous number of insect species and of the large variety of diet types. Nonetheless, the comprehension of the insect digestive processes is a crucial challenge for the future: i) to develop appropriate strategies against agriculture pests and vectors of pathogens, while maintaining biodiversity; ii) to improve the efficacy of insect-dependent industrial production (e.g. silkworm); iii) to set alternative models aimed at investigating human health and diseases. To this end, future investigations should evaluate the relative contribution of passive- versus transporter-dependent lipid uptake, and precisely identify the lipases that are active in the midgut as well as their substrate specificity. More importantly, to provide an integrated view of lipid digestion for a given species, it will be essential to clarify the contribution of each midgut sub-region in the different digestive processes and to further determine to which extent the microbiota influences these processes. While setting a uniform picture of insect lipid digestion is obviously a futile challenge, these studies will certainly improve our understanding of the digestive physiology of key insect species.
References
- Agarwal HC, Nair AM (1976) Studies on the cholesterol ester hydrolase of trogoderma (coleoptera). Biochem J. 157:111-116. [CrossRef]
- Alborn HT, Turlings TCJ, Jones TH, Stenhagen G, Loughrin JH, Tumlison JH (1997) An elicitor of plant volatiles from beet armyworm oral secretion. Science. 276:945-949.
- Albuquerque-Cunha JM, Gonzalez MS, Garcia ES, Mello CB, Azambuja P, Almeida JC, de Souza W, Nogueira NF (2009) Cytochemical characterization of microvillar and perimicrovillar membranes in the posterior midgut epithelium of rhodnius prolixus. Arthropod Struct Dev. 38:31-44. [CrossRef]
- Arrese EL, Canavoso LE, Jouni ZE, Pennington JE, Tsuchida K, Wells MA (2001) Lipid storage and mobilization in insects: Current status and future directions. Insect Biochem Mol Biol. 31:7-17.
- Atella GC, Gondim C, Masuda H (1995) Loading of lipophorin particles with phospholipids at the midgut of rhodnius prolixus. Arch Insect Biochem Physiol. 30:337-350. [CrossRef]
- Azevedo DO, Neves CA, Mallet JR, Goncalves TC, Zanuncio JC, Serrao JE (2009) Notes on midgut ultrastructure of cimex hemipterus (hemiptera: Cimicidae). J Med Entomol. 46:435-441. [CrossRef]
- Barroso IG, Cardoso C, Ferreira C, Terra WR (2021) Transcriptomic and proteomic analysis of the underlying mechanisms of digestion of triacylglycerols and phosphatides and absorption and fate of fatty acids along the midgut of musca domestica. Comp Biochem Physiol Part D Genomics Proteomics. 39:100826. [CrossRef]
- Bateta R, Wang J, Wu Y, Weiss BL, Warren WC, Murilla GA, Aksoy S, Mireji PO (2017) Tsetse fly (glossina pallidipes) midgut responses to trypanosoma brucei challenge. Parasit Vectors. 10:614. [CrossRef]
- Beenakkers AM, Van der Horst DJ, Van Marrewijk WJ (1985) Insect lipids and lipoproteins, and their role in physiological processes. Prog Lipid Res. 24:19-67. [CrossRef]
- Biagio FP, Tamaki FK, Terra WR, Ribeiro AF (2009) Digestive morphophysiology of gryllodes sigillatus (orthoptera: Gryllidae). J Insect Physiol. 55:1125-1133. [CrossRef]
- Billingsley PF (1990) Blood digestion in the mosquito, anopheles stephensi liston (diptera: Culicidae): Partial characterization and post-feeding activity of midgut aminopeptidases. Arch Insect Biochem Physiol. 15:149-163. [CrossRef]
- Blitzer EJ, Vyazunova I, Lan Q (2005) Functional analysis of aescp-2 using gene expression knockdown in the yellow fever mosquito, aedes aegypti. Insect Mol Biol. 14:301-307. [CrossRef]
- Blomquist G, Borgeson CE, Vundla M (1991) Polyunsaturated fatty acids and ecosanoids in insects. Insect Biochem. 21:99-106.
- Bollade D, Paris R, Moulins M (1970) Origine et mode d'action de la lipase intestinale chez les blattes. J Insect Physiol. 16:45-53.
- Bolognesi R, Terra WR, Ferreira C (2008) Peritrophic membrane role in enhancing digestive efficiency. Theoretical and experimental models. J Insect Physiol. 54:1413-1422. [CrossRef]
- Bonelli M, Bruno D, Caccia S, Sgambetterra G, Cappellozza S, Jucker C, Tettamanti G, Casartelli M (2019) Structural and functional characterization of hermetia illucens larval midgut. Front Physiol. 10:204. [CrossRef]
- Bownes M (1992) Why is there sequence similarity between insect yolk proteins and vertebrate lipases? J Lipid Res. 33:777-790.
- Breznak JA (1982) Intestinal microbiota of termites and other xylophagous insects. Annu Rev Microbiol. 36:323-343. [CrossRef]
- Brune A (2014) Symbiotic digestion of lignocellulose in termite guts. Nat Rev Microbiol. 12:168-180. [CrossRef]
- Brune A, Dietrich C (2015) The gut microbiota of termites: Digesting the diversity in the light of ecology and evolution. Annu Rev Microbiol. 69:145-166. [CrossRef]
- Buchon N, Osman D, David FP, Fang HY, Boquete JP, Deplancke B, Lemaitre B (2013) Morphological and molecular characterization of adult midgut compartmentalization in drosophila. Cell Rep. 3:1725-1738. [CrossRef]
- Caccia S, Casartelli M, Tettamanti G (2019) The amazing complexity of insect midgut cells: Types, peculiarities, and functions. Cell Tissue Res. 377:505-525. [CrossRef]
- Canavoso LE, Frede S, Rubiolo ER (2004) Metabolic pathways for dietary lipids in the midgut of hematophagous panstrongylus megistus (hemiptera: Reduviidae). Insect Biochem Mol Biol. 34:845-854. [CrossRef]
- Canavoso LE, Jouni ZE, Karnas KJ, Pennington JE, Wells MA (2001) Fat metabolism in insects. Annu Rev Nutr. 21:23-46.
- Canavoso LE, Wells MA (2000) Metabolic pathways for diacylglycerol biosynthesis and release in the midgut of larval manduca sexta. Insect Biochem Mol Biol. 30:1173-1180. [CrossRef]
- Canavoso LE, Wells MA (2001) Role of lipid transfer particle in delivery of diacylglycerol from midgut to lipophorin in larval manduca sexta. Insect Biochem Mol Biol. 31:783-790. [CrossRef]
- Canton PE, Bonning BC (2020) Extraoral digestion: Outsourcing the role of the hemipteran midgut. Curr Opin Insect Sci. 41:86-91. [CrossRef]
- Carvalho M, Sampaio JL, Palm W, Brankatschk M, Eaton S, Shevchenko A (2012) Effects of diet and development on the drosophila lipidome. Mol Syst Biol. 8:600. [CrossRef]
- Carvalho M, Schwudke D, Sampaio JL, Palm W, Riezman I, Dey G, Gupta GD, Mayor S, Riezman H, Shevchenko A et al. (2010) Survival strategies of a sterol auxotroph. Development. 137:3675-3685.
- Chapman RF, Simpson SJ, Douglas AE. (2013). The insects. Structure and function.
- Chintapalli VR, Wang J, Dow JA (2007) Using flyatlas to identify better drosophila melanogaster models of human disease. Nat Genet. 39:715-720.
- Chng WA, Sleiman MSB, Schupfer F, Lemaitre B (2014) Transforming growth factor beta/activin signaling functions as a sugar-sensing feedback loop to regulate digestive enzyme expression. Cell Rep. 9:336-348. [CrossRef]
- Chow SL, Hollander D (1979) A dual, concentration-dependent absorption mechanism of linoleic acid by rat jejunum in vitro. J Lipid Res. 20:349-356.
- Christeller JT, Amara S, Carriere F (2011) Galactolipase, phospholipase and triacylglycerol lipase activities in the midgut of six species of lepidopteran larvae feeding on different lipid diets. J Insect Physiol. 57:1232-1239. [CrossRef]
- Christeller JT, Poulton J, Markwick NM, Simpson RM (2010) The effect of diet on the expression of lipase genes in the midgut of the lightbrown apple moth (epiphyas postvittana walker; tortricidae). Insect Mol Biol. 19:9-25. [CrossRef]
- Chu BS, Rich GT, Ridout MJ, Faulks RM, Wickham MS, Wilde PJ (2009) Modulating pancreatic lipase activity with galactolipids: Effects of emulsion interfacial composition. Langmuir. 25:9352-9360. [CrossRef]
- Cifarelli V, Abumrad NA (2018) Intestinal cd36 and other key proteins of lipid utilization: Role in absorption and gut homeostasis. Compr Physiol. 8:493-507. [CrossRef]
- Cioffi M (1984) Ultrastructure of arthropod transporting epithelia. Amer Zool. 24:139-156.
- Ciufo LF, Murray PA, Thompson A, Rigden DJ, Rees HH (2011) Characterisation of a desmosterol reductase involved in phytosterol dealkylation in the silkworm, bombyx mori. PLoS One. 6:e21316. [CrossRef]
- Clark AJ, Block K (1959) The absence of sterol synthesis in insects. J Biol Chem. 234:2578-2582.
- Clayton RB (1964) The utilization of sterols by insects. J Lipid Res. 5:3-19.
- Cleveland LR (1923) Symbiosis between termites and their intestinal protozoa. Proc Natl Acad Sci U S A. 9:424-428. [CrossRef]
- Cobbs C, Heath J, Stireman JO, 3rd, Abbot P (2013) Carotenoids in unexpected places: Gall midges, lateral gene transfer, and carotenoid biosynthesis in animals. Mol Phylogenet Evol. 68:221-228. [CrossRef]
- Coleman RA, Mashek DG (2011) Mammalian triacylglycerol metabolism: Synthesis, lipolysis, and signaling. Chem Rev. 111:6359-6386. [CrossRef]
- Collatz K-G, Mommsen T (1974) Die struktur der emulgierenden substanzen verschicdener invertebraten. J comp Physiol. 94:339-352.
- Consuegra J, Grenier T, Akherraz H, Rahioui I, Gervais H, da Silva P, Leulier F (2020a) Metabolic cooperation among commensal bacteria supports drosophila juvenile growth under nutritional stress. iScience. 23:101232. [CrossRef]
- Consuegra J, Grenier T, Baa-Puyoulet P, Rahioui I, Akherraz H, Gervais H, Parisot N, da Silva P, Charles H, Calevro F et al. (2020b) Drosophila-associated bacteria differentially shape the nutritional requirements of their host during juvenile growth. PLoS Biol. 18:e3000681. [CrossRef]
- Cooper MK, Wassif CA, Krakowiak PA, Taipale J, Gong R, Kelley RI, Porter FD, Beachy PA (2003) A defective response to hedgehog signaling in disorders of cholesterol biosynthesis. Nat Genet. 33:508-513. [CrossRef]
- Coutinho-Abreu IV, Serafim TD, Meneses C, Kamhawi S, Oliveira F, Valenzuela JG (2020) Leishmania infection induces a limited differential gene expression in the sand fly midgut. BMC Genomics. 21:608. [CrossRef]
- Cruden DL, Markovetz AJ (1987) Microbial ecology of the cockroach gut. Annu Rev Microbiol. 41:617-643. [CrossRef]
- da Silva G, Costa Ramos LF, Dos Santos Seckler H, Mendonca Gomes F, Reis Cortines J, Ramos I, Dinis Anobom C, de Alcantara Machado E, Perpetua de Oliveira DM (2019) Biochemical characterization of digestive membrane-associated alkaline phosphatase from the velvet bean caterpillar anticarsia gemmatalis. Arch Insect Biochem Physiol. 102:e21591. [CrossRef]
- Dadd RH (1961) The nutritional requirements of locusts. J Ins Physiol. 6:126-145.
- Dadd RH, Kleinjan JE (1979) Essential fatty acid for the mosquito culex pipiens: Arachidonic acid. J Insect Physiol. 25:495-502. [CrossRef]
- de Oliveira Souza A, Couto-Lima CA, Catalao CHR, Santos-Junior NN, Dos Santos JF, da Rocha MJA, Alberici LC (2019) Neuroprotective action of eicosapentaenoic (epa) and docosahexaenoic (dha) acids on paraquat intoxication in drosophila melanogaster. Neurotoxicology. 70:154-160. [CrossRef]
- de Sousa ME, Wanderley-Teixeira V, Teixeira AA, de Siqueira HA, Santos FA, Alves LC (2009) Ultrastructure of the alabama argillacea (hubner) (lepidoptera: Noctuidae) midgut. Micron. 40:743-749. [CrossRef]
- Derewenda ZS (1994) Structure and function of lipases. Adv Protein Chem. 45:1-52. [CrossRef]
- Dewett D, Lam-Kamath K, Poupault C, Khurana H, Rister J (2021) Mechanisms of vitamin a metabolism and deficiency in the mammalian and fly visual system. Dev Biol. 476:68-78. [CrossRef]
- Dicke M, Sabelis MW, Takabayashi J, Bruin J, Posthumus MA (1990) Plant strategies of manipulating predatorprey interactions through allelochemicals: Prospects for application in pest control. J Chem Ecol. 16:3091-3118. [CrossRef]
- Dragh MA, Xu Z, Al-Allak ZS, Hong L (2017) Vitamin k2 prevents lymphoma in drosophila. Sci Rep. 7:17047. [CrossRef]
- Dubreuil RR (2004) Copper cells and stomach acid secretion in the drosophila midgut. Int J Biochem Cell Biol. 36:745-752. [CrossRef]
- Dutta D, Dobson AJ, Houtz PL, Glasser C, Revah J, Korzelius J, Patel PH, Edgar BA, Buchon N (2015) Regional cell-specific transcriptome mapping reveals regulatory complexity in the adult drosophila midgut. Cell Rep. 12:346-358. [CrossRef]
- Eguchi M (1995) Alkaline phosphatase isozymes in insects and comparison with mammalian enzyme. Comp Biochem Physiol B Biochem Mol Biol. 111:151-162. [CrossRef]
- Engel P, Moran NA (2013) The gut microbiota of insects - diversity in structure and function. FEMS Microbiol Rev. 37:699-735. [CrossRef]
- Entringer PF, Majerowicz D, Gondim KC (2021) The fate of dietary cholesterol in the kissing bug rhodnius prolixus. Front Physiol. 12:654565. [CrossRef]
- Erkosar B, Leulier F (2014) Transient adult microbiota, gut homeostasis and longevity: Novel insights from the drosophila model. FEBS Lett. 588:4250-4257. [CrossRef]
- Faber V, Komnick H (1989) Peroxisomes of the midgut epithelium, malpighian tubules and fat body of larvae of the dragonfly, aeshna cyanea. Tissue Cell. 21:917-924. [CrossRef]
- Ferreira C, Capella AN, Sitnik R, Terra WR (1994) Properties of the digestive enzymes and the permeability of the peritrophic membrane of spodoptera frugiperda (lepidoptera) larvae. Comp Biochem Physiol. 107A:631-640.
- Fialho MCQ, Terra WR, Moreira NR, Zanuncio JC, Serrao JE (2013) Ultrastructure and immunolocalization of digestive enzymes in the midgut of podisus nigrispinus (heteroptera: Pentatomidae). Arthropod Struct Dev. 42:277-285. [CrossRef]
- Fluegel ML, Parker TJ, Pallanck LJ (2006) Mutations of a drosophila npc1 gene confer sterol and ecdysone metabolic defects. Genetics. 172:185-196. [CrossRef]
- Flybase (2003) The flybase database of the drosophila genome projects and community literature. Nucleic Acids Res. 31:172-175. [CrossRef]
- Ford PS, Van Heusden MC (1994) Triglyceride-rich lipophorin in aedes aegypti (diptera: Culicidae). J Med Entomol. 31:435-441. [CrossRef]
- Fraenkel G, Blewett M (1946) Linoleic acid, vitamin e and other fat-soluble substances in the nutrition of certain insects, ephestia kuehniella, e. Elutella, e. Cautella and plodia interpunctella (lep.). J Exp Biol. 22:172-190. [CrossRef]
- Fruttero LL, Rubiolo ER, Canavoso LE (2009) Biochemical and cellular characterization of lipophorin-midgut interaction in the hematophagous panstrongylus megistus (hemiptera: Reduviidae). Insect Biochem Mol Biol. 39:322-331. [CrossRef]
- Fukuda S, Toh H, Hase K, Oshima K, Nakanishi Y, Yoshimura K, Tobe T, Clarke JM, Topping DL, Suzuki T et al. (2011) Bifidobacteria can protect from enteropathogenic infection through production of acetate. Nature. 469:543-547. [CrossRef]
- Fuzita FJ, Pimenta DC, Palmisano G, Terra WR, Ferreira C (2019) Detergent-resistant domains in spodoptera frugiperda midgut microvillar membranes and their relation to microapocrine secretion. Comp Biochem Physiol B Biochem Mol Biol. 235:8-18. [CrossRef]
- Galbiati F, Volonte D, Goltz JS, Steele Z, Sen J, Jurcsak J, Stein D, Stevens L, Lisanti MP (1998) Identification, sequence and developmental expression of invertebrate flotillins from drosophila melanogaster. Gene. 210:229-237. [CrossRef]
- Gandotra S, Bhuyan PM, Gogoi DK, Kumar A, Subramanian S (2016) Screening of nutritionally important gut bacteria from the lepidopteran insects through qualitative enzyme assays. Proc Natl Acad Sci, India. 88:329-337. [CrossRef]
- Garrido D, Rubin T, Poidevin M, Maroni B, Le Rouzic A, Parvy JP, Montagne J (2015) Fatty acid synthase cooperates with glyoxalase 1 to protect against sugar toxicity. PLoS Genet. 11:e1004995. [CrossRef]
- Gazara RK, Cardoso C, Bellieny-Rabelo D, Ferreira C, Terra WR, Venancio TM (2017) De novo transcriptome sequencing and comparative analysis of midgut tissues of four non-model insects pertaining to hemiptera, coleoptera, diptera and lepidoptera. Gene. 627:85-93. [CrossRef]
- Ge L, Qi W, Wang LJ, Miao HH, Qu YX, Li BL, Song BL (2011) Flotillins play an essential role in niemann-pick c1-like 1-mediated cholesterol uptake. Proc Natl Acad Sci U S A. 108:551-556. [CrossRef]
- Gondim KC, Wells MA (2000) Characterization of lipophorin binding to the midgut of larval manduca sexta. Insect Biochem Mol Biol. 30:405-413. [CrossRef]
- Gong J, Hou Y, Zha XF, Lu C, Zhu Y, Xia QY (2006) Molecular cloning and characterization of bombyx mori sterol carrier protein x/sterol carrier protein 2 (scpx/scp2) gene. DNA Seq. 17:326-333. [CrossRef]
- Goudriaan JR, Dahlmans VE, Febbraio M, Teusink B, Romijn JA, Havekes LM, Voshol PJ (2002) Intestinal lipid absorption is not affected in cd36 deficient mice. Mol Cell Biochem. 239:199-202.
- Grenier T, Leulier F (2020) How commensal microbes shape the physiology of drosophila melanogaster. Curr Opin Insect Sci. 41:92-99. [CrossRef]
- Grillo LA, Majerowicz D, Gondim KC (2007) Lipid metabolism in rhodnius prolixus (hemiptera: Reduviidae): Role of a midgut triacylglycerol-lipase. Insect Biochem Mol Biol. 37:579-588. [CrossRef]
- Grillo LA, Pontes EG, Gondim KC (2003) Lipophorin interaction with the midgut of rhodnius prolixus: Characterization and changes in binding capacity. Insect Biochem Mol Biol. 33:429-438. [CrossRef]
- Gronke S, Mildner A, Fellert S, Tennagels N, Petry S, Muller G, Jackle H, Kuhnlein RP (2005) Brummer lipase is an evolutionary conserved fat storage regulator in drosophila. Cell Metab. 1:323-330. [CrossRef]
- Guedes BAM, Zanuncio JC, Ramalho FS, Serrao JE (2007) Midgut morphology and enzymes of the obligate zoophytophagous stinkbug brontocoris tabidus (signoret, 1963) (heteroptera: Pentatomidae). Pan-Pac Entomol. 83:66-74.
- Guo XR, Zheng SC, Liu L, Feng QL (2009) The sterol carrier protein 2/3-oxoacyl-coa thiolase (scpx) is involved in cholesterol uptake in the midgut of spodoptera litura: Gene cloning, expression, localization and functional analyses. BMC Mol Biol. 10:102. [CrossRef]
- Hakim RS, Baldwin K, Smagghe G (2010) Regulation of midgut growth, development, and metamorphosis. Annu Rev Entomol. 55:593-608. [CrossRef]
- Halitschke R, Schittko U, Pohnert G, Boland W, Baldwin IT (2001) Molecular interactions between the specialist herbivore manduca sexta (lepidoptera, sphingidae) and its natural host nicotiana attenuata. Iii. Fatty acid-amino acid conjugates in herbivore oral secretions are necessary and sufficient for herbivore-specific plant responses. Plant Physiol. 125:711-717. [CrossRef]
- Harrison JF (2001) Insect acid-base physiology. Annu Rev Entomol. 46:221-250. [CrossRef]
- Harrison JF, Kaiser A, VandenBrooks JM (2010) Atmospheric oxygen level and the evolution of insect body size. Proc Biol Sci. 277:1937-1946. [CrossRef]
- Hegedus D, Erlandson M, Gillott C, Toprak U (2009) New insights into peritrophic matrix synthesis, architecture, and function. Annu Rev Entomol. 54:285-302. [CrossRef]
- Hobson RP (1935) On a fat-soluble growth factor required by blow-fly larvae: Identity of the growth factor with cholesterol. Biochem J. 29:2023-2026. [CrossRef]
- Hoffman AGD, Downer RGH (1976) The crop as an organ of glyceride absorption in the american cockroach, periplaneta americana l. Can J Zool. 54:1165-1171.
- Hoffman AGD, Downer RGH (1979) End product specificity of triacylglycerol lipases from intestine, fat body, muscle and haemolymph of the american cockroach, periplaneta americana l. Lipids. 14:893-899.
- Holmquist M (2000) Alpha/beta-hydrolase fold enzymes: Structures, functions and mechanisms. Curr Protein Pept Sci. 1:209-235. [CrossRef]
- Holtof M, Lenaerts C, Cullen D, Vanden Broeck J (2019) Extracellular nutrient digestion and absorption in the insect gut. Cell Tissue Res. 377:397-414. [CrossRef]
- Horne I, Haritos VS, Oakeshott JG (2009) Comparative and functional genomics of lipases in holometabolous insects. Insect Biochem Mol Biol. 39:547-567. [CrossRef]
- House HL (1966) Effects of vitamins e and a on growth and development, and the necessity of vitamin e for reproduction in the parasitoid agria affinis (fallen) (diptera, sarcophagidae). J Insect Physiol. 12:409-417. [CrossRef]
- Hu X, Chen L, Xiang X, Yang R, Yu S, Wu X (2012) Proteomic analysis of peritrophic membrane (pm) from the midgut of fifth-instar larvae, bombyx mori. Mol Biol Rep. 39:3427-3434. [CrossRef]
- Huang X, Ma J, Qin X, Tu X, Cao G, Wang G, Nong X, Zhang Z (2017) Biology, physiology and gene expression of grasshopper oedaleus asiaticus exposed to diet stress from plant secondary compounds. Sci Rep. 7:8655. [CrossRef]
- Huang X, Suyama K, Buchanan J, Zhu AJ, Scott MP (2005) A drosophila model of the niemann-pick type c lysosome storage disease: Dnpc1a is required for molting and sterol homeostasis. Development. 132:5115-5124. [CrossRef]
- Huang X, Warren JT, Buchanan J, Gilbert LI, Scott MP (2007) Drosophila niemann-pick type c-2 genes control sterol homeostasis and steroid biosynthesis: A model of human neurodegenerative disease. Development. 134:3733-3742.
- Ian De Veau EJ, Schultz JC (1992) Reassessment of interaction between gut detergents and tannins in lepidoptera and significance for gypsy moth larvae. J Chem Ecol. 18:1437-1453. [CrossRef]
- Issaq HJ, Chan KC, Blonder J, Ye X, Veenstra TD (2009) Separation, detection and quantitation of peptides by liquid chromatography and capillary electrochromatography. J Chromatogr A. 1216:1825-1837. [CrossRef]
- Jing TZ, Qi FH, Wang ZY (2020) Most dominant roles of insect gut bacteria: Digestion, detoxification, or essential nutrient provision? Microbiome. 8:38. [CrossRef]
- Jing X, Behmer ST (2020) Insect sterol nutrition: Physiological mechanisms, ecology, and applications. Annu Rev Entomol. 65:251-271. [CrossRef]
- Jing X, Vogel H, Grebenok RJ, Zhu-Salzman K, Behmer ST (2012) Dietary sterols/steroids and the generalist caterpillar helicoverpa zea: Physiology, biochemistry and midgut gene expression. Insect Biochem Mol Biol. 42:835-845. [CrossRef]
- Jouni ZE, Zamora J, Wells MA (2002) Absorption and tissue distribution of cholesterol in manduca sexta. Arch Insect Biochem Physiol. 49:167-175. [CrossRef]
- Jugder BE, Kamareddine L, Watnick PI (2021) Microbiota-derived acetate activates intestinal innate immunity via the tip60 histone acetyltransferase complex. Immunity. 54:1683-1697 e1683. [CrossRef]
- Jupatanakul N, Sim S, Dimopoulos G (2014) Aedes aegypti ml and niemann-pick type c family members are agonists of dengue virus infection. Dev Comp Immunol. 43:1-9. [CrossRef]
- Jurzitza G (1974) [furnishing of sterol by the yeast-like endosymbionts of lasioderma serricorne f. (coleoptera, anobiidae) and its ecological importance for the host]. Oecologia. 16:163-172. [CrossRef]
- Kamareddine L, Robins WP, Berkey CD, Mekalanos JJ, Watnick PI (2018) The drosophila immune deficiency pathway modulates enteroendocrine function and host metabolism. Cell Metab. 28:449-462 e445. [CrossRef]
- Kesnerova L, Mars RAT, Ellegaard KM, Troilo M, Sauer U, Engel P (2017) Disentangling metabolic functions of bacteria in the honey bee gut. PLoS Biol. 15:e2003467. [CrossRef]
- Kiefer C, Sumser E, Wernet MF, Von Lintig J (2002) A class b scavenger receptor mediates the cellular uptake of carotenoids in drosophila. Proc Natl Acad Sci U S A. 99:10581-10586. [CrossRef]
- Kim MS, Lan Q (2010) Sterol carrier protein-x gene and effects of sterol carrier protein-2 inhibitors on lipid uptake in manduca sexta. BMC Physiol. 10:9. [CrossRef]
- Kim Y, Stanley D (2021) Eicosanoid signaling in insect immunology: New genes and unresolved issues. Genes (Basel). 12. [CrossRef]
- Kirfel G, Komnick H (1999) Differential absorption and esterification of dietary long-chain fatty acids by larvae of the dragonfly, aeshna cyanea. Arch Insect Biochem Physiol. 40:183-193. [CrossRef]
- Ko CW, Qu J, Black DD, Tso P (2020) Regulation of intestinal lipid metabolism: Current concepts and relevance to disease. Nat Rev Gastroenterol Hepatol. 17:169-183. [CrossRef]
- Komnick H, Bauerfeind R (1991) Intestinal absorption of defined lipids by the larval dragonfly aeshna cyanea (insecta, odonata): Wax ester and fatty alcohols. J Insect Physiol. 37:179-191.
- Kong HG, Kim HH, Chung JH, Jun J, Lee S, Kim HM, Jeon S, Park SG, Bhak J, Ryu CM (2019) The galleria mellonella hologenome supports microbiota-independent metabolism of long-chain hydrocarbon beeswax. Cell Rep. 26:2451-2464 e2455. [CrossRef]
- Kuhnlein RP (2012) Thematic review series: Lipid droplet synthesis and metabolism: From yeast to man. Lipid droplet-based storage fat metabolism in drosophila. J Lipid Res. 53:1430-1436.
- Kuhns EH, Seidl-Adams I, Tumlinson JH (2012) A lepidopteran aminoacylase (l-acy-1) in heliothis virescens (lepidoptera: Noctuidae) gut lumen hydrolyzes fatty acid-amino acid conjugates, elicitors of plant defense. Insect Biochem Mol Biol. 42:32-40. [CrossRef]
- Kunding AH, Christensen SM, Danielsen EM, Hansen GH (2010) Domains of increased thickness in microvillar membranes of the small intestinal enterocyte. Mol Membr Biol. 27:170-177. [CrossRef]
- Kvietys PR, Specian RD, Grisham MB, Tso P (1991) Jejunal mucosal injury and restitution: Role of hydrolytic products of food digestion. Am J Physiol. 261:G384-391. [CrossRef]
- Kwong WK, Medina LA, Koch H, Sing KW, Soh EJY, Ascher JS, Jaffe R, Moran NA (2017) Dynamic microbiome evolution in social bees. Sci Adv. 3:e1600513. [CrossRef]
- LaJeunesse DR, Johnson B, Presnell JS, Catignas KK, Zapotoczny G (2010) Peristalsis in the junction region of the drosophila larval midgut is modulated by dh31 expressing enteroendocrine cells. BMC Physiol. 10:14. [CrossRef]
- Lan Q, Massey RJ (2004) Subcellular localization of the mosquito sterol carrier protein-2 and sterol carrier protein-x. J Lipid Res. 45:1468-1474. [CrossRef]
- Lan Q, Wessely V (2004) Expression of a sterol carrier protein-x gene in the yellow fever mosquito, aedes aegypti. Insect Mol Biol. 13:519-529. [CrossRef]
- Lee WJ, Hase K (2014) Gut microbiota-generated metabolites in animal health and disease. Nat Chem Biol. 10:416-424. [CrossRef]
- Lehane MJ (1977) Transcellular absorption of lipids in the midgut of the stablefly, stomoxys calcitrans. J Insect Physiol. 23:945-954. [CrossRef]
- Lehane MJ (1997) Peritrophic matrix structure and function. Annu Rev Entomol. 42:525-550. [CrossRef]
- Li S, Jing X (2020) Fates of dietary sterols in the insect alimentary canal. Curr Opin Insect Sci. 41:106-111. [CrossRef]
- Lin X, Wen X, Wei Z, Guo K, Shi F, Huang T, Wang W, Zheng J (2021) Vitamin k2 protects against abeta42-induced neurotoxicity by activating autophagy and improving mitochondrial function in drosophila. Neuroreport. 32:431-437. [CrossRef]
- Listenberger LL, Han X, Lewis SE, Cases S, Farese RV, Jr., Ory DS, Schaffer JE (2003) Triglyceride accumulation protects against fatty acid-induced lipotoxicity. Proc Natl Acad Sci U S A. 100:3077-3082. [CrossRef]
- Male KB, Storey KB (1981) Enzyme activities and isozyme composition of triglyceride, diglyceride and monoglyceride lipases in periplaneta americana, locusta migratoria and polia adjuncta. Insect Biochem. 11:423-427.
- Marianes A, Spradling AC (2013) Physiological and stem cell compartmentalization within the drosophila midgut. Elife. 2:e00886. [CrossRef]
- Markvoort AJ, Marrink SJ (2011) Lipid acrobatics in the membrane fusion arena. Curr Top Membr. 68:259-294. [CrossRef]
- Matic D, Vlahovic M, Ilijin L, Grcic A, Filipovic A, Todorovic D, Peric-Mataruga V (2021) Implications of long-term exposure of a lymantria dispar l. Population to pollution for the response of larval midgut proteases and acid phosphatases to chronic cadmium treatment. Comp Biochem Physiol C Toxicol Pharmacol. 250:109172. [CrossRef]
- Mattila J, Havula E, Suominen E, Teesalu M, Surakka I, Hynynen R, Kilpinen H, Vaananen J, Hovatta I, Kakela R et al. (2015) Mondo-mlx mediates organismal sugar sensing through the gli-similar transcription factor sugarbabe. Cell Rep. 13:350-364.
- McFarlane JE (1976) Vitamin k: Growth factor for the house cricket (orthoptera: Gryllidae). Can Entomol. 108:391-394.
- McFarlane JE, Alli I (1985) Volatile fatty acids of frass of certain omnivorous insects. J Chem Ecol. 11:59-63. [CrossRef]
- Mei Y, Jing D, Tang S, Chen X, Chen H, Duanmu H, Cong Y, Chen M, Ye X, Zhou H et al. (2022) Insectbase 2.0: A comprehensive gene resource for insects. Nucleic Acids Res. 50:D1040-D1045. [CrossRef]
- Meikle JES, McFarlane JE (1965) The role of lipid in the nutrition of the house cricket, acheta domesticus l. Can J Zool. 43:87-98.
- Merritt RW, Dadd RH, Walker ED (1992) Feeding behavior, natural food, and nutritional relationships of larval mosquitoes. Annu Rev Entomol. 37:349-376. [CrossRef]
- Miao Z, Cao X, Jiang H (2020) Digestion-related proteins in the tobacco hornworm, manduca sexta. Insect Biochem Mol Biol. 126:103457. [CrossRef]
- Miguel-Aliaga I, Jasper H, Lemaitre B (2018) Anatomy and physiology of the digestive tract of drosophila melanogaster. Genetics. 210:357-396. [CrossRef]
- Miled N, Canaan S, Dupuis L, Roussel A, Riviere M, Carriere F, de Caro A, Cambillau C, Verger R (2000) Digestive lipases: From three-dimensional structure to physiology. Biochimie. 82:973-986. [CrossRef]
- Milger K, Herrmann T, Becker C, Gotthardt D, Zickwolf J, Ehehalt R, Watkins PA, Stremmel W, Fullekrug J (2006) Cellular uptake of fatty acids driven by the er-localized acyl-coa synthetase fatp4. J Cell Sci. 119:4678-4688. [CrossRef]
- Moadeli T, Mainali B, Ponton F, Taylor PW (2020) Effects of fatty acids and vitamin e in larval diets on development and performance of queensland fruit fly. J Insect Physiol. 125:104058. [CrossRef]
- Montigny C, Lyons J, Champeil P, Nissen P, Lenoir G (2016) On the molecular mechanism of flippase- and scramblase-mediated phospholipid transport. Biochim Biophys Acta. 1861:767-783. [CrossRef]
- Moran NA, Jarvik T (2010) Lateral transfer of genes from fungi underlies carotenoid production in aphids. Science. 328:624-627. [CrossRef]
- Nassir F, Wilson B, Han X, Gross RW, Abumrad NA (2007) Cd36 is important for fatty acid and cholesterol uptake by the proximal but not distal intestine. J Biol Chem. 282:19493-19501. [CrossRef]
- Nauli AM, Nassir F, Zheng S, Yang Q, Lo CM, Vonlehmden SB, Lee D, Jandacek RJ, Abumrad NA, Tso P (2006) Cd36 is important for chylomicron formation and secretion and may mediate cholesterol uptake in the proximal intestine. Gastroenterology. 131:1197-1207. [CrossRef]
- Neira Oviedo M, Vanekeris L, Corena-McLeod MD, Linser PJ (2008) A microarray-based analysis of transcriptional compartmentalization in the alimentary canal of anopheles gambiae (diptera: Culicidae) larvae. Insect Mol Biol. 17:61-72. [CrossRef]
- Niot I, Poirier H, Tran TT, Besnard P (2009) Intestinal absorption of long-chain fatty acids: Evidence and uncertainties. Prog Lipid Res. 48:101-115. [CrossRef]
- Nor Aliza AR, Othman M, Sabri M, Stanley DW (2018) A midgut digestive phospholipase a2 in larval mosquitoes, aedes albopictus and culex quinquefasciatus. Enzyme Res. 2018:9703413. [CrossRef]
- Nor Aliza AR, Stanley D (1998) A digestive phospholipase a2 in larval mosquitoes, aedes aegypti. Insect Biochem Mol Biol. 28:561-569.
- Noriega FG (2014) Juvenile hormone biosynthesis in insects: What is new, what do we know, and what questions remain? Int Sch Res Notices. 2014:967361. [CrossRef]
- Palm W, Sampaio JL, Brankatschk M, Carvalho M, Mahmoud A, Shevchenko A, Eaton S (2012) Lipoproteins in drosophila melanogaster--assembly, function, and influence on tissue lipid composition. PLoS Genet. 8:e1002828. [CrossRef]
- Parvy JP, Napal L, Rubin T, Poidevin M, Perrin L, Wicker-Thomas C, Montagne J (2012) Drosophila melanogaster acetyl-coa-carboxylase sustains a fatty acid-dependent remote signal to waterproof the respiratory system. PLoS Genet. 8:e1002925. [CrossRef]
- Peller CR, Bacon EM, Bucheger JA, Blumenthal EM (2009) Defective gut function in drop-dead mutant drosophila. J Insect Physiol. 55:834-839. [CrossRef]
- Pennington JE, Wells MA (2002) Triacylglycerol-rich lipophorins are found in the dipteran infraorder culicomorpha, not just in mosquitoes. J Insect Sci. 2:15. [CrossRef]
- Perez-Salas U, Garg S, Gerelli Y, Porcar L (2021) Deciphering lipid transfer between and within membranes with time-resolved small-angle neutron scattering. Curr Top Membr. 88:359-412. [CrossRef]
- Petersen TN, Brunak S, von Heijne G, Nielsen H (2011) Signalp 4.0: Discriminating signal peptides from transmembrane regions. Nat Methods. 8:785-786. [CrossRef]
- Plateau F (1876). Les phenomenes de la digestion chez la blatte americaine (periplaneta americana). In: Hayez F, editor.: Imprimeur de l’Académie Royale de Belgique.
- Rana RL, Hoback WW, Rahim NAR, Bedick J, Stanley DW (1997) Pre-oral digestion: A phospholipase a2 associated with oral secretions in adult burying beetles, nicrophorus marginatus. Comp Biochem Physiol. 118:375-380.
- Rana RL, Stanley DW (1999) In vitro secretion of digestive phospholipase a(2) by midguts isolated from tobacco hornworms, manduca sexta. Arch Insect Biochem Physiol. 42:179-187. [CrossRef]
- Reboul E, Klein A, Bietrix F, Gleize B, Malezet-Desmoulins C, Schneider M, Margotat A, Lagrost L, Collet X, Borel P (2006) Scavenger receptor class b type i (sr-bi) is involved in vitamin e transport across the enterocyte. J Biol Chem. 281:4739-4745. [CrossRef]
- Reiher W, Shirras C, Kahnt J, Baumeister S, Isaac RE, Wegener C (2011) Peptidomics and peptide hormone processing in the drosophila midgut. J Proteome Res. 10:1881-1892. [CrossRef]
- Renne MF, Hariri H (2021) Lipid droplet-organelle contact sites as hubs for fatty acid metabolism, trafficking, and metabolic channeling. Front Cell Dev Biol. 9:726261. [CrossRef]
- Rewitz KF, Rybczynski R, Warren JT, Gilbert LI (2006) The halloween genes code for cytochrome p450 enzymes mediating synthesis of the insect moulting hormone. Biochem Soc Trans. 34:1256-1260.
- Ribeiro JM, Genta FA, Sorgine MH, Logullo R, Mesquita RD, Paiva-Silva GO, Majerowicz D, Medeiros M, Koerich L, Terra WR et al. (2014) An insight into the transcriptome of the digestive tract of the bloodsucking bug, rhodnius prolixus. PLoS Negl Trop Dis. 8:e2594. [CrossRef]
- Rietveld A, Neutz S, Simons K, Eaton S (1999) Association of sterol- and glycosylphosphatidylinositol-linked proteins with drosophila raft lipid microdomains. J Biol Chem. 274:12049-12054. [CrossRef]
- Rodriguez-Vazquez M, Vaquero D, Parra-Peralbo E, Mejia-Morales JE, Culi J (2015) Drosophila lipophorin receptors recruit the lipoprotein ltp to the plasma membrane to mediate lipid uptake. PLoS Genet. 11:e1005356. [CrossRef]
- Rost-Roszkowska MM, Pilka M, Szymska R, Klag J (2007) Ultrastructural studies of midgut epithelium formation in lepisma saccharina l. (insecta, zygentoma). J Morphol. 268:224-231. [CrossRef]
- Rudin W, Hecker H (1979) Functional morphology of the midgut of aedes aegypti l. (insecta, diptera) during blood digestion. Cell Tissue Res. 200:193-203. [CrossRef]
- Sahaka M, Amara S, Wattanakul J, Gedi MA, Aldai N, Parsiegla G, Lecomte J, Christeller JT, Gray D, Gontero B et al. (2020) The digestion of galactolipids and its ubiquitous function in nature for the uptake of the essential alpha-linolenic acid. Food Funct. 11:6710-6744. [CrossRef]
- Sajjadian SM, Vatanparast M, Stanley D, Kim Y (2019) Secretion of secretory phospholipase a2 into spodoptera exigua larval midgut lumen and its role in lipid digestion. Insect Mol Biol. 28:773-784. [CrossRef]
- Santos HP, Rost-Roszkowska M, Vilimova J, Serrao JE (2017) Ultrastructure of the midgut in heteroptera (hemiptera) with different feeding habits. Protoplasma. 254:1743-1753. [CrossRef]
- Schmidt K, Engel P (2021) Mechanisms underlying gut microbiota-host interactions in insects. J Exp Biol. 224. [CrossRef]
- Shen LR, Lai CQ, Feng X, Parnell LD, Wan JB, Wang JD, Li D, Ordovas JM, Kang JX (2010) Drosophila lacks c20 and c22 pufas. J Lipid Res. 51:2985-2992. [CrossRef]
- Shin SC, Kim SH, You H, Kim B, Kim AC, Lee KA, Yoon JH, Ryu JH, Lee WJ (2011) Drosophila microbiome modulates host developmental and metabolic homeostasis via insulin signaling. Science. 334:670-674. [CrossRef]
- Shukla SP, Plata C, Reichelt M, Steiger S, Heckel DG, Kaltenpoth M, Vilcinskas A, Vogel H (2018) Microbiome-assisted carrion preservation aids larval development in a burying beetle. Proc Natl Acad Sci U S A. 115:11274-11279. [CrossRef]
- Siddiqi S, Sheth A, Patel F, Barnes M, Mansbach CM, 2nd (2013) Intestinal caveolin-1 is important for dietary fatty acid absorption. Biochim Biophys Acta. 1831:1311-1321. [CrossRef]
- Sieber MH, Thummel CS (2009) The dhr96 nuclear receptor controls triacylglycerol homeostasis in drosophila. Cell Metab. 10:481-490.
- Sieber MH, Thummel CS (2012) Coordination of triacylglycerol and cholesterol homeostasis by dhr96 and the drosophila lipa homolog magro. Cell Metab. 15:122-127. [CrossRef]
- Silva W, Cardoso C, Ribeiro AF, Terra WR, Ferreira C (2013) Midgut proteins released by microapocrine secretion in spodoptera frugiperda. J Insect Physiol. 59:70-80. [CrossRef]
- Smith AF, Tsuchida K, Hanneman E, Suzuki TC, Wells MA (1992) Isolation, characterization, and cdna sequence of two fatty acid-binding proteins from the midgut of manduca sexta larvae. J Biol Chem. 267:380-384.
- Somerville HJ, Pockett HV (1976) Phospholipase activity in gut juice of lepidopterous larvae. Insect Biochem. 6:351-353.
- Song W, Veenstra JA, Perrimon N (2014) Control of lipid metabolism by tachykinin in drosophila. Cell Rep. 9:40-47. [CrossRef]
- Spates GE, Bull DL, Chen AC (1990) Hydrolysis of sphingomyelin and phosphatidylcholine by midgut homogenates of the stable fly. Arch Insect Biochem Physiol. 14:1-12. [CrossRef]
- Spiteller D, Boland W (2003) N-(17-acyloxy-acyl)-glutamines: Novel surfactants from oral secretions of lepidopteran larvae. J Org Chem. 68:8743-8749. [CrossRef]
- Spiteller D, Dettner K, Boland W (2000) Gut bacteria may be involved in interactions between plants, herbivores and their predators: Microbial biosynthesis of n-acylglutamine surfactants as elicitors of plant volatiles. Biol Chem. 381:755-762.
- Stahl A, Hirsch DJ, Gimeno RE, Punreddy S, Ge P, Watson N, Patel S, Kotler M, Raimondi A, Tartaglia LA et al. (1999) Identification of the major intestinal fatty acid transport protein. Mol Cell. 4:299-308. [CrossRef]
- Stanley-Samuelson DW (1994) Prostaglandins and related eicosanoids in insects. Adv Insect Physiol. 24:115-212.
- Stanley D (2006) The non-venom insect phospholipases a2. Biochim Biophys Acta. 1761:1383-1390. [CrossRef]
- Stanley DW, Sarath G, Rana RL (1998) A digestive phospholipase a(2) in midguts of tobacco hornworms, manduca sexta l. J Insect Physiol. 44:297-303. [CrossRef]
- Stark AH, Crawford MA, Reifen R (2008) Update on alpha-linolenic acid. Nutr Rev. 66:326-332. [CrossRef]
- Storelli G, Defaye A, Erkosar B, Hols P, Royet J, Leulier F (2011) Lactobacillus plantarum promotes drosophila systemic growth by modulating hormonal signals through tor-dependent nutrient sensing. Cell Metab. 14:403-414. [CrossRef]
- Storelli G, Strigini M, Grenier T, Bozonnet L, Schwarzer M, Daniel C, Matos R, Leulier F (2018) Drosophila perpetuates nutritional mutualism by promoting the fitness of its intestinal symbiont lactobacillus plantarum. Cell Metab. 27:362-377 e368. [CrossRef]
- Svoboda JA (1999) Variability of metabolism and function of sterols in insects. Crit Rev Biochem Mol Biol. 34:49-57. [CrossRef]
- Tamaki FK, Pimentel AC, Dias AB, Cardoso C, Ribeiro AF, Ferreira C, Terra WR (2014) Physiology of digestion and the molecular characterization of the major digestive enzymes from periplaneta americana. J Insect Physiol. 70:22-35. [CrossRef]
- Teixeira A, Fialho Mdo C, Zanuncio JC, Ramalho Fde S, Serrao JE (2013) Degeneration and cell regeneration in the midgut of podisus nigrispinus (heteroptera: Pentatomidae) during post-embryonic development. Arthropod Struct Dev. 42:237-246. [CrossRef]
- Terra WR (2001) The origin and functions of the insect peritrophic membrane and peritrophic gel. Arch Insect Biochem Physiol. 47:47-61. [CrossRef]
- Terra WR, Costa RH, Ferreira C (2006) Plasma membranes from insect midgut cells. An Acad Bras Cienc. 78:255-269. [CrossRef]
- Terra WR, Ferreira C (2020) Evolutionary trends of digestion and absorption in the major insect orders. Arthropod Struct Dev. 56:100931. [CrossRef]
- Terra WR, Ferreira C, Garcia ES (1988) Origin, distribution, properties and functiosn of the major rhodnius prolixus midgut hydrolases. Insect Biochem. 18:423-434.
- Tinker KA, Ottesen EA (2016) The core gut microbiome of the american cockroach, periplaneta americana, is stable and resilient to dietary shifts. Appl Environ Microbiol. 82:6603-6610. [CrossRef]
- Toprak U, Hegedus D, Dogan C, Guney G (2020) A journey into the world of insect lipid metabolism. Arch Insect Biochem Physiol. 104:e21682. [CrossRef]
- Treherne JE (1958) The digestion and absorption of tripalmitin in the cockroach, periplanata americana l. J Exp Biol. 35:862-870.
- Trigatti BL, Anderson RG, Gerber GE (1999) Identification of caveolin-1 as a fatty acid binding protein. Biochem Biophys Res Commun. 255:34-39. [CrossRef]
- Tsuchida K, Wells MA (1988) Digestion, absoprtion, transport and storage of fat during the last larval stadium of manduca sexta. Changes in the role of lipophorin in the delivery of dietary lipid to the fat body. Insect Biochem. 18:263-268.
- Turunen S (1975) Absorption and transport of dietary lipid in pieris brassicae. J Insect Physiol. 21:1521-1529.
- Turunen S (1988a) Digestion and absorption of glycerophospholipid in pieris brassicae. Comp Biochem Physiol. 89A:19-24.
- Turunen S (1988b) Uptake of dietary lipids: A novel pathway in pieris brassicae. Insect Biochem. 18:499-505.
- Turunen S (1990) Absorption of choline, myo-inositol, and oleic acid in th emidgut of pieris brassicae: Sectional differentiation and uptake into the haemolymph. J Insect Physiol. 36:737-741.
- Turunen S (1993) Metabolic pathways in the midgut epithelium of pieris brassicae during carbohydrate and lipid assimilation. Insect Biochem Molec Biol. 23:681-689.
- Turunen S, Chippendale GM (1977) Lipid absorption and transport: Sectional analysis of the larval midgut of the corn borer, diatraea grandiosella. Insect Biochem. 7:203-208.
- Turunen S, Chippendale GM (1989) Relationship between dietary lipids, midgut lipids, and lipid absorption in eight species of lepidoptera reared on artificial and natural diets J Insect Physiol. 35:627-633.
- Turunen S, Kastari T (1979) Digestion and absorption of lecithin in larvae of the cabbage butterfly, pieris brassicae. Comp Biochem Physiol. 62A:933-937.
- Uscian JM, Miller JS, Sarah G, Stanley-Samuelson DW (1995) A digestive phospholipase a2 in the tiger beetle cicindella circumpicta. J Insect Physiol. 41:135-141.
- Valzania L, Coon KL, Vogel KJ, Brown MR, Strand MR (2018) Hypoxia-induced transcription factor signaling is essential for larval growth of the mosquito aedes aegypti. Proc Natl Acad Sci U S A. 115:457-465. [CrossRef]
- Vance JE (2015) Phospholipid synthesis and transport in mammalian cells. Traffic. 16:1-18. [CrossRef]
- Vasconcelos MA, Orsolin PC, Oliveira VC, Lima P, Naves MPC, de Morais CR, Nicolau-Junior N, Bonetti AM, Spano MA (2020) Modulating effect of vitamin d3 on the mutagenicity and carcinogenicity of doxorubicin in drosophila melanogaster and in silico studies. Food Chem Toxicol. 143:111549. [CrossRef]
- Vatanparast M, Ahmed S, Herrero S, Kim Y (2018) A non-venomous spla2 of a lepidopteran insect: Its physiological functions in development and immunity. Dev Comp Immunol. 89:83-92. [CrossRef]
- Veenstra JA (2009) Peptidergic paracrine and endocrine cells in the midgut of the fruit fly maggot. Cell Tissue Res. 336:309-323. [CrossRef]
- Veenstra JA, Agricola HJ, Sellami A (2008) Regulatory peptides in fruit fly midgut. Cell Tissue Res. 334:499-516. [CrossRef]
- Verger R, Mieras MC, de Haas GH (1973) Action of phospholipase a at interfaces. J Biol Chem. 248:4023-4034.
- Visotto LE, Oliveira MG, Guedes RN, Ribon AO, Good-God PI (2009) Contribution of gut bacteria to digestion and development of the velvetbean caterpillar, anticarsia gemmatalis. J Insect Physiol. 55:185-191. [CrossRef]
- Vogel H, Shukla SP, Engl T, Weiss B, Fischer R, Steiger S, Heckel DG, Kaltenpoth M, Vilcinskas A (2017a) The digestive and defensive basis of carcass utilization by the burying beetle and its microbiota. Nat Commun. 8:15186. [CrossRef]
- Vogel KJ, Valzania L, Coon KL, Brown MR, Strand MR (2017b) Transcriptome sequencing reveals large-scale changes in axenic aedes aegypti larvae. PLoS Negl Trop Dis. 11:e0005273. [CrossRef]
- Voght SP, Fluegel ML, Andrews LA, Pallanck LJ (2007) Drosophila npc1b promotes an early step in sterol absorption from the midgut epithelium. Cell Metab. 5:195-205. [CrossRef]
- Volonte D, Galbiati F, Li S, Nishiyama K, Okamoto T, Lisanti MP (1999) Flotillins/cavatellins are differentially expressed in cells and tissues and form a hetero-oligomeric complex with caveolins in vivo. Characterization and epitope-mapping of a novel flotillin-1 monoclonal antibody probe. J Biol Chem. 274:12702-12709. [CrossRef]
- Vos M, Esposito G, Edirisinghe JN, Vilain S, Haddad DM, Slabbaert JR, Van Meensel S, Schaap O, De Strooper B, Meganathan R et al. (2012) Vitamin k2 is a mitochondrial electron carrier that rescues pink1 deficiency. Science. 336:1306-1310. [CrossRef]
- Wada-Katsumata A, Zurek L, Nalyanya G, Roelofs WL, Zhang A, Schal C (2015) Gut bacteria mediate aggregation in the german cockroach. Proc Natl Acad Sci U S A. 112:15678-15683. [CrossRef]
- Wegener C, Veenstra JA (2015) Chemical identity, function and regulation of enteroendocrine peptides in insects. Curr Opin Insect Sci. 11:8-13. [CrossRef]
- Weidlich S, Hoffmann KH, Woodring J (2015) Secretion of lipases in the digestive tract of the cricket gryllus bimaculatus. Arch Insect Biochem Physiol. 90:209-217. [CrossRef]
- Weiher B, Komnick H (1997) Digestion of phosphatidylcholines, absorption, and esterification of lipolytic products by aeshna cyanea larvae as studied in vivo and in vitro. Arch Insect Biochem Physiol. 36:273-293. [CrossRef]
- Weintraub H, Tietz A (1973) Triglyceride digestion and absorption in the locust, locusta migratoria. Biochim Biophys Acta. 306:31-41. [CrossRef]
- Weintraub H, Tietz A (1978) Lipid absorption by isolated intestinal preparations. Insect Biochem. 8:267-274.
- Wicker-Thomas C, Garrido D, Bontonou G, Napal L, Mazuras N, Denis B, Rubin T, Parvy JP, Montagne J (2015) Flexible origin of hydrocarbon/pheromone precursors in drosophila melanogaster. J Lipid Res. 56:2094-2101. [CrossRef]
- Woodring J, Lorenz MW (2007) Feeding, nutrient flow, and functional gut morphology in the cricket gryllus bimaculatus. J Morphol. 268:815-825. [CrossRef]
- Yamanaka N, Rewitz KF, O'Connor MB (2013) Ecdysone control of developmental transitions: Lessons from drosophila research. Annu Rev Entomol. 58:497-516.
- Yoshinaga N, Aboshi T, Abe H, Nishida R, Alborn HT, Tumlinson JH, Mori N (2008) Active role of fatty acid amino acid conjugates in nitrogen metabolism in spodoptera litura larvae. Proc Natl Acad Sci U S A. 105:18058-18063. [CrossRef]
- Yun HK, Jouni ZE, Wells MA (2002) Characterization of cholesterol transport from midgut to fat body in manduca sexta larvae. Insect Biochem Mol Biol. 32:1151-1158. [CrossRef]
- Zeng X, Hou SX (2015) Enteroendocrine cells are generated from stem cells through a distinct progenitor in the adult drosophila posterior midgut. Development. 142:644-653. [CrossRef]
- Zhang T, Yuan D, Xie J, Lei Y, Li J, Fang G, Tian L, Liu J, Cui Y, Zhang M et al. (2019) Evolution of the cholesterol biosynthesis pathway in animals. Mol Biol Evol. [CrossRef]
- Zhao X, Karpac J (2020) The drosophila midgut and the systemic coordination of lipid-dependent energy homeostasis. Curr Opin Insect Sci. 41:100-105. [CrossRef]
- Zheng H, Powell JE, Steele MI, Dietrich C, Moran NA (2017) Honeybee gut microbiota promotes host weight gain via bacterial metabolism and hormonal signaling. Proc Natl Acad Sci U S A. 114:4775-4780. [CrossRef]
- Zheng JC, Sun SL, Yue XR, Liu TX, Jing X (2018) Phylogeny and evolution of the cholesterol transporter npc1 in insects. J Insect Physiol. 107:157-166. [CrossRef]
- Zinke I, Schutz CS, Katzenberger JD, Bauer M, Pankratz MJ (2002) Nutrient control of gene expression in drosophila: Microarray analysis of starvation and sugar-dependent response. EMBO J. 21:6162-6173. [CrossRef]
- Zwick RK, Ohlstein B, Klein OD (2019) Intestinal renewal across the animal kingdom: Comparing stem cell activity in mouse and drosophila. Am J Physiol Gastrointest Liver Physiol. 316:G313-G322. [CrossRef]
Figure 1.
The insect digestive insect. (A) Scheme of an insect digestive tract. The PM (black dashed line) allows bidirectional fluxes (white arrows). (B) The epithelium of the midgut comprises three main cell types: intestinal stem cells, enteroendocrine cells and columnar cells.
Figure 1.
The insect digestive insect. (A) Scheme of an insect digestive tract. The PM (black dashed line) allows bidirectional fluxes (white arrows). (B) The epithelium of the midgut comprises three main cell types: intestinal stem cells, enteroendocrine cells and columnar cells.
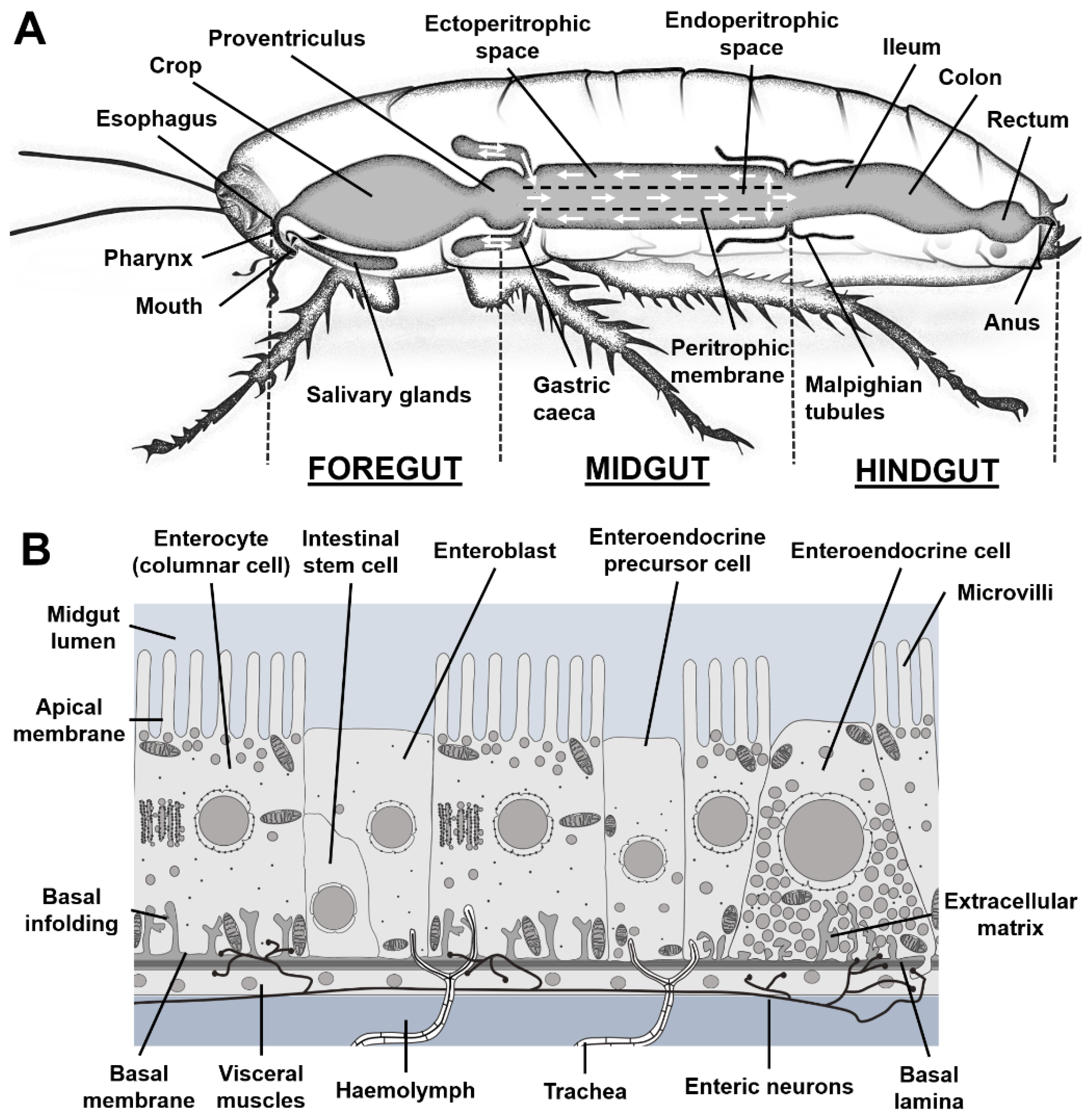
Figure 2.
Essential and non-essential lipids. (A) Galactolipids are the main components of membrane thylakoids; R1 and R2 represent the esterified FA moieties; Gal the galactose residues. (B) Essential and non-essential PUFAs. (C) The synthesis of cholesterol from farnesylpyrophsophate does not operate in insects.
Figure 2.
Essential and non-essential lipids. (A) Galactolipids are the main components of membrane thylakoids; R1 and R2 represent the esterified FA moieties; Gal the galactose residues. (B) Essential and non-essential PUFAs. (C) The synthesis of cholesterol from farnesylpyrophsophate does not operate in insects.
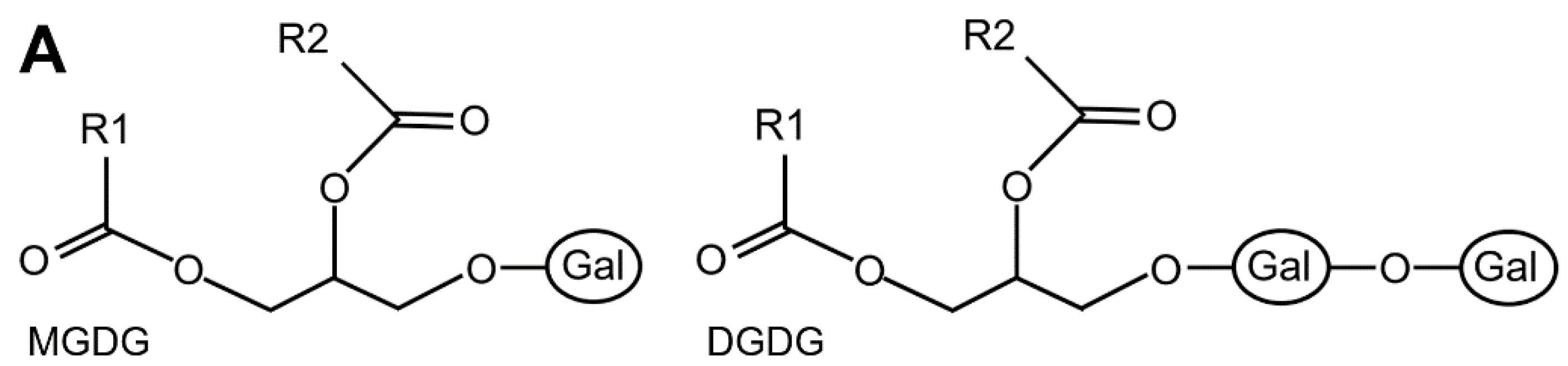
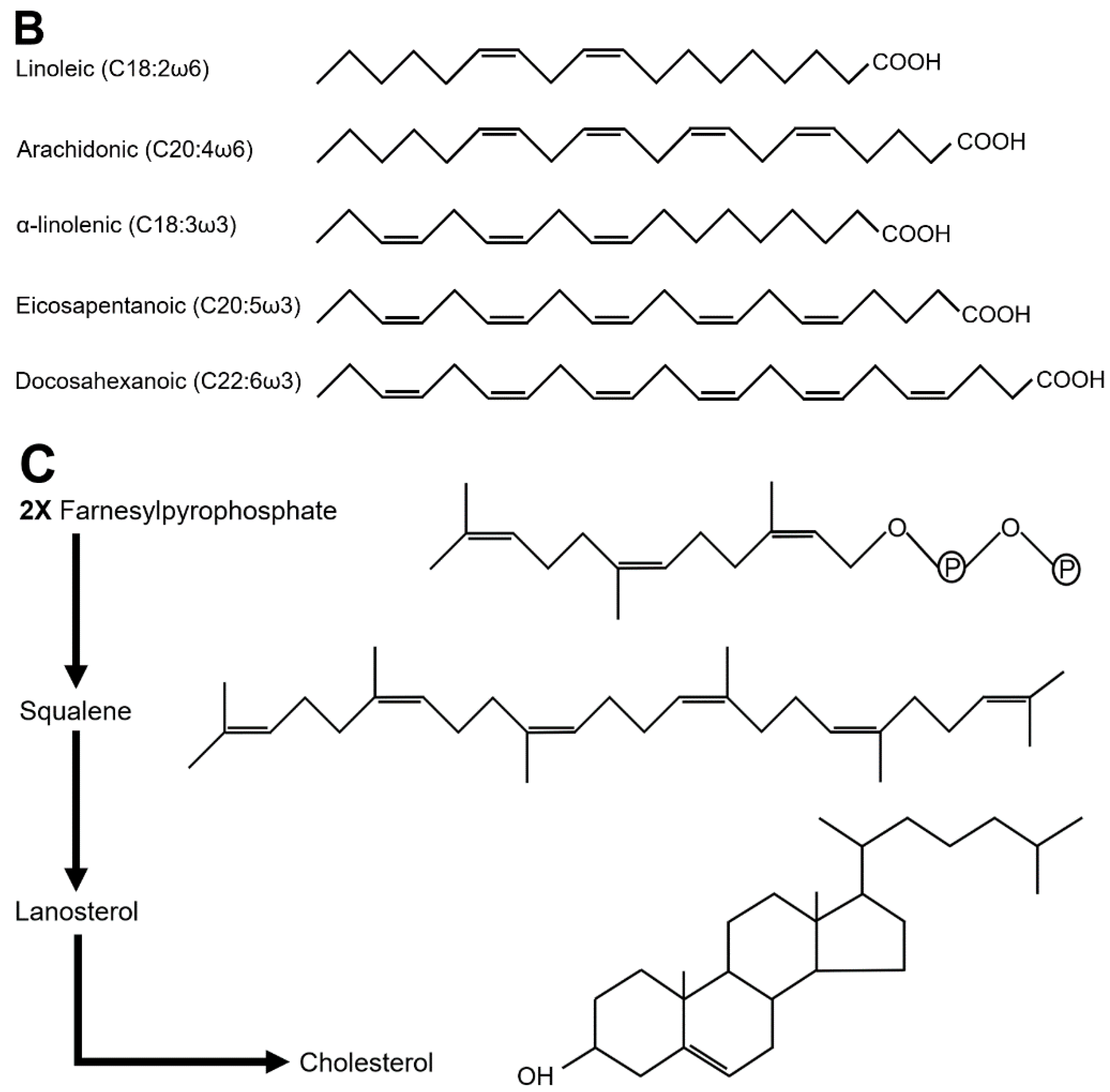
Figure 3.
Lipase activities. (A) TAG lipases hydrolyze the ester bounds (arrowheads) between glycerol and FA moieties (R1, R2, R3). (B) PLA1, PLA2, PLC and PLD cleave phospholipids at specific sites (arrowheads).
Figure 3.
Lipase activities. (A) TAG lipases hydrolyze the ester bounds (arrowheads) between glycerol and FA moieties (R1, R2, R3). (B) PLA1, PLA2, PLC and PLD cleave phospholipids at specific sites (arrowheads).
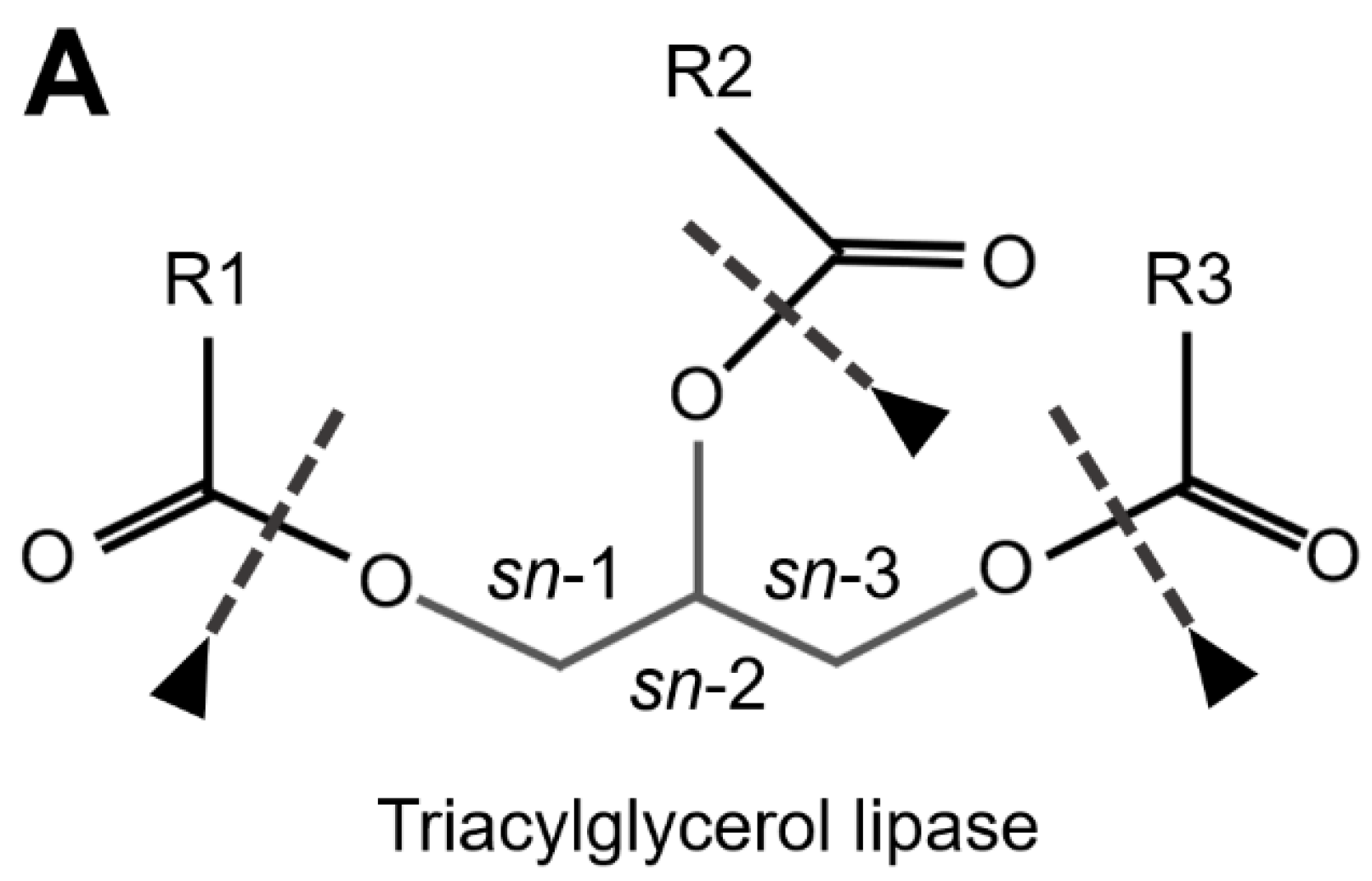
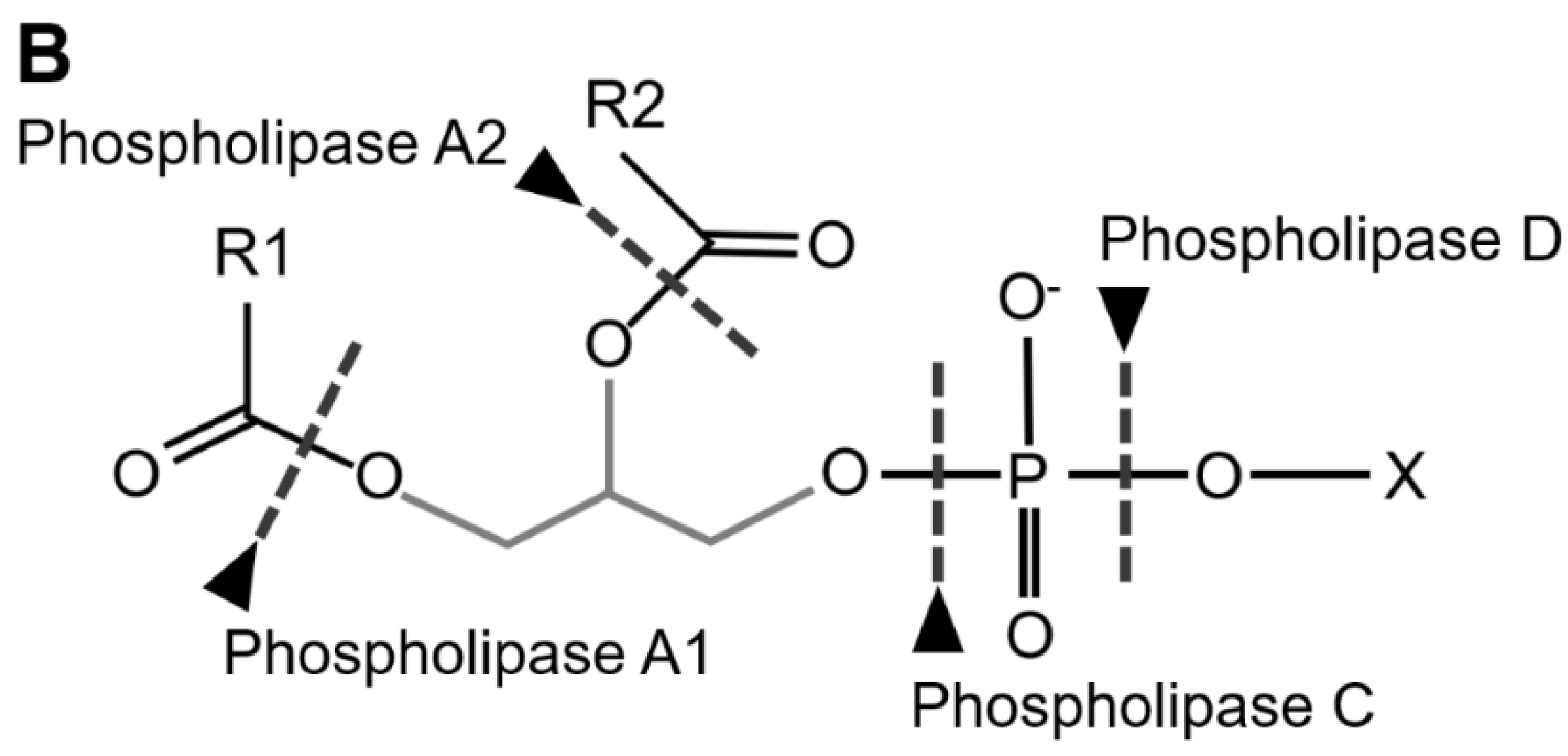
Figure 4.
phospholipid and TAG biosynthesis. The DHAP, Glycerol-3-phosphate and MAG pathways to TAG biosynthesis operate in an organ-specific manner. DAG is the precursor of the phospholipids PC, PE and PS, whereas PA is the precursor of PI, PG and cardiolipin.
Figure 4.
phospholipid and TAG biosynthesis. The DHAP, Glycerol-3-phosphate and MAG pathways to TAG biosynthesis operate in an organ-specific manner. DAG is the precursor of the phospholipids PC, PE and PS, whereas PA is the precursor of PI, PG and cardiolipin.
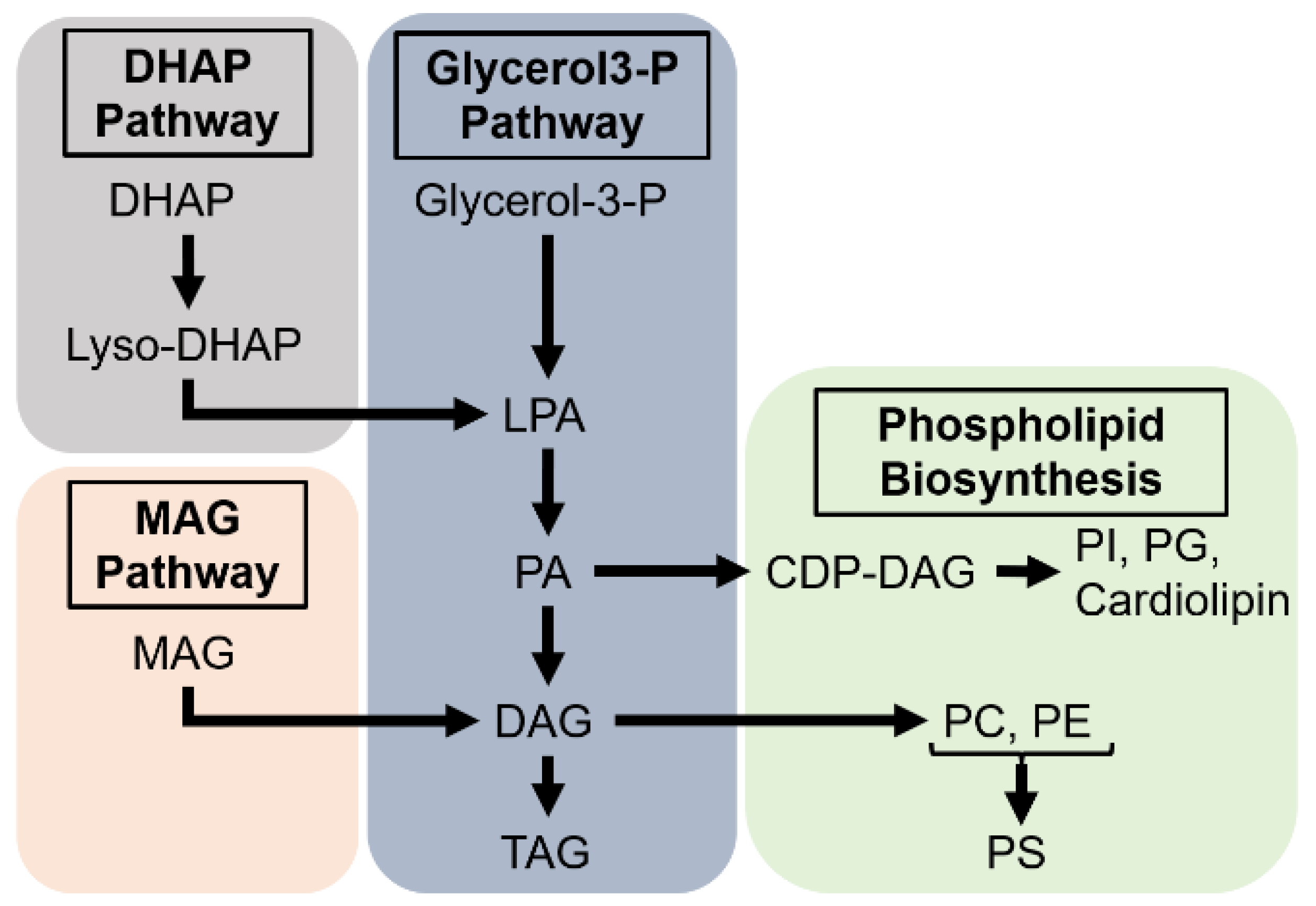
Figure 5.
Lipid digestive processes of insects. Enzymatic digestion of dietary lipids mostly takes place in the lumen. Although lipid uptake has been proposed to proceed passively, specific transporters have been reported. In the enterocytes, lipids may be used to sustain homeostasis, stored in specialized organelles or exported via Lpp via the haemolymph.
Figure 5.
Lipid digestive processes of insects. Enzymatic digestion of dietary lipids mostly takes place in the lumen. Although lipid uptake has been proposed to proceed passively, specific transporters have been reported. In the enterocytes, lipids may be used to sustain homeostasis, stored in specialized organelles or exported via Lpp via the haemolymph.
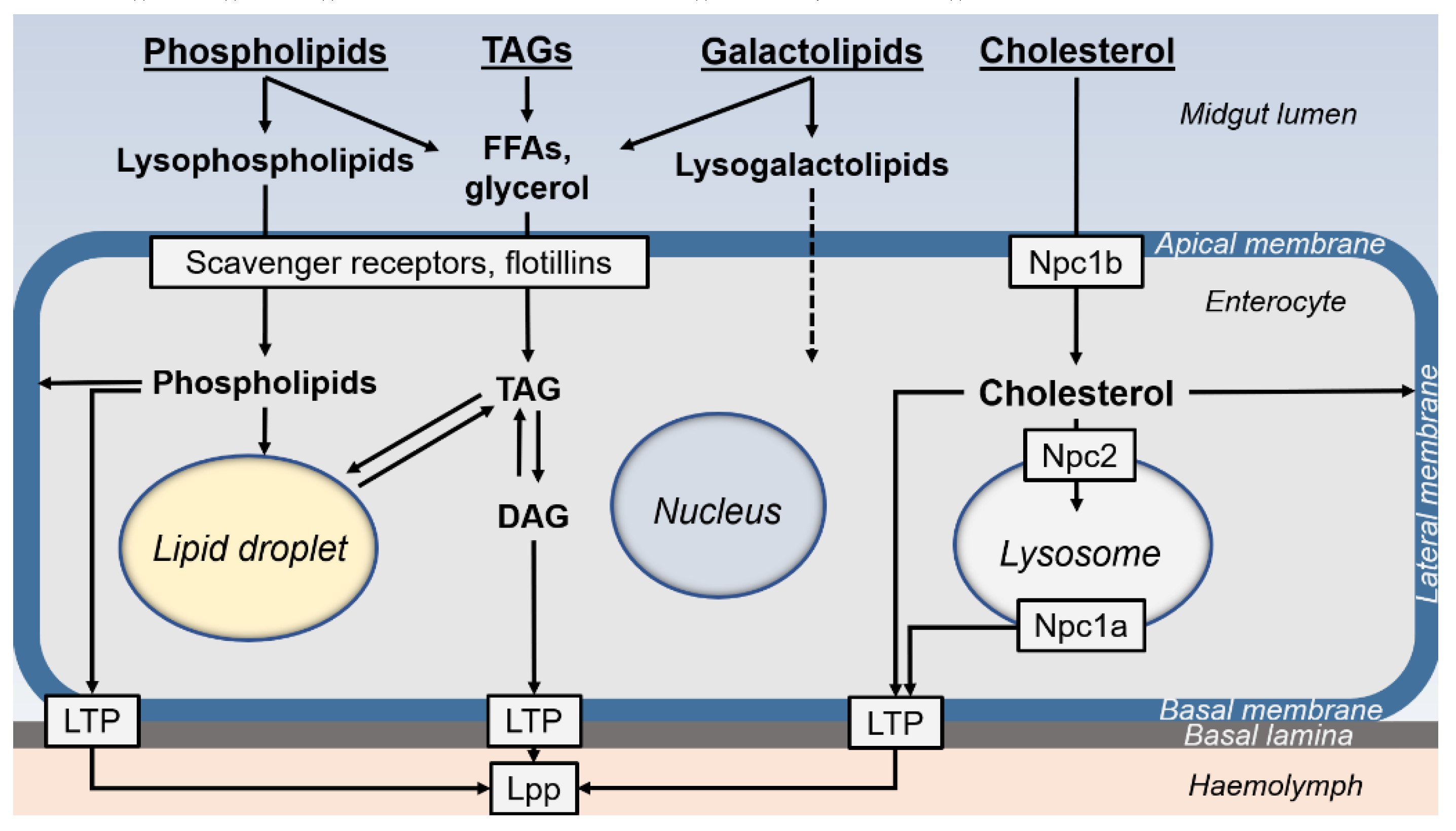

Table 1.
Number of lipases per class in various insect species. These numbers have been established using Flybase (Flybase 2003) and insectBase (Mei et al. 2022). Lipases2 that are absent in insects, yolk proteins that contain inactive catalytic lipase sites and most phospholipases are not represented.
Table 1.
Number of lipases per class in various insect species. These numbers have been established using Flybase (Flybase 2003) and insectBase (Mei et al. 2022). Lipases2 that are absent in insects, yolk proteins that contain inactive catalytic lipase sites and most phospholipases are not represented.
| Neutral | Acid | Lipase3 | GDSL | HSL | ATGL | Total | |
| Aedes aegypti | 42 | 23 | 2 | 4 | 1 | 0 | 72 |
| Anopheles gambiae | 23 | 13 | 2 | 5 | 1 | 1 | 45 |
| Apis mellifera | 14 | 5 | 1 | 5 | 1 | 1 | 27 |
| Bombyx mori | 25 | 17 | 1 | 1 | 1 | 1 | 46 |
| Cimex lectularius | 14 | 12 | 2 | 1 | 1 | 2 | 32 |
| Drosophila melanogaster | 37 | 25 | 1 | 2 | 1 | 2 | 68 |
| Locusta migratoria | 22 | 14 | 2 | 3 | 1 | 1 | 43 |
| Manduca sexta | 45 | 28 | 1 | 1 | 1 | 1 | 77 |
| Musca domestica | 54 | 39 | 1 | 6 | 1 | 1 | 102 |
| Rhodnius prolixus | 18 | 6 | 3 | 2 | 1 | 0 | 30 |
| Spodoptera frugiperda | 57 | 22 | 1 | 2 | 1 | 0 | 83 |
| Tenebrio molitor | 28 | 25 | 2 | 1 | 1 | 1 | 58 |
Table 2.
Lipase activities in the midgut lumen. A number of studies (column reference) have investigated enzymatic activities (column activity) in the midgut of various insect species (column species).
Table 2.
Lipase activities in the midgut lumen. A number of studies (column reference) have investigated enzymatic activities (column activity) in the midgut of various insect species (column species).
| Activity | Species | Reference |
| TAG to sn-2-MAG |
Periplaneta americana; Blabera craniifer Pieris brassicae Periplaneta americana |
(Bollade et al. 1970) (Turunen 1975) (Hoffman and Downer 1979) |
| TAG to FFAs+glycerol |
Locusta migratoria Periplaneta americana; Locusta migratoria Manduca sexta Panstrongylus megistus |
(Weintraub and Tietz 1973) (Male and Storey 1981) (Tsuchida and Wells 1988) (Canavoso et al. 2004) |
| Phospholipase A1 |
Drosophila melanogaster Lepidoptera species Musca domestica |
Table 3 (Canavoso et al. 2001) (Barroso et al. 2021) |
| Phospholipase A2 |
Cicindella circumpicta Manduca sexta Aedes aegypti Spodoptera exigua Musca domestica |
(Uscian et al. 1995) (Stanley et al. 1998) (Nor Aliza and Stanley 1998) (Sajjadian et al. 2019) (Barroso et al. 2021) |
| Phospholipase B | Drosophila melanogaster | Table 3 |
| Phospholipase C |
Stomoxys calcitrans Aeshna cyanea Musca domestica |
(Spates et al. 1990) (Weiher and Komnick 1997) (Barroso et al. 2021) |
| Phospholipase D | Pieris brassicae | (Turunen 1988a) |
| Galactolipase | Epiphyas postvittana; Helicoverpa armigera | (Christeller et al. 2011) |
| Sterol dealkilation | Bombyx mori | (Ciufo et al. 2011) |
| Sterol esterase | Drosophila melanogaster | (Sieber and Thummel 2012) |
| Wax hydrolysis |
Aeshna cyanea Galleria mellonella |
(Komnick and Bauerfeind 1991) (Kong et al. 2019) |
| Lipid synthesis |
Locusta migratoria Pieris brassicae |
(Weintraub and Tietz 1978) (Turunen 1988b) |
Table 3.
Enzymatic activities of Drosophila melanogaster lipases potentially involved in lipid digestion in the midgut lumen. The list of genes and their activities (CG number) have been established mining Flybase (Flybase 2003) for lipases and phospholipases. The final selection retained the genes that were significantly expressed in the midgut based on FlyAtlas (Chintapalli et al. 2007) and contain a signal peptide based on signalP-4.1 (Petersen et al. 2011).
Table 3.
Enzymatic activities of Drosophila melanogaster lipases potentially involved in lipid digestion in the midgut lumen. The list of genes and their activities (CG number) have been established mining Flybase (Flybase 2003) for lipases and phospholipases. The final selection retained the genes that were significantly expressed in the midgut based on FlyAtlas (Chintapalli et al. 2007) and contain a signal peptide based on signalP-4.1 (Petersen et al. 2011).
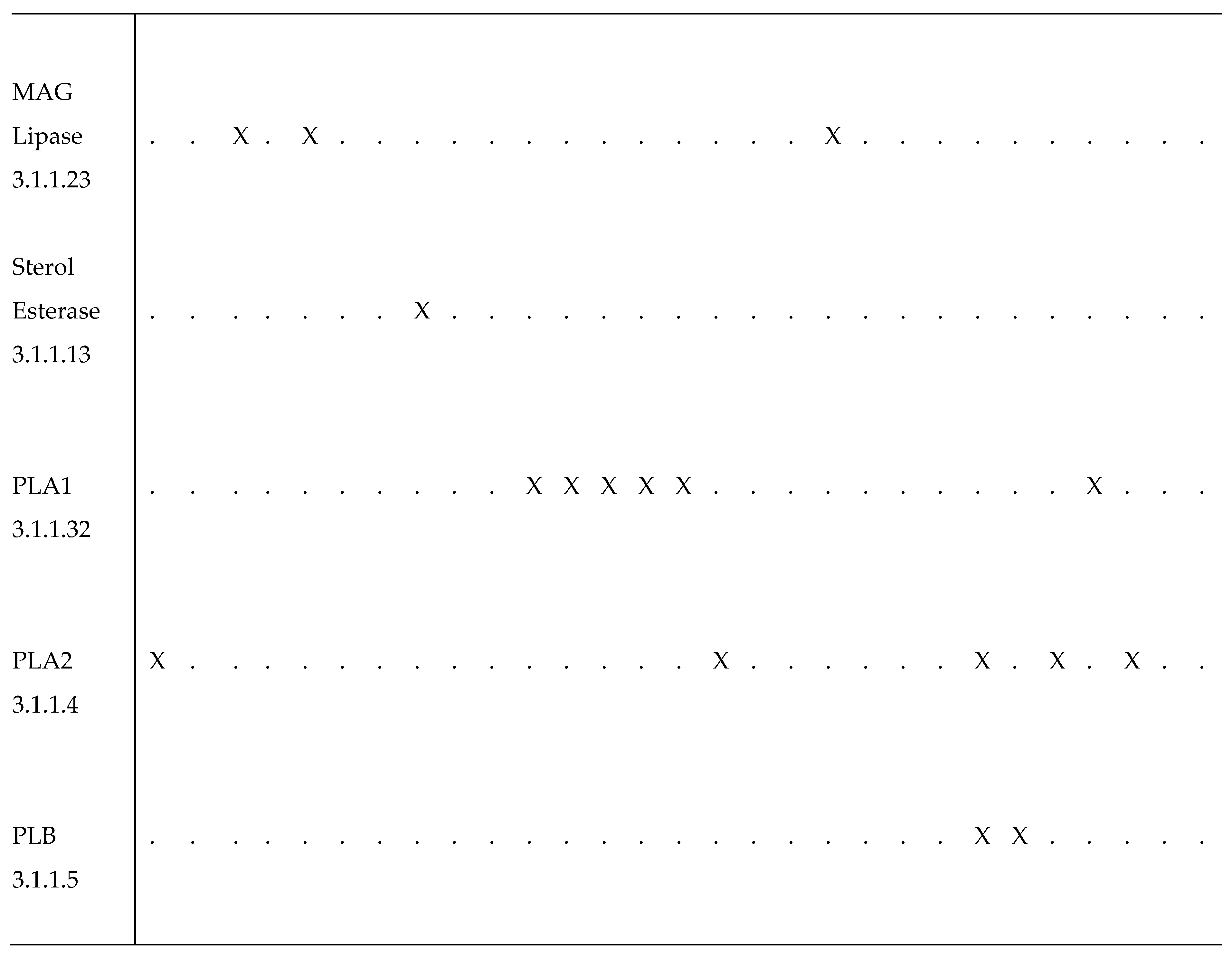
Table 4.
Number of receptors or transporters involved in lipid uptake or transfer in various insect species. These numbers were established using Flybase (Flybase 2003) and insectBase (Mei et al. 2022) and could potentially increase based on future studies.
Table 4.
Number of receptors or transporters involved in lipid uptake or transfer in various insect species. These numbers were established using Flybase (Flybase 2003) and insectBase (Mei et al. 2022) and could potentially increase based on future studies.
| Scavenger receptor | FABP | FATP | Npc1a | Npc1b | Npc2 | Flo | |
| Aedes aegypti | 3 | 1 | 2 | 0 | 1 | 4 | 2 |
| Anopheles gambiae | 4 | 2 | 2 | 1 | 1 | 4 | 3 |
| Apis mellifera | 1 | 2 | 3 | 1 | 1 | 2 | 2 |
| Bombyx mori | 1 | 5 | 5 | 1 | 1 | 1 | 3 |
| Cimex lectularius | 3 | 2 | 4 | 1 | 1 | 3 | 2 |
| Drosophila melanogaster | 3 | 1 | 3 | 1 | 1 | 8 | 2 |
| Locusta migratoria | 3 | 7 | 4 | 1 | 1 | 1 | 2 |
| Manduca sexta | 1 | 18 | 5 | 1 | 1 | 2 | 2 |
| Musca domestica | 4 | 2 | 2 | 1 | 1 | 9 | 3 |
| Rhodnius prolixus | 2 | 1 | 7 | 1 | 1 | 1 | 2 |
| Spodoptera frugiperda | 2 | 6 | 4 | 2 | 1 | 1 | 2 |
| Tenebrio molitor | 3 | 3 | 2 | 2 | 1 | 4 | 3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



