
Submitted:
25 June 2024
Posted:
26 June 2024
You are already at the latest version
Abstract
Dark pigmentation can be observed in various parts of the plant, ranging from foliage to petals to berries. The molecular basis of black pigmentation appears to vary among species, with anthocyanins playing a significant role, although specific anthocyanin types and their mechanisms differ. These findings suggest that the development of phenotypes is species-specific or varies between larger taxonomic groups; this is further supported by the polyphyletic nature of dark pigmentation. Additionally, several different regulatory mechanisms have been described for the occurrence of dark pigmentation. Firstly, the repression or knockout of the competing flavone biosynthesis has been shown to lead to darker pigmentation while another mechanism is based on the activation and upregulation of the anthocyanin biosynthesis genes in the presence of MYB transcription factors. Potential ecological functions of dark pigmentation were identified as the protection of the photosynthesis apparatus, camouflage against herbivores, and the attraction of pollinators. Promising industrial applications include microbial factories for the production of natural food colourants, induction of novel phenotypes for the ornamental plant industry and lastly the increase of anthocyanins within agriculturally relevant crops.
Keywords:
pigmentation
; secondary plant metabolites
; specialized plant metabolites
; flower colour
; stress response
; flavonoids
; anthocyanins
Introduction
The process by which plants produce colour is a fascinating realm of scientific inquiry that has captivated botanists, molecular biologists, and ecologists alike. The diverse range of colourful pigments found in plants serve vital functions, from photosynthesis facilitation to ecological interactions and stress response. Flavonoids are one of the most studied specialised metabolites within plants and responsible for the introduction of colour into various parts of the plant such as petals, berries, or foliage (Winkel-Shirley, 2001). Anthocyanins form a diverse group of compounds derived from the flavonoid biosynthesis. They are able to cause a colour spectrum from red to blue, based on slight structural variations. The visual phenotype has led to the emergence of anthocyanin biosynthesis as one of the best studied specialised metabolite pathways in plants.
Structure and Biosynthesis of Anthocyanins
Anthocyanins have a basic structure consisting of C6-C3-C6 as the basic C skeleton structure. Six major anthocyanin groups have been reported in plants: delphinidin, pelargonidin, cyanidin, malvidin, petunidin and peonidin (Houghton et al., 2021). The distinction is based on the number of hydroxyl groups, and their specific modifications through sugar or acetyl groups resulting in their respective colour properties (Winkel-Shirley, 2001). The biosynthetic pathway responsible for the formation of various anthocyanins is a well-studied mechanism that initiates with the amino acid phenylalanine. Anthocyanins are synthesised through a series of enzymatic catalytic reactions (Figure 1). In the initial six steps starting from phenylalanine, the enzymatic reactions involved in the construction of anthocyanins are consistent across the different classes. These reactions lead to the formation of dihydrokaempferol, which serves as a precursor for the subsequent steps. The pathway splits up into three parallel branches to form the three anthocyanins cyanidin, pelargonidin and delphinidin, which in turn are differentiated into malvidin, petunidin and peonidin based on specialised reactions (Schwinn et al., 2014). Each subclass of the anthocyanins is able to produce a specific colour range, which is further influenced based on the environmental conditions, especially the pH-value, and additional modifications (Van Buren et al., 1974).
The general structure and decoration groups of the specialised anthocyanins are shown in Table 1. While the main structure of all 6 anthocyanins remains the same, they display varying residues at the positions R3′ and R5′. These subtle differences are responsible for the ability of each anthocyanin to provide a certain colour.
Dark Pigmentation in Plants - Where Does It Occur?
Most plants display a variety of colours and hues in their flowers ranging from yellow, orange, red and purple to blue, but there are few plants showing a much darker or even black colouration within the petal (Markham et al., 2004). While the occurrence of true black in nature is debated, it was suggest that the definition of black colouration of plants is a function of the CIE (Commission internationale de l’éclairage) L*a*b* coordinate, namely lightness (L*) and chroma [c*: calculated as c* = (a*2 + b*2)1/2] (Deguchi et al., 2013; Van Buren et al., 1974) (Figure 2). L* indicates the lightness level of the colour, therefore the lower L* is the darker the colour, while a high L* indicated white. c* is a function of a* and b*, where a* and b* indicate magenta and yellow, respectively, if a* and b* have a positive value and green and blue if the value is negative. It has been shown, that the chroma values are not sufficient to differentiate between purple and black plant cultivars, however the L* is lower for black plants. This led Deguchi to propose a hypothetical threshold based on L* to determine an objective black definition for plants. Previously, Weatherall and Lee determined the term “black” based on a lightness value below 25 in berries (Weatherall & Lee, 1991).
Dark colouration can be found in several parts of the plant (Figure 3). For example, the occurrence of black pigmentation in seeds is very widespread within the plant kingdom, while dark pigmentation of flowers, fruits, and leaves is rare. The functional implications of this divide are not fully understood and might be associated with varying functions based on the molecular mechanism behind the dark pigmentation. For plant seeds there are known mechanisms based on so called allomelanins, which is a term describing a group of dark pigment derived from a large number of different precursors (Glagoleva et al., 2020). They often co-occur with other dark pigments such as oxidised proanthocyanidins, located in the seed coat (Devic et al., 1999).
While fully dark pigmentation of flowers, fruits, or leaves is rare it is not unheard of in nature. Especially within berries there are multiple examples for the occurrence of exceptionally dark fruits. Prominent examples for black fruits are the blackberry (Rubus armeniacus), black nightshade (Solanum nigrum) (Figure 3A), mulberry (Morus nigra) and black elderberry (Sambucus nigra). Additionally, there are several plant species which show partially dark pigmented flowers in the form of seemingly black petal spots and structures. Several potential functions have been proposed for this phenomenon, including an increase in attraction and guidance of pollinating insects, leading to a higher rate of pollination (Davies et al., 2012; Sasaki & Takahashi, 2002). A well known example of petal spots is the poppy (Papaver rhoeas) characterised by a bright red petal with black markings at the bottom of the petal (Figure 3B) (van der Kooi & Stavenga, 2019). Another example for black petal spot formation is Gorteria diffusa which displays a bright orange petal colour with a complex dark petal spot towards the bottom of the petal. The spots increase the flower’s attractiveness to potential pollinators (de Jager et al., 2017; Johnson & Midgley, 1997). Lastly, Tulipia julia displays prominent black petal spots at the base of the petals. There are also a few naturally occurring species which display a fully dark flower. One example for a dark to black flower colour would be Tacca chitari or commonly known as the bat orchid. This plant belongs to the orchid family and is naturally occurring within Southeast Asia. Another example is Lisianthius nigrescens, this plant is native to Mexico and Guatemala and is regarded as the plant with the darkest petal colour world wide (Markham et al., 2004). In this context it is noteworthy that there are non-naturally occurring garden cultivars available which were specifically bred to achieve a black flower colour (Hsu et al., 2019; Okitsu et al., 2018). There are many plant species which are available as a dark cultivar, such as tulips, petunias, dahlias and roses to name only a few. Furthermore, there are also plant species which naturally display a very dark foliage or leaves (Figure 3C). The dark pigmentation of leaves can be found both in trees as well as grasses.
To understand the phylogenetic distribution of dark pigmentation across plantae, an extensive literature screening was performed. The results were summarised and mapped on a phylogenetic family tree (Figure 4). Families that include at least one species displaying dark pigmentation in any of the selected plant tissue are highlighted. Species which display dark pigmentation can be found in several families. There is no clear clustering of the trait indicating that it has evolved independently within multiple clades thus the trait of dark pigmentation appears to be polyphyletic.
What Is Causing Dark Pigmentation?
The molecular basis of black pigmentation was studied for many different species and could be caused by different metabolites in these species. However, many reports revealed a contribution of anthocyanins to the dark pigmentation (Table 2). For example, Ophiopogon planiscapus is a plant which displays black foliage and it has been shown that the chlorophyll content and the anthocyanin content of black leaves grown under sunlight exposure was substantially higher than that of green leaves, while the carotenoid content was lower in black leaves (Hatier et al., 2013). The authors present anthocyanins as the most likely explanation for the black leaf colour. Similarly for many darkly pigmented plants the analyses revealed an exceptionally high content of anthocyanins within darkly pigmented plants ranging up to 24% of petal dry weight for L. nigrescense (Markham et al., 2004), leading to the assumption that black colour is induced by the sheer density of anthocyanin accumulation in the petals. However, a study in Dahlia variabilis has revealed that a sole upregulation of anthocyanin content is not sufficient to induce black colour in petals (Thill et al., 2012). It was identified that for Dahlia variabilis, specific cyanidin derivatives must coincide with a transcriptional block of the flavone biosynthesis to cause black flower colour (Deguchi et al., 2013). The presence of anthocyanins alone or a lack of flavone biosynthesis alone were not sufficient to resemble the black flower phenotype. Deguchi et al., 2013 speculate that substrate competition between flavone and anthocyanin biosynthesis has to be switched off to enable the formation of the black phenotype in dahlia. Similarly, studies focusing on the increased production of anthocyanins for the establishment of microbial cell factories significantly increased the amount of anthocyanins within the cell by the introduction of an AmRosea1/AmDelila1 transcription factor complex (Appelhagen et al., 2018). Another study revealed that Cy3M5G is more important for black colour than Pg3M5G (Deguchi et al., 2016). However, there does not appear to be a consensus as to which anthocyanins are responsible for the black pigmentation, as the main anthocyanin determined for black cultivars varies depending on the species (Table 2). For example, a study of the “Sun Black” Tomato line identified petanin and negretein as the most abundant anthocyanins with petanin making up 56.6 % and negretein 21.4% of total anthocyanin content in the black peel of the tomato variety, while no significant amount of cyanidin based anthocyanins were detected (Blando et al., 2019), directly contradicting the hypothesis that cyanidin derivatives have the strongest impact on black colouration. In summary, these findings indicate that the molecular mechanisms underlying the development of a dark or black phenotype might be species-specific or at least differ between larger taxonomic groups. This aligns with our phylogenetic analysis of dark to black pigmentation that revealed a polyphyletic nature of this trait (Figure 4).
How Is the Formation of Dark Pigmentation Regulated?
The regulation of the anthocyanin metabolism is a complex web involving many different transcription factors and regulators with the MYB-bHLH-WD40 complex being the best studied component (Albert et al., 2023; Gonzalez et al., 2008, 2016) Understanding the underlying mechanisms can unlock the ability to establish new traits in various plant parts. The following chapter will highlight different regulations associated with the introduction, change or enhancement of the anthocyanin accumulation.
The Role of Transcription Factors in Dark Pigmentation
The anthocyanin biosynthesis is regulated by a transcription factor complex composed of three key regulatory proteins: MYB, bHLH, and WD40 (Gonzalez et al., 2008; Ramsay & Glover, 2005). This complex is referred to as the MBW complex. MYB proteins form the largest family of transcription regulators within plants and are characterised as a highly conserved structure including two or three imperfect repeat motives of the MYB DNA-binding motif at the N-terminus (Dubos et al., 2010). Depending on the number of repeats they are classified, e.g., as R2R3-MYBs or R3-MYBs. Each repeat is made up of about 52 amino acids which form three alpha helices each (Rosinski & Atchley, 1998). These helices and repeats have regularly spaced tryptophan residues. The third helix is regarded as the “recognition” helix and allows for an interaction and binding to specific DNA structure (L. Jia et al., 2004). Additionally, MYBs can contain an activation or repression domain at the C-terminal end conveying their regulatory function (Dubos et al., 2010).
The bHLH transcription factor family is characterised by the structural feature of a basic helix-loop-helix (bHLH) domain and represents the second largest transcription factor family within plants (Murre et al., 1994). A stretch of basic amino acids located at the N-terminal end of the bHLH domain is responsible for the DNA binding, while the helix-loop-helix facilitates protein-protein interaction based on hydrophobic residues (Murre et al., 1994). This also enables bHLH transcription factors to be involved in the formation of transcription factor complexes such as the MBW complex (Lloyd et al., 2017).
The WDR family is based on the presence of a WD repeat motif, which is defined as an approximately 40 amino acid structural repeat ending with W-D (Smith et al., 1999). This loose definition makes it much harder to identify WDR proteins as they can vary greatly in not only the position but also the length of the WDR element. The main function of the WDR proteins is the facilitation of protein-protein interactions to form complexes (Lloyd et al., 2017). In the MBW complex, the WDR protein is considered to serve as scaffolding protein for the interaction of a bHLH and a MYB. It has been shown in several species that the MBW is responsible for the activation of the structural genes involved in the proanthocyanidin and anthocyanin biosynthesis (Baudry et al., 2004; Gonzalez et al., 2008). The specificity of the activation is based on the identity of the MYB and bHLH involved in the complex formation.
Several studies aimed to identify the role and identity of transcription factors involved in the occurrence of dark anthocyanin pigmentation. One striking example is the complex formation of petal spots in different cultivars of the Beetle Daisy (Gorteria diffusa). By comparing the different cultivars and the metabolites present in the dark petal spots versus the brightly coloured petal it was revealed that malonylated cyanidins were almost exclusively present in the dark spots, as the most abundant anthocyanin (Fattorini et al., 2024). Further, three subgroup 6 R2R3-MYBs namely GdMYBSG6-1, GdMYBSG6-2, and GdMYBSG6-3 were identified in connection to the spot formation (Fattorini et al., 2024). The expression of these transcription factors was significantly increased in spotted tissue and it is assumed that they could act redundantly to promote anthocyanin production with GdMYBSG6-2 being the most important (Fattorini et al., 2024). Further, it was shown that the expression of GdMYBSG6-1,2,3 lead to an upregulation of GdANS, GdDFR, and GdMAT1 in spotted tissue compared to unspotted petals (Fattorini et al., 2024).
Similarly, in dark Phalaenopsis orchids the PeMYB11 was identified as the most likely cause of dark pigmentation (Hsu et al., 2019). Here, it was discovered, that the dark phenotype was based on the insertion of a retrotransposon, named Harlequin Orchid RetroTransposon 1 (HORT1), which further increased the expression of PeMYB11 leading to an enhanced activation of the anthocyanin biosynthesis genes (Hsu et al., 2019).
Solanum Lycopersicum a Model to Understand the Regulation of Anthocyanin Accumulation in Dark Phenotypes
While the typical tomato plant (Solanum lycopersicum) does not produce anthocyanins in their fruit, wild relatives such as the wild Solanum species S. chilense and S. lycopersicoides are capable of producing and accumulating anthocyanins in the fruit peel. Solanum chilense develops a distinct phenotype of dark spotted fruits on green fruit peel in response to low temperatures or high light exposure (Mes et al., 2008). The genetic region responsible for the induction of anthocyanin synthesis is the Anthocyanin Fruit (Aft) locus, located on chromosome 10 and encoding a R2R3-MYB (Colanero et al., 2020). The MYB has been identified as SlAN2-like and has been shown to activate the expression of DFR based on the formation of a MBW complex consisting of SlAN2-like, WDR, bHLH1, and bHLH2 (Sun et al., 2020). Comparison of the SlAN2-like sequence with that of a red-fruited tomato revealed a small sequence variation leading to alternative splice sites (Sapir et al., 2008). While the spliced version from S. chilense leads to a functional R2R3-MYB, the alternatively spliced SlAN2-like in S. lycopersicum cannot activate the anthocyanin biosynthesis pathway (Colanero et al., 2020). If SIAN2-like from S. chilense is introduced into S. lycopersicum an accumulation of anthocyanins in the fruit peel can be observed further indicating SlAN2-like is a key factor on the introduction of anthocyanin biosynthesis in Solanum plants (Sapir et al., 2008). A similar mechanism was discovered for Solanum lycopersicoides, with the responsible locus called ‘Aubergine’ or Agb (Mes et al., 2008; Rick et al., 1994). Homozygous plants for agb develop infertile, deformed, very dark fruits, while heterozygotes produce fruits with the expected shape and size but still presenting a dark phenotype. Menconi et al. identified the location of Abg on Chromosome 10 and detected the same splice site variation as in aft, along with another splicing variant leading to reduced anthocyanin accumulation (Menconi et al., 2023). Further, the absence or reduced activity of an anthocyanin repressor, SlMYB-ATV, may play a role in the accumulation of anthocyanins in Solanum species. For the genetically engineered purple tomato species indigo rose, a 4 bp deletion in the repressor led to a significant increase in anthocyanin accumulation, while other dark tomato varieties with an intact repressor showed a lower anthocyanin content (Cao et al., 2017). These studies of different close relatives of tomato demonstrated that the capability to produce anthocyanins was lost due to the introduction of an alternative splicing site in the R2R3-MYB gene SlAN2-like, an activator of late anthocyanin biosynthesis.
Genetic Modification Resulting in a Dark Phenotype
Besides naturally occurring dark phenotypes there have been attempts to introduce the phenotype via genetic engineering (Zhang et al., 2014, 2016). The main interest in the studies were the health-promoting properties of anthocyanins, with the introduction of a dark phenotype being a secondary effect. There are two distinct approaches and targets for the enrichment of anthocyanins. The first target is to produce dark phenotypes in commonly consumed fruits and vegetables such as the tomato. As previously described, there have been long standing breeding programs with naturally occurring anthocyanin producing wild species to generate a dark tomato cultivar (Mazzucato et al., 2013). The success through breeding avoids the production of a genetically modified plant making it particularly interesting for the European market. Simultaneously, the use of genetic engineering has also been applied to induce the production of anthocyanins within the tomato clade (Butelli et al., 2008, 2021).
The introduction of the anthocyanin biosynthesis is based on the introduction of new regulatory genes encoding transcription factors activating the relevant anthocyanin biosynthesis genes. By introducing the bHLH protein Delila (AmDel) and the MYB protein Rosea1 (AmRos1) to a wildtype tomato species the anthocyanin biosynthesis is reactivated and the dark phenotype is introduced (Butelli et al., 2008). The transcripts are under the control of the fruit-specific E8-promoter leading to the accumulation within the entire fruit, both pulp and peel, leading to an increased amount of anthocyanins in comparison to the bred purple tomato varieties previously described, which only accumulate anthocyanins within the peel (Butelli et al., 2008; Mazzucato et al., 2013). Del and Ros1 activate several genes required for anthocyanin biosynthesis including PAL, CHS and DFR explaining the high accumulation of anthocyanins (Butelli et al., 2008; Naing et al., 2017). This leads to a dark purple phenotype. An additional expression of MYB12 from Arabidopsis thaliana resulted in an increased accumulation of anthocyanins leading to a more intensely coloured tomato fruit, due to enhanced activation of the PAL and CHS, this variant is referred to as ‘indigo’ (Butelli et al., 2021). A similar approach has been used to establish microbial cell factories, in order to build a stable high-yield production of specifically decorated anthocyanins for the application in the food industry as colourants or food additives. Applehagen and colleagues presented a strategy to induce anthocyanin in Nicotiana tabacum cv. Samsun cells, by introducing the bHLH Delila (AmDel) and the MYB Rosea1 (AmRos1) from Antirrhinum majus (Appelhagen et al., 2018). In N. tabacum the accumulation of anthocyanins is restricted to the flower petals, while producing almost exclusively cyanidin 3-O-rutenoside (Appelhagen et al., 2018). When the AmRos1 and AmDel are constitutively expressed in N. tabacum plants, the anthocyanin accumulation expands to the rest of the plant with C3R remaining the main anthocyanin produced (Appelhagen et al., 2018). A cell culture was established and it showed a stable production throughout the cultivation reaching anthocyanin levels higher then the amount of C3R found within anthocyanin rich fruits such as blackberries or blueberries (Appelhagen et al., 2018). Furthermore, by introducing additional genes like the F3’5’H from Petunia or 3-O-rutinoside-4’’’-hydroxycinnamoyl transferase from Solanum lycopersicum (Sl3AT) the range of anthocyanins can be expanded allowing to the production of several anthocyanins at the same time (Appelhagen et al., 2018).
Stability of Anthocyanin Pigmentation
The stability of anthocyanins can be influenced by a variety of parameters such as the pH-value, temperatur, light conditions, co-pigmentation with flavones and iron metals as well as additional modifications such as glycosyl and aromatic or aliphatic acyl moieties (Rowan et al., 2009; Zhao et al., 2021). For example, light conditions are an important factor to consider when storing anthocyanins, as it has a significant negative effect on their stability (Amr & Al-Tamimi, 2007; Kearsley & Rodriguez, 1981). By storing anthocyanin extracts in the dark the half life of the molecules is significantly increased. A study showed that extracts from black currant fruits lose half of their antioxidant potential and associated anthocyanins within 8.25 days when stored in natural light conditions (N. Jia et al., 2013). Under artificial light the half life is increased to 18.81 days while a storage in complete darkness further increases the half life to 21 days (N. Jia et al., 2013). Similar results were reported for extracts from both mulberry as well as acai berry (Aramwit et al., 2010; de Rosso & Mercadante, 2007).
Moreover, high temperature has also been reported to significantly decrease the stability of anthocyanins, with the anthocyanin content of grape extracts being reduced to less than half (Mori et al., 2007). For blueberries it has been reported that the half life in pH 3.0 of anthocyanins slowly decreased with a temperature increase from 25°C to 50°C (Liu et al., 2018). However, further increasing the temperature to 60°C and above leads to a rapid decrease in half life, resulting in ¼ of the half life at 60°C compared to 25°C (Liu et al., 2018).
Interestingly, not all anthocyanins appear to be degrading at the same rate. In a study focusing on the stability of anthocyanins in black carrot, it was found, that acylated anthocyanins from black carrot remain more stable to temperature increase of 20-50°C compared to non-acetylated anthocyanins extracted from blackberry (Zozio et al., 2011). Similarly, diacylated anthocyanins provide significantly high blue colour stability to red cabbage at 50°C as compared to non-acylated anthocyanins (Fenger et al., 2021). However, the increased stabilisation of anthocyanins through acylation is dependent on the type of acyl group, their position or attachment side as well as the number of acylations (Fenger et al., 2021). In general, it is differentiated between aromatic and aliphatic acyl groups. The stabilisation based on acylation by aromatic acid is based on enabling intra- and intermolecular copigmentation or self-association of anthocyanins. Acylation with aliphatic acid, however, stabilises the anthocyanins based on the introduction of steric hindrance (Jokioja et al., 2021).
However, the effect of the stabilisation is greatly dependent on the identity of the residue added. Luo et al., 2007 showed that for anthocyanins extracted from a transgenic tobacco line, the highest stability of anthocyanin was conveyed by the addition of (coumaroyl) glucoside with a half life of 48h, followed by cyanidin 3-rutinoside (t ½= 24 h), cyanidin 3-glucoside (t ½ = 10 h) and lastly cyanidin 3-glucoside 5-malonylglucoside (t ½ = 5.5 h) (Luo et al., 2007).
What Are the Proposed Ecological Functions and Potential Industrial Applications of Dark Pigmentation?
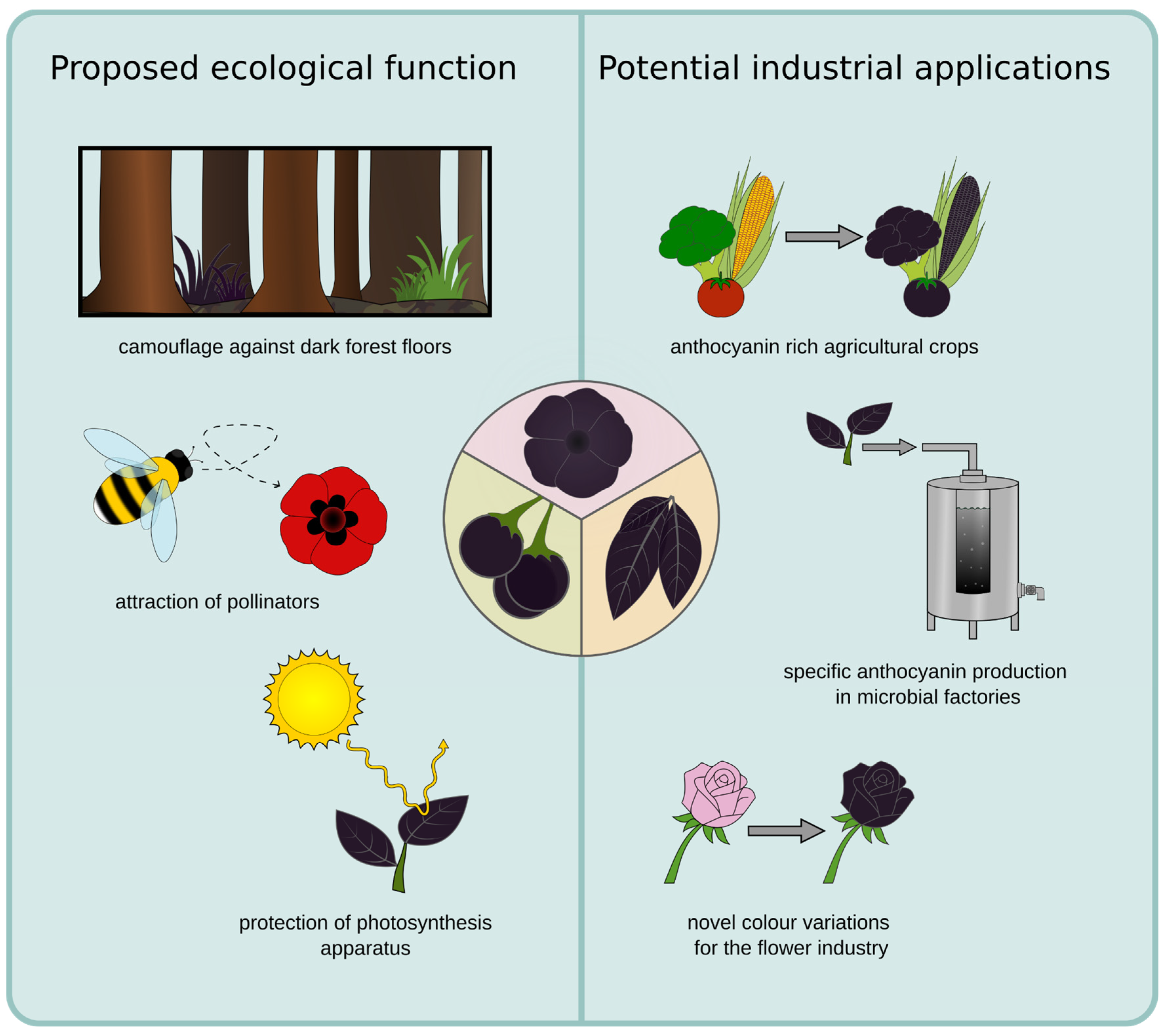
Anthocyanins have been associated with many beneficial characteristics based on their antioxidant properties and serve several essential functions which contribute to plant survival, development, and interaction with the environment (Chalker-Scott, 1999; Gould, 2004). Additionally, vibrant colours give plants an advantage in attracting different pollinators, however the repeated occurrence of dark petals, foliage and berries suggests that there are potential benefits to dark colours as well. One proposed advantage of black foliage is an indirect protection against herbivores, as the dark foliage looks reminiscent of a dead plant and is in general less visible to herbivores against exposed soil of a dark forest floor (Figure 5) (Gould, 2004). This could potentially explain the occurrence of dark leaves and foliage as seen in Ophiopogon planiscapus. Furthermore, it has been proposed that dark flowers might help with pollination due to an increased nectar temperature caused by the poor reflection of sunlight which warms up the flower (Büdel, 1959; Lacey et al., 2010; Tikhomirov et al., 1960). This could be beneficial in cold environments, as insects are more likely to visit dark flowers in these conditions (Norgate et al., 2010). A study involving the Australian native bee confirmed that an increased nectar temperature is only an advantage if the environmental temperature is low (Norgate et al., 2010). However, a test on Ophiopogon planiscapus did not reveal a significant influence of the dark pigmentation on the leaf temperature, while revealing that an intense pigmentation of the leaves reduces the carbon fixation and biomass formation under intense light conditions (Hatier et al., 2013). At the same time, the photo inhibitory stress was lowered (Gould, Neill, et al., 2002; Hatier et al., 2013). These findings suggest that dark pigmentation has a potential to reduce the photosynthesis of a plant while simultaneously providing protection of the photosynthesis apparatus. While the mechanism for the enhanced photoprotection in the black leaves of O. planiscapus is not clear (Hatier et al., 2013), it is most likely due to the high accumulation of anthocyanins, as the functions of anthocyanins are the protection against UV radiation from the sun and scavenging of reactive oxygen species (Gould, 2004). Anthocyanins act as a natural sunblock, by absorbing UV-B and UV-A light (Landi et al., 2015). This protects the plant tissue from potential negative effects of the UV rays and is particularly important for plants in areas with intense sunlight or at high altitudes (Gould, 2004). Further, they could protect through the absorbance of excess photons while additionally being antioxidants that scavenge reactive oxygen species (ROS) and prevent damage of the photosynthesis apparatus (Figure 5)(Gould, 2004).
Another natural function of anthocyanins lies within the attraction of pollinators and seed dispersers. For example several plant species utilise dark pigmentation in order to mimic their respective pollinator in order to attract them to the flower (Figure 5) (Johnson & Midgley, 1997). Examples for this mimicry or sexual deception can be observed for the beatle daisy as well as the genus Ophrys. The dark petal spot of G. diffusia mimics the bee fly Megapalpus capensis as this is their main pollinator, with male flies preferring the more complex petal spots (Ellis et al., 2014; Thomas et al., 2009). Similarly, the genus Ophrys is imitating several different species of insect pollinators and several shifts and adaptations to new pollinators can be traced within the genus (Breitkopf et al., 2015). This genus displays an exceptionally high rate of specification which has led to the emergence of several hundred species in the Mediterranean region of the western Palaearctic (Baguette et al., 2020). By imitating sexually receptive females of one particular species of insect, the Ophrys genus is capable of attracting male pollinators and increasing their pollination rate. Both the structure of the flower as well as the dark pigmentation play a key role in this process (Bradshaw et al., 2010). Additionally, in berries the anthocyanin content often increases with the ripening of the berry. For example blackberries change colour from a light green to a black appearance over the course of ripening. Although red pigmentation appears often more effective, the darkened colour attracts birds, who consume the berries and then distribute the seed (Duan et al., 2014; Enaru et al., 2021; Schaefer et al., 2006).
Anthocyanin production greatly increases in the presence of stress factors (Chalker-Scott, 1999; Liu et al., 2020), particularly in response to low temperatures (Christie et al., 1994), drought (Chalker-Scott, 1999), nutrient deficiencies (Peng et al., 2008) or pathogen attacks (Liu et al., 2020). Their connection to stress response is assumed to be due to their antioxidant properties (Gould, McKelvie, et al., 2002). Exactly these antioxidant properties are what makes anthocyanins interesting to the field of medicine and anthocyanins have been implicated as preventive or curative measures regarding several health conditions such as cardiovascular disease, diabetes, and obesity (Belwal et al., 2017; Dong et al., 2022; Lee et al., 2017).
While anthocyanins are often present in commonly consumed fruits and vegetables, their concentration and therefore uptake may not be sufficient to convey these health benefits. In order to overcome this challenge several attempts have been undertaken to increase the concentration within popular agricultural crops (Butelli et al., 2008). While there are approaches to selectively breed for the desired increase of anthocyanin concentration, this is a very time consuming process (Mazzucato et al., 2013). Alternatively, for the tomato plant the increase of anthocyanins and induction of a dark phenotype was also achieved by the introduction of novel transcription factors in the form of genetic engineering, this allows for a more targeted introduction of desired anthocyanins (Butelli et al., 2021). Another interesting industrial application for revealing the mechanism of dark pigmentation is that it would allow for the targeted introduction of a dark phenotype in ornamental plants or flowers for the cut flower industry. Previously, there have been studies introducing novel colour variants of Chrysanthemum morifolium Ramat. By expanding the anthocyanin biosynthesis pathway to include delphinidin derived anthocyanins a blue and purple phenotype was introduced to the species (N. Noda et al., 2013). A similar approach can be followed to expand colour variations within desirable ornamental plants to include a dark phenotype.
The natural colour properties of anthocyanins make them highly desirable as alternatives to synthetic dyes in various industries, including food, cosmetics, textiles, and nutraceuticals (Alappat & Alappat, 2020). In the food industry, anthocyanins are used as natural colourants to enhance the visual appeal of a wide range of products, such as beverages, dairy products, confectioneries, and baked goods (Appelhagen et al., 2018; Ghosh et al., 2022). However, despite their promising scientific and industrial potential, there are several challenges associated with the industrial use of anthocyanins. For example, the currently commercially available colour spectrum of anthocyanins is still limited (Appelhagen et al., 2018). At the moment, anthocyanins are extracted from waste products from the wine industry such as grapes or other vegetables such as red cabbage and purple sweet potato (Francis & Markakis, 1989; Ghareaghajlou et al., 2021; A. Li et al., 2019). While this presents an economically useful process, it also means that no batch of extracted anthocyanins will be exactly the same. Additionally, it is not possible to easily extract just one specific anthocyanin with the desired colour properties but rather a mix of all anthocyanins present in the fruit in varying quantities. This leads to a slight colour variation between batches of extracted anthocyanins (Díaz-García et al., 2015).
Anthocyanins provide an attractive alternative to synthetic dyes, as they are perceived as natural and healthier options by consumers who prefer clean-label products. In regards to dark food colouration, a clean alternative to synthetic colourants is currently lacking. To the best of our knowledge, the only available natural black food colourant on the market is called vegetable carbon or E153. This food colourant is created by charring wood and vegetable parts to turn these into activated charcoal, which is added to foods to convey black colouration (EFSA Panel on Food Additives and Nutrient Sources added to Food (ANS), 2012). The use of vegetable carbon, as any food additives, is under strict regulation by several authorities such as the FDA in the United States or the EFSA in Europe. However, there are consumer concerns about the potential presence of carcinogenic compounds within E153 (Hilber et al., 2022). Answering the scientific question regarding the underlying mechanism of dark pigmentation in plants could enable the production of a natural colourant allowing to substitute E153. Additionally to the food industry anthocyanins are similarly used in the cosmetics and textile industry to produce natural colourants which meet the current consumer demand to strive for sustainable and non-toxic alternatives.
Figure 5.
Schematic overview of proposed ecological functions and potential industrial applications of dark pigmentation.
Figure 5.
Schematic overview of proposed ecological functions and potential industrial applications of dark pigmentation.
Conclusion and Future Perspectives
Dark pigmentation in plants is a rare and fascinating phenomenon with many unanswered research questions. Currently, a strong expression and accumulation of anthocyanins are the most likely explanation for its occurrence. However, the reason behind the accumulation, the underlying mechanism and the identity of the necessary anthocyanin are not yet fully discovered.
The main anthocyanin for dark phenotypes varies from species to species indicating that there are different mechanisms leading to dark colour. This idea is also supported by the independent evolution of dark phenotypes across different plant families.
Additionally, several mechanisms of the regulation associated with the overexpression of anthocyanins have been identified. One mechanism is based on the repression or knockout of the competing flavone biosynthesis leading to darker pigmentation while another mechanism is based on the activation and upregulation of the anthocyanin biosynthesis genes in the presence of MYB transcription factors.
However, there are still several key questions which should be addressed to fully understand the occurrence of dark pigmentation in plants. Firstly, the composition of the anthocyanins present within darkly pigmented plant parts should be analysed and compared between closely related species. Is there a consensus on the base anthocyanin or the modifications present? Further, it would be necessary to shed light on both the genes involved in the production of the anthocyanins associated with dark pigmentation and also the regulatory elements involved.
Understanding the underlying cause of dark pigmentation in plants can have an impact on several industrial applications.
By revealing the regulatory elements and genes involved in this distinct phenotype it opens up the possibility of either selectively breeding or screening for the desired traits in flowers or even inducing the traits with the help of genetic engineering or genome editing. Being able to develop dark flowers for popular ornamental plants would be an interesting application for the cut flower industry. Similarly this approach could be expanded to agriculturally interesting plants, as shown in tomato, to increase the anthocyanin content and therefore the antioxidant properties of the targeted crop. This could lead to health benefits for the consumers or be utilised to increase the yield of anthocyanin extractions of foods for an application in natural food colourants. Additionally, it would unlock heterologous production of specific anthocyanins within yeast or even the establishment of a plant cell culture. Heterologous expression is a promising approach for targeted production of specific anthocyanins associated with dark colouration with the potential of establishing upscaling to industrial standards and parallelisation, leading to increased yield of natural colourants to be used in the food industry.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Abbreviations
| PAL | phenylalanine ammonia-lyase |
| C4H | cinnamic acid 4-hydroxylase |
| 4CL | 4-coumarate-CoA ligase |
| CHS | chalcone synthase |
| CHI | chalcone isomerase |
| F3H | flavanone 3-hydroxylase |
| F3’H | flavonoid 3’-hydroxylase |
| F3’5’H | flavonoid 3’,5’-hydroxylase |
| DFR | dihydroflavonol 4-reductase |
| LDOX | leucoanthocyanidin dioxygenase |
| arGST | anthocyanin-related Glutathione S-transferase |
| CIE | Commission internationale de l’éclairage |
| MYB | Myeloblastosis |
| bHLH | basic helix-loop-helix |
References
- Alappat, B., & Alappat, J. (2020). Anthocyanin Pigments: Beyond Aesthetics. Molecules, 25(23), Article 23. [CrossRef]
- Albert, N. W., Iorizzo, M., Mengist, M. F., Montanari, S., Zalapa, J., Maule, A., Edger, P. P., Yocca, A. E., Platts, A. E., Pucker, B., & Espley, R. V. (2023). Vaccinium as a comparative system for understanding of complex flavonoid accumulation profiles and regulation in fruit. Plant Physiology, 192(3), 1696–1710. [CrossRef]
- Amr, A., & Al-Tamimi, E. (2007). Stability of the crude extracts of Ranunculus asiaticus anthocyanins and their use as food colourants. International Journal of Food Science & Technology, 42(8), 985–991. [CrossRef]
- Appelhagen, I., Wulff-Vester, A. K., Wendell, M., Hvoslef-Eide, A.-K., Russell, J., Oertel, A., Martens, S., Mock, H.-P., Martin, C., & Matros, A. (2018). Colour bio-factories: Towards scale-up production of anthocyanins in plant cell cultures. Metabolic Engineering, 48, 218–232. [CrossRef]
- Aramwit, P., Bang, N., & Srichana, T. (2010). The properties and stability of anthocyanins in mulberry fruits. Food Research International, 43(4), 1093–1097. [CrossRef]
- Awika, J. M., Rooney, L. W., & Waniska, R. D. (2005). Anthocyanins from black sorghum and their antioxidant properties. Food Chemistry, 90(1), 293–301. [CrossRef]
- Baguette, M., Bertrand, J. A. M., Stevens, V. M., & Schatz, B. (2020). Why are there so many bee-orchid species? Adaptive radiation by intra-specific competition for mnesic pollinators. Biological Reviews, 95(6), 1630–1663. [CrossRef]
- Baudry, A., Heim, M. A., Dubreucq, B., Caboche, M., Weisshaar, B., & Lepiniec, L. (2004). TT2, TT8, and TTG1 synergistically specify the expression of BANYULS and proanthocyanidin biosynthesis in Arabidopsis thaliana. The Plant Journal: For Cell and Molecular Biology, 39(3), 366–380. [CrossRef]
- Belwal, T., Nabavi, S. F., Nabavi, S. M., & Habtemariam, S. (2017). Dietary Anthocyanins and Insulin Resistance: When Food Becomes a Medicine. Nutrients, 9(10), Article 10. [CrossRef]
- Blando, F., Berland, H., Maiorano, G., Durante, M., Mazzucato, A., Picarella, M. E., Nicoletti, I., Gerardi, C., Mita, G., & Andersen, Ø. M. (2019). Nutraceutical Characterization of Anthocyanin-Rich Fruits Produced by “Sun Black” Tomato Line. Frontiers in Nutrition, 6. [CrossRef]
- Bradshaw, E., Rudall, P. J., Devey, D. S., Thomas, M. M., Glover, B. J., & Bateman, R. M. (2010). Comparative labellum micromorphology of the sexually deceptive temperate orchid genus Ophrys: Diverse epidermal cell types and multiple origins of structural colour. Botanical Journal of the Linnean Society, 162(3), 504–540. [CrossRef]
- Breitkopf, H., Onstein, R. E., Cafasso, D., Schlüter, P. M., & Cozzolino, S. (2015). Multiple shifts to different pollinators fuelled rapid diversification in sexually deceptive Ophrys orchids. New Phytologist, 207(2), 377–389. [CrossRef]
- Büdel, A. (1959). The microclimate of flowers blooming near the ground. https://scholar.google.com/scholar_lookup?journal=Z.+Bienenforsch.&title=The+microclimate+of+flowers+blooming+near+the+ground&author=A.+B%C3%BCdel&volume=4&publication_year=1959&pages=131-140&.
- Butelli, E., Bulling, K., Hill, L., & Martin, C. (2021). Beyond Purple Tomatoes: Combined Strategies Targeting Anthocyanins to Generate Crimson, Magenta, and Indigo Fruit. Horticulturae, 7(9), Article 9. [CrossRef]
- Butelli, E., Titta, L., Giorgio, M., Mock, H.-P., Matros, A., Peterek, S., Schijlen, E. G. W. M., Hall, R. D., Bovy, A. G., Luo, J., & Martin, C. (2008). Enrichment of tomato fruit with health-promoting anthocyanins by expression of select transcription factors. Nature Biotechnology, 26(11), 1301–1308. [CrossRef]
- Cao, X., Qiu, Z., Wang, X., Van Giang, T., Liu, X., Wang, J., Wang, X., Gao, J., Guo, Y., Du, Y., Wang, G., & Huang, Z. (2017). A putative R3 MYB repressor is the candidate gene underlying atroviolacium, a locus for anthocyanin pigmentation in tomato fruit. Journal of Experimental Botany, 68(21–22), 5745–5758. [CrossRef]
- Chalker-Scott, L. (1999). Environmental Significance of Anthocyanins in Plant Stress Responses. Photochemistry and Photobiology, 70(1), 1–9. [CrossRef]
- Christie, P. J., Alfenito, M. R., & Walbot, V. (1994). Impact of low-temperature stress on general phenylpropanoid and anthocyanin pathways: Enhancement of transcript abundance and anthocyanin pigmentation in maize seedlings. Planta, 194(4), 541–549. [CrossRef]
- Colanero, S., Tagliani, A., Perata, P., & Gonzali, S. (2020). Alternative Splicing in the Anthocyanin Fruit Gene Encoding an R2R3 MYB Transcription Factor Affects Anthocyanin Biosynthesis in Tomato Fruits. Plant Communications, 1(1), 100006. [CrossRef]
- Davies, K. M., Albert, N. W., Schwinn, K. E., Davies, K. M., Albert, N. W., & Schwinn, K. E. (2012). From landing lights to mimicry: The molecular regulation of flower colouration and mechanisms for pigmentation patterning. Functional Plant Biology, 39(8), 619–638. [CrossRef]
- de Jager, M. L., Willis-Jones, E., Critchley, S., & Glover, B. J. (2017). The impact of floral spot and ring markings on pollinator foraging dynamics. Evolutionary Ecology, 31(2), 193–204. [CrossRef]
- de Rosso, V. V., & Mercadante, A. Z. (2007). Evaluation of colour and stability of anthocyanins from tropical fruits in an isotonic soft drink system. Innovative Food Science & Emerging Technologies, 8(3), 347–352. [CrossRef]
- Deguchi, A., Ohno, S., Hosokawa, M., Tatsuzawa, F., & Doi, M. (2013). Endogenous post-transcriptional gene silencing of flavone synthase resulting in high accumulation of anthocyanins in black dahlia cultivars. Planta, 237(5), 1325–1335. [CrossRef]
- Deguchi, A., Tatsuzawa, F., Hosokawa, M., Doi, M., & Ohno, S. (2016). Quantitative Evaluation of the Contribution of Four Major Anthocyanins to Black Flower Coloring of Dahlia Petals. The Horticulture Journal, advpub, MI–121. [CrossRef]
- Devic, M., Guilleminot, J., Debeaujon, I., Bechtold, N., Bensaude, E., Koornneef, M., Pelletier, G., & Delseny, M. (1999). The BANYULS gene encodes a DFR-like protein and is a marker of early seed coat development. The Plant Journal: For Cell and Molecular Biology, 19(4), 387–398. [CrossRef]
- Díaz-García, M. C., Castellar, M. R., Obón, J. M., Obón, C., Alcaraz, F., & Rivera, D. (2015). Production of an anthocyanin-rich food colourant from Thymus moroderi and its application in foods. Journal of the Science of Food and Agriculture, 95(6), 1283–1293. [CrossRef]
- Dong, Y., Wu, X., Han, L., Bian, J., He, C., El-Omar, E., Gong, L., & Wang, M. (2022). The Potential Roles of Dietary Anthocyanins in Inhibiting Vascular Endothelial Cell Senescence and Preventing Cardiovascular Diseases. Nutrients, 14(14), Article 14. [CrossRef]
- Duan, Q., Goodale, E., & Quan, R. (2014). Bird fruit preferences match the frequency of fruit colours in tropical Asia. Scientific Reports, 4(1), 5627. [CrossRef]
- Dubos, C., Stracke, R., Grotewold, E., Weisshaar, B., Martin, C., & Lepiniec, L. (2010). MYB transcription factors in Arabidopsis. Trends in Plant Science, 15(10), 573–581. [CrossRef]
- EFSA Panel on Food Additives and Nutrient Sources added to Food (ANS). (2012). Scientific Opinion on the re-evaluation of vegetable carbon (E 153) as a food additive. EFSA Journal, 10(4), 2592. [CrossRef]
- Ellis, A. G., Brockington, S. F., de Jager, M. L., Mellers, G., Walker, R. H., & Glover, B. J. (2014). Floral trait variation and integration as a function of sexual deception in Gorteria diffusa. Philosophical Transactions of the Royal Society B: Biological Sciences, 369(1649), 20130563. [CrossRef]
- Enaru, B., Drețcanu, G., Pop, T. D., Stǎnilǎ, A., & Diaconeasa, Z. (2021). Anthocyanins: Factors Affecting Their Stability and Degradation. Antioxidants, 10(12), Article 12. [CrossRef]
- Fattorini, R., Khojayori, F. N., Mellers, G., Moyroud, E., Herrero, E., Kellenberger, R. T., Walker, R., Wang, Q., Hill, L., & Glover, B. J. (2024). Complex petal spot formation in the Beetle Daisy (Gorteria diffusa) relies on spot-specific accumulation of malonylated anthocyanin regulated by paralogous GdMYBSG6 transcription factors. New Phytologist, n/a(n/a). [CrossRef]
- Fenger, J.-A., Sigurdson, G. T., Robbins, R. J., Collins, T. M., Giusti, M. M., & Dangles, O. (2021). Acylated Anthocyanins from Red Cabbage and Purple Sweet Potato Can Bind Metal Ions and Produce Stable Blue Colors. International Journal of Molecular Sciences, 22(9), Article 9. [CrossRef]
- Francis, F. J., & Markakis, P. C. (1989). Food colorants: Anthocyanins. Critical Reviews in Food Science & Nutrition. [CrossRef]
- Ghareaghajlou, N., Hallaj-Nezhadi, S., & Ghasempour, Z. (2021). Red cabbage anthocyanins: Stability, extraction, biological activities and applications in food systems. Food Chemistry, 365, 130482. [CrossRef]
- Ghosh, S., Sarkar, T., Das, A., & Chakraborty, R. (2022). Natural colorants from plant pigments and their encapsulation: An emerging window for the food industry. LWT, 153, 112527. [CrossRef]
- Glagoleva, A. Y., Shoeva, O. Y., & Khlestkina, E. K. (2020). Melanin Pigment in Plants: Current Knowledge and Future Perspectives. Frontiers in Plant Science, 11. [CrossRef]
- Gonzalez, A., Brown, M., Hatlestad, G., Akhavan, N., Smith, T., Hembd, A., Moore, J., Montes, D., Mosley, T., Resendez, J., Nguyen, H., Wilson, L., Campbell, A., Sudarshan, D., & Lloyd, A. (2016). TTG2 controls the developmental regulation of seed coat tannins in Arabidopsis by regulating vacuolar transport steps in the proanthocyanidin pathway. Developmental Biology, 419(1), 54–63. [CrossRef]
- Gonzalez, A., Zhao, M., Leavitt, J. M., & Lloyd, A. M. (2008). Regulation of the anthocyanin biosynthetic pathway by the TTG1/bHLH/Myb transcriptional complex in Arabidopsis seedlings. The Plant Journal, 53(5), 814–827. [CrossRef]
- Goto, T., Takase, S., & Kondo, T. (1978). PMR spectra of natural acylated anthocyanins determination of stereostructure of awobanin, shisonin and violanin. Tetrahedron Letters, 19(27), 2413–2416. [CrossRef]
- Gould, K. S. (2004). Nature’s Swiss Army Knife: The Diverse Protective Roles of Anthocyanins in Leaves. Journal of Biomedicine and Biotechnology, 2004(5), 314–320. [CrossRef]
- Gould, K. S., McKelvie, J., & Markham, K. R. (2002). Do anthocyanins function as antioxidants in leaves? Imaging of H2O2 in red and green leaves after mechanical injury. Plant, Cell & Environment, 25(10), 1261–1269. [CrossRef]
- Gould, K. S., Neill, S. O., & Vogelmann, T. C. (2002). A unified explanation for anthocyanins in leaves? In Advances in Botanical Research (Bd. 37, S. 167–192). Academic Press. [CrossRef]
- Hatier, J.-H. B., Clearwater, M. J., & Gould, K. S. (2013). The Functional Significance of Black-Pigmented Leaves: Photosynthesis, Photoprotection and Productivity in Ophiopogon planiscapus ‘Nigrescens’. PLOS ONE, 8(6), e67850. [CrossRef]
- Hilber, I., Blum, F., Schmidt, H.-P., & Bucheli, T. D. (2022). Current analytical methods to quantify PAHs in activated carbon and vegetable carbon (E153) are not fit for purpose. Environmental Pollution, 309, 119599. [CrossRef]
- Houghton, A., Appelhagen, I., & Martin, C. (2021). Natural Blues: Structure Meets Function in Anthocyanins. Plants, 10(4), Article 4. [CrossRef]
- Hsu, C.-C., Su, C.-J., Jeng, M.-F., Chen, W.-H., & Chen, H.-H. (2019). A HORT1 Retrotransposon Insertion in the PeMYB11 Promoter Causes Harlequin/Black Flowers in Phalaenopsis Orchids. Plant Physiology, 180(3), 1535–1548. [CrossRef]
- Jia, L., Clegg, M. T., & Jiang, T. (2004). Evolutionary dynamics of the DNA-binding domains in putative R2R3-MYB genes identified from rice subspecies indica and japonica genomes. Plant Physiology, 134(2), 575–585. [CrossRef]
- Jia, N., Kong, B. H., & Liu, Q. (2013). Influence of Light on the Anthocyanins Stability and Antioxidant Activity of Black Currant (Ribes nigrum L.) Extract. Applied Mechanics and Materials, 448–453, 1119–1122. [CrossRef]
- Johnson, S. D., & Midgley, J. J. (1997). Fly pollination of Gorteria diffusa (Asteraceae), and a possible mimetic function for dark spots on the capitulum. American Journal of Botany, 84(4), 429–436. [CrossRef]
- Jokioja, J., Yang, B., & Linderborg, K. M. (2021). Acylated anthocyanins: A review on their bioavailability and effects on postprandial carbohydrate metabolism and inflammation. Comprehensive Reviews in Food Science and Food Safety, 20(6), 5570–5615. [CrossRef]
- Kearsley, M. W., & Rodriguez, N. (1981). The stability and use of natural colours in foods: Anthocyanin, β-carotene and riboflavin. International Journal of Food Science & Technology, 16(4), 421–431. [CrossRef]
- Lacey, E. P., Lovin, M. E., Richter, S. J., & Herington, D. A. (2010). Floral Reflectance, Color, and Thermoregulation: What Really Explains Geographic Variation in Thermal Acclimation Ability of Ectotherms? The American Naturalist, 175(3), 335–349. [CrossRef]
- Landi, M., Tattini, M., & Gould, K. S. (2015). Multiple functional roles of anthocyanins in plant-environment interactions. Environmental and Experimental Botany, 119, 4–17. [CrossRef]
- Lee, Y.-M., Yoon, Y., Yoon, H., Park, H.-M., Song, S., & Yeum, K.-J. (2017). Dietary Anthocyanins against Obesity and Inflammation. Nutrients, 9(10), Article 10. [CrossRef]
- Li, A., Xiao, R., He, S., An, X., He, Y., Wang, C., Yin, S., Wang, B., Shi, X., & He, J. (2019). Research Advances of Purple Sweet Potato Anthocyanins: Extraction, Identification, Stability, Bioactivity, Application, and Biotransformation. Molecules, 24(21), 3816. [CrossRef]
- Li, H.-T., Luo, Y., Gan, L., Ma, P.-F., Gao, L.-M., Yang, J.-B., Cai, J., Gitzendanner, M. A., Fritsch, P. W., Zhang, T., Jin, J.-J., Zeng, C.-X., Wang, H., Yu, W.-B., Zhang, R., van der Bank, M., Olmstead, R. G., Hollingsworth, P. M., Chase, M. W., … Li, D.-Z. (2021). Plastid phylogenomic insights into relationships of all flowering plant families. BMC Biology, 19(1), 232. [CrossRef]
- Lightbourn, G. J., Griesbach, R. J., Novotny, J. A., Clevidence, B. A., Rao, D. D., & Stommel, J. R. (2008). Effects of Anthocyanin and Carotenoid Combinations on Foliage and Immature Fruit Color of Capsicum annuum L. Journal of Heredity, 99(2), 105–111. [CrossRef]
- Liu, Y., Li, M., Li, T., Chen, Y., Zhang, L., Zhao, G., Zhuang, J., Zhao, W., Gao, L., & Xia, T. (2020). Airborne fungus-induced biosynthesis of anthocyanins in Arabidopsis thaliana via jasmonic acid and salicylic acid signaling. Plant Science, 300, 110635. [CrossRef]
- Liu, Y., Liu, Y., Tao, C., Liu, M., Pan, Y., & Lv, Z. (2018). Effect of temperature and pH on stability of anthocyanin obtained from blueberry. Journal of Food Measurement and Characterization, 12(3), 1744–1753. [CrossRef]
- Lloyd, A., Brockman, A., Aguirre, L., Campbell, A., Bean, A., Cantero, A., & Gonzalez, A. (2017). Advances in the MYB–bHLH–WD Repeat (MBW) Pigment Regulatory Model: Addition of a WRKY Factor and Co-option of an Anthocyanin MYB for Betalain Regulation. Plant and Cell Physiology, 58(9), 1431–1441. [CrossRef]
- Luo, J., Nishiyama, Y., Fuell, C., Taguchi, G., Elliott, K., Hill, L., Tanaka, Y., Kitayama, M., Yamazaki, M., Bailey, P., Parr, A., Michael, A. J., Saito, K., & Martin, C. (2007). Convergent evolution in the BAHD family of acyl transferases: Identification and characterization of anthocyanin acyl transferases from Arabidopsis thaliana. The Plant Journal, 50(4), 678–695. [CrossRef]
- Maleknia, S. D., & Downard, K. M. (2016). New Anthocyanins from Black Elderberry of Inhibitory Potential Revealed by Mass Spectrometry. The Natural Products Journal, 6(2), 94–102.
- Markham, K. R., Bloor, S. J., Nicholson, R., Rivera, R., Shemluck, M., Kevan, P. G., & Michener, C. (2004). Black Flower Coloration in Wild Lisianthius nigrescens: Its Chemistry and Ecological Consequences. Zeitschrift Für Naturforschung C, 59(9–10), 625–630. [CrossRef]
- Mazzucato, A., Willems, D., Bernini, R., Picarella, M. E., Santangelo, E., Ruiu, F., Tilesi, F., & Soressi, G. P. (2013). Novel phenotypes related to the breeding of purple-fruited tomatoes and effect of peel extracts on human cancer cell proliferation. Plant Physiology and Biochemistry, 72, 125–133. [CrossRef]
- Menconi, J., Perata, P., & Gonzali, S. (2023). Novel R2R3 MYB transcription factors regulate anthocyanin synthesis in Aubergine tomato plants. BMC Plant Biology, 23(1), 148. [CrossRef]
- Mes, P. J., Boches, P., Myers, J. R., & Durst, R. (2008). Characterization of Tomatoes Expressing Anthocyanin in the Fruit. Journal of the American Society for Horticultural Science, 133(2), 262–269. [CrossRef]
- Mori, K., Goto-Yamamoto, N., Kitayama, M., & Hashizume, K. (2007). Loss of anthocyanins in red-wine grape under high temperature. Journal of Experimental Botany, 58(8), 1935–1945. [CrossRef]
- Murre, C., Bain, G., van Dijk, M. A., Engel, I., Furnari, B. A., Massari, M. E., Matthews, J. R., Quong, M. W., Rivera, R. R., & Stuiver, M. H. (1994). Structure and function of helix-loop-helix proteins. Biochimica Et Biophysica Acta, 1218(2), 129–135. [CrossRef]
- Naing, A. H., Park, K. I., Ai, T. N., Chung, M. Y., Han, J. S., Kang, Y.-W., Lim, K. B., & Kim, C. K. (2017). Overexpression of snapdragon Delila (Del) gene in tobacco enhances anthocyanin accumulation and abiotic stress tolerance. BMC Plant Biology, 17, 65. [CrossRef]
- Noda, N., Aida, R., Kishimoto, S., Ishiguro, K., Fukuchi-Mizutani, M., Tanaka, Y., & Ohmiya, A. (2013). Genetic Engineering of Novel Bluer-Colored Chrysanthemums Produced by Accumulation of Delphinidin-Based Anthocyanins. Plant and Cell Physiology, 54(10), 1684–1695. [CrossRef]
- Noda, Y., Kneyuki, T., Igarashi, K., Mori, A., & Packer, L. (2000). Antioxidant activity of nasunin, an anthocyanin in eggplant peels. Toxicology, 148(2), 119–123. [CrossRef]
- Norgate, M., Boyd-Gerny, S., Simonov, V., Rosa, M. G. P., Heard, T. A., & Dyer, A. G. (2010). Ambient Temperature Influences Australian Native Stingless Bee (Trigona carbonaria) Preference for Warm Nectar. PLOS ONE, 5(8), e12000. [CrossRef]
- Okitsu, N., Noda, N., Chandler, S., & Tanaka, Y. (2018). Flower Color and Its Engineering by Genetic Modification. In J. Van Huylenbroeck (Hrsg.), Ornamental Crops (S. 29–62). Springer International Publishing. [CrossRef]
- Peng, M., Hudson, D., Schofield, A., Tsao, R., Yang, R., Gu, H., Bi, Y.-M., & Rothstein, Steven. J. (2008). Adaptation of Arabidopsis to nitrogen limitation involves induction of anthocyanin synthesis which is controlled by the NLA gene. Journal of Experimental Botany, 59(11), 2933–2944. [CrossRef]
- Ramsay, N. A., & Glover, B. J. (2005). MYB-bHLH-WD40 protein complex and the evolution of cellular diversity. Trends in Plant Science, 10(2), 63–70. [CrossRef]
- Rick, C., Cisneros, P., Chetelat, R., & DeVerna, J. (1994). Abg—A gene on chromosome 10 for purple fruit derived from S. lycopersicoides.
- Rosinski, J. A., & Atchley, W. R. (1998). Molecular evolution of the Myb family of transcription factors: Evidence for polyphyletic origin. Journal of Molecular Evolution, 46(1), 74–83. [CrossRef]
- Rowan, D. D., Cao, M., Lin-Wang, K., Cooney, J. M., Jensen, D. J., Austin, P. T., Hunt, M. B., Norling, C., Hellens, R. P., Schaffer, R. J., & Allan, A. C. (2009). Environmental regulation of leaf colour in red 35S:PAP1 Arabidopsis thaliana. New Phytologist, 182(1), 102–115. [CrossRef]
- Sapir, M., Oren-Shamir, M., Ovadia, R., Reuveni, M., Evenor, D., Tadmor, Y., Nahon, S., Shlomo, H., Chen, L., Meir, A., & Levin, I. (2008). Molecular Aspects of Anthocyanin fruit Tomato in Relation to high pigment-1. Journal of Heredity, 99(3), 292–303. [CrossRef]
- Sasaki, K., & Takahashi, T. (2002). A flavonoid from Brassica rapa flower as the UV-absorbing nectar guide. Phytochemistry, 61(3), 339–343. [CrossRef]
- Schaefer, H. M., Levey, D. J., Schaefer, V., & Avery, M. L. (2006). The role of chromatic and achromatic signals for fruit detection by birds. Behavioral Ecology, 17(5), 784–789. [CrossRef]
- Schwinn, K., Miosic, S., Davies, K., Thill, J., Gotame, T. P., Stich, K., & Halbwirth, H. (2014). The B-ring hydroxylation pattern of anthocyanins can be determined through activity of the flavonoid 3′-hydroxylase on leucoanthocyanidins. Planta, 240(5), 1003–1010. [CrossRef]
- Smith, T. F., Gaitatzes, C., Saxena, K., & Neer, E. J. (1999). The WD repeat: A common architecture for diverse functions. Trends in Biochemical Sciences, 24(5), 181–185. [CrossRef]
- Sun, C., Deng, L., Du, M., Zhao, J., Chen, Q., Huang, T., Jiang, H., Li, C.-B., & Li, C. (2020). A Transcriptional Network Promotes Anthocyanin Biosynthesis in Tomato Flesh. Molecular Plant, 13(1), 42–58. [CrossRef]
- Thill, J., Miosic, S., Ahmed, R., Schlangen, K., Muster, G., Stich, K., & Halbwirth, H. (2012). „Le Rouge et le Noir“: A decline in flavone formation correlates with the rare color of black dahlia (Dahlia variabilis hort.) flowers. BMC Plant Biology, 12(1), 225. [CrossRef]
- Thomas, M. M., Rudall, P. J., Ellis, A. G., Savolainen, V., & Glover, B. J. (2009). Development of a complex floral trait: The pollinator-attracting petal spots of the beetle daisy, Gorteria diffusa (Asteraceae). American Journal of Botany, 96(12), 2184–2196. [CrossRef]
- Tikhomirov, B., Shamurin, V., & Shtepa, V. (1960). The temperature of arctic plants. https://scholar.google.com/scholar_lookup?journal=Russ.+Acad.+Sci.&title=The+temperature+of+arctic+plants&author=B.+Tikhomirov&author=V.+Shamurin&author=V.+Shtepa&volume=3&publication_year=1960&pages=429-442&.
- Van Buren, J. P., Hrazdina, G., & Robinson, W. B. (1974). COLOR OF ANTHOCYANIN SOLUTIONS EXPRESSED IN LIGHTNESS AND CHROMATICITY TERMS. Effect of pH and Type of Anthocyanin. Journal of Food Science, 39(2), 325–328. [CrossRef]
- van der Kooi, C. J., & Stavenga, D. G. (2019). Vividly coloured poppy flowers due to dense pigmentation and strong scattering in thin petals. Journal of Comparative Physiology A, 205(3), 363–372. [CrossRef]
- Wang, X., Hansen, C., & Allen, K. (2013). Identification of Anthocyanins Isolated from Black Bean Canning Wastewater by Macroporous Resin Using Optimized Conditions. Food and Nutrition Sciences, 4(8), Article 8. [CrossRef]
- Weatherall, I. L., & Lee, W. G. (1991). Instrumental evaluation of some New Zealand fruit colours using CIELAB values. New Zealand Journal of Botany, 29(2), 197–205. [CrossRef]
- Winkel-Shirley, B. (2001). Flavonoid Biosynthesis. A Colorful Model for Genetics, Biochemistry, Cell Biology, and Biotechnology. Plant Physiology, 126(2), 485–493. [CrossRef]
- Zhang, Y., Butelli, E., & Martin, C. (2014). Engineering anthocyanin biosynthesis in plants. Current Opinion in Plant Biology, 19, 81–90. [CrossRef]
- Zhang, Y., Chu, G., Hu, Z., Gao, Q., Cui, B., Tian, S., Wang, B., & Chen, G. (2016). Genetically engineered anthocyanin pathway for high health-promoting pigment production in eggplant. Molecular Breeding, 36(5), 54. [CrossRef]
- Zhao, Y.-W., Wang, C.-K., Huang, X.-Y., & Hu, D.-G. (2021). Anthocyanin stability and degradation in plants. Plant Signaling & Behavior, 16(12), 1987767. [CrossRef]
- Zozio, S., Pallet, D., & Dornier, M. (2011). Evaluation of anthocyanin stability during storage of a coloured drink made from extracts of the Andean blackberry ( Rubus glaucus Benth.), açai ( Euterpe oleracea Mart.)and black carrot ( Daucus carota L.). Fruits, 66(3), 203–215. [CrossRef]
Figure 1.
Simplified overview of the main reactions of the general anthocyanin pathway. The enzymes involved in the synthesis are abbreviated as follows: PAL- phenylalanine ammonia-lyase, C4H - cinnamic acid 4-hydroxylase, 4CL - 4-coumarate-CoA ligase, CHS - chalcone synthase, CHI - chalcone isomerase, F3H - flavanone 3-hydroxylase, F3’H - flavonoid 3’-hydroxylase, F3’5’H - flavonoid 3’,5’-hydroxylase, DFR - dihydroflavonol 4-reductase, LDOX - leucoanthocyanidin dioxygenase, arGST - anthocyanin-related Glutathione S-transferase.
Figure 1.
Simplified overview of the main reactions of the general anthocyanin pathway. The enzymes involved in the synthesis are abbreviated as follows: PAL- phenylalanine ammonia-lyase, C4H - cinnamic acid 4-hydroxylase, 4CL - 4-coumarate-CoA ligase, CHS - chalcone synthase, CHI - chalcone isomerase, F3H - flavanone 3-hydroxylase, F3’H - flavonoid 3’-hydroxylase, F3’5’H - flavonoid 3’,5’-hydroxylase, DFR - dihydroflavonol 4-reductase, LDOX - leucoanthocyanidin dioxygenase, arGST - anthocyanin-related Glutathione S-transferase.
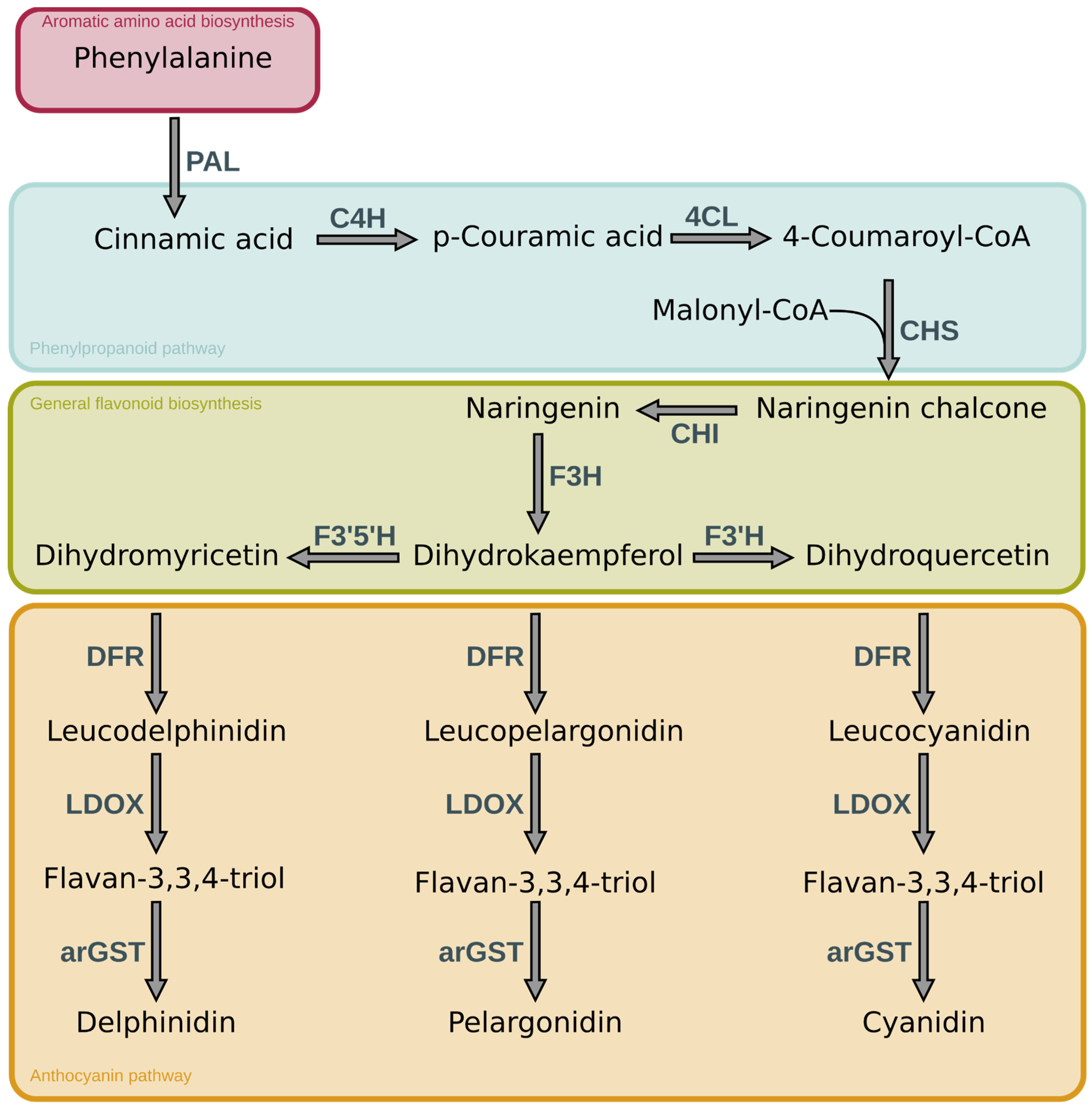
Figure 2.
Visual representation of the CIE L*a*b* coordinate with example colours and corresponding CIE values. L* indicates the lightness level of the colour, while a* and b* indicate magenta and yellow respectively, if a* and b* have a positive value and green and blue if the value is negative.
Figure 2.
Visual representation of the CIE L*a*b* coordinate with example colours and corresponding CIE values. L* indicates the lightness level of the colour, while a* and b* indicate magenta and yellow respectively, if a* and b* have a positive value and green and blue if the value is negative.

Figure 3.
Examples of dark pigmentation in plants. A: Berries (left to right): Blackberry (Rubus armeniacus), Black nightshade (Solanum nigrum L.), black currant (Ribes nigrum L.). B:Petal spots (left to right): Poppy (Papaver rhoeas), orange black-eyed Susan vine (Thunbergia alata), white black-eyed Susan vine (Thunbergia alata) C: Leaves (left to right): Black elderberry (Sambucus nigra ‘black lace’), black elephant ear (Colocasia rubra ‘Black Magic’), sweet potato vine (Ipomoea batatas ‘SolarTower Black’).
Figure 3.
Examples of dark pigmentation in plants. A: Berries (left to right): Blackberry (Rubus armeniacus), Black nightshade (Solanum nigrum L.), black currant (Ribes nigrum L.). B:Petal spots (left to right): Poppy (Papaver rhoeas), orange black-eyed Susan vine (Thunbergia alata), white black-eyed Susan vine (Thunbergia alata) C: Leaves (left to right): Black elderberry (Sambucus nigra ‘black lace’), black elephant ear (Colocasia rubra ‘Black Magic’), sweet potato vine (Ipomoea batatas ‘SolarTower Black’).

Figure 4.
Dark pigmentation occurrence mapped to a phylogenetic tree of plant families. For the highlighted families at least one species with dark pigmentation has been found. The colours represent the location of the pigmentation: green = foliage, pink = fruit and purple = flower. In total 36 species were included in the highlighting of relevant families. Tree topology is derived from Li et al. (H.-T. Li et al., 2021). A full list of species can be found in Supplementary file 1.
Figure 4.
Dark pigmentation occurrence mapped to a phylogenetic tree of plant families. For the highlighted families at least one species with dark pigmentation has been found. The colours represent the location of the pigmentation: green = foliage, pink = fruit and purple = flower. In total 36 species were included in the highlighting of relevant families. Tree topology is derived from Li et al. (H.-T. Li et al., 2021). A full list of species can be found in Supplementary file 1.
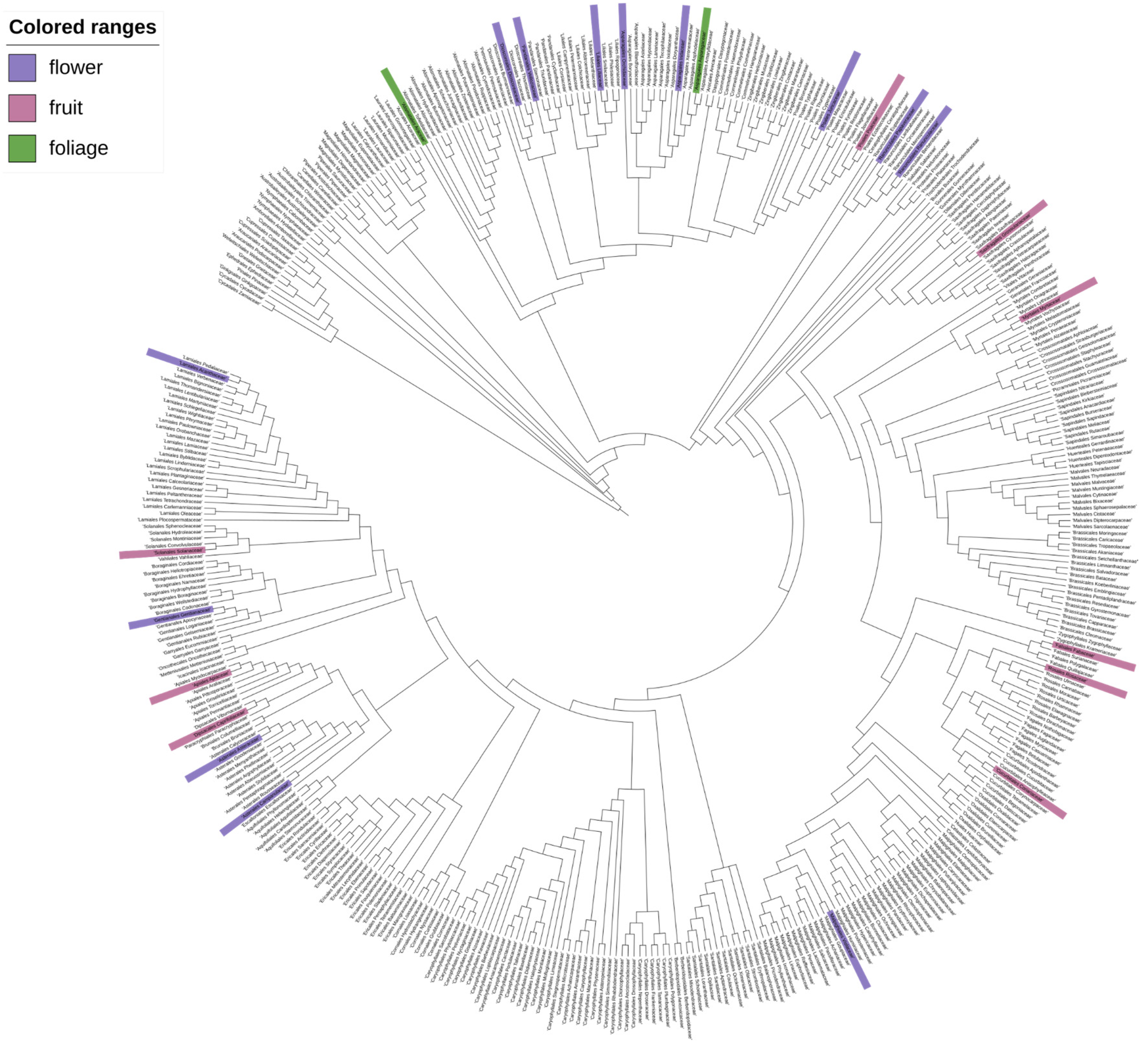

Table 1.
Overview of the most abundant anthocyanins and their chemical structure.
| Anthocyanin | R3′ | R5′ | chemical structure |
|---|---|---|---|
| Pelargonidin | −H | −H | 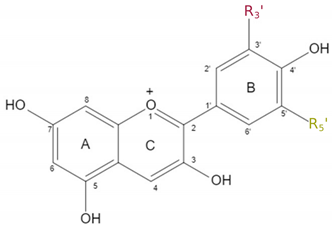 |
| Cyanidin | −OH | −H | |
| Delphinidin | −OH | −OH | |
| Peonidin | −OCH3 | −H | |
| Petunidin | −OH | −OCH3 | |
| Malvidin | −OCH3 | −OCH3 |
Table 2.
Examples of anthocyanins associated with black pigmentation in fruits and plants.
| Species | Anthocyanin | Reference |
|---|---|---|
|
Viola tricolor (pansy) |
delphinidin-5-O-glucoside-3-O-[4-p-coumaroylrhamnosyl(1-6)-glucoside] | (Goto et al., 1978) |
|
Lisianthius nigrescens (Flower of Death) |
delphinidin-3-O-rhamnol(1-6)galactoside | (Markham et al., 2004) |
| Dahlia variabilis (garden dahlia) | cyanidin-3-(6″-malonylglucoside)-5-glucoside | (Deguchi et al., 2016) |
| Capsicum annuum (pepper) | delphinidin-3-p-coumaroyl-rutinoside-5-glucoside | (Lightbourn et al., 2008) |
| Solanum melongena (Eggplant) | Delphinidin-3-(p-coumaroylrutinoside)-5-glucoside (nasunin) | (Y. Noda et al., 2000) |
| Solanum lycopersicum (Tomato) | petunidin-3-O-[6″-O-(4‴-O-E-p-coumaroyl-α-rhamnopyranosyl)-β-glucopyranoside]-5-O-β-glucopyranoside (petanin) malvidin-3-O-[6″-O-(4‴-O-E-p-coumaroyl-α-rhamnopyranosyl)-β-glucopyranoside]-5-O-β-glucopyranoside (negretein) |
(Blando et al., 2019) |
| black sorghum (sorghum) | Luteolinidin and apigeninidin * | (Awika et al., 2005) |
| Phaseolus vulgaris (Black Bean) | delphinidin 3-glucoside, petunidin 3- glucoside and maldvidin 3-glucoside | (Wang et al., 2013) |
| Sambucus nigra (black elderberry) | cyanidin-3-O-sambubioside, prenylated and other derivatives of cyanidin-3-acetylrutinoside | (Maleknia & Downard, 2016) |
*sugar moiety decoration unclear.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



