
Submitted:
25 June 2024
Posted:
27 June 2024
You are already at the latest version
Abstract
The conversion of caatinga vegetation areas into pastures in the Brazilian semiarid region has depleted soil fertility due to degradation. The natural regeneration of the caatinga has been the alternative adopted to restore soil fertility. However, the real effectiveness of this practice in re-covering organic carbon (SOC) and nutrient stocks is unknown. This work evaluated the effect of natural regeneration of the caatinga vegetation on the restoration of levels and stocks of SOC, N, and P as a strategy to maintain the sustainability of the environment. We collected soil samples every 5 cm, up to 20 cm deep, in areas of degraded pasture, caatinga in regeneration (10 years), and regenerated caatinga (35 years) to evaluate the levels and stocks of SOC, N, and P. The depth sig-nificantly affects the levels and stocks of SOC, N, and P. Stocks and levels of SOC, N, and P were higher in regenerated caatinga > regenerating caatinga > degraded pasture. The regenerated caatinga increased SOC, N, and P stocks by 115%, 110%, and 117%, respectively, compared to the degraded pasture. Although 35 years of regeneration of the caatinga has improved soil fertility, the levels and stocks of SOC, N, and P remain low.
Keywords:
dry forest soil
; land use change
; soil degradation and regeneration
; environmental sustainability
1. Introduction
Arid and semiarid landscapes represent 45% of the Earth's surface [1], and in these areas, dry forests are crucial for providing important ecosystem services locally and globally [2]. The native vegetation is an essential regulator of climate, biodiversity, biogeochemical cycles, and the soil's physical, chemical, and biological properties. Additionally, native vegetation acts as a sink for greenhouse gases [3,4]. However, most dry forest areas have been converted into human-modified landscapes [5].
In Brazil, the Caatinga dry forest has enormous endemic biodiversity and is considered one of the dry forests with the most incredible biodiversity around the globe, containing more than 4,000 plant species [6,7,8,9]. Despite this, the Caatinga has been severely devastated due to its replacement by pasture areas, which has led to severe environmental problems such as loss of biodiversity and desertification [8,10,11,12,13]. Overgrazing is one of the practices with the most degraded soils in the Brazilian semiarid region. Soil compaction caused by excessive animal trampling leads to the loss of organic matter by increasing soil erosion, which directly affects microbial communities, as well as the availability of nutrients in the soil, and an increase in greenhouse gas emissions [14,15,16,17].
The contents and stocks of soil organic carbon (SOC), nitrogen (N), and phosphorus (P) in Brazilian semiarid soils are strongly controlled by the rates of addition and removal of organic matter, which is significantly affected by the management adopted. SOC plays a vital role in several soil properties (physical, chemical, and biological), as well as nitrogen and phosphorus, which are essential for the nutrition of native vegetation. Thus, understanding the effects caused by changes in land use on the content of these elements becomes important since efforts have been made to identify soil management systems that favor their increases and/or maintain their concentrations in order to create strategies for soil management that reduce the impact of agriculture on the environment.
Brazilian semiarid soils are generally poor in organic matter [18]. The conversion of caatinga into degraded pasture areas drastically reduces carbon stocks and nutrient contents such as N and P, resulting in the loss of fertility and productive potential of Brazilian semiarid soils [19]. After degradation, these areas are generally abandoned and left fallow. Therefore, restoring soil fertility and increasing organic matter and nutrient levels is crucial to maintaining environmental sustainability.
Reforestation and natural regeneration are essential plant restoration strategies used in the Caatinga. These ecological restoration strategies effectively sequester SOC and soil nutrients, restore degraded ecosystems, and improve soil quality [20]. However, the natural regeneration of the Caatinga occurs slowly due to the loss of organic matter, nutrients, and water scarcity. Thus, there is a wide range of information about the effects of natural regeneration of the caatinga on SOC and soil nutrients. This study aimed to evaluate the effectiveness of caatinga natural regeneration over a degraded pasture to restore the concentrations and stocks of SOC, N, and P, and their distributions in depth, in soils representative of the Brazilian semiarid region, as a strategy to maintain the sustainability of the Brazilian semiarid soils.
2. Materials and Methods
2.1. Study Area
The research was conducted at Fazenda Lagoinha (Figure 1), located in the municipality of Serra Talhada, in the microregion of Vale do Pajeú, in the hinterland of Pernambuco. According to the Köppen classification system, the climate is BShw' semiarid, hot and dry, with precipitation occurring between December and May. The highest precipitation values are observed in March, with an annual average of 642 mm [21]. The predominant soils in the study area are Chromic Luvissol and Leptosol (Figure 2), which are shallow and have a low degree of weathering.
Currently, the study area consists of areas with degraded pasture, fallow areas with the presence of caatinga vegetation under natural regeneration at an intermediate size (10 years in regeneration), and areas with regenerated caatinga vegetation, predominantly angico (Anadenanthera colubrina (Vell.) Brenan) in tree-shrub form, with 35 years from fallow until regeneration (Figure 1).
2.2. Soil Sampling
This study evaluated the contents and stocks of SOC, N, and P in the top 20 cm layer because this layer is more sensitive to changes in land use compared to deeper layers [22]. Soil samples were collected from three different land use conditions: degraded pasture, regenerating caatinga, and regenerated caatinga. The samples were gathered using a sampling grid with 10 x 10 m equidistant points in a 50 x 50 m area. In each area studied, soil samples were taken at four depth layers: 0-5, 5-10, 10-15, and 15-20 cm. There were 36 sampling points at each depth, resulting in 144 soil samples for each area. The sampling grids were set up under the same topographic conditions in all three areas.
2.3. Chemical and Physical Properties Analysis
To evaluate the chemical attributes of the soils (Table 1), we measured the pH in water (1:2.5). Available P was extracted using the methodology of Olsen et al. [23] and measured by colorimetry. The exchangeable cations – Ca2+, Mg2+, Na+ e K+, were extracted with 1 mol L-1 ammonium acetate solution, with Ca2+ and Mg2+ determined by atomic absorption spectrophotometry and Na+ and K+ by flame emission photometry. The cation exchange capacity was determined by the sodium acetate and ammonium acetate 1 mol L-1 method [24]. We determined the SOC content using the Walkey Black method, adapted by Mendonça and Matos [25], and total nitrogen using the Embrapa method [26]. In the physical characterization (Table 1), the granulometric composition was determined [26] for textural comparison of the areas and to verify the influence of texture on the organic carbon content of the study areas.
2.4. SOC, P and N Stocks
SOC, N, and P stocks were calculated separately for each soil layer studied in the three areas and expressed on an area basis, as described by Carvalho et al. [27], using the equation:
where X represents the stock of SOC, N, or P in the studied layer (Mg ha-1); Xi is the SOC, N, or P content (g kg-1) in the study layer; BD is the soil density in the studied layer (kg dm-3); and H is the thickness of the layer considered (cm).
X Stock = (Xi x BD x H)/10,
Don et al. [28] emphasized the crucial need to correct carbon stock calculations due to variations in soil density following land use changes. Therefore, the stocks of SOC, N, and P were subsequently corrected for an equivalent soil mass in both study areas, according to Ellert and Bettany [29]:
where Xc Stock is the carbon stock run in the layer; X is the uncorrected carbon stock of the layer of interest; BDref is the soil density of the layer in the reference area (regenerated caatinga); BD is the soil density in the layer of the area of interest.
Xc Stock = X x (BDref/BD),
Finally, the corrected carbon stock was meticulously calculated for the entire surface layer - 20 cm, by summing the corrected carbon stock of each layer for each area separately.
2.5. Statistical Analysis
The data underwent ANOVA analysis, followed by a comparison of means using the Tukey test (p < 0.05) to assess differences in contents between the studied areas and depths. Additionally, correlation analysis was conducted using the Pearson method.
3. Results
3.1. Effects of Caatinga Natural Regeneration on SOC, P and N Distribution
The depth significantly influences (p < 0.05) on the SOC, P, and N contents in the studied areas (Figure 3, Figure 4 and Figure 5). In all areas, the SOC, P, and N contents were higher on the surface and significantly reduced in the subsurface.
In the degraded pasture area (Figure 3A), the highest SOC content observed was 1.21 dag kg-1 (0-5 cm), and the lowest one was 0.56 dag kg-1 (15-20 cm), and this means that in degraded pasture areas, SOC contents are reduced by approximately 54% from the surface to 20 cm deep. In the regenerating caatinga area (Figure 3B), SOC varied from 2.04 dag kg-1 (0-5 cm) to 0.95 dag kg-1 (15-20 cm), implying percentage reductions of approximately 52% of surface up to 20 cm deep. In the regenerated caatinga (Figure 3C), the surface SOC content was 2.93 dag kg-1 (0-5 cm), while the lowest value observed was 1.08 dag kg-1(15-20 cm), representing reductions of approximately 63 % in SOC contents from the surface to 20 cm depth.
Regarding phosphorus (P) content, in the degraded pasture area, the highest observed content was 20.06 mg kg-1 in the surface layer (0-5 cm), while the lowest was 6.99 mg kg-1 in the 15-20 cm depth layer, indicating a reduction of approximately 65% in P content from the surface to the subsurface (Figure 4A). In the regenerating Caatinga area, P contents ranged from 45.64 mg kg-1 (0-5 cm) to 15.62 mg kg-1 (15-20 cm), corresponding to a decrease of about 66% down to a depth of 20 cm (Figure 4B). In the regenerated Caatinga, P varied from 54.69 mg kg-1 (0-5 cm) to 18.06 mg kg-1 (15-20 cm), reflecting a reduction of approximately 67%.
In the degraded pasture area, N showed a significant reduction, with the highest level found at 0.107 g kg-1 (0-5 cm) and the lowest at 0.05 g kg-1 (15-20 cm), indicating a decrease of approximately 53% from the surface to 20 cm depth (Figure 5A). Similarly, in the regenerating Caatinga area, N content also decreased significantly, ranging from 0.179 g kg-1 (0-5 cm) to 0.08 g kg-1 (15-20 cm), representing a reduction of about 55% over the same depth range (Figure 5B). In the regenerated Caatinga, the reduction in N content was even more pronounced, with a decrease of around 63% from the surface to a depth of 20 cm, where the highest content observed was 0.247 g kg-1 (0-5 cm), and the lowest was 0. 09 g kg-1 (15-20 cm) (Figure 5C).
3.2. SOC, P and N Concentrations as Influenced by Caatinga Natural Regeneration
The data shows that natural regeneration of the caatinga significantly (p<0.05) improves soil fertility. The SOC, P, and N contents were higher in the regenerated caatinga compared to regenerating caatinga and degraded pasture. SOC varied from 2.93 (in the 0 – 5 cm layer of the regenerated caatinga) to 0.56 dag kg-1 (in the 15 – 20 cm layer of the degraded pasture) (Figure 6A). Compared to degraded pasture, SOC increased by 66% and 140% in the 0 – 5 cm layer of the regenerating and regenerated caatinga areas, respectively. In the 5 – 10 cm layer, SOC gains were 59 and 107% for the regenerating caatinga and regenerated caatinga areas, respectively. For the 10 – 15 cm layer, the increases in SOC of regenerating caatinga and regenerated caatinga, in relation to degraded pasture, were 55 and 98%, while for the 15 – 20 cm layer, the SOC gains were 69 and 92%, respectively.
The phosphorus (P) varied across the study areas (Figure 6B), ranging from 54.7 mg kg-1 in the regenerated caatinga (0 – 5 cm) to 6.9 mg kg-1 in the degraded pasture (15 – 20 cm). The data from this study suggest that P are positively influenced (p < 0.05) by caatinga regeneration. At 0 – 5 cm layer, there were increases of 127% and 172% in P content compared to degraded pasture, in the regenerating and regenerated caatinga areas, respectively. In the 5 – 10 cm layer, the increases in P content were 93% and 61% in the regenerating and regenerated caatinga areas, respectively. In the 10 – 15 cm layer, only the regenerating caatinga showed significant increases in P, with a gain of 71% compared to the degraded pasture. However, at 15 – 20 cm, both regenerating and regenerated caatinga areas showed significant increases in P content – 123% and 158%, respectively, compared to the degraded pasture area.
The N contents varied between the study areas, ranging from 0.247 mg kg-1 in the regenerated caatinga (0 – 5 cm) to 0.04 mg kg-1 in the degraded pasture (15 – 20 cm) (Figure 6C). Caatinga regeneration significantly (p < 0.05) increased the N contents compared to degraded pasture. The results show that the increases in N content were as follows: 69% and 133% (0 – 5 cm), 67% and 108% (5 – 10 cm), 60% and 100% (10 – 15 cm), and 72% and 100% (15 – 20 cm) for the regenerating caatinga and regenerated caatinga, respectively.
3.3. SOC, P and N Stocks
In general, the Caatinga regeneration significantly (p < 0.05) increased SOC, N, and P stocks (Figure 7). The contents of SOC, N, and P stocks supported these findings, with higher concentrations observed in regenerated caatinga > regenerating caatinga > degraded pasture. Specifically, compared to degraded pasture, SOC stocks increased significantly by 64% and 115% in the regenerating and regenerated Caatinga areas, respectively (Figure 7A). While N stocks did not differ significantly between regenerating Caatinga and degraded pasture, the regenerated Caatinga exhibited a substantial increase of approximately 110% compared to degraded pasture (Figure 7B). By the other hand, P stocks showed no significant difference between regenerating and regenerated Caatinga (Figure 7C). This suggests that Caatinga is more effective in recovering P stocks compared to N or SOC. Specifically, P stocks increased by 109% and 117% in regenerating and regenerated Caatinga, respectively, relative to degraded pasture.
4. Discussion
Our results revealed that soil anthropization drastically reduces SOC, P, and N contents. We observed a greater accumulation of these elements on the topsoil (0 – 5 cm), which is expected due to the soil surface being the layer with greater deposition and accumulation of litter [30]. These findings are consistent with those reported by Gava et al. [19], who also found higher contents of SOC, N, and P in the surface layer (0 – 5 cm) under different land use conditions in the Brazilian semi-arid region. Since no fertilizers are applied in the study areas, the N and P distribution patterns in the soil align with the results for SOC. This suggests that the dynamics of N and P in Brazilian semiarid soils, under conditions of natural regeneration of the caatinga, are similar to that of SOC since organic matter is the only source of nutrients in the soils of these areas.
After forest regeneration, SOC and soil nutrient rates depend on climate, land use conditions, previous land use, soil clay content, type and density of vegetation cover, and sampling depth [22]. In our study, it was possible to verify the contribution of the natural regeneration of the caatinga on the SOC, P, and N contents, which resulted in the improvement of the natural fertility of semiarid soils through the reestablishment of vegetation cover. The higher accumulation of SOC, P, and N in the regenerated caatinga area compared to other areas is due to the increased vegetation, which contributes more organic matter to the soil. The contribution of plant residues to the soil surface promotes slow and gradual decomposition, ensuring continuous incorporation of organic material into the soil. The trend of increasing soil organic matter over time is consistent with the effects of reforestation and forest succession on soil organic matter accumulation.
The lower contents of SOC, P, and N observed in this study in degraded pasture areas occur mainly due to the removal of vegetation cover, which results in a low production of litter, reducing the supply of organic matter to the soil surface. In many cases, the limited amount of litter produced is consumed by animals as food, leaving the soil surface unprotected. The lack of soil cover, combined with excessive trampling, leads to soil compaction and increased susceptibility to water erosion, especially during heavy rainfall in the rainy season [31,32]. Consistent with our findings, Almeida et al. [10] conducted a 10-year comparison study between degraded pasture and regenerating Caatinga areas, observing reduced soil erosion and organic carbon losses in the regenerating Caatinga. They also noted higher nutrient contents in the regenerating Caatinga, with increases of 75% for nitrogen, 73% for phosphorus, and 34% for potassium.
The SOC, P, and N contents positively correlate with the clay content (Table 2), so the more clay there is, the higher the content of these elements in the soil. Conversely, the higher the silt content, the lower the SOC, P, and N contents due to their negative correlation with these nutrients. Additionally, P content was negatively correlated with Mg2+ and Na+ content (Table 2).
Clay is responsible for stabilizing colloidal or humic compounds in soil organic matter due to the adsorption process, providing protection due to the high specific surface area involved and the strong interaction between organic and inorganic colloids, which results in the formation of aggregates such as protection and the presence of stable organomineral complexes [33,34], which explains the positive correlation between clay and soil organic carbon content. Although the degraded pasture area had a higher clay content, the silt, the Mg2+, and Na+ contents were also higher than in the other areas (Table 1), and this also contributed strongly to the reductions in SOC, and consequently, the P and N contents, as their levels depend on soil organic matter, their main source.
The conversion of degraded areas into forests reestablishes nutrient cycling. Perennial land cover in native and regenerating forests provides greater and constant deposition of litter, which can contribute to the maintenance of soil moisture and lower soil surface temperatures, which can increase carbon stocks, phosphorus, and nitrogen in the topsoil [35]. Our study found that caatinga regeneration was more effective in recovering P stocks since, in 7 years of forest regeneration, P stocks did not differ from the regenerated caatinga. However, this was not the case for SOC and N, where 7 years of caatinga regeneration was insufficient to raise SOC and N stocks to the same level as in the 25-year-old regenerated caatinga. These findings suggest that for elements with low levels in Brazilian semi-arid soils, such as phosphorus in our study, caatinga regeneration can restore the element's stocks in a shorter period than elements that are naturally more abundant in semiarid soils.
It is important to note that, even with the caatinga regenerated (25 years regenerated), the SOC, N, and P stocks were low compared to other biomes. For example, Azevedo et al. [36] found SOC stocks of 136.68 Mg ha-1 for the Atlantic Forest, representing a SOC stock three times higher than in this study. Villela et al. [37] found N stocks varying from 0.98 to 1.86 Mg ha-1. Also, these values were much higher for the Atlantic Forest than those verified in our study for N stocks. Thus, the data verified in this study indicate that the slow Caatinga regeneration, as it is a dry forest, makes it difficult to reestablish soil nutrients. This occurs due to climatic conditions characterized by low rainfall and high temperatures, which limit plant growth and biomass production, leading to reduced litter deposition and, consequently, a decreased supply of nutrients to the soil.
Althoff et al. [38] suggest that SOC recovery in deforested caatinga areas may take up to 60 years to reach the original levels of dense vegetation before deforestation. Additionally, Araújo Filho et al. [39] indicate that in caatinga areas, the average time for SOC to return to its initial value in the soil is approximately 65 years. Like SOC, in our study, P and N followed the same tendency to return to the soil as the vegetation establishment time after deforestation. This implies that actions need to be taken to prevent deforestation of the caatinga, support reforestation efforts, and therefore preserve and recover SOC and nutrients in the soil. Such actions should include financial compensation to local producers who rely on income generated by livestock production for their subsistence to avoid deforestation of the caatinga.
5. Conclusions
The findings of this study highlight the importance of revegetating the caatinga to enhance and restore the natural soil fertility in the semiarid region of Brazil, particularly in the topsoil. Our research showed a significant increase in SOC, P, and N concentrations and stocks due to natural caatinga regeneration compared to degraded pasture, emphasizing the importance of this technique to restore soil fertility and contribute to the sustainability of the Brazilian semiarid soils. Our data also revealed that nutrient contents and stocks remained relatively low even after 35 years of caatinga regeneration.
For nutrients with deficient concentrations in Brazilian semiarid soils, such as those found for P in this study, a shorter regeneration time in the caatinga is sufficient to recover their concentrations and stocks. Given the caatinga's slow growth, we suggest additional research to explore methods to enhance its growth and establishment, thereby accelerating the restoration of SOC and soil nutrients and ensuring the soil's sustainability.
Author Contributions
Conceptualization, L.F.d.S.S. and L.G.M.P.; methodology, M.B.G.d.S.F., L.F.d.S.S. and L.G.M.P.; Field collections in laboratory work, L.F.d.S.S.; data curation, E.S.d.S. and G.B.J.; writing—original draft, L.F.d.S.S., L.G.M.P., D.O. and E.M.d.S.; writing—review and editing, J.O.N.d.S.; supervision, A.C.B., E.S.d.S.; funding acquisition, L.G.M.P. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the “National Council for Scientific and Technological Development-CNPq” (grants: 409937/2021-5).
Institutional Review Board Statement
Not applicable
Data Availability Statement
Data may be made available upon request to the corresponding author.
Acknowledgments
To INCT: NOWCDCB National Observatory of Water and Carbon Dynamics in the Caatinga Biome network, for providing areas to carry out the research.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Soper, F.M.; McCalley, C.K.; Sparks, K.; Sparks, J.P. Soil carbon dioxide emissions from the Mojave desert: Isotopic evidence for a carbonate source. Geophys. Res. Lett. 2017, 44, 245–251. [Google Scholar] [CrossRef]
- Powers, J.S.; Feng, X.; Sanchez-Azofeifa, A.; Medvigy, D. Focus on tropical dry forest ecosystems and ecosystem services in the face of global change. Environ. Res. Lett. 2018, 13, 1–5. [Google Scholar] [CrossRef]
- Chapuis-Lardy, L.; Wrage, N.; Metay, A.; Chotte, J.; Bernoux, M. Soils, a sink for N2O? A review. Glob. Chang. Biol. 2007, 13, 1–17. [Google Scholar] [CrossRef]
- Kirschbaum, M.U.F. To sink or burn? A discussion of the potential contributions of forests to greenhouse gas balances through storing carbon or providing biofuels. Biomass and Bioenergy 2003, 24, 297–310. [Google Scholar] [CrossRef]
- Tabarelli, M.; Filgueiras, B.K.C.; Ribeiro, E.M.S.; Lopes, A.V.; Leal, I.R. Tropical dry forests. in Encyclopedia of Biodiversity, Elsevier, 2024, 294–312. [CrossRef]
- Fernandes, M.F.; Cardoso, D.; Queiroz, L.P. An updated plant checklist of the Brazilian Caatinga seasonally dry forests and woodlands reveals high species richness and endemism. J. Arid Environ. 2020, 174, 1–10. [Google Scholar] [CrossRef]
- Koch, R.; Almeida-Cortez, J.S.; Kleinschmit, B. Revealing areas of high nature conservation importance in a seasonally dry tropical forest in Brazil: Combination of modelled plant diversity hot spots and threat patterns. J. Nat. Conserv. 2017, 35, 24–39. [Google Scholar] [CrossRef]
- Leal, I.R.; Silva, J.M.C.; Tabarelli, M.; Lacher, T.E. Changing the course of biodiversity conservation in the caatinga of northeastern Brazil. Conserv. Biol. 2005, 19, 701–706. [Google Scholar] [CrossRef]
- Santos, M.G.; Oliveira, M.T.; Figueiredo, K.V.; Falcao, H.M.; Arruda, E.C.; Almeida-Cortez, J.; Antonino, A.C. Caatinga, the Brazilian dry tropical forest: can it tolerate climate changes? Theor. Exp. Plant Physiol. 2014, 26, 83–99. [Google Scholar] [CrossRef]
- Almeida, C.L.; Araújo, J.C.; Costa, M.C.G.; Almeida, A.M.M.; Andrade, E.M. Fallow reduces soil losses and increases carbon stock in Caatinga. Floresta e Ambient. 2017, 24, 1–10. [Google Scholar] [CrossRef]
- Oliveira Filho, J.S.; Vieira, J.N.; Silva, E.M.R.; Oliveira, J.G.B.; Pereira, M.G.; Brasileiro, F.G. Assessing the effects of 17 years of grazing exclusion in degraded semi-arid soils: Evaluation of soil fertility, nutrients pools and stoichiometry. J. Arid Environ. 2019, 166, 1–10. [Google Scholar] [CrossRef]
- Silva, A.R.A.; Bezerra, F.M.L.; Lacerda, C.F.; Sousa, C.H.C.; Bezerra, M.A. Physiological responses of dwarf coconut plants under water deficit in salt-affected soils. Rev. Caatinga 2017, 30, 447–457. [Google Scholar] [CrossRef]
- Sousa, F.P.; Ferreira, T.O.; Mendonça, E.S.; Romero, R.E.; Oliveira, J.G.B. Carbon and nitrogen in degraded Brazilian semi-arid soils undergoing desertification. Agric. Ecosyst. Environ. 2012, 148, 11–21. [Google Scholar] [CrossRef]
- Dong, L.; Li, J.; Sun, J.; Yang, C. Soil degradation influences soil bacterial and fungal community diversity in overgrazed alpine meadows of the Qinghai-Tibet Plateau. Sci. Rep. 2021, 11, 1–11. [Google Scholar] [CrossRef]
- Li, Y.; Wang, S.; Jiang, L.; Zhang, L.; Cui, S.; Meng, F.; Zhou, Y. Changes of soil microbial community under different degraded gradients of alpine meadow. Agric. Ecosyst. Environ. 2016, 222, 213–222. [Google Scholar] [CrossRef]
- Wang, L. Delgado-Baquerizo, M.; Zhao, X.; Zhang, M.; Song, Y.; Cai, J.; Xin, X. Livestock overgrazing disrupts the positive associations between soil biodiversity and nitrogen availability. Funct. Ecol. 2020, 34, 1713–1720. [Google Scholar] [CrossRef]
- Zhou, H.; Zhang, D.; Jiang, Z.; Sun, P.; Xiao, H.; Yuxin, W.; Chen, J. Changes in the soil microbial communities of alpine steppe at Qinghai-Tibetan Plateau under different degradation levels. Sci. Total Environ. 2019, 651, 2281–2291. [Google Scholar] [CrossRef] [PubMed]
- Macedo, R.S.; Moro, L.; Lambais, É.O.; Lambais, G.R.; Bakker, A.P. Effects of degradation on soil attributes under caatinga in the Brazilian semi-arid. Rev. Árvore 2023, 47, 1–11. [Google Scholar] [CrossRef]
- Gava, C.A.T.; Giongo, V.; Signor, D.; Fernandes-Júnior, P.I. Land-use change alters the stocks of carbon, nitrogen, and phosphorus in a Haplic Cambisol in the Brazilian semi-arid region. Soil Use Manag. 2022, 38, 953–963. [Google Scholar] [CrossRef]
- Cheng, H.; Zhou, X.; Dong, R.; Wang, X.; Liu, G.; Li, Q. Natural vegetation regeneration facilitated soil organic carbon sequestration and microbial community stability in the degraded karst ecosystem. CATENA 2023, 222, 1–10. [Google Scholar] [CrossRef]
- Souza, R.; Feng, X.; Antonino, A.; Montenegro, S.; Souza, E.; Porporato, A. Vegetation response to rainfall seasonality and interannual variability in tropical dry forests. Hydrol. Process. 2016, 30, 3583–3595. [Google Scholar] [CrossRef]
- Lin, Y.-H.; Lee, P.-C.; Menyailo, O.V.; Cheng, C.-H. Changes in soil organic carbon concentration and stock after forest regeneration of agricultural fields in Taiwan. Forests 2021, 12, 1–12. [Google Scholar] [CrossRef]
- Olsen, S.R.; Cole, C.V.; Watandbe, F.; Dean, L. Estimation of available phosphorus in soil by extraction with sodium bicarbonate; US Department of Agriculture, 1954; Volume 53.
- USSL. Diagnosis and Improvement of Saline and Alkali Soils, vol. 46, no. 6. Washington, 1954. [CrossRef]
- Mendonça, E.S.; Matos, E.S. Matéria orgânica do solo: métodos de análises. Universidade Federal de Viçosa, Brasil, Viçosa - MG, 2005, pag. 81.
- EMBRAPA, Manual de métodos de análise de solos. 3rd ed. Embrapa, Rio de Janeiro, Brazil, 2017.
- Carvalho, J.L.N.; Cerri, C.E.P.; Feigl, B.J.; Píccolo, M.C.; Godinho, V.P.; Cerri, C.C. Carbon sequestration in agricultural soils in the Cerrado region of the Brazilian Amazon. Soil Tillage Res. 2009, 103, 342–349. [Google Scholar] [CrossRef]
- Don, A.; Schumacher, J.; Freibauer, A. Impact of tropical land-use change on soil organic carbon stocks - a meta-analysis. Glob. Chang. Biol. 2011, 17, 1658–1670. [Google Scholar] [CrossRef]
- Ellert, B.H.; Bettany, J.R. Calculation of organic matter and nutrients stored in soils under contrasting management regimes. Can. J. Soil Sci. 1995, 75, 529–538. [Google Scholar] [CrossRef]
- Pessoa, L.G.M.; Miranda, M.F.A.; Calheiros, A.S.; Freire, F.J.; Freire, M.B.G.S.; Barbosa, M.D. Marangon, L.C. Soil and litter properties from a riparian to non-riparian zone in a tropical dry forest. J. Environ. Anal. Prog 2023, 285–298. [Google Scholar] [CrossRef]
- Ebrahimi, M.; Khosravi, H.; Rigi, M. Short-term grazing exclusion from heavy livestock rangelands affects vegetation cover and soil properties in natural ecosystems of southeastern Iran. Ecol. Eng. 2016, 95, 10–18. [Google Scholar] [CrossRef]
- Schulz, K.; Voigt, K.; Beusch, C.; Almeida-Cortez, J.S.; Kowarik, I.; Walz, A.; and Cierjacks, A. Grazing deteriorates the soil carbon stocks of Caatinga forest ecosystems in Brazil. For. Ecol. Manage. 2016, 367, 62–70. [Google Scholar] [CrossRef]
- Berhongaray, G.; Alvarez, R. Soil carbon sequestration of Mollisols and Oxisols under grassland and tree plantations in South America - A review. Geoderma Reg. 2019, 18, 1–10. [Google Scholar] [CrossRef]
- Hewins, D.B.; Lyseng, M.P.; Schoderbek, D.F.; Alexander, M.; Willms, W.D.; Carlyle, C.N.; Bork, E.W. Grazing and climate effects on soil organic carbon concentration and particle-size association in northern grasslands. Sci. Rep. 2018, 8, 1–9. [Google Scholar] [CrossRef]
- Stockmann, U.; Adams, M.A.; Crawford, J.W.; Field, D.J.; Henakaarchchi, N.; Jenkins, M.; Zimmermann, M. The knowns, known unknowns and unknowns of sequestration of soil organic carbon. Agric. Ecosyst. Environ. 2013, 164, 80–99. [Google Scholar] [CrossRef]
- Azevedo, A.D.; Camara, R.; Francelino, M.R.; Pereira, M.G.; Leles, P.S.S. Estoque de carbono em áreas de restauração florestal da mata atlântica. FLORESTA 2018, 48, 1–12. [Google Scholar] [CrossRef]
- Villela, D.; Mattos, E.; Pinto, A.; Vieira, S.; Martinelli, L. Carbon and nitrogen stock and fluxes in coastal Atlantic Forest of southeast Brazil: potential impacts of climate change on biogeochemical functioning. Brazilian J. Biol. 2012, 72, 633–642. [Google Scholar] [CrossRef] [PubMed]
- Althoff, T.D.; Menezes, R.S.C.; Pinto, A.S.; Pareyn, F.G.C.; Carvalho, A.L.; Martins, J.C.R.; Sampaio, E.V.D.S.B. Adaptation of the century model to simulate C and N dynamics of Caatinga dry forest before and after deforestation. Agric. Ecosyst. Environ. 2018, 254, 26–34. [Google Scholar] [CrossRef]
- Araújo Filho, R.N.; Freire, M.B.G.S.; Wilcox, B.P.; West, J.B.; Freire, F.J.; Marques, F.A. Recovery of carbon stocks in deforested caatinga dry forest soils requires at least 60 years. For. Ecol. Manage. 2018, 407, 210–220. [Google Scholar] [CrossRef]
Figure 1.
Location map of the study areas – (a) Caatinga in regeneration, (b) Regenerated Caatinga, and (c) Degraded pasture.
Figure 1.
Location map of the study areas – (a) Caatinga in regeneration, (b) Regenerated Caatinga, and (c) Degraded pasture.
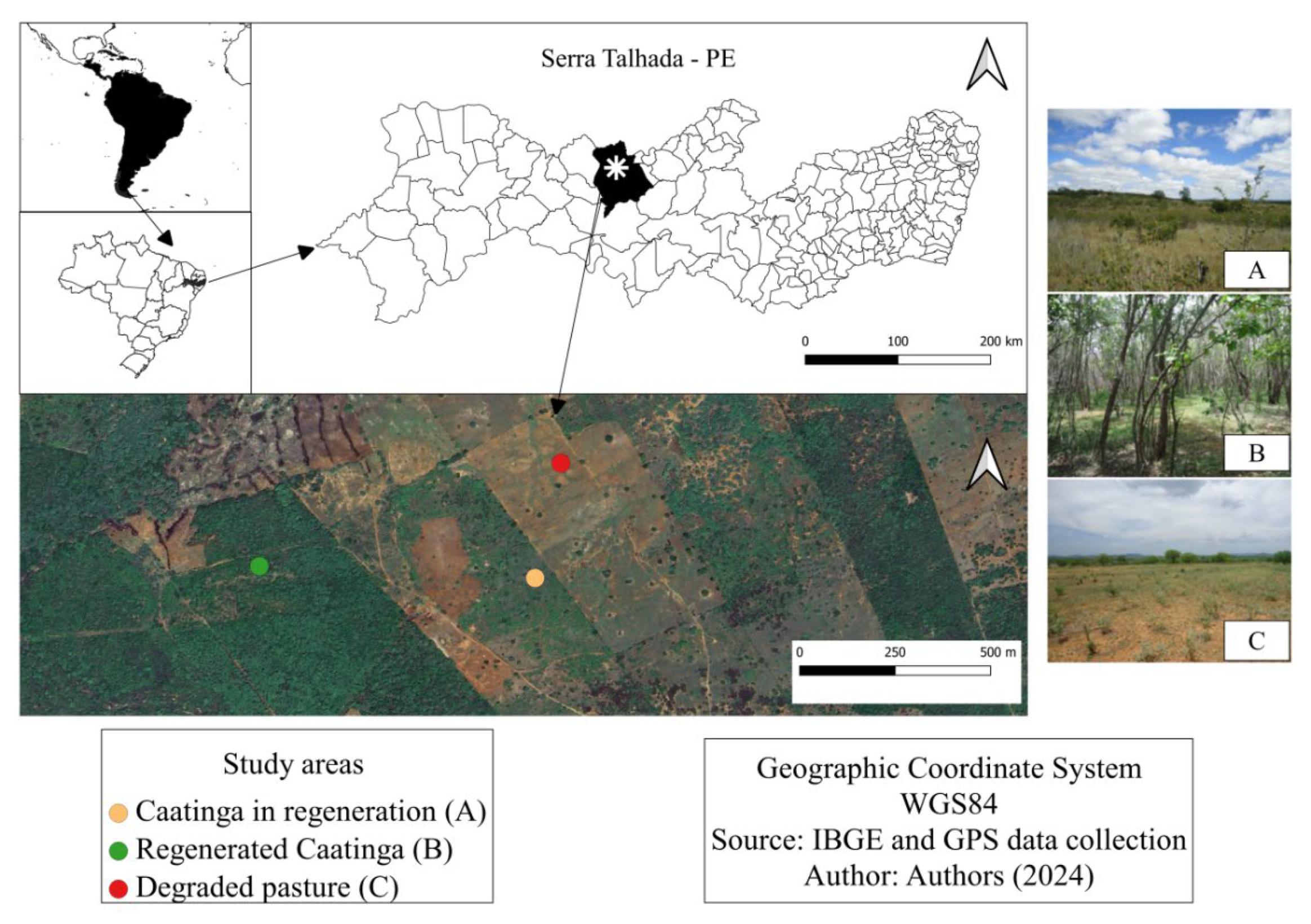
Figure 2.
Soil profiles of the study areas: (a) Degraded pasture – Luvissolo Crômico (Luvisol), (b) Caatinga in regeneration – Luvissolo Crômico (Luvisol), and (c) Regenerated Caatinga – Neossolo Litólico (Leptosol).
Figure 2.
Soil profiles of the study areas: (a) Degraded pasture – Luvissolo Crômico (Luvisol), (b) Caatinga in regeneration – Luvissolo Crômico (Luvisol), and (c) Regenerated Caatinga – Neossolo Litólico (Leptosol).

Figure 3.
Soil organic carbon contents as a function of depth for each study area: (A) Degraded Pasture, (B) Caatinga in regeneration, and (C) Regenerated Caatinga. Different letters at the depths differ by the Tukey test at 0.05 probability level.
Figure 3.
Soil organic carbon contents as a function of depth for each study area: (A) Degraded Pasture, (B) Caatinga in regeneration, and (C) Regenerated Caatinga. Different letters at the depths differ by the Tukey test at 0.05 probability level.
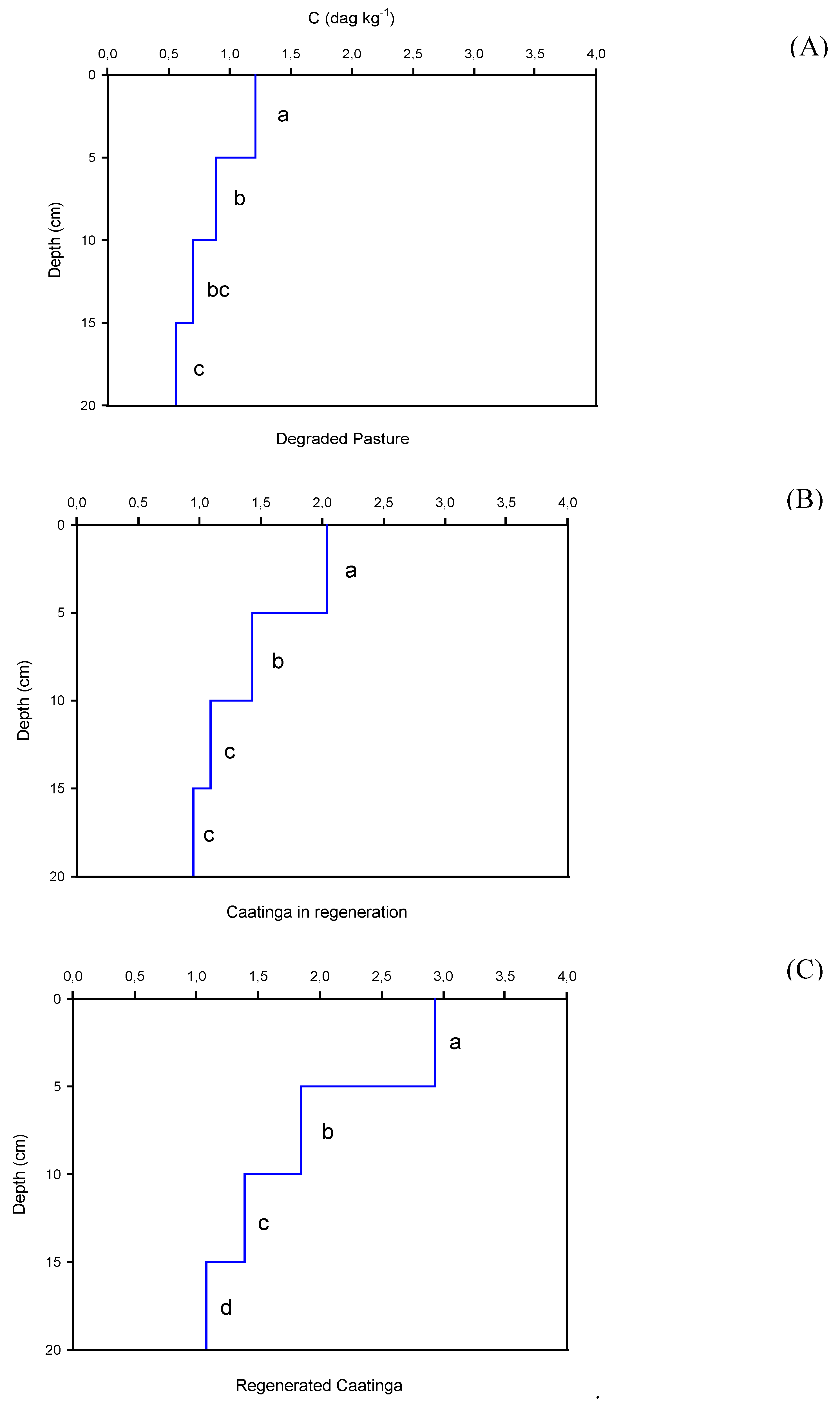
Figure 4.
Available phosphorous contents as a function of depth for each study area: (A) Degraded Pasture, (B) Caatinga in regeneration, and (C) Regenerated Caatinga. Different letters at the depths differ by the Tukey test at 0.05 probability level.
Figure 4.
Available phosphorous contents as a function of depth for each study area: (A) Degraded Pasture, (B) Caatinga in regeneration, and (C) Regenerated Caatinga. Different letters at the depths differ by the Tukey test at 0.05 probability level.
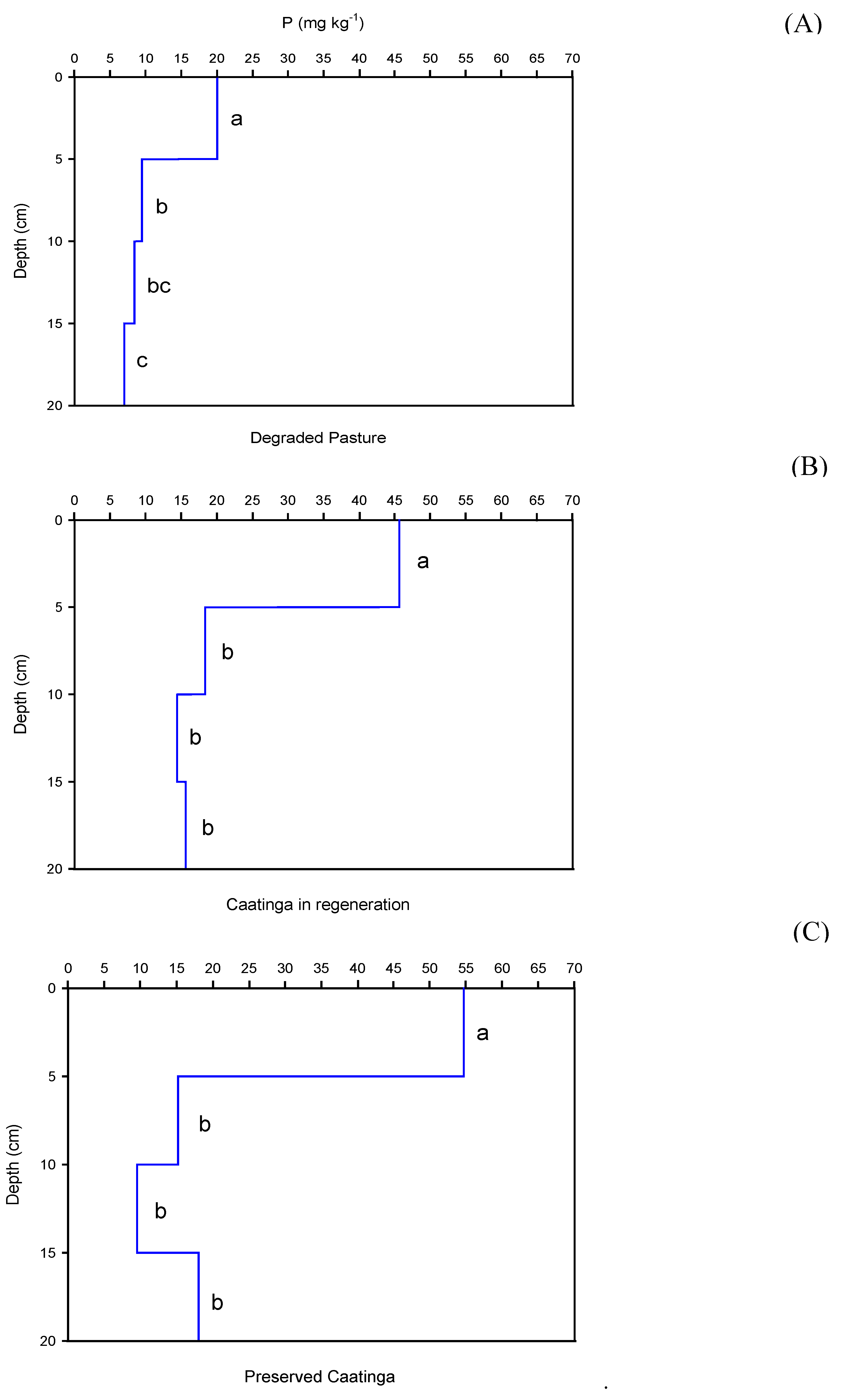
Figure 5.
Total nitrogen contents as a function of depth for each study area: (A) Degraded Pasture, (B) Caatinga in regeneration, and (C) Regenerated Caatinga. Different letters at the depths differ by the Tukey test at 0.05 probability level.
Figure 5.
Total nitrogen contents as a function of depth for each study area: (A) Degraded Pasture, (B) Caatinga in regeneration, and (C) Regenerated Caatinga. Different letters at the depths differ by the Tukey test at 0.05 probability level.
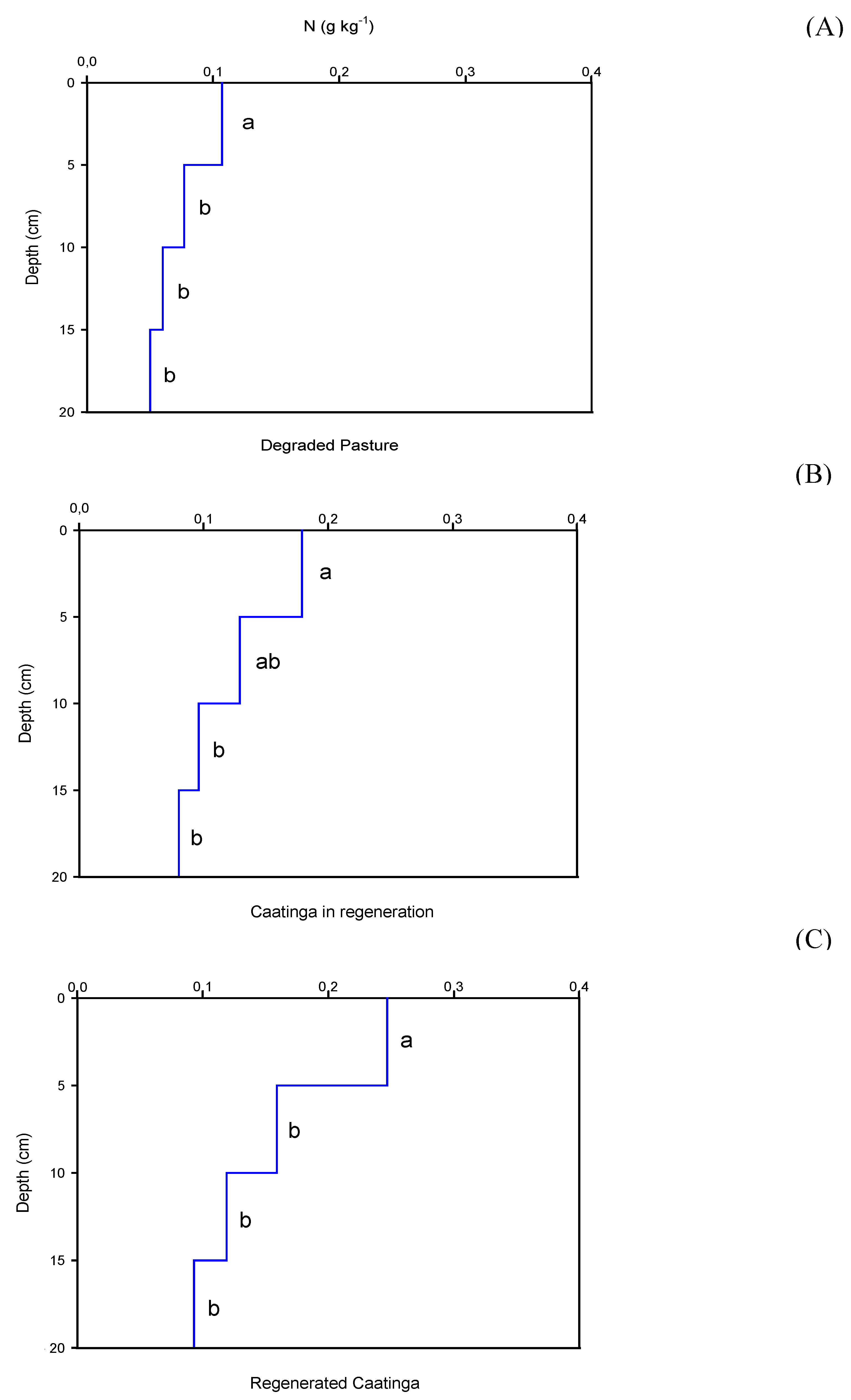
Figure 6.
Carbon (A), phosphorus (B), and nitrogen (C) contents in the soil depend on the land uses of the studied areas at the sampled depths. Different letters at each soil depth differ by the Tukey test at 0.05 probability level.
Figure 6.
Carbon (A), phosphorus (B), and nitrogen (C) contents in the soil depend on the land uses of the studied areas at the sampled depths. Different letters at each soil depth differ by the Tukey test at 0.05 probability level.
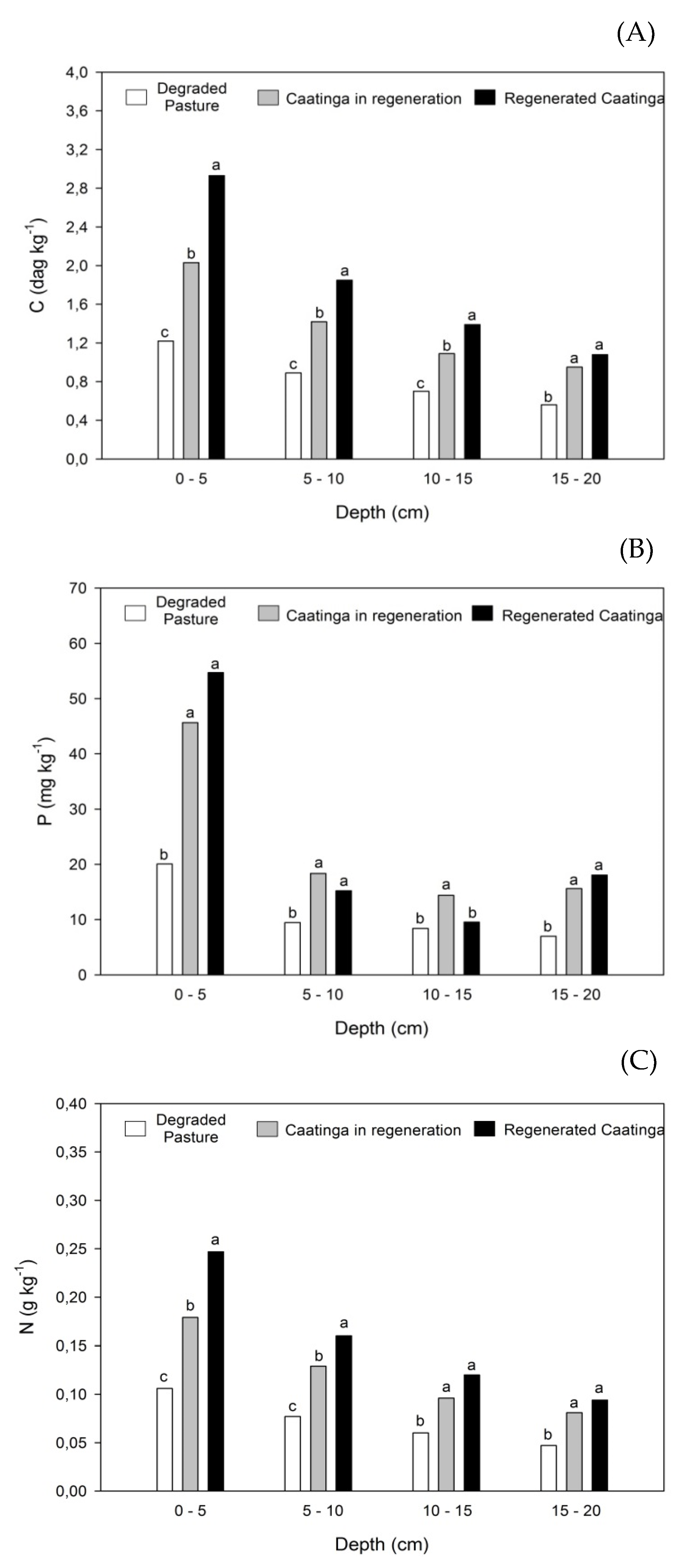
Figure 7.
Stocks of total soil organic carbon (A), nitrogen (B), and phosphorus (C) as influenced by the land-use of the studied sites, at 20 cm depth. Different letters differ by the Tukey test at 0.05 probability level.
Figure 7.
Stocks of total soil organic carbon (A), nitrogen (B), and phosphorus (C) as influenced by the land-use of the studied sites, at 20 cm depth. Different letters differ by the Tukey test at 0.05 probability level.
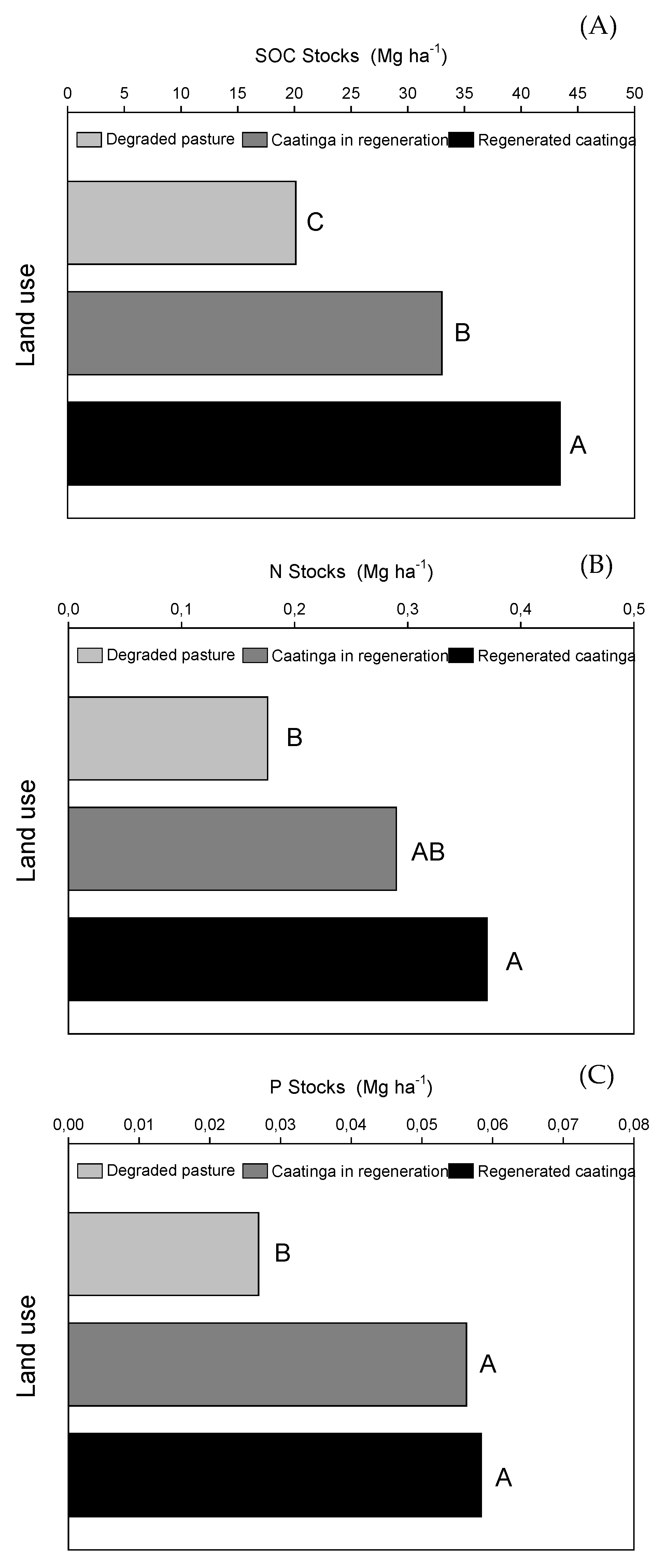

Table 1.
Chemical and physical attributes of soils in the study areas.
| Study area | pH | Ca2+ | Mg2+ | Na+ | K+ | CEC1 | CS2 | TS3 | Silt | Clay |
|---|---|---|---|---|---|---|---|---|---|---|
| ________________ cmolc kg-1 ________________ | ______________ % _____________ | |||||||||
| Degraded Pasture | 6.78 | 16.38 | 4.99 | 0.44 | 0.41 | 22.22 | 38.8 | 20.8 | 21.8 | 18.6 |
| Caatinga in regeneration | 6.95 | 15.33 | 2.91 | 0.28 | 0.39 | 18.91 | 43.2 | 20.6 | 18.8 | 17.4 |
| Regenerated Caatinga | 6.48 | 11.96 | 1.81 | 0.31 | 0.41 | 14.48 | 50.2 | 22.2 | 17.4 | 10.2 |
1CEC = Cation Exchange Capacity; 2CS = Coarse Sand; 3TS = Thin Sand.
Table 2.
Correlation coefficients for organic carbon, P, and N contents with chemical and physical attributes verified in the study areas.
Table 2.
Correlation coefficients for organic carbon, P, and N contents with chemical and physical attributes verified in the study areas.
| pH | Ca2+ | Mg2+ | Na+ | K+ | TS2 | CS3 | Silt | Clay | |
|---|---|---|---|---|---|---|---|---|---|
| SOC1 | -0.24 | -0.51 | -0.94 | -0.55 | -0.53 | 0.45 | 0.98 | -0.99* | 0.93* |
| P | 0.15 | -0.13 | -0.99* | -0.83* | -0.44 | 0.69 | 0.82 | -0.94* | 0.72* |
| N | -0.19 | -0.46 | -0.96 | -0.59 | -0.11 | 0.40 | 0.96 | -0.99* | 0.91* |
1SOC: Organic Carbon; 2TS: Thin Sand; 3CS: Coarse Sand. *Significant at p < 0.05.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



