
Submitted:
26 June 2024
Posted:
26 June 2024
You are already at the latest version
Abstract
The present study aimed to elucidate the neural mechanisms underpinning the visual recognition of morphologically complex verbs in Korean, a morphologically rich, agglutinative language with inherent polymorphemic characteristics. In an fMRI experiment with a lexical decision paradigm, we investigated whether verb inflection types (base, regular, and irregular) are processed through separate mechanisms or a single system. Furthermore, we explored the semantic influence in processing inflectional morphology by manipulating semantic ambiguity (homonymous vs. unambiguous) of inflected verbs. The results showed equivalent activation levels in the left inferior frontal gyrus for both regular and irregular verbs, challenging the dichotomy. Graded effects of verb regularity were observed in the occipitotemporal regions, with regular inflections eliciting increased activation in the fusiform and lingual gyri. In the middle occipital gyrus, homonyms showed decreased activation relative to unambiguous words, specifically for base and irregular forms. Furthermore, the angular gyrus exhibited significant modulation by all verb types, indicating a semantic influence during morphological processing. These findings support single-system theories and the connectionist framework, challenging the assumptions of purely orthographic morphological decomposition and dual-mechanism accounts. Furthermore, they provided evidence for a semantic influence during morphological processing, with differential reliance on semantic activation for regular and irregular inflections.
Keywords:
visual word recognition
; inflectional morphology
; morphologically rich language
; fMRI
1. Introduction
The issue of how various types of inflection, such as regular (e.g., work–worked) and irregular (e.g., catch–caught) forms, are represented and processed in the brain remains a central topic in psycholinguistics. Classical full-listing models argue that all morphologically complex words are stored in their entirety, regardless of regularity [1]. On the other hand, full-parsing models propose that these forms undergo an obligatory decomposition into their morphemic units, with only morphemes being stored [2,3]. Bridging these two opposing views, dual-mechanism accounts argue that there are categorical differences between regular and irregular forms, suggesting they are processed by distinct mechanisms [4,5,6]. Regular forms, which follow predictable patterns of inflection (e.g., adding ‘–ed’ to form the past tense), are processed through a rule-based system, while irregular forms are stored and processed in the mental lexicon through lexical-semantic memory. In contrast, single-system accounts challenge this dichotomy, suggesting continuous rather than categorical distinctions between regular and irregular inflections [7,8,9]. These theories, based on earlier connectionist models [10,11], advocate for a single associative system that maps orthographic or phonological forms to semantic meanings, leading to graded effects in inflectional morphology.
In the functional magnetic resonance imaging (fMRI) domain, recent studies have investigated brain regions involved in processing morphologically complex words. Initial fMRI research focused on identifying brain regions specifically tuned to morphological processing [12,13]. Devlin et al. [13] observed reduced activation in temporal and parietal regions, such as the bilateral angular gyrus (AG), left occipitotemporal cortex, and left middle temporal gyrus (MTG) for morphologically related word pairs compared to unrelated pairs, suggesting that morphology results from the convergence of form and meaning. Similarly, in an fMRI study using a masked priming paradigm, Gold and Rastle [14] reported the involvement of occipital regions, including the fusiform gyrus (FG) and middle occipital gyrus (MOG), in both morphological and orthographic relationships, with semantic conditions reducing activation in the middle temporal gyrus (MTG), indicating a structural nature of early morphological decomposition. The debate continues about whether morphologically complex words are processed as whole units or decomposed into morphemes. Some neuroimaging evidence supports whole-word processing [13,15], while other evidence favors morpheme decomposition [16]. Whole-word processing highlights lexical-semantic effects in widespread bilateral frontotemporal regions, including the middle temporal gyrus (MTG), superior temporal gyrus (STG), and inferior frontal gyrus pars orbitalis (IFG, BA 47) [15,17]. Conversely, decomposition emphasizes the role of the posterior left inferior frontal gyrus (LIFG, BA 44/45) [16].
The LIFG has been consistently implicated in the literature as a core region for processing inflectional morphology, particularly the English past tense, demonstrating distinct activation patterns for regular and irregular inflections. Both types show increased activity in temporal and hippocampal regions, including the fusiform gyrus (FG), middle temporal gyrus (MTG), and parahippocampal gyrus (PHG). Regular verbs, however, exhibit greater activation in the LIFG, along with additional regions such as the middle frontal gyrus (MFG), basal ganglia, and cerebellum [18,19,20,21]. This binary distinction aligns with the dual-route theory, positing two memory systems: the procedural memory system for regular inflections and the declarative memory system for irregular ones [6,22,23]. Neuroimaging studies corroborate this theory, showing selective activation for regular inflections in the procedural memory network, characterized by the left-lateralized frontotemporal network, including the LIFG, basal ganglia, and cerebellum [24,25,26].
However, some researchers provide evidence for a single-system theory, which suggests continuous differences in inflection processing [7,27]. Joanisse and Seidenberg [7] found greater activation for regular compared to irregular verbs in the bilateral IFG during a past-tense generation task. They observed that phonologically similar irregulars elicited similar activation to regulars, while irregulars with no phonological similarity showed increased activity. Desai et al. [27] also reported no additional activation for regular inflected verbs compared to irregulars when phonological complexity was matched. These findings suggest that inflectional morphology is influenced by phonological, semantic, and probabilistic factors rather than a binary rule-based system.
Albeit the heavy focus on English past tense, some neuroimaging studies have also investigated the neural correlates of inflectional morphology employing morphologically rich languages like Finnish, Japanese, and Korean. Finnish studies showed that inflected nouns elicited an increased activation in the left inferior frontal gyrus (LIFG, BA 47), superior temporal gyrus (STG, BA 22), middle temporal gyrus (MTG, BA 21), and angular gyrus (AG, BA 39), indicating that morphological processing involves suffix stripping at the semantic-syntactic level rather than the visual word form level [28,29]. Japanese studies indicated greater activation in the left MTG (BA 21) for verbs compared to nouns, with selective activation for inflected verbs in the LIFG (BA 44/45) [30]. Yim et al. [31], the only fMRI study on Korean inflected verbs to the best of our knowledge, found similar activation patterns for regular and irregular verbs in temporal regions, including the left middle temporal gyrus (MTG), medial frontal gyrus (MFG), superior temporal gyrus (STG), and parahippocampal gyrus PHG), suggesting that Korean relies on memory and meaning irrespective of verb regularity, supporting the single-system theory.
In sum, previous fMRI literature has highlighted the distinct neural correlates involved in processing morphologically complex words, identifying widespread activation across occipital, temporal, and frontal regions [13,14,15,16,25,32]. Notably, the left frontotemporal regions, particularly the left inferior frontal gyrus (LIFG), have been proposed as the core network (procedural memory network) for the rule-based computation of regular verbs. In contrast, temporal-hippocampal regions are implicated as the declarative memory network for processing irregular verbs, which relies on the retrieval of semantic information, similar to whole-word processing [20,21,24]. However, some researchers have reported no significant difference between regular and irregular verbs after controlling for phonological complexity [7,27] or found equivalent activation for both inflection types in the temporal regions [31]. These findings suggest that verb regularity effects may be gradually modulated by the convergence of orthographic, semantic, and phonological information rather than by categorical differences among inflection types.
With respect to semantic processing, the LIFG has been also highlighted for its role of top-down control over activating and selecting meanings of homonyms [33,34,35]. Other relevant regions include the middle temporal gyrus (MTG), angular gyrus (AG), and supramarginal gyrus (SMG), associated with accessing and encoding lexical-semantic representations [34,36,37,38,39,40]. For instance, Hoffman and Tamm [38] found that the inferior frontal gyrus (IFG) and posterior middle temporal gyrus (pMTG) play roles in semantic control and representation, respectively. Nevertheless, it remains uncertain whether the semantic information engages in the processing of inflected verbs, and if so, whether it is processed in a distinct mechanism depending on the regularity.
In the present study using rapid event-related fMRI design with the lexical decision task (LDT), we aimed to investigate if different inflection types in Korean verbs show distinct brain activation patterns and how semantic information affects morphological processing. In this regard, we manipulated three inflection types including the uninflected base form as well as regularly and irregularly inflected verbs to examine whether inflected forms incur a processing cost and whether base and inflected forms exhibit graded effects or distinct differences [41,42]. Furthermore, by orthogonally manipulating the semantic ambiguity of verbs and comparing homonymous verbs with those possessing a single meaning within each type of inflection, we sought to elucidate the underlying processes involved in morphological and semantic processing. In Korean, morphologically complex words convey semantic and syntactic information at the single-word level. Moreover, Korean verbs are inherently polymorphemic, composed of a stem and various suffixes. These unique characteristics make the Korean writing system suitable for investigation of the complex interplay of form and meaning during the processing of inflectional morphology.
We hypothesized that memory-based retrieval of inflected forms would activate widespread bilateral frontotemporal regions, including the middle temporal gyrus (MTG), superior temporal gyrus (STG), and anterior inferior frontal gyrus (IFG, BA 47). In contrast, rule-based decomposition was expected to activate the posterior left inferior frontal gyrus (LIFG, BA 44/45). If regular and irregular inflections showed categorical differences, regular inflections would increase left frontotemporal activation, especially in the posterior LIFG. If graded effects were assumed, both inflections would increase activation in the left frontotemporal network, with regular verbs showing the most pronounced activation. Semantic effects were expected to modulate activation in the inferior parietal lobe (IPL), including the angular gyrus (AG) and supramarginal gyrus (SMG).
2. Materials and Methods
2.1. Participants
Twenty-four healthy native Korean speakers participated in the experiment (12 female; 23.8 ± 2.5 years, M ± SD). All had normal or corrected-to-normal vision and were rated as right-handed by the Edinburgh Handedness Inventory [43]. They signed and were informed with written consent and compensated with payment for their participation. Two participants’ data were excluded from the further analyses due to their poor behavioral performance, showing accuracy below 70%. The remaining twenty-two participants (11 female) had a mean age of 23.77 ± 2.62 years.
2.2. Materials
The experimental stimuli set comprised 240 items, including 120 Korean words and 120 pseudowords, selected from Korean Sejong Corpus [44] with a 15-million size. All word stimuli were verbs with 2–3 syllables, as confirmed by the Standard Korean dictionary. Only pure Korean words were utilized to avoid potential confounding effects of Sino-Korean words (i.e., Chinese-derived words; see [45]).
The words were categorized into three inflection types, including uninflected base form (n = 40), regularly inflected form (n = 40), and irregularly inflected form (n = 40). As suggested in previous literature on Korean inflectional morphology [46,47], base forms consisted of a verb stem and a basic ending suffix (–다), as in 씻다 (to wash) with the stem 씻– and suffix -다. Regular inflections were formed by the simple addition of a suffix to the verb stem (e.g., 먹어 – to eat and, with the stem 먹 and the suffix 어), while irregular inflections involved a transformation of the stem when suffixed as in 도와 (to help and) with the stem 돕– transformed to 도와 when suffixed with –아.
Semantic ambiguity was manipulated by comparing verbs with single meanings (i.e., control; n = 60) and those with multiple unrelated meanings (i.e., homonyms; n = 60). The classification was based on objective measures from the given corpus and the Standard Korean dictionary. Subjective measures were also utilized, in which three sets of subjective ratings were conducted on the experimental stimuli: familiarity, the number of meanings (NOM), and the related of meanings (ROM). Twenty volunteers (all native Korean speakers, 13 female; mean age 23.1 years, SD = 3.7). For familiarity rating, participants rated the subjective familiarity and frequency of use on a 7-point Likert scale.
The subjective ratings of the NOM and ROM were carried out following the procedure of Azuma [48]. For the NOM Rating, they indicated whether a presented stimulus had no meaning (0), a single meaning (1), or multiple meanings (2). Stimuli were presented in their base form (e.g., stem suffixed with the basic ending suffix ‘-다’), including 60 control stems, 60 homonymous stems, and 60 pseudostems. The base form of pseudostems was created by randomly selecting syllables from real verbs. NOM ratings showed that homonymous stems had significantly more meanings compared to control stems (homonym: M = 1.43, SD = 0.43; control: M = 1.18, SD = 0.17; t(59) = -6.40, p < .001). In the ROM rating, participants judged the degree of relatedness of meanings on a 7-point Likert scale, ranging from 1 (unrelated meanings) to 7 (same meaning). A base form of the stem and exemplar sentences of its meaning pair were presented. For stems with more than two meanings, the most frequently used meaning was paired with another meaning in multiple trials, and the rating score was averaged over all pairwise comparisons. The ROM rating set included 60 homonymous stems and 10 polysemous stems as a filler condition. ROM ratings confirmed that homonymous stems had relatively unrelated meanings (M = 1.89, SD = 1.50), compared to the highly related meanings of polysemous stems (M = 5.35, SD = 1.62).
Moreover, the meaning dominance of homonyms was also assessed through additional subjective ratings. Here, twenty native Korean speakers (16 female; mean age 27.7 years, SD = 4.59) who did not participate in the main experiment or the three sets of subjective ratings, were asked to judge the subjective familiarity of each meaning of the homonyms on a 7-point Likert scale. The results demonstrated no significant difference among conditions in the meaning dominance, showing the mean familiarity ratings for dominant and subordinate meanings: base form homonymy, 6.12 (SD = 0.53) for dominant meanings and 4.54 (SD = 1.05) for subordinate meanings; regular form homonymy, 5.95 (SD = 0.52) for dominant meanings and 4.25 (SD = 1.07) for subordinate meanings; irregular form homonymy, 5.98 (SD = 0.46) for dominant meanings and 4.11 (SD = 0.94) for subordinate meanings. Other sublexical and lexical variables known to affect behavioral and brain responses during visual word recognition, including word frequency [49], stem length [50], neighborhood density (i.e., type frequency), and cumulative frequency (i.e., token frequency) of a first syllable [51] were also statistically matched between experimental conditions (all ps > .05, see Supplementary Table S1).
Finally, 120 pronounceable pseudowords were created for the filler word condition by randomly selecting and concatenating verb stems and suffixes used in the word stimuli set. The length of stimuli of each condition, including pseudowords, was matched so that half of the stimuli had 2 syllables, and the rest half had 3 syllables.
2.3. Procedure
A rapid event-related fMRI design was employed for the lexical decision task (Figure 1). Each trial commenced with a fixation point (+) displayed for 100 ms, followed by the presentation of a target stimulus for 1000 ms. Participants responded using their index and middle fingers to indicate whether the stimulus was a word or nonword, respectively. The responding hand was counterbalanced across participants: half (N=12) used their right hand, while the other half (N=12) used their left hand. Following each trial, a blank screen was presented during the inter-trial interval (ITI), which varied randomly in duration from 1000 ms to 7000 ms.
All stimuli were presented in white font (Courier New, size 28) on a black background, arranged in a pseudo-random order. The duration of the jittered intervals and the sequence of target stimuli were optimized using optseq2 software [52], and the presentation was controlled with the E-prime 2.0 Professional program (Psychology Software Tools, Inc.). Prior to the experimental session, the participants completed 30 practice trials. These included 5 baseline masks, 10 words, and 10 pseudowords not part of the experiment session. All participants achieved an accuracy rate above 80% in the practice session, ensuring their understanding of the procedure.
2.4. Image Acquisition
MRI scanning was performed using a Siemens Magnetom Trio 3T MRI scanner (Erlangen, Germany) in the Korea University Brain Imaging Center. Functional images were acquired using T2*-weighted gradient EPI (Echo Planar Imaging) sequences (TR = 2000 ms; TE = 20 ms; Flip Angle = 90º; Field of View = 240 mm; slice thickness = 3mm; no gap 42 slices; matrix size = 80 × 80; voxel size = 3 mm × 3 mm × 3 mm). T1-weighted structural images were acquired with a 3D MP-RAGE (Magnetization-Prepared Rapid Gradient-Echo) sequence (TR = 1900 ms; TE = 2.52 ms; Flip Angle = 90º; Field of View = 256 ms; matrix size = 256 × 256; voxel size = 1 mm × 1 mm × 1 mm) covering the whole head.
2.5. Behavioral Analysis
A two-way repeated measures analysis of variance (ANOVA) with the factors Ambiguity (2: control vs. homonym) and Morphology (3: base vs. regular vs. irregular) was conducted for response time (RT) and error rate in the LDT. Here, Greenhouse-Geisser correction was used when the presumption of sphericity was violated (p < .05) according to Mauchly’s Test of Sphericity. Only correct responses were used for the RT analysis. A post-hoc paired t-test with Bonferroni correction for multiple comparisons was carried out when any significant effect was found. R software [53] was used for the statistical analyses of behavioral and fMRI data.
2.6. fMRI Analysis
The fMRI data were processed and analyzed using the SPM12 toolbox (Wellcome Trust Centre for Neuroimaging, London, UK; http://www.fil.ion.ucl.ac.uk/spm/software/spm12/) alongside custom-built MATLAB scripts [54]. The first three images from each session were discarded to mitigate the transition effects of hemodynamic responses. The remaining functional images underwent realignment for motion correction, followed by slice timing correction to adjust for acquisition time differences between slices. The structural images were then coregistered to the mean functional images and segmented. Subsequently, the functional images were spatially normalized to a standard MNI (Montreal Neurological Institute) template, using the parameters obtained during segmentation. To reduce spatial noise, all images were smoothed with an isotropic Gaussian kernel of 6 mm full-width half-maximum (FWHM).
2.6.1. Statistical Analysis
Statistical analyses were conducted using a general linear model (GLM) with a two-stage mixed-effect approach. At the individual level, contrast images between the experimental conditions and baseline masks were generated, resulting in six contrasts: Base Control > Mask, Base Homonym > Mask, Regular Control > Mask, Regular Homonym > Mask, Irregular Control > Mask, Irregular Homonym > Mask. BOLD signals were convolved with a standard hemodynamic response function (HRF). Only correct responses in the lexical decision task were included in the analysis. Movement parameters obtained during realignment were entered as regressors in the model specification. Additionally, response time (RT) for each condition and participant was included as a parametric modulator, following the method suggested by Taylor et al. [54]. At the group level, one-sample t-tests were conducted on the contrast images estimated at the individual level. A whole-brain statistical parametric map was constructed for each Morphology and Semantic Ambiguity condition. The voxel-level statistical threshold was set at p < .001, and the cluster-level threshold was set at q < .05 with False Discovery Rate (FDR) correction, requiring a minimum cluster extent of 30 contiguous voxels (kE > 30).
To further investigate the activation patterns observed in the GLM results, follow-up analyses of variance (ANOVA) were performed with the factors Morphology (3: base vs. regular vs. irregular) and Semantic Ambiguity (2: control vs. homonym) using beta estimates extracted from significant clusters. Post-hoc pairwise t-tests with Bonferroni correction for multiple comparisons were carried out if significant effects were found.
2.6.2. ROI Analysis
Region of interest (ROI) analysis focused on brain regions previously associated with morphological and semantic processing [19,56]. ROIs for morphological processing included the left inferior frontal gyrus (LIFG), specifically the pars triangularis (BA 44/45, [–46, 30, 14]) and pars orbitalis (BA 47, [–37, 31, –12]). Semantic processing regions included the bilateral angular gyrus (AG, BA 39; left AG: [–45, –61, 36]; right AG: [44, –59, 39]) and the middle temporal gyrus (MTG, BA 40; left MTG: [–56, –34, –2]; right MTG: [56, –37, –2]). All ROIs were defined as spheres with a 5-mm radius centered at the corresponding MNI coordinates, selected from the Automatic Anatomical Labeling Atlas 3 (AAL3) [57].
Raw average ROI parameter estimates for each of the six experimental conditions contrasted to baseline masks (e.g., Base Control > Mask) were extracted using the MarsBaR toolbox [58]. Separate 2 × 3 repeated measures ANOVAs were conducted for each ROI with the factors Morphology (Base vs. Regular vs. Irregular) and Ambiguity (Control vs. Homonym), followed by post-hoc pairwise t-tests. Additionally, one-sample t-tests were conducted to assess (de)activation of each inflection type relative to the non-linguistic baseline mask (e.g., Regular > Mask).
3. Results
3.1. Behavioral Results
Behavioral performance on the lexical decision task is displayed in Table 1. Trials with reaction times exceeding 3 SDs (standard deviations) from the mean or error rates above 30% were excluded from the analysis. This accounted for 9.17% of the responses to word targets. Only correct responses were included in the RT analysis. The overall accuracy rate was 90.94 ± 28.72%, and the RT was 580 ± 117 ms on average.
A repeated measures ANOVA conducted on the reaction time (RT) data revealed a significant main effect of Morphology [F(2, 42) = 21.05, p < .001, η² = .06]. Specifically, the base form elicited faster responses compared to both regularly and irregularly inflected forms. Post-hoc pairwise t-tests confirmed that responses were significantly faster for the base form compared to the regular form [t(43) = –5.54, p < .001] and the irregular form [t(44) = –6.05, p < .001]. There was no significant difference in RTs between the regular and irregular forms [t(43) = 0.44, p > .1].
Additionally, a significant interaction between Morphology and Ambiguity was found [F(2, 42) = 4.13, p = .023, η² = .01]. Subsequent post-hoc pairwise t-tests revealed a significant ambiguity advantage for the base form, where homonyms elicited faster latencies compared to control words [t(22) = –2.24, p = .04]. Conversely, a marginally significant ambiguity disadvantage was observed in the regular form, with homonyms resulting in slower latencies compared to control words [t(22) = 1.87, p = .076].
An analysis of error rates also demonstrated a significant main effect of Morphology [F(2, 42) = 12.9, p < .001, η² = .09], consistent with the RT analysis. Participants responded more accurately for the base form compared to the regular form [t(44) = 4.24, p < .001]. There was a trend-toward-significance in the comparison between the base and irregular forms, with the base form showing a higher accuracy [t(44) = 2.39, p = .063]. No further significant effects were observed in the error rate analysis.
3.2. GLM Results
The results of the two-staged mixed effect general linear model (GLM) analysis on whole-brain activation are detailed in Table 2 and Table 3 and illustrated in Figure 2 and Figure 3. A broad array of brain regions associated with language processing showed activation for each type of morphology and semantic ambiguity condition compared to the non-linguistic baseline mask condition. These regions included the precentral gyrus, thalamus, supplementary motor area, middle occipital gyrus, lingual gyrus, and fusiform gyrus.
Follow-up repeated-measures ANOVAs were conducted on significant clusters identified in the whole-brain analysis, focusing on occipitotemporal regions including the left fusiform gyrus (FG), lingual gyrus (LG), and middle occipital gyrus (MOG). The factors were Morphology (3 levels: base, regular, irregular) and Semantic Ambiguity (2 levels: control, homonym). When significant effects were detected, post-hoc pairwise t-tests with Bonferroni correction for multiple comparisons were performed, as illustrated in Figure 4.
The main effect of Morphology was significant in the FG [F(2, 42) = 10.74, p < .001, η² = .09] and MOG [F(2, 42) = 14.85, p < .001, η² = .09], and marginally significant in the LG [F(2, 42) = 3.11, p = .055, η² = .03]. Post-hoc pairwise t-tests revealed that the regular form elicited increased activation in the FG compared to the irregular form [t(44) = –4.26, p < .001]. There was also a marginally significant increase in activation for the regular form compared to the base form [t(44) = –2.47, p = .052]. In the MOG, there was a significant decrease in activation for irregular forms compared to both the base form [t(44) = 3.56, p = .003] and the regular form [t(44) = –4.58, p < .001]. The LG showed a marginally significant increase in activation for the regular form compared to the irregular form [t(44) = –2.34, p = .071].
The main effect of Semantic Ambiguity was significant in all regions examined. In the FG [F(1, 21) = 21.17, p < .001, η² = .07], LG [F(1, 21) = 10.99, p = .003, η² = .03], and MOG [F(1, 21) = 33.37, p < .001, η² = .13], homonyms showed reduced activation relative to control words. The differences were statistically significant with t(65) = 3.69, p < .001 for FG, t(65) = 3.00, p = .004 for LG, and t(65) = 5.19, p < .001 for MOG.
The interaction between Morphology and Semantic Ambiguity was significant in the LG [F(2, 42) = 6.04, p = .005, η² = .04], and marginally significant in the MOG [F(2, 42) = 2.47, p = .096, η² = .03]. An ambiguity advantage, characterized by increased activation for control words relative to homonyms, was observed in the MOG for both the base form [t(21) = 3.53, p = .002] and the irregular form [t(21) = 5.42, p < .001]. In the LG, a significant increase in activation for control words compared to homonyms was found for the base form [t(21) = 3.81, p = .001]. However, no significant differences between control and homonyms were found in the regular condition for any region (all ps > .1).
3.3. ROI Results
Region of interest (ROI) analyses were conducted on brain regions implicated in previous fMRI literature [19,56], including the left inferior frontal gyrus pars triangularis (LIFG tri) and pars orbitalis (LIFG orb), bilateral angular gyrus (AG), and middle temporal gyrus (MTG). Figure 5 illustrates the ROIs that showed a significant main effect of Morphology in the repeated measures ANOVAs, which included factors Morphology and Ambiguity.
Significant main effects of Morphology were observed in the LIFG tri [F(2, 42) = 5.52, p = .007, η² = .031] and bilateral AG [left: F(2, 46) = 3.83, p = .03, η² = .03; right: F(2, 46) = 4.17, p = .02, η² = .031]. Post-hoc paired t-tests indicated a significantly increased activation for regular verbs compared to base forms in the LIFG tri [t(43) = –2.95, p = .016]. Conversely, the regular form showed decreased activation relative to the base form in the bilateral AG [left: t(43) = 2.91, p = .006; right: t(43) = 2.58, p = .04].
One-sample t-tests were performed to test for activation of each inflection type relative to the baseline mask. The results revealed that processing inflected forms (regular and irregular) selectively increased activation in the LIFG tri [regular: t(43) = 3.52, p = .001; irregular: t(43) = 2.09, p = .04], with regular forms also showing increased activation in the LIFG orb [t(43) = 2.45, p = .02]. In contrast, processing verbs, regardless of inflection type, was associated with increased deactivation in the left AG [base: t(43) = –2.12, p = .04; regular: t(43) = –4.89, p < .001; irregular: t(43) = –2.55, p = .014] and right AG [base: t(43) = –2.05, p = .046; regular: t(43) = –5.12, p < .001; irregular: t(43) = –3.56, p = .001]. A similar pattern of deactivation was also found in the right MTG for all types of verbs [base: t(43) = –2.98, p = .004; regular: t(43) = –2.29, p = .027; irregular: t(43) = –3.07, p = .003].
As illustrated in Figure 6, repeated measures ANOVAs revealed a significant main effect of semantic ambiguity in the right middle temporal gyrus (MTG) [F(1, 21) = 5.84, p = .025, η² = .014]. Specifically, homonyms showed a marginally significant increase in deactivation relative to control words [t(65) = 1.91, p = .061]. No other regions exhibited significant differences in response to semantic ambiguity. Further analysis using one-sample t-tests indicated that homonyms were significantly deactivated compared to the baseline mask in the bilateral angular gyrus (AG) [left: t(65) = –3.49, p < .001; right: t(65) = –4.09, p < .001] and the right MTG [t(65) = –4.25, p < .001]. Similarly, control words also showed significant deactivation in the same regions [left AG: t(65) = –4.16, p < .001; right AG: t(65) = –4.5, p < .001; right MTG: t(65) = –2.42, p = .018]. Importantly, the left inferior frontal gyrus (LIFG) displayed an opposite pattern of activation. Both homonymous and control words elicited increased activation in the LIFG pars triangularis (LIFG tri) [control: t(65) = 3.09, p = .002; homonym: t(65) = 2.11, p = .039].
4. Discussion
The present study aimed to investigate the spatial localization of morphological processing using a rapid event-related fMRI design combined with a lexical decision task. The experiment explored how morphological inflection (base, regular, and irregular) and semantic ambiguity (control vs. homonymous) influence both behavioral performance and brain activation patterns during the visual recognition of morphologically complex Korean verbs. It was hypothesized that verb regularity and inflectional cost would be reflected in differential activation in the left frontotemporal regions, particularly the left inferior frontal gyrus (LIFG), with regularly inflected verbs showing increased activity relative to base or irregular forms. An interaction between form and meaning was also anticipated, with regularly inflected homonyms expected to show increased activation relative to control words in temporal regions, while base and irregular forms were expected to elicit the reverse pattern.
The behavioral results revealed a significant inflectional cost on reaction times and accuracy, indicating that participants responded faster and more accurately to base forms compared to both regular and irregular inflected forms. Additionally, a significant interaction was observed between Morphology and Ambiguity. Base forms exhibited an ambiguity advantage, whereas regular forms displayed a trend towards an ambiguity disadvantage. These findings align with previous research on inflected words in morphologically rich languages, suggesting that uninflected forms are processed more efficiently due to their direct access to semantic representations, while inflected forms require additional processing effort, leading to increased processing time and error rates [28,59]. Furthermore, the significant interaction between form and meaning suggests that the processing of homonymous words can be facilitated or impeded depending on their morphological form, potentially due to the interplay between morphological parsing and access to semantic representations [60].
The whole-brain fMRI results revealed activations predominantly in occipitotemporal regions associated with morphology and semantics, including the fusiform gyrus (FG), lingual gyrus (LG), and middle occipital gyrus (MOG). Significant modulation of activation by inflection type was observed, with regular forms showing increased activation in the FG and LG compared to base and irregular forms. In contrast, irregular forms exhibited significantly reduced activation in the MOG relative to base and regular forms. These findings are consistent with previous studies implicating the occipitotemporal cortex in visual word form processing and morphological analysis [14,61].
The occipitotemporal cortex, especially the visual word form area (VWFA) located within the left FG, has been consistently associated with orthographic processing during visual recognition of words [14,62,63,64], and thus often suggested as evidence for a morphological decomposition driven by orthography in a blind-to semantic manner. For instance, in a masked priming fMRI experiment, Gold and Rastle [14] found an overlapping activation in occipitotemporal regions, including the left fusiform and lingual gyri for pseudo-morphological (e.g., corner–CORN) and orthographic (e.g., brothel–BROTH) conditions, but not for the semantic (bucket-PAIL) condition, related and more importantly, a distinctive selective activation of lingual gyrus (BA 19) for morphologically word pairs compared to unrelated condition, suggesting early morphological decomposition is driven by orthography in a blind to semantics manner. Indeed, the current data demonstrated the contribution of the FG, LG, and MOG to morphological effects, however with regular forms showing increased activation compared to either base or irregular forms, challenging the notion of obligatory segmentation processes for all morphologically complex words [2,64].
Furthermore, semantic effects were observed in occipitotemporal regions, with homonyms showing decreased activation relative to unambiguous words. A form-with-meaning interaction was found, where decreased MOG activation for homonyms relative to unambiguous words was evident for base and irregular forms, but not for regular forms. This pattern is consistent with behavioral findings showing a reversal of the semantic ambiguity effect for base and regular forms. The FG’s role in lexical-semantic processing, as suggested by previous literature [61,66] fits well with the current data, which demonstrated both morphological and semantic effects, indicating the occipitotemporal regions’ role in interfacing form and meaning in morphologically complex Korean verbs.
ROI analyses, based on previous fMRI literature [19,56], further elucidated the neural substrates underlying the processing of different morphological and semantic properties of verbs. The analyses focused on the LIFG pars triangularis and pars orbitalis, bilateral AG, and MTG. A significant main effect of Morphology was observed in the LIFG tri and bilateral AG. Regular forms showed increased activation in the LIFG tri compared to base forms, while the regular form showed decreased activation relative to the base form in the bilateral AG. One-sample t-tests indicated that processing inflected forms (regular and irregular) selectively increased LIFG activation in the LIFG tri, with regular forms also showing increased activation in the LIFG orb. In contrast, all verb types, regardless of inflection, were associated with increased deactivation in the bilateral AG and right MTG.
In the dual-route model of inflectional morphology [6,22,23], the declarative memory network involving the temporal-hippocampal regions is expected to modulate both types of inflection. In contrast, rule-based processing of regularly inflected verbs is associated with distinctive activation in the procedural memory network, which includes the LIFG, cerebellum, and basal ganglia. According to this model, regular inflections should uniquely engage the LIFG and related structures, while both regular and irregular inflections should activate the temporal-hippocampal network. We indeed observed a significant reduction in AG activation for all verb types compared to the baseline mask, with the regular inflections showing reduced activity relative to base forms. Given the functional association of AG and the inferior parietal lobule (IPL) with semantic processing [56,67], these results suggest that processing base form verbs benefited more from memory-based retrieval processes relative to regularly inflected verbs and that these processes are involved in processing Korean verbs, regardless of whether they are inflected, and if so, whether they are regularly or irregularly inflected.
Contrary to the expectation of the dual-route model, the posterior division of the LIFG (i.e., LIFG pars triangularis, BA 44) showed selective activation for both inflected forms but not for uninflected base forms. Importantly, regular and irregular verbs did not differ significantly in LIFG activation, with irregular verbs eliciting equivalent levels of activation, challenging the binary distinction of LIFG activation patterns for regular and irregular inflections. Furthermore, the current results contradict the notion that all morphologically complex words are processed as whole words, which would predict similar activation levels for regular and irregular inflections across widespread bilateral regions, including the IFG pars orbitalis, MTG, and occipitotemporal regions. Instead, the present findings support a single-system mechanism where continuous rather than categorical differences between regular and irregular inflections are expected. These results align with a more general function of the LIFG, which involves processing morphologically complex words, as suggested by previous fMRI literature on derivation and inflection [16,20,28,29].
Finally, semantic effects were reflected in the modulation of the temporal region, particularly the right MTG, along with the occipitotemporal regions observed in the whole-brain results. The functional contribution of temporal regions to semantic processing is well-documented [36,37]. Surprisingly, increased activation in these regions, interpreted as engaging greater neural resources to resolve semantic ambiguity in previous research [34], contradicts the behavioral findings of this study, where facilitative effects for homonyms were observed for base and irregular forms. However, a significant difference in right MTG activation between homonymous and unambiguous words suggests that facilitative effects for homonyms may be due to greater activation of abundant lexical-semantic representations [68,69]. Furthermore, both homonymous and unambiguous words elicited equivalent activation in the LIFG pars triangularis, suggesting that processing homonyms relies more on lexical-semantic representations than on the top-down regulation of multiple competitors.
5. Conclusions
The present study investigated the spatial dynamics underlying the visual recognition of morphologically complex Korean verbs using a rapid event-related fMRI design. The findings challenge the dual-route model of inflectional morphology [6,22,23], which posits distinct processing pathways for regular and irregular verbs. Contrary to this model, both regular and irregular verbs showed equivalent levels of activation in the LIFG, a core region in the proposed procedural memory network. Instead, these results support the general role of the LIFG in handling morphological processing demands, a role primarily demonstrated in studies of morphologically rich languages [28,59]. Importantly, graded effects for verb regularity were observed in the bilateral occipitotemporal regions, with regular inflections eliciting increased activation in the left FG compared to base and irregular forms. The AG, a region in the IPL consistently associated with semantic processing, differentiated between the three inflection types, with base forms showing the least deactivation, followed by regular and then irregular inflections. These results provide evidence for the single-system mechanism [7,27], which argues for a continuous effect of verb regularity, with orthographic, semantic, and phonological properties converging in the processing of words. Furthermore, the facilitative effect of semantic ambiguity was reflected in the selective deactivation of the MTG and MOG for homonyms, while the LIFG showed equivalent activation for both homonymous and unambiguous words. These findings suggest that processing Korean homonyms may benefit from facilitated access to lexical-semantic information rather than being inhibited by the processes of selection and regulation among multiple meanings.
The pattern of activation observed across widespread bilateral inferior frontal, temporal, and occipitotemporal regions for processing morphologically complex Korean verbs suggests that the integration of morphological and semantic information involves a complex network of brain regions. This supports the notion that multiple cortical areas, distributed over a vast network, engage in morphological processing and, more generally, in language processing. These results align with the view that posits a distributed function involving extensive neural networks involved in morphological processing [70], and more generally, in language processing [71,72].
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Table S1: Experimental conditions and statistical comparisons of sublexical and lexical variables.
Author Contributions
Conceptualization, J.K. and K.N.; Formal analysis, J.K.; Funding acquisition, J.K.; Investigation, J.K. and S.K.; Methodology, J.K. and K.N.; Resources, J.K. and S.K.; Supervision, K.N.; Validation, S.K. and K.N.; Visualization, J.K. and S.K.; Writing—original draft, J.K.; Writing—review & editing, S.K. and K.N. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Ministry of Education of the Republic of Korea and the National Research Foundation of Korea, grant number NRF-2019H1A2A1076382.
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki, and approved by the Institutional Review Board (or Ethics Committee) of Korea University (protocol code KUIRB-2021-0427-01, 2021.12.21.).
Informed Consent Statement
Informed consent was obtained from all participants involved in the study.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Butterworth, B. Lexical Representation. In Language Production Vol. 2.; Butterworth, B., Ed.; Academic Press: London, 1983; pp. 257–294. [Google Scholar]
- Taft, M. Morphological Decomposition and the Reverse Base Frequency Effect. Q. J. Exp. Psychol. Sect. A: Hum. Exp. Psychol. 2004, 57, 745–765. [Google Scholar] [CrossRef] [PubMed]
- Taft, M.; Forster, K.I. Lexical Storage and Retrieval of Prefixed Words. J Verbal Learn. Verbal Behav 1975, 14, 638–647. [Google Scholar] [CrossRef]
- Clahsen, H. Lexical Entries and Rules of Language: A Multidisciplinary Study of German Inflection. Behav. Brain Sci. 1999, 22, 991–1013. [Google Scholar] [CrossRef] [PubMed]
- Pinker, S. Rules of Language. Science (1979) 1991, 253, 530–535. [Google Scholar] [CrossRef] [PubMed]
- Ullman, M.T. Contributions of Memory Circuits to Language: The Declarative/Procedural Model. Cognition 2004, 92, 231–270. [Google Scholar] [CrossRef] [PubMed]
- Joanisse, M.F.; Seidenberg, M.S. Imaging the Past: Neural Activation in Frontal and Temporal Regions during Regular and Irregular Past-Tense Processing. Cogn Affect Behav Neurosci 2005, 5, 282–296. [Google Scholar] [CrossRef] [PubMed]
- Marslen-Wilson, W.; Tyler, L.K. Rules, Representations, and the English Past Tense. Trends Cogn Sci 1998, 2, 428–435. [Google Scholar] [CrossRef] [PubMed]
- McClelland, J.L.; Patterson, K. Rules or Connections in Past-Tense Inflections: What Does the Evidence Rule Out? Trends Cogn Sci 2002, 6, 465–472. [Google Scholar] [CrossRef] [PubMed]
- McClelland, J.L.; Rumelhart, D.E. On Learning the Past Tenses of English Verbs. In Parallel Distributed Processing: Explorations in the Microstructure of Cognition: Psychological and Biological Models; MIT Press: Cambridge, 1987; ISBN 9780262291262. [Google Scholar]
- Seidenberg, M.S.; McClelland, J.L. A Distributed, Developmental Model of Word Recognition and Naming. Psychol Rev 1989, 96, 523–568. [Google Scholar] [CrossRef] [PubMed]
- Davis, M.H.; Meunier, F.; Marslen-Wilson, W.D. Neural Responses to Morphological, Syntactic, and Semantic Properties of Single Words: An FMRI Study. Brain Lang 2004, 89, 439–449. [Google Scholar] [CrossRef]
- Devlin, J.T.; Jamison, H.L.; Matthews, P.M.; Gonnerman, L.M. Morphology and the Internal Structure of Words. Proc. Natl. Acad. Sci. 2004, 101, 14984–14988. [Google Scholar] [CrossRef] [PubMed]
- Gold, B.T.; Rastle, K. Neural Correlates of Morphological Decomposition during Visual Word Recognition. J Cogn Neurosci 2007, 19, 1983–1993. [Google Scholar] [CrossRef] [PubMed]
- Bozic, M.; Tyler, L.K.; Li, S.; Wingfield, C.; Marslen-Wilson, W.D. Neurobiological Systems for Lexical Representation and Analysis in English. 2013. [CrossRef]
- Bozic, M.; Marslen-Wilson, W.D.; Stamatakis, E.A.; Davis, M.H.; Tyler, L.K. Differentiating Morphology, Form, and Meaning: Neural Correlates of Morphological Complexity. J Cogn Neurosci 2007, 19, 1464–1475. [Google Scholar] [CrossRef] [PubMed]
- Vannest, J.; Newport, E.L.; Newman, A.J.; Bavelier, D. Interplay between Morphology and Frequency in Lexical Access: The Case of the Base Frequency Effect. Brain Res 2011, 1373, 144–159. [Google Scholar] [CrossRef] [PubMed]
- Bozic, M.; Marslen-Wilson, W. Neurocognitive Contexts for Morphological Complexity: Dissociating Inflection and Derivation. Linguist. Lang. Compass 2010, 4, 1063–1073. [Google Scholar] [CrossRef]
- Bulut, T. Neural Correlates of Morphological Processing: An Activation Likelihood Estimation Meta-Analysis. Cortex 2022, 151, 49–69. [Google Scholar] [CrossRef] [PubMed]
- Tyler, L.K.; Bright, P.; Fletcher, P.; Stamatakis, E.A. Neural Processing of Nouns and Verbs: The Role of Inflectional Morphology. Neuropsychologia 2004, 42, 512–523. [Google Scholar] [CrossRef]
- Marslen-Wilson, W.D.; Tyler, L.K. Morphology, Language and the Brain: The Decompositional Substrate for Language Comprehension. Philos. Trans. R. Soc. B: Biol. Sci. 2007, 362, 823–836. [Google Scholar] [CrossRef]
- Pinker, S.; Ullman, M.T. The Past and Future of the Past Tense. Trends Cogn Sci 2002, 6, 456–463. [Google Scholar] [CrossRef]
- Ullman, M.T. A Neurocognitive Perspective on Language: The Declarative/Procedural Model. Nat Rev Neurosci 2001, 2, 717–726. [Google Scholar] [CrossRef] [PubMed]
- Pliatsikas, C.; Johnstone, T.; Marinis, T. FMRI Evidence for the Involvement of the Procedural Memory System in Morphological Processing of a Second Language. PLoS One 2014, 9, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Vannest, J.; Polk, T.A.; Lewis, R.L. Dual-Route Processing of Complex Words: New FMRI Evidence from Derivational Suffixation. Cogn Affect Behav Neurosci 2005, 5, 67–76. [Google Scholar] [CrossRef] [PubMed]
- Beretta, A.; Campbell, C.; Carr, T.H.; Huang, J.; Schmitt, L.M.; Christianson, K.; Cao, Y. An ER-FMRI Investigation of Morphological Inflection in German Reveals That the Brain Makes a Distinction between Regular and Irregular Forms. Brain Lang 2003, 85, 67–92. [Google Scholar] [CrossRef] [PubMed]
- Desai, R.; Conant, L.L.; Waldron, E.; Binder, J.R. FMRI of Past Tense Processing: The Effects of Phonological Complexity and Task Difficulty. J Cogn Neurosci 2006, 18, 278–297. [Google Scholar] [CrossRef] [PubMed]
- Lehtonen, M.; Vorobyev, V.A.; Hugdahl, K.; Tuokkola, T.; Laine, M. Neural Correlates of Morphological Decomposition in a Morphologically Rich Language: An FMRI Study. Brain Lang 2006, 98, 182–193. [Google Scholar] [CrossRef] [PubMed]
- Lehtonen, M.; Vorobyev, V.; Soveri, A.; Hugdahl, K.; Tuokkola, T.; Laine, M. Language-Specific Activations in the Brain: Evidence from Inflectional Processing in Bilinguals. J Neurolinguistics 2009, 22, 495–513. [Google Scholar] [CrossRef]
- Yokoyama, S.; Miyamoto, T.; Riera, J.; Kim, J.; Akitsuki, Y.; Iwata, K.; Yoshimoto, K.; Horie, K.; Sato, S.; Kawashima, R. Cortical Mechanisms Involved in the Processing of Verbs: An FMRI Study. J Cogn Neurosci 2006, 18, 1304–1313. [Google Scholar] [CrossRef] [PubMed]
- Yim, H.; Park, C.; Lim, H.; Nam, K. Mental Representation and Processing Involved in Comprehending Korean Regular and Irregular Verb Eojeols: An FMRI and Reaction Time Study. In Proceedings of the Neural Information Processing: 13th International Conference, ICONIP 2006, Hong Kong, China, October 3-6, 2006. Proceedings, Part I 13; 2006; Vol. 4232, pp. 247–254.
- Russo, A.G.; Esposito, F.; Laudanna, A.; Mancuso, A.; Di Salle, F.; Elia, A.; De Martino, M. The Neural Substrate of Noun Morphological Inflection: A Rapid Event-Related FMRI Study in Italian. Neuropsychologia 2021, 151. [Google Scholar] [CrossRef] [PubMed]
- Grindrod, C.M.; Garnett, E.O.; Malyutina, S.; den Ouden, D.B. Effects of Representational Distance between Meanings on the Neural Correlates of Semantic Ambiguity. Brain Lang 2014, 139, 23–35. [Google Scholar] [CrossRef] [PubMed]
- Rodd, J.M.; Davis, M.H.; Johnsrude, I.S. The Neural Mechanisms of Speech Comprehension: FMRI Studies of Semantic Ambiguity. Cereb. Cortex 2005, 15, 1261–1269. [Google Scholar] [CrossRef] [PubMed]
- Vitello, S.; Rodd, J.M. Resolving Semantic Ambiguities in Sentences: Cognitive Processes and Brain Mechanisms. Lang Linguist Compass 2015, 9, 391–405. [Google Scholar] [CrossRef]
- Binder, J.R.; Desai, R.H. The Neurobiology of Semantic Memory. Trends Cogn Sci 2011, 15, 527–536. [Google Scholar] [CrossRef] [PubMed]
- Lambon Ralph, M.A.; Jefferies, E.; Patterson, K.; Rogers, T.T. The Neural and Computational Bases of Semantic Cognition. Nat Rev Neurosci 2017, 18, 42–55. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, P.; Tamm, A. Barking up the Right Tree: Univariate and Multivariate FMRI Analyses of Homonym Comprehension. Neuroimage 2020, 219, 117050. [Google Scholar] [CrossRef]
- Humphreys, G.F.; Lambon Ralph, M.A. Mapping Domain-Selective and Counterpointed Domain-General Higher Cognitive Functions in the Lateral Parietal Cortex: Evidence from FMRI Comparisons of Difficulty-Varying Semantic Versus Visuo-Spatial Tasks, and Functional Connectivity Analyses. Cereb. Cortex 2017, 27, 4199–4212. [Google Scholar] [CrossRef] [PubMed]
- Zempleni, M.Z.; Renken, R.; Hoeks, J.C.J.; Hoogduin, J.M.; Stowe, L.A. Semantic Ambiguity Processing in Sentence Context: Evidence from Event-Related FMRI. Neuroimage 2007, 34, 1270–1279. [Google Scholar] [CrossRef] [PubMed]
- Soveri, A.; Lehtonen, M.; Laine, M. Word Frequency and Morphological Processing in Finnish Revisited. Ment Lex 2007, 2, 359–385. [Google Scholar] [CrossRef]
- Vartiainen, J.; Aggujaro, S.; Lehtonen, M.; Hultén, A.; Laine, M.; Salmelin, R. Neural Dynamics of Reading Morphologically Complex Words. Neuroimage 2009, 47, 2064–2072. [Google Scholar] [CrossRef] [PubMed]
- Oldfield, R.C. The Assessment and Analysis of Handedness: The Edinburgh Inventory. Neuropsychologia 1971, 9, 97–113. [Google Scholar] [CrossRef] [PubMed]
- Kang, B.; Kim, H. Frequency of Korean Language: Based on 15 Million Size Eojeol Sejong Syntactic and Semantic Corpus; Handkook munhwasa: Seoul, 2009; ISBN 9788957266366. [Google Scholar]
- Pae, H.K.; Bae, S.; Yi, K. Lexical Properties Influencing Visual Word Recognition in Hangul. Read Writ 2020, 33, 2391–2412. [Google Scholar] [CrossRef]
- Lee, E.; Lee, S.; Kim, Y. The Mechanism for Processing Regular and Irregular Inflections of Korean Verbs -A Priming Study-. Korean Lang. Lit. 2018, 183, 67–124. [Google Scholar] [CrossRef]
- Park, H.; Nam, K.; Koo, M. The Lexical Access of Regular and Irregular Korean Verbs in the Mental Lexicon. Korean J. Cogn. Sci. 2012, 23, 1–23. [Google Scholar] [CrossRef]
- Azuma, T. Familiarity and Relatedness of Word Meanings: Ratings for 110 Homographs. Behav. Res. Methods Instrum. Comput. 1996, 28, 109–124. [Google Scholar] [CrossRef]
- Rubenstein, H.; Garfield, L.; Millikan, J.A. Homographic Entries in the Internal Lexicon. J Verbal Learn. Verbal Behav 1970, 9, 487–494. [Google Scholar] [CrossRef]
- Kim, J.; Kang, J.; Kim, J.; Nam, K. Temporal Dynamics of Form and Meaning in Morphologically Complex Word Processing: An ERP Study on Korean Inflected Verbs. J Neurolinguistics 2022, 64, 101098. [Google Scholar] [CrossRef]
- Conrad, M.; Carreiras, M.; Jacobs, A.M. Contrasting Effects of Token and Type Syllable Frequency in Lexical Decision. Lang Cogn Process 2008, 23, 296–326. [Google Scholar] [CrossRef]
- Dale, A.M. Optimal Experimental Design for Event-Related FMRI. Hum. Brain Mapp. 1999, 8, 109–114. [Google Scholar] [CrossRef]
- R Core Team R: A Language and Environment for Statistical Computing 2022.
- The MathWorks Inc. MATLAB Version: 23.2.0 (R2023b) 2023.
- Taylor, J.S.H.; Rastle, K.; Davis, M.H. Interpreting Response Time Effects in Functional Imaging Studies. Neuroimage 2014, 99, 419–433. [Google Scholar] [CrossRef] [PubMed]
- Vigneau, M.; Beaucousin, V.; Hervé, P.Y.; Duffau, H.; Crivello, F.; Houdé, O.; Mazoyer, B.; Tzourio-Mazoyer, N. Meta-Analyzing Left Hemisphere Language Areas: Phonology, Semantics, and Sentence Processing. Neuroimage 2006, 30, 1414–1432. [Google Scholar] [CrossRef] [PubMed]
- Rolls, E.T.; Huang, C.C.; Lin, C.P.; Feng, J.; Joliot, M. Automated Anatomical Labelling Atlas 3. Neuroimage 2020, 206, 116189. [Google Scholar] [CrossRef]
- Brett, M.; Anton, J.-L.; Valabregue, R.; Poline, J.-B. Region of Interest Analysis Using the MarsBar Toolbox for SPM 99. Neuroimage 2002, 16, S497. [Google Scholar]
- Leminen, A.; Leminen, M.; Lehtonen, M.; Nevalainen, P.; Ylinen, S.; Kimppa, L.; Sannemann, C.; Mäkelä, J.P.; Kujala, T. Spatiotemporal Dynamics of the Processing of Spoken Inflected and Derived Words: A Combined EEG and MEG Study. Front Hum Neurosci 2011, 5. [Google Scholar] [CrossRef] [PubMed]
- Marslen-Wilson, W.; Tyler, L.K.; Waksler, R.; Older, L. Morphology and Meaning in the English Mental Lexicon; 1994; Vol. 101.
- Devlin, J.T.; Jamison, H.L.; Gonnerman, L.M.; Matthews, P.M. The Role of the Posterior Fusiform Gyrus in Reading. J Cogn Neurosci 2006, 18, 911–922. [Google Scholar] [CrossRef] [PubMed]
- Cohen, L.; Dehaene, S.; Naccache, L.; Lehéricy, S.; Dehaene-Lambertz, G.; Hénaff, M.-A.; Michel, F. The Visual Word Form Area. Brain 2000, 123, 291–307. [Google Scholar] [CrossRef] [PubMed]
- Cohen, L.; Lehéricy, S.; Chochon, F.; Lemer, C.; Rivaud, S.; Dehaene, S. Language-specific Tuning of Visual Cortex? Functional Properties of the Visual Word Form Area. Brain 2002, 125, 1054–1069. [Google Scholar] [CrossRef] [PubMed]
- Dehaene, S.; Naccache, L.; Cohen, L.; Bihan, D.L.; Mangin, J.-F.; Poline, J.-B.; Rivière, D. Cerebral Mechanisms of Word Masking and Unconscious Repetition Priming. Nat Neurosci 2001, 4, 752–758. [Google Scholar] [CrossRef] [PubMed]
- Rastle, K.; Davis, M.H.; New, B. The Broth in My Brother’s Brothel: Morpho-Orthographic Segmentation in Visual Word Recognition. Psychon Bull Rev 2004, 11, 1090–1098. [Google Scholar] [CrossRef] [PubMed]
- Kronbichler, M.; Hutzler, F.; Wimmer, H.; Mair, A.; Staffen, W.; Ladurner, G. The Visual Word Form Area and the Frequency with Which Words Are Encountered: Evidence from a Parametric FMRI Study. Neuroimage 2004, 21, 946–953. [Google Scholar] [CrossRef] [PubMed]
- Kuhnke, P.; Chapman, C.A.; Cheung, V.K.M.; Turker, S.; Graessner, A.; Martin, S.; Williams, K.A.; Hartwigsen, G. The Role of the Angular Gyrus in Semantic Cognition: A Synthesis of Five Functional Neuroimaging Studies. Brain Struct Funct 2023, 228, 273–291. [Google Scholar] [CrossRef] [PubMed]
- Bedny, M.; McGill, M.; Thompson-Schill, S.L. Semantic Adaptation and Competition during Word Comprehension. Cereb. Cortex 2008, 18, 2574–2585. [Google Scholar] [CrossRef] [PubMed]
- Tyler, L.K.; Cheung, T.P.L.; Devereux, B.J.; Clarke, A. Syntactic Computations in the Language Network: Characterizing Dynamic Network Properties Using Representational Similarity Analysis. Front Psychol 2013, 4. [Google Scholar] [CrossRef] [PubMed]
- Cavalli, E.; Colé, P.; Badier, J.M.; Zielinski, C.; Chanoine, V.; Ziegler, J.C. Spatiotemporal Dynamics of Morphological Processing in Visual Word Recognition. J Cogn Neurosci 2016, 28, 1228–1242. [Google Scholar] [CrossRef] [PubMed]
- Friederici, A.D. The Cortical Language Circuit: From Auditory Perception to Sentence Comprehension. Trends Cogn Sci 2012, 16, 262–268. [Google Scholar] [CrossRef] [PubMed]
- Hagoort, P. On Broca, Brain, and Binding: A New Framework. Trends Cogn Sci 2005, 9, 416–423. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Rapid event-related fMRI paradigm used in Experiment 2.

Figure 2.
The results of whole-brain analysis. for each type of condition contrasted with the baseline mask condition. (a) The results of whole-brain analysis for each type of Morphology, where highlighted regions indicate significant effects for base (green), regular (red), and irregular (blue) conditions, and overlapping regions (yellow). (b) The results of whole-brain analysis for each type of Semantic ambiguity condition, where highlighted regions represent significant effects for control (green) and homonym (red) conditions. The yellow color indicates overlapping regions.
Figure 2.
The results of whole-brain analysis. for each type of condition contrasted with the baseline mask condition. (a) The results of whole-brain analysis for each type of Morphology, where highlighted regions indicate significant effects for base (green), regular (red), and irregular (blue) conditions, and overlapping regions (yellow). (b) The results of whole-brain analysis for each type of Semantic ambiguity condition, where highlighted regions represent significant effects for control (green) and homonym (red) conditions. The yellow color indicates overlapping regions.
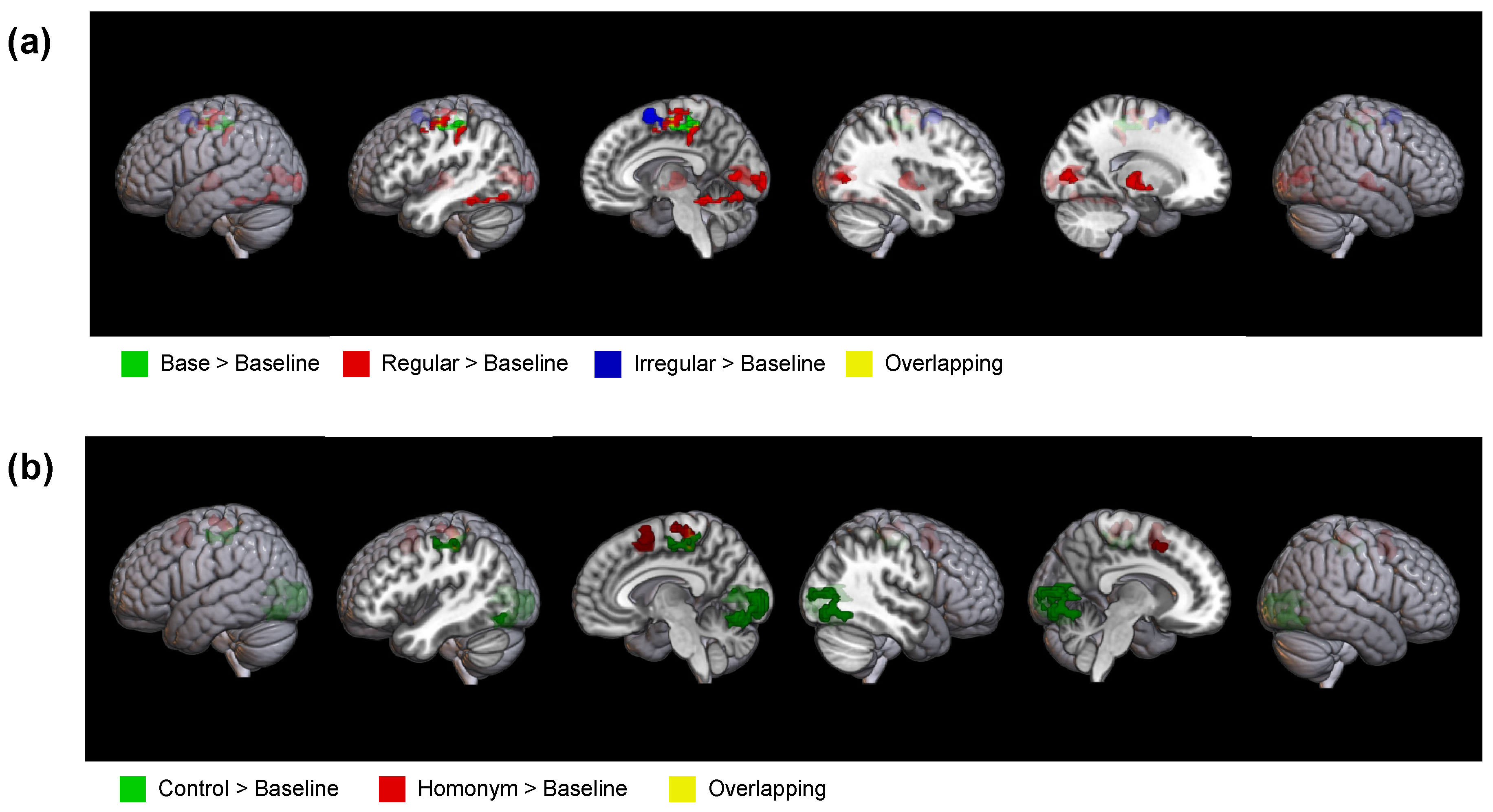
Figure 3.
(a) The whole brain activation map of the fusiform gyrus (FG), lingual gyrus (LG), and middle occipital gyrus (MOG). The highlighted regions indicate FG (red), LG (blue), and MOG (yellow). (b) Post-hoc analysis results on the main effect of Morphology. (c) Post-hoc analysis results on the main effect of semantic ambiguity. (d) Post-hoc analysis results on the two-way interaction. Note. *** p < .001, ** p < .01, * p < .05, † p < .1.
Figure 3.
(a) The whole brain activation map of the fusiform gyrus (FG), lingual gyrus (LG), and middle occipital gyrus (MOG). The highlighted regions indicate FG (red), LG (blue), and MOG (yellow). (b) Post-hoc analysis results on the main effect of Morphology. (c) Post-hoc analysis results on the main effect of semantic ambiguity. (d) Post-hoc analysis results on the two-way interaction. Note. *** p < .001, ** p < .01, * p < .05, † p < .1.
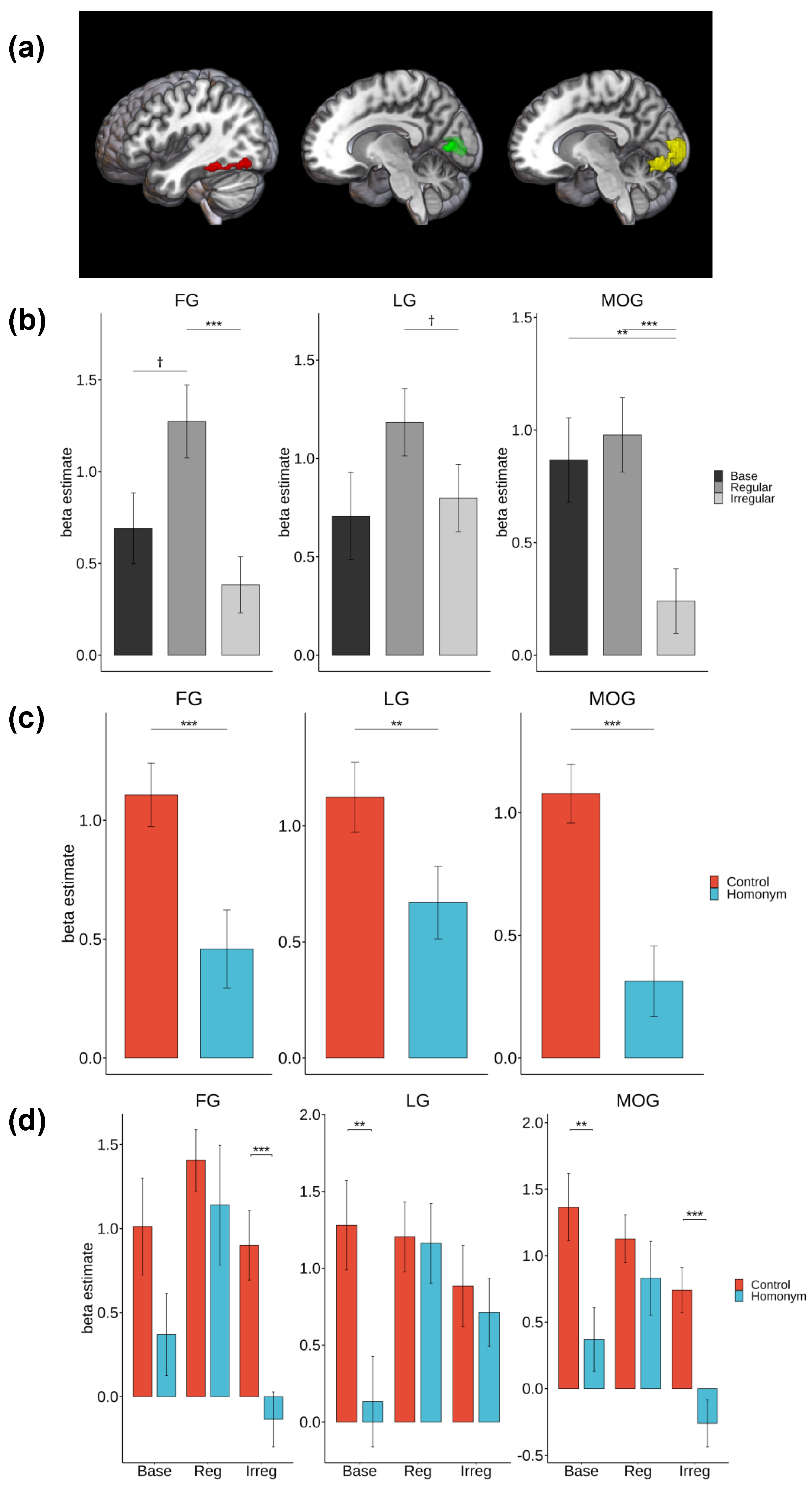
Figure 4.
ROI analysis results that showed a significant main effect of Morphology. (a) The regions highlighted in red, blue, green, and yellow represent LIFG pars triangluaris, LIFG pars orbitalis, AG, and MTG, respectively. (b) The beta estimates for each condition and statistical comparisons among inflection types. Note. *** p < .001, ** p < .01, * p < .05, † p < .1.
Figure 4.
ROI analysis results that showed a significant main effect of Morphology. (a) The regions highlighted in red, blue, green, and yellow represent LIFG pars triangluaris, LIFG pars orbitalis, AG, and MTG, respectively. (b) The beta estimates for each condition and statistical comparisons among inflection types. Note. *** p < .001, ** p < .01, * p < .05, † p < .1.
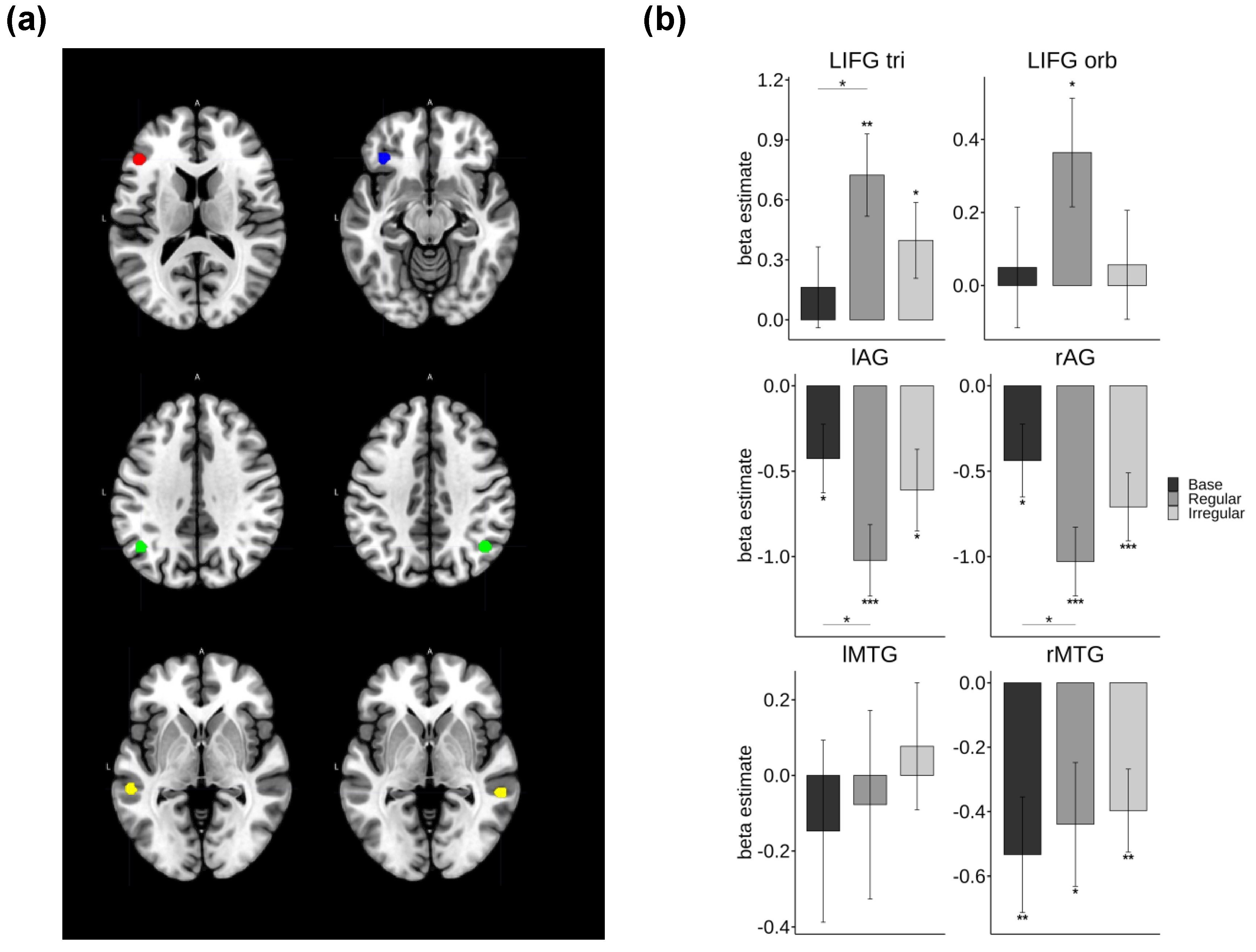
Figure 5.
ROI analysis results illustrating a significant main effect of Ambiguity. The left panel shows the corresponding regions highlighted in red (SMG), blue (left MOG), and yellow (right MTG). The right panel presents the beta estimates for each Ambiguity condition and statistical comparisons between control and homonymous words. Note. *** p < .001, ** p < .01, * p < .05, † p < .1.
Figure 5.
ROI analysis results illustrating a significant main effect of Ambiguity. The left panel shows the corresponding regions highlighted in red (SMG), blue (left MOG), and yellow (right MTG). The right panel presents the beta estimates for each Ambiguity condition and statistical comparisons between control and homonymous words. Note. *** p < .001, ** p < .01, * p < .05, † p < .1.
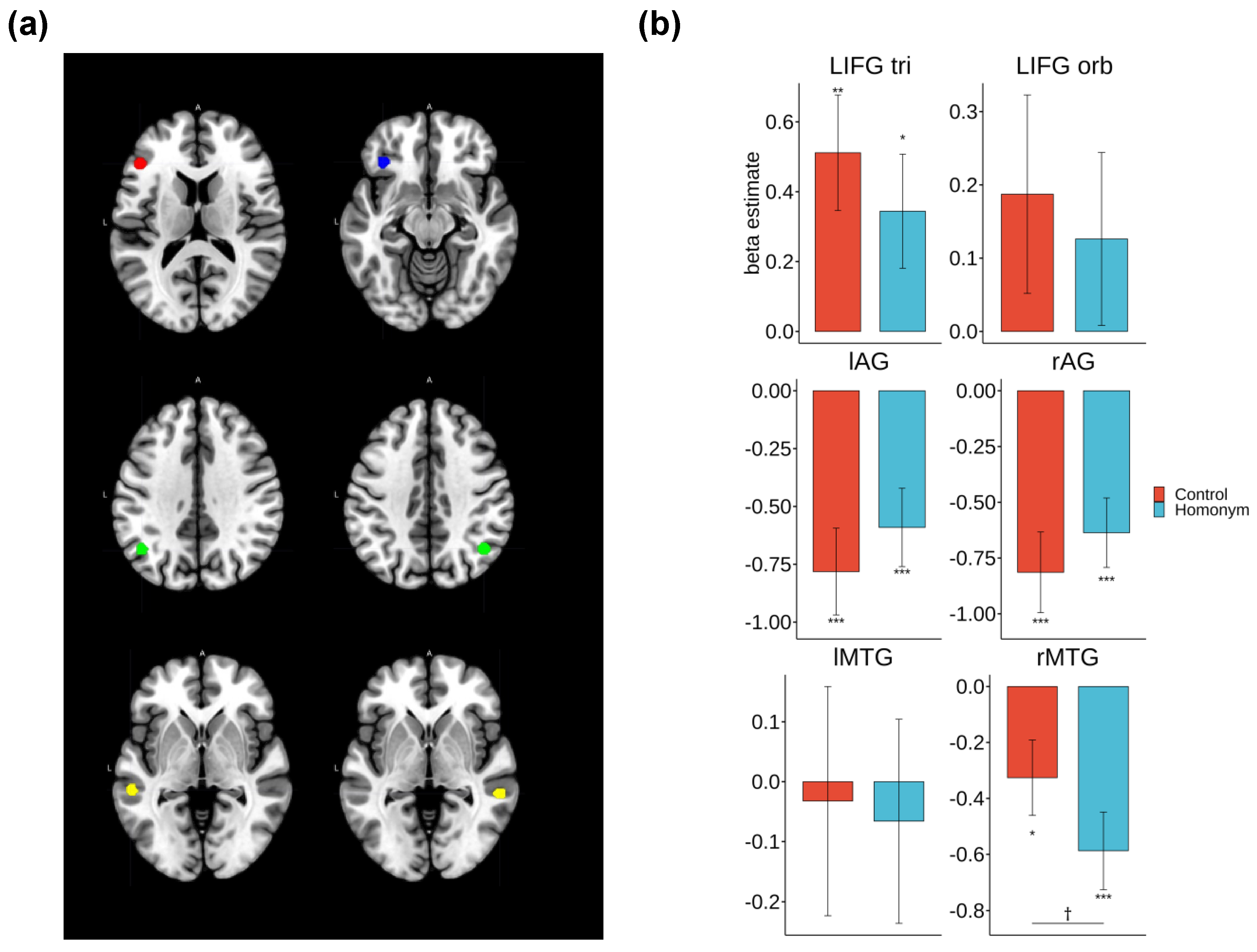

Table 1.
Mean reaction time (RT, in ms), percentage of accuracy (ACC, %), and standard deviations (SD, in parenthesis) for six conditions.
Table 1.
Mean reaction time (RT, in ms), percentage of accuracy (ACC, %), and standard deviations (SD, in parenthesis) for six conditions.
| Inflection type |
Semantic ambiguity | |||
| Control | Homonym | |||
| RT (ms) | ACC (%) | RT (ms) | ACC (%) | |
| Base | 568.66 (113.13) |
93.38 (24.89) |
554.9 (105.14) |
96.78 (17.67) |
| Regular | 583.21 (114.33) |
87.98 (32.56) |
601.97 (121.54) |
85.19 (35.58) |
| Irregular | 587.87 (121.57) |
90.85 (28.87) |
593.92 (121.29) |
90.12 (29.87) |
Note. Reaction times (RTs) for correct responses only.
Table 2.
Brain regions that showed significant effect for each type of morphological condition, relative to the baseline mask condition.
Table 2.
Brain regions that showed significant effect for each type of morphological condition, relative to the baseline mask condition.
| Contrast | Cluster size |
Brain regions | Hemisphere | MNI coordinates | Z | ||
| x | y | z | |||||
| Base > Mask | 89 | Precentral gyrus | L | –36 | –28 | 56 | 5.75 |
| Reg > Mask | 71 | Calcarine | L | –12 | –100 | –4 | 5.10 |
| 32 | Middle occipital gyrus | R | 36 | –76 | 2 | 3.40 | |
| 116 | Calcarine | R | 9 | –82 | 8 | 4.60 | |
| Lingual gyrus | L | 0 | –73 | 5 | 3.97 | ||
| Calcarine | R | 6 | –88 | 2 | 3.94 | ||
| 55 | Postcentral gyrus | L | –48 | –16 | 56 | 4.54 | |
| Precentral gyrus | L | –33 | –10 | 65 | 4.00 | ||
| –51 | –4 | 50 | 3.81 | ||||
| 98 | Thalamus | R | 21 | –22 | 2 | 4.23 | |
| Putamen | R | 30 | –4 | –7 | 3.87 | ||
| 65 | Fusiform gyrus | L | –42 | –40 | –19 | 4.16 | |
| –39 | –76 | –13 | 3.91 | ||||
| –42 | –55 | –19 | 3.55 | ||||
| 39 | Inferior parietal gyrus | L | –39 | –28 | 38 | 3.98 | |
| Postcentral gyrus | L | –42 | –34 | 50 | 3.85 | ||
| Irreg > Mask | 55 | Supplementary motor area | L | –3 | 8 | 65 | 3.64 |
| Middle cingulate cortex | L | –3 | –4 | 50 | 3.44 | ||
Note. All regions were significant to q < .05 FDR-corrected, kE ≥ 30 at the cluster level, and p < .001 at a voxel level. Reg = regular inflection; Irreg = irregular inflection.
Table 3.
Brain regions that showed significant effect for each type of semantic ambiguity condition, relative to the mask condition.
Table 3.
Brain regions that showed significant effect for each type of semantic ambiguity condition, relative to the mask condition.
| Contrast | Cluster size |
Brain regions | Hemisphere | MNI coordinates | Z | ||
| x | y | z | |||||
| Con > Mask | 285 | Calcarine | L | –12 | –100 | –4 | 5.18 |
| Fusiform gyrus | L | –36 | –79 | –16 | 4.67 | ||
| Middle occipital gyrus | L | –27 | –88 | 2 | 4.65 | ||
| 105 | Postcentral gyrus | L | –36 | –34 | 56 | 4.68 | |
| L | –42 | –16 | 50 | 3.53 | |||
| Precentral gyrus | L | –33 | –16 | 50 | 4.27 | ||
| 316 | Calcarine | R | 9 | –82 | 8 | 4.26 | |
| Lingual gyrus | R | 15 | –94 | –7 | 4.23 | ||
| Hom > Mask | 119 | Supplementary motor area | R | 9 | 14 | 47 | 4.55 |
| L | –6 | 5 | 56 | 4.42 | |||
| 57 | Postcentral gyrus | L | –36 | –34 | 56 | 4.10 | |
| Precentral gyrus | L | –24 | –19 | 68 | 3.46 | ||
| –33 | –22 | 65 | 3.46 | ||||
Note. All regions were significant to q < .05 FDR-corrected, kE ≥ 30 at the cluster level, and p < .001 at a voxel level.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



