
Submitted:
26 June 2024
Posted:
27 June 2024
You are already at the latest version
Abstract
Bile acids (BAs) play a crucial role in the human body's defense against infections caused by bacteria, fungi, and viruses. BAs counteract infections not only through interactions with intestinal bacteria exhibiting bile salt hydrolases (BSHs) activity but also directly combat infections. Building upon our research group's previous discoveries highlighting the role of BAs in combating infections, we have initiated an in-depth investigation into the interactions between BAs and microbiota. Leveraging existing literature, we offer a comprehensive analysis of the relationships between BAs and 16 key microbiota. This investigation encompasses bacteria (e.g., Clostridioides difficile, Staphylococcus aureus, Escherichia coli, Enterococcus, Pseudomonas aeruginosa, Mycobacterium tuberculosis, Bacteroides, Clostridium scindens, Streptococcus thermophilus, Saccharomyces boulardii, Clostridium butyricum, Lactic Acid Bacteria), fungi (e.g., Candida albicans), and viruses (e.g., Coronavirus SARS-CoV-2, influenza virus, norovirus). Our research underscores the critical role of the interplay between BAs and intestinal microbiota, including Bacteroides, Clostridium scindens, Streptococcus thermophilus, Saccharomyces boulardii, Clostridium butyricum, and Lactic Acid Bacteria, in maintaining intestinal homeostasis and combating infections. It is imperative to note that Primary bile acid (PBA) and Secondary bile acid (SBA) often exhibit distinct roles in the anti-infection process. In the antimicrobial action of BAs, SBA demonstrate antagonistic properties against a wide range of microbiota, with the exception of Norovirus. Given the intricate interplay between BAs and intestinal microbiota, and their regulatory effects on infections, we assert that BAs hold significant potential as a novel approach for preventing and treating intestinal microbial infections.
Keywords:
bile acid
; infection
; microbiota
1. Introduction
The modulation of BAs in mammalian systems constitutes an intricate procedure, jointly orchestrated by the liver, intestines, and some intestinal microbiota [1]. PBA, predominantly comprised of cholic acid and chenodeoxycholic acid (CDCA) [2], are largely reabsorbed within the enterohepatic circulation. These are subsequently converted by the intestinal microbiota, yielding SBA, largely comprised of lithocholic acid (LCA) and ursodeoxycholic acid (UDCA) [3,4]. In this metabolic cascading, enzymes engendered by the intestinal microbiota, particularly BSHs mediated by the bile salt hydrolase (BSH) gene and 7α hydroxylase facilitated by the bai operon, assume a pivotal role [5,6,7,8,9,10].
Both BAs and intestinal microbiota emerge as keystones in host metabolism, wherein their synthesized or regulated metabolites frequently function as signaling molecules, precluding the colonization of pathogens within the host [5,11].
The changes in the concentration and composition of intestinal BAs are not only pivotal in affecting the growth and colonization of various pathogens but also play a significant role in the mechanisms of disease prevention and pathogenesis [11,12].Some studies have found the vital roles of PBA and SBA in maintaining intestinal homeostasis and combating infections [13,14,15]. For example, PBA have been shown to facilitate the germination of Clostridioides difficile spores, while SBA play a role in inhibiting its proliferation [16]. Interestingly, alterations in the intestinal microbiota significantly affect the host’s health and disease progression by profoundly influencing BAs conversion dynamics [17,18]. A wide array of intestinal microbiota with BSHs activity, plays a crucial role in maintaining the balance of bile acid pools [9,10,19,20,21,22,23]. Moreover, the association between BSHs activity and various health conditions, such as obesity, cancer, and inflammatory bowel disease, has become a burgeoning research hotspot [24,25,26,27,28,29,30], and BSHs are emerging as potential therapeutic targets for metabolic diseases [31,32,33,34,35]. Recent studies have highlighted the critical role of the bai operon-mediated 7α-dehydroxylation reaction in the intestinal microbiota, predominantly carried out by members of Clostridium cluster XIVa, particularly Clostridium hiranonis and Clostridium scindens [36], which not only increases the hydrophobicity of bile acids but also triggers a range of significant biological effects, including alterations in intestinal permeability, antibiotic biosynthesis, and activation of the Farnesoid X Receptor (FXR) [37,38,39]. The bai operon has shown effectiveness in reducing intestinal inflammation [40]. Furthermore, Clostridium scindens, equipped with the bai operon, has shown promise in combatting Clostridioides difficile infections [41].
The dynamic interaction between BAs and the intestinal microbiota not only leads to changes in bile acid pools but also allows BAs to influence the structural composition of the intestinal microbiota [42]. Although earlier research focused on the interactions of PBA or SBA with specific intestinal microbiota, BAs transformation is an ongoing process facilitated by intestinal microbiota exhibiting BSHs activity [43]. Increasing evidence suggests that the structure and function of the intestinal microbiota can exert long-lasting impacts on the host [44,45]. This review aims to offer a comprehensive exploration of the interactions between BAs and key intestinal microbes from the perspective of the intestinal microbiota. In the current era of widespread antibiotic use and rising microbial resistance [46], the role of BAs as preventive and therapeutic agents is becoming increasingly important.
2. Regulatory Mechanisms of Bile Acids in Maintaining Intestinal Homeostasis and Counteracting Infections
BAs play a pivotal role in regulating intestinal homeostasis[47]. Some studies have shown that BAs can enhance intestinal epithelial permeability, thereby increasing susceptibility to infections [48]. Interestingly, natural BAs have demonstrated significant antimicrobial properties against a variety of organisms including bacteria, parasites, and fungi [49,50,51,52]. That is becasue the roles of PBA and SBA are different. For instance, PBA like taurocholic acid (TCA), can promotes Clostridioides difficile proliferation and facilitates Candida albicans colonization [12,28,53,54,55]. In contrast, SBA such as taurodeoxycholic acid, can mitigate sepsis-induced intestinal inflammation, and deoxycholic acid and LCA encourage Clostridium scindens proliferation and inhibit Clostridioides difficile spore germination [56,57,58,59]. These diverse effects could be attributed to specific bile acid species, the unique receptors they activate and their interactions with intestinal microbiota.
BAs interact with various cellular receptors, including FXR, TGR5 (Takeda G Protein-Coupled Receptor 5), Pregnane X Receptor, Sphingosine-1-Phosphate Receptor 2, and Vitamin D Receptor. FXR is activated primarily by CDCA. FXR activation strengthens the intestinal barrier, influences microbial community composition, and modulates inflammatory responses [60,61,62]. Moreover, FXR promotes the proliferation of regulatory T cells, enhancing their antiviral capabilities [63,64,65]. Taurodeoxycholic acid-induced TGR5 activation, which can reduce cAMP levels, inhibit the Myosin Light Chain Kinase pathway and thus mitigate Escherichia coli epithelial barrier damage [66,67]. Other receptors such as Sphingosine-1-Phosphate Receptor 2, Pregnane X Receptor, and Vitamin D Receptor also play important roles in inflammatory response modulation when activated by BAs [68,69].
2.1. BAs and Fungal
Interactions Between BAs and Candida albicans
Candida albicans (C.albicans), an opportunistic fungus, primarily originates from its endogenous populations in the gastrointestinal tract [70,71,72,73,74,75,76]. C.albicans frequently causes invasive infections, particularly in immunocompromised individuals or in those with dysbiosis of the intestinal microbiota [53,73,77,78,79,80].
TCA, a primary bile acid, can modulate immune responses and microbial balance within the intestine, influencing the colonization and spread of fungi like C.albicans [81]. Specifically, TCA has been shown to suppress key immune molecules, such as angiogenin-4 and CX3CR1, which are crucial for maintaining intestinal barrier integrity [82,83,84]. Additionally, TCA is associated with reduced expression of tight junction proteins [85,86,87]. This may promote the growth of harmful microbes like C. difficile and facilitate C.albicans over-colonization [28,88,89,90,91]. In contrast, SBA, specifically LCA and deoxycholic acid, can prevent the C.albicans’s morphological transformation and restricting its proliferation in the intestine [92].
During the mouse experiments investigations into intestinal microbiota composition following C.albicans infection, there’s an increase in Bacteroides, Proteobacteria, Pseudomonas, and Enterococcus, while Firmicutes levels decrease [93,94]. These changes may facilitate enhanced C.albicans colonization by altering BSHs activity and SBA concentrations in the intestine. Moreover, TCA supplementation can heighten C.albicans ‘s invasiveness and virulence by increasing specific bacterial populations, like enterohemorrhagic Escherichia col [88] (Figure 1).
2.2. BAs and Bacteria
2.2.1. Interactions Between BAs and Clostridioides difficile
Clostridioides difficile (C. difficile), is a gram-positive bacterium. Its spores can detect specific environmental cues in the gastrointestinal tract and initiat germination processes [95,96]. C. difficile can produce two major protein toxins, TcdA and TcdB, which can disrupt host cell signaling pathways and lead to apoptosis [97]. In clinical settings, C. difficile infections can range from mild diarrhea to severe pseudomembranous colitis [98]..
In the lifecycle of C. difficile, BAs play a regulatory role [99,100]. Some studies have identified that BAs can affect the proliferation of C. difficile by influencing both the structural and functional aspects of the TcdB toxin [93,101]. In addition, BAs also can cause disruptions in the equilibrium of intestinal microbiota. Specifically, TCA, a primary bile acid, has been implicated in facilitating the in vitro germination of C. difficile spores, which can promote the subsequent release of toxins [102]. Conversely, SBA like LCA and deoxycholic acid are known to inhibit the growth and toxic effects of C. difficile [99,103,104]. This inhibition includes: 1) The activation of bile acid receptors such as FXR and TGR5 by SBA, which enhances the innate immune response and inhibits C. difficile proliferation through signaling pathways, notably NF-κB [105]; 2) The direct interaction of SBA with the C-terminal region of TcdB, leading to conformational changes in the toxin and preventing its binding and toxic effects on host cells [106] (Figure 2).
2.2.2. Interactions Between BAs and Staphylococcus aureus
Staphylococcus aureus (S. aureus), a Gram-positive bacterium, presents significant clinical management challenges exacerbated by indiscriminate antibiotic use [107]. Recent research, though limited in number with only two studies identified so far, has begun to elucidate the significant role of SBA in the response to S. aureus infections.
Deoxycholic acids, a secondary bile acid, has been observed to promote the repair of tight junction proteins and substantially reduces the expression of inflammation-associated markers in mouse experiments [8]. Furthermore, deoxycholic acids can also alleviat S. aureus-induced endometritis discovered in Hu, J’s studies [108]. Its protective effects are thought to stem from deoxycholic acid’s influence on the TGR5/PKA-NF-κB-NLRP3 signaling axis [109]. However, deoxycholic acid does not directly suppress the proliferation of S. aureus [8].
Studies indicate that an imbalance in intestinal microbiota leads to an exacerbated response to mastitis in mouse experiments challenged with S. aureus, thereby intensifying the clinical symptoms [110,111]. Remarkably, supplementing the intestinal microbiota of infected mouse with BSHs-active organisms, such as Clostridium scindens, significantly reduces the inflammatory response to mastitis [8] (Figure 3).
2.2.3. Interactions Between BAs and Extended-Spectrum Beta-Lactamase-Resistant Escherichia coli
The overuse of antibiotics has led to a widespread increase in the prevalence of extended-spectrum beta-lactamase-resistant Escherichia coli (ESBL-EAEC). This particular E. coli strain is highly pathogenic, often resulting in severe diarrheal diseases. The pathological hallmarks of ESBL-EAEC infection include inflammation, epithelial cell exfoliation, and compromised epithelial barrier functionality [112].
UDCA, a secondary bile acid, has shown significant inhibitory effects on ESBL-EAEC in mouse experiments. In the context of ESBL-EAEC infection, a notable reduction in the abundance of key intestinal microbial families with BSHs activity such as Corbacteriaceae, Ruminococcaceae, and Lachnospiraceae has been observed. However, this change is effectively countered by UDCA treatment by repairing microbial imbalances [113]. Moreover, UDCA enhances tight junction functionality by upregulating TGR5 transcription and inhibiting IκB α phosphorylation[14,114] (Figure 4).
2.2.4. Interactions Between BAs and Enterococci
In the gastrointestinal tract, Enterococcus faecalis, is a commensal bacterium. However, under conditions of intestinal microbiota dysbiosis, E. faecalis may transition to a pathogenic state, particularly in elderly or immunocompromised individuals [115,116,117]. Recent clinical studies have elucidated that through the elevation of deoxycholic acid levels or the reduction of TCA, can effectively curtail the proliferation of Enterococcus faecalis. Further research suggests that deoxycholic acid ‘s growth-inhibitory effect on E. faecalis could be due to its impact on the expression of various ribosomal protein genes [118].
Vancomycin-resistant enterococci (VRE) present significant challenges in clinical settings due to their antibiotic resistance. The formation of biofilms is critical for the colonization of enterococci in various host environments [119]. Rahman’s study has revealed that LCA can curtail the growth of VRE by maintain VRE in a diplococcal state and inhibiting the morphological transformation of VRE. Additionally, LCA exposure induces genetic mutations in VRE that result in persistent diplococcal morphology, reduced biofilm production [120] (Figure 5).
2.2.5. Interactions Between BAs and Other Bacteria (Pseudomonas aeruginosa, Mycobacterium tuberculosis, etc.)
Pseudomonas aeruginosa is known for its diverse infection profiles [121]. Surprisingly, TCA as a primary bile acid, demonstrates a significant inhibitory effect on Pseudomonas aeruginosa. In detail, TCA is particularly effective in inhibiting biofilm formation and dispersing existing biofilms [122,123]. This effect is believed to originate from TCA’s modulation of Pseudomonas aeruginosa ‘s virulence factors, including its impact on metabolites like the siderophore pyochelin, thereby altering its toxicity and biofilm dynamics [124].
Mycobacterium tuberculosis, the causative agent of tuberculosis, shows a unique susceptibility pattern in the gastrointestinal tract [125]. Regions with lower bile acid concentrations, such as the terminal ileum and cecum, are more susceptible to intestinal tuberculosis [126]. BAs like CDCA, deoxycholic acid, and cholic acid have demonstrated inhibitory effects on the proliferation of Mycobacterium tuberculosis. This inhibition could be due to the detrimental impact of BAs on the distinctive lipid-rich cell wall of Mycobacterium tuberculosis [127].
Moreover, BAs influence various other pathogenic bacteria. For example, deoxycholic acid has been shown to induce the transcription of genes involved in DNA repair and recombination in response to infections by bacteria such as Escherichia coli, Salmonella enterica serovar Typhimurium, Bacillus cereus, and Listeria monocytogenes [128]. However, BAs also have a dual role; their presence has been linked to increased virulence in Shigella dysenteriae, promoting infection [129] (Figure 6).
2.2.6. Interactions Between BAs and Bacteroidetes
The Bacteroidetes phylum significantly contributes to gastrointestinal health and the prevention of infections [130]. It has reported that Bacteroides thetaiotaomicron (B. thetaiotaomicron), Bacteroides ovatus and Bacteroides fragilis can alleviate colitis in mouse experiments by regulating BA metabolism to inhibit the proliferation of C .difficile [131,132,133,134].
In related research, Bacteroides dorei strain (BDX-01) and its therapeutic effects in a colitis mouse model by regulating BA metabolism, indicated by changes in total fecal bile acid levels and bile acid ratios, and by affecting the FXR-NLRP3 signaling pathway which lead to reduced proinflammatory cytokine expression and diminished IL-1β secretion in the colon, thereby mitigating DSS-induced experimental colitis [9,135,136,137,138,139].
However, it has revealed a potential adverse role of Bacteroides fragilis NCTC9343 in gastrointestinal health, particularly concerning their BSHs activity [140]. Elevated BSH gene expression in colonizing Bacteroidetes strains can lead to an increased influx of bile acids, which may activate signaling pathways like WNT/β-catenin and NF-κB resulting in oxidative DNA damage and enhanced cellular proliferation, eventually exacerbating colorectal cancer progression in mouse experiments [9,34,141](Figure 7).
2.2.7. Interactions Between BAs and Clostridium scindens
Clostridium scindens (C. scindens) harbors a bile acid-inducible operon, bai [56]. This operon is essential for the synthesis of SBA by regulating the expression of 7α-dehydroxylase [7,57]. Some studies have discovered that C. scindens plays a crucial role in preventing the colonization and proliferation of Clostridioides difficile (C. difficile) [40]. In cases of acute C. difficile Infection, a marked decrease in both BSHs and 7α-dehydroxylase expression is observed in the cecal contents of mouse, aligning with reduced gene expressions in the Lachnospiraceae and Clostridiaceae families [142].However, introducing C. scindens into the gut of mouse with acute C. difficile Infection significantly enhances intestinal health. Particularly, C. scindens has been shown to suppress TcdA/TcdB toxin production by C. difficile and reduce its overall cellular count [40,99,143,144]. Therefore, the synergistic action of SBA and C. scindens is increasingly recognized as a critical strategy in countering intestinal colonization by this pathogenic bacterium [145] (Figure 8).
2.2.8. Interactions Between BAs and Streptococcus thermophilus
Streptococcus thermophilus MN002 (S. thermophilus), acknowledged as an efficacious probiotic [146,147], has shown promising potential in mitigating the risks associated with metabolic syndrome and colorectal tumors [148,149,150], as well as reducing the incidence of obesity, neonatal bacteremia, and meningitis caused by Escherichia coli K1 [151]. The consumption of a high-fat diet is known to disrupt the intestinal microbial equilibrium, leading to both intestinal and systemic inflammation [152,153,154].Intriguingly, deoxycholic acid can reduce the inflammatory symptoms in high-fat diet mouse experiments. Specifically, S. thermophilus is capable of optimizing BAs configurations and fostering a balanced intestinal microbiota [155,156]. This is achieved by augmenting the relative abundance of bacteria proficient in producing SBA, including members of the Ruminococcaceae, Bacteroides, Clostridium, and Blautia families[157] (Figure 9).
2.2.9. Interactions Between BAs and Saccharomyces boulardii
Saccharomyces boulardii CNCM I-745 (SB) has been shown to effectively mitigate the risk of Clostridioides difficile enteritis following antibiotic therapy in a clinical randomized controlled trial [158,159]. Central to the protective mechanism of SB is its ability to inhibit bacterial proliferation while rapidly restoring the balance of the intestinal microbiota [160]. In detail, SB not only can thwart bacterial adhesion,but also can accelerate the neutralization of enteric toxins and bolster the immune response within the intestinal mucosa [161,162,163]. Furthermore, research involving healthy volunteer cohorts has illuminated SB can safeguard the health of intestine by promoting the proliferation of microbiota with BSHs activity [28]. Complementing this, vitro studies also have discovered that SB can hinder the germination of Clostridioides difficile spores [164,165,166] (Figure 10).
2.2.10. Interactions Between BAs and Clostridium butyricum
Clostridium butyricum (C. butyricum) can modulate lipid metabolism by influencing the bile acid profile within the liver and ileum [167,168]. Research has been shown that C. butyricum supplementation can reshape the gut microbiota composition and BA distribution of intrauterine growth restricted piglets, thereby optimizing their lipid metabolism. At the same time, it significantly reduces the abundance of specific gut microbiota Streptococcus and Enterococcus in the ileum of these piglets, leading to an increase in conjugated BAs, thereby activating key liver receptors like liver X receptor α and FXR, that is crucial for reducing inflammatory response and protecting normal liver function [166,169,170,171,172,173,174,175,176].
Clostridium butyricum strain CCFM1299 administration significantly leads to an increase in UDCA levels in feces and taurocholic acid levels in serum, thereby activating TGR5 and inhibiting FXR, subsequently enhancing GLP-1 production in the intestine, which helps regulate blood sugar and reduce obesity [177,178,179,180]. Furthermore, C. butyricum reshapes the microbiota by increasing butyric acid levels, maintaining secondary bile acid balance, and attenuating the inhibitory effects of the FXR/SHP pathway on lipid synthesis [181]. And it also activates the butyrate/GPR43 pathway, reducing damage to the intestinal barrier and restoring the intestinal immune microenvironment in CP rabbits [182] (Figure 11).
2.2.11. Interactions Between BAs and Lactic Acid Bacteria
Pediococcus pentosaceus Li05 belong to the Pediococcus genus of the Lactobacillaceae family. Li05 can improve tight junction proteins and downregulates inflammatory responses in mouse experiments by modulating intestinal microbiota and bile acid metabolism [183]. And it can promote the growth of beneficial microbial taxa such as Lactobacillus, Prevotella, and Paraprevotella, while inhibiting opportunistic pathogens, thereby altering the BAs composition and influencing liver injury processes [54,184]. It has reported that Li05 treatment notably reduced weight loss, liver damage, and bile stasis in DDC-induced cholestasis mouse experiments[185,186], which is likely linked to Li05’s modulation of the intestinal microbiota, particularly enhancing propionate and butyrate-producing bacteria like Anaerostipes and Eubacterium. Anaerostipes and Eubacterium that all of them known for metabolizing inositol into propionic and butyric acids and converting PBA into SBA via 7α-dehydroxylation [19,187,188].
Liu L et al also revealed Lactiplantibacillus plantarum LPJZ-658 modulates intestinal microbiota and BA metabolism in mouse model which reveals the potential for treating non-alcoholic fatty liver [189]. Furthermore, Lactiplantibacillus plantarum LPJZ-658 increased the abundance of Firmicutes and Actinobacteria, suggesting a healthier intestinal environment conducive to non-alcoholic steatohepatitis mitigation [190,191,192] (Figure 12).
2.3. BAs and Viruses
2.3.1. Interactions Between BAs and Coronavirus SARS-CoV-2
Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2) utilizes the receptor binding domain within its spike protein to engage the host’s angiotensin-converting enzyme 2 (ACE2) receptor, facilitating cellular entry [193,194,195]. Recent investigations have revealed the potential of BAs, particularly UDCA and CDCA, in disrupting this critical virus-host interaction.
Some studies have identified that UDCA can directly bind the receptor binding domain of SARS-CoV-2, thereby diminishing its affinity for ACE2 and potentially mitigating cellular damage [196,197,198]. Specifically, UDCA appear to alter the virus’s structural integrity, allowing the penetration of polar inhibitors and solvents into the viral cells, which could impede replication [198,199].
Beyond direct antiviral effects, UDCA also can modulate the host’s immune response. The cytokine storm, a critical factor in severe COVID-19 cases, can be mitigated by UDCA’s anti-inflammatory, antioxidant, immunomodulatory, and anti-apoptotic properties [200,201,202,203,204,205]. Notably, UDCA can also reduce ACE2 expression in various human and animal tissues by regulating ACE2 transcription [206,207,208] [209,210]. In addition, retrospective studies have indicated that UDCA can improve clinical outcomes in patients [211]. However, UDCA did not demonstrate a reduction in susceptibility to SARS-CoV-2 infection in pediatric populations [212].
Emerging research suggests a correlation between the intestinal microbiome, particularly the Collinsella genus, and COVID-19 outcomes. Hirayama M. et al. employed machine learning to uncover a potential link between intestinal Collinsella and reduced COVID-19 severity [213]. UDCA produced by Collinsella may prevent COVID-19 infection and ameliorate acute respiratory distress syndrome in COVID-19 by suppressing cytokine storm syndrome in clinical setting [214] (Figure 13).
2.3.2. Interactions Between BAs and Other Viruses (Influenza Virus, Norovirus, etc.)
Influenza A virus (IAV), a significant respiratory pathogen. Recent studies have uncovered the antiviral potential of CDCA and sodium taurocholate against IAV. They attenuate IAV infection by inhibiting the nuclear export of viral ribonucleoproteins and modulating the Toll-like receptor 4/NF-κB signaling pathway [215,216]. Specifically, CDCA, a secondary bile acid, shows promise in inhibiting IAV subtypes, including H5N1, H9N2, and H1N1, by interfering with viral ribonucleoproteins nuclear export and inhibiting viral replication [215]. Sodium taurocholate, a primary bile acid derivative , surprisingly exhibits antiviral efficacy against various influenza strains, including H5N6 and H3N2, by targeting the early stages of viral transcription and replication via the TLR4/NF-κB pathway [217].
BAs play a interesting role in norovirus infection [218,219]. Glycine deoxycholic acid, a secondary BA, could enhance murine noroviruses infectivity [220]. In addition, The intestinal microbiota distinctly modulates norovirus infection dynamics in different intestinal regions, with BAs mediating their inhibitory effect in the proximal small intestine, while bile acid receptors regulate infection in the distal small intestine [221,222].
3. In Conclusion
The regulation of BA is a complex process in mammalian systems. Intestinal microbiota play a crucial role in converting PBA to SBA by regulating the metabolic activities of BSHs and 7α-hydroxylase. Here, we explored the interactions between BAs and a comprehensive array of 16 key intestinal microbiota. We reveal that SBA demonstrate a robust resistance against infections induced by these microbiota, with the notable exception of norovirus. Furthermore, the interplay between BAs and specific intestinal microbiota, including Bacteroides, Clostridium scindens, Streptococcus thermophilus, Saccharomyces boulardii, Clostridium butyricum, and Lactic Acid Bacteria, plays a crucial role in maintaining intestinal homeostasis and combating infections.
SBA combat infections in several ways. First, SBA slow down the growth of harmful microbiota, inhibit the transformation of Candida albicans, reduce Clostridioides difficile spore sprouting, disrupt VRE biofilms, and weaken Mycobacterium tuberculosis cell walls. SBA also reduce SARS-CoV-2’s binding to ACE2 receptors and inhibit influenza virus replication. Second, SBA modify the structure of Clostridioides difficile’s TcdB toxin and trigger the NF-κB signaling pathway via bile acid receptors like FXR and TGR5. This interaction boosts the body’s immune defenses, enhancing responses against pathogens like Clostridioides difficile and SARS-CoV-2. Last, the synergy between SBA and some specific intestinal microbiota is crucial, particularly in enhancing their anti-infective potential. Clostridium butyricum, for example, promotes intestinal health through enterohepatic circulation, reducing BSH-active microbiota and increasing conjugated BAs production. However, certain Bacteroidetes strains with high BSH gene expression may inadvertently increase BAs entry into the colon, potentially triggering colorectal cancer.
The interaction between viruses and BAs is complex. Most SBA preserving intestinal mucosal health, but glycine deoxycholic acid, a secondary bile acid potentially exacerbating norovirus infections. In addition, sodium taurocholate, a primary bile acid derivative, surprisingly shows efficacy against the influenza virus.
BAs are diverse in type, each with unique physical structures and biological properties. The dynamic metabolism of BAs in the human body results in fluctuations in types and concentrations along the intestinal tract. Current research, often utilizing fixed BAs formulations, may not fully capture these variations. Nonetheless, it is evident that SBA generally exert a favorable anti-infectious influence against most intestinal microorganism-induced infections. Given the intricate interplay between BAs and intestinal microbiota, and their regulatory effects on infections, we assert that BAs hold significant potential as a novel approach for preventing and treating intestinal microbial infections.
Author Contributions
Wenweiran Li, was the main contributor to thein writing of the manuscript. Hui Chen was responsible for the collection and collation of reference articles and participated in the writing of the manuscript. Jianguo Tang made great contributions to the subsequent revision and improvement work. The author order was determined on the basis of seniority. All authors read and approved the final manuscript.
Funding
Not applicable.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data were created or analyzed in this study.
Acknowledgments
Not applicable.
Conflicts of Interest
The authors declare no competing interests.
Abbreviations
| BAs: Bile acids |
| BSHs: Bile salt hydrolases |
| BSH: Bile salt hydrolase |
| PBA: Primary bile acid |
| CDCA: Chenodeoxycholic acid |
| TCA: Taurocholic acid |
| SBA: Secondary bile acid |
| LCA: Lithocholic acid |
| UDCA: Ursodeoxycholic acid |
| FXR: Farnesoid X Receptor |
| TGR5: Takeda G Protein-Coupled Receptor 5 |
| ACE2: Angiotensin-converting enzyme 2 |
| C. albicans: Candida albicans |
| C. difficile: Clostridioides difficile |
| C. scindens: Clostridium scindens |
| E. coli: Escherichia coli |
| S. aureus: Staphylococcus aureus |
| ESBL-EAEC: Extended-spectrum beta-lactamase-resistant Escherichia coli |
| VRE: Vancomycin-resistant enterococci |
| M. tuberculosis: Mycobacterium tuberculosis |
| S. thermophilus: Streptococcus thermophilus |
| SB: Saccharomyces boulardii |
| C. butyricum: Clostridium butyricum |
| SARS-CoV-2: Severe Acute Respiratory Syndrome Coronavirus 2 |
| IAV: Influenza A virus |
| COVID-19: Coronavirus Disease 2019 |
| PBA: Cholic acid, Chenodeoxycholic acid, Taurocholic acid, Sodium.taurocholate. |
| SBA: Lithocholic acid, Deoxycholic acid, Ursodeoxycholic acid, Taurodeoxycholic acid, Glycine deoxycholic acid. |
References
- Guzior, D.V.; Quinn, R.A. Review: microbial transformations of human bile acids. Microbiome 2021, 9, 140. [Google Scholar] [CrossRef] [PubMed]
- Martoni, C.J.; Labbe, A.; Ganopolsky, J.G.; Prakash, S.; Jones, M.L. Changes in bile acids, FGF-19 and sterol absorption in response to bile salt hydrolase active L. reuteri NCIMB 30242. Gut Microbes 2015, 6, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Di Ciaula, A.; Garruti, G.; Lunardi Baccetto, R.; Molina-Molina, E.; Bonfrate, L.; Wang, D.Q.; Portincasa, P. Bile Acid Physiology. Ann Hepatol 2017, 16, s4–s14. [Google Scholar] [CrossRef]
- Schoeler, M.; Caesar, R. Dietary lipids, gut microbiota and lipid metabolism. Rev Endocr Metab Disord 2019, 20, 461–472. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zheng, L.; Zhou, Z.; Yao, D.; Huang, Y.; Liu, B.; Duan, Y.; Li, Y. Review article: insights into the bile acid-gut microbiota axis in intestinal failure-associated liver disease-redefining the treatment approach. Aliment Pharmacol Ther 2022, 55, 49–63. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Hong, Y.; Zheng, N.; Xie, G.; Lyu, Y.; Gu, Y.; Xi, C.; Chen, L.; Wu, G.; Li, Y.; et al. Gut microbiota remodeling reverses aging-associated inflammation and dysregulation of systemic bile acid homeostasis in mice sex-specifically. Gut Microbes 2020, 11, 1450–1474. [Google Scholar] [CrossRef] [PubMed]
- Jose, S.; Mukherjee, A.; Horrigan, O.; Setchell, K.D.R.; Zhang, W.; Moreno-Fernandez, M.E.; Andersen, H.; Sharma, D.; Haslam, D.B.; Divanovic, S.; et al. Obeticholic acid ameliorates severity of Clostridioides difficile infection in high fat diet-induced obese mice. Mucosal Immunol 2021, 14, 500–510. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Wang, J.; Li, X.; Li, Y.; Ye, C. Deoxycholic acid inhibits Staphylococcus aureus-induced endometritis through regulating TGR5/PKA/NF-kappaB signaling pathway. Int Immunopharmacol 2023, 118, 110004. [Google Scholar] [CrossRef] [PubMed]
- Jia, W.; Xie, G.; Jia, W. Bile acid-microbiota crosstalk in gastrointestinal inflammation and carcinogenesis. Nat Rev Gastroenterol Hepatol 2018, 15, 111–128. [Google Scholar] [CrossRef]
- Ridlon, J.M.; Daniel, S.L.; Gaskins, H.R. The Hylemon-Bjorkhem pathway of bile acid 7-dehydroxylation: history, biochemistry, and microbiology. J Lipid Res 2023, 64, 100392. [Google Scholar] [CrossRef] [PubMed]
- Jia, W.; Li, Y.; Cheung, K.C.P.; Zheng, X. Bile acid signaling in the regulation of whole body metabolic and immunological homeostasis. Sci China Life Sci 2023. [Google Scholar] [CrossRef] [PubMed]
- Guinan, J.; Thangamani, S. Antibiotic-induced alterations in taurocholic acid levels promote gastrointestinal colonization of Candida albicans. FEMS Microbiol Lett 2018, 365. [Google Scholar] [CrossRef] [PubMed]
- Winkler, E.S.; Shrihari, S.; Hykes, B.L., Jr.; Handley, S.A.; Andhey, P.S.; Huang, Y.S.; Swain, A.; Droit, L.; Chebrolu, K.K.; Mack, M.; et al. The Intestinal Microbiome Restricts Alphavirus Infection and Dissemination through a Bile Acid-Type I IFN Signaling Axis. Cell 2020, 182, 901–918. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Winglee, K.; Gharaibeh, R.Z.; Gauthier, J.; He, Z.; Tripathi, P.; Avram, D.; Bruner, S.; Fodor, A.; Jobin, C. Microbiota-Derived Metabolic Factors Reduce Campylobacteriosis in Mice. Gastroenterology 2018, 154, 1751–1763. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Latorre, J.D.; Bansal, M.; Abraha, M.; Al-Rubaye, B.; Tellez-Isaias, G.; Hargis, B.; Sun, X. Microbial metabolite deoxycholic acid controls Clostridium perfringens-induced chicken necrotic enteritis through attenuating inflammatory cyclooxygenase signaling. Sci Rep 2019, 9, 14541. [Google Scholar] [CrossRef] [PubMed]
- Woollett, L.A.; Buckley, D.D.; Yao, L.; Jones, P.J.; Granholm, N.A.; Tolley, E.A.; Tso, P.; Heubi, J.E. Cholic acid supplementation enhances cholesterol absorption in humans. Gastroenterology 2004, 126, 724–731. [Google Scholar] [CrossRef] [PubMed]
- Missmer, S.A.; Tu, F.F.; Agarwal, S.K.; Chapron, C.; Soliman, A.M.; Chiuve, S.; Eichner, S.; Flores-Caldera, I.; Horne, A.W.; Kimball, A.B.; et al. Impact of Endometriosis on Life-Course Potential: A Narrative Review. Int J Gen Med 2021, 14, 9–25. [Google Scholar] [CrossRef]
- As-Sanie, S.; Black, R.; Giudice, L.C.; Gray Valbrun, T.; Gupta, J.; Jones, B.; Laufer, M.R.; Milspaw, A.T.; Missmer, S.A.; Norman, A.; et al. Assessing research gaps and unmet needs in endometriosis. Am J Obstet Gynecol 2019, 221, 86–94. [Google Scholar] [CrossRef] [PubMed]
- Ridlon, J.M.; Kang, D.J.; Hylemon, P.B. Bile salt biotransformations by human intestinal bacteria. J Lipid Res 2006, 47, 241–259. [Google Scholar] [CrossRef]
- Xu, F.; Hu, X.J.; Singh, W.; Geng, W.; Tikhonova, I.G.; Lin, J. The complex structure of bile salt hydrolase from Lactobacillus salivarius reveals the structural basis of substrate specificity. Sci Rep 2019, 9, 12438. [Google Scholar] [CrossRef]
- Lundeen, S.G.; Savage, D.C. Multiple forms of bile salt hydrolase from Lactobacillus sp. strain 100-100. J Bacteriol 1992, 174, 7217–7220. [Google Scholar] [CrossRef]
- Tian, Y.; Gui, W.; Koo, I.; Smith, P.B.; Allman, E.L.; Nichols, R.G.; Rimal, B.; Cai, J.; Liu, Q.; Patterson, A.D. The microbiome modulating activity of bile acids. Gut Microbes 2020, 11, 979–996. [Google Scholar] [CrossRef] [PubMed]
- Long, S.L.; Gahan, C.G.M.; Joyce, S.A. Interactions between gut bacteria and bile in health and disease. Mol Aspects Med 2017, 56, 54–65. [Google Scholar] [CrossRef] [PubMed]
- Ma, C.; Han, M.; Heinrich, B.; Fu, Q.; Zhang, Q.; Sandhu, M.; Agdashian, D.; Terabe, M.; Berzofsky, J.A.; Fako, V.; et al. Gut microbiome-mediated bile acid metabolism regulates liver cancer via NKT cells. Science 2018, 360. [Google Scholar] [CrossRef]
- Burgess, S.L.; Leslie, J.L.; Uddin, J.; Oakland, D.N.; Gilchrist, C.; Moreau, G.B.; Watanabe, K.; Saleh, M.; Simpson, M.; Thompson, B.A.; et al. Gut microbiome communication with bone marrow regulates susceptibility to amebiasis. J Clin Invest 2020, 130, 4019–4024. [Google Scholar] [CrossRef]
- Alavi, S.; Mitchell, J.D.; Cho, J.Y.; Liu, R.; Macbeth, J.C.; Hsiao, A. Interpersonal Gut Microbiome Variation Drives Susceptibility and Resistance to Cholera Infection. Cell 2020, 181, 1533–1546. [Google Scholar] [CrossRef] [PubMed]
- Wilson, K.H. Efficiency of various bile salt preparations for stimulation of Clostridium difficile spore germination. J Clin Microbiol 1983, 18, 1017–1019. [Google Scholar] [CrossRef]
- Theriot, C.M.; Koenigsknecht, M.J.; Carlson, P.E., Jr.; Hatton, G.E.; Nelson, A.M.; Li, B.; Huffnagle, G.B.; J, Z.L.; Young, V.B. Antibiotic-induced shifts in the mouse gut microbiome and metabolome increase susceptibility to Clostridium difficile infection. Nat Commun 2014, 5, 3114. [Google Scholar] [CrossRef]
- Mullish, B.H.; McDonald, J.A.K.; Pechlivanis, A.; Allegretti, J.R.; Kao, D.; Barker, G.F.; Kapila, D.; Petrof, E.O.; Joyce, S.A.; Gahan, C.G.M.; et al. Microbial bile salt hydrolases mediate the efficacy of faecal microbiota transplant in the treatment of recurrent Clostridioides difficile infection. Gut 2019, 68, 1791–1800. [Google Scholar] [CrossRef]
- Stenman, L.K.; Holma, R.; Eggert, A.; Korpela, R. A novel mechanism for gut barrier dysfunction by dietary fat: epithelial disruption by hydrophobic bile acids. Am J Physiol Gastrointest Liver Physiol 2013, 304, G227–G234. [Google Scholar] [CrossRef]
- Chiang, J.Y.L.; Ferrell, J.M. Discovery of farnesoid X receptor and its role in bile acid metabolism. Mol Cell Endocrinol 2022, 548, 111618. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Xie, C.; Wang, G.; Wu, Y.; Wu, Q.; Wang, X.; Liu, J.; Deng, Y.; Xia, J.; Chen, B.; et al. Gut microbiota and intestinal FXR mediate the clinical benefits of metformin. Nat Med 2018, 24, 1919–1929. [Google Scholar] [CrossRef] [PubMed]
- Qi, X.; Yun, C.; Sun, L.; Xia, J.; Wu, Q.; Wang, Y.; Wang, L.; Zhang, Y.; Liang, X.; Wang, L.; et al. Gut microbiota-bile acid-interleukin-22 axis orchestrates polycystic ovary syndrome. Nat Med 2019, 25, 1225–1233. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Zhang, Y.; Cai, J.; Rimal, B.; Rocha, E.R.; Coleman, J.P.; Zhang, C.; Nichols, R.G.; Luo, Y.; Kim, B.; et al. Bile salt hydrolase in non-enterotoxigenic Bacteroides potentiates colorectal cancer. Nat Commun 2023, 14, 755. [Google Scholar] [CrossRef] [PubMed]
- Joyce, S.A.; MacSharry, J.; Casey, P.G.; Kinsella, M.; Murphy, E.F.; Shanahan, F.; Hill, C.; Gahan, C.G. Regulation of host weight gain and lipid metabolism by bacterial bile acid modification in the gut. Proc Natl Acad Sci U S A 2014, 111, 7421–7426. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.D.; Myers, C.J.; Harris, S.C.; Kakiyama, G.; Lee, I.K.; Yun, B.S.; Matsuzaki, K.; Furukawa, M.; Min, H.K.; Bajaj, J.S.; et al. Bile Acid 7alpha-Dehydroxylating Gut Bacteria Secrete Antibiotics that Inhibit Clostridium difficile: Role of Secondary Bile Acids. Cell Chem Biol 2019, 26, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Chiang, J.Y. Nuclear receptors in bile acid metabolism. Drug Metab Rev 2013, 45, 145–155. [Google Scholar] [CrossRef] [PubMed]
- Moschetta, A.; Bookout, A.L.; Mangelsdorf, D.J. Prevention of cholesterol gallstone disease by FXR agonists in a mouse model. Nat Med 2004, 10, 1352–1358. [Google Scholar] [CrossRef] [PubMed]
- Jin, W.B.; Li, T.T.; Huo, D.; Qu, S.; Li, X.V.; Arifuzzaman, M.; Lima, S.F.; Shi, H.Q.; Wang, A.; Putzel, G.G.; et al. Genetic manipulation of gut microbes enables single-gene interrogation in a complex microbiome. Cell 2022, 185, 547–562. [Google Scholar] [CrossRef]
- Reed, A.D.; Nethery, M.A.; Stewart, A.; Barrangou, R.; Theriot, C.M. Strain-Dependent Inhibition of Clostridioides difficile by Commensal Clostridia Carrying the Bile Acid-Inducible (bai) Operon. J Bacteriol 2020, 202. [Google Scholar] [CrossRef]
- Hylemon, P.B.; Zhou, H.; Pandak, W.M.; Ren, S.; Gil, G.; Dent, P. Bile acids as regulatory molecules. J Lipid Res 2009, 50, 1509–1520. [Google Scholar] [CrossRef] [PubMed]
- van Best, N.; Rolle-Kampczyk, U.; Schaap, F.G.; Basic, M.; Olde Damink, S.W.M.; Bleich, A.; Savelkoul, P.H.M.; von Bergen, M.; Penders, J.; Hornef, M.W. Bile acids drive the newborn's gut microbiota maturation. Nat Commun 2020, 11, 3692. [Google Scholar] [CrossRef] [PubMed]
- Cai, J.; Rimal, B.; Jiang, C.; Chiang, J.Y.L.; Patterson, A.D. Bile acid metabolism and signaling, the microbiota, and metabolic disease. Pharmacol Ther 2022, 237, 108238. [Google Scholar] [CrossRef]
- Saffouri, G.B.; Shields-Cutler, R.R.; Chen, J.; Yang, Y.; Lekatz, H.R.; Hale, V.L.; Cho, J.M.; Battaglioli, E.J.; Bhattarai, Y.; Thompson, K.J.; et al. Small intestinal microbial dysbiosis underlies symptoms associated with functional gastrointestinal disorders. Nat Commun 2019, 10, 2012. [Google Scholar] [CrossRef] [PubMed]
- Lynch, S.V.; Pedersen, O. The Human Intestinal Microbiome in Health and Disease. N Engl J Med 2016, 375, 2369–2379. [Google Scholar] [CrossRef] [PubMed]
- Pilmis, B.; Le Monnier, A.; Zahar, J.R. Gut Microbiota, Antibiotic Therapy and Antimicrobial Resistance: A Narrative Review. Microorganisms 2020, 8. [Google Scholar] [CrossRef]
- Calzadilla, N.; Comiskey, S.M.; Dudeja, P.K.; Saksena, S.; Gill, R.K.; Alrefai, W.A. Bile acids as inflammatory mediators and modulators of intestinal permeability. Front Immunol 2022, 13, 1021924. [Google Scholar] [CrossRef]
- di Gregorio, M.C.; Cautela, J.; Galantini, L. Physiology and Physical Chemistry of Bile Acids. Int J Mol Sci 2021, 22. [Google Scholar] [CrossRef]
- Luu, T.H.; Bard, J.M.; Carbonnelle, D.; Chaillou, C.; Huvelin, J.M.; Bobin-Dubigeon, C.; Nazih, H. Lithocholic bile acid inhibits lipogenesis and induces apoptosis in breast cancer cells. Cell Oncol (Dordr) 2018, 41, 13–24. [Google Scholar] [CrossRef]
- Goldberg, A.A.; Titorenko, V.I.; Beach, A.; Sanderson, J.T. Bile acids induce apoptosis selectively in androgen-dependent and -independent prostate cancer cells. PeerJ 2013, 1, e122. [Google Scholar] [CrossRef]
- Huang, F.; Zheng, X.; Ma, X.; Jiang, R.; Zhou, W.; Zhou, S.; Zhang, Y.; Lei, S.; Wang, S.; Kuang, J.; et al. Theabrownin from Pu-erh tea attenuates hypercholesterolemia via modulation of gut microbiota and bile acid metabolism. Nat Commun 2019, 10, 4971. [Google Scholar] [CrossRef]
- Ma, J.; Huo, H.; Zhang, H.; Wang, L.; Meng, Y.; Jin, F.; Wang, X.; Zhao, Y.; Zhao, Y.; Tu, P.; et al. 2-(2-phenylethyl)chromone-enriched extract of the resinous heartwood of Chinese agarwood (Aquilaria sinensis) protects against taurocholic acid-induced gastric epithelial cells apoptosis through Perk/eIF2alpha/CHOP pathway. Phytomedicine 2022, 98, 153935. [Google Scholar] [CrossRef] [PubMed]
- Datta, A.; Hernandez-Franco, J.F.; Park, S.; Olson, M.R.; HogenEsch, H.; Thangamani, S. Bile Acid Regulates Mononuclear Phagocytes and T Helper 17 Cells to Control Candida albicans in the Intestine. J Fungi (Basel) 2022, 8. [Google Scholar] [CrossRef]
- Theriot, C.M.; Bowman, A.A.; Young, V.B. Antibiotic-Induced Alterations of the Gut Microbiota Alter Secondary Bile Acid Production and Allow for Clostridium difficile Spore Germination and Outgrowth in the Large Intestine. mSphere 2016, 1. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.X.; Ren, C.; Yao, R.Q.; Luo, Y.N.; Yin, Y.; Wu, Y.; Dong, N.; Zhu, X.M.; Yao, Y.M. Sestrin2 protects against lethal sepsis by suppressing the pyroptosis of dendritic cells. Cell Mol Life Sci 2021, 78, 8209–8227. [Google Scholar] [CrossRef] [PubMed]
- Shen, A. A Gut Odyssey: The Impact of the Microbiota on Clostridium difficile Spore Formation and Germination. PLoS Pathog 2015, 11, e1005157. [Google Scholar] [CrossRef] [PubMed]
- Greathouse, K.L.; Harris, C.C.; Bultman, S.J. Dysfunctional families: Clostridium scindens and secondary bile acids inhibit the growth of Clostridium difficile. Cell Metab 2015, 21, 9–10. [Google Scholar] [CrossRef] [PubMed]
- Marion, S.; Studer, N.; Desharnais, L.; Menin, L.; Escrig, S.; Meibom, A.; Hapfelmeier, S.; Bernier-Latmani, R. In vitro and in vivo characterization of Clostridium scindens bile acid transformations. Gut Microbes 2019, 10, 481–503. [Google Scholar] [CrossRef] [PubMed]
- He, Z.; Ma, Y.; Yang, S.; Zhang, S.; Liu, S.; Xiao, J.; Wang, Y.; Wang, W.; Yang, H.; Li, S.; et al. Gut microbiota-derived ursodeoxycholic acid from neonatal dairy calves improves intestinal homeostasis and colitis to attenuate extended-spectrum beta-lactamase-producing enteroaggregative Escherichia coli infection. Microbiome 2022, 10, 79. [Google Scholar] [CrossRef]
- Sorribas, M.; Jakob, M.O.; Yilmaz, B.; Li, H.; Stutz, D.; Noser, Y.; de Gottardi, A.; Moghadamrad, S.; Hassan, M.; Albillos, A.; et al. FXR modulates the gut-vascular barrier by regulating the entry sites for bacterial translocation in experimental cirrhosis. J Hepatol 2019, 71, 1126–1140. [Google Scholar] [CrossRef]
- Xiao, Y.; Wang, Y.; Liu, Y.; Wang, W.; Tian, X.; Chen, S.; Lu, Y.; Du, J.; Cai, W. A nonbile acid farnesoid X receptor agonist tropifexor potently inhibits cholestatic liver injury and fibrosis by modulating the gut-liver axis. Liver Int 2021, 41, 2117–2131. [Google Scholar] [CrossRef] [PubMed]
- Campbell, C.; McKenney, P.T.; Konstantinovsky, D.; Isaeva, O.I.; Schizas, M.; Verter, J.; Mai, C.; Jin, W.B.; Guo, C.J.; Violante, S.; et al. Bacterial metabolism of bile acids promotes generation of peripheral regulatory T cells. Nature 2020, 581, 475–479. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Chang, K.O. Inhibitory effects of bile acids and synthetic farnesoid X receptor agonists on rotavirus replication. J Virol 2011, 85, 12570–12577. [Google Scholar] [CrossRef] [PubMed]
- Chang, K.O.; George, D.W. Bile acids promote the expression of hepatitis C virus in replicon-harboring cells. J Virol 2007, 81, 9633–9640. [Google Scholar] [CrossRef]
- Song, M.; Zhang, F.; Fu, Y.; Yi, X.; Feng, S.; Liu, Z.; Deng, D.; Yang, Q.; Yu, M.; Zhu, C.; et al. Tauroursodeoxycholic acid (TUDCA) improves intestinal barrier function associated with TGR5-MLCK pathway and the alteration of serum metabolites and gut bacteria in weaned piglets. J Anim Sci Biotechnol 2022, 13, 73. [Google Scholar] [CrossRef] [PubMed]
- Andrade, J.M.; Faustino, C.; Garcia, C.; Ladeiras, D.; Reis, C.P.; Rijo, P. Rosmarinus officinalis L.: an update review of its phytochemistry and biological activity. Future Sci OA 2018, 4, FSO283. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Chen, S.; Xiang, H.; Wang, X.; Xiao, J.; Zhao, S.; Shu, Z.; Ouyang, J.; Liang, Z.; Deng, M.; et al. S1PR2/RhoA/ROCK1 pathway promotes inflammatory bowel disease by inducing intestinal vascular endothelial barrier damage and M1 macrophage polarization. Biochem Pharmacol 2022, 201, 115077. [Google Scholar] [CrossRef] [PubMed]
- Venkatesh, M.; Mukherjee, S.; Wang, H.; Li, H.; Sun, K.; Benechet, A.P.; Qiu, Z.; Maher, L.; Redinbo, M.R.; Phillips, R.S.; et al. Symbiotic bacterial metabolites regulate gastrointestinal barrier function via the xenobiotic sensor PXR and Toll-like receptor 4. Immunity 2014, 41, 296–310. [Google Scholar] [CrossRef]
- Zangerolamo, L.; Vettorazzi, J.F.; Rosa, L.R.O.; Carneiro, E.M.; Barbosa, H.C.L. The bile acid TUDCA and neurodegenerative disorders: An overview. Life Sci 2021, 272, 119252. [Google Scholar] [CrossRef]
- Low, C.Y.; Rotstein, C. Emerging fungal infections in immunocompromised patients. F1000 Med Rep 2011, 3, 14. [Google Scholar] [CrossRef]
- Perfect, J.R.; Hachem, R.; Wingard, J.R. Update on epidemiology of and preventive strategies for invasive fungal infections in cancer patients. Clin Infect Dis 2014, 59 Suppl 5, S352–355. [Google Scholar] [CrossRef]
- Zhai, B.; Ola, M.; Rolling, T.; Tosini, N.L.; Joshowitz, S.; Littmann, E.R.; Amoretti, L.A.; Fontana, E.; Wright, R.J.; Miranda, E.; et al. High-resolution mycobiota analysis reveals dynamic intestinal translocation preceding invasive candidiasis. Nat Med 2020, 26, 59–64. [Google Scholar] [CrossRef] [PubMed]
- Thangamani, S.; Monasky, R.; Lee, J.K.; Antharam, V.; HogenEsch, H.; Hazbun, T.R.; Jin, Y.; Gu, H.; Guo, G.L. Bile Acid Regulates the Colonization and Dissemination of Candida albicans from the Gastrointestinal Tract by Controlling Host Defense System and Microbiota. J Fungi (Basel) 2021, 7. [Google Scholar] [CrossRef] [PubMed]
- Nucci, M.; Anaissie, E. Revisiting the source of candidemia: skin or gut? Clin Infect Dis 2001, 33, 1959–1967. [Google Scholar] [CrossRef] [PubMed]
- Miranda, L.N.; van der Heijden, I.M.; Costa, S.F.; Sousa, A.P.; Sienra, R.A.; Gobara, S.; Santos, C.R.; Lobo, R.D.; Pessoa, V.P., Jr.; Levin, A.S. Candida colonisation as a source for candidaemia. J Hosp Infect 2009, 72, 9–16. [Google Scholar] [CrossRef]
- Krause, R.; Krejs, G.J.; Wenisch, C.; Reisinger, E.C. Elevated fecal Candida counts in patients with antibiotic-associated diarrhea: role of soluble fecal substances. Clin Diagn Lab Immunol 2003, 10, 167–168. [Google Scholar] [CrossRef] [PubMed]
- Samonis, G.; Gikas, A.; Anaissie, E.J.; Vrenzos, G.; Maraki, S.; Tselentis, Y.; Bodey, G.P. Prospective evaluation of effects of broad-spectrum antibiotics on gastrointestinal yeast colonization of humans. Antimicrob Agents Chemother 1993, 37, 51–53. [Google Scholar] [CrossRef] [PubMed]
- Netea, M.G.; Joosten, L.A.; van der Meer, J.W.; Kullberg, B.J.; van de Veerdonk, F.L. Immune defence against Candida fungal infections. Nat Rev Immunol 2015, 15, 630–642. [Google Scholar] [CrossRef] [PubMed]
- Koh, A.Y.; Kohler, J.R.; Coggshall, K.T.; Van Rooijen, N.; Pier, G.B. Mucosal damage and neutropenia are required for Candida albicans dissemination. PLoS Pathog 2008, 4, e35. [Google Scholar] [CrossRef]
- Drummond, R.A.; Gaffen, S.L.; Hise, A.G.; Brown, G.D. Innate Defense against Fungal Pathogens. Cold Spring Harb Perspect Med 2014, 5. [Google Scholar] [CrossRef]
- Leonardi, I.; Li, X.; Semon, A.; Li, D.; Doron, I.; Putzel, G.; Bar, A.; Prieto, D.; Rescigno, M.; McGovern, D.P.B.; et al. CX3CR1(+) mononuclear phagocytes control immunity to intestinal fungi. Science 2018, 359, 232–236. [Google Scholar] [CrossRef] [PubMed]
- Hooper, L.V.; Stappenbeck, T.S.; Hong, C.V.; Gordon, J.I. Angiogenins: a new class of microbicidal proteins involved in innate immunity. Nat Immunol 2003, 4, 269–273. [Google Scholar] [CrossRef] [PubMed]
- Medina-Contreras, O.; Geem, D.; Laur, O.; Williams, I.R.; Lira, S.A.; Nusrat, A.; Parkos, C.A.; Denning, T.L. CX3CR1 regulates intestinal macrophage homeostasis, bacterial translocation, and colitogenic Th17 responses in mice. J Clin Invest 2011, 121, 4787–4795. [Google Scholar] [CrossRef] [PubMed]
- Zarrinpar, A.; Chaix, A.; Xu, Z.Z.; Chang, M.W.; Marotz, C.A.; Saghatelian, A.; Knight, R.; Panda, S. Antibiotic-induced microbiome depletion alters metabolic homeostasis by affecting gut signaling and colonic metabolism. Nat Commun 2018, 9, 2872. [Google Scholar] [CrossRef]
- Wahlstrom, A.; Sayin, S.I.; Marschall, H.U.; Backhed, F. Intestinal Crosstalk between Bile Acids and Microbiota and Its Impact on Host Metabolism. Cell Metab 2016, 24, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Fan, D.; Coughlin, L.A.; Neubauer, M.M.; Kim, J.; Kim, M.S.; Zhan, X.; Simms-Waldrip, T.R.; Xie, Y.; Hooper, L.V.; Koh, A.Y. Activation of HIF-1alpha and LL-37 by commensal bacteria inhibits Candida albicans colonization. Nat Med 2015, 21, 808–814. [Google Scholar] [CrossRef]
- Wilson, K.H.; Kennedy, M.J.; Fekety, F.R. Use of sodium taurocholate to enhance spore recovery on a medium selective for Clostridium difficile. J Clin Microbiol 1982, 15, 443–446. [Google Scholar] [CrossRef]
- Sorg, J.A.; Sonenshein, A.L. Bile salts and glycine as cogerminants for Clostridium difficile spores. J Bacteriol 2008, 190, 2505–2512. [Google Scholar] [CrossRef]
- Hsieh, S.H.; Brock, M. Lipid components of bile increase the protective effect of conjugated bile salts against antifungal drugs. Fungal Biol 2017, 121, 929–938. [Google Scholar] [CrossRef]
- Hsieh, S.H.; Brunke, S.; Brock, M. Encapsulation of Antifungals in Micelles Protects Candida albicans during Gall-Bladder Infection. Front Microbiol 2017, 8, 117. [Google Scholar] [CrossRef]
- Guinan, J.; Villa, P.; Thangamani, S. Secondary bile acids inhibit Candida albicans growth and morphogenesis. Pathog Dis 2018, 76. [Google Scholar] [CrossRef] [PubMed]
- Bertolini, M.; Ranjan, A.; Thompson, A.; Diaz, P.I.; Sobue, T.; Maas, K.; Dongari-Bagtzoglou, A. Candida albicans induces mucosal bacterial dysbiosis that promotes invasive infection. PLoS Pathog 2019, 15, e1007717. [Google Scholar] [CrossRef] [PubMed]
- Hiengrach, P.; Panpetch, W.; Worasilchai, N.; Chindamporn, A.; Tumwasorn, S.; Jaroonwitchawan, T.; Wilantho, A.; Chatthanathon, P.; Somboonna, N.; Leelahavanichkul, A. Administration of Candida Albicans to Dextran Sulfate Solution Treated Mice Causes Intestinal Dysbiosis, Emergence and Dissemination of Intestinal Pseudomonas Aeruginosa and Lethal Sepsis. Shock 2020, 53, 189–198. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Zhou, Y.; Wu, C.; Tang, J. Enterohemorrhagic Escherichia coli promotes the invasion and tissue damage of enterocytes infected with Candida albicans in vitro. Sci Rep 2016, 6, 37485. [Google Scholar] [CrossRef] [PubMed]
- Shrestha, R.; Sorg, J.A. Hierarchical recognition of amino acid co-germinants during Clostridioides difficile spore germination. Anaerobe 2018, 49, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Shen, A. Clostridium difficile toxins: mediators of inflammation. J Innate Immun 2012, 4, 149–158. [Google Scholar] [CrossRef] [PubMed]
- Smits, W.K.; Lyras, D.; Lacy, D.B.; Wilcox, M.H.; Kuijper, E.J. Clostridium difficile infection. Nat Rev Dis Primers 2016, 2, 16020. [Google Scholar] [CrossRef]
- Icho, S.; Ward, J.S.; Tam, J.; Kociolek, L.K.; Theriot, C.M.; Melnyk, R.A. Intestinal bile acids provide a surmountable barrier against C. difficile TcdB-induced disease pathogenesis. Proc Natl Acad Sci U S A 2023, 120, e2301252120. [Google Scholar] [CrossRef]
- Winston, J.A.; Rivera, A.J.; Cai, J.; Thanissery, R.; Montgomery, S.A.; Patterson, A.D.; Theriot, C.M. Ursodeoxycholic Acid (UDCA) Mitigates the Host Inflammatory Response during Clostridioides difficile Infection by Altering Gut Bile Acids. Infect Immun 2020, 88. [Google Scholar] [CrossRef]
- Jose, S.; Mukherjee, A.; Abhyankar, M.M.; Leng, L.; Bucala, R.; Sharma, D.; Madan, R. Neutralization of macrophage migration inhibitory factor improves host survival after Clostridium difficile infection. Anaerobe 2018, 53, 56–63. [Google Scholar] [CrossRef]
- Pike, C.M.; Tam, J.; Melnyk, R.A.; Theriot, C.M. Tauroursodeoxycholic Acid Inhibits Clostridioides difficile Toxin-Induced Apoptosis. Infect Immun 2022, 90, e0015322. [Google Scholar] [CrossRef] [PubMed]
- Winston, J.A.; Theriot, C.M. Diversification of host bile acids by members of the gut microbiota. Gut Microbes 2020, 11, 158–171. [Google Scholar] [CrossRef] [PubMed]
- Thanissery, R.; Winston, J.A.; Theriot, C.M. Inhibition of spore germination, growth, and toxin activity of clinically relevant C. difficile strains by gut microbiota derived secondary bile acids. Anaerobe 2017, 45, 86–100. [Google Scholar] [CrossRef] [PubMed]
- Winston, J.A.; Theriot, C.M. Impact of microbial derived secondary bile acids on colonization resistance against Clostridium difficile in the gastrointestinal tract. Anaerobe 2016, 41, 44–50. [Google Scholar] [CrossRef] [PubMed]
- Tam, J.; Icho, S.; Utama, E.; Orrell, K.E.; Gomez-Biagi, R.F.; Theriot, C.M.; Kroh, H.K.; Rutherford, S.A.; Lacy, D.B.; Melnyk, R.A. Intestinal bile acids directly modulate the structure and function of C. difficile TcdB toxin. Proc Natl Acad Sci U S A 2020, 117, 6792–6800. [Google Scholar] [CrossRef] [PubMed]
- Donta, I.; Karayannacos, P.E.; Boudoulas, H.; Kostakis, A.; Sechas, M.; Varonos, D.; Scalkeas, G.R. Effect of beta-adrenergic blockade on physiologic growth in the Wistar rat. Res Commun Chem Pathol Pharmacol 1982, 37, 147–150. [Google Scholar] [CrossRef]
- Zhao, C.; Wu, K.; Hao, H.; Zhao, Y.; Bao, L.; Qiu, M.; He, Y.; He, Z.; Zhang, N.; Hu, X.; et al. Gut microbiota-mediated secondary bile acid alleviates Staphylococcus aureus-induced mastitis through the TGR5-cAMP-PKA-NF-kappaB/NLRP3 pathways in mice. NPJ Biofilms Microbiomes 2023, 9, 8. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Wang, C.; Huang, X.; Yi, S.; Pan, S.; Zhang, Y.; Yuan, G.; Cao, Q.; Ye, X.; Li, H. Gut microbiota-mediated secondary bile acids regulate dendritic cells to attenuate autoimmune uveitis through TGR5 signaling. Cell Rep 2021, 36, 109726. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Guo, J.; Zhao, C.; Jiang, P.; Maimai, T.; Yanyi, L.; Cao, Y.; Fu, Y.; Zhang, N. The gut microbiota contributes to the development of Staphylococcus aureus-induced mastitis in mice. ISME J 2020, 14, 1897–1910. [Google Scholar] [CrossRef]
- Zhao, C.; Hu, X.; Bao, L.; Wu, K.; Feng, L.; Qiu, M.; Hao, H.; Fu, Y.; Zhang, N. Aryl hydrocarbon receptor activation by Lactobacillus reuteri tryptophan metabolism alleviates Escherichia coli-induced mastitis in mice. PLoS Pathog 2021, 17, e1009774. [Google Scholar] [CrossRef]
- Valat, C.; Forest, K.; Auvray, F.; Metayer, V.; Meheut, T.; Polizzi, C.; Gay, E.; Haenni, M.; Oswald, E.; Madec, J.Y. Assessment of Adhesins as an Indicator of Pathovar-Associated Virulence Factors in Bovine Escherichia coli. Appl Environ Microbiol 2014, 80, 7230–7234. [Google Scholar] [CrossRef] [PubMed]
- Vavassori, P.; Mencarelli, A.; Renga, B.; Distrutti, E.; Fiorucci, S. The bile acid receptor FXR is a modulator of intestinal innate immunity. J Immunol 2009, 183, 6251–6261. [Google Scholar] [CrossRef] [PubMed]
- Verbeke, L.; Farre, R.; Verbinnen, B.; Covens, K.; Vanuytsel, T.; Verhaegen, J.; Komuta, M.; Roskams, T.; Chatterjee, S.; Annaert, P.; et al. The FXR agonist obeticholic acid prevents gut barrier dysfunction and bacterial translocation in cholestatic rats. Am J Pathol 2015, 185, 409–419. [Google Scholar] [CrossRef] [PubMed]
- Biagioli, M.; Carino, A.; Cipriani, S.; Francisci, D.; Marchiano, S.; Scarpelli, P.; Sorcini, D.; Zampella, A.; Fiorucci, S. The Bile Acid Receptor GPBAR1 Regulates the M1/M2 Phenotype of Intestinal Macrophages and Activation of GPBAR1 Rescues Mice from Murine Colitis. J Immunol 2017, 199, 718–733. [Google Scholar] [CrossRef] [PubMed]
- Miller, W.R.; Munita, J.M.; Arias, C.A. Mechanisms of antibiotic resistance in enterococci. Expert Rev Anti Infect Ther 2014, 12, 1221–1236. [Google Scholar] [CrossRef]
- Arias, C.A.; Murray, B.E. The rise of the Enterococcus: beyond vancomycin resistance. Nat Rev Microbiol 2012, 10, 266–278. [Google Scholar] [CrossRef]
- Repoila, F.; Le Bohec, F.; Guerin, C.; Lacoux, C.; Tiwari, S.; Jaiswal, A.K.; Santana, M.P.; Kennedy, S.P.; Quinquis, B.; Rainteau, D.; et al. Adaptation of the gut pathobiont Enterococcus faecalis to deoxycholate and taurocholate bile acids. Sci Rep 2022, 12, 8485. [Google Scholar] [CrossRef]
- Paganelli, F.L.; Willems, R.J.; Leavis, H.L. Optimizing future treatment of enterococcal infections: attacking the biofilm? Trends Microbiol 2012, 20, 40–49. [Google Scholar] [CrossRef]
- McKenney, P.T.; Yan, J.; Vaubourgeix, J.; Becattini, S.; Lampen, N.; Motzer, A.; Larson, P.J.; Dannaoui, D.; Fujisawa, S.; Xavier, J.B.; et al. Intestinal Bile Acids Induce a Morphotype Switch in Vancomycin-Resistant Enterococcus that Facilitates Intestinal Colonization. Cell Host Microbe 2019, 25, 695–705. [Google Scholar] [CrossRef] [PubMed]
- Rahman, L.; Sarwar, Y.; Khaliq, S.; Inayatullah; Abbas, W.; Mobeen, A.; Ullah, A.; Hussain, S.Z.; Khan, W.S.; Kyriazi, M.E.; et al. Surfactin-Conjugated Silver Nanoparticles as an Antibacterial and Antibiofilm Agent against Pseudomonas aeruginosa. ACS Appl Mater Interfaces 2023, 15, 43321–43331. [CrossRef]
- Sanchez, L.M.; Cheng, A.T.; Warner, C.J.; Townsley, L.; Peach, K.C.; Navarro, G.; Shikuma, N.J.; Bray, W.M.; Riener, R.M.; Yildiz, F.H.; et al. Biofilm Formation and Detachment in Gram-Negative Pathogens Is Modulated by Select Bile Acids. PLoS One 2016, 11, e0149603. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, L.M.; Wong, W.R.; Riener, R.M.; Schulze, C.J.; Linington, R.G. Examining the fish microbiome: vertebrate-derived bacteria as an environmental niche for the discovery of unique marine natural products. PLoS One 2012, 7, e35398. [Google Scholar] [CrossRef] [PubMed]
- Condren, A.R.; Kahl, L.J.; Boelter, G.; Kritikos, G.; Banzhaf, M.; Dietrich, L.E.P.; Sanchez, L.M. Biofilm Inhibitor Taurolithocholic Acid Alters Colony Morphology, Specialized Metabolism, and Virulence of Pseudomonas aeruginosa. ACS Infect Dis 2020, 6, 603–612. [Google Scholar] [CrossRef] [PubMed]
- Escalante, P.; Arias-Guillen, M.; Palacios Gutierrez, J.J. New Research Strategies in Latent Tuberculosis Infection. Arch Bronconeumol (Engl Ed) 2021, 57, 151–153. [Google Scholar] [CrossRef] [PubMed]
- Sharma, M.P.; Bhatia, V. Abdominal tuberculosis. Indian J Med Res 2004, 120, 305–315. [Google Scholar]
- Epstein, D.; Mistry, K.; Whitelaw, A.; Watermeyer, G.; Pettengell, K.E. The effect of physiological concentrations of bile acids on in vitro growth of Mycobacterium tuberculosis. S Afr Med J 2012, 102, 522–524. [Google Scholar] [CrossRef]
- Merritt, M.E.; Donaldson, J.R. Effect of bile salts on the DNA and membrane integrity of enteric bacteria. J Med Microbiol 2009, 58, 1533–1541. [Google Scholar] [CrossRef]
- Sistrunk, J.R.; Nickerson, K.P.; Chanin, R.B.; Rasko, D.A.; Faherty, C.S. Survival of the Fittest: How Bacterial Pathogens Utilize Bile To Enhance Infection. Clin Microbiol Rev 2016, 29, 819–836. [Google Scholar] [CrossRef]
- Fu, T.; Wang, Y.; Ma, M.; Dai, W.; Pan, L.; Shang, Q.; Yu, G. Isolation of Alginate-Degrading Bacteria from the Human Gut Microbiota and Discovery of Bacteroides xylanisolvens AY11-1 as a Novel Anti-Colitis Probiotic Bacterium. Nutrients 2023, 15. [Google Scholar] [CrossRef]
- Yao, L.; Seaton, S.C.; Ndousse-Fetter, S.; Adhikari, A.A.; DiBenedetto, N.; Mina, A.I.; Banks, A.S.; Bry, L.; Devlin, A.S. A selective gut bacterial bile salt hydrolase alters host metabolism. Elife 2018, 7. [Google Scholar] [CrossRef]
- Li, X.; Kang, Y.; Huang, Y.; Xiao, Y.; Song, L.; Lu, S.; Ren, Z. A strain of Bacteroides thetaiotaomicron attenuates colonization of Clostridioides difficile and affects intestinal microbiota and bile acids profile in a mouse model. Biomed Pharmacother 2021, 137, 111290. [Google Scholar] [CrossRef] [PubMed]
- Yoon, S.; Yu, J.; McDowell, A.; Kim, S.H.; You, H.J.; Ko, G. Bile salt hydrolase-mediated inhibitory effect of Bacteroides ovatus on growth of Clostridium difficile. J Microbiol 2017, 55, 892–899. [Google Scholar] [CrossRef] [PubMed]
- Deng, H.; Yang, S.; Zhang, Y.; Qian, K.; Zhang, Z.; Liu, Y.; Wang, Y.; Bai, Y.; Fan, H.; Zhao, X.; et al. Bacteroides fragilis Prevents Clostridium difficile Infection in a Mouse Model by Restoring Gut Barrier and Microbiome Regulation. Front Microbiol 2018, 9, 2976. [Google Scholar] [CrossRef] [PubMed]
- Garrett, W.S. The gut microbiota and colon cancer. Science 2019, 364, 1133–1135. [Google Scholar] [CrossRef]
- Sun, X.; Chen, Z.; Yu, L.; Zeng, W.; Sun, B.; Fan, H.; Bai, Y. Bacteroides dorei BDX-01 alleviates DSS-induced experimental colitis in mice by regulating intestinal bile salt hydrolase activity and the FXR-NLRP3 signaling pathway. Front Pharmacol 2023, 14, 1205323. [Google Scholar] [CrossRef] [PubMed]
- Tiratterra, E.; Franco, P.; Porru, E.; Katsanos, K.H.; Christodoulou, D.K.; Roda, G. Role of bile acids in inflammatory bowel disease. Ann Gastroenterol 2018, 31, 266–272. [Google Scholar] [CrossRef]
- Zhen, Y.; Zhang, H. NLRP3 Inflammasome and Inflammatory Bowel Disease. Front Immunol 2019, 10, 276. [Google Scholar] [CrossRef] [PubMed]
- Thomas, J.P.; Modos, D.; Rushbrook, S.M.; Powell, N.; Korcsmaros, T. The Emerging Role of Bile Acids in the Pathogenesis of Inflammatory Bowel Disease. Front Immunol 2022, 13, 829525. [Google Scholar] [CrossRef]
- Chen, M.L.; Takeda, K.; Sundrud, M.S. Emerging roles of bile acids in mucosal immunity and inflammation. Mucosal Immunol 2019, 12, 851–861. [Google Scholar] [CrossRef]
- Mullish, B.H.; Allegretti, J.R. The contribution of bile acid metabolism to the pathogenesis of Clostridioides difficile infection. Therap Adv Gastroenterol 2021, 14, 17562848211017725. [Google Scholar] [CrossRef]
- Nusse, R.; Clevers, H. Wnt/beta-Catenin Signaling, Disease, and Emerging Therapeutic Modalities. Cell 2017, 169, 985–999. [Google Scholar] [CrossRef]
- Allegretti, J.R.; Mullish, B.H.; Kelly, C.; Fischer, M. The evolution of the use of faecal microbiota transplantation and emerging therapeutic indications. Lancet 2019, 394, 420–431. [Google Scholar] [CrossRef] [PubMed]
- Aguirre, A.M.; Yalcinkaya, N.; Wu, Q.; Swennes, A.; Tessier, M.E.; Roberts, P.; Miyajima, F.; Savidge, T.; Sorg, J.A. Bile acid-independent protection against Clostridioides difficile infection. PLoS Pathog 2021, 17, e1010015. [Google Scholar] [CrossRef] [PubMed]
- Saenz, C.; Fang, Q.; Gnanasekaran, T.; Trammell, S.A.J.; Buijink, J.A.; Pisano, P.; Wierer, M.; Moens, F.; Lengger, B.; Brejnrod, A.; et al. Clostridium scindens secretome suppresses virulence gene expression of Clostridioides difficile in a bile acid-independent manner. Microbiol Spectr 2023, 11, e0393322. [Google Scholar] [CrossRef] [PubMed]
- Aguirre, A.M.; Adegbite, A.O.; Sorg, J.A. Clostridioides difficile bile salt hydrolase activity has substrate specificity and affects biofilm formation. NPJ Biofilms Microbiomes 2022, 8, 94. [Google Scholar] [CrossRef]
- Kumar, A.; Sundaram, K.; Mu, J.; Dryden, G.W.; Sriwastva, M.K.; Lei, C.; Zhang, L.; Qiu, X.; Xu, F.; Yan, J.; et al. High-fat diet-induced upregulation of exosomal phosphatidylcholine contributes to insulin resistance. Nat Commun 2021, 12, 213. [Google Scholar] [CrossRef]
- Li, Q.; Hu, W.; Liu, W.X.; Zhao, L.Y.; Huang, D.; Liu, X.D.; Chan, H.; Zhang, Y.; Zeng, J.D.; Coker, O.O.; et al. Streptococcus thermophilus Inhibits Colorectal Tumorigenesis Through Secreting beta-Galactosidase. Gastroenterology 2021, 160, 1179–1193. [Google Scholar] [CrossRef]
- Wang, D.; Zhao, R.; Qu, Y.Y.; Mei, X.Y.; Zhang, X.; Zhou, Q.; Li, Y.; Yang, S.B.; Zuo, Z.G.; Chen, Y.M.; et al. Colonic Lysine Homocysteinylation Induced by High-Fat Diet Suppresses DNA Damage Repair. Cell Rep 2018, 25, 398–412. [Google Scholar] [CrossRef]
- Wang, D.; Fu, L.; Wei, J.; Xiong, Y.; DuBois, R.N. PPARdelta Mediates the Effect of Dietary Fat in Promoting Colorectal Cancer Metastasis. Cancer Res 2019, 79, 4480–4490. [Google Scholar] [CrossRef]
- Zhao, M.; Jiang, Z.; Cai, H.; Li, Y.; Mo, Q.; Deng, L.; Zhong, H.; Liu, T.; Zhang, H.; Kang, J.X.; et al. Modulation of the Gut Microbiota during High-Dose Glycerol Monolaurate-Mediated Amelioration of Obesity in Mice Fed a High-Fat Diet. mBio 2020, 11. [Google Scholar] [CrossRef]
- Kim, J.D.; Yoon, N.A.; Jin, S.; Diano, S. Microglial UCP2 Mediates Inflammation and Obesity Induced by High-Fat Feeding. Cell Metab 2019, 30, 952–962. [Google Scholar] [CrossRef] [PubMed]
- Tian, B.; Zhao, J.; Zhang, M.; Chen, Z.; Ma, Q.; Liu, H.; Nie, C.; Zhang, Z.; An, W.; Li, J. Lycium ruthenicum Anthocyanins Attenuate High-Fat Diet-Induced Colonic Barrier Dysfunction and Inflammation in Mice by Modulating the Gut Microbiota. Mol Nutr Food Res 2021, 65, e2000745. [Google Scholar] [CrossRef] [PubMed]
- Wei, M.; Gao, X.; Liu, L.; Li, Z.; Wan, Z.; Dong, Y.; Chen, X.; Niu, Y.; Zhang, J.; Yang, G. Visceral Adipose Tissue Derived Exosomes Exacerbate Colitis Severity via Pro-inflammatory MiRNAs in High Fat Diet Fed Mice. ACS Nano 2020, 14, 5099–5110. [Google Scholar] [CrossRef] [PubMed]
- Bisanz, J.E.; Upadhyay, V.; Turnbaugh, J.A.; Ly, K.; Turnbaugh, P.J. Meta-Analysis Reveals Reproducible Gut Microbiome Alterations in Response to a High-Fat Diet. Cell Host Microbe 2019, 26, 265–272. [Google Scholar] [CrossRef] [PubMed]
- Wan, Y.; Wang, F.; Yuan, J.; Li, J.; Jiang, D.; Zhang, J.; Li, H.; Wang, R.; Tang, J.; Huang, T.; et al. Effects of dietary fat on gut microbiota and faecal metabolites, and their relationship with cardiometabolic risk factors: a 6-month randomised controlled-feeding trial. Gut 2019, 68, 1417–1429. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Cheng, R.; Liang, H.; Miao, Z.; Wang, J.; Zhou, Q.; Chen, J.; He, F.; Shen, X. Influence of high-fat diet on host animal health via bile acid metabolism and benefits of oral-fed Streptococcus thermophilus MN-ZLW-002. Exp Anim 2022, 71, 468–480. [Google Scholar] [CrossRef] [PubMed]
- McFarland, L.V.; Surawicz, C.M.; Greenberg, R.N.; Fekety, R.; Elmer, G.W.; Moyer, K.A.; Melcher, S.A.; Bowen, K.E.; Cox, J.L.; Noorani, Z.; et al. A randomized placebo-controlled trial of Saccharomyces boulardii in combination with standard antibiotics for Clostridium difficile disease. JAMA 1994, 271, 1913–1918. [Google Scholar] [CrossRef] [PubMed]
- Surawicz, C.M.; McFarland, L.V.; Greenberg, R.N.; Rubin, M.; Fekety, R.; Mulligan, M.E.; Garcia, R.J.; Brandmarker, S.; Bowen, K.; Borjal, D.; et al. The search for a better treatment for recurrent Clostridium difficile disease: use of high-dose vancomycin combined with Saccharomyces boulardii. Clin Infect Dis 2000, 31, 1012–1017. [Google Scholar] [CrossRef]
- Kelly, C.P.; Chong Nguyen, C.; Palmieri, L.J.; Pallav, K.; Dowd, S.E.; Humbert, L.; Seksik, P.; Bado, A.; Coffin, B.; Rainteau, D.; et al. Saccharomyces boulardii CNCM I-745 Modulates the Fecal Bile Acids Metabolism During Antimicrobial Therapy in Healthy Volunteers. Front Microbiol 2019, 10, 336. [Google Scholar] [CrossRef]
- Castagliuolo, I.; LaMont, J.T.; Nikulasson, S.T.; Pothoulakis, C. Saccharomyces boulardii protease inhibits Clostridium difficile toxin A effects in the rat ileum. Infect Immun 1996, 64, 5225–5232. [Google Scholar] [CrossRef]
- Tasteyre, A.; Barc, M.C.; Karjalainen, T.; Bourlioux, P.; Collignon, A. Inhibition of in vitro cell adherence of Clostridium difficile by Saccharomyces boulardii. Microb Pathog 2002, 32, 219–225. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Kokkotou, E.G.; Mustafa, N.; Bhaskar, K.R.; Sougioultzis, S.; O'Brien, M.; Pothoulakis, C.; Kelly, C.P. Saccharomyces boulardii inhibits ERK1/2 mitogen-activated protein kinase activation both in vitro and in vivo and protects against Clostridium difficile toxin A-induced enteritis. J Biol Chem 2006, 281, 24449–24454. [Google Scholar] [CrossRef] [PubMed]
- Sorg, J.A.; Sonenshein, A.L. Inhibiting the initiation of Clostridium difficile spore germination using analogs of chenodeoxycholic acid, a bile acid. J Bacteriol 2010, 192, 4983–4990. [Google Scholar] [CrossRef] [PubMed]
- Gurung, B.; Stricklin, M.; Wang, S. Gut Microbiota-Gut Metabolites and Clostridioides difficile Infection: Approaching Sustainable Solutions for Therapy. Metabolites 2024, 14. [Google Scholar] [CrossRef]
- Weingarden, A.R.; Dosa, P.I.; DeWinter, E.; Steer, C.J.; Shaughnessy, M.K.; Johnson, J.R.; Khoruts, A.; Sadowsky, M.J. Changes in Colonic Bile Acid Composition following Fecal Microbiota Transplantation Are Sufficient to Control Clostridium difficile Germination and Growth. PLoS One 2016, 11, e0147210. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.W.; Wang, J.; Zhang, H.J.; Wu, S.G.; Qi, G.H. Supplemental Clostridium butyricum Modulates Lipid Metabolism Through Shaping Gut Microbiota and Bile Acid Profile of Aged Laying Hens. Front Microbiol 2020, 11, 600. [Google Scholar] [CrossRef]
- Seo, M.; Inoue, I.; Tanaka, M.; Matsuda, N.; Nakano, T.; Awata, T.; Katayama, S.; Alpers, D.H.; Komoda, T. Clostridium butyricum MIYAIRI 588 improves high-fat diet-induced non-alcoholic fatty liver disease in rats. Dig Dis Sci 2013, 58, 3534–3544. [Google Scholar] [CrossRef]
- Zhang, X.; Yun, Y.; Lai, Z.; Ji, S.; Yu, G.; Xie, Z.; Zhang, H.; Zhong, X.; Wang, T.; Zhang, L. Supplemental Clostridium butyricum modulates lipid metabolism by reshaping the gut microbiota composition and bile acid profile in IUGR suckling piglets. J Anim Sci Biotechnol 2023, 14, 36. [Google Scholar] [CrossRef]
- Bergogne-Berezin, E.; Towner, K.J. Acinetobacter spp. as nosocomial pathogens: microbiological, clinical, and epidemiological features. Clin Microbiol Rev 1996, 9, 148–165. [Google Scholar] [CrossRef]
- Haenni, M.; Lupo, A.; Madec, J.Y. Antimicrobial Resistance in Streptococcus spp. Microbiol Spectr 2018, 6. [Google Scholar] [CrossRef]
- Torres, C.; Alonso, C.A.; Ruiz-Ripa, L.; Leon-Sampedro, R.; Del Campo, R.; Coque, T.M. Antimicrobial Resistance in Enterococcus spp. of animal origin. Microbiol Spectr 2018, 6. [Google Scholar] [CrossRef] [PubMed]
- Ren, D.; Pichichero, M.E. Vaccine targets against Moraxella catarrhalis. Expert Opin Ther Targets 2016, 20, 19–33. [Google Scholar] [CrossRef] [PubMed]
- Fatahi-Bafghi, M. Characterization of the Rothia spp. and their role in human clinical infections. Infect Genet Evol 2021, 93, 104877. [Google Scholar] [CrossRef] [PubMed]
- Panzitt, K.; Wagner, M. FXR in liver physiology: Multiple faces to regulate liver metabolism. Biochim Biophys Acta Mol Basis Dis 2021, 1867, 166133. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Guo, Y.; Guo, S.; Tan, J. Effects of Clostridium butyricum and Enterococcus faecium on growth performance, lipid metabolism, and cecal microbiota of broiler chickens. Appl Microbiol Biotechnol 2013, 97, 6477–6488. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.C.; Kuo, C.Y.; Tzang, B.S.; Chen, H.M.; Kao, S.H. IL-6 augmented motility of airway epithelial cell BEAS-2B via Akt/GSK-3beta signaling pathway. J Cell Biochem 2012, 113, 3567–3575. [Google Scholar] [CrossRef]
- Liao, J.; Liu, Y.; Yao, Y.; Zhang, J.; Wang, H.; Zhao, J.; Chen, W.; Lu, W. Clostridium butyricum Strain CCFM1299 Reduces Obesity via Increasing Energy Expenditure and Modulating Host Bile Acid Metabolism. Nutrients 2023, 15. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Liang, X.; Wang, K.; Lin, J.; Wang, X.; Wang, P.; Zhang, Y.; Nie, Q.; Liu, H.; Zhang, Z.; et al. Intestinal hypoxia-inducible factor 2alpha regulates lactate levels to shape the gut microbiome and alter thermogenesis. Cell Metab 2021, 33, 1988–2003. [Google Scholar] [CrossRef]
- Wang, K.; Liao, M.; Zhou, N.; Bao, L.; Ma, K.; Zheng, Z.; Wang, Y.; Liu, C.; Wang, W.; Wang, J.; et al. Parabacteroides distasonis Alleviates Obesity and Metabolic Dysfunctions via Production of Succinate and Secondary Bile Acids. Cell Rep 2019, 26, 222–235. [Google Scholar] [CrossRef]
- Zheng, X.; Chen, T.; Jiang, R.; Zhao, A.; Wu, Q.; Kuang, J.; Sun, D.; Ren, Z.; Li, M.; Zhao, M.; et al. Hyocholic acid species improve glucose homeostasis through a distinct TGR5 and FXR signaling mechanism. Cell Metab 2021, 33, 791–803. [Google Scholar] [CrossRef]
- Li, Z.; Chen, M.; Zhang, R.; Wang, Z.; He, H.; Wan, Z.; Li, H.; Cai, H.; Chen, Z.; Li, M.; et al. Clostridium butyricum Ameliorates the Effect of Coprophagy Prevention on Hepatic Lipid Synthesis in Rabbits via the Gut-Liver Axis. Int J Mol Sci 2023, 24. [Google Scholar] [CrossRef] [PubMed]
- Huang, P.; Cui, X.; Wang, Z.; Xiao, C.; Ji, Q.; Wei, Q.; Huang, Y.; Bao, G.; Liu, Y. Effects of Clostridium butyricum and a Bacteriophage Cocktail on Growth Performance, Serum Biochemistry, Digestive Enzyme Activities, Intestinal Morphology, Immune Responses, and the Intestinal Microbiota in Rabbits. Antibiotics (Basel) 2021, 10. [Google Scholar] [CrossRef] [PubMed]
- Xu, Q.; Gu, S.; Chen, Y.; Quan, J.; Lv, L.; Chen, D.; Zheng, B.; Xu, L.; Li, L. Protective Effect of Pediococcus pentosaceus LI05 Against Clostridium difficile Infection in a Mouse Model. Front Microbiol 2018, 9, 2396. [Google Scholar] [CrossRef] [PubMed]
- Antharam, V.C.; McEwen, D.C.; Garrett, T.J.; Dossey, A.T.; Li, E.C.; Kozlov, A.N.; Mesbah, Z.; Wang, G.P. An Integrated Metabolomic and Microbiome Analysis Identified Specific Gut Microbiota Associated with Fecal Cholesterol and Coprostanol in Clostridium difficile Infection. PLoS One 2016, 11, e0148824. [Google Scholar] [CrossRef] [PubMed]
- Fickert, P.; Pollheimer, M.J.; Beuers, U.; Lackner, C.; Hirschfield, G.; Housset, C.; Keitel, V.; Schramm, C.; Marschall, H.U.; Karlsen, T.H.; et al. Characterization of animal models for primary sclerosing cholangitis (PSC). J Hepatol 2014, 60, 1290–1303. [Google Scholar] [CrossRef] [PubMed]
- Han, S.; Wang, K.; Shen, J.; Xia, H.; Lu, Y.; Zhuge, A.; Li, S.; Qiu, B.; Zhang, S.; Dong, X.; et al. Probiotic Pediococcus pentosaceus Li05 Improves Cholestasis through the FXR-SHP and FXR-FGF15 Pathways. Nutrients 2023, 15. [Google Scholar] [CrossRef]
- Bui, T.P.N.; Manneras-Holm, L.; Puschmann, R.; Wu, H.; Troise, A.D.; Nijsse, B.; Boeren, S.; Backhed, F.; Fiedler, D.; deVos, W.M. Conversion of dietary inositol into propionate and acetate by commensal Anaerostipes associates with host health. Nat Commun 2021, 12, 4798. [Google Scholar] [CrossRef] [PubMed]
- Chiang, J.Y. Bile acids: regulation of synthesis. J Lipid Res 2009, 50, 1955–1966. [Google Scholar] [CrossRef]
- Jones, B.V.; Begley, M.; Hill, C.; Gahan, C.G.; Marchesi, J.R. Functional and comparative metagenomic analysis of bile salt hydrolase activity in the human gut microbiome. Proc Natl Acad Sci U S A 2008, 105, 13580–13585. [Google Scholar] [CrossRef]
- Deng, L.; Liu, L.; Fu, T.; Li, C.; Jin, N.; Zhang, H.; Li, C.; Liu, Y.; Zhao, C. Genome Sequence and Evaluation of Safety and Probiotic Potential of Lactiplantibacillus plantarum LPJZ-658. Microorganisms 2023, 11. [Google Scholar] [CrossRef]
- Zhao, D.; Cao, J.; Jin, H.; Shan, Y.; Fang, J.; Liu, F. Beneficial impacts of fermented celery (Apium graveolens L.) juice on obesity prevention and gut microbiota modulation in high-fat diet fed mice. Food Funct 2021, 12, 9151–9164. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Hui, S.; Lang, H.; Zhou, M.; Zhang, Y.; Kang, C.; Zeng, X.; Zhang, Q.; Yi, L.; Mi, M. SIRT3 Deficiency Promotes High-Fat Diet-Induced Nonalcoholic Fatty Liver Disease in Correlation with Impaired Intestinal Permeability through Gut Microbial Dysbiosis. Mol Nutr Food Res 2019, 63, e1800612. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Berardi, M.; Li, W.; Farzan, M.; Dormitzer, P.R.; Harrison, S.C. Conformational states of the severe acute respiratory syndrome coronavirus spike protein ectodomain. J Virol 2006, 80, 6794–6800. [Google Scholar] [CrossRef]
- Hoffmann, M.; Kleine-Weber, H.; Schroeder, S.; Kruger, N.; Herrler, T.; Erichsen, S.; Schiergens, T.S.; Herrler, G.; Wu, N.H.; Nitsche, A.; et al. SARS-CoV-2 Cell Entry Depends on ACE2 and TMPRSS2 and Is Blocked by a Clinically Proven Protease Inhibitor. Cell 2020, 181, 271–280. [Google Scholar] [CrossRef] [PubMed]
- Zhou, P.; Yang, X.L.; Wang, X.G.; Hu, B.; Zhang, L.; Zhang, W.; Si, H.R.; Zhu, Y.; Li, B.; Huang, C.L.; et al. A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature 2020, 579, 270–273. [Google Scholar] [CrossRef] [PubMed]
- Thuy, P.X.; Bao, T.D.D.; Moon, E.Y. Ursodeoxycholic acid ameliorates cell migration retarded by the SARS-CoV-2 spike protein in BEAS-2B human bronchial epithelial cells. Biomed Pharmacother 2022, 150, 113021. [Google Scholar] [CrossRef] [PubMed]
- Rodal Canales, F.J.; Perez-Campos Mayoral, L.; Hernandez-Huerta, M.T.; Sanchez Navarro, L.M.; Matias-Cervantes, C.A.; Martinez Cruz, M.; Cruz Parada, E.; Zenteno, E.; Ramos-Martinez, E.G.; Perez-Campos Mayoral, E.; et al. Interaction of Spike protein and lipid membrane of SARS-CoV-2 with Ursodeoxycholic acid, an in-silico analysis. Sci Rep 2021, 11, 22288. [Google Scholar] [CrossRef]
- Carino, A.; Moraca, F.; Fiorillo, B.; Marchiano, S.; Sepe, V.; Biagioli, M.; Finamore, C.; Bozza, S.; Francisci, D.; Distrutti, E.; et al. Hijacking SARS-CoV-2/ACE2 Receptor Interaction by Natural and Semi-synthetic Steroidal Agents Acting on Functional Pockets on the Receptor Binding Domain. Front Chem 2020, 8, 572885. [Google Scholar] [CrossRef]
- Yadav, R.; Choudhury, C.; Kumar, Y.; Bhatia, A. Virtual repurposing of ursodeoxycholate and chenodeoxycholate as lead candidates against SARS-Cov2-Envelope protein: A molecular dynamics investigation. J Biomol Struct Dyn 2022, 40, 5147–5158. [Google Scholar] [CrossRef]
- Subramanian, S.; Iles, T.; Ikramuddin, S.; Steer, C.J. Merit of an Ursodeoxycholic Acid Clinical Trial in COVID-19 Patients. Vaccines (Basel) 2020, 8. [Google Scholar] [CrossRef]
- Kim, Y.J.; Jeong, S.H.; Kim, E.K.; Kim, E.J.; Cho, J.H. Ursodeoxycholic acid suppresses epithelial-mesenchymal transition and cancer stem cell formation by reducing the levels of peroxiredoxin II and reactive oxygen species in pancreatic cancer cells. Oncol Rep 2017, 38, 3632–3638. [Google Scholar] [CrossRef] [PubMed]
- Lapenna, D.; Ciofani, G.; Festi, D.; Neri, M.; Pierdomenico, S.D.; Giamberardino, M.A.; Cuccurullo, F. Antioxidant properties of ursodeoxycholic acid. Biochem Pharmacol 2002, 64, 1661–1667. [Google Scholar] [CrossRef] [PubMed]
- Talebian, R.; Panahipour, L.; Gruber, R. Ursodeoxycholic acid attenuates the expression of proinflammatory cytokines in periodontal cells. J Periodontol 2020, 91, 1098–1104. [Google Scholar] [CrossRef] [PubMed]
- Ko, W.K.; Lee, S.H.; Kim, S.J.; Jo, M.J.; Kumar, H.; Han, I.B.; Sohn, S. Anti-inflammatory effects of ursodeoxycholic acid by lipopolysaccharide-stimulated inflammatory responses in RAW 264.7 macrophages. PLoS One 2017, 12, e0180673. [Google Scholar] [CrossRef] [PubMed]
- Ko, W.K.; Kim, S.J.; Jo, M.J.; Choi, H.; Lee, D.; Kwon, I.K.; Lee, S.H.; Han, I.B.; Sohn, S. Ursodeoxycholic Acid Inhibits Inflammatory Responses and Promotes Functional Recovery After Spinal Cord Injury in Rats. Mol Neurobiol 2019, 56, 267–277. [Google Scholar] [CrossRef] [PubMed]
- Willart, M.A.; van Nimwegen, M.; Grefhorst, A.; Hammad, H.; Moons, L.; Hoogsteden, H.C.; Lambrecht, B.N.; Kleinjan, A. Ursodeoxycholic acid suppresses eosinophilic airway inflammation by inhibiting the function of dendritic cells through the nuclear farnesoid X receptor. Allergy 2012, 67, 1501–1510. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.K.; Cho, J.H.; Kim, E.; Kim, Y.J. Ursodeoxycholic acid inhibits the proliferation of colon cancer cells by regulating oxidative stress and cancer stem-like cell growth. PLoS One 2017, 12, e0181183. [Google Scholar] [CrossRef]
- Brevini, T.; Maes, M.; Webb, G.J.; John, B.V.; Fuchs, C.D.; Buescher, G.; Wang, L.; Griffiths, C.; Brown, M.L.; Scott, W.E. , 3rd, et al. FXR inhibition may protect from SARS-CoV-2 infection by reducing ACE2. Nature 2023, 615, 134–142. [Google Scholar] [CrossRef]
- Kumar, Y.; Yadav, R.; Bhatia, A. Can natural detergent properties of bile acids be used beneficially in tackling coronavirus disease-19? Future Virology 2020, 15, 779–782. [Google Scholar] [CrossRef]
- Abdulrab, S.; Al-Maweri, S.; Halboub, E. Ursodeoxycholic acid as a candidate therapeutic to alleviate and/or prevent COVID-19-associated cytokine storm. Med Hypotheses 2020, 143, 109897. [Google Scholar] [CrossRef]
- Xiao, X.; Chen, H.; Yang, L.; Xie, G.; Shimuzu, R.; Murai, A. Concise review: Cancer cell reprogramming and therapeutic implications. Transl Oncol 2022, 24, 101503. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Wang, J.S. Ursodeoxycholic acid administration did not reduce susceptibility to SARS-CoV-2 infection in children. Liver Int 2023, 43, 1950–1954. [Google Scholar] [CrossRef]
- Liu, L.; Aigner, A.; Schmid, R.D. Identification, cloning, heterologous expression, and characterization of a NADPH-dependent 7beta-hydroxysteroid dehydrogenase from Collinsella aerofaciens. Appl Microbiol Biotechnol 2011, 90, 127–135. [Google Scholar] [CrossRef] [PubMed]
- Hirayama, M.; Nishiwaki, H.; Hamaguchi, T.; Ito, M.; Ueyama, J.; Maeda, T.; Kashihara, K.; Tsuboi, Y.; Ohno, K. Intestinal Collinsella may mitigate infection and exacerbation of COVID-19 by producing ursodeoxycholate. PLoS One 2021, 16, e0260451. [Google Scholar] [CrossRef] [PubMed]
- Luo, L.; Han, W.; Du, J.; Yang, X.; Duan, M.; Xu, C.; Zeng, Z.; Chen, W.; Chen, J. Chenodeoxycholic Acid from Bile Inhibits Influenza A Virus Replication via Blocking Nuclear Export of Viral Ribonucleoprotein Complexes. Molecules 2018, 23. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Hu, X.; Zhang, Q.; Zhao, L.; Sun, X.; Yang, L.; Jin, M. Sodium taurocholate hydrate inhibits influenza virus replication and suppresses influenza a Virus-triggered inflammation in vitro and in vivo. Int Immunopharmacol 2023, 122, 110544. [Google Scholar] [CrossRef]
- Kumar, N.; Xin, Z.T.; Liang, Y.; Ly, H.; Liang, Y. NF-kappaB signaling differentially regulates influenza virus RNA synthesis. J Virol 2008, 82, 9880–9889. [Google Scholar] [CrossRef]
- Chang, K.O.; Sosnovtsev, S.V.; Belliot, G.; Kim, Y.; Saif, L.J.; Green, K.Y. Bile acids are essential for porcine enteric calicivirus replication in association with down-regulation of signal transducer and activator of transcription 1. Proc Natl Acad Sci U S A 2004, 101, 8733–8738. [Google Scholar] [CrossRef] [PubMed]
- Ettayebi, K.; Crawford, S.E.; Murakami, K.; Broughman, J.R.; Karandikar, U.; Tenge, V.R.; Neill, F.H.; Blutt, S.E.; Zeng, X.L.; Qu, L.; et al. Replication of human noroviruses in stem cell-derived human enteroids. Science 2016, 353, 1387–1393. [Google Scholar] [CrossRef]
- Nelson, C.A.; Wilen, C.B.; Dai, Y.N.; Orchard, R.C.; Kim, A.S.; Stegeman, R.A.; Hsieh, L.L.; Smith, T.J.; Virgin, H.W.; Fremont, D.H. Structural basis for murine norovirus engagement of bile acids and the CD300lf receptor. Proc Natl Acad Sci U S A 2018, 115, E9201–E9210. [Google Scholar] [CrossRef]
- Grau, K.R.; Zhu, S.; Peterson, S.T.; Helm, E.W.; Philip, D.; Phillips, M.; Hernandez, A.; Turula, H.; Frasse, P.; Graziano, V.R.; et al. The intestinal regionalization of acute norovirus infection is regulated by the microbiota via bile acid-mediated priming of type III interferon. Nat Microbiol 2020, 5, 84–92. [Google Scholar] [CrossRef] [PubMed]
- Baldridge, M.T.; Nice, T.J.; McCune, B.T.; Yokoyama, C.C.; Kambal, A.; Wheadon, M.; Diamond, M.S.; Ivanova, Y.; Artyomov, M.; Virgin, H.W. Commensal microbes and interferon-lambda determine persistence of enteric murine norovirus infection. Science 2015, 347, 266–269. [Google Scholar] [CrossRef] [PubMed]
- Yan, H.; Peng, B.; Liu, Y.; Xu, G.; He, W.; Ren, B.; Jing, Z.; Sui, J.; Li, W. Viral entry of hepatitis B and D viruses and bile salts transportation share common molecular determinants on sodium taurocholate cotransporting polypeptide. J Virol 2014, 88, 3273–3284. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Interactions Between BAs and Candida albicans.SBA:(A) Inhibit the morphological transformation of Candida albicans; (B) Direct antimicrobial activity against Candida albicans; (C) Inhibit the growth of Clostridioides difficile; (D) Increased the abundance of Firmicutes. PBA:(E) Inhibit the production of immune active substances angiogenin-4 and CX3CR1; (F) Reduce the tight junction proteins in the intestine. PBA: Primary bile acid; SBA: Secondary bile acid.
Figure 1.
Interactions Between BAs and Candida albicans.SBA:(A) Inhibit the morphological transformation of Candida albicans; (B) Direct antimicrobial activity against Candida albicans; (C) Inhibit the growth of Clostridioides difficile; (D) Increased the abundance of Firmicutes. PBA:(E) Inhibit the production of immune active substances angiogenin-4 and CX3CR1; (F) Reduce the tight junction proteins in the intestine. PBA: Primary bile acid; SBA: Secondary bile acid.
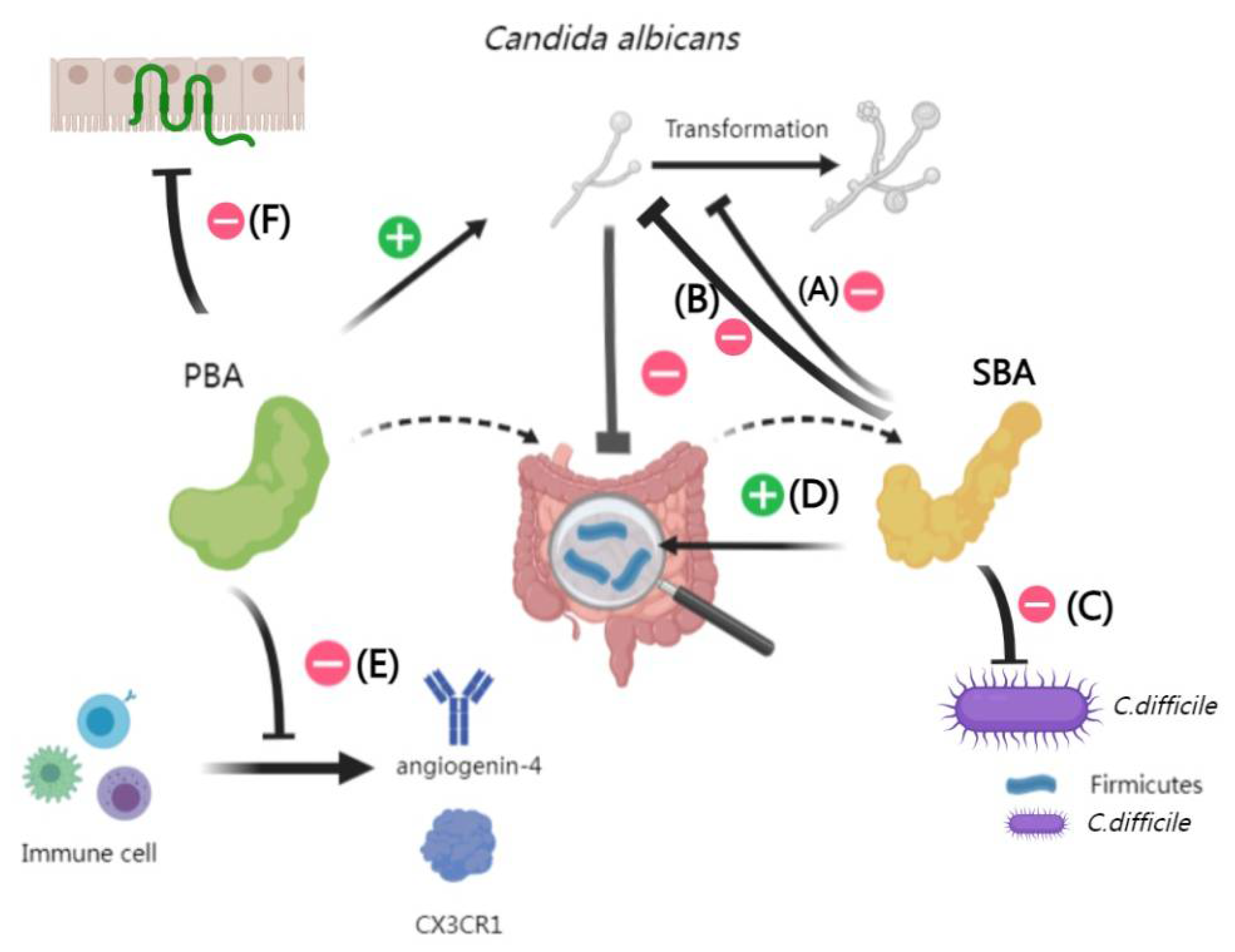
Figure 2.
Interactions Between BAs and Clostridioides difficile. SBA: (A) Binding to FXR and TGR5 receptors, activating NF-κB and other signaling pathways, enhancing innate immunity, and inhibiting the growth of Clostridioides difficile; (B) Directly interacting with the C-terminus of toxin TcdB, inducing toxin structural changes, and preventing toxin binding with host cells. PBA: (C) Promote the spore germination of Clostridioides difficile; (D) Promote the release of Clostridioides difficile toxins TcdA and TcdB. PBA: Primary bile acid; SBA: Secondary bile acid; FXR: Farnesoid X Receptor; TGR5: Takeda G Protein-Coupled Receptor 5.
Figure 2.
Interactions Between BAs and Clostridioides difficile. SBA: (A) Binding to FXR and TGR5 receptors, activating NF-κB and other signaling pathways, enhancing innate immunity, and inhibiting the growth of Clostridioides difficile; (B) Directly interacting with the C-terminus of toxin TcdB, inducing toxin structural changes, and preventing toxin binding with host cells. PBA: (C) Promote the spore germination of Clostridioides difficile; (D) Promote the release of Clostridioides difficile toxins TcdA and TcdB. PBA: Primary bile acid; SBA: Secondary bile acid; FXR: Farnesoid X Receptor; TGR5: Takeda G Protein-Coupled Receptor 5.
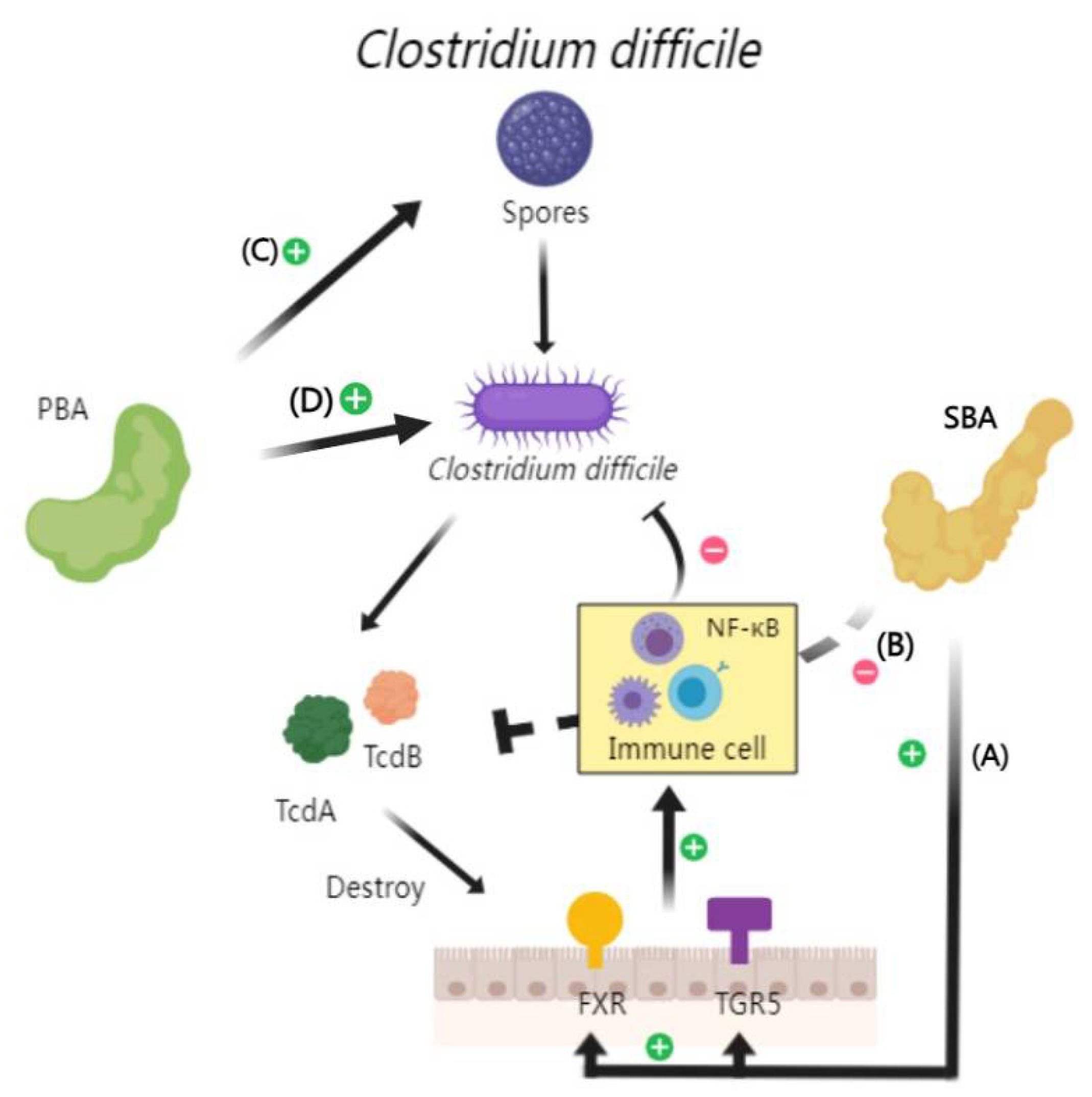
Figure 3.
Interactions Between BAs and Staphylococcus aureus. SBA:(A) Inhibit infection through the TGR5/PKA—NF-κB and NLRP3 signaling pathways; (B) Repair tight junction proteins in the intestinal epithelium; (C) Reduce the expression of inflammatory markers; (D) Promote the release of Clostridioides difficile toxins TcdA and TcdB; (E) Cannot directly inhibit the growth of Staphylococcus aureus. SBA: Secondary bile acid; TGR5: Takeda G Protein-Coupled Receptor 5.
Figure 3.
Interactions Between BAs and Staphylococcus aureus. SBA:(A) Inhibit infection through the TGR5/PKA—NF-κB and NLRP3 signaling pathways; (B) Repair tight junction proteins in the intestinal epithelium; (C) Reduce the expression of inflammatory markers; (D) Promote the release of Clostridioides difficile toxins TcdA and TcdB; (E) Cannot directly inhibit the growth of Staphylococcus aureus. SBA: Secondary bile acid; TGR5: Takeda G Protein-Coupled Receptor 5.
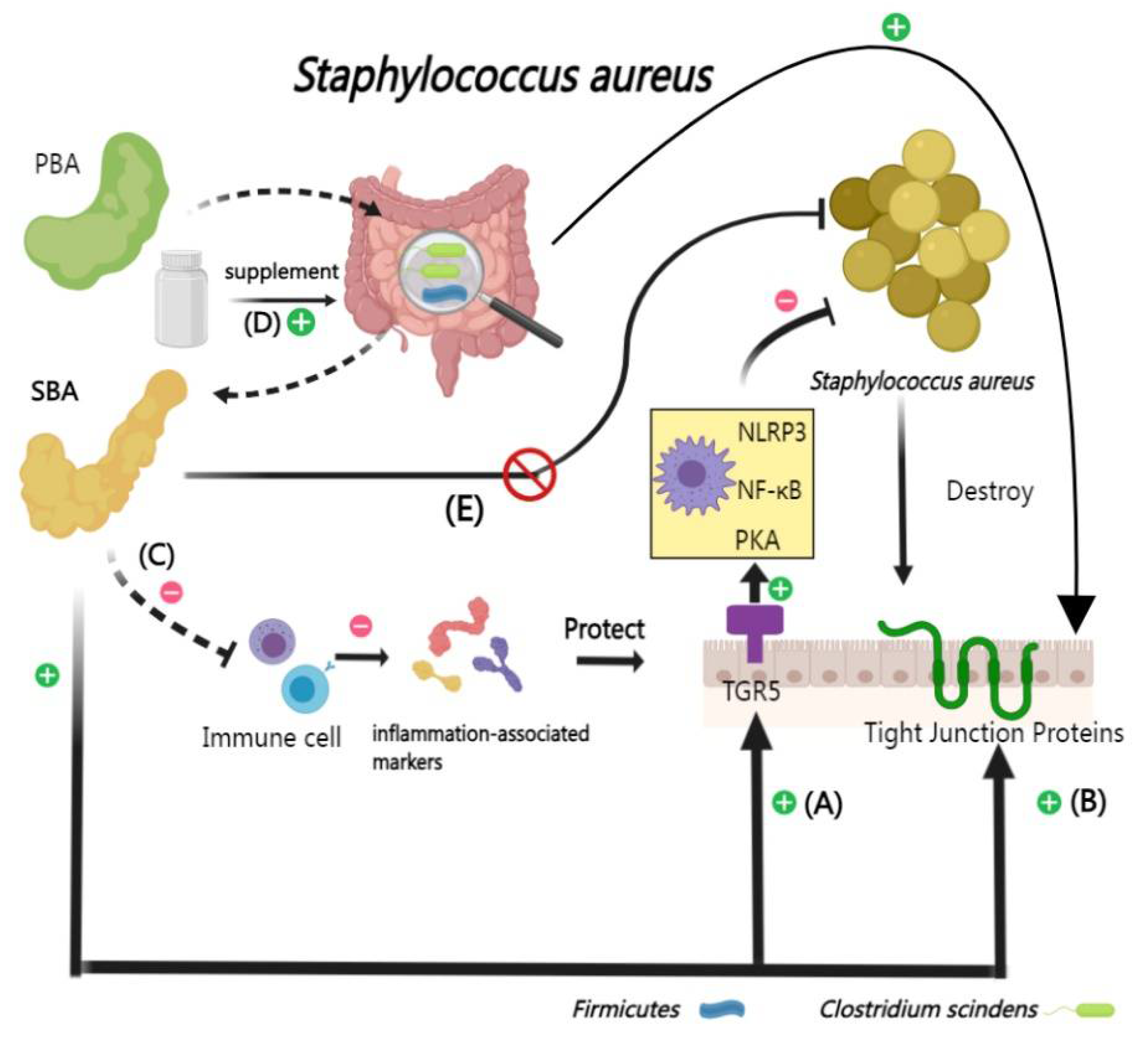
Figure 4.
Interactions Between BAs and Extended-Spectrum Beta-Lactamase-Resistant Escherichia coli. SBA:(A) By optimizing the structure of the intestinal microbiota; (B) Resist infection by promoting the production of SCFA; (C) Increase TGR5 transcription, activate the NF-κB signaling pathway, enhance innate immunity, and strengthen the intestinal barrier. SBA: Secondary bile acid; SCFA: short-chain fatty acids; TGR5: Takeda G Protein-Coupled Receptor 5.
Figure 4.
Interactions Between BAs and Extended-Spectrum Beta-Lactamase-Resistant Escherichia coli. SBA:(A) By optimizing the structure of the intestinal microbiota; (B) Resist infection by promoting the production of SCFA; (C) Increase TGR5 transcription, activate the NF-κB signaling pathway, enhance innate immunity, and strengthen the intestinal barrier. SBA: Secondary bile acid; SCFA: short-chain fatty acids; TGR5: Takeda G Protein-Coupled Receptor 5.
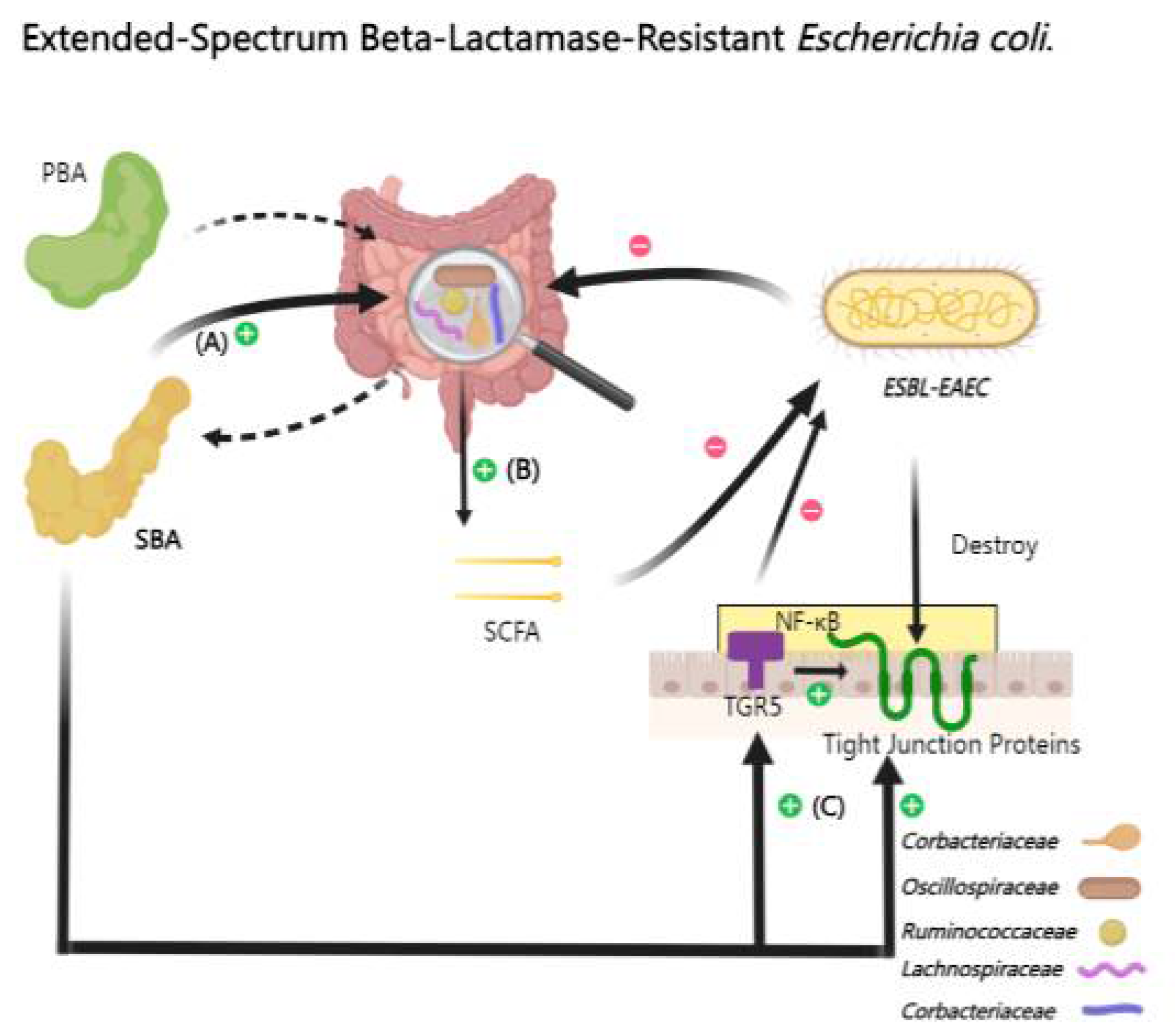
Figure 5.
Interactions Between BAs and Enterococci. SBA:(A) Inhibit the expression of ribosomal protein genes, suppress the growth of Enterococcus faecium; (B) Maintain VRE in a diplococcal state, and inhibit the morphological transformation of VRE; (C) Inhibit the formation of VRE biofilms. SBA: Secondary bile acid; VRE: Vancomycin-resistant enterococci.
Figure 5.
Interactions Between BAs and Enterococci. SBA:(A) Inhibit the expression of ribosomal protein genes, suppress the growth of Enterococcus faecium; (B) Maintain VRE in a diplococcal state, and inhibit the morphological transformation of VRE; (C) Inhibit the formation of VRE biofilms. SBA: Secondary bile acid; VRE: Vancomycin-resistant enterococci.
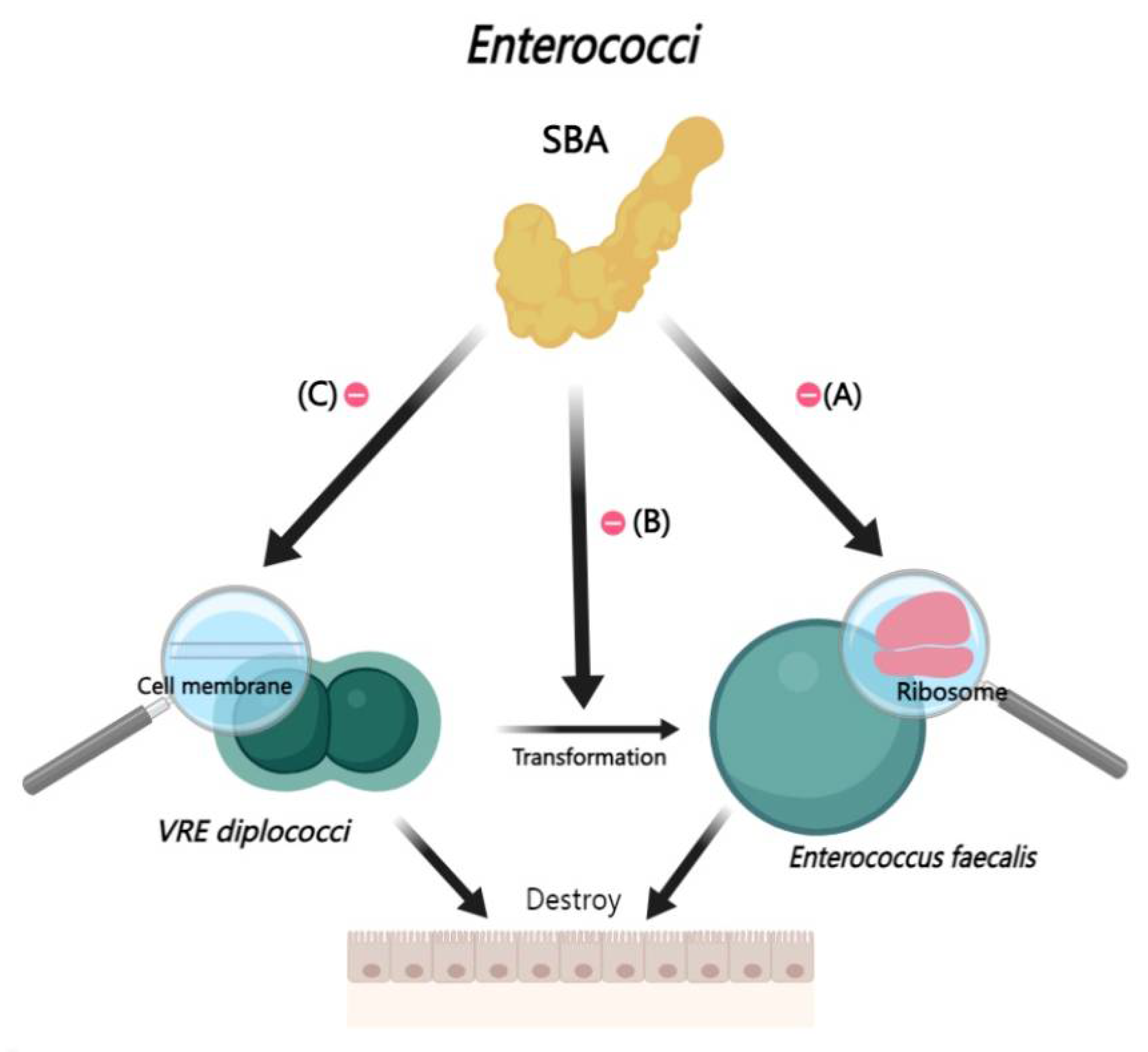
Figure 6.
Interactions Between BAs and Other Pathogenic Bacteria. PBA:(A) Regulate the synthesis of virulence-related metabolites, such as the iron chelator pyochelin, thereby affecting Pseudomonas aeruginosa’s toxicity and inhibit its biofilm formation. SBA:(B) Disrupt the cell wall of lipid-rich Mycobacterium tuberculosis.; (C) Suppression of Infections by Escherichia coli, Salmonella enterica serovar Typhimurium, Bacillus cereus, and Listeria monocytogenes through the Induction of DNA Repair Damage and Transcription of Recombination-Related Genes. PBA: Primary bile acid; SBA: Secondary bile acid.
Figure 6.
Interactions Between BAs and Other Pathogenic Bacteria. PBA:(A) Regulate the synthesis of virulence-related metabolites, such as the iron chelator pyochelin, thereby affecting Pseudomonas aeruginosa’s toxicity and inhibit its biofilm formation. SBA:(B) Disrupt the cell wall of lipid-rich Mycobacterium tuberculosis.; (C) Suppression of Infections by Escherichia coli, Salmonella enterica serovar Typhimurium, Bacillus cereus, and Listeria monocytogenes through the Induction of DNA Repair Damage and Transcription of Recombination-Related Genes. PBA: Primary bile acid; SBA: Secondary bile acid.
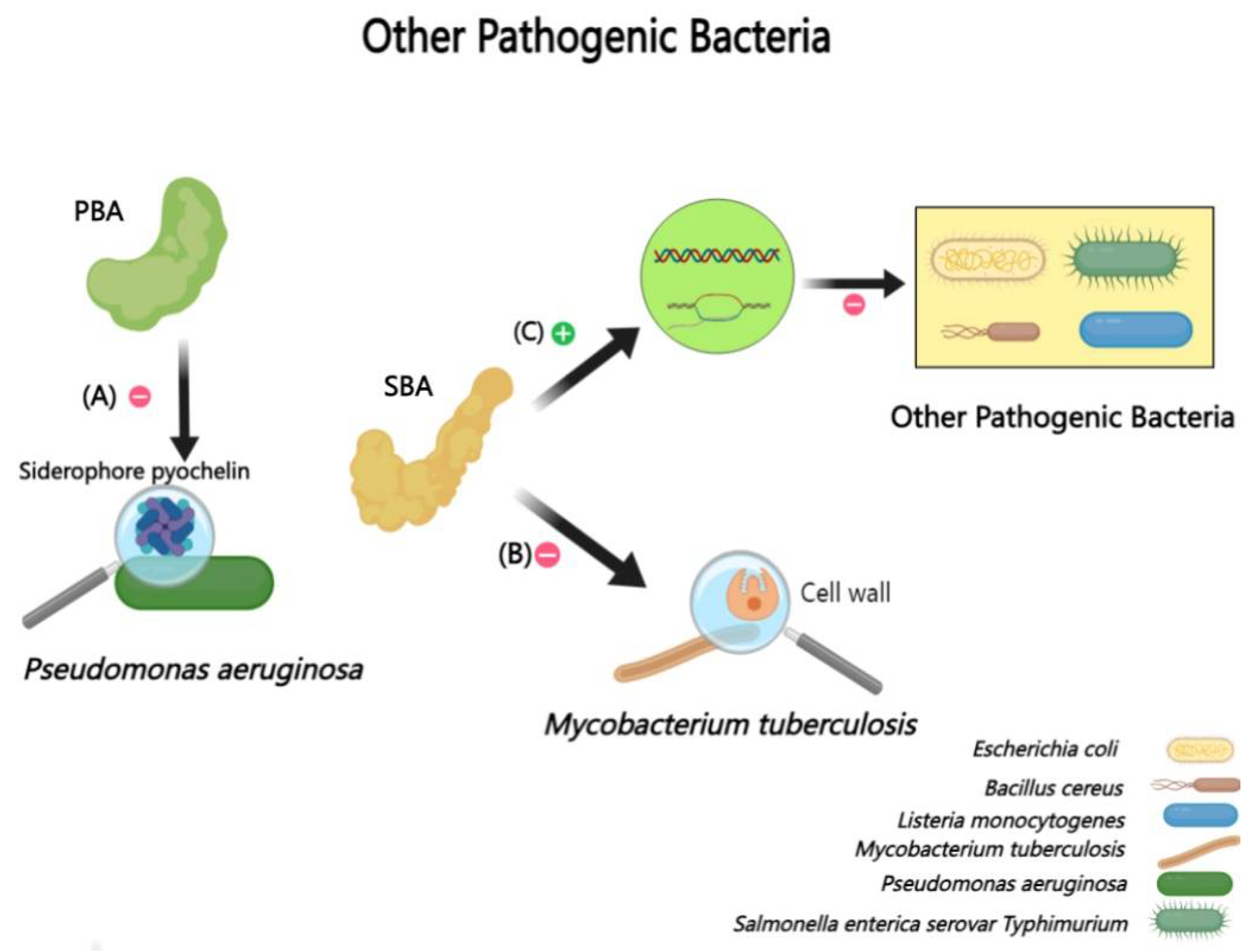
Figure 7.
Interactions Between BAs and Bacteroidetes. (A) Bacteroidetes exhibit BSHs activity, facilitate SBA production, and inhibit Clostridioides difficile infection; (B) Bacteroides with high BSH gene expression will promote the massive production of SBA, which can induce colorectal cancer; (C) BDX-01 enhances intestinal health by modulating bile acid metabolism and the FXR-NLRP3 signaling pathway, leading to lower proinflammatory cytokine levels and reduced IL-1β in the colon, thus mitigating experimental colitis. SBA: Secondary bile acid; BSHs: Bile salt hydrolases; FXR: Farnesoid X Receptor;.
Figure 7.
Interactions Between BAs and Bacteroidetes. (A) Bacteroidetes exhibit BSHs activity, facilitate SBA production, and inhibit Clostridioides difficile infection; (B) Bacteroides with high BSH gene expression will promote the massive production of SBA, which can induce colorectal cancer; (C) BDX-01 enhances intestinal health by modulating bile acid metabolism and the FXR-NLRP3 signaling pathway, leading to lower proinflammatory cytokine levels and reduced IL-1β in the colon, thus mitigating experimental colitis. SBA: Secondary bile acid; BSHs: Bile salt hydrolases; FXR: Farnesoid X Receptor;.
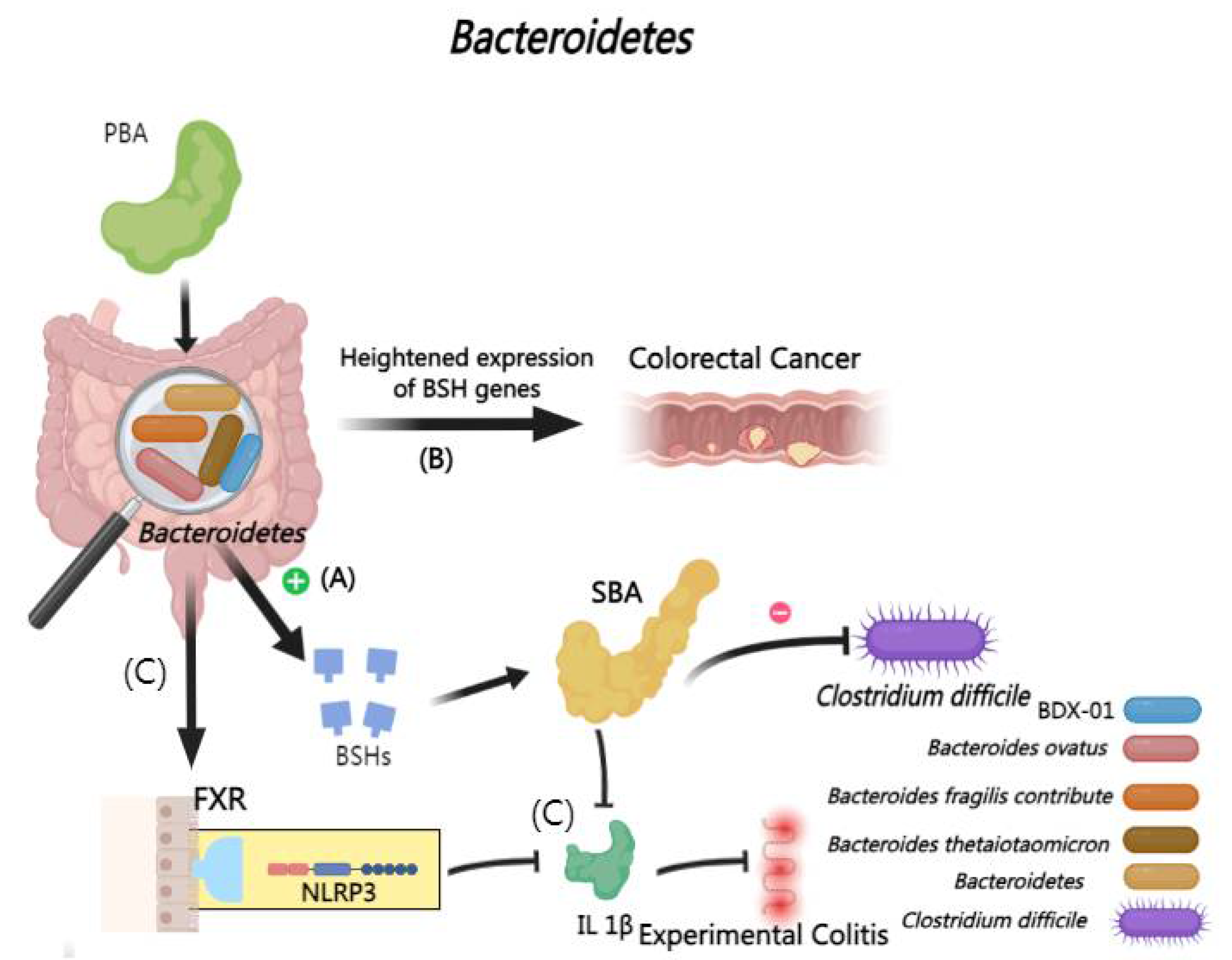
Figure 8.
Interactions Between BAs and Clostridium scindens. (A) Clostridium scindens exhibit BSHs and 7α-dehydroxylase activity, facilitate SBA production, and inhibit Clostridium difficile infection; (B) Clostridium scindens inhibit the toxin production of Clostridium difficile; (C) Clostridium difficile also inhibits the formation of Clostridium scindens biofilms. SBA: Secondary bile acids; BSHs: Bile salt hydrolases.
Figure 8.
Interactions Between BAs and Clostridium scindens. (A) Clostridium scindens exhibit BSHs and 7α-dehydroxylase activity, facilitate SBA production, and inhibit Clostridium difficile infection; (B) Clostridium scindens inhibit the toxin production of Clostridium difficile; (C) Clostridium difficile also inhibits the formation of Clostridium scindens biofilms. SBA: Secondary bile acids; BSHs: Bile salt hydrolases.
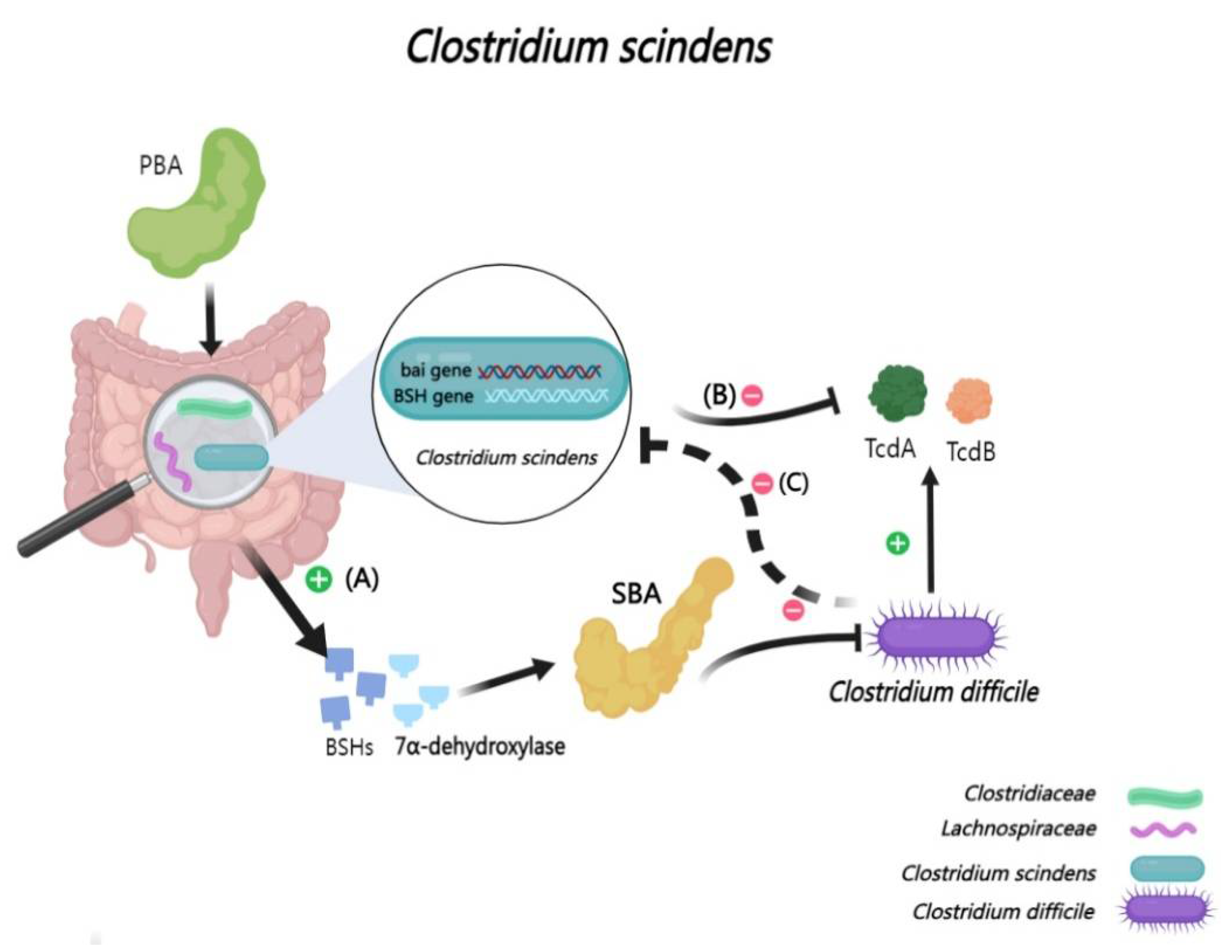
Figure 9.
Interactions Between BAs and Streptococcus thermophilus. (A) Streptococcus thermophilus can regulate the structure of the intestinal microbiota, increase the bacterial populations that produce SBA, and modulate the bile acid spectrum. SBA Secondary bile acid.
Figure 9.
Interactions Between BAs and Streptococcus thermophilus. (A) Streptococcus thermophilus can regulate the structure of the intestinal microbiota, increase the bacterial populations that produce SBA, and modulate the bile acid spectrum. SBA Secondary bile acid.
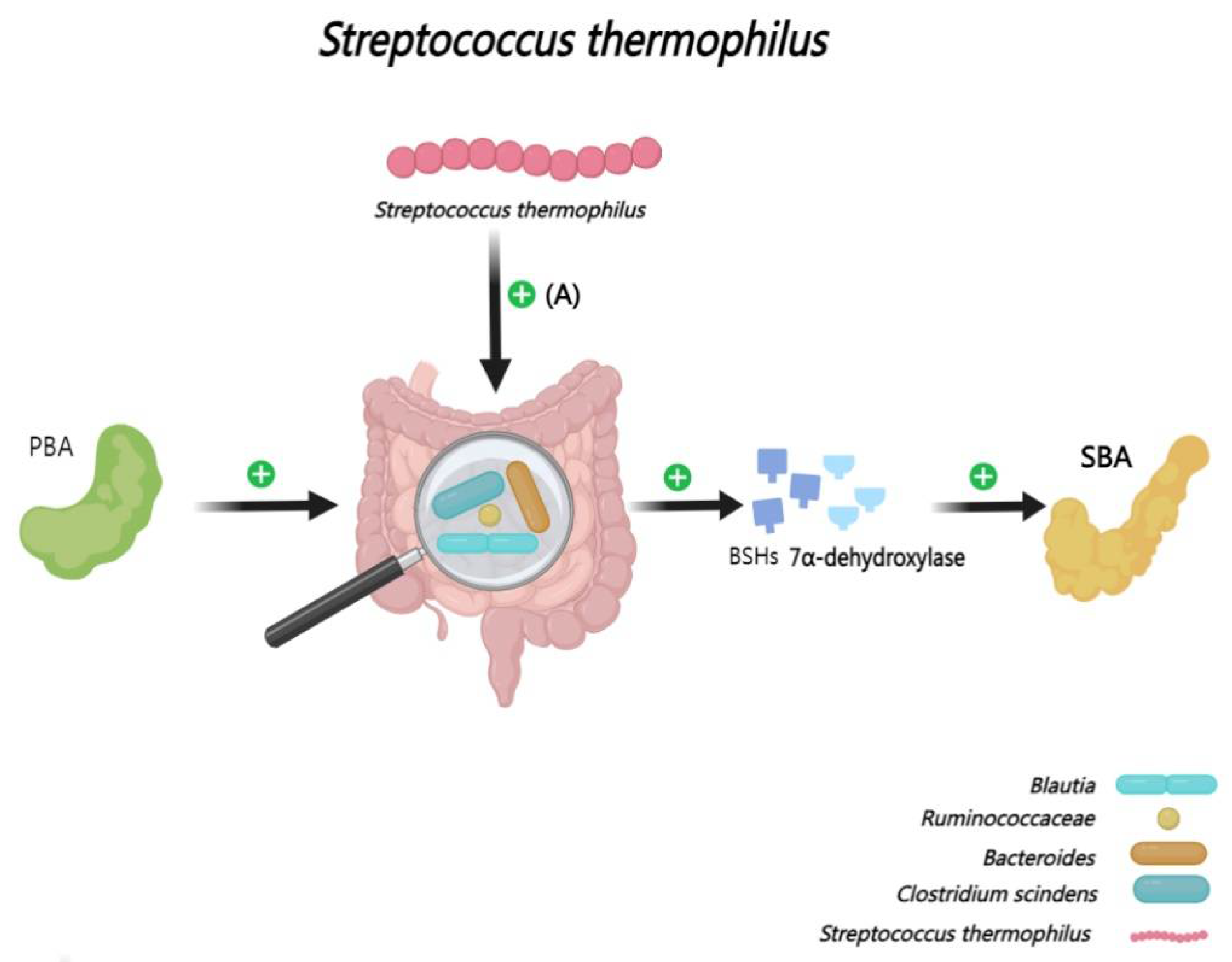
Figure 10.
Interactions Between BAs and Saccharomyces boulardii. (A) Saccharomyces boulardii can protect the intestinal barrier; (B) Saccharomyces boulardii can facilitate the proliferation of intestinal microbiota endowed with BSHs activity; (C) Saccharomyces boulardii aids in the neutralization of enteric toxins. BSHs: Bile salt hydrolases.
Figure 10.
Interactions Between BAs and Saccharomyces boulardii. (A) Saccharomyces boulardii can protect the intestinal barrier; (B) Saccharomyces boulardii can facilitate the proliferation of intestinal microbiota endowed with BSHs activity; (C) Saccharomyces boulardii aids in the neutralization of enteric toxins. BSHs: Bile salt hydrolases.
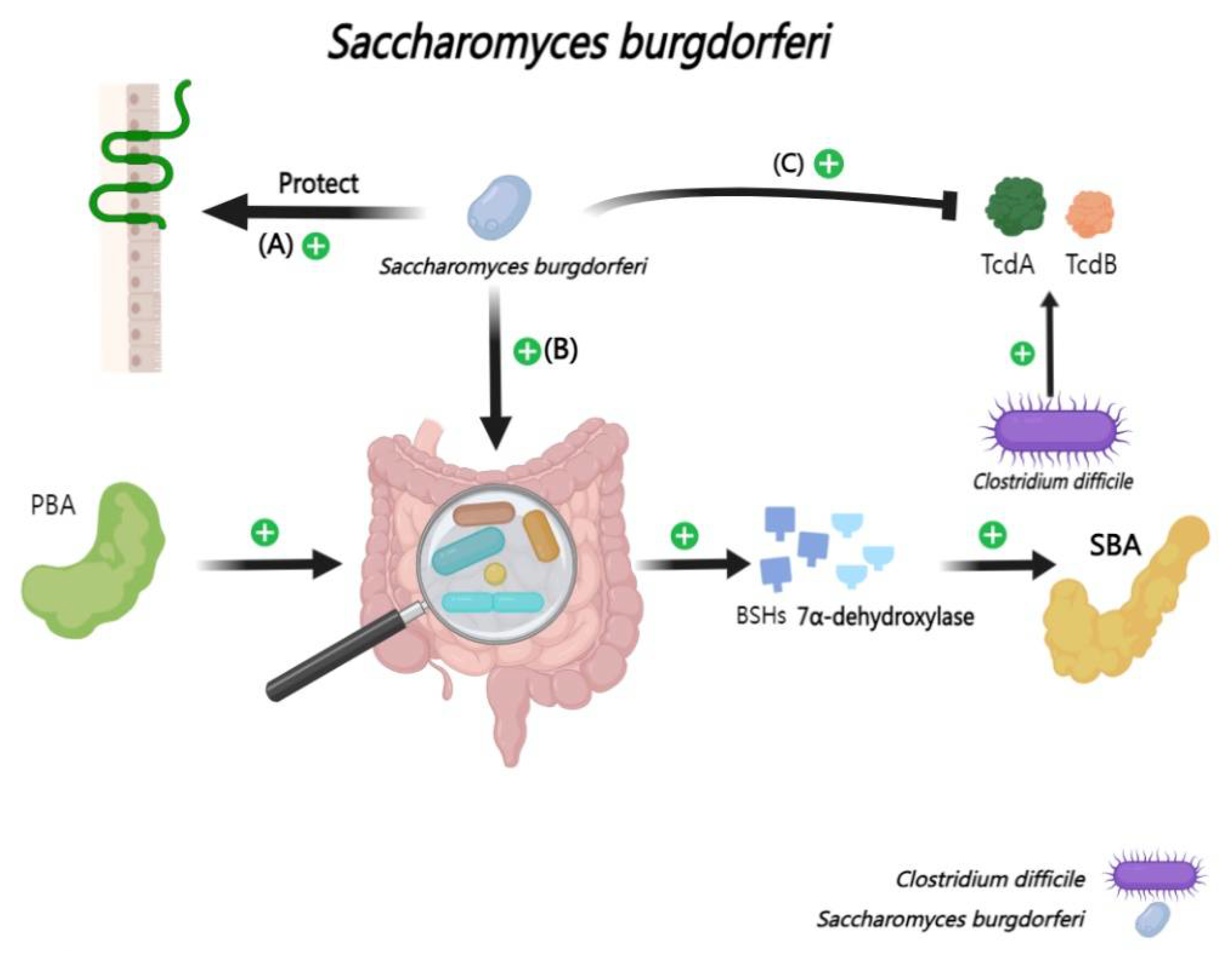
Figure 11.
Interactions Between BAs and Clostridium butyricum. (A) Clostridium butyricum can decrease the abundance of intestinal bacterial populations with BSHs activity, leading to an increase in conjugated bile acid levels; (B) Conjugated bile acids stimulate LXRα and FXR receptors, maintaining intestinal health; (C) Clostridium butyricum reshapes the microbiota by attenuating the inhibitory effects of the FXR/SHP pathway on lipid synthesis; (D) Clostridium butyricum activates the butyrate/GPR43 pathway, reducing damage to the intestinal barrier and restoring the intestinal immune microenvironment. BSHs: Bile salt hydrolases; LXRα: liver X receptor alpha; FXR: Farnesoid X Receptor.
Figure 11.
Interactions Between BAs and Clostridium butyricum. (A) Clostridium butyricum can decrease the abundance of intestinal bacterial populations with BSHs activity, leading to an increase in conjugated bile acid levels; (B) Conjugated bile acids stimulate LXRα and FXR receptors, maintaining intestinal health; (C) Clostridium butyricum reshapes the microbiota by attenuating the inhibitory effects of the FXR/SHP pathway on lipid synthesis; (D) Clostridium butyricum activates the butyrate/GPR43 pathway, reducing damage to the intestinal barrier and restoring the intestinal immune microenvironment. BSHs: Bile salt hydrolases; LXRα: liver X receptor alpha; FXR: Farnesoid X Receptor.
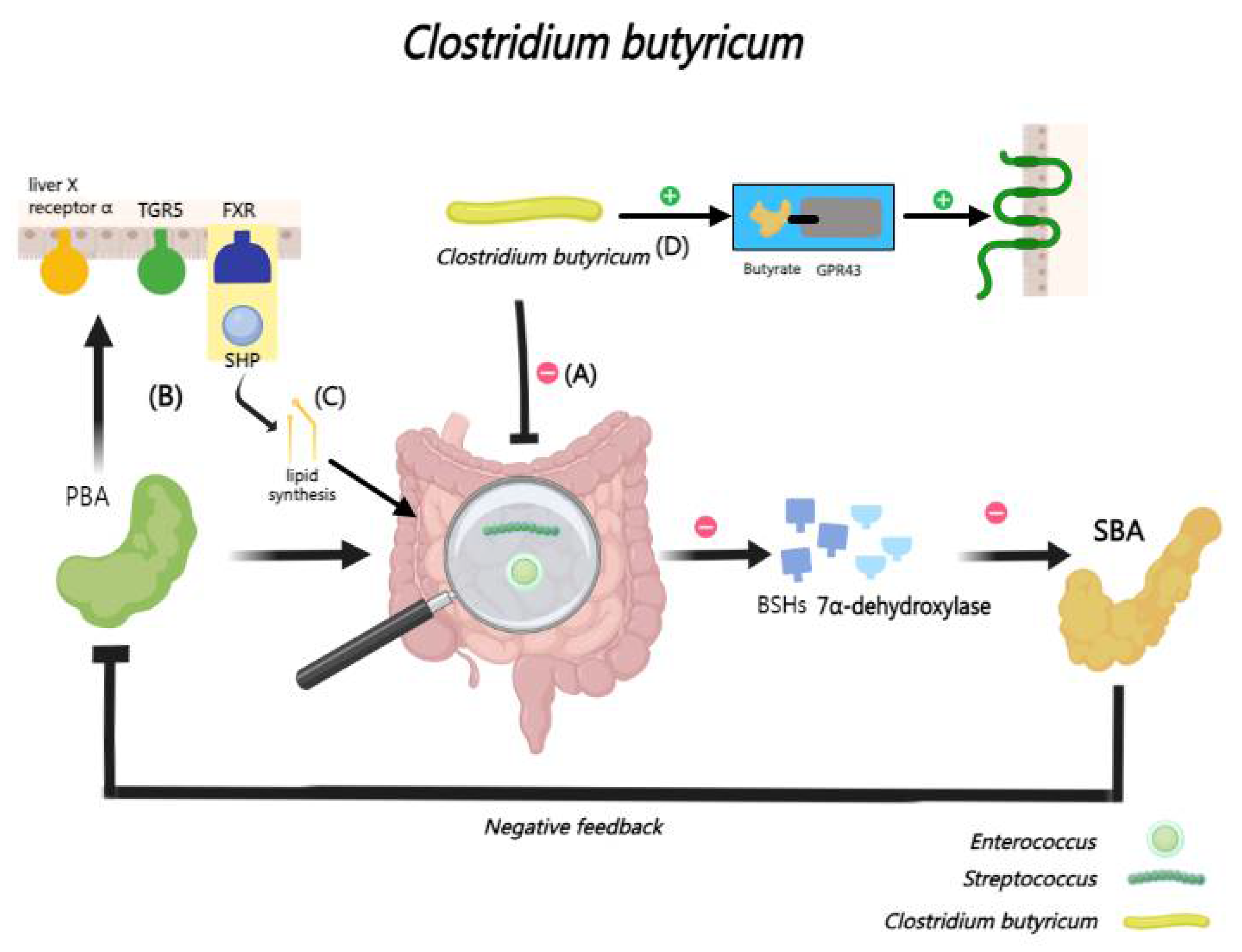
Figure 12.
Interactions Between BAs and Lactic Acid Bacteria. (A) Lactic Acid Bacteria enhances intestinal microbiota, particularly increasing Anaerostipes and Eubacterium, which metabolize inositol into propionic and butyric acids and convert PBA into SBA via 7α-dehydroxylation, potentially inhibiting primary sclerosing cholangitis; (B) Lactic Acid Bacteria improves tight junction proteins ,downregulates inflammatory responses by promoting the growth of beneficial microbial taxa such as Lactobacillus, Prevotella, and Paraprevotella during Clostridioides difficile infection ,thereby altering the bile acid composition and influencing liver injury processe; (C) Lactic Acid Bacteria increased the abundance of Firmicutes and Actinobacteria, thereby conducting to non-alcoholic steatohepatitis mitigation. PBA: Primary bile acid; SBA: Secondary bile acid.
Figure 12.
Interactions Between BAs and Lactic Acid Bacteria. (A) Lactic Acid Bacteria enhances intestinal microbiota, particularly increasing Anaerostipes and Eubacterium, which metabolize inositol into propionic and butyric acids and convert PBA into SBA via 7α-dehydroxylation, potentially inhibiting primary sclerosing cholangitis; (B) Lactic Acid Bacteria improves tight junction proteins ,downregulates inflammatory responses by promoting the growth of beneficial microbial taxa such as Lactobacillus, Prevotella, and Paraprevotella during Clostridioides difficile infection ,thereby altering the bile acid composition and influencing liver injury processe; (C) Lactic Acid Bacteria increased the abundance of Firmicutes and Actinobacteria, thereby conducting to non-alcoholic steatohepatitis mitigation. PBA: Primary bile acid; SBA: Secondary bile acid.
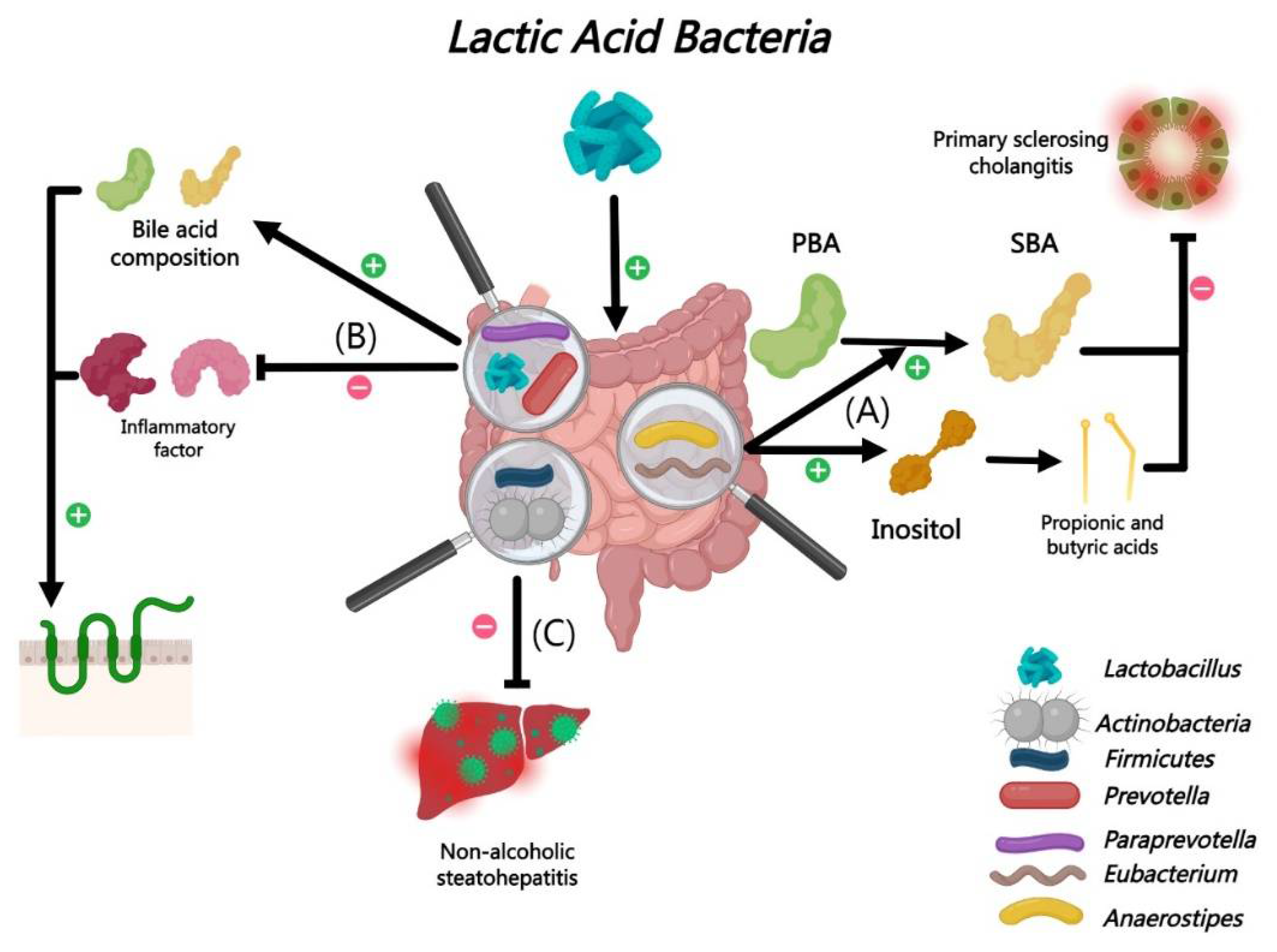
Figure 13.
Interactions Between BAs and Coronavirus SARS-CoV-2. UDCA directly damages the virus structure, inhibiting its replication; (B) UDCA reduces the affinity between the receptor binding domain of Coronavirus SARS-CoV-2 and the host ACE2; (C) UDCA can inhibit FXR gene expression, thereby suppressing ACE2 expression; (D) UDCA possesses anti-inflammatory, antioxidative, immunomodulatory, and anti-apoptotic properties; (E) Increasing the abundance of Collinsella promotes the synthesis of 7β-Hydroxysteroid dehydrogenase, enhancing SBA synthesis and protecting the intestinal barrier. UDCA: Ursodeoxycholic acid; SARS-CoV-2: Severe Acute Respiratory; FXR: Farnesoid X Receptor; ACE2: Angiotensin-converting enzyme 2; SBA: Secondary bile acid.
Figure 13.
Interactions Between BAs and Coronavirus SARS-CoV-2. UDCA directly damages the virus structure, inhibiting its replication; (B) UDCA reduces the affinity between the receptor binding domain of Coronavirus SARS-CoV-2 and the host ACE2; (C) UDCA can inhibit FXR gene expression, thereby suppressing ACE2 expression; (D) UDCA possesses anti-inflammatory, antioxidative, immunomodulatory, and anti-apoptotic properties; (E) Increasing the abundance of Collinsella promotes the synthesis of 7β-Hydroxysteroid dehydrogenase, enhancing SBA synthesis and protecting the intestinal barrier. UDCA: Ursodeoxycholic acid; SARS-CoV-2: Severe Acute Respiratory; FXR: Farnesoid X Receptor; ACE2: Angiotensin-converting enzyme 2; SBA: Secondary bile acid.
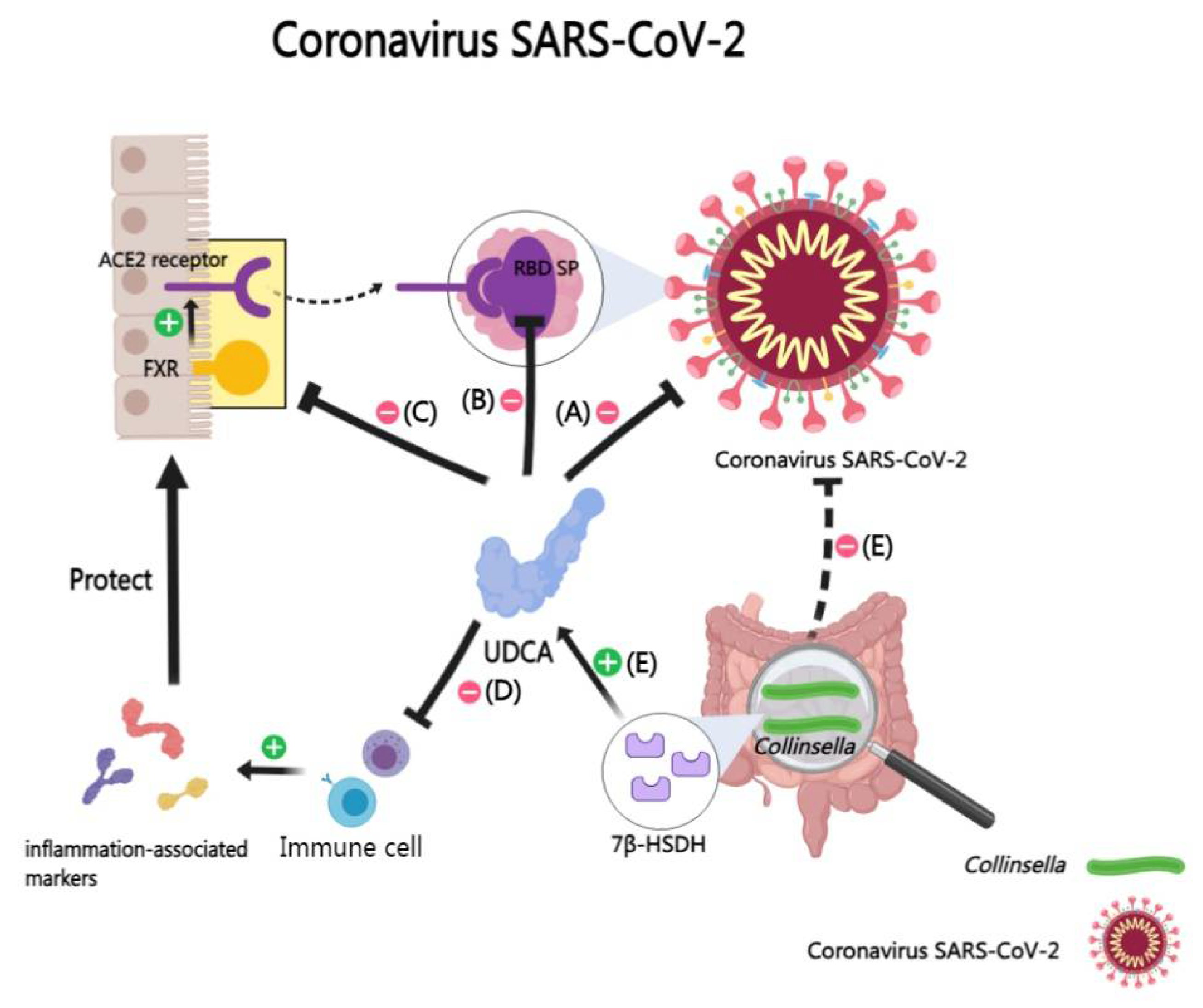
Figure 14.
Interactions Between BAs and Other Viruses. (A) STH manifests antiviral activity against IVA infections through the modulation of signaling pathways, including TLR4, NF-κB, and MAPK; (B) CDCA demonstrates the capacity to attenuate IAV infections by inhibiting the nuclear export of vRNPs; (C) GCDCA enhances the virulence of Norovirus through a mechanism that is not yet clarified; (D) The intestinal microbiota modulates Norovirus-induced infections; (E) CDCA can reduce virus-induced lipid synthesis, inhibiting the replication of Rotavirus; (F) CDCA activates FXR and TGR5 receptors to counteract HBV infection. STH: Sodium taurocholate; TLR4: Toll-like receptor 4; CDCA Chenodeoxycholic acid; vRNPs: Viral ribonucleoproteins; IAV: Influenza A virus; GCDCA: Glycine deoxycholic acid; FXR: Farnesoid X Receptor; TGR5: Takeda G Protein-Coupled Receptor 5; HBV: Hepatitis B virus.
Figure 14.
Interactions Between BAs and Other Viruses. (A) STH manifests antiviral activity against IVA infections through the modulation of signaling pathways, including TLR4, NF-κB, and MAPK; (B) CDCA demonstrates the capacity to attenuate IAV infections by inhibiting the nuclear export of vRNPs; (C) GCDCA enhances the virulence of Norovirus through a mechanism that is not yet clarified; (D) The intestinal microbiota modulates Norovirus-induced infections; (E) CDCA can reduce virus-induced lipid synthesis, inhibiting the replication of Rotavirus; (F) CDCA activates FXR and TGR5 receptors to counteract HBV infection. STH: Sodium taurocholate; TLR4: Toll-like receptor 4; CDCA Chenodeoxycholic acid; vRNPs: Viral ribonucleoproteins; IAV: Influenza A virus; GCDCA: Glycine deoxycholic acid; FXR: Farnesoid X Receptor; TGR5: Takeda G Protein-Coupled Receptor 5; HBV: Hepatitis B virus.
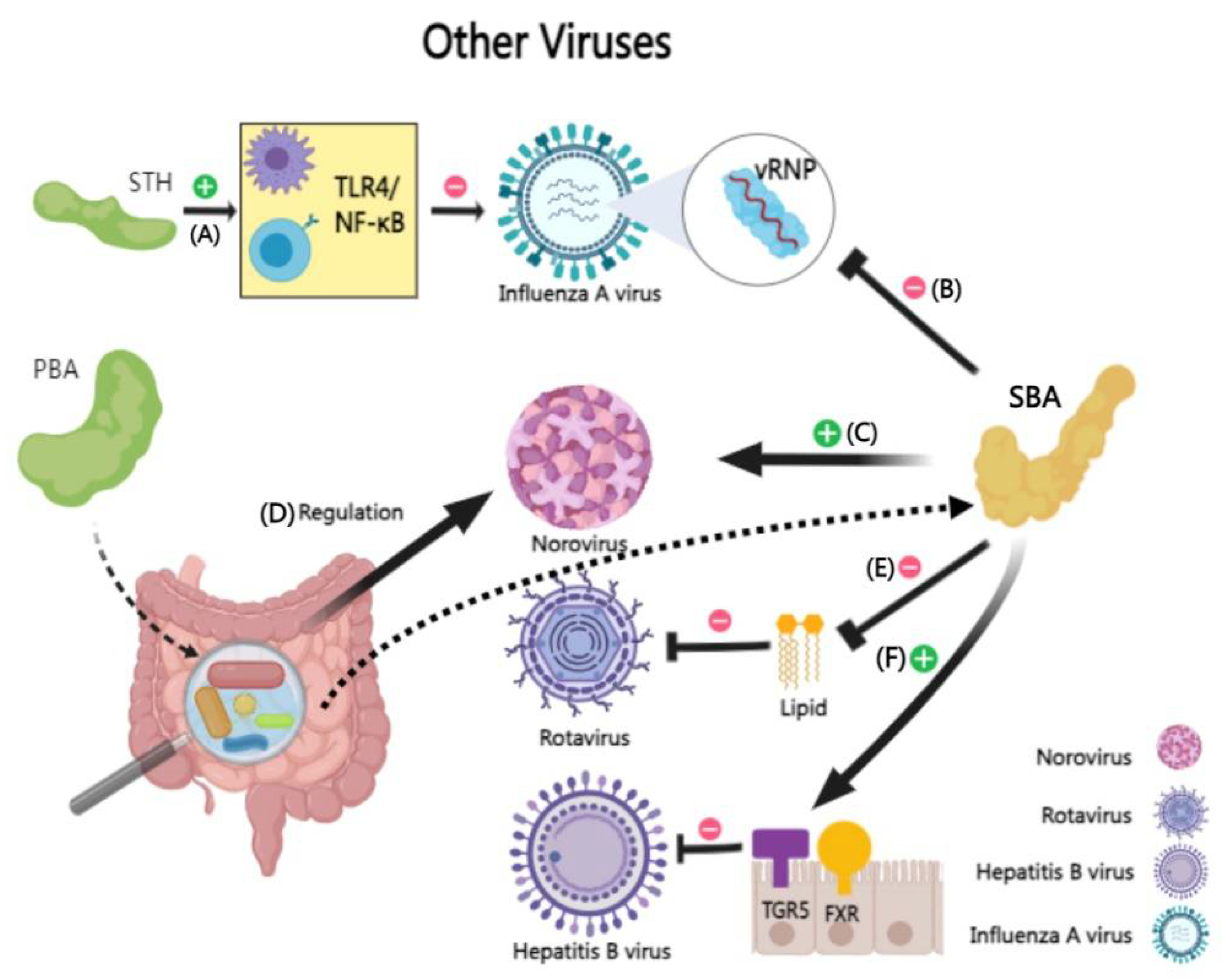
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



