
Submitted:
28 June 2024
Posted:
01 July 2024
You are already at the latest version
Abstract
This article explores the transformative potential of fermentation in elevating the quality of plant-based matrices to match the desirable attributes of traditional dairy and meat products. As the demand for sustainable products without animal welfare issues increases, fermentation emerges as a key process to enhance the organoleptic properties and nutritional content of plant-based analogs. This study explores the effect of fermentation when applied to legume matrices, focusing on the resulting texture, flavor, and nutritional value. A selection of Bacillus subtilis, lactic acid bacteria (LAB) strains, and combinations thereof, showed potential for improving the aforementioned organoleptic and nutritional characteristics of fermented plant bases. In four different legume-derived matrices, fermentation improved texture, degraded undesirable plant carbohydrates, and removed off-flavor compounds, while producing desirable dairy-associated compounds. The degradation of the undesirable beany off-flavor-causing compound hexanal appears to be a universal phenomenon, as every tested strain as well as their combinations exhibited the capability to decrease the hexanal content albeit with varying efficiency. Some LAB strains were found to be capable of producing carotenoids and might hence have the potential for tailoring fermented plant-based matrices for specific applications, such as yellow cheese or red meat analogs.

Keywords:
Bacillus
; lactic acid bacteria
; fermentation
; plant-based
; flavor
1. Introduction
A growing number of people are adopting vegetarian or vegan lifestyles for ethical, environmental, and health reasons. As a result, there is an increased demand for diverse and satisfying plant-based alternatives to traditional dairy and meat products. As consumer preferences evolve, the food industry continues to innovate and expand its offerings to meet the changing demands of an increasingly diverse and conscientious consumer base. Ongoing developments in food technology and research have enabled the creation of more sophisticated plant-based analogs mimicking the sensory attributes of traditional dairy and meat products [1]. However, dairy- and meat-alternative products are often nutritionally unbalanced [2], and their flavor profiles have limited their consumer acceptance. Additionally, texturizing agents such as hydrocolloids (e.g., carrageenan or xanthan gum) are often added to enhance the structural integrity and mouthfeel of the product. However, increasing the number of additives also impacts consumer acceptance negatively. Fermentation can improve the sensory and nutritional attributes, texture, microbial safety, and antimicrobial properties of plant-based dairy and meat alternatives, potentially removing the need for flavor masking and texturing ingredients thus simultaneously improving product quality and moving towards clean-label specifications ([3,4,5,6,7]).
With regards to adopting legume-based foods, the challenge is often related to the presence of undesirable 'beany', ‘green’, ‘grassy’ off-flavors associated with aldehydes, like hexanal and 3-Z-hexenal, alcohols like n-hexanol, n-pentanol, and n-heptanol, as well as ketones (e.g., ethyl vinyl ketone) and furans (e.g., 2-pentyl furan and 2-(1-pentenyl) furan) ([5,8,9,36]). Fermentation of plant-based matrices can result in the reduction or elimination of beany off-flavor and introduce desirable flavor compounds, such as dairy (e.g. diacetyl) and umami (e.g. glutamate) associated metabolites [9]. Additionally, it has been demonstrated that fermentation can lead to the breakdown of plant-specific antinutritional factors like tannins, saponins, phytic acid, α-galactosides, and trypsin inhibitors. As an example, the raffinose family of oligosaccharides (RFO) such as sucrose-core RFOs stachyose, raffinose, verbascose glucose-core RFOs melibiose, manninotriose and manninotetraose can be effectively digested by certain microbes ([10,11]). RFOs are considered anti-nutritional factors since they cause discomfort (flatulence and diarrhea) in humans and animals. Flatulence is the singlemost important deterrant in consumption and utilization of legumes in human and animal diets [11]. Consequently, the incorporation of fermentation and the development of fermented products present a promising avenue for elevating both the nutritional quality and overall acceptability of plant-based dairy and meat analogs.
The color of meat analogs is expected to resemble meat. Most-used ingredients for meat analogs have a beige or yellow-brown color. Therefore, colorants like red beet, red berries, or carrots are added to the ingredient mix, often together with citric, acetic, or lactic acid, in order to match the pH range of the colorant [12]. Pigments can be produced by the fermenting microorganisms, or they may originate in the plant bases themselves, e.g. leghemogobins such as the heme-like complex of iron in soybean [13] or anthocyanin-like structures [14]. The use of pigment-producing microorganisms can naturally introduce desirable colors into the fermented plant bases, e.g. yellow, orange, or red color for applications such as orange Cheddar or meat analogs or purple color for the production of e.g. “blueberry” vegurts. Heme-like complexes of transition metals such as the zinc-protoporpyrin IX complex produced by Lc. lactis subsp. cremoris was shown to improve the color of fermented meat products [19]. Carminic acid, a commonly used natural red food colorant, and a group of flavonoids known as anthocyanins, which are responsible for the color of many fruits, vegetables, and flowers, can be synthesized by microbes ([20,21,22,23,24]). Carotenoids, the most widely distributed pigments in nature, that exhibit yellow, orange, red, and purple colors, are naturally produced by yeasts Rhodotorula, Rhodosporidium, Sporobolomyces, Xanthophyllomyces dendrorhous (former Phaffia rhodozyma) ([25,26]), LAB [27], and filamentous fungi such as Phycomyces blakesleeanus, Blakeslea trispora and Mucor circinelloides ([28,29,30]). These filamentous fungi, capable of producing carotenoids, generate sporangiophores. Consequently, besides imparting desirable color, the filamentous fungi can also provide meat analogs with a muscle-tissue-like texture. Seaweed and microalgae that are already used for flavor enhancement in plant-based seafood analogs ([15,16]) can also modify matrix color as a result of carotenoid production ([17,18]).
Historically, LAB have been employed in prolonging the shelf life and enhancing the flavor and texture of fermented milk. More recently, their efficacy was demonstrated in enhancing both the flavor and texture of plant-based products ([3,5,6]). Some Bacillus species have a known history of safe use in foods in fermented plant-based foods in Africa and Asia. Natto is a traditional Japanese fermented product consisting of cooked soybeans fermented with B. subtilis var. natto. Most European legumes are suitable substrates to produce Natto [31]. In addition, a combination of LAB with B. subtilis strains that do not form spores was shown to improve the texture and the acidification speed in milk [32]. The sporulation-negative phenotype is important for generating products such as plant-based alternatives of yogurts, sour cream, and the like. In this work, we show that some LAB and two sporulation-negative B. subtilis strains and their combinations can be used to improve the texture, flavor, and nutritional value of a variety of legume-derived plant-based matrices.
Materials and Methods
Materials
Four different protein concentrates such as yellow pea (Pisum sativum) (from two different suppliers), chickpea and faba bean (Vicium faba) were used as matrices for fermentation. VITESSENCE™ 1803 Pea Protein containing a minimum of 80% protein content on a dry-matter basis and 3.4% carbohydrates was used to prepare a 4% solution. ADM ProFam™ Pea Protein 580 with 80% protein isolate content and 1.0% carbohydrates was used to prepare a 4% solution. VITESSENCE® Prista P 360 faba bean protein, which is 60% protein concentrate (on a dry basis) and 24.8% carbohydrates, was used to prepare an 8% solution. CP-PRO70® Chickpea Protein Concentrate from Innovapro containing at least 68% protein and 5.7% carbohydrates was used to prepare a 7% solution.
Base Preparation and Fermentation
The legume matrices were prepared by the addition of distilled water to protein concentrates, homogenization, and pasteurization at 90°C for 20 min while stirring. The suspensions were allowed to stand at ambient temperature overnight, followed by a repetition of the pasteurization step. This iterative pasteurization step was undertaken with the aim of eliminating any vegetative cells that may have arisen as a result of the initial pasteurization and thatcould potentially trigger spore germination. When the matrices were cooled to the incubation temperature, the carbohydrate solution (1% sucrose and 1% glucose) and the 1% overnight inoculum of Bacillus and/or 1% overnight inoculum of LAB were added to the matrices, and the matrices were fermented overnight at 30°C or 37°C.
Bacillus subtilis Natto spore(-) strains were incubated in LB medium overnight at 37°C while shaking at 250 rpm. Lactiplantibacillus plantarum, Lacticaseibacillus paracasei, Lactocaseibacillus rhamnosus, Latilactobacillus fragifolii and Leuconostoc mesenteroides were incubated overnight in MRS-Difco at 30°C. S. thermophilus was incubated overnight in M17 + 1% sucrose and 1% glucose at 37°C.
The fermentation was performed in four different formats: 2 ml samples in 20 ml headspace vials for volatile organic compound (VOC) analysis, 0.2 ml samples in a microtiter plate (MTP) for Raffinose Family Oligosaccharides (RFO) analysis, 1 ml samples in 1 ml 96-well MTP for texture measurements using the high-throughput (HTP) compression test [5], and 2 ml samples in 2 ml 96-well MTP for texture measurements using total aspiration and dispense monitoring (TADM) [33].
Before subjecting the samples to the VOC analysis, the fermentation was stopped by adding diluted sulfuric acid (0.4 ml of 2M H2SO4 to each vial containing 2 ml sample) and freezing the samples at -20°C. For RFO measurements, 2 volumes of 60% ethanol were added after the fermentation. The experimental setup is shown in Figure 1.
Analysis of Volatile Organic Compounds
Two grams of the different emulsions were aliquoted directly into 20 ml headspace vials. Uninoculated emulsions were used as negative controls and were not incubated. All samples were prepared in duplicates. After incubation, 400µl of 2M sulfuric acid were added to the headspace vials, and samples were stored at -20°C until analysis.
Volatile organic compounds (VOCs) produced during fermentation were determined by headspace solid phase microextraction gas chromatography coupled to mass spectrometry (HS-SPME-GC-MS) as in [5]. VOCs were calculated as peak height divided by baseline noise (signal-to-noise, S/N). The removal ratio for the beany off-flavors and the enhancement formation ratio for dairy notes compounds were calculated by comparing the detected S/N values of those in the fermented samples to those in the respective uninoculated matrix. Standards solutions for hexanal and diacetyl were included for calculation of the concentration (mg/L). The precision was calculated based on the relative standard deviation of a Cheddar cheese used as control sample that was analyzed at intermittent points throughout the analytical sequence (n=14).
Raffinose Family Type, Alpha-Galactooligosaccharide Analysis
The quantification of sucrose-based α-galactooligosaccharides such as melibiose, raffinose, stachyose, and verbascose from plant-based fermented products was performed. The method quantifies glucose and fructose typically present in the samples as well. The analyses were performed by HPAEC-PAD (High-Performance Anion Exchange Chromatography with Pulsed Amperometry Detection) on a Dionex ICS-6000 system (ThermoFisher, Sunnyvale, CA, USA). The α-galactooligosaccharides were separated with a 250 x 1mm PA-200 column plus guard column and quantified by conductivity detection. The gradient elution was from the generated potassium hydroxide (A). 0-4 min 10 mM KOH, 4 - 4.5 min. 100 mM KOH, 4-5-13 min 200 mM KOH, 13-13.5 min 10 mM KOH, 13.5 min – 30 min 10 mM KOH. The flow rate of the gradient elution was set to 63 µL min-1 with a total run time of 30 min. Retention time and peak area were used to quantify the carbohydrates in the samples. Standards from 2.5 µg mL-1 to 500 mg L-1 were used for calibration curves.
Carotenoid Production
BLAST analysis using crtN, crtM sequences from Lactobacillaceae (GenBank BBA82255.1 and BBA82256.1) was used to find strains that have a genetic potential for producing carotenoids. Carotenoids were extracted from bacterial cells grown overnight in 50 ml MRS and separated from the supernatant by centrifugation. 10 ml Y-PER™ (Thermo Fisher Scientific, Cat. No. 78990) was added to the cell pellet and left at room temperature in the dark for 1 h. Then 20 mL of 96% ethanol were added, and the suspension was stored at -20 °C in the dark overnight. Afterward, the material was thawed at room temperature and 5 mL of n-hexane was added to the suspension, and the suspension was left to extract for 2 h with gentle agitation every 20 min. After the final 20 min, the n-hexane fraction was transferred to a new tube. The extract was concentrated by passive evaporation in the dark and then the spectrum of the concentrated extract was measured between 400 nm and 500 nm on a Shimadzu UV-1900i spectrophotometer in quartz cuvettes [34] .
Results and Discussion
Texture Formation
For fermented milk and plant-based products, texture is an important quality parameter and depends on both the microbes driving the fermentation as well as the process conditions. When growing in legume-derived bases, LABs produce organic acids, and that results in gel formation leading to changes in texture. Four different protein concentrates such as pea from two different suppliers, chickpea and faba bean were used as matrices for fermentation using two B. subtilis Natto spore(-) strains, two Lb. paracasei strains, Lb. plantarum, Lb. rhamnosus, Leuconostoc mesenteroides and the various combinations thereof. The texture of the fermented matrices was evaluated using the HTP compression test and TADM (Figure 2). B. subtilis strains did not acidify the matrices, and thus the gel was only formed in the presence of LAB (see “control”, Figure 2). Some LAB strains are known for their exocellular polysaccharide production, which can enhance the texture of milk and plant-based matrices [3]. A combination of Bacillus and Lb. paracasei 1 resulted in the highest overall texture, as assessed by the positive compression area and TADM. As expected, the thermophilic S. thermophilus had the highest texture formation when tested at 37°C, while the mesophilic Leuconostoc had the highest texture formation when tested at 30°C.
Reduction of Off-Flavors
Off-flavors in legumes are commonly associated with volatile compounds such as aldehydes, pyrazines, furans,sulfide compounds or non-volatile compounds such as saponins, phenolic acids, peptides, amino acids, or alkaloids [35]. Here, we accessed if B. subtilis, LAB and their combinations can improve the flavor of the four different legume matrices by removing these off-flavors (Figure 3, Supplementary Figures S1–S4).
We focused on four different compounds as representative of off-flavors: 3-methyl-butanal, pentanal, hexanal and 1-penten-3-ol. Hexanal is one of the main off-flavors in legumes (pea, soybean and faba bean) and it is selected together with the pentanal due to their reported involvement in the green, grassy and beany off-flavor ([9,36]). 3-Methyl-butanal has a malty flavor [37]. 1-Penten-3-ol contributes with a grassy, vegetable-like perception in legume protein matrices ([38,39]) or as earthy, mushroom-like or bitter off-notes [35].
In most samples, the off-flavor related compounds were reduced as the result of fermentation. However, the degradation patterns varied depending on the matrix and strains used (Figure 3, Supplementary Figures S1–S4). In general, the abundance of pentanal, hexanal, 1-penten-3-ol and 3-methyl-butanal were reduced in the samples fermented by B. subtilis, LAB, or their combinations at the two different temperatures. In general, pentanal and hexanal were reduced more when B. subtilis was combined with Leuconostoc, Lb. plantarum, Lb. rhamnosus and Lb. paracasei than B. subtilis alone. The degradation patterns were different, depending on the matrix. In Vitessence pea, no degradation was observed for the four off-flavor associated compounds when B. subtilis was fermented at 30°C. The 1-penten-3-ol concentration was reduced the most when B. subtilis was combined with Leuconostoc. Pentanal and hexanal were degraded to a higher extent when B. subtilis was combined with Leuconostoc, Lb. paracasei and Lb. rhamnosus. 3-Methyl-butanal was degraded to a higher extent when B. subtilis was combined with Lb. rhamnosus. In ADM pea, 1-penten-3-ol was degraded more efficiently when B. subtilis was combined with Leuconostoc, Lb. paracasei and Lb. rhamnosus. In chickpea, in most fermented samples the degradation of hexanal and pentanal took place, with slightly higher efficiency when B. subtilis was combined with LAB, especially Leuconostoc, Lb. paracasei and S. thermophilus. 3-Methyl-butanal was degraded to a higher extent when B. subtilis fermented the matrix alone or in combination with S. thermophilus in both chickpea and faba. The combination of B. subtilis with S. thermophilus was efficient for the removal of 1-penten-3-ol in chickpea. However, this compound was not found in faba.
Formation of Desirable Dairy Note Compounds
Four compounds were selected to demonstrate the potential of using fermentation as a means of producing dairy-associated flavor compounds: diacetyl, acetoin, 2,3-pentanedione, and 2-nonanone. Diacetyl and acetoin are important aroma compounds in dairy products due to their buttery flavor [40], while 2,3-pentanedione and 2-nonanone are related to a buttery, creamy, and sweet flavor [41].
In all the samples, the dairy-associated compounds were produced as the result of fermentation. However, the production patterns varied depending on the matrix and strains used (Figure 3, Supplementary Figures S5–S8). As an example, the mixed culture of B. subtilis and Lb. paracasei 2 or B. subtilis and Lb. rhamnosus produced comparable or higher amounts of the desirable flavor compound diacetyl than the single LAB strains. Additionally, the mixed cultures degraded more efficiently the undesirable off-flavor compound hexanal (Figure 3). This exemplifies a synergistic interaction, where the combined effect exceeds the average of the amounts of dairy-associated compounds produced or off-flavor-associated compounds degraded by individual strains.
Overall, B. subtilis and LAB combinations resulted in higher diacetyl production compared to B. subtilis alone (Supplementary Figures S5–S8). In faba, diacetyl and acetoin were produced by all B. subtilis and LAB combinations, except for B. subtilis combined with Leuconostoc and Lb. plantarum. In Vitessense pea, the production of diacetyl and acetoin was more pronounced for most B. subtilis - LAB combinations compared to B. subtilis alone. In ADM pea, diacetyl and acetoin were formed for all B. subtilis - LAB combinations except for Leuconostoc. In chickpea, the diacetyl was formed for all combinations except for B. subtilis combined with Leuconostoc or Lb. plantarum and acetoin production was not observed for any of the combinations. 2,3-Pentanedionee was produced to a greater extent when B. subtilis was combined with S. thermophilus in faba, chickpea, and ADM pea. In Vitessense pea, 2,3-pentanedione and 2-nonanone were not formed under any of the tested conditions. 2-Nonanone was produced to a larger extent when B. subtilis and Lb. rhamnosus were combined, and degraded when combinations of B. subtilis and Leuconostoc were used. In ADM pea, 2-nonanone was generated to a greater extent when B. subtilis was combined with Lb. paracasei (at 37°C) and Lb. rhamnosus.
Ethanol and Ester Formation
The formation of ethyl esters is considered an important contributor to cheese flavor. This reaction is dependent on available ethanol and other factors such as esterase activity and free fatty acids [42]. Here, ethyl-acetate and ethyl-hexanoate were selected as representatives of this compound class [43]. Those compounds could be detected in a much larger formation in all of the matrices when fermented using a combination of B. subtilis and the heterofermentative Leuconostoc, as expected (Supplementary Figures S9–S12).
Degradation of Raffinose-Family Oligosaccharides
We have investigated if the fermentation of the four legume matrices can result in the degradation of poorly digestible or undigestible oligosaccharides (RFOs), whose fermentation can lead to digestive discomfort (e.g., flatulence, diarrhea). The degradation of RFOs varied depending on the matrix and strains used (Figure 4, Supplementary Figures S13–S16).
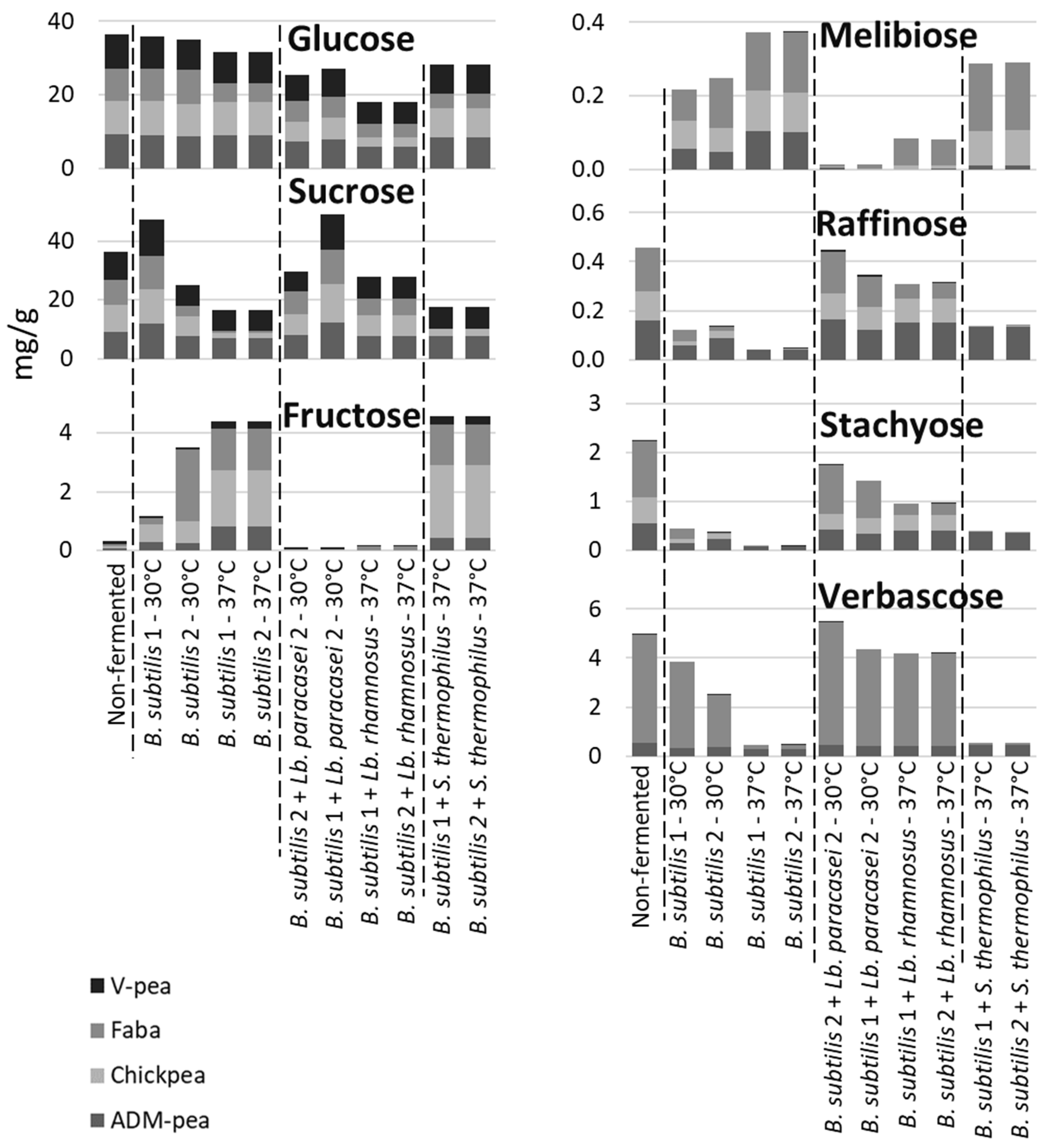
In samples containing B. subtilis strains alone, or in combination with S. thermophilus, the content of verbascose, stachyose, and raffinose decreased, presumably as a result of α-galactosidase activity, while the content of melibiose, fructose, and in some cases sucrose increased (Figure 4). The increase in the content of fructose may be attributed to the hydrolysis of verbascose, stachyose, raffinose and sucrose during fermentation, while the accumulation of melibiose was likely the result of the degradation of verbascose, stachyose and raffinose. These results are in agreement with previous reports on S. thermophilus in soy, where it was shown to consume sucrose, release fructose [44], and degrade the indigestible α-oligogalactosides ([45,46,47]).
The degradation of RFOs through fermentation by a combination of B. subtilis with Lb. paracasei 2 or Lb. rhamnosus strains was less efficient, particularly when mono- and disaccharides like glucose and sucrose, which are often preferred by the bacteria, were still abundant. If sucrose and glucose were not present in the matrices after fermentation, a different carbohydrate pattern might have been observed, with RFOs potentially being degraded to a greater extent.
Pigment Production
Pigment-producing microorganisms are of interest for some plant-based product segments. Carotenoid production was detected in Lb. fragifolii strain that contained dehydrosqualene desaturase (crtN) and dehydrosqualene synthase (crtM) sequences described in LAB ([27,48]). The spectrum of the n-hexane extract showed the characteristic absorbance peaks of β-carotene (Supplementary Figure S17) as reported [34].
Despite we detected the two genes from the carotenoid pathway in the Lb. fragifolii strain, and we were able to extract carotenoids from the bacterial cells, we could not detect a significant color change of several plant-based matrices when fermented using Lb. fragifolii, compared to a non-carotenoid-producing control strain Lb. paracasei 1 (data not shown). As carotenoids are produced intracellularly, they might not give a visible color effect on the matrix, unless the cells are lysed. Also, carotenoids might be produced in small amounts, and it would require optimization to enhance their production, as described in ([28,29,30]).
Conclusion
The increasing popularity of plant-based diets has created a competitive market for dairy and meat analogs. To stay relevant and attract a wide consumer base, manufacturers are diversifying their product portfolio to offer unique and appealing alternatives. Fermentation of plant-based matrices was shown to be a powerful tool for the improvement of flavor, texture, and nutritional value of such matrices and has the potential for generating more sustainable food with great taste and texture. Here we show that some B. subtilis, LAB strains, and their combinations were able to reduce the amount of the beany-associated off-flavor compounds such as hexanal, while producing metabolites associated with pleasant dairy notes such as diacetyl. Moreover, some strains and strain combinations could degrade the indigestible α-oligogalactosides found in different legume matrices such as pea, chickpea and faba.
Although legume protein has been widely explored for dairy and meat alternatives, its consumption is still limited by undesirable sensory characteristics and low solubility. All these properties can be modified during the protein extraction process. Fermentation of plant-based matrices can be used to generate the final product, e.g. plant-based cheese, vegurt, meat, or seafood analog, but also to improve the properties of plant proteins before their use. Nutritional, sensory, and technological properties of plant proteins can be influenced by the protein extraction process. In a process for plant protein isolation from flour, acid is often used to precipitate and isolate the proteins [49]. Fermentation has been used to improve the properties of pea protein prior to precipitation [50]. The reduction in pH throughout the protein precipitation process can be attained through the natural generation of lactic acid by LAB during fermentation. This natural acid production can serve as a substitute for the addition of mineral acids. LAB-based cultures not only have the ability to produce the required lactic acid for protein precipitation but also improve the protein flavor and nutritional properties, alone or in combination with e.g. B. subtilis or yeasts.
Meat analogs might be produced through a two-step process. Initially, a LAB / B. subtilis fermentation is employed to swiftly lower the pH, inhibiting contaminants within the matrix while enhancing the taste, texture, and nutritional value. In the subsequent step, filamentous fungi such as e.g. Aspergillus oryzae, Phycomyces blakesleeanus, Blakeslea trispora and Mucor circinelloides may be utilized. These fungi have the capability to consume the acids generated in the first step, which might be undesirable for the taste of meat analogs. Moreover, they might contribute to the development of an even more appealing flavor, color, and texture closely resembling meat, given that their filaments mimic meat muscles.
However, additional research is needed to further investigate how the fermentation and the change in the profile of RFOs, flavor compounds and nutritional factors can impact the overall sensory perception and liking in fermented plant-based products to facilitate the production of higher quality plant based end-products with greater consumer acceptance.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Acknowledgments
The authors would like to thank Helle Simonsen for her help with the GC-MS (VOCs) analysis.
Author Details
Novonesis A/S, Gammel Venlighedsvej 14, DK-2970 Hoersholm, Denmark.
Competing Interests
All authors are employed by Novonesis A/S.
References
- D. J. McClements, Meat Less: The next food revolution, vol. 30, no. 1. 2023.
- A. A. Nolden and C. G. Forde, “The Nutritional Quality of Plant-Based Foods,” Sustainability (Switzerland), vol. 15, no. 4. 2023. doi: 10.3390/su15043324.
- V. K. Poulsen et al., “Versatile Lactococcus lactis strains improve texture in both fermented milk and soybean matrices,” FEMS Microbiol Lett, vol. 369, no. 1, 2022, doi: 10.1093/femsle/fnac117.
- M. Tangyu et al., “Flavour by design: food-grade lactic acid bacteria improve the volatile aroma spectrum of oat milk, sunflower seed milk, pea milk, and faba milk towards improved flavour and sensory perception,” Microb Cell Fact, vol. 22, no. 1, 2023, doi: 10.1186/s12934-023-02147-6.
- C. Masiá, R. Fernández-Varela, V. K. Poulsen, P. E. Jensen, and K. I. Sørensen, “Composition of bacterial blends for fermentation-induced pea protein emulsion gels using multi-property screening of lactic acid bacteria,” Food Biosci, vol. 56, 2023, doi: 10.1016/j.fbio.2023.103333.
- M. Tangyu, J. Muller, C. J. Bolten, and C. Wittmann, “Fermentation of plant-based milk alternatives for improved flavour and nutritional value,” Applied Microbiology and Biotechnology, vol. 103, no. 23–24. 2019. doi: 10.1007/s00253-019-10175-9.
- F. Beck, N. R. Pedersen, and D. S. Nielsen, “Fermented Rapeseed and Soybean Alone and in Combination with Macro Algae Inhibit Human and Pig Pathogenic Bacteria In Vitro,” Microorganisms, vol. 12, no. 5, 2024, doi: 10.3390/microorganisms12050891.
- A. R. Harper, R. C. J. Dobson, V. K. Morris, and G. J. Moggré, “Fermentation of plant-based dairy alternatives by lactic acid bacteria,” Microbial Biotechnology, vol. 15, no. 5. 2022. doi: 10.1111/1751-7915.14008.
- W. Engels, J. Siu, S. van Schalkwijk, W. Wesselink, S. Jacobs, and H. Bachmann, “Metabolic Conversions by Lactic Acid Bacteria during Plant Protein Fermentations,” Foods, vol. 11, no. 7, 2022, doi: 10.3390/foods11071005.
- K. M. Heravi, H. Watzlawick, and J. Altenbuchner, “The melredca operon encodes a utilization system for the raffinose family of oligosaccharides in bacillus subtilis,” J Bacteriol, vol. 201, no. 15, 2019, doi: 10.1128/JB.00109-19.
- D. Elango et al., “Raffinose Family Oligosaccharides: Friend or Foe for Human and Plant Health?,” Frontiers in Plant Science, vol. 13. 2022. doi: 10.3389/fpls.2022.829118.
- S. van Vliet et al., “A metabolomics comparison of plant-based meat and grass-fed meat indicates large nutritional differences despite comparable Nutrition Facts panels,” Sci Rep, vol. 11, no. 1, 2021, doi: 10.1038/s41598-021-93100-3.
- M. I. Ahmad, S. Farooq, Y. Alhamoud, C. Li, and H. Zhang, “Soy Leghemoglobin: A review of its structure, production, safety aspects, and food applications,” Trends in Food Science and Technology, vol. 141. 2023. doi: 10.1016/j.tifs.2023.104199.
- H. E. Khoo, A. Azlan, S. T. Tang, and S. M. Lim, “Anthocyanidins and anthocyanins: Colored pigments as food, pharmaceutical ingredients, and the potential health benefits,” Food and Nutrition Research, vol. 61. 2017. doi: 10.1080/16546628.2017.1361779.
- B. Coleman et al., “Potential of microalgae as flavoring agents for plant-based seafood alternatives,” Future Foods, vol. 5, 2022, doi: 10.1016/j.fufo.2022.100139.
- M. Kazir and Y. D. Livney, “Plant-based seafood analogs,” Molecules, vol. 26, no. 6. 2021. doi: 10.3390/molecules26061559.
- M. García-Vaquero, N. Brunton, and T. Lafarga, “Microalgae as a source of pigments for food applications,” in Cultured Microalgae for the Food Industry: Current and Potential Applications, 2021. doi: 10.1016/B978-0-12-821080-2.00014-9.
- V. Balasubramaniam, L. June Chelyn, S. Vimala, M. N. Mohd Fairulnizal, I. A. Brownlee, and I. Amin, “Carotenoid composition and antioxidant potential of Eucheuma denticulatum, Sargassum polycystum and Caulerpa lentillifera,” Heliyon, vol. 6, no. 8, 2020, doi: 10.1016/j.heliyon.2020.e04654.
- M. Kauser-Ul-alam, Y. Toba, S. Hioki, T. Hayakawa, H. Kumura, and J. I. Wakamatsu, “Lactococcus lactis subsp. cremoris Produces Zinc Protoporphyrin IX Both Aerobically and Anaerobically and Improves the Bright Red Color of Fermented Meat Products,” Foods, vol. 9, no. 11, 2020, doi: 10.3390/foods9111583.
- A. Solopova et al., “Engineering Lactococcus lactis for the production of unusual anthocyanins using tea as substrate,” Metab Eng, vol. 54, 2019, doi: 10.1016/j.ymben.2019.04.002.
- R. P. Pandey, P. Parajuli, M. A. G. Koffas, and J. K. Sohng, “Microbial production of natural and non-natural flavonoids: Pathway engineering, directed evolution and systems/synthetic biology,” Biotechnology Advances, vol. 34, no. 5. 2016. doi: 10.1016/j.biotechadv.2016.02.012.
- J. Zha, X. Wu, and M. A. Koffas, “Making brilliant colors by microorganisms,” Current Opinion in Biotechnology, vol. 61. 2020. doi: 10.1016/j.copbio.2019.12.020.
- Q. Zhang et al., “De novo biosynthesis of carminic acid in Saccharomyces cerevisiae,” Metab Eng, vol. 76, 2023, doi: 10.1016/j.ymben.2023.01.005.
- R. J. N. Frandsen et al., “Heterologous production of the widely used natural food colorant carminic acid in Aspergillus nidulans,” Sci Rep, vol. 8, no. 1, 2018, doi: 10.1038/s41598-018-30816-9.
- G. Sandmann, H. Pollmann, S. Gassel, and J. Breitenbach, “Xanthophyllomyces dendrorhous, a Versatile Platform for the Production of Carotenoids and Other Acetyl-CoA-Derived Compounds,” in Advances in Experimental Medicine and Biology, vol. 1261, 2021. doi: 10.1007/978-981-15-7360-6_11.
- P. Buzzini, M. Innocenti, B. Turchetti, D. Libkind, M. Van Broock, and N. Mulinacci, “Carotenoid profiles of yeasts belonging to the genera Rhodotorula, Rhodosporidium, Sporobolomyces, and Sporidiobolus,” Can J Microbiol, vol. 53, no. 8, 2007, doi: 10.1139/W07-068.
- M. Legein, S. Wittouck, and S. Lebeer, “Latilactobacillus fragifolii sp. nov., isolated from leaves of a strawberry plant (Fragaria x ananassa),” Int J Syst Evol Microbiol, vol. 72, no. 1, 2022, doi: 10.1099/ijsem.0.005193.
- V. Kuzina and E. Cerdá-Olmedo, “Ubiquinone and carotene production in the Mucorales Blakeslea and Phycomyces,” Appl Microbiol Biotechnol, vol. 76, no. 5, 2007, doi: 10.1007/s00253-007-1069-7.
- E. Cerdá-Olmedo, “ Phycomyces and the biology of light and color ,” FEMS Microbiol Rev, vol. 25, no. 5, 2001, doi: 10.1111/j.1574-6976.2001.tb00588.x.
- M. J. Ruiz-Hidalgo, M. A. López-Matas, A. Velayos, P. D. Fraser, P. M. Bramley, and A. P. Eslava, “Carotenoid Mutants of Mucor circinelloides,” Botanica Acta, vol. 108, no. 4, 1995, doi: 10.1111/j.1438-8677.1995.tb00511.x.
- R. Rocchi, J. Zwinkels, M. Kooijman, A. Garre, and E. J. Smid, “Development of novel natto using legumes produced in Europe,” Heliyon, vol. 10, no. 5, 2024, doi: 10.1016/j.heliyon.2024.e26849.
- K. Bjerre, M. D. Cantor, T. Janzen, and P. Derkx, “Fermented milk inoculated with both lactic acid bacteria (LAB) and Bacillus,” US11684073B2, 2017.
- V. K. Poulsen, P. Derkx, and G. Oregaard, “High-throughput screening for texturing Lactococcus strains,” FEMS Microbiol Lett, vol. 366, no. 2, 2019, doi: 10.1093/femsle/fnz001.
- S. Lara-Abia, G. Lobo-Rodrigo, J. Welti-Chanes, and M. Pilar Cano, “Carotenoid and carotenoid ester profile and their deposition in plastids in fruits of new papaya (Carica papaya l.) varieties from the canary islands,” Foods, vol. 10, no. 2, 2021, doi: 10.3390/foods10020434.
- S. Saffarionpour, “Off-Flavors in Pulses and Grain Legumes and Processing Approaches for Controlling Flavor-Plant Protein Interaction: Application Prospects in Plant-Based Alternative Foods,” Food and Bioprocess Technology. 2023. doi: 10.1007/s11947-023-03148-4.
- E. Fischer, R. Cachon, and N. Cayot, “Impact of Ageing on Pea Protein Volatile Compounds and Correlation with Odor,” Molecules, vol. 27, no. 3, 2022, doi: 10.3390/molecules27030852.
- Y. Wang et al., “Flavor challenges in extruded plant-based meat alternatives: A review,” Compr Rev Food Sci Food Saf, vol. 21, no. 3, 2022, doi: 10.1111/1541-4337.12964.
- C. El Youssef, P. Bonnarme, S. Fraud, A. C. Péron, S. Helinck, and S. Landaud, “Sensory improvement of a pea protein-based product using microbial co-cultures of lactic acid bacteria and yeasts,” Foods, vol. 9, no. 3, 2020, doi: 10.3390/foods9030349.
- M. Xu, Z. Jin, Y. Lan, J. Rao, and B. Chen, “HS-SPME-GC-MS/olfactometry combined with chemometrics to assess the impact of germination on flavor attributes of chickpea, lentil, and yellow pea flours,” Food Chem, vol. 280, 2019, doi: 10.1016/j.foodchem.2018.12.048.
- V. Macciola, G. Candela, and A. De Leonardis, “Rapid gas-chromatographic method for the determination of diacetyl in milk, fermented milk and butter,” Food Control, vol. 19, no. 9, 2008, doi: 10.1016/j.foodcont.2007.08.014.
- M. Zhao et al., “Variation of Aroma Components of Pasteurized Yogurt with Different Process Combination before and after Aging by DHS/GC-O-MS,” Molecules, vol. 28, no. 4, 2023, doi: 10.3390/molecules28041975.
- T. B. Pedersen, D. Ristagno, P. L. H. McSweeney, F. K. Vogensen, and Y. Ardö, “Potential impact on cheese flavour of heterofermentative bacteria from starter cultures,” Int Dairy J, vol. 33, no. 2, 2013, doi: 10.1016/j.idairyj.2013.03.003.
- S. Rajendran, P. Silcock, and P. Bremer, “Flavour Volatiles of Fermented Vegetable and Fruit Substrates: A Review,” Molecules, vol. 28, no. 7. 2023. doi: 10.3390/molecules28073236.
- M. Boulay, M. Al Haddad, and F. Rul, “Streptococcus thermophilus growth in soya milk: Sucrose consumption, nitrogen metabolism, soya protein hydrolysis and role of the cell-wall protease PrtS,” Int J Food Microbiol, vol. 335, 2020, doi: 10.1016/j.ijfoodmicro.2020.108903.
- O. N. Donkor, A. Henriksson, T. Vasiljevic, and N. P. Shah, “α-Galactosidase and proteolytic activities of selected probiotic and dairy cultures in fermented soymilk,” Food Chem, vol. 104, no. 1, 2007, doi: 10.1016/j.foodchem.2006.10.065.
- Y. C. Wang, R. C. Yu, H. Y. Yang, and C. C. Chou, “Sugar and acid contents in soymilk fermented with lactic acid bacteria alone or simultaneously with bifidobacteria,” Food Microbiol, vol. 20, no. 3, 2003, doi: 10.1016/S0740-0020(02)00125-9.
- M. S. Garro, G. F. De Valdez, G. Oliver, and G. S. De Giori, “Growth characteristics and fermentation products of Streptococcus salivarius subsp. thermophilus, Lactobacillus casei and L. fermentum in soymilk,” European Food Research and Technology, vol. 206, no. 1, 1998, doi: 10.1007/s002170050217.
- M Kim M, DH Jung, DH Seo, WH Chung, MJ Seo, “Genome analysis of Lactobacillus plantarum subsp. plantarum KCCP11226 reveals a well-conserved C30 carotenoid biosynthetic pathway,”. 3 Biotech, vol 10, no 4, 2020,. doi: 10.1007/s13205-020-2149-y.
- C. Tanger, J. Engel, and U. Kulozik, “Influence of extraction conditions on the conformational alteration of pea protein extracted from pea flour,” Food Hydrocoll, vol. 107, 2020, doi: 10.1016/j.foodhyd.2020.105949.
- M. Emkani, B. Oliete, and R. Saurel, “Pea protein extraction assisted by lactic fermentation: Impact on protein profile and thermal properties,” Foods, vol. 10, no. 3, 2021, doi: 10.3390/foods10030549.
Figure 1.
Experimental setup. Several LAB and B. subtilis strains and their combinations were used to ferment four different legume matrices. Acidification was monitored using the color-of-pH method. Texture was measured using total aspiration and dispense monitoring (TADM) and high-throughput (HTP) compression test. The raffinose family of oligosaccharides (RFO) and volatile organic compounds (VOC) were assessed before and after the fermentation.
Figure 1.
Experimental setup. Several LAB and B. subtilis strains and their combinations were used to ferment four different legume matrices. Acidification was monitored using the color-of-pH method. Texture was measured using total aspiration and dispense monitoring (TADM) and high-throughput (HTP) compression test. The raffinose family of oligosaccharides (RFO) and volatile organic compounds (VOC) were assessed before and after the fermentation.
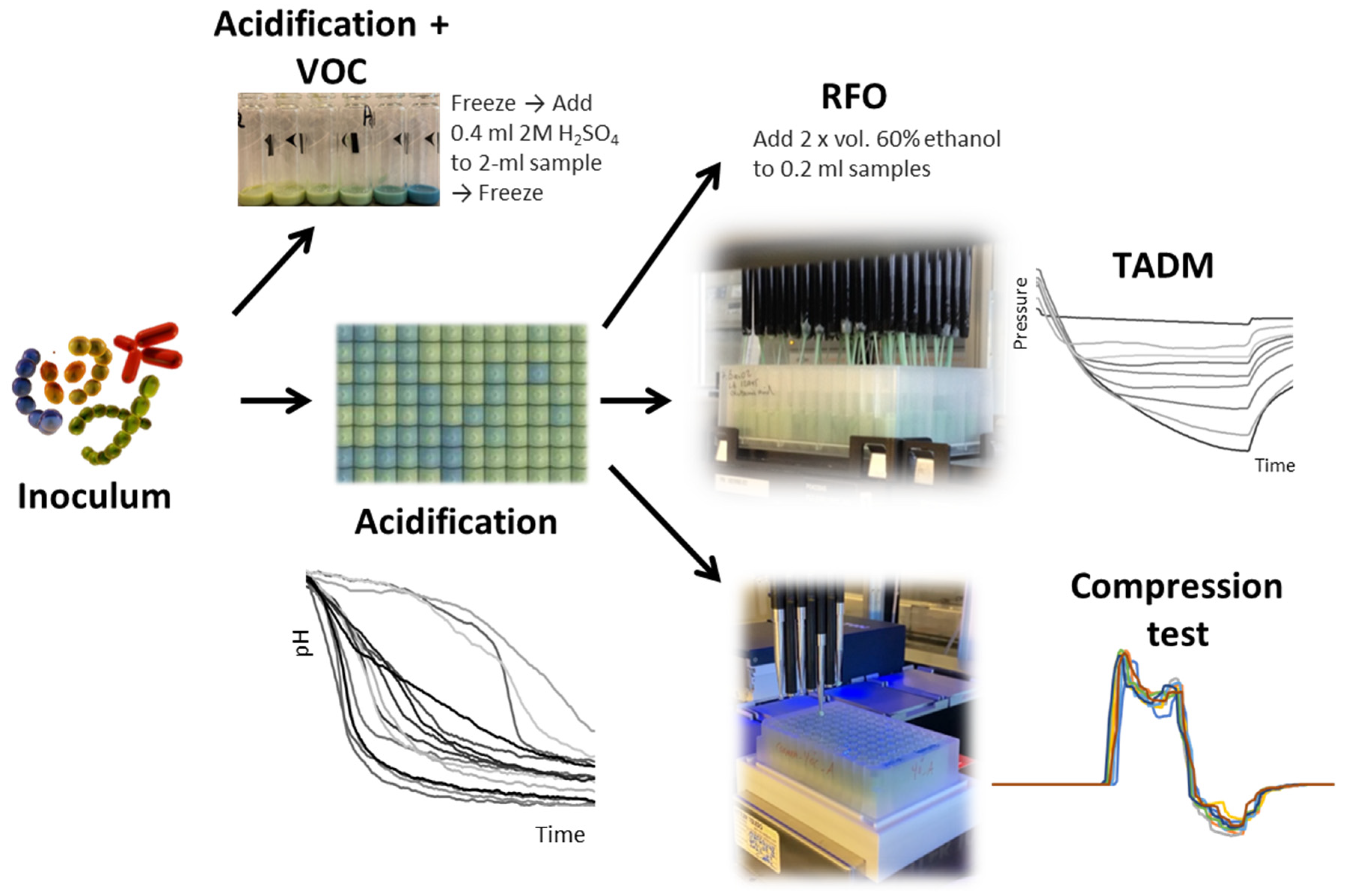
Figure 2.
Texture of legume matrices fermented by combinations of a LAB strain and a B. subtilis at two different temperatures, 30 or 37°C. Texture were measured using compression test (left panel) and TADM (right panel) for the four matrices indicated in the graphs. “Control” represents matrices fermented using B. subtilis alone.
Figure 2.
Texture of legume matrices fermented by combinations of a LAB strain and a B. subtilis at two different temperatures, 30 or 37°C. Texture were measured using compression test (left panel) and TADM (right panel) for the four matrices indicated in the graphs. “Control” represents matrices fermented using B. subtilis alone.
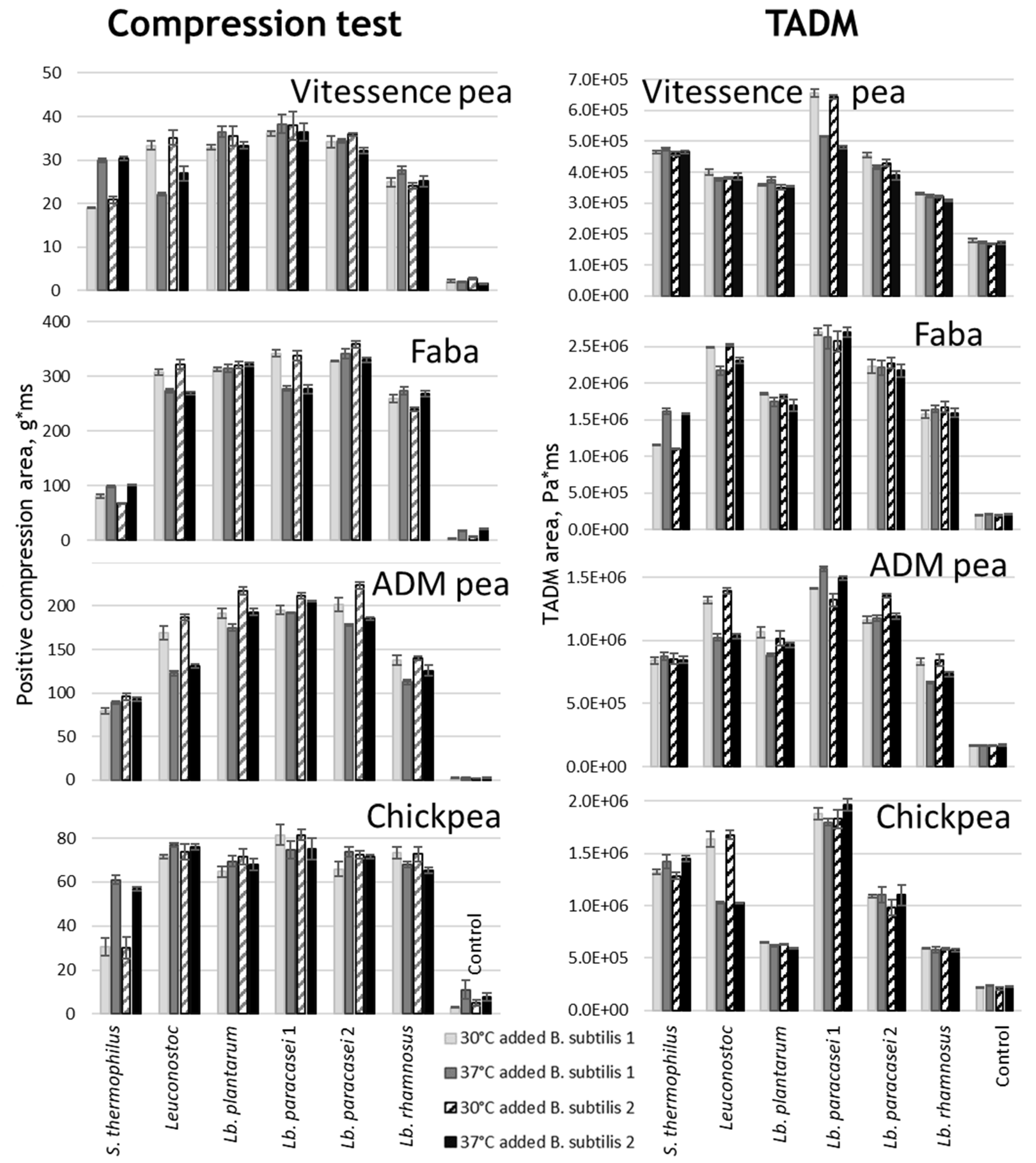
Figure 3.
Examples of B. subtilis - LAB combinations with a potential for flavor improvement. Each stacked column represents the sum of the averaged values from the four matrices based on two biological replicates. Diacetyl and hexanal showed a RSD value for the QC of 16% and 7%, respectively.
Figure 3.
Examples of B. subtilis - LAB combinations with a potential for flavor improvement. Each stacked column represents the sum of the averaged values from the four matrices based on two biological replicates. Diacetyl and hexanal showed a RSD value for the QC of 16% and 7%, respectively.
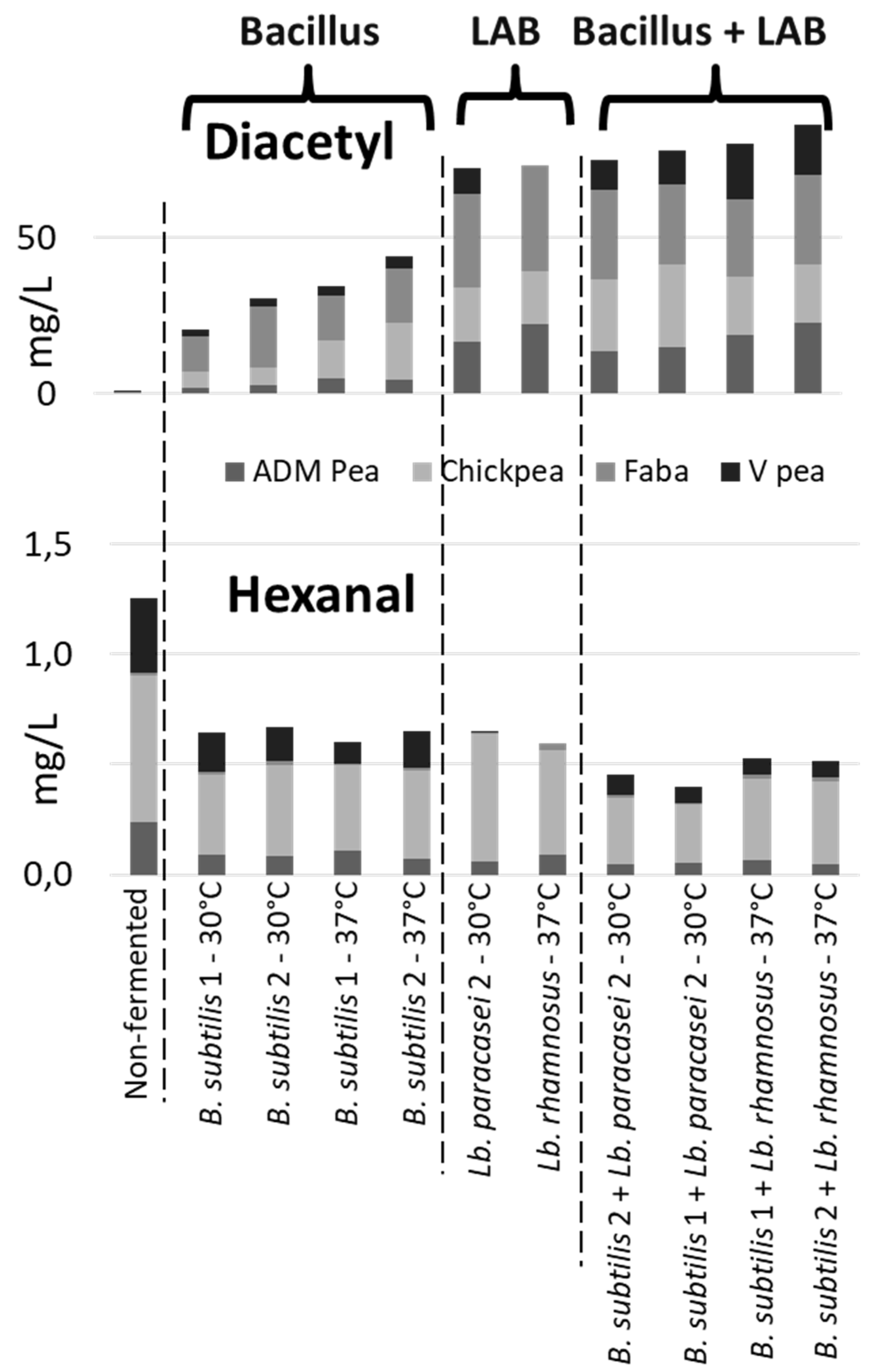
Figure 4.
Carbohydrate levels in legume matrices fermented at two different temperatures, 30 or 37°C, compared to the non-fermented matrix. Errors based on 3 biological replicates in each matrix were between 0.01 and 0.45 for glucose, 0.00 and 0.94 for sucrose, 0.00 and 0.01 for melibiose, 0.00 and 0.01 for raffinose, 0.00 and 0.08 for stachyose, and 0.00 and 0.37 for vesbascose.
Figure 4.
Carbohydrate levels in legume matrices fermented at two different temperatures, 30 or 37°C, compared to the non-fermented matrix. Errors based on 3 biological replicates in each matrix were between 0.01 and 0.45 for glucose, 0.00 and 0.94 for sucrose, 0.00 and 0.01 for melibiose, 0.00 and 0.01 for raffinose, 0.00 and 0.08 for stachyose, and 0.00 and 0.37 for vesbascose.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



