
Submitted:
01 July 2024
Posted:
01 July 2024
You are already at the latest version
Abstract
Recent omics technologies have revolutionized the study of molecular plant responses to air pollution, overcoming previous limitations. This review synthesizes the latest advancements in molecular plant responses to major air pollutants, emphasizing ozone (O3), nitrogen oxides (NOX) and particulate matter (PM) research. These pollutants induce stress responses common to other abiotic and biotic stresses, including the activation of reactive oxygen species (ROS)-scavenging enzymes and hormone signaling pathways. New evidence showed the central role of antioxidant phenolic compound biosynthesis via phenylpropanoids pathway in air pollution stress responses. Transcription factors like WRKY, AP2/ERF, and MYB, which connect hormones signaling to antioxidant biosynthesis, were also affected. To date, research has predominantly focused on laboratory studies analyzing individual pollutants. This review highlights the need for comprehensive field studies and the identification of molecular tolerance traits, which are crucial for the identification of tolerant plant species aimed to the development of sustainable Nature Based Solutions (NBS) to mitigate urban air pollution.
Keywords:
plant species
; ozone
; nitrogen dioxide
; particulate matter
; photosynthesis
; reactive oxygen species
; phenylpropanoids
; transcription factors
1. Introduction
Air pollution is defined as the pollution of indoor or outdoor environments caused by chemical, physical, or biological factors that alter the natural characteristics of the atmosphere [1]. Since 19th century, the global trend toward urbanization and industrialization has been dramatically accompanied by a detrimental increase in air pollution [2]. Latest release of the World Health Organization (WHO) Ambient Air Quality Database indicates that almost the entire global population (99%) breathes air exceeding quality limits (World Health Organization, 2021, 2024), which lead to augmented health risks. Adverse effects of air pollution on human health are associated with increased mortality linked to cardiovascular and respiratory diseases, including infections related to respiratory tract [1].
Air pollution is a mixture of hazardous substances from both anthropic activity and natural sources. Traffic and mobility, industrial production and heating systems are the main source of urban air pollution [1].
A general classification divides air pollutants in primary and secondary pollutants, depending on whether they are directly discharged in the atmosphere or derive from the interaction between primary pollutants and other molecules present in the air. Primary pollutants include gaseous molecules such as sulphur oxides (SOX) and nitrogen oxides (NOX) that are mainly generated from the combustion of different substrates, for instance fossil fuels. Further primary pollutants include the particulate matter (PM), a heterogeneous solid–liquid mixture composed of various particles of different size and origins that was shown to be tightly related to health issues in cities [4]. Sulphur dioxide (SO2), NOX and PM are among the main pollutants found in urban areas (Figure 1). Sunlight triggers photochemical reactions between primary pollutants in the atmosphere, such as carbon monoxide (CO), volatile organic compounds (VOCs) and NOx, resulting in the production of secondary pollutants, such as ground-level ozone (O3), sulfuric (H2SO4), nitric acid (HNO3), secondary VOCs and PM, increasing the complexity of air pollution and resulting in acid rain formation, further affecting air quality and human health [1,3].
According to their aerodynamic diameter, PM is divided into coarse particles (≤10 μm, PM10), fine particles (≤2.5 μm, PM2.5) and ultrafine particles (≤0.1 μm, PM0.1) [5]. Among the different air pollutants, PM is the most widespread due to its longer atmospheric duration and its presence in both urban and rural areas [1,3].
Particulate matter could contain different chemical elements including heavy metals (HMs) such as lead (Pb), copper (Cu), cadmium (Cd), chromium (Cr), mercury (Hg), arsenic (As) and antimony (Sb), mainly deriving from agriculture, industrial activities and traffic emissions [6,7]. For instance, analysis of road dusts collected in urban parks of Beijing (China) revealed the presence of nickel (Ni), Zinc (Zn), Cr, Cu, Cd and Pb at a concentration of 25.97, 219.20, 69.33, 72.13, 0.64 and 201.82 mg kg-1, respectively [8]. Furthermore, an extensive study conducted in 210 locations in 16 countries showed that the main components of PM2.5 were sulphate, nitrate, ammonium, black carbon, organic carbon, mineral dust and sea salt, which are associated with increased health risk [9].
Diesel exhaust (DE) is a complex mixture of gases and fine particles emitted from diesel engines in vehicles, as well as off-road diesel engines used in agricultural, maintenance, and construction equipment [10]. Diesel exhaust significantly contributes to urban pollution and health diseases since it contains various toxic chemicals including CO, CO2, SO2, NOX, aldehydes (formaldehyde, acrolein, acetaldehyde), benzene, polycyclic aromatic hydrocarbons (PAHs) and PM (Partial List of Chemicals Associated with Diesel Exhaust https://www.osha.gov/diesel-exhaust/chemical).
Urban and rural vegetation acts as a natural sponge absorbing pollutants from air. Plants morphological and physiological features, like the extended leaf area and the microstructure on leaf surface, promote PM deposition on leaves [11,12]. However, air pollution may affect plant health inducing different phytotoxic responses depending on the physical and chemical features of the pollutant. Foliar injury, pigment loss, premature senescence, and decreased photosynthetic/growth rates, are the main symptoms induced by O3 and NO2 in plants. Particulate matter accumulation on leaves can alter their optical properties, affecting the absorption and reflection of photosynthetically active radiation (PAR), clogging stomata, reducing photosynthesis and respiration, and ultimately decreasing plant growth and yields due to disrupted stomatal movement [13,14]. Depending on physical and chemical composition, PM may induce different phytotoxic responses in plants. Additionally, PM can increase leaf surface temperature and indirectly impact plant health by disturbing beneficial microbial communities in the phyllosphere and leaf endosphere, thus negatively affecting plant growth and health [15,16].
Air pollution may also affect plants growth inducing a plethora of stress responses, including oxidative stress that is among the most common effects caused by air pollution and triggers the deregulation of reactive oxygen species (ROS)-scavenging enzymes, such as catalase (CAT), superoxide dismutase (SOD), ascorbate peroxidase (APX) and peroxidase (POD). At the same time, the oxidative stress is generally counteracted by an enhanced biosynthesis of antioxidant molecules, flavonoids derived from phenylpropanoid pathway [17].
So far, most of the research has been conducted in laboratory conditions, focusing on the effects of individual pollutants and rarely analysing the impact of their combined presence. Few recent studies investigated the effect of air pollution on plants molecular mechanisms, performing the analysis on vegetation grown in urbanised and polluted areas. Thus, new data is expected for both laboratory and field scale, to unveil the effects induced by the combination of various air pollutants and the related plant molecular responses that could drive adaptation to air pollution. Several plant species evolved tolerance mechanisms towards air pollution paving the way for an assigned application of sustainable Nature-Based Solutions (NBS) to mitigate air pollution negative effects in urban areas. Indeed, the implementation of vegetation in the form of green infrastructure (GI) is a promising strategy recently implemented worldwide. The GI take advantage of the ability of plants to absorb, adsorb, accumulate and degrade atmospheric pollutants [18]. Therefore, the study of the molecular mechanisms activated by plants in response to air pollution and the identification of the molecular tolerance traits exhibited by different plant species, become of utmost importance to provide refined tools to support the diffusion of the GI.
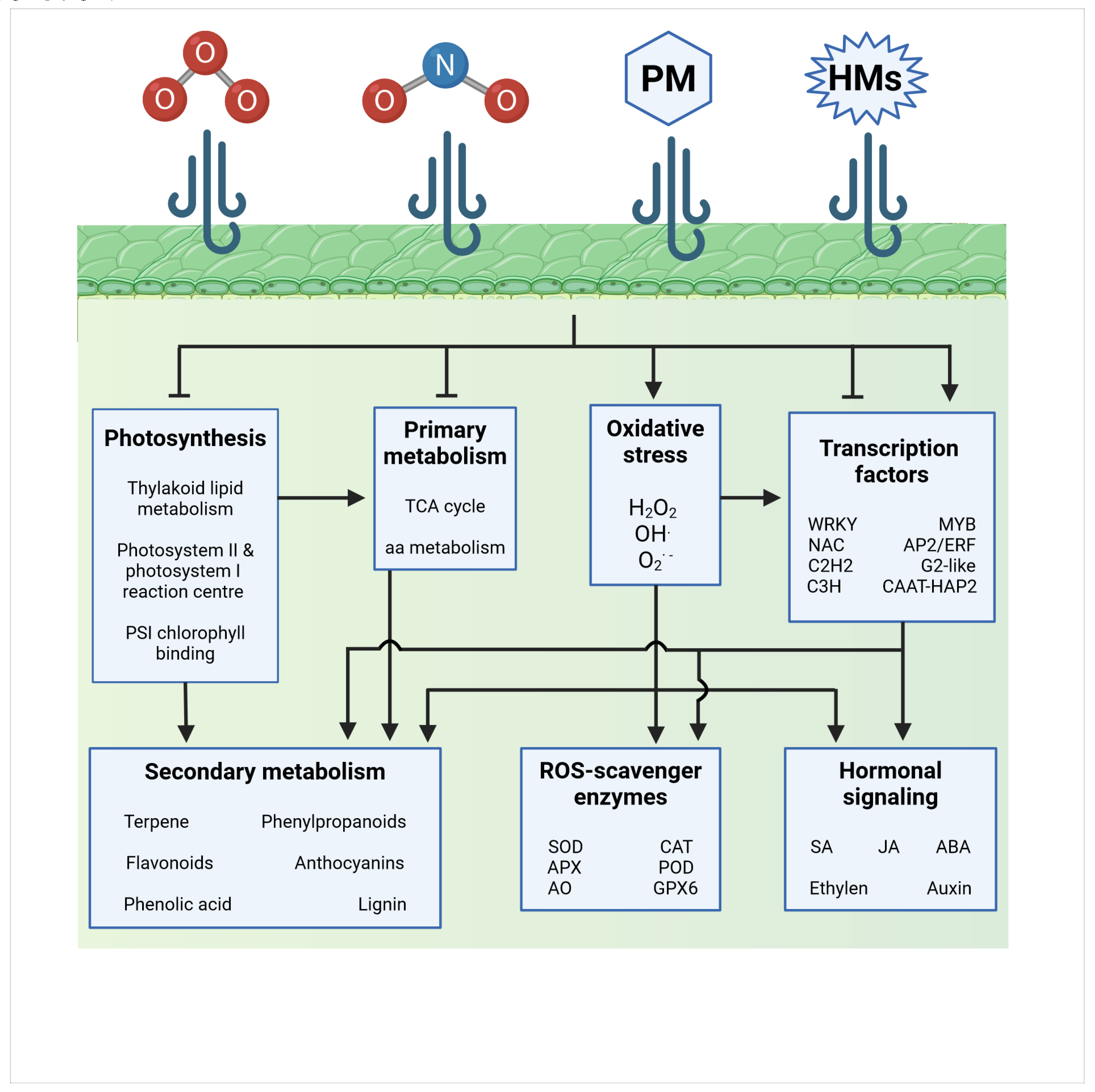
Adaptation of tolerant plants to air pollution involves a complex regulation of phytohormones signalling pathways, such as that of abscisic acid (ABA), jasmonic acid (JA), salicylic acid (SA), ethylene, cytokinin and auxin, which are generally involved in plant responses to abiotic and biotic stress [19,20,21,22]. Besides, several transcription factors (TFs) involved in stress responses, such as WRKY (named after WRKYGQK heptapeptide at the N-terminal end), MYB (first identified as oncogene derived from avian MYeloBlastosis virus), APETALA2/ethylene responsive factor (APA2/ERF), are also involved in air pollution responses, activating gene expression and bridging the signalling pathway of different hormones, like ABA and JA, induced by stresses [23]. Amino acid metabolism has also a primary role in stress responses, indeed phenylalanine is used as a substrate by enzymes involved in phenylpropanoids production, like phenylalanine ammonia-lyase (PAL) and cinnamate 4-hydroxylase (C4H). Downstream of PAL and C4H, chalcone synthase (CHS) catalyses the first step in the flavonoid pathway (Figure 2). Interestingly, in-depth analysis of gene expression uncovered recurring patterns, outlining the presence of conserved mechanisms activated in plants to cope with air pollution.
This review focuses on latest findings in the study of the effects of air pollution on plant growth and molecular responses induced in plant species. The bibliographic research highlighted that: i) O3, NOx, PM2.5, PM10 and HMs are the most studied pollutants; ii) most of the studies are based on fumigation experiments; iii) field analysis are limited so far. However, the small number of evidence and the variety of experimental approaches result in a matrix of fragmented information that need to be further investigated to fill the gaps. Besides laboratory scale experiments, future studies should include open field trials, which could lead to the identification of deregulated pathways and tolerance traits in different plant species and within the same species.
2. Air pollution induces similar but not overlapping molecular responses in plants
In the past decades, several studies attempted to investigate the molecular responses induced in plants by air pollution; however, most of the research were conducted before the advent of modern high-throughput technologies, thus several gaps in knowledge still need to be fulfilled. Nowadays, omics approaches offer the possibility to address unsolved questions regarding the specific responses induced in plants by air pollution and may help in the selection of plants for the mitigation of air pollution. Most of the findings highlighted that air pollutants, such as O3, NO2 and PM, activated general stress responses common to abiotic and biotic stress in plants, such as the induction of ROS-scavenger enzymes [24,25,26]. Recent transcriptomics, proteomics and metabolomics analysis shed light on the consequences of air pollution on plant growth, providing insightful cues for the identification of useful air pollution tolerance traits (Table1).
Most of the pollutants enhanced the formation of ROS, such as hydrogen peroxide (H2O2), superoxide ion (O2· -) and hydroxyl radical (OH.) and strongly activating oxidative stress-responsive pathways. Among them, the expression and activity of ROS-scavenger enzymes often resulted impaired, as well as the phenylpropanoid biosynthetic pathway that is crucial for the synthesis of antioxidant phenolic compounds, such as flavonoids.

Table 1.
Summary of omic techniques and affected pathways induced by air pollution in plant species.
Table 1.
Summary of omic techniques and affected pathways induced by air pollution in plant species.
| Species | Experimental set-up |
Pollutant | Exposure | Omics Platform | Enriched Pathways | Reference |
|---|---|---|---|---|---|---|
|
Arabidopsis thaliana L. |
Controlled environment growth chambers |
O3 350–423 nL L–1 |
2-6 h | Transcriptomics (RNA-seq) | Photosynthesis Response to SA Response to ROS Response to JA Response to ethylene ABA signalling pathway |
[23] |
|
Medicago truncatula L. |
Controlled environment growth chambers |
O3 70 nmol mol-1 |
6 h d-1 for 6 d | Transcriptomics (Microarray hybridization) |
Phenylalanine biosynthesis Sugar metabolism Photosynthetic electron transport Responses to inorganic substances |
[30] |
| Malus L. | Open-top growth chamber |
O3 300 nL L−1 |
3 h | Transcriptomics (RNA-seq) Metabolomics (ultra-performance liquid chromatography coupled to tandem mass spectrometry UPLC MS/MS) |
Chloroplast thylakoid membrane Chloroplast photosystem I H2O2 dehydratase activity Chalcone synthase activity Flavonoid metabolism Hormone pathways |
[31] |
|
Pisum sativum L. Glycine max L. Phaseolus vulgaris L. |
Controlled environment growth chambers |
O3 ~151.2 nL L−1 |
8 h d-1 for 45 d | Transcriptomics (RNA-seq) | Phenilpropanoid metabolism Ascorbate–glutathione cycling Glycolysis TCA cycle |
[32] |
|
Rosa hybrida L. |
Controlled environment growth chambers |
O3 80 ppb |
10 h | Transcriptomics (RNA-seq) | Phenylpropanoid biosynthesis, Starch and sucrose metabolism Sesquiterpenoid biosynthesis Triterpenoid biosynthesis. |
[33] |
|
Abies religiosa Schltdl. & Cham. |
Urban environment |
O3 87-170 ppb |
3 years | Transcriptomics (RNA-seq) | Carbohydrate metabolism Gene regulation Transcription factors Defense regulation Terpenes |
[36] |
|
Bougainvillea Spectabilis Willd. |
Controlled environment growth chambers |
NO2 8 μL L− 1 |
8 h | Metabolomics (UPLC) | Biosynthesis of amino acids Phenylalanine metabolism Phenylpropanoid biosynthesis Starch and sucrose metabolism Glutathione metabolism TCA cycle |
[37] |
|
Arabidopsis thaliana L. |
Controlled environment growth chambers |
O3 350 ppb NO2 10-30 ppm |
6 h O3 1 h NO2 |
Transcriptomic data Microarray data |
Pathogen resistance Cell death Ethylene signalling |
[38] |
|
Ambrosia artemisiifolia L. |
Controlled environment growth chambers |
O3 NO2 40-80 ppb |
61 d | Transcriptomics (RNA-seq) | Jasmonic acid pathway Response to ethylene stimulus Response to auxin stimulus Abscisic acid signalling pathway |
[39] |
|
Wrightia religiosa (Teijsm.&BINN.)Hook.F. |
Controlled environment growth chambers |
PM 470–500 μg m-3 |
24 h | Proteomics (LC-MS/MS) | Photosynthetic proteins |
[44] |
| Sansevieria trifasciata (Dracaena trifasciata Prain.) | Controlled environment growth chambers |
PM Up to 980 μg m-3 |
24 h | Proteomics (LC-MS/MS) Metabolomics |
Precursor metabolites Photosynthesis Alternative carbon metabolism Brassinosteroid signalling Stress-related proteins Metal and cadmium ion stimuli |
[43, 45] |
|
Laurus nobilis L. |
Urban environment |
PM up to 150 μg m-2 |
3 months | Transcriptomics (RNA-seq) |
Primary metabolism Secondary metabolism Hormone-related pathways Environmental stress response Transcription factors |
[41] |
|
Photinia x fraseri L. |
Urban environment |
PM up to 150 μg m-2 |
3 months | Transcriptomics (RNA-seq) |
Leaf primary metabolism Biotic stress response Abiotic stress response Cell cycle and cell division Transcription factors |
[40] |
2.1. Ozone
The Effects and responses induced by O3 in plants have been extensively investigated in both model and non-model plant species through fumigation experiments. Studies conducted on different Arabidopsis thaliana L. accessions provided significant information on responses induced upon short-term exposure (2-6 hours). Compared to control plants treated with 10-20 nL L−1 O3, treatments using 350-423 nL L−1 O3 revealed that the ecotype Columbia (Col) was tolerant to O3, while the ecotypes Shahdara (Sha) and Cape Verde islands (Cvi) displayed significant signs of leaf damages, such as reduced photosynthetic performance and cell death. Transcriptomic data showed that the three accessions shared the upregulation of several genes related to hormones signalling, including SA, JA ethylene and ABA, which are typically involved in responses induced by oxidative stress. Interestingly, genes coding for H2O2 catabolism, such as CAT and SOD, were downregulated in Sha and Cvi, while those involved in flavonoid biosynthesis, such as Phenylalanine Ammonia-Lyase (AtPAL) and Cinnamate-4-Hydroxylase (AtC4H) were upregulated in Col, indicating that the upregulation of ROS-scavenger enzymes and flavonoid biosynthesis are at the base of the tolerance in Col.
In addition, O3 enhanced the expression of several members of the TF families WRKY, AP2/ERF and MYB, which are known to bind to the promoters of O3-responsive genes [23]. WRKY family Transcription factors are involved in the stress-induced signalling cascade of JA and SA, AP2/ERFs participate in responsive mechanisms to various stresses, hormone signal transduction and metabolite regulation, while MYBs have been shown to be key factors in the biosynthesis of secondary metabolites in plants, including anthocyanins, flavonols and lignin, in response to multiple abiotic stresses [27,28,29].
Detrimental effects were observed in 50-days-old Medicago truncatula Gaertn. plants treated with 70 nmol mol-1 O3 for 6 hours per day for 6 days, compared to controls grown using environmental O3 concentration (~40 nmol mol-1). After fumigation with O3, M. truncatula showed significant decrease in photosynthetic performance, increase in ROS production, upregulation of genes coding for the transcription factors WRKY42, WRKY50 and MYB62, and genes related to JA signalling [30]. These results confirm that, similarly to other stresses, WRKYs expression and JA signalling cascade are tightly connected also during responses to air pollution stress.
Interestingly, comparable results were obtained exposing apple plants (Malus L. crabapple cv. Hongjiu) to 300 nL L−1 O3 for 3 hours in an open-top chamber, which best simulate environmental conditions. Leaves from O3 treated plants displayed significant foliar damages, reduced chlorophyll content, about 60% increased anthocyanin amount compared to controls, and increased SOD and POD activity [31]. Similarly to A. thaliana, O3 induced the expression of ERF genes in apple plants too. Among several WRKY deregulated genes, WRKY75 expression increased in plants treated with O3, as well as that of McC4H, dihydroflavonol-4-reductase (McDFR) and anthocyanidin reductase (McANR), which code for pivotal enzymes for the flavonoid pathway. Accordingly, metabolomic analysis reported an increased amount of metabolites involved in the phenylpropanoids and hormones biosynthetic pathways, confirming data from transcriptomic analysis [31].
Long-term exposure to high levels of O3 induces the upregulation of the entire phenylpropanoid biosynthetic pathway in leaves of garden pea (Pisum sativum L.), soybean (Glycine max L. Merr.) and common bean (Phaseolus vulgaris L.) plants, under controlled conditions [32]. This pathway is involved in the synthesis of different antioxidant molecules, such as flavonoids and phenolic acids, which contribute to the defence mechanisms against oxidative stress. After 45 days treatment with a O3 concentration of about 151.2 nL L−1, key enzymes for phenylpropanoid metabolism, including PAL, chalcone synthase (CHS), isoflavone reductase (IFR) and DFR, showed increased transcript abundance in all three legume species compared to control (O3 concentration ~ 12.5 nL L−1). Accordingly, phenolic content in leaves increased upon O3 treatment. However, none of the garden pea varieties investigated exhibited a negative physiological response, such as reduced photosynthetic activity, indicating that these species hold mechanisms that promote tolerance to O3. Conversely, soybean and common bean exposed to O3 displayed damages on leaves and increased expression of ascorbate oxidase (AO), which negatively affected the amount of ascorbate, thus reducing the amount of active antioxidant molecules. Interestingly, expression of genes coding for ROS-scavenger enzymes was mainly unaffected in garden pea, while glutathione peroxidase 6 (GPX6) and SOD2 transcript levels increased in soybean and common bean, indicating an active oxidative stress response in the latter plant species [32].
A recent study evaluated tolerance to O3 using four rose cultivars (Rosa hybrida L. cv. “Schloss Mannheim,” “Iceberg,” “Lüye,” and “Spectra”) grown in open-top chambers and treated with unfiltered air supplemented with 40 and 80 ppb O3 for up to 120 days. Out of the four cultivars, only Schloss Mannheim was sensitive to O3, showing detrimental effects including foliar injury, reduced chlorophyll content and reduced net photosynthetic rate [33]. In this cultivar compared to others, the expression of several heat stress transcription factors (HSF) - which mediate responses to several abiotic stresses [34] - WRKY and MYB, such as HSF24, WRKY42, WRKY75, MYB36 and MYB62, was upregulated by exposure to O3 (H. Wang, Li, et al., 2023).Besides, the expression of the ABA-related genes NCED1, PP2Cs, PYR/PYL and UGTs increased in Schloss Mannheim, sustaining the hypothesis that during O3 stress, ABA acts as a developmental signal integrating responses from different pathways [33].
A field study on the coniferous Abies religiosa ([Kunth] Schlechtendahl et Chamisso) trees grown in an area sensitive to O3 contamination in Mexico City (Mexico), evaluated the effect of O3 on leaves during different periods of the year. Each timepoint was characterized by different O3 concentrations, corresponding to moderate (87 ppb), intermediate (120–94 ppb) and high (170 ppb). Histologic, metabolomic and transcriptomic analysis revealed that within individuals from the same plant species, different tolerance mechanisms were visible. Symptomatic leaves showed thicker epidermis and collapsed cells in the palisade parenchyma compared to asymptomatic ones. The expression of the peroxidase 72 (POD72) and that of several TFs of the NAC family - commonly involved in multiple stresses such as drought, high salinity, and in ABA and JA signalling [35] - were upregulated in symptomatic trees. Genes coding for flavonoids, such as flavonol synthase (FLS) and IFR, were downregulated in symptomatic trees, while the expression of genes related to the terpene biosynthesis and BVOC emission, was induced in asymptomatic leaves. Metabolomic analysis confirmed the deregulation in genes involved in terpene metabolism, showing significant differences in terpene composition among individuals, particularly in sesquiterpenes such as β-pinene, δ-cadinene and β-caryophyllene. Sesquiterpenes are involved in the degradation of ROS and were higher in asymptomatic trees [36].
Overall, the fast induction of the ROS detoxifying machinery highlighted its key role as a first line defensive response to air pollutants in plants. However, the trend in the increasing ROS-scavenger enzymes and antioxidant production was not detected after prolonged exposure (> 72 hours) or high doses of specific pollutant (> 200 ppm) suggesting that these mechanisms may participate to early responses to air pollution [24,25,32].
2.2. Nitrogen Dioxide
Increasing concerns related to air quality in urban areas are rising due to the high amount of nitrogen dioxide (NO2) generated by anthropic activities. However, few studies investigated the impact of this molecule on plant development, with most of them focusing on injury symptoms, physiological effects and photosynthetic performance. Recent research on Bougainvillea spectabilis Willd. seedlings exposed to short-term high-concentration fumigation with up to 8 μL⋅L-1 NO2 for 8 hours, showed the induction of yellow-brown spotting on the leaves, which was likely related to oxidative stress [37]. Indeed, POD, SOD and CAT activity was significantly increased in seedlings treated with NO2 compared to controls, suggesting the activation of ROS-induced stress responses. In addition, metabolomic analysis highlighted significant differences in metabolites related to flavonoid and stilbene biosynthesis, amino acids metabolism and tricarboxylic acid (TCA) cycle among treated and control plants, supporting the hypothesis for an increased oxidative stress upon NO2 exposure in B. spectabilis [37].
Details regarding the effects of air pollution on gene expression in plants were provided by an in-depth study on approximatively 372 different accessions from A. thaliana plants treated with up to 30 ppm of NO2 for 1 hour or up to 400 ppm of O3 for 2-6 hours. Depending on the accession, plants displayed different degrees of tolerance to O3 and NO2, which were investigated by genome wide association study (GWAS). Comparison of transcriptomic and microarray data revealed that O3 and NO2 induced similar expression responses that included genes involved in ROS metabolism, ethylene signalling, pathogen resistance and cell death [38]. Transcripts for marker genes related to JA and ethylene signalling, such as cooperatively regulated by ethylene and jasmonate 1 (CEJ1), and to SA signalling, like glutaredoxin 480 (GRX480) and flavin dependent monooxygenase 1 (FMO1), were significantly more abundant in plants treated with NO2 and O3 compared to controls. Conversely, the expression of respiratory burst oxidase homolog F (RBOH), which encodes for a NADPH oxidases involved in ROS synthesis, was upregulated upon O3 treatment while decreased after NO2 treatment. [38].
Consequences of NO2 and O3 were further investigated during pollen development, with the aim of better understanding the transcriptional changes in pollens from allergenic plant species, such as the common ragweed (Ambrosia artemisiifolia L.), upon NO2 and O3 treatments [39]. Ragweed plants were exposed to a long-term fumigation (61 days) with 40 ppb (control) and 80 ppb NO2 (treatment), as well as 40 ppb (control), 80 ppb and 120 ppb O3 (treatments), thus pollen was collected. Transcriptomic data reported a significant enrichment in Gene Ontology (GO) terms related to responses to abiotic and biotic stress. Jasmonic acid biosynthetic processess and cell homeostasis of phosphate were found to be affected by higher levels of NO2 and O3. In treated pollens, GO terms including responses to ethylene stimulus, ABA and auxin signalling pathways were highly enriched mainly in upregulated transcripts. Interestingly, this study reported a significant increase in pollen allergen transcript amounts following NO2 fumigation. Conversely, a significant decrease of allergen transcripts was found in pollens treated with high levels of O3, supporting the notion that NO2 and O3 regulate the expression of the same genes in an opposite way [39].
2.3. Particulate matter and heavy metals
Among the air pollutants found in highly urbanized areas, PM have a high-risk health potential, thus deciphering how plants cope with this stress could provide insightful information to identify tolerant plants, which may help in mitigating air pollution.
Recently, a transcriptomic analysis performed on leaves from the ornamental shrub Photinia x fraseri Dress. and Laurus nobilis L. grown for three months in a rural area and near to a busy road in the city of Altopascio (Lucca, Italy), demonstrated a significant deregulation of the genes involved in different pathwys such as TCA, phenylpropanoids and phenols biosynthesis, PSI and PSII assembly, ROS production, brassinosteroid signalling pathways in response to air pollution [40,41]. The rural and urban area were characterized by a PM10 concentration up to 233.93 and 266.32 μg m-3, while that of PM2.5 reached up to 133.06 and 185.48 μg m-3, respectively. According to previous data, Photinia and Laurus plants grown along the road showed a significant impairment in key genes involved photosynthesis as well as in those involved in the TCA cycle. Expression of succinate dehydrogenase [ubiquinone] iron-sulphur subunit 1 (SDH2–1), Succinate dehydrogenase [ubiquinone] flavoprotein subunit 2 (SDH1–2), malate synthase (MLS), isocitrate lyase (ICL) and Glycine decarboxylase complex (GDCH), was down-regulated in plants exposed to high PM concentration. A clear downregulation was observed for genes related to desaturase enzymes including fatty acid desaturase family protein, 16:0delta9 desaturase 2, delta 9 desaturase 1 and Acyl-coenzyme a desaturase-like2, that play pivotal roles in thylakoid lipid metabolism and could affect the photosynthetic machinery of both Photinia and Laurus. Genes coding for several enzymes involved in the biosynthesis of phenylpropanoids and phenols, such as cinnamyl alcohol dehydrogenase homolog, was downregulated in both plant species grown in the urban area. Conversely, plants from the rural area showed an up-regulation of genes involved in terpene and phenylpropanoid-related pathways such as terpene synthase 14, flavin-monooxygenase glucosinolate s-oxygenase 5, nicotinamidase 3 and cinnamyl-alcohol dehydrogenase, as well as an upregulation of genes coding for cytochrome B5 isoform C (CB5-C), CAT2, CAT3 and peroxiredoxin type 2 and APX5. These data suggest that ROS-scavenging enzymes are more effective in low PM environments, such as rural areas, rather than highly air polluted urban centres. On the other hand, ROS scavenging could be an early stress response to high PM concentration, thus not being detected in plants long-term exposed to PM. Expression of Heat shock proteins (HSPs) - which act downstream of HSFs and regulate response to stresses (ul Haq et al., 2019) - was mainly induced in plants grown in the urban area and genes coding for HSP17.8, HSP17.6II, HSP15.7, HSPA2, and HSP70 were all upregulated, compared to plants grown in the rural area. Interestingly, pathogenesis-related genes such as putative pathogenesis-related thaumatin superfamily protein (ATLP-1) and putative basic pathogenesis-related protein 1 (ATPRB1) and Disease resistance protein (Q19e69) were induced in Photinia plants grown at high PM level [40]. Similar results were obtained for A. thaliana plants exposed to 30 mg m-3 of SO2, which showed up-regulation of generic HSPs and pathogen-related protein [25,42].
Finally, several TFs involved in plant development and stress responses, including G2-like, C3H, MYB, WRKYs, C2H2 and CAAT-HAP2, were deregulated in P. fraseri and L. nobilis in response to PM stress. On the other hand, there was no clear differential modulation of any class of hormone-related genes [40,41].
Further insights on plant responses to PM were obtained by the exposure of the ornamental plant species Wrightia religiosa (Teijsm.&BINN.)Hook.F. and Sansevieria trifasciata (Dracaena trifasciata Prain.) to burning cigarettes as a source of pollution [43,44,45]. Plants were placed in an enclosed chamber and exposed to concentration of PM1, PM2.5, and PM10 up to 900–945, 900–945, and 950–980 μg m-3, respectively, for about 7 days. Interestingly, also priming - i.e., the attitude of a plant exposed to a certain stress, to better tolerate subsequent stress treatments [46] - to PM stress in plants was investigated [43,44,45]. Proteomic analysis revealed that in W. religiosa most of the proteins related to photosystem II (PSII), photosystem I (PSI) reaction centre and PSI chlorophyll binding, were downregulated. Consistently, leaf chlorophyl content decreased after the treatment [44]. On the other hand, in S. trifasciata plants exposed to cigarette smoke, photosynthesis performance was unaffected, and an up-regulation of proteins involved in PSI and PSII assembly was observed [43]. In detail, leaves of S. trifasciata treated plants showed specifically expressed unique proteins involved in biological processes related to photosynthesis, chlorophyll-binding and electron transport chains, which were not identified in control leaves, suggesting a specific activation of the photosynthetic process in S. trifasciata exposed to air pollution. Among ROS-scavenger enzymes, only SOD amount decreased compared to other enzymes, like CAT, that were mainly unaffected in S. trifasciata. This could be due to the increased protein abundance related to brassinosteroid signalling, such as brassinosteroid insensitive 1-associated receptor kinase 1 (BAK1), which is involved in the repression of ROS production by stimulating of antioxidant activity [43]. Moreover, GO classification identified differentially abundant proteins (DAPs) involved in responses to metal ion and cadmium ion stimuli [43]. This result is consistent with previous reports of HMs presence in cigarette smoke, which is especially rich in Cd (1–2 µg g-1) [47]. In S. trifasciata leaves of treated plants, the activation of an alternative carbon metabolisms was found. Modulation of alcohol dehydrogenase (ADH), serine hydroxymethyltransferase (SHM) and glycolate oxidase (GLO) enzymes, which allow organic compound assimilation through the folate cycle to produce serine, probably enabling S. trifasciata to use absorbed PM as a carbon source [43]. Upon cigarette smoke treatment, the relative water content was found to be increased in W. religiosa leaves in agreement with the observed upregulation of the putative homolog of the aquaporin-related gene TIP2-2 in S. trifasciata, suggesting a probable increase in water channels amount in response to air pollution (Permana et al., 2023, 2024; Treesubsuntorn et al., 2021). Although this response remains partially unclear, altogether these data indicate that compared to other plant species, S. trifasciata could be a good candidate for mitigation of air pollution characterized by high PM concentration.
3. Conclusions
The raise of air pollution is increasing the risks associated with human and environmental health. The development of NBS centred on the greening of urbanized regions could be a sustainable strategy to mitigate air pollution. Unveiling the molecular mechanisms that promote plant tolerance to air pollution could provide useful insights to select appropriate plant species for strategies aimed to the improvement of air quality. Common pathways were induced in response to treatments using different air pollutants, such as the upregulation of ROS-scavenger enzymes activity and gene expression in sensitive plants. The upregulation of antioxidants molecules like flavonoids was confirmed by transcriptomic, proteomic and metabolomic analysis in different plant species, both sensitive and tolerant to air pollution stress. These data suggest a prominent role for phenylpropanoids metabolism in tolerance to air pollution. A recurring involvement of AP2/ERF, WRKY and MYB TFs families was observed along different plant species, as well as a main impairment of phytohormones signalling, including that of ABA, JA and ethylene.
Altogether, these data provide crucial tools for the evaluation of tolerance traits in plants towards several molecules that affect air quality, such as O3, NO2 and PM, posing the basis for a detailed understanding of responses induced by air pollution stress.
Author Contributions
Conceptualization, P.B., D.M. and M.L.A.; writing—original draft preparation, M.L.A., C.C. and F.M.C.; writing—review and editing, P.B. and D.M.; figure production, M.L.A. All authors have read and agreed to the published version of the manuscript.
Funding
Project funded under the National Recovery and Resilience Plan (NRRP), Mission 4 Component 2 Investment 1.4 - Call for tender No. 3138 of 16 December 2021, rectified by Decree n.3175 of 18 December 2021 of Italian Ministry of University and Research funded by the European Union – NextGenerationEU; Award Number: Project code CN_00000033, Concession Decree No. 1034 of 17 June 2022 adopted by the Italian Ministry of University and Research, CUP CUPB83C22002930006, Project title “National Biodiversity Future Center - NBFC”.
Conflicts of Interest
The authors declare no conflicts of interest
References
- World Health Organization WHO Global Air Quality Guidelines: Particulate Matter (PM2. 5 and PM10), Ozone, Nitrogen Dioxide, Sulfur Dioxide and Carbon Monoxide; World Health Organization: Geneva, 2021; ISBN 9789240034228. [Google Scholar]
- Wang, Q.; Kwan, M.P.; Zhou, K.; Fan, J.; Wang, Y.; Zhan, D. The Impacts of Urbanization on Fine Particulate Matter (PM2.5) Concentrations: Empirical Evidence from 135 Countries Worldwide. Environmental Pollution 2019, 247, 989–998. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization WHO Ambient Air Quality Database (Update 2024). Version 6.1. 2024.
- Addis Alemayehu, Y.; Leta Asfaw, S.; Alemu Terfie, T. Exposure to Urban Particulate Matter and Its Association with Human Health Risks. Environmental Science and Pollution Research 2020, 27491–27506. [Google Scholar] [CrossRef] [PubMed]
- Mühlfeld, C.; Rothen-Rutishauser, B.; Blank, F.; Vanhecke, D.; Ochs, M.; Gehr, P.; hlfeld, M.C. Interactions of Nanoparticles with Pulmonary Structures and Cellular Responses. Am J Physiol Lung Cell Mol Physiol 2008, 294, 817–829. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Zhan, C.; Liu, H.; Liu, S.; Quan, J.; Liu, X.; Zhang, J.; Qu, C. Source-Specific Health Risk of PM2.5-Bound Metals in a Typical Industrial City, Central China, 2021–2022. Atmosphere (Basel) 2023, 14. [Google Scholar] [CrossRef]
- Lee, J.K.; Woo, S.Y.; Kwak, M.J.; Park, S.H.; Kim, H.D.; Lim, Y.J.; Park, J.H.; Lee, K.A. Effects of Elevated Temperature and Ozone in Brassica Juncea L.: Growth, Physiology, and Ros Accumulation. Forests 2020, 11. [Google Scholar] [CrossRef]
- Du, Y.; Gao, B.; Zhou, H.; Ju, X.; Hao, H.; Yin, S. Health Risk Assessment of Heavy Metals in Road Dusts in Urban Parks of Beijing, China. Procedia Environ Sci 2013, 18, 299–309. [Google Scholar] [CrossRef]
- Masselot, P.; Sera, F.; Schneider, R.; Kan, H.; Lavigne, É.; Stafoggia, M.; Tobias, A.; Chen, H.; Burnett, R.T.; Schwartz, J.; et al. Differential Mortality Risks Associated With PM2.5 Components A Multi-Country, Multi-City Study. Epidemiology 2022, 33, 167–175. [Google Scholar] [CrossRef]
- Lloyd, A.C.; Cackette, T.A. Diesel Engines: Environmental Impact and Control. J Air Waste Manage Assoc 2001, 51, 809–847. [Google Scholar] [CrossRef]
- Prigioniero, A.; Zuzolo, D.; Niinemets, Ü.; Postiglione, A.; Mercurio, M.; Izzo, F.; Trifuoggi, M.; Toscanesi, M.; Scarano, P.; Tartaglia, M.; et al. Particulate Matter and Polycyclic Aromatic Hydrocarbon Uptake in Relation to Leaf Surface Functional Traits in Mediterranean Evergreens: Potentials for Air Phytoremediation. J Hazard Mater 2022, 435. [Google Scholar] [CrossRef]
- Tan, X.Y.; Liu, L.; Wu, D.Y. Relationship between Leaf Dust Retention Capacity and Leaf Microstructure of Six Common Tree Species for Campus Greening. Int J Phytoremediation 2022, 24, 1213–1221. [Google Scholar] [CrossRef]
- Lee, J.K.; Kwak, M.J.; Park, S.H.; Han Dong Kim, H.D.; Lim, Y.J.; Jeong, S.G.; Choi, Y.S.; Woo, S.Y. Ozone Response of Leaf Physiological and Stomatal Characteristics in Brassica Juncea L. at Supraoptimal Temperatures. Land (Basel) 2021, 10. [Google Scholar] [CrossRef]
- Lee, J.K.; Kim, D.Y.; Park, S.H.; Woo, S.Y.; Nie, H.; Kim, S.H. Particulate Matter (Pm) Adsorption and Leaf Characteristics of Ornamental Sweet Potato (Ipomoea Batatas l.) Cultivars and Two Common Indoor Plants (Hedera Helix l. and Epipremnum Aureum Lindl. & Andre). Horticulturae 2022, 8. [Google Scholar] [CrossRef]
- Weyens, N.; Thijs, S.; Popek, R.; Witters, N.; Przybysz, A.; Espenshade, J.; Gawronska, H.; Vangronsveld, J.; Gawronski, S.W. The Role of Plant–Microbe Interactions and Their Exploitation for Phytoremediation of Air Pollutants. Int J Mol Sci 2015, 16, 25576–25604. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Liu, Y.; Feng, S.; Liu, C.; Zhong, X.; Ren, Y.; Liu, Y.; Huang, Y.; Yang, M. The Relationship between Atmospheric Particulate Matter, Leaf Surface Microstructure, and the Phyllosphere Microbial Diversity of Ulmus L. BMC Plant Biol 2024, 24. [Google Scholar] [CrossRef]
- Huchzermeyer, B.; Menghani, E.; Khardia, P.; Shilu, A. Metabolic Pathway of Natural Antioxidants, Antioxidant Enzymes and ROS Providence. Antioxidants 2022, 11. [Google Scholar] [CrossRef]
- Prigioniero, A.; Zuzolo, D.; Niinemets, Ü.; Guarino, C. Nature-Based Solutions as Tools for Air Phytoremediation: A Review of the Current Knowledge and Gaps. Environmental Pollution 2021, 277. [Google Scholar] [CrossRef] [PubMed]
- Li, S.M.; Zheng, H.X.; Zhang, X.S.; Sui, N. Cytokinins as Central Regulators during Plant Growth and Stress Response. Plant Cell Rep 2021, 40, 271–282. [Google Scholar] [CrossRef]
- Chen, S.; Zhao, C.B.; Ren, R.M.; Jiang, J.H. Salicylic Acid Had the Potential to Enhance Tolerance in Horticultural Crops against Abiotic Stress. Front Plant Sci 2023, 14. [Google Scholar] [CrossRef]
- Chen, H.; Bullock, D.A.; Alonso, J.M.; Stepanova, A.N. To Fight or to Grow: The Balancing Role of Ethylene in Plant Abiotic Stress Responses. Plants 2022, 11. [Google Scholar] [CrossRef]
- Marzi, D.; Brunetti, P.; Saini, S.S.; Yadav, G.; Puglia, G.D.; Dello Ioio, R. Role of Transcriptional Regulation in Auxin-Mediated Response to Abiotic Stresses. Front Genet 2024, 15. [Google Scholar] [CrossRef] [PubMed]
- Morales, L.O.; Shapiguzov, A.; Safronov, O.; Leppälä, J.; Vaahtera, L.; Yarmolinsky, D.; Kollist, H.; Brosché, M. Ozone Responses in Arabidopsis: Beyond Stomatal Conductance. Plant Physiol 2021, 186, 180–192. [Google Scholar] [CrossRef] [PubMed]
- Muneer, S.; Kim, T.H.; Choi, B.C.; Lee, B.S.; Lee, J.H. Effect of CO, NOx and SO2 on ROS Production, Photosynthesis and Ascorbate-Glutathione Pathway to Induce Fragaria×annasa as a Hyperaccumulator. Redox Biol 2014, 2, 91–98. [Google Scholar] [CrossRef]
- Li, L.; Yi, H. Differential Expression of Arabidopsis Defense-Related Genes in Response to Sulfur Dioxide. Chemosphere 2012, 87, 718–724. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Yi, H. Genome-Wide Transcriptome Analysis of Arabidopsis Response to Sulfur Dioxide Fumigation. Mol Genet Genomics 2014, 289, 989–999. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Cheng, X.; Yin, D.; Chen, D.; Luo, C.; Liu, H.; Huang, C. Advances in the Research on Plant WRKY Transcription Factors Responsive to External Stresses. Curr Issues Mol Biol 2023, 45, 2861–2880. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Niu, Y.; Zheng, Y. Multiple Functions of Myb Transcription Factors in Abiotic Stress Responses. Int J Mol Sci 2021, 22. [Google Scholar] [CrossRef] [PubMed]
- Feng, K.; Hou, X.L.; Xing, G.M.; Liu, J.X.; Duan, A.Q.; Xu, Z.S.; Li, M.Y.; Zhuang, J.; Xiong, A.S. Advances in AP2/ERF Super-Family Transcription Factors in Plant. Crit Rev Biotechnol 2020, 40, 750–776. [Google Scholar] [CrossRef]
- Iyer, N.J.; Tang, Y.; Mahalingam, R. Physiological, Biochemical and Molecular Responses to a Combination of Drought and Ozone in Medicago Truncatula. Plant Cell Environ 2013, 36, 706–720. [Google Scholar] [CrossRef]
- Wu, S.; Wang, Y.; Zhang, J.; Wang, Y.; Yang, Y.; Chen, X.; Wang, Y. How Does Malus Crabapple Resist Ozone? Transcriptomics and Metabolomics Analyses. Ecotoxicol Environ Saf 2020, 201. [Google Scholar] [CrossRef]
- Yendrek, C.R.; Koester, R.P.; Ainsworth, E.A. A Comparative Analysis of Transcriptomic, Biochemical, and Physiological Responses to Elevated Ozone Identifies Species-Specific Mechanisms of Resilience in Legume Crops. J Exp Bot 2015, 66, 7101–7112. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Li, M.; Yang, Y.; Sun, P.; Zhou, S.; Kang, Y.; Xu, Y.; Yuan, X.; Feng, Z.; Jin, W. Physiological and Molecular Responses of Different Rose (Rosa Hybrida L.) Cultivars to Elevated Ozone Levels. Plant Direct 2023, 7. [Google Scholar] [CrossRef] [PubMed]
- Guo, M.; Liu, J.H.; Ma, X.; Luo, D.X.; Gong, Z.H.; Lu, M.H. The Plant Heat Stress Transcription Factors (HSFS): Structure, Regulation, and Function in Response to Abiotic Stresses. Front Plant Sci 2016, 7. [Google Scholar] [CrossRef]
- Nakashima, K.; Takasaki, H.; Mizoi, J.; Shinozaki, K.; Yamaguchi-Shinozaki, K. NAC Transcription Factors in Plant Abiotic Stress Responses. Biochim Biophys Acta Gene Regul Mech 2012, 1819, 97–103. [Google Scholar] [CrossRef] [PubMed]
- Reyes-Galindo, V.; Jaramillo-Correa, J.P.; Shishkova, S.; Sandoval-Zapotitla, E.; Flores-Ortiz, C.M.; Piñero, D.; Spurgin, L.G.; Martin, C.A.; Torres-Jardón, R.; Zamora-Callejas, C.; et al. Histologic, Metabolomic, and Transcriptomic Differences in Fir Trees from a Peri-Urban Forest under Chronic Ozone Exposure. Ecol Evol 2024, 14. [Google Scholar] [CrossRef]
- Sheng, Q.; Zhou, C.; Liang, Y.; Zhang, H.; Song, M.; Zhu, Z. Elevated NO2 Induces Leaf Defensive Mechanisms in Bougainvillea Spectabilis Seedlings. Ecotoxicol Environ Saf 2022, 248. [Google Scholar] [CrossRef] [PubMed]
- Leppälä, J.; Gaupels, F.; Xu, E.; Morales, L.O.; Durner, J.; Brosché, M. Ozone and Nitrogen Dioxide Regulate Similar Gene Expression Responses in Arabidopsis but Natural Variation in the Extent of Cell Death Is Likely Controlled by Different Genetic Loci. Front Plant Sci 2022, 13. [Google Scholar] [CrossRef]
- Zhao, F.; Durner, J.; Winkler, J.B.; Traidl-Hoffmann, C.; Strom, T.M.; Ernst, D.; Frank, U. Pollen of Common Ragweed (Ambrosia Artemisiifolia L.): Illumina-Based de Novo Sequencing and Differential Transcript Expression upon Elevated NO2/O3. Environmental Pollution 2017, 224, 503–514. [Google Scholar] [CrossRef] [PubMed]
- Vergata, C.; Contaldi, F.; Baccelli, I.; Santini, A.; Pecori, F.; Buti, M.; Mengoni, A.; Vaccaro, F.; Moura, B.B.; Ferrini, F.; et al. How Does Particulate Matter Affect Plant Transcriptome and Microbiome? Environ Exp Bot 2023, 209. [Google Scholar] [CrossRef]
- Vergata, C.; Contaldi, F.; Baccelli, I.; Buti, M.; Vangelisti, A.; Giordani, T.; Moura, B.; Ferrini, F.; Martinelli, F. The Transcriptional Mechanism Responding to Air Particulate Matter in Laurus Nobilis (L.). Environ Exp Bot 2023, 210. [Google Scholar] [CrossRef]
- Zhao, J.; Yi, H. Genome-Wide Transcriptome Analysis of Arabidopsis Response to Sulfur Dioxide Fumigation. Mol Genet Genomics 2014, 289, 989–999. [Google Scholar] [CrossRef] [PubMed]
- Permana, B.H.; Krobthong, S.; Yingchutrakul, Y.; Saithong, T.; Thiravetyan, P.; Treesubsuntorn, C. Evidence of Brassinosteroid Signalling and Alternate Carbon Metabolism Pathway in the Particulate Matter and Volatile Organic Compound Stress Response of Sansevieria Trifasciata. Environ Exp Bot 2023, 205. [Google Scholar] [CrossRef]
- Treesubsuntorn, C.; Setiawan, G.D.; Permana, B.H.; Citra, Y.; Krobthong, S.; Yingchutrakul, Y.; Siswanto, D.; Thiravetyan, P. Particulate Matter and Volatile Organic Compound Phytoremediation by Perennial Plants: Affecting Factors and Plant Stress Response. Science of the Total Environment 2021, 794. [Google Scholar] [CrossRef]
- Permana, B.H.; Krobthong, S.; Yingchutrakul, Y.; Thiravetyan, P.; Treesubsuntorn, C. Sansevieria Trifasciata’s Specific Metabolite Improves Tolerance and Efficiency for Particulate Matter and Volatile Organic Compound Removal. Environmental Pollution 2024, 355. [Google Scholar] [CrossRef] [PubMed]
- Kerchev, P.; van der Meer, T.; Sujeeth, N.; Verlee, A.; Stevens, C. V.; Van Breusegem, F.; Gechev, T. Molecular Priming as an Approach to Induce Tolerance against Abiotic and Oxidative Stresses in Crop Plants. Biotechnol Adv 2020, 40. [Google Scholar] [CrossRef] [PubMed]
- Dinh, Q.P.; Novirsa, R.; Jeong, H.; Cahya Nugraha, W.; Addai-Arhin, S.; Viet, P.H.; Tominaga, N.; Ishibashi, Y.; Arizono, K. Mercury, Cadmium, and Lead in Cigarettes from International Markets: Concentrations, Distributions and Absorption Ability of Filters; 2021.
Figure 1.
Representative image of main natural and human sources of the primary and secondary air pollutants.
Figure 1.
Representative image of main natural and human sources of the primary and secondary air pollutants.
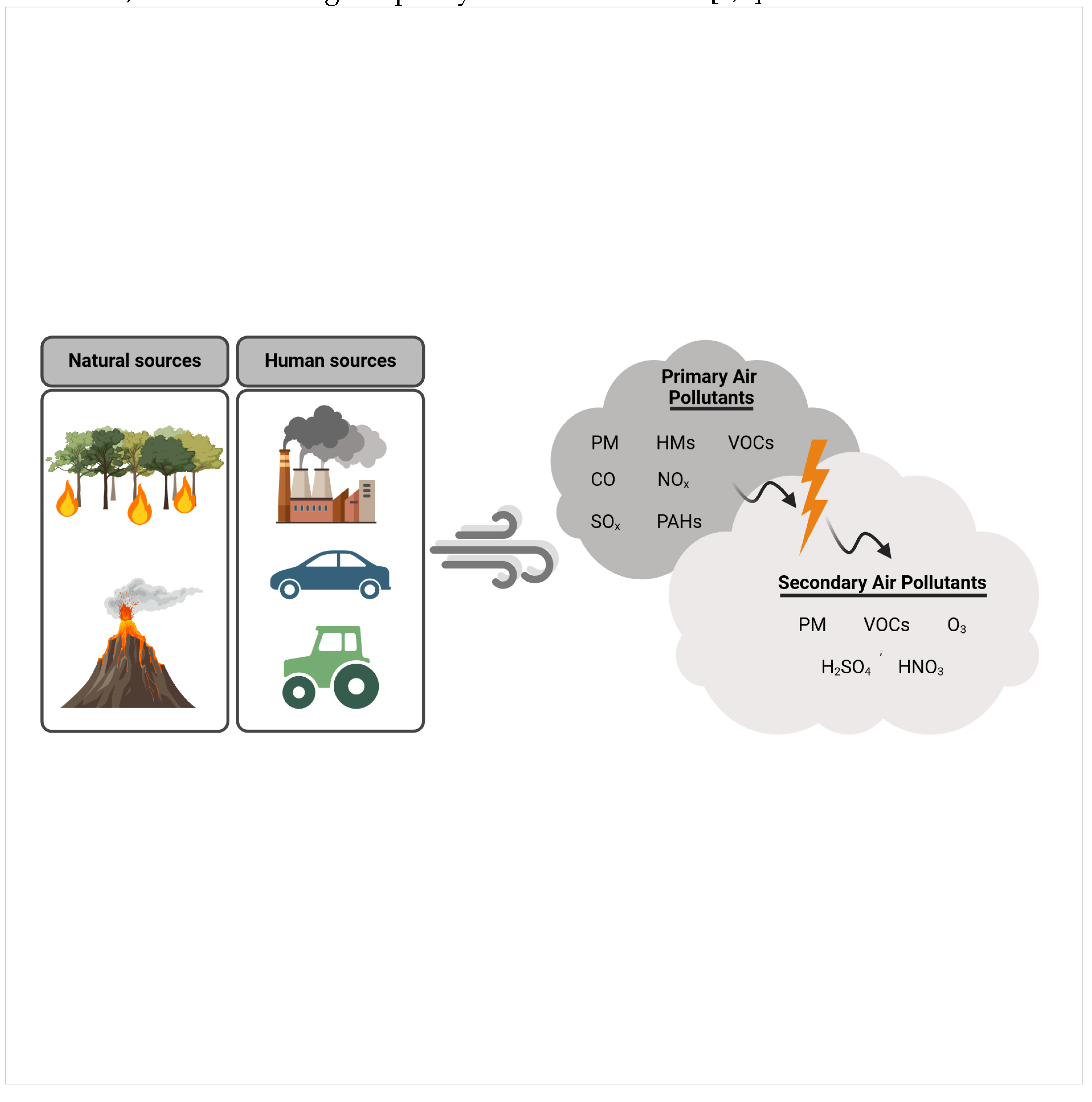
Figure 2.
Schematic representation of the main modulated pathways in response to O3, NO2, PM and HMs air pollutants.
Figure 2.
Schematic representation of the main modulated pathways in response to O3, NO2, PM and HMs air pollutants.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



