
Submitted:
01 July 2024
Posted:
02 July 2024
You are already at the latest version
Abstract
Endophytes are ubiquitous organisms that reside within plant organs and contribute to the growth and well-being of plants. Nevertheless, there is limited knowledge regarding the plant growth-promoting endophytes found in medicinal plants. Therefore, the aims of the present work were to isolate some endophytic bacteria associated with roots, stems, leaves and inflorescences of Aerva javanica, collected from the Shada Alasfal Mountain at Albaha region, Saudi Arabia. After that, they were screened for some of their direct and indirect plant growth-promoting mechanisms. Finally, based upon their internal transcribed spacer 16S rRNA gene sequencing, the isolates were identified. The results showed that the selected endophytes differentially produced IAA and ammonia, and solubilized phosphate as direct mechanisms. In addition, they had exhibited variable capacities to produce some lytic enzymes and HCN beside their antifungal activities. Furthermore, they showed abilities to tolerate some stress conditions. The bacterial endophytes were identified by molecular methods and belonged to the genera of Micrococcus, Enterobacter, Pseudomonas, Delftia and Bacillus. The findings of this study indicated that the bacterial endophytes isolated from Aerva javanica are possessing crucial functions to improve plant growth and could be utilized as biofertilizers or bioagents to establish a sustainable crop production system.
Keywords:
aerva javanica
; endophytic bacteria
; stress tolerance
; enzymes
; antifungal
1. Introduction
The medicinal plants and their valuable secondary metabolites have extensive applications as raw materials in agriculture and industry due to their high economic value. Over the past two decades, numerous research has concentrated on the endophytic microbiota by examining their presence within the organs of medicinal plants [1]. These findings contribute to the comprehension of the colonization and diversity of endophytic bacteria in medicinal plants. The arrangement and makeup of endophytic communities are influenced by environmental conditions and characteristics linked with plants, such as the genetic makeup of the plant, its stage of development, and its phenology [2]. Endophytic bacteria were identified from many phyla of bacteria, including Actinobacteria, Firmicutes, Proteobacteria, and Bacteroidetes [3]. Plant metabolites possess potent physiological actions; yet, their production is hindered by unpredictable environmental conditions and frequent climatic fluctuations, resulting in challenges for both quality and productivity [4]. Therefore, the exploration of bioactive metabolites from microorganisms through bioprospecting has emerged as a very promising alternative approach for drug development [5]. Endophytes are renowned for their role in enhancing and sustaining the health and growth of plants [6]. In turn, plants offer a diverse environment consisting of various abiotic and biotic elements that facilitate the colonization of endophytes. Nevertheless, as a result of distinct nutritional requirements throughout various phases of growth, the physiological characteristics of plants differ throughout their life cycle. [7].
The use of endophytes is following two directions which are the direct and indirect mechanisms by which these microorganisms can aid in plant health [8]. Converting endophytic bacteria into bio-factories for the sustainable production of a variety of materials using renewable sources is the goal of agricultural and industrial biotechnologies such as producing enzymes, HCN, which are considered indirect mechanisms for plant growth, and other metabolic compounds, in agriculture field, nutrition and solve environmental issues, etc. In this regard, the wide metabolic potential of endophytes has resulted in a huge attention being attracted to these microorganisms in a biotechnological perspective [1]. These systems can serve as both direct and indirect means of enhancing plant development. For instance, many strains of endophytic bacteria have demonstrated advantageous characteristics that serve as direct methods for promoting plant growth. These include nitrogen fixation, inorganic phosphate solubilization, secretion of siderophores, and production of IAA [9]. Moreover, the synthesis of antimicrobial substances is regarded as a very effective and extensively researched method employed by endophytes to restrict the growth of disease-causing microorganisms in plants. This mechanism has been widely comprehended in the last twenty years [10].
Aerva javanica is a perennial herbaceous plant that is classified under the Amaranthaceae family and is known for its therapeutic properties. This plant species exhibits a high level of adaptation that enables it to thrive in the unique environment found in the southern west parts of Saudi Arabia, which are known for their abundant plant variety [11]. The present investigation will concentrate on Shada Alasfal Mountain, situated in the Albaha region of Saudi Arabia [12]. Aerva javanica is a plant with a tall, upright, and sturdy stem that can reach a height of 1.6 meters [13]. It was studied ethno-botanically, chemically, and biologically to assess their medicinal uses, phytochemical constituents, and biological activities. In a study by Suleiman [14], the plant organs ethanolic extract had revealed weak antimicrobial activities against some pathogens. Another investigation carried out on different extracts of A. javanica revealed that methanolic extracts showed potential antibacterial activities [15]. Singh and Jha [16] had isolated Enterobacter cloaceae as an endophytic bacterium for A. javanica roots that had the ability to produce ACC deaminase activity, nitrogen fixation and ammonia production. The strain had shown the tolerance to NaCl concentrations up to 6% (w/v), growing ability up to pH 11 and temperature of 50°C. In another study, an endophytic fungus Cercospora sp. PM018 that was isolated from A. Javanica, had produced palmitic acid and stearic acid as antibacterial compounds [17].
Hence, this study was focused on the isolation and identification by molecular method of putative endophytic bacteria isolates from Aerva javanica, which is a native inhabitant of Shada Alasfal Mountain. Direct and indirect plant growth promoting (PGP) properties of the selected bacterial endophytes involving extracellular enzymes (amylase, protease, cellulase, gelatinase, lipase and esterase), IAA, HCN and ammonia productions, phosphate solubilization abilities, their antifungal activities and abilities to tolerate some stress conditions were evaluated as some beneficial roles for plant growth.
2. Materials and Methods
2.1. Plant Sampling and Study Area
Aerva javanica (Burm. f.) Juss. ex Schult. (Family Amaranthaceae) (Figure 1) was collected from Shada Al-Asfal Mountain (19°44'07.7"N 41°22'52.5"E) in Albaha region, Saudi Arabia (Figure 2). Twelve individual plant samples were collected from the site. The plant samples were transferred to the lab in sterile plastic bags and stored at 4ºC until used. After that, they were subjected to isolation procedures within 48 hrs [18]. A sample of the plant was formally identified and deposed at the herbarium of Biological Sciences Department of King Abdul-Aziz University.
2.2. Isolation of Endophytic Bacteria
Aerva javanica samples were divided into stems, roots, leaves, and inflorescences and sterilized by the five-step surface sterilization method, as outlined by Qin et al. [18] and Musa et al. [19]. To isolate endophytic bacteria from each plant organ, the organ was aseptically fragmented into small segments and placed directly on seven different agar media. These agar media included Starch Casein Agar, Starch Nitrate Agar, Humic Acid-Vitamin Agar, Tryptic Soy Agar, Tap Water Yeast Extract Agar, Inorganic Salts-Starch Agar (ISP 4), and Potato Dextrose Agar. The petri dishes were placed in an incubator set at a temperature of 28°C for a duration of 1-2 weeks, as described by Phongsopitanuna et al. [20]. To purify a single colony of bacterial endophyte, it was selected and streaked over freshly prepared Tryptic Soy Agar in order to get pure isolates.
2.3. Direct Mechanisms by Endophytic Bacteria
2.3.1. Solubilization of phosphate
The approach described by Islam et al. [21] was employed to assess the capacity of the chosen endophytes to solubilize phosphate. The isolates were inoculated onto Pikovskaya's agar plates supplemented with tricalcium phosphate (TCP) and incubated at a temperature of 28°C for a duration of 24 hours. Phosphate solubilization was suggested by the presence of a clear zone surrounding a developing colony. The formulas used to compute the phosphate solubilization index (PSI) was as follows:
PSI = (colony diameter (mm) + halo zone diameter (mm)) / colony diameter (mm)
2.3.2. Production of Indole-3-acetic acid (IAA)
The determination of IAA production was conducted using the standard approach established by Ndeddy Aka and Babalola [22]. The strains were introduced into a solution of tryptic Soy broth supplemented with 2 mg/ml of L-tryptophan and subjected to incubation on an orbital shaker at a speed of 150 rpm. The bacterial strains were cultured at a temperature of 28°C for a duration of 72 hours. The culture broths underwent centrifugation at a speed of 10,000 revolutions per minute for a duration of 10 minutes. Following centrifugation, 1 ml of the resulting liquid above the sediment was combined with 2 ml of Salkowski reagent. The presence of a pink color indicates the formation of IAA.
2.3.3. Ammonia Production
The ammonia generation by endophytic isolates was evaluated using the methodology described by Islam et al. [21]. Ten microliters of recently prepared strains cultures were introduced into test tubes containing 10 milliliters of peptone water broth and placed in an incubator at a temperature of 28 degrees Celsius for a duration of 24 hours. The test tubes were placed on a rotary shaker and rotated at a speed of 150 revolutions per minute. After the incubation period, 1 ml of Nessler's reagent was introduced into each test tube, and any alterations in color were noted. A shift in the media's hue to yellow or brown indicates a beneficial outcome for ammonia production.
2.4. Indirect Mechanisms by Endophytic Bacteria
2.4.1. Antifungal Activities
The bacterial isolates were tested for their ability to inhibit the development of three phytopathogens, namely Lasiodiplodia sp., Pinicillium sp., and Aspergillus flavus. This was done using a dual-culture technique that measures the percentage of inhibition of radial growth (PIRG) [23]. Each phytopathogen was inoculated with a mycelial plug (5 mm) on one side of a Petri dish containing tryptic soy agar (TSA), positioned 20 mm away from the edge. On the opposite side, each endophytic bacterial isolate was streaked as a line on the medium, also 20 mm away from the edge. The antagonistic activity of the chosen bacterial isolates against these phytopathogens was evaluated after 7 days of incubation by measuring the radius of the phytopathogens colony using the specified formula:
Percentage (%) of inhibition of radial growth (PIRG) = (R1 − R2/R1) × 100
Where R1 is the radial growth of the phytopathogen fungal colony on the control plate and R2 is the radial growth of the phytopathogen fungal colony in the dual culture.
2.4.2. Production of Extracellular Enzymes
The selected isolates were tested for their capacity to produce hydrolytic enzymes (amylase, protease, gelatinase, lipase, esterase, and cellulase) on Tryptic Soy Agar supplemented with 1% starch, skimmed milk, gelatin, Tween 80, Tween 20, and carboxy methylcellulose (CMC) correspondingly. The chosen bacterial isolates were streaked on the designated media and placed in an incubator for a period of 72 hours at a temperature of 28 degrees Celsius. The control treatments comprised of the identical media devoid of bacterial inoculation [24]. Furthermore, the presence of catalase activity was identified by introducing 1 mL of a 3% hydrogen peroxide (H2O2) solution to the bacterial colonies following the incubation time. The addition of hydrogen peroxide resulted in a positive reaction characterized by the creation of bubbles, which released O2 [25]. The experiments were conducted three times to ensure accuracy and reliability.
An assistant performed the detection of certain enzymes using specified solutions. To produce amylase, the plates were submerged in an Iodine solution, left for one minute, and then drained. The reaction between iodine and starch results in the formation of a chemical that exhibits a blue tint. The rate of fading of this blue color is high. Therefore, the absence of color in the area surrounding colonies shows the synthesis of amylase [26]. Furthermore, gelatinase synthesis was evaluated by saturating the petri dishes with a 20% solution of trichloroacetic acid (TCA). The presence of a distinct area devoid of growth within a minimum of 4 minutes highlights the excellent outcome [27]. In addition, 1% solution of Congo Red was applied to the surface of mature cellulase test cells for a duration of 20 minutes. Subsequently, the plate surfaces were immersed in a 1 M NaCl solution and allowed to remain undisturbed for a duration of 30 minutes. The presence of halos surrounding colonies serves as an indication of the deterioration of CMC and is indicative of the synthesis of cellulase [28]. The activities of protease, lipase, and esterase were assessed by directly measuring the ensuing clear zone after inoculation, without the use of immersion solutions.
Enzymatic activity was observed for all the enzymes examined, except for catalase. This was determined by measuring the hydrolysis halos in millimeters and the diameter of the colonies at the conclusion of the incubation period [24]. The experiments were conducted in duplicate. Subsequently, the results were assessed by determining the enzymatic index (EI). The enzyme index (EI) was determined using the following equation: EI is defined as the ratio of the diameter of the halo to the diameter of the colony. The isolates were categorized according on their production levels as non-producer (EI=0), low producer (0>EI≤2), moderate producer (2>EI≤3), and high producer (EI>3). The calculation was performed using the formula:
EI = Diameter of hydrolysis zone (mm) divided by Diameter of colony (mm)
2.4.3. Production of Hydrogen Cyanide (HCN)
The determination of hydrogen cyanide activity was conducted following the procedure established by Bakker and Schippers [29]. The endophytic bacterial cultures were streaked individually on tryptic soy agar supplemented with 0.4% (w/v) glycine. A Whatman no. 1 filter paper, saturated with a solution of picric acid, was positioned on the surface of the Petri dishes. Subsequently, the plates were securely sealed using parafilm and placed in an incubator set at a temperature of 28°C for a duration of 24 hrs. The transition of the filter paper's hue from yellow to orange to reddish-brown, as visually seen, signifies a positive outcome.
2.4.4. Abiotic Stress Tolerance
Various abiotic stress tolerance experiments were performed on fresh tryptic soy agar cultures of endophytic bacteria at different levels. These tests included exposure to Sodium chloride (NaCl), heavy metals such as copper sulphate (CuSO4) and cobalt sulphate (CoSO4), as well as high and low temperature degrees and pH values. The salinity resistance test cultures were assessed by monitoring their growth on a medium containing various concentrations of NaCl (0.5%, 2.5%, 5%, 7.5%, and 10%) [30]. The isolates were subjected to heavy metal tolerance stress by cultivating them on TSA plates that were supplemented with various soluble heavy metal sulphate salts (copper-Cu and cobalt-Co) at different concentration ranges (4, 10, 20, 30, 50, 100, 150, 200, 300, 500, 700, 1000, 1250, and 1500 ppm) [31]. The temperature resistance was evaluated using a range of temperatures (4, 10, 20, 28, 37, 45, 50, 55, and 60∘C). The pH values ranged from 4 to 11, encompassing both acidic and alkaline conditions. The stress components' levels were examined in order to identify the maximum tolerance concentration (MTC). Each of these petri dishes underwent incubation for a duration of 48 hours. The results were classified as positive (+) or negative (-) based on the bacterial growth observed in each treatment.
2.5. Molecular Identification of Endophytic Bacteria
The identification of bacterial chosen isolates was performed using the partial sequencing of 16S rRNA, as described by Nxumalo et al. [32]. The genomic DNA of each bacterial isolate was extracted using the methodology described by Weiland [33]. In summary, individual colonies were collected from a Tryptic Soy Agar plate using a sterile toothpick and then placed in a 50 μL solution consisting of 25 μL of Mastermix (including 50 units/ml of Taq polymerase, 400μL of moles DNTPs, and 3mM of MgCl2), 19μL of ultrapure PCR water, 2 μL of forward primer, and 2μL of RNA template. The thermocycler used for the reaction was the PTC 200 model from MJ Research. The reaction conditions were as follows: an initial denaturation step at 95°C for 5 minutes, followed by 30 cycles of denaturation at 95°C for 1 minute, annealing at 58°C for 1 minute, and elongation at 72°C for 2 minutes. The final elongation step was performed at 72°C for 5 minutes. These conditions were used to amplify a partial 16S rDNA fragment using the bacterial universal primers 27f (5′-AGAGTTTGATCCTGGCTCAG-3′) and 1492r (5′-GGTTACCTTGTTACGACTT-3′). The amplification process was verified through the detection of ethidium bromide fluorescence in a 1% agarose gel. PCR results were sent to Macrogen, Inc. (Seoul, Korea) for sequencing. The obtained sequences were then compared to similar sequences in their 16S rRNA databases using the NCBI BLAST nucleotide search, which is based on the GenBank database [34].
3. Results
3.1. Isolation of Endophytic Bacteria
A total of 1129 bacterial colonies were isolated from the roots, stems, leaves, and inflorescences of healthy Aerva javanica plants obtained from Shada Alasfal Mountain in the Albaha region of Saudi Arabia. The roots and stems exhibited the greatest abundance of bacterial colonies, with 359 and 311 respectively, while inflorescences displayed the lowest number of endophytic bacteria, with 199 colonies. Moreover, tryptic soy agar and starch casein agar exhibited the highest efficacy in promoting the growth of endophytic bacteria, with 225 and 191 isolates, respectively. The humic acid-vitamins agar exhibited the lowest number of bacterial isolates, with a total of 128 strains (Figure 3). All the data from the following experiments were presented as the average of three separate replicates, with the standard deviation (SD) indicated as mentioned earlier. Mean values were deemed statistically significant when p < 0.05.
3.2. Direct PGP Activities of A. javanica Bacterial Endophytes
The PGP activities were directly assessed by evaluating the capacity of the chosen bacterial isolates to solubilize phosphate, create indole acetic acid (IAA), and generate ammonia. 50% of the bacterial isolates exhibited phosphate solubilization capabilities, while IAA was detected in 66.7% of the bacterial samples. Nevertheless, only one isolate exhibited a complete lack of ammonia production capability. BAB6 had the highest potential for phosphate solubilization, with an EI (Effectiveness Index) of 2.75. On the other hand, IAA showed the highest production rate among BAB1, BAB4, and BAB6. Furthermore, both BAB2 and BAB5 exhibited the most elevated levels of ammonia generation, as indicated in Table 1.
3.3. Indirect PGP Activities of A. javanica Bacterial Endophytes
All bacterial isolates tested positive for HCN, with BAB1 exhibiting the greatest production rate (Table 1). According to figure 4., all bacterial isolates exhibited antagonistic effects against two pathogenic fungi, namely Penicillium sp. and Aspergillus flavus. However, only one bacterial isolate was unable to suppress the growth of Lasiodiplodia sp. BAB3 had the strongest antagonistic activity, inhibiting Lasiodiplodia sp. by 81.6%. BAB5 also showed significant inhibitory activity, with a rate of 66.7%. However, BAB1 did not demonstrate any ability to inhibit Lasiodiplodia sp. In addition, BAB3 exhibited a 61% inhibition of the phytopathogenic fungus Penicillium sp. All of the bacterial isolates exhibited a limited capacity to inhibit Aspergillus flavus, with inhibition rates ranging from 28.8% to 37.2%.
The endophytic bacteria were examined for their capacity to synthesize various lytic enzymes, including amylase, protease, gelatinase, cellulase, lipase, and esterase (Figure 5). The Enzymatic Index (EI) for all of these isolates was determined by assessing their capacity to synthesize a minimum of 3 out of the 6 examined enzymes. All of the isolates exhibited amylolytic activity. Four isolates produced protease, gelatinase, and esterase enzymes apiece. Only two isolates showed lipolytic activity, whereas three isolates had end cellulolytic activity. Certain isolates demonstrated exceptional enzymatic efficiency. The amylase and gelatinase activity of BAB1 were found to be the greatest, with an Enzyme Index (EI) of 1.38 and 4.94, respectively. BAB3 had the highest levels of protease and esterase production, with an Enzyme Index (EI) of 2.59 and 2.00, respectively. On the other hand, BAB5 demonstrated the highest EI for cellulase production, with a value of 2.00. In contrast, only BAB3 and BAB4 isolates exhibited high EI readings of 0.96 and 1.00 respectively.
The bacterial isolates were assessed for their abiotic stress resistance by observing their development at varying levels of sodium chloride, temperatures, pH, and heavy metals such as copper sulphate and cobalt sulphate. The majority of the bacterial isolates demonstrated the capacity to thrive in the presence of 2.5% salt, with the exception of BAB2 and BAB6, which exhibited an inability to grow under conditions of salinity stress. Nevertheless, BAB1 and BAB1 reached their maximum growth rates at 7.5% and 10% respectively, as seen in Figure 6. Regarding temperature stress, it was observed that only three isolates were able to develop at a minimum temperature of 20°C, despite the fact that all bacterial isolates were capable of growing within the range of 28-37°C. In addition, BAB2, BAB3, BAB5, and BAB6 exhibited tolerance to temperatures up to 50°C, as shown in Figure 7. Furthermore, the capabilities of the chosen bacterial isolates were evaluated for their ability to thrive in both low and high pH environments. The findings indicated that the isolates exhibited growth at a minimum pH of 6, while their tolerance to higher pH levels varied. For example, BAB2 and BAB6 reached a pH of 9 and 10, respectively, while the other isolates grew up to a pH of 11 (Figure 8).
In addition, the bacterial endophytes demonstrated resistance to copper sulphate and cobalt sulphate, which were used as heavy metal tests. BAB5 exhibited the lowest levels of resistance to copper sulphate, with maximum growth observed at a concentration of 200 ppm. In contrast, BAB2 and BAB3 shown resistance up to a concentration of 300 ppm. Nevertheless, BAB1, BAB4, and BAB6 exhibited greater resistance to concentrations of 500, 700, and 1000 ppm, respectively, as shown in Figure 9. However, the chosen isolates exhibited a greater resistance to cobalt sulphate compared to copper sulphate. BAB6 exhibited the greatest level of resistance to 1500 ppm of cobalt sulfate, followed by BAB4 and BAB1 with concentrations of 1250 and 1000 ppm, respectively. At a concentration of 500 ppm, both BAB3 and BAB5 exhibited their maximum tolerance to copper sulphate, as shown in Figure 10. BAB2, on the other hand, reached its maximum tolerance at a concentration of 700 ppm.
3.4. Molecular Identification of Selected Endophytic Bacteria
Six bacterial isolates were chosen for further investigation based on the variations seen among the colonies. In summary, BAB1 and BAB2 were extracted from the roots, whereas BAB3 was extracted from the stems. However, the remaining isolates were obtained from leaves. In addition, BAB1, BAB5, and BAB6 were obtained from SNA, SCA, and TWYEA, respectively, while the remaining isolates were obtained from ISP4 Table (2). The bacterial isolates BAB1, BAB2, BAB3, BAB4, BAB5, and BAB6 were identified molecularly by comparing their 16S rRNA gene sequences to known sequences. The results showed that BAB1 had a 99% similarity to the sequence of Micrococcus sp. Mcap_H18, BAB2 had a 99% similarity to the sequence of Enterobacter cloacae HSNJ4, BAB3 had a 99% similarity to the sequence of Pseudomonas aeruginosa AAII-2, BAB4 had a 99% similarity to the sequence of Delftia sp. lp09, BAB5 had a 99% similarity to the sequence of Bacillus licheniformis VTM1R80, and BAB6 had a 99% similarity to the sequence of Bacillus parabrevis strain NAP3. Table 1 and Figure 11 displayed the source organ for each chosen bacterial isolate, along with their genetic identity. The sequences of closely related organisms were collected from Gen Bank in order to reconstruct the phylogenetic tree.

Table 2.
Isolated organ and medium, and sequence similarities of endophytic bacteria strains with sequences registered in GenBank.
Table 2.
Isolated organ and medium, and sequence similarities of endophytic bacteria strains with sequences registered in GenBank.
 |
4. Discussion
Plant growth promoting bacteria are currently being used as a promising approach to enhance plant growth and increase crop output in sustainable agriculture. Endophytic bacteria are valuable bioresources due to their capacity to inhabit the internal tissues of plants through direct contact [35]. Plants face ongoing challenges from their surroundings, including living organisms and non-living factors including infections, high and low pH levels, extreme temperatures, and heavy metals. Endophytes can assist these plants through various direct and indirect means, offering possible support. The study aimed to separate and categorize the chosen endophytic bacteria from Aerva javanica based on their physical features and molecular classification. The following six Plant Growth Promoting Bacteria (PGPB) were isolated from various parts of A. javanica, including roots, stems, leaves, and inflorescences: Micrococcus sp. Mcap_H18, Enterobacter cloacae, Pseudomonas aeruginosa, Delftia sp., Bacillus licheniformis, and Brevibacillus parabrevis. Bacillus, Enterobacter, and Pseudomonas are the predominant genera of bacterial endophytes, as described by Hassan [36], Ben Slama et al. [37], and Pinto et al. [38].
Endophytes can contribute to plant health through two distinct processes: direct and indirect. These strategies involve the active participation of bacteria in promoting plant well-being [8]. Direct interactions with endophytes involve enhanced nutrient uptake and increased phytohormone levels in the plant. These interactions directly contribute to increased biomass production, root system development, plant height, weight reproduction, and yield. Due to these advantages, they can be labeled as biofertilizers [39]. Initially, the direct actions of PGP have been conducted by screening for the solubilization of phosphate and the formation of ammonia and IAA. 50% of the bacteria that were separated were capable of exhibiting phosphate solubilization. The genera that exhibited the highest level of productivity were Bacillus and Pseudomonas. Pinto et al. [38] have documented comparable findings. Endophytic bacteria have the ability to produce indole acetic acid (IAA), which is an important hormone that regulates plant growth. They do this by using different routes and using tryptophan as the major building block [40]. Our analysis revealed that all the isolates tested, with the exception of BAB1 and BAB5, were capable of producing IAA. Extensive evidence exists to support the fact that the majority of endophytic bacteria are capable of synthesizing indole-3-acetic acid (IAA) [36,37,40].
The generation of ammonia is a significant characteristic of bacteria that promote plant growth. It collects and provides nitrogen to their host plants, hence enhancing plant development. Several investigations have documented the ammonia generation by endophytic bacteria. All bacterial strains, save one, exhibited positive ammonia production in this investigation. The isolates have been classified as diazotrophic bacteria due to their capacity to convert gaseous nitrogen into a useful form of ammonia [41]. The results align with the discoveries made by Hassan [36] and Pinto et al. [38]. All of the bacterial strains that were examined showed positive results for the HCN production test. It was thought that the synthesis of HCN contributes to the stimulation of plant growth by inhibiting plant diseases [42]. Nevertheless, this notion has undergone recent modifications. HCN synthesis is thought to enhance phosphorus availability by binding and isolating metals, hence indirectly increasing nutrient availability to rhizobacteria and host plants [43,44].
Furthermore, indirect PGP activities encompass the synthesis of secondary metabolites, such as antimicrobial agents and enzymes. Additionally, these activities provide protection for plants against biotic threats, such as pests, and abiotic challenges, including salt salinity, extreme pH levels, temperatures, and heavy metal exposure. This work investigated indirect pathways by conducting screenings for antifungal activity against pathogenic fungi, lytic enzymes, and abiotic stimuli such as salt salinity, high and low pH, temperatures, and heavy metals like copper sulfate and cobalt sulphate. Endophytic bacteria engage in several interactions with plants, including the control of plant diseases through antagonism, which indirectly enhances plant development characteristics. Endophytes possessing plant growth promoting properties are employed as a substitute for chemical pesticides in the agricultural sector [45,46]. Our results indicated that every isolated endophytic bacterium exhibited zones of inhibition against at least one of the fungal infections that were examined. These endophytes have the capacity to act as biocontrol agents by suppressing the growth of pathogenic fungus.
Endophytes possess enzymatic capabilities that enable them to defend their host plants by breaking down the cell walls of harmful bacteria, as described by Glick [9]. Endophytes acquire nutrients from plants through the secretion of enzymes. These enzymes enhance plant nutrition and contribute to plant aging by breaking down certain organic compounds within the plants [47]. Amylases are employed for the process of breaking down starch into sugars on a large scale in industries. On the other hand, cellulase is utilized in the manufacturing of biofuels, specifically bioethanol, by utilizing lignocellulosic substrates. Lipases, also known as triacylglycerol ester hydrolyses, facilitate the hydrolysis of triglycerides into glycerol and free fatty acids at the interface between oil and water. Under conditions without water, they facilitate the formation of glycerides from glycerol and fatty acids by catalyzing the reverse processes, namely esterification, interesterification, and transesterification [48]. In addition, protease is an enzyme that is found in all living species and is necessary for cell signaling, differentiation, cell proliferation, and metabolism. The majority of proteases are obtained from Bacillus sp. [48]. Antioxidant enzymes, like catalase, have a crucial function in removing reactive oxygen species (ROS). Research has shown that during drought conditions, these enzymes are activated as a response to stress. This suggests that a strong antioxidant capacity is linked to the ability to tolerate stress [49]. Catalase catalyzes the conversion of hydrogen peroxide (H2O2) into water and molecular oxygen (O2) [50].
Abiotic stress refers to any non-living environmental element that restricts the growth and metabolic processes of plants. The research focused on studying temperatures, salt saltiness, pH, and heavy metals like copper sulphate and cobalt sulphate. These parameters have significant impacts on agricultural output and serve as important limitations. The prevalence of salt salinity is extensive, impacting around 10% of the Earth's land surface, particularly in regions with irrigation [51]. Furthermore, fluctuations in temperature can induce several forms of stress in plants, including osmotic damage, desiccation, loss of stomatal control, and decreased efficiency of the photosynthetic machinery [52]. Research has demonstrated that a rapid rise in the surrounding temperature of 5-7 degrees Celsius induces heat stress in plants. According to Ljubej et al. [53], it disrupts the process of photosynthesis, decreases plant water availability, hinders flowering and fruiting, and attracts pests and illnesses. The pH of soil primarily affects the water in the soil and has a crucial role in influencing soil biology, chemistry, and physical processes. These factors directly affect the growth and development of plants and the productivity of crops [54], while there is variation among plants in their ability to withstand severe pH levels, the majority of agricultural plants thrive best when the pH is close to neutral [55].
Soil contamination by heavy metals is a type of mineral toxicity that has physiological significance in plants. The presence of abnormally high levels of heavy metals in soils can be attributed to mining activities, the composition of the parent rocks, and the processing of metals. Plant growth is significantly impacted by a high concentration of heavy metals, as it results in toxic effects that hamper the plants' ability to absorb nutrients. This, in turn, leads to damage to the membrane integrity and enzyme activity of the plants' cells [56].
5. Conclusions
Currently, the study found that the A. Javanica medicinal plant, which is a native of Shada Al-Asfal Mountain, serves as an ecological niche for a variety of putative bacterial endophytes. In this study, A. Javanica selected bacterial had a variety of production of IAA, ammonia, and phosphate solubilization factors as direct mechanisms, while indirect mechanisms were demonstrated by these bacteria which had produced HCN and some lytic enzymes with antifungal activities. Moreover, the bacterial isolates had good stress tolerance towards some tested factors like pH, temperatures, salt salinity and heavy metals which were copper sulphate and cobalt sulphate. Future studies will examine these isolates' further biocontrol capabilities, though.
Author Contributions
Conceptualization, KZ, MA, SJ and RA; methodology, KZ and MA; software, KZ; validation, SJ and RA; formal analysis, MA; resources, KZ, MA, SJ and RA; writing-original draft preparation, KZ; writing-review and editing, KZ; visualization, MA, SJ and RA; supervision, MA, SJ and RA; project administration, MA; funding acquisition, KZ, MA, SJ and RA. All authors have read and agreed to the published version of this article.
Funding
No external funding was received for this work.
Acknowledgments
The authors wish to thank Mr. Nasser Alshadawi, the historical researcher, for his help during the samples collecting from Shada Alasfal Mountain.
Conflicts of Interest
The authors declare that they have no known competing financial interests nor personal relationships that could have appeared to influence the work reported in this article.
References
- Alzahrani, K., Jastaniah, S., Amasha, R., et al. Diversity and Colonization of Endophytic Actinomycetes in some Medicinal Plants: Review. Journal of Contemporary Medical Sciences 2022, 8(3). [CrossRef]
- Goulart, M., Cueva-Yesquén, L., Hidalgo Martinez, K., et al. Comparison of specific endophytic bacterial communities in different developmental stages of Passiflora incarnata using culture-dependent and culture-independent analysis. Microbiology Open 2019, 8, e896. [CrossRef]
- Suman, A., Yadav, A. and Verma, P. Endophytic Microbes in Crops: Diversity and Beneficial Impact for Sustainable Agriculture. In: Singh D., Singh H., Prabha R. (eds) Microbial Inoculants in Sustainable Agricultural Productivity. Springer, New Delhi, 2016.
- Singh, M., Kumar, A., Singh, R., et al. Endophytic bacteria: A new source of bioactive compounds. 3 Biotech 2017, 7(5), 315. [CrossRef]
- Buatong, J., Phongpaichit, S., Rukachaisirikul, V., et al. Antimicrobial activity of crude extracts from mangrove fungal endophytes. World J Microbiol Biotechnol. 2011, 27(12), 3005–8. [CrossRef]
- Santoyo, G., Moreno-Hagelsieb, G., del Orozco-Mosqueda, M., et al. Plant growth-promoting bacterial endophytes. Microbiol. Res. 2016, 183, 92–99. [CrossRef]
- López-Arredondo, D., Leyva-González, M., Alatorre-Cobos, F., et alBiotechnology of nutrient uptake and assimilation in plants. Int. J. Dev. Biol. 2013, 57, 595–610.
- Noemi, C. and Everlon, C. (2022) Endophytic fungi: a tool for plant growth promotion and sustainable agriculture, Mycology 2022, 13(1), 39-55.
- Glick, B. Plant growth-promoting bacteria: mechanisms and applications. Scientifica 2012, 1–15. [CrossRef]
- Gupta, P., Samant, K., and Sahu, A. Isolation of cellulose-degrading bacteria and determination of their cellulolytic potential. International journal of microbiology 2012, 578925. [CrossRef]
- Nagei, T., Alshaeri, S., Alsulaimany, F., et al. The Role of Aerva Javanica (Burm.F.) Juss. Growing in Jazan, Saudi Arabia as Antimicrobial and Coagulating Factor. European Journal of Applied Sciences, 2021, 9(5), 456-471. [CrossRef]
- Thomas, J., El-Sheikh, M. and Alatar, A. Endemics and endangered species in the biodiversity hotspot of the Shada Mountains, Saudi Arabia. Journal of Arid Land 2017, 9(1), 109–121. [CrossRef]
- El-Tayeh, N., Galal, H., Soliman, M., et al. Association of Morphological, Ecological, and Genetic Diversity of Aerva javanica Populations Growing in the Eastern Desert of Egypt. Agronomy 2020, 10, 402.
- Suleiman, M. Ethnobotanical, Phytochemical, and Biological Study of Tamarix aphylla and Aerva javanica Medicinal Plants Growing in the Asir Region, Saudi Arabia. Tropical Conservation Science 2019, 12.
- Srinivas, P. and Reddy S. Screening for antibacterial principle and activity of Aerva javanica (Burm.f) Juss. ex Schult. Asian Pacific Journal of Tropical Biomedicine 2012, S838-S845. [CrossRef]
- Singh, R. and Jha, P. Plant Growth Promoting Potential of ACC Deaminase Rhizospheric Bacteria Isolated from Aerva javanica: A Plant Adapted to Saline Environments. Int.J.Curr.Microbiol.App.Sci 2015, 4(7), 142-152.
- Mookherjee, A., Mitra, M., Kutty, N., et al. Characterization of endo-metabolome exhibiting antimicrobial and antioxidant activities from endophytic fungus Cercospora sp. PM018. South African journal of botany 2020, 134. [CrossRef]
- Qin, S., Li, J., Chen, H., et al. Isolation, diversity, and antimicrobial activity of rare actinobacteria from medicinal plants of tropical rain forests in Xishuangbanna, China. Appl. Environ. Microbiol. 2009, 75, 6176-6186. [CrossRef]
- Musa, Z., Ma, J., Egamberdieva, D., et al. Diversity and Antimicrobial Potential of Cultivable Endophytic Actinobacteria Associated with the Medicinal Plant Thymus roseus. Frontiers in Microbiology. 2020, 11. [CrossRef]
- Phongsopitanuna, W., Sripreechasakc, P., Rueangsawang, K., et al. Diversity and antimicrobial activity of culturable endophytic actinobacteria associated with Acanthaceae plants. Science Asia 2020, 46, 288-296.
- Islam, M., Madhaiyan, M., Deka Boruah, H., et al. Characterization of plant growth-promoting traits of free-living diazotrophic bacteria and their inoculation effects on growth and nitrogen uptake of crop plants. J Microbiol Biotechnol. 2009, 19(10), 1213–1222.
- Ndeddy Aka, R. and Babalola, O. Effect of bacterial inoculation of strains of Pseudomonas aeruginosa, Alcaligenes feacalis and Bacillus subtilis on germination, growth and heavy metal (Cd, Cr, and Ni) uptake of Brassica juncea. Int J Phytorem. 2016, 18, 200–209.
- Pitiwittayakul N, Wongsorn D, Tanasupawat S. Characterisation of Plant Growth-Promoting Endophytic Bacteria from Sugarcane and Their Antagonistic Activity against Fusarium moniliforme. Trop Life Sci Res. 2021, 32(3), 97-118. [CrossRef]
- Mohamed, A., Abd El-Megeed, F., Hassanein, N., et al. Native Rhizospheric and Endophytic Fungi as Sustainable Sources of Plant Growth Promoting Traits to Improve Wheat Growth under Low Nitrogen Input. J. Fungi 2022, 8, 94. [CrossRef]
- Minotto, E., Milagre, L., Oliveira, M., et al. Enzyme Characterization of Endophytic Actinobacteria Isolated from Tomato Plants. Journal of Advanced Scientific Research 2014, 5(02), 16-23.
- Pranay, K., Padmadeo, S., Jha, V., et al. Screening and identification of amylase producing strains of Bacillus. J. Appl. Biol. Biotech 2019, 7, 57–62.
- Medina, P. and Baresi, L. (2007). Rapid identification of gelatin and casein hydrolysis using TCA. J Microbiol Meth. 2007, 69, 391-3. [CrossRef]
- Mahdi, I., Fahsi, N., Hafidi, M., et al. Plant Growth Enhancement using Rhizospheric Halotolerant Phosphate Solubilizing Bacterium Bacillus licheniformis QA1 and Enterobacter asburiae QF11 Isolated from Chenopodium quinoa Willd. Microorganisms 2020, 8, 948. [CrossRef]
- Bakker, A. and Schippers, B. Microbial cyanide production in the rhizosphere in relation to potato yield reduction and Pseudomonas spp. mediated plant growth-stimulation. Soil Biol Biochem. 1987, 19, 451–457. [CrossRef]
- Kumar, A., Singh, R., Yadav, A., et al. Isolation and characterization of bacterial endophytes of Curcuma longa L. 3 Biotech 2016, 6, 60. [CrossRef]
- Perelomov, L., Sizova, O., Rahman, M., et al. Metal-Tolerant Bacteria of Wastewater Treatment Plant in a Large City. Sustainability 2022, 14, 11335. [CrossRef]
- Nxumalo, C., Ngidi, L., Shandu, J., et al. Isolation of endophytic bacteria from the leaves of Anredera cordifolia CIX1 for metabolites and their biological activities. BMC Complement Med Ther. 2020, 20, 300. [CrossRef]
- Weiland, J. Rapid procedure for the extraction of DNA from fungal spores and mycelia. Fungal Genet. Rep. 1997, 44, 60–63. [CrossRef]
- Altschul, S., Gish, W., Miller, W., et al. Basic local alignment search tool. J Mol Biol. 1990, 215,3, 403–410. [CrossRef]
- Chaturvedi, H., Singh, V. and Gupta, G. Potential of bacterial endophytes as plant growth promoting factors. Journal of Plant Pathology Microbiology 2016, 7, 376. [CrossRef]
- Hassan, S. Plant growth-promoting activities for bacterial and fungal endophytes isolated from medicinal plant of Teucrium polium L. J. Adv. Res. 2017, 8, 687–695. [CrossRef]
- Ben Slama, H., Triki, M., Chenari Bouket, A., et al. Screening of the High-Rhizosphere Competent Limoniastrum monopetalum’ Culturable Endophyte Microbiota Allows the Recovery of Multifaceted and Versatile Biocontrol Agents. Microorganisms 2019, 7(8), 249. [CrossRef]
- Pinto, M., Inocente, L., Oliveira, P., et al. Plant Growth-Promoting (PGP) Traits of Endophytic Bacteria from In Vitro Cultivated Tectona grandis L.f. Forests 2022, 13(10), 1539. [CrossRef]
- Bamisile, B., Dash, C., Akutse, K., et al. Fungal endophytes: beyond herbivore management. Front Microbiol. 2018, 9(11).
- Belbahri, L., Chenari Bouket, A., Rekik, I., et al. Comparative genomics of Bacillus amyloliquefaciens strains reveals a core genome with traits for habitat adaptation and a secondary metabolites rich accessory genome. Front. Microbiol 2017, 8(1438).
- Ji, S., Gururani, M. and Chun, S. Isolation and characterization of plant growth promoting endophytic diazotrophic bacteria from Korean rice cultivars. Microbiol. Res. 2014, 169, 83–98.
- Defago, G., Berling, C., Burger, U., et al. Suppression of black root rot of tobacco and other root diseases by strains of Pseudomonas fluorescens: Potential applications and mechanisms. Biol. Control Soil Borne Plant Pathog. 1990, 34, 93–108.
- Rijavec, T. and Lapanje, A. Hydrogen cyanide in the rhizosphere: not suppressing plant pathogens, but rather regulating a vailability of phosphate. Front. Microbiol. 2016, 7, 1785. [CrossRef]
- Agbodjato, N., Noumavo, P., Baba Moussa, F., et al. Characterization of potential plant growth promoting rhizobacteria isolated from Maize (Zea mays L.) in central and Northern Benin (West Africa). Appl. Environ. Soil Sci. 2015, 01-09.
- Kloepper, J., Tuzun, S., Liu, L., et al. Plant growth promoting rhizobacteria as inducers of systemic disease resistance. Pest Management: Biologically Based Technologies. American Chemical Society Books, Washington, DC. 1993, 156–165.
- Mendes, R., Kruijt, M., De Bruijn, I., et al. (2011). Deciphering the rhizosphere microbiome for disease suppressive bacteria. Science 2011, 332, 1097–1100. [CrossRef]
- Choi, Y., Hodgkiss, I. and Hyde, K. Enzyme production by endophytes of Brucea javanica. J Agric Tech 2005, 1.
- Singh, R., Singh, T. and Pandey, A. Chapter 1, Microbial Enzymes-An Overview. In Biomass, Biofuels, Biochemicals. Advances in Enzyme Technology, Elsevier, 2019, 1-40.
- Zandalinas, S., Mittler, R., Balfagón, D., et al. Plant adaptations to the combination of drought and high temperatures. Physiol. Plant 2018, 162 (1), 2–12.
- Singh, D., Kaur, S. and Kumar, A. In vitro drought tolerance in selected elite clones of Eucalyptus tereticornis Sm. Acta Physiol. Plant 2020, 42, 1–9.
- Koza, N., Adedayo, A. Babalola, O., et al. Microorganisms in Plant Growth and Development: Roles in Abiotic Stress Tolerance and Secondary Metabolites Secretion. Microorganisms 2022, 10, 1528. [CrossRef]
- Balliu, A., Zheng, Y., Sallaku, G., et al. Environmental and Cultivation Factors Affect the Morphology, Architecture and Performance of Root Systems in Soilless Grown Plants. Horticulturae 2021, 7(8), 243.
- Ljubej, V., Karalija, E., Salopek-Sondi, B., et al. Effects of Short-Term Exposure to Low Temperatures on Proline, Pigments, and Phytochemicals Level in Kale (Brassica oleracea var. acephala). Horticulturae 2021, 7, 341. [CrossRef]
- Msimbira, L. and Smith, D. The Roles of Plant Growth Promoting Microbes in Enhancing Plant Tolerance to Acidity and Alkalinity Stresses. Front. Sustain. Food Syst. 2020, 4, 106. [CrossRef]
- Burt, R. Soil Survey Staff: Soil Survey Field and Laboratory Methods Manual- Soil Survey Investigations Report. Washington, DC: US Department of Agriculture 2014, 51, 227–234.
- Läuchli, A. and Grattan, S. “Soil pH extremes,” in Plant Stress Physiology; ed. S. Shabala; Wallingford: Centre for Agriculture and Bioscience International, 2012, 194.
Figure 1.
Aerva javanica plant sample in field.

Figure 2.
A map of Shada Al-Asfal Mountain.
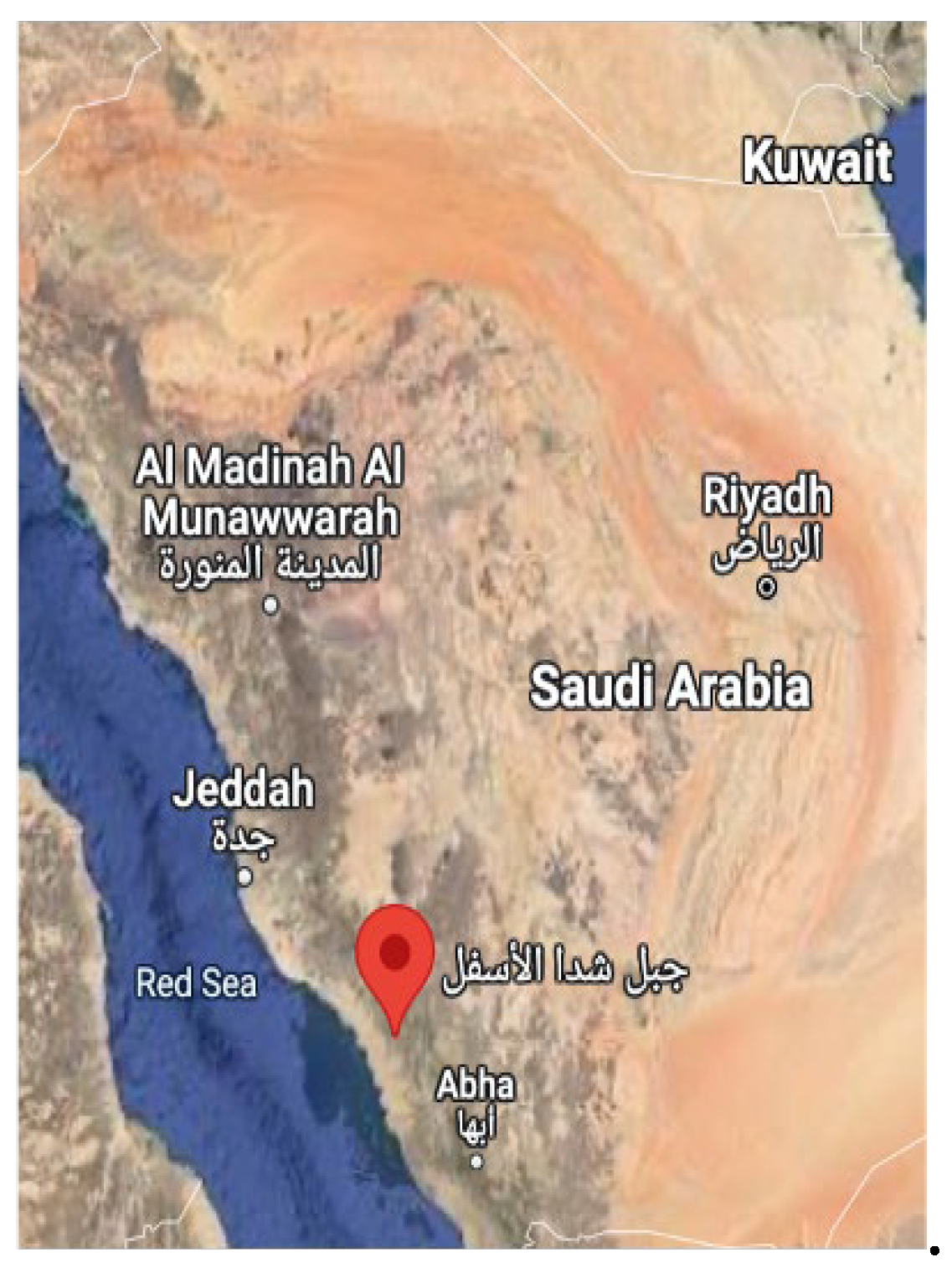
Figure 3.
The total percentage of the appeared bacterial colonies on the isolation media for each organ of A. javanica.
Figure 3.
The total percentage of the appeared bacterial colonies on the isolation media for each organ of A. javanica.
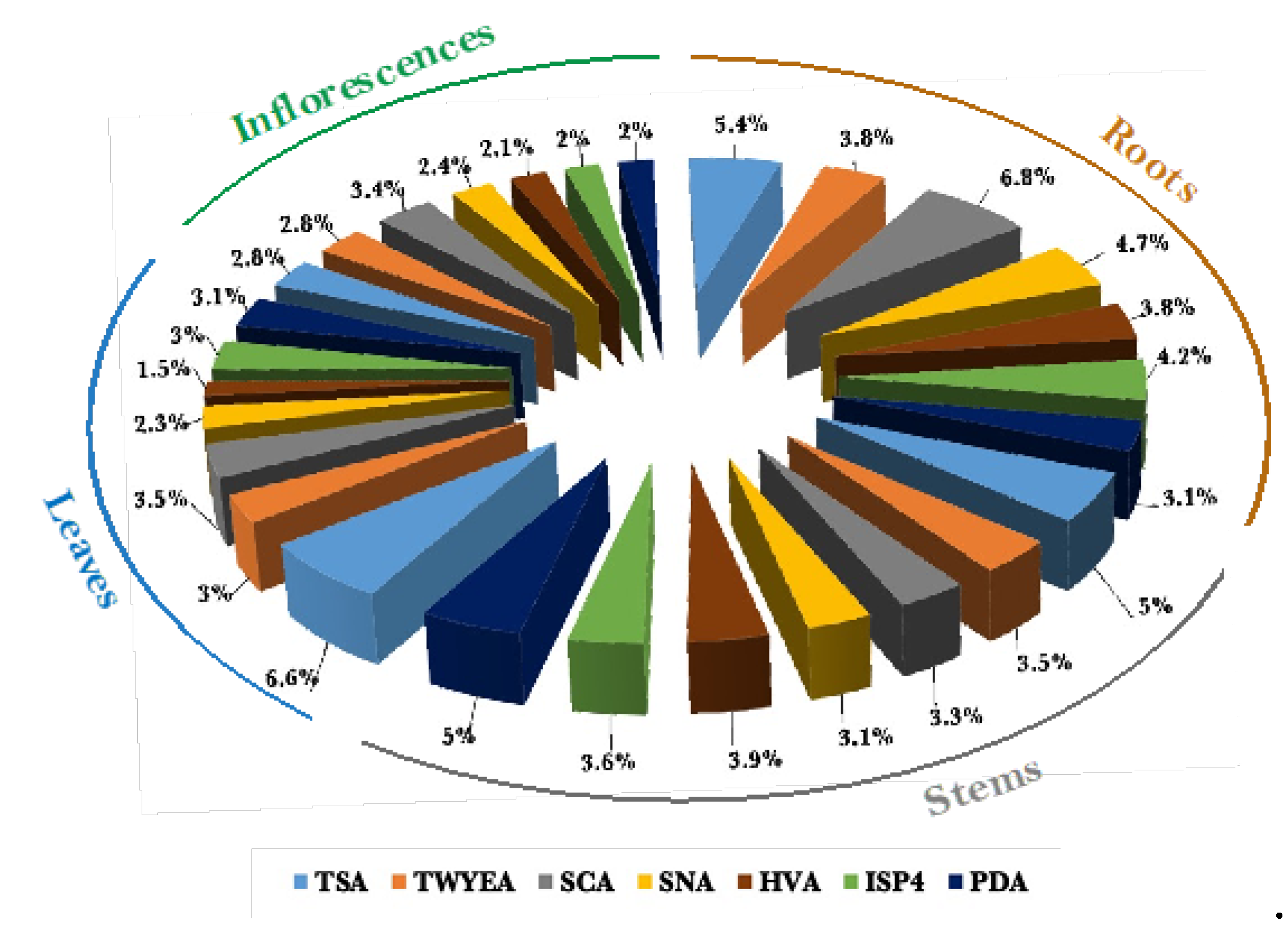
Figure 4.
Percentage of inhibition of radial growth (PIRG%) for some pathogenic fungal strains by the endophytic bacteria isolated from Aerva javanica.
Figure 4.
Percentage of inhibition of radial growth (PIRG%) for some pathogenic fungal strains by the endophytic bacteria isolated from Aerva javanica.
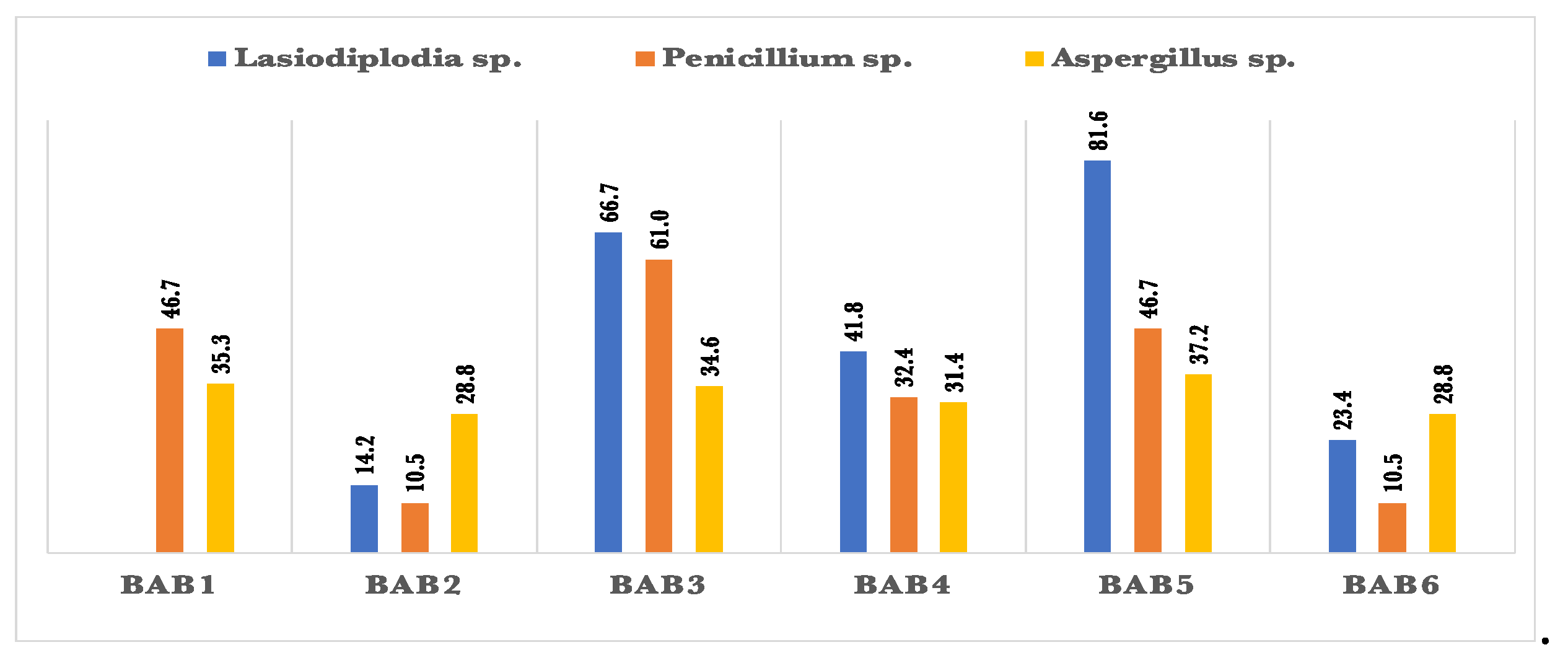
Figure 5.
Enzymatic index (EI) for the selected bacterial isolates.
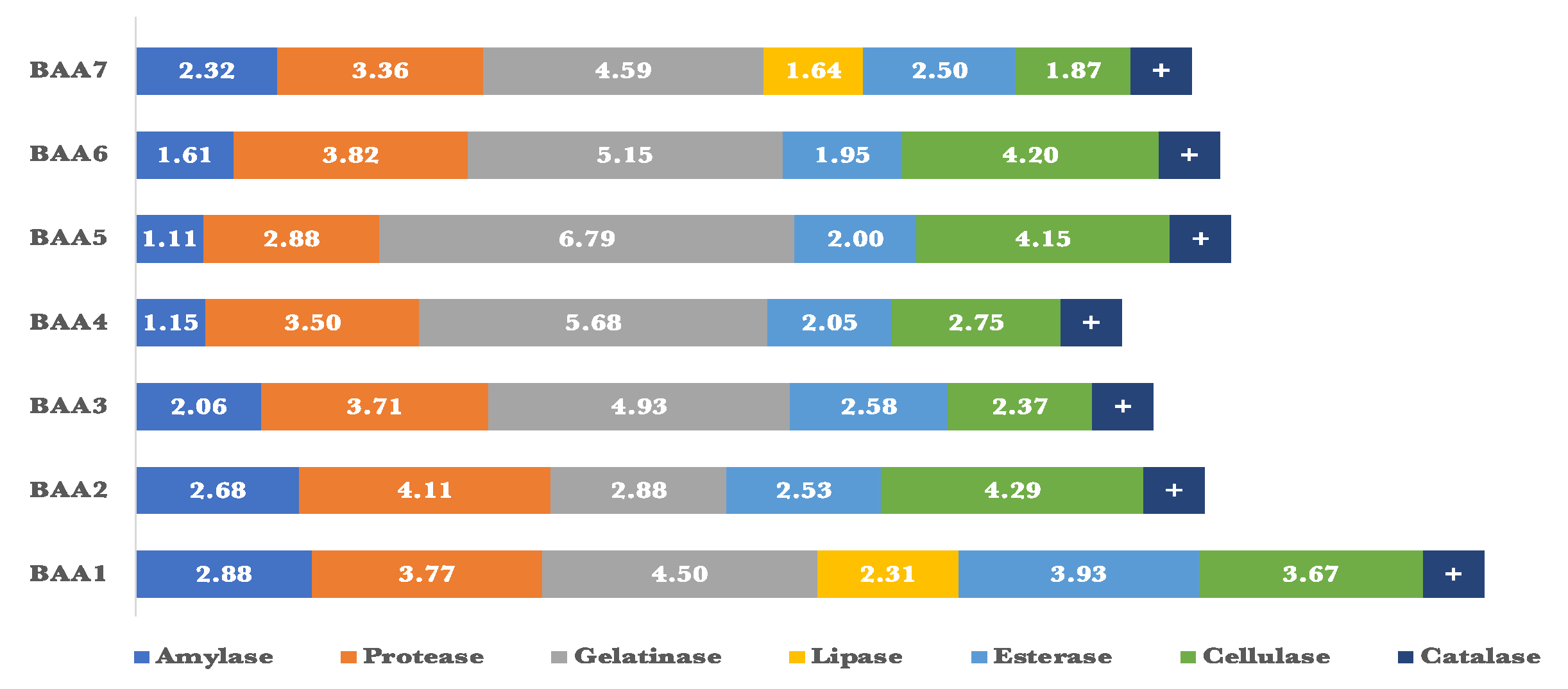
Figure 6.
Salt salinity tolerance by the selected bacterial isolates.
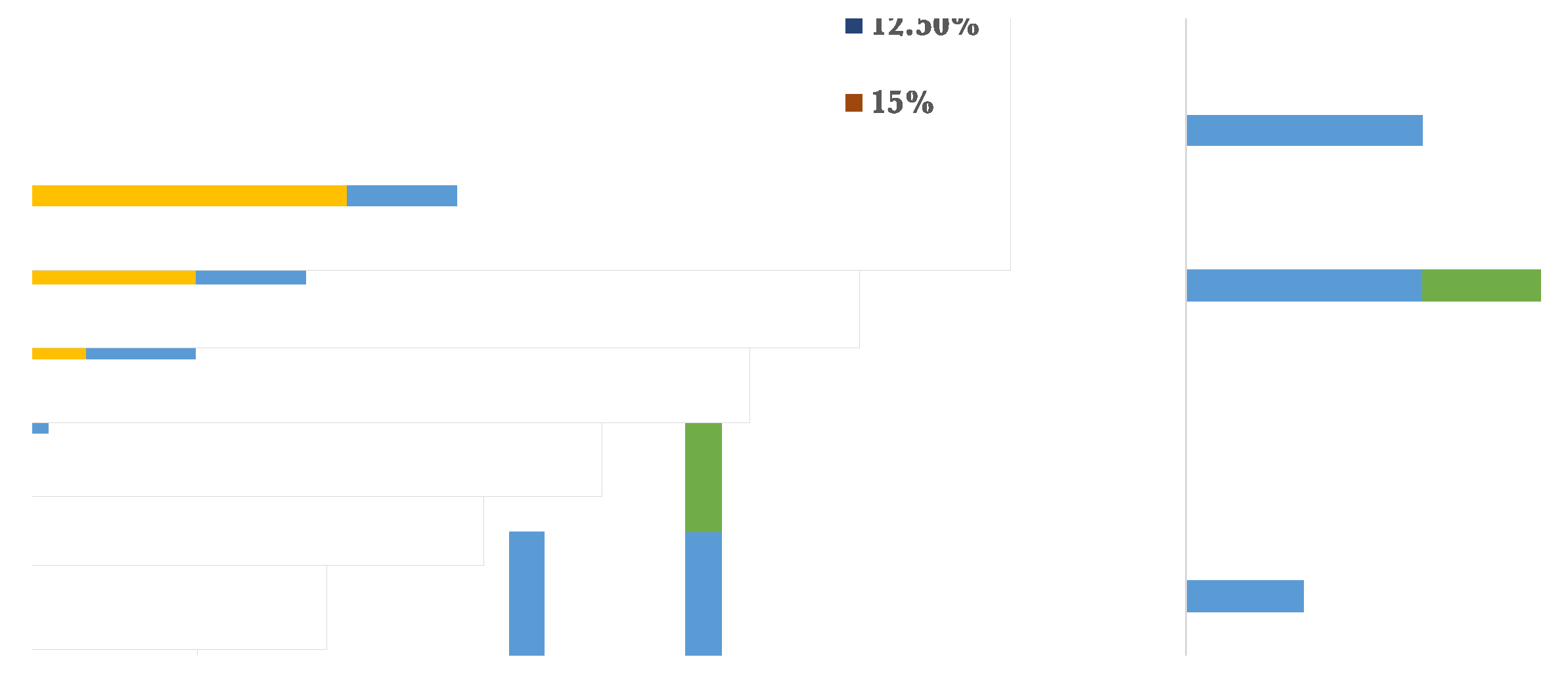
Figure 7.
High temperatures tolerance by the selected bacterial isolates.
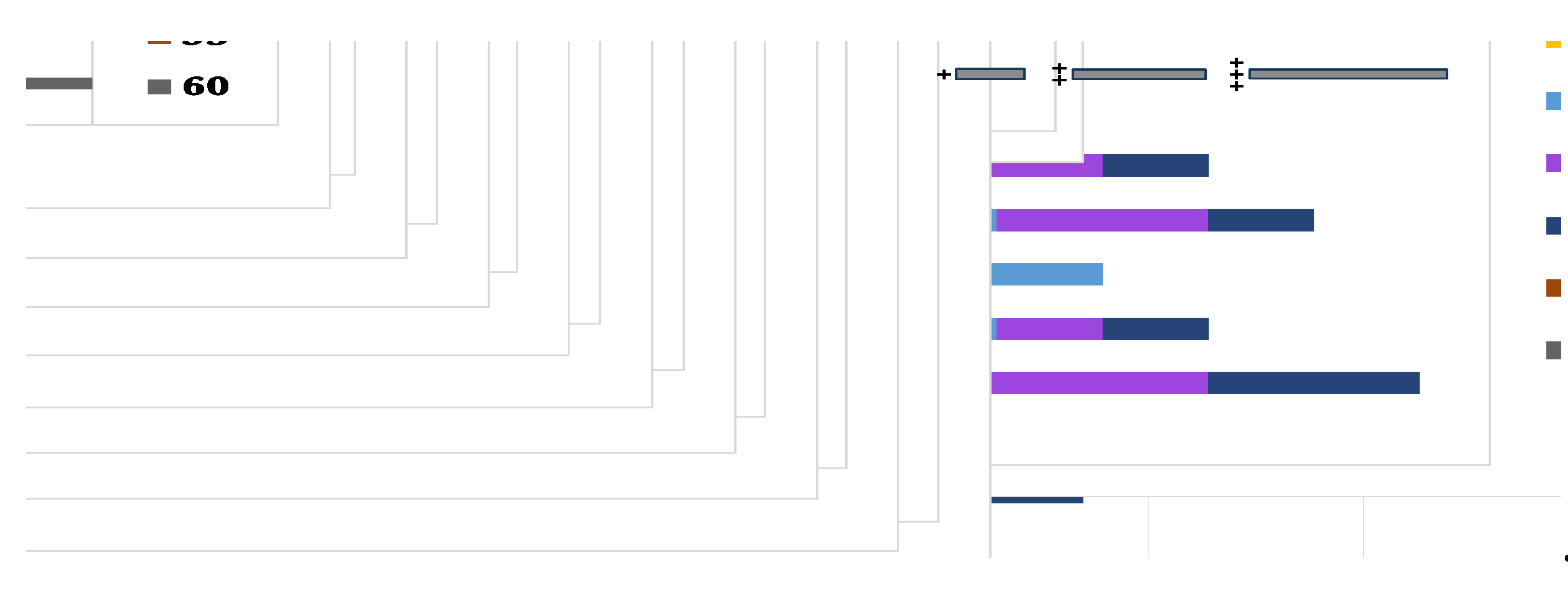
Figure 8.
pH tolerance by the selected bacterial isolates.

Figure 9.
Stress tolerance of Copper Sulphate as a heavy metal by the selected bacterial isolates.
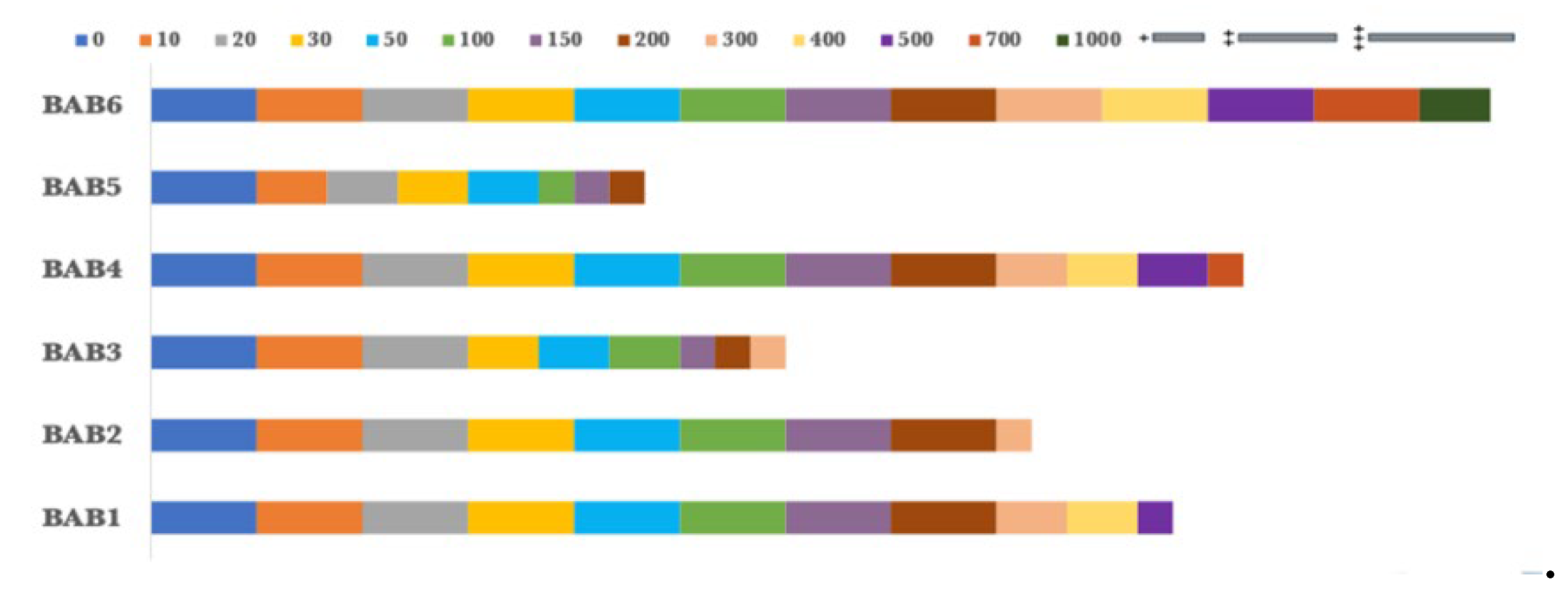
Figure 10.
Stress tolerance of Cobalt Sulphate as a heavy metal by the selected bacterial isolates.
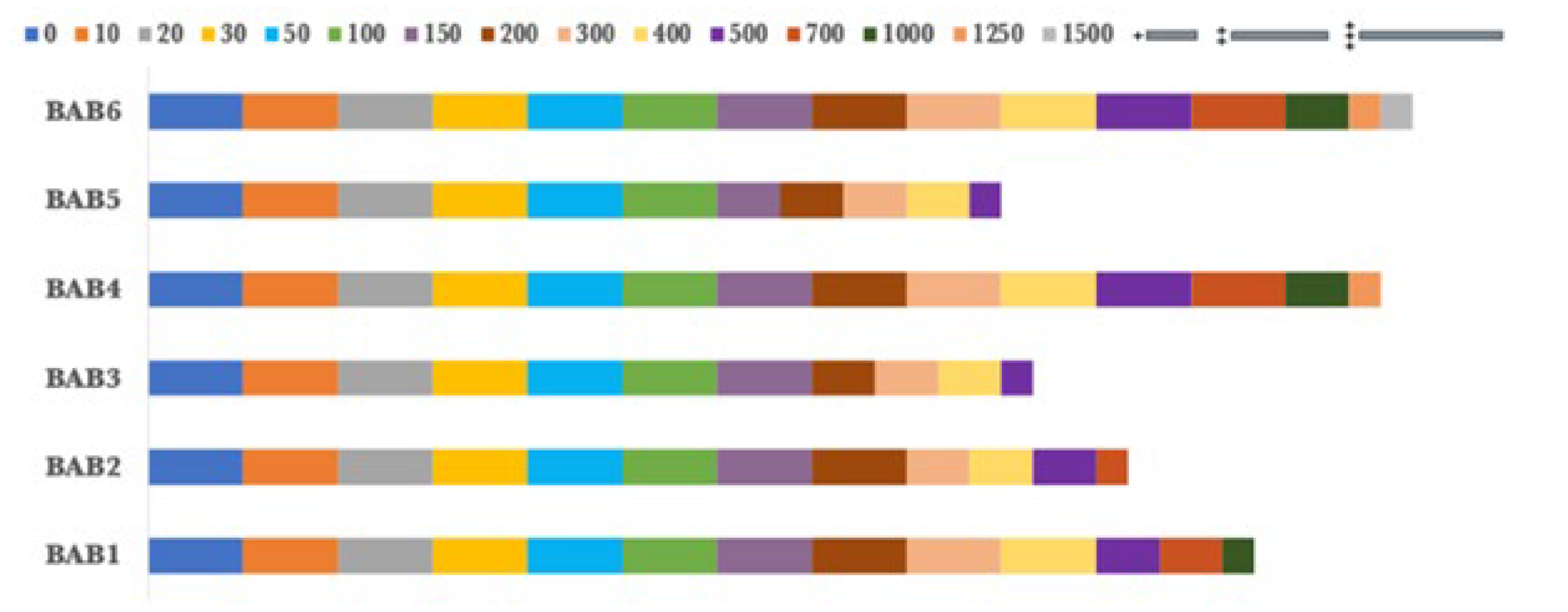
Figure 11.
Phylogenetic analysis of 16S rRNA sequences of the bacterial isolates with the sequences retrieved from NCBI (National Center for Biotechnology Information).
Figure 11.
Phylogenetic analysis of 16S rRNA sequences of the bacterial isolates with the sequences retrieved from NCBI (National Center for Biotechnology Information).

Table 1.
Phosphate solubilization index (PSI) and IAA, ammonia and HCN production rates by the selected endophytic bacteria strains.
Table 1.
Phosphate solubilization index (PSI) and IAA, ammonia and HCN production rates by the selected endophytic bacteria strains.
 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



