
Submitted:
02 July 2024
Posted:
03 July 2024
You are already at the latest version
Preprints on COVID-19 and SARS-CoV-2
Abstract
The COVID-19 pandemic was characterized by the emergence and succession of SARS-CoV-2 variants able to evade the antibody response induced by natural infection and vaccination. In this study, IgG reactivity and neutralizing capacity of the serum of individuals vaccinated with Sputnik V was determined against the spike glycoprotein (S) of the ancestral and different viral variants. At 42 days’ post vaccination, the frequency of recognition and reactivity to S protein of the omicron variant was lower compared to the other variants. There was higher reactivity to Alpha and Delta S protein compared to ancestral S protein. A higher neutralization titer was seen against the ancestral variant compared to the variants, especially Omicron. Some sera exhibited a higher neutralization titer to the Gamma variant, compared to the ancestral variant. This may be due to exposure to this variant during the period of its spread in the country, which coincided with the period of the clinical trial. A correlation was found between reactivity to S and neutralization titer to each variant. These results suggest that antibodies induced by Sputnik-V can recognize, persist and neutralize the SARS-CoV-2 variants circulating in Venezuela, with Omicron being the one that best evades this response.
Keywords:
SARS-CoV-2
; Sputnik-V
; antibodies
; variants
; ELISA
; neutralization
1. Introduction
By March 2024, SARS-CoV-2, the virus responsible for the COVID-19 pandemic, had caused more than 700 million cases and claimed over 7 million lives [1], despite estimations suggesting even higher figures [2,3]. Notably, this viral family possesses a unique non-structural protein with proof-reading capacity, setting it apart from other RNA viruses. The virus has exhibited many mutations, due to numerous replication cycles in millions of hosts, a high frequency of recombination, and the effect of host editing enzymes [4].
Consequently, variants with higher transmission capacity and immune evasion have been selected [5]. Five variants of concern (VOCs) of SARS-CoV-2 have emerged since the end of 2020, resulting in new waves of infections, deaths, and chronic sequelae [6]: Alpha (Original lineage B.1.1.7), which arose in the UK, Beta (B.1.351), which appeared in South Africa, Gamma (B.1.1.28.2, P.1), which originated in Brazil, Delta (B.1.617.2), from India and Omicron (B.1.1.529) first identified in South Africa and whose sub-lineages are the only ones circulating at present [7]. Furthermore, the antigenic evolution of the S protein has dampened the efficacy and effectiveness of many vaccines [8].
Non-replicating adenoviral vectors, mRNA, whole inactivated viruses, and protein subunit-based vaccines are powerful tools in the fight against emergent pathogens, including SARS-CoV-2. Prior to the introduction of bivalent adapted boosters, most vaccine-approved candidates targeted the S protein of the ancestral variant first identified in China [9]. The Gam-COVID-Vac (Sputnik V) vaccine has played a significant role in vaccination campaigns in Venezuela and other Latin American countries. Studies have consistently shown that it is as effective as mRNA vaccines [10]. Sputnik V is composed of two non-replicative recombinant human adenoviruses (serotypes 26 and 5), both carrying the coding sequence for the ancestral S protein. The vectors have deletions in the early expression genes E1 and E3, which prevents its replication and admit the introduction of the gene coding for SARS-CoV-2 S glycoprotein. Additionally, the heterologous design of Sputnik-V is advantageous as it reduces immunogenicity issues arising from immunity against the vector [11]. Upon delivery, each antigenic stimulus produces antibodies through plasmatic cells, diversifies the B cell receptor (BCR) repertoire, and generates specific B memory cells against the entire S glycoprotein [12-14].
Understanding how the recognition and neutralization of antibodies generated after Sputnik-V vaccination might be affected by variants of this virus is an issue that requires further research. In contrast to mRNA-based vaccines, the effect of Sputnik V vaccine has been significantly less studied. In fact, a PubMed query shows the scarce number of publications about the Sputnik V vaccine compared to other most common ones, even though its use was approved in more than 70 countries (Table 1). In Venezuela, a clinical trial was conducted using the Sputnik V vaccine, from December 2020 to June 2021, time which coincide with the emergence of the Gamma VOC in the country [15]. The aim of this study was to evaluate the antibody reactivity and neutralization ability to SARS-CoV-2 variants and to assess the correlation between these parameters in the vaccinees in Venezuela.
2. Materials and Methods
2.1. Experimental Design, population and sample
Sera samples from the prospective, double-blind, randomized, placebo-controlled clinical trial of Sputnik V in Venezuela from December 2020 to July 2021 (NCT04642339) were evaluated. The inclusion and exclusion criteria are detailed on the clinical trial website (clinicaltrials.gov/ct2/show/NCT04642339). Volunteers were residents of Caracas, aged 20-80 years, and all signed an informed consent form approved by the National Ethics Committee for Research in COVID-19. Samples were collected 42 days after vaccination (dpv), counting after the first dose and 21 days after the second dose. Matching samples from 46 vaccinees were also collected at 180 dpv. Of the 133 volunteers, 105 were vaccinated and 28 received placebo. Sera from 85/105 vaccinees without serological evidence of exposure, as determined by nucleoprotein (N) reactivity [18], were analyzed for reactivity to S and RBD antigens at 42 dpv. For comparisons of 42 versus 180 dpv reactivity, 20 sera from N-negative vaccinees were selected. 18 samples from this subgroup were used for neutralization assays at 42 and 180 dpv.
2.2. Antigens
The recombinant antigens for the enzyme-linked immunosorbent assays (ELISA) were acquired from MyBioSource Inc. (San Diego, CA, USA). The ancestral S protein (MBS8574721) and RBD (MBS8574741), Alpha S (MBS184025), Beta S (MBS184021), Gamma S (MBS184022) and RBD (MBS434292), Delta S (MBS184024) and Omicron S (MBS553745) antigens, were employed. The concentration of antigens was determined using Qubit™ Protein Assay according to manufacturer’s specifications (catalog number Q32866, Thermo Fisher, Waltham, MA, USA).
2.3. Reactivity towards SARS-CoV-2 antigens
To assess the differences between reactivity against S or RBD from SARS-CoV-2 variants, an adapted version of the protocol proposed by Stadlbauer et al., 2020 was used [19] and thoroughly described by Cornejo et al., 2024 [18]. Briefly, 96-wells microtiter plates were coated using 1 µg/mL of each capture antigen. To prevent unspecific antibody binding commercial blocking solution was employed (Abcam, Cambridge, United Kingdom, ab126587). All samples were diluted 1/100 and incubated for 2 hours at 37 °C. Horseradish peroxidase (HRP) IgG-linked (Jackson ImmunoResearch Inc, West Grove, PA, USA,) was used as a detection antibody (1/70,000) and incubated for 1 hour. TMB (3,3′,5,5′-tetramethylbenzidine) was used as a chromogenic substrate of HRP. All washes were done with 0.01% PBS-Tween. HCl [3M] was used to stop the reaction. Data gathering was achieved with a spectrophotometer (SpectraMax 250, Hampton, NA, USA) at 450 nm. Reactivity against SARS-CoV-2 antigens was assessed within this group of 134 individuals. The optical densities (O.D.) of the blank were subtracted from the O.D of the samples. O.D exceeding the cut-off established with negative control mean plus 3 standard deviations were considered as responders to each antigen. The negative control was established with 18 prepandemic sera from apparently healthy individuals. A mixture of two sera from individuals with hybrid immunity (Sputnik V-vaccinated and two confirmed infections by PCR) was used as a positive control. Both controls were used during normalization to estimate the relative IgG levels using the sample-to-positive ratio (S/P) previously described [20]:
2.4. Plaque Reduction Neutralization Test of Sputnik V vaccinees sera
A plaque reduction neutralization test (PRNT) was conducted on 18 samples of serum collected at 42 days post-vaccination (dpv) and 180 dpv. PRNT was performed according to a previously reported procedure [18,21]. VERO C1008 cells (Vero 76, clone E6, vero. ATCC, Manassas, VA, USA), were maintained at 37 °C and 5% CO2 in RPMI medium (Thermo Fisher Scientific, 11875093) supplemented with 10% fetal bovine serum (FBS, Thermo Fisher Scientific, 16000044) and 1% penicillin–streptomycin (Thermo Fisher Scientific, Waltham, MA, USA). All procedures were performed in a biosafety cabinet class II (ESCO®, Airstream®, Singapore) within a level 3 biosecurity laboratory. Viral seeds of the different SARS-CoV-2 variant strains: ancestral (B.1.1.33), P.1 (Gamma VOC), AY122 (Delta VOC), B.1.621 (Mu VOI) and BA.1.1 (Omicron VOC) [18], were incubated for 30 min at 37 °C with different dilutions of vaccinated and control sera. The infection process was performed for one hour at 37 °C in a 5% CO2 atmosphere, and the cells were washed twice with PBS to remove the non-internalized viruses. The cells were overlayed with 0.5% carboxymethyl cellulose in a culture medium and incubated for 72 h. Then, the cells were fixed with 4% formaldehyde and stained with crystal violet to count the number of lytic plaques under each condition. Highly responsive mixed of hybrid sera was used as a positive control whereas culture medium without serum or viral seeds was used as the mock control.
The PRNT50 of each serum was determined through a nonlinear regression test, defining this value as the reverse of the dilution at which 50% of the virus is neutralized. After lytic plaque counting the percentage of neutralization for each serum dilution was calculated using the following formula:
Afterwards, Log2 transformed reciprocal of each serum dilution (X axis) vs the percentage of neutralization (Y axis) was plotted and fitted to a non-linear regression variable slope dose-response inhibition model to determine the dilution needed to reduce 50% of lytic plaque formation observed in the infection control (IC).
2.5. Statistical analysis
To evaluate differences between reactivities against the different antigens at 42 dpv the Kruskal Wallis test (p < 0.05) was used with Dunn’s post-hoc analysis. For comparisons between 42 and 180 dpv a Wilcoxon matched-pairs signed rank test was carried out to estimate significant differences of reactivity between both times (p < 0.05). To establish differences among neutralization titers towards the different SARS-CoV-2 isolates within 42 and 180 dpv a Wilcoxon matched-pairs signed rank test was conducted (p < 0.05, PRISM GraphPad 9.0 ©, California, USA). Statistical difference was assessed with Chi square test (Epi Info™, Centers for Disease Control and Prevention, Atlanta, USA) for frequency comparisons among groups. Association between vaccinees ages, IgG antibody reactivity and PRNT50 was determined through a hierarchical clustering correlation matrix heatmap by the Spearman method with a confidence interval (CI) of 95% employing the “visdat” and “ggcorrplot” packages of R version 4.3.3. To explore patterns of antibody response an ascending hierarchical classification of the individuals was performed using the “shiny” package of R version 4.3.3.
3. Results
3.1. Reactivity to SARS-CoV-2 variants S protein
The reactivity to S from the 5 VOCs was assessed by ELISA in 85 vaccinated volunteers, without evidence of exposure to the virus. Statistically significant differences were observed between the mean reactivities of the S protein of the different variants. Reactivity to the S protein of the Alpha and Delta variants was higher than that observed for the ancestral variant (p <00001). No significant differences were observed between the reactivity to the S protein of the ancestral variant and that of the beta and gamma variants. The mean reactivity to the ancestral variant was higher (3-fold) than the one to the Omicron variant (p <00001). Almost all the sera reacted to the S protein of the different variants, except for the Omicron one, which was only recognized by 67% of the sera, a frequency statistically lower (p<0.0001) (Figure 1).
The reactivity to S VOCs was also assessed in follow up sera. The S/P values were lower at 180 dpv, with a reduction of 3-fold (S ancestral), 2-fold (S alpha, beta, gamma and delta) and 3-fold (S omicron). However, no reduction was observed in the number of sera recognizing each variant: an even higher frequency of sera recognizing the S protein was observed. The exception was the Omicron variant, for which even less sera recognized the protein, although this difference was not significant (Figure 2).
3.2. Neutralizing activity of Sputnik-V-induced antibodies against SARS-CoV-2 variants at 42 and 180 dpv.
Plaque reduction neutralization titer was assessed on 18 sera taken at 42 and 180 dpv from vaccinees with no signs of infection by the virus (Figure 3A). A modest reduction in the neutralization titer was observed with the Delta variants at 42 and 180 dpv, compared to the ancestral one, while the mean titer of neutralization against the Gamma variant was higher compared to the ancestral one, although this difference was not significant. In the case of Omicron, a significant reduction in neutralization titer was observed at 42 and 180 dpv, of 6 and 7 times, respectively, in addition to a significant reduction in the number of sera neutralizing the variant.
When dissecting the neutralizing activity against the Gamma variant, a group of sera (n = 10) exhibited a modest decrease in neutralizing titers against it at 42 and 180 dpv (Figure 3B), while a 3.7-fold increase in neutralization titer was observed against this variant in the sera of the other group (n = 8), at 42 and 180 dpv compared to the ancestral variant (Figure 3C).
Because of the unexpected higher neutralization activity in some sera of the Gamma variant respect the ancestral strain, the reactivity to the S and RBD antigens of the SARS-CoV-2 variants was evaluated for the sera analyzed for neutralizing activity. A statistically significant higher reactivity to the S Gamma variant, compared to the ancestral protein, was observed in the sera exhibiting higher neutralizing activity against this variant. This difference was not observed in the reactivity to RBD (Figure 4B and D).
3.3. Correlation between age, reactivity and PRNT50.
The Spearman test was performed to correlate all the parameters analyzed in this study. There was no significant correlation between antibody levels, neutralization titers and the age of the vaccinees. Regarding associations between ELISA reactivity and PRNT50, strong (S gamma vs PRNT50 gamma) to moderate positive correlations were observed at 42 dpv. Conversely, moderate (S omicron vs PRNT50 omicron) to low correlations were observed at 180 dpv (Figure 5).
3.5. Signatures of antibody immune response in Sputnik V vaccinees.
An ascending hierarchical classification of the vaccinees was performed to compare their pattern of reactivity and neutralization. The classification made on individuals revealed two main patterns of response. One main cluster grouped most of the vaccinees (low responders, n=14), and the remaining (high responders, n=4) with a high reactivity and PRNT50 titer to Gamma, Delta or Omicron variants (Figure 6).
4. Discussion
As vaccines were based on ancestral antigens and because of the evolution of SARS-CoV-2, it was critical to determine whether the binding, neutralization and life span of antibodies induced after vaccination were affected by the new variants. Sputnik-V played a pivotal role in the vaccination initiatives throughout Venezuela and Latin America, so it was essential to assess the antibody response elicited by it in the face of the sequential emergence of variants of concern, considering moreover the lesser amount of information available for this vaccine compared to mRNA ones. In order to perform comparisons of reactivity among antigens and to assess how these changed over time, we used an in-house ELISA. On the other hand, we used the PRNT to estimate the neutralization titers of these sera. We also evaluated the possible correlation between these variables.
No differences in reactivity to Beta and Gamma compared to ancestral antigens were reported. Other authors have described a reduced reactivity and neutralization of Sputnik-V-induced antibodies against these variants [13,22]. Interestingly, the S antigen from the Alpha and Delta variants were recognized with a higher reactivity compared to the ancestral S protein at 42 dpv, and for Delta also at 180 dpv, consistent with the presence of high affinity and neutralizing antibodies generated by this vaccine against SARS-CoV-2 variant antigens [23]. With the exception of Omicron, where a significant decrease in reactivity to S and RBD antigens (RBD, data not shown) was also observed at both time points, seroconversion to all other antigens ranged from 88 to 99% and correlated with the efficacy of this vaccine as previously reported [24,25]. Although reactivity tended to decline over time, the vaccine maintained an effective response at six months. The durable antibody response induced by this vaccine has also been described by others [18,26,27]. These observations support the notion that the antibody response generated by immunization with Sputnik-V persists for at least 6 months after administration of the first vaccine component.
Higher neutralization titers were reported against the ancestral variant compared to the other variants, and Omicron was the variant for which the highest reduction in neutralization titer was observed. Omicron and its sub-lineages have exhibited greater antigenic variation than previous variants, significantly altering the patterns of recognition and neutralization of vaccine-induced antibodies [28]. This change in viral behavior threatens potential outbreaks from other emerging SARS-CoV-2 variants or new coronaviruses. Although the neutralization titer against Delta and Omicron is reduced, we observed that some sera were able to recognize and neutralize these variants. A possible explanation is that after Sputnik-V injection, naïve B cells undergo germinal center (GC) selection by somatic hypermutation (SHM) of variable heavy (VH) and variable light (VL) chain genes, thus stimulating B cell receptor (BCR) repertoire diversification, increasing binding affinity and neutralizing activity, which are considered indicators of antibody affinity maturation [29]. Indeed, mRNA vaccines induce a robust B cell, germinal center and follicular helper T cell (Tfh) response, which increases after the second dose [30]. Furthermore, Tfhs have been detected in blood and lymph nodes for at least 200 days after vaccination [31]. In addition, memory B cell clones that interact with the RBD of the alpha, beta and delta variants have higher levels of SHM than those that bind only to the RBD of the ancestral variant after mRNA vaccination [32]. In addition, the persistence of GC activity after immunization, and therefore SHM, appears to promote a protective antibody response against variants of this virus [33] by increasing the likelihood of cross-neutralization of other variants, a response already reported during the convalescent phase [34,35,36].
This persistent GC activity could be explained by antigenic persistence of adenovirus vectors or mRNA after immunization in different tissues [37,38]. In addition, persistence of S and S1 antigens has been reported in some mRNA vaccinated individuals for up to 60 days after immunization [39,40]. The persistence, distribution in primary and secondary lymphoid organs and prolonged activation of GC due to the capture and prolonged presentation of S, S1, S2, RBD and peptides could support that GC activity persists for periods of time yet to be defined after vaccination, thus promoting the generation of antibodies with greater diversity and affinity for epitopes of the S protein. Indeed, there is evidence that structurally undamaged antigens are preserved in B cell follicles due to low protease activity in dendritic cells located in this compartment, which may explain the long-term availability of antigens after immunization, thus facilitating antigen presentation for a prolonged period [41,42]. Furthermore, the presence of B cells in germinal centers that recognize protein S for at least 30 weeks after immunization with mRNA has been described [30,33].
To our knowledge, GC activity in tissues from Sputnik-V vaccinated individuals remains to be elucidated. However, the same mechanisms observed in mRNA vaccines may be operating to counteract SARS-CoV-2n variants. GC activity has been described in other vaccines using both adenoviruses used in Sputnik-V [43,44,45]. Furthermore, an increase in memory, antibody-secreting, FcRL4+ and Tfh cells has been reported after immunization with BNT162b2, AZD1222 and Sputnik-V [46]. In addition, an increase in cytokines that mediate B- and Tfh cell proliferation, differentiation, localization, interaction, and antibody class switching [47] such as IL-6, IL-10, IFN-γ, IL-2, IL-17A, and sCD40L, has been identified [48], suggesting GC activity after administration of this adenoviral vaccine.
A subset of sera exhibited higher neutralization titers to the Gamma VOC compared to the ancestral variant. This observation suggests that these individuals might have been exposed to this variant during the study period, as it was disseminating in Venezuela during March-August 2021 [15,49]. None of these volunteers reported an infectious event during the clinical trial. These sera showed no reactivity to the N protein [18], which would have been expected in the event of infection, and a modest increase in reactivity to S but not to RBD Gamma VOC. We suggest that they might had an asymptomatic or subclinical infection, related to the protective effect of the vaccine received, manifested only at the presence of neutralizing antibodies against the infecting variant, and a modest increase in reactivity to the Gamma S protein variant. Other authors have found neutralizing antibodies in asymptomatic or mildly affected COVID-19 individuals with no detectable IgG antibodies [50]. Similar results (higher titer against the Gamma variant without apparent previous infection) were obtained by another group who studied the neutralization of Sputnik V-induced antibodies in a cohort of volunteers in Argentina [51]. On the other hand, the fact the sera from our study exhibited just a modest increase in reactivity towards the S protein, and not the RBD region, might be due to the fact that other regions of the S protein, like the N-terminal region, might be contributing in inducing neutralizing antibodies against the virus, as suggested by our previous study [18].
Taken together, our results suggest that the Sputnik-V vaccine induces antibodies that are similarly able to interact with S and neutralizing SARS-CoV-2 variants, with the Omicron VOC being the most resistant to antibody immunity, as previously reported [14,25]. Some volunteers may have been exposed to the Gamma variant during the clinical trial, without showing symptoms. This reinforces the significant beneficial effect of the COVID-19 vaccines in protecting, if not completely against infection, against symptomatic infection due to variant emergence, and then against severe disease manifestation, death and long-term sequelae [52,53]. The antigenic distance between different variants enhances our understanding of how immunized populations may be susceptible to the emergence of new variants, and has been and should be continued to be considered in the design of the next generation of vaccines [8,54]. Therefore, continuous monitoring of SARS-CoV-2 evolutionary trajectories should be a priority in the design of new vaccines that include immunogens that promote GC responses to provide broader protection against infection and disease caused by emerging variants. We provide further evidence on the magnitude, breadth and persistence of antibodies induced by this adenoviral vaccine in the context of SARS-CoV-2 evolution, providing insights for the design of future vaccines against new potential coronaviruses.
Author Contributions
Conceptualization, supervision and project administration, F.H.P., A.G., H.R.R., J.L.Z. and F.L.; investigation, C.F., A.C., M.R.N., D.J.G. and R.C.J.; resources, I.B., S.M., M.H. and N.P.G., writing—original draft preparation, C.F.; writing—review and editing, C.F., A.C., F.H.P., F.L., R.C.J. and J.L.Z.; funding acquisition, F.H.P., H.R.R. and A.G. All authors have made substantial contributions to: the conception or design of the work or the acquisition, analysis or interpretation of data for the work; drafting the work or revising it critically for important intellectual content; final approval of the version to be published; agreement to be accountable for all aspects of the work in ensuring that questions related to the accuracy or integrity of any part of the work are appropriately investigated and resolved. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Ministerio del Poder Popular de Ciencia, Tecnología e Innovación of Venezuela.
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki and approval was obtained from the National Ethics Committee for Research in COVID-19 of Venezuela, G.O.41.962.
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
The complete genome sequences of the SARS-CoV-2 variants used for neutralization assays have been deposited in the GISAID database accession numbers: ancestral (EPI_ISL_6980947), gamma (EPI_ISL_2628299), delta (EPI_ISL_6976265) and omicron (EPI_ISL_17389567).
Acknowledgments
The authors are indebted to all members of the clinical trial team. We are also indebted to the volunteers who acceded to participate in the clinical trial. This study was supported by the Ministerio del Poder Popular de Ciencia, Tecnología e Innovación of Venezuela.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study, in the collection, analyses or interpretation of data, in the writing of the manuscript or in the decision to publish the results.
References
- World Health Organization WHO Data. Available online: https://data.who.int/dashboards/covid19/cases?n=c (accessed on 26 March 2024).
- Barber, R.M.; Sorensen, R.J.D.; Pigott, D.M.; Bisignano, C.; Carter, A.; Amlag, J.O.; Collins, J.K.; Abbafati, C.; Adolph, C.; Allorant, A.; et al. Estimating Global, Regional, and National Daily and Cumulative Infections with SARS-CoV-2 through Nov 14, 2021: A Statistical Analysis. The Lancet 2022, 399, 2351–2380. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Paulson, K.R.; Pease, S.A.; Watson, S.; Comfort, H.; Zheng, P.; Aravkin, A.Y.; Bisignano, C.; Barber, R.M.; Alam, T.; et al. Estimating Excess Mortality Due to the COVID-19 Pandemic: A Systematic Analysis of COVID-19-Related Mortality, 2020–21. The Lancet 2022, 399, 1513–1536. [Google Scholar] [CrossRef]
- Wei, L. Retrospect of the Two-Year Debate: What Fuels the Evolution of SARS-CoV-2: RNA Editing or Replication Error? Curr Microbiol 2023, 80, 151. [Google Scholar] [CrossRef] [PubMed]
- Carabelli, A.M.; Peacock, T.P.; Thorne, L.G.; Harvey, W.T.; Hughes, J.; de Silva, T.I.; Peacock, S.J.; Barclay, W.S.; de Silva, T.I.; Towers, G.J.; et al. SARS-CoV-2 Variant Biology: Immune Escape, Transmission and Fitness. Nat Rev Microbiol 2023, 21, 162–177. [Google Scholar] [PubMed]
- Sherif, Z.A.; Gomez, C.R.; Connors, T.J.; Henrich, T.J.; Reeves, W.B. Pathogenic Mechanisms of Post-Acute Sequelae of SARS-CoV-2 Infection (PASC). Elife 2023, 12. [Google Scholar]
- Ghildiyal, T.; Rai, N.; Mishra Rawat, J.; Singh, M.; Anand, J.; Pant, G.; Kumar, G.; Shidiki, A. Challenges in Emerging Vaccines and Future Promising Candidates against SARS-CoV-2 Variants. J Immunol Res 2024, 2024. [Google Scholar] [CrossRef]
- Mykytyn, A.Z.; Rissmann, M.; Kok, A.; Rosu, M.E.; Schipper, D.; Breugem, T.I.; van den Doel, P.B.; Chandler, F.; Bestebroer, T.; de Wit, M.; et al. Antigenic Cartography of SARS-CoV-2 Reveals That Omicron BA.1 and BA.2 Are Antigenically Distinct. Sci. Immunol 2022, 7, 4450. [Google Scholar] [CrossRef] [PubMed]
- Mendonça, S.A.; Lorincz, R.; Boucher, P.; Curiel, D.T. Adenoviral Vector Vaccine Platforms in the SARS-CoV-2 Pandemic. NPJ Vaccines 2021, 6. [Google Scholar] [CrossRef]
- Fiolet, T.; Kherabi, Y.; MacDonald, C.J.; Ghosn, J.; Peiffer-Smadja, N. Comparing COVID-19 Vaccines for Their Characteristics, Efficacy and Effectiveness against SARS-CoV-2 and Variants of Concern: A Narrative Review. Clinical Microbiology and Infection 2022, 28, 202–221. [Google Scholar] [PubMed]
- Logunov, D.Y.; Dolzhikova, I.V.; Zubkova, O.V.; Tukhvatullin, A.I.; Shcheblyakov, D.V.; Dzharullaeva, A.S.; Grousova, D.M.; Erokhova, A.S.; Kovyrshina, A.V.; Botikov, A.G.; et al. Safety and Immunogenicity of an RAd26 and RAd5 Vector-Based Heterologous Prime-Boost COVID-19 Vaccine in Two Formulations: Two Open, Non-Randomised Phase 1/2 Studies from Russia. The Lancet 2020, 396, 887–897. [Google Scholar] [CrossRef]
- Martynova, E.; Hamza, S.; Garanina, E.E.; Kabwe, E.; Markelova, M.; Shakirova, V.; Khaertynova, I.M.; Kaushal, N.; Baranwal, M.; Rizvanov, A.A.; et al. Long Term Immune Response Produced by the Sputnikv Vaccine. Int J Mol Sci 2021, 22. [Google Scholar] [CrossRef] [PubMed]
- Ikegame, S.; Siddiquey, M.N.A.; Hung, C.T.; Haas, G.; Brambilla, L.; Oguntuyo, K.Y.; Kowdle, S.; Chiu, H.P.; Stevens, C.S.; Vilardo, A.E.; et al. Neutralizing Activity of Sputnik V Vaccine Sera against SARS-CoV-2 Variants. Nat Commun 2021, 12. [Google Scholar] [CrossRef] [PubMed]
- Gushchin, V.A.; Dolzhikova, I.V.; Shchetinin, A.M.; Odintsova, A.S.; Siniavin, A.E.; Nikiforova, M.A.; Pochtovyi, A.A.; Shidlovskaya, E.V.; Kuznetsova, N.A.; Burgasova, O.A.; et al. Neutralizing Activity of Sera from Sputnik V-Vaccinated People against Variants of Concern (VOC: B.1.1.7, B.1.351, P.1, B.1.617.2, B.1.617.3) and Moscow Endemic SARS-CoV-2 Variants. Vaccines (Basel) 2021, 9. [Google Scholar] [CrossRef] [PubMed]
- Jaspe, R.C.; Loureiro, C.L.; Sulbaran, Y.; Moros, Z.C.; D’Angelo, P.; Rodríguez, L.; Zambrano, J.L.; Hidalgo, M.; Vizzi, E.; Alarcón, V.; et al. Introduction and Rapid Dissemination of SARS-CoV-2 Gamma Variant of Concern in Venezuela. Infection, Genetics and Evolution 2021, 96. [Google Scholar] [CrossRef] [PubMed]
- Kudlay, D.; Svistunov, A. COVID-19 Vaccines: An Overview of Different Platforms. Bioengineering 2022, 9. [Google Scholar]
- Basta, N.; Moodie, E. COVID19 VACCINE TRACKER. Available online: https://covid19.trackvaccines.org/vaccines/approved/ (accessed on 30 June 2024).
- Cornejo, A.; Franco, C.; Rodríguez, M.; García, A.; Belisario, I.; Mayoría, S.; Garzaro, D.J.; Zambrano, J.L.; Jaspe, R.C.; Hidalgo, M.; et al. Humoral Immunity across the SARS-CoV-2 Spike after Sputnik V (Gam-COVID-Vac) Vaccination. Antibodies 2024, 13, 41. [Google Scholar] [CrossRef] [PubMed]
- Stadlbauer, D.; Amanat, F.; Chromikova, V.; Jiang, K.; Strohmeier, S.; Arunkumar, G.A.; Tan, J.; Bhavsar, D.; Capuano, C.; Kirkpatrick, E.; et al. SARS-CoV-2 Seroconversion in Humans: A Detailed Protocol for a Serological Assay, Antigen Production, and Test Setup. Curr Protoc Microbiol 2020, 57. [Google Scholar] [CrossRef]
- Claro, F.; Silva, D.; Rodriguez, M.; Rangel, H.R.; de Waard, J.H. Immunoglobulin G Antibody Response to the Sputnik V Vaccine: Previous SARS-CoV-2 Seropositive Individuals May Need Just One Vaccine Dose. International Journal of Infectious Diseases 2021, 111, 261–266. [Google Scholar] [CrossRef]
- Rodriguez-Nuñez, M.; Cepeda, M. del V.; Bello, C.; Lopez, M.A.; Sulbaran, Y.; Loureiro, C.L.; Liprandi, F.; Jaspe, R.C.; Pujol, F.H.; Rangel, H.R. Neutralization of Different Variants of SARS-CoV-2 by a F(Ab′)2 Preparation from Sera of Horses Immunized with the Viral Receptor Binding Domain. Antibodies 2023, 12. [Google Scholar] [CrossRef]
- Gonzalez, M.M.; Ledesma, L.; Sanchez, L.; Ojeda, D.S.; Rouco, S.O.; Rossi, A.H.; Varese, A.; Mazzitelli, I.; Pascuale, C.A.; Miglietta, E.A.; et al. Longitudinal Study after Sputnik V Vaccination Shows Durable SARS-CoV-2 Neutralizing Antibodies and Reduced Viral Variant Escape to Neutralization over Time. mBio 2022, 13, e03442–21. [Google Scholar]
- Röltgen, K.; Nielsen, S.C.A.; Silva, O.; Younes, S.F.; Zaslavsky, M.; Costales, C.; Yang, F.; Wirz, O.F.; Solis, D.; Hoh, R.A.; et al. Immune Imprinting, Breadth of Variant Recognition, and Germinal Center Response in Human SARS-CoV-2 Infection and Vaccination. Cell 2022, 185, 1025–1040.e14. [Google Scholar] [CrossRef] [PubMed]
- Logunov, D.Y.; Dolzhikova, I.V.; Shcheblyakov, D.V.; Tukhvatulin, A.I.; Zubkova, O.V.; Dzharullaeva, A.S.; Kovyrshina, A.V.; Lubenets, N.L.; Grousova, D.M.; Erokhova, A.S.; et al. Safety and Efficacy of an RAd26 and RAd5 Vector-Based Heterologous Prime-Boost COVID-19 Vaccine: An Interim Analysis of a Randomised Controlled Phase 3 Trial in Russia. The Lancet 2021, 397, 671–681. [Google Scholar] [CrossRef] [PubMed]
- Radion, E.I.; Mukhin, V.E.; Kholodova, A.V.; Vladimirov, I.S.; Alsaeva, D.Y.; Zhdanova, A.S.; Ulasova, N.Y.; Bulanova, N.V.; Makarov, V.V.; Keskinov, A.A.; et al. Functional Characteristics of Serum Anti-SARS-CoV-2 Antibodies against Delta and Omicron Variants after Vaccination with Sputnik V. Viruses 2023, 15. [Google Scholar] [CrossRef] [PubMed]
- Claro, F.; Silva, D.; Pérez Bogado, J.A.; Rangel, H.R.; de Waard, J.H. Lasting SARS-CoV-2 Specific IgG Antibody Response in Health Care Workers from Venezuela, 6 Months after Vaccination with Sputnik V. International Journal of Infectious Diseases 2022, 122, 850–854. [Google Scholar] [CrossRef] [PubMed]
- Lapa, D.; Grousova, D.M.; Matusali, G.; Meschi, S.; Colavita, F.; Bettini, A.; Gramigna, G.; Francalancia, M.; Garbuglia, A.R.; Girardi, E.; et al. Retention of Neutralizing Response against SARS-CoV-2 Omicron Variant in Sputnik V-Vaccinated Individuals. Vaccines (Basel) 2022, 10. [Google Scholar] [CrossRef] [PubMed]
- Greaney, A.J.; Starr, T.N.; Gilchuk, P.; Zost, S.J.; Binshtein, E.; Loes, A.N.; Hilton, S.K.; Huddleston, J.; Eguia, R.; Crawford, K.H.D.; et al. Complete Mapping of Mutations to the SARS-CoV-2 Spike Receptor-Binding Domain That Escape Antibody Recognition. Cell Host Microbe 2021, 29, 44–57.e9. [Google Scholar] [CrossRef] [PubMed]
- Victora, G.D.; Nussenzweig, M.C. Germinal Centers. Annu Rev Immunol 2022, 30, 429–457. [Google Scholar] [CrossRef]
- Turner, J.S.; O’Halloran, J.A.; Kalaidina, E.; Kim, W.; Schmitz, A.J.; Zhou, J.Q.; Lei, T.; Thapa, M.; Chen, R.E.; Case, J.B.; et al. SARS-CoV-2 MRNA Vaccines Induce Persistent Human Germinal Centre Responses. Nature 2021, 596, 109–113. [Google Scholar] [CrossRef]
- Mudd, P.A.; Minervina, A.A.; Pogorelyy, M.V.; Turner, J.S.; Kim, W.; Kalaidina, E.; Petersen, J.; Schmitz, A.J.; Lei, T.; Haile, A.; et al. SARS-CoV-2 MRNA Vaccination Elicits a Robust and Persistent T Follicular Helper Cell Response in Humans. Cell 2022, 185, 603–613.e15. [Google Scholar] [CrossRef]
- Goel, R.R.; Painter, M.M.; Apostolidis, S.A.; Mathew, D.; Meng, W.; Rosenfeld, A.M.; Lundgreen, K.A.; Reynaldi, A.; Khoury, D.S.; Pattekar, A.; et al. MRNA Vaccines Induce Durable Immune Memory to SARS-CoV-2 and Variants of Concern. Science (1979) 2021, 374. [Google Scholar] [CrossRef]
- Kim, W.; Zhou, J.Q.; Horvath, S.C.; Schmitz, A.J.; Sturtz, A.J.; Lei, T.; Liu, Z.; Kalaidina, E.; Thapa, M.; Alsoussi, W.B.; et al. Germinal Centre-Driven Maturation of B Cell Response to MRNA Vaccination. Nature 2022, 604, 141–145. [Google Scholar] [CrossRef] [PubMed]
- Sakharkar, M.; Rappazzo, G.; Wieland-Alter, W.F.; Hsieh, C.L.; Wrapp, D.; Esterman, E.S.; Kaku, C.I.; Wec, A.Z.; Geoghegan, J.C.; McLellan, J.S.; et al. Prolonged Evolution of the Human B Cell Response to SARS-CoV-2 Infection. Sci Immunol 2021, 6. [Google Scholar] [CrossRef] [PubMed]
- Moriyama, S.; Adachi, Y.; Sato, T.; Tonouchi, K.; Sun, L.; Fukushi, S.; Yamada, S.; Kinoshita, H.; Nojima, K.; Kanno, T.; et al. Temporal Maturation of Neutralizing Antibodies in COVID-19 Convalescent Individuals Improves Potency and Breadth to Circulating SARS-CoV-2 Variants. Immunity 2021, 54, 1841–1852.e4. [Google Scholar] [CrossRef] [PubMed]
- Gaebler, C.; Wang, Z.; Lorenzi, J.C.C.; Muecksch, F.; Finkin, S.; Tokuyama, M.; Cho, A.; Jankovic, M.; Schaefer-Babajew, D.; Oliveira, T.Y.; et al. Evolution of Antibody Immunity to SARS-CoV-2. Nature 2021, 591, 639–644. [Google Scholar] [CrossRef] [PubMed]
- Krauson, A.J.; Casimero, F.V.C.; Siddiquee, Z.; Stone, J.R. Duration of SARS-CoV-2 MRNA Vaccine Persistence and Factors Associated with Cardiac Involvement in Recently Vaccinated Patients. NPJ Vaccines 2023, 8. [Google Scholar] [CrossRef] [PubMed]
- Stebbings, R.; Armour, G.; Pettis, V.; Goodman, J. AZD1222 (ChAdOx1 NCov-19): A Single-Dose Biodistribution Study in Mice. Vaccine 2022, 40, 192–195. [Google Scholar] [CrossRef] [PubMed]
- Ogata, A.F.; Cheng, C.A.; Desjardins, M.; Senussi, Y.; Sherman, A.C.; Powell, M.; Novack, L.; Von, S.; Li, X.; Baden, L.R.; et al. Circulating Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2) Vaccine Antigen Detected in the Plasma of MRNA-1273 Vaccine Recipients. Clinical Infectious Diseases 2022, 74, 715–718. [Google Scholar] [CrossRef]
- Yonker, L.M.; Swank, Z.; Bartsch, Y.C.; Burns, M.D.; Kane, A.; Boribong, B.P.; Davis, J.P.; Loiselle, M.; Novak, T.; Senussi, Y.; et al. Circulating Spike Protein Detected in Post-COVID-19 MRNA Vaccine Myocarditis. Circulation 2023, 147, 867–876. [Google Scholar] [CrossRef] [PubMed]
- Tam, H.H.; Melo, M.B.; Kang, M.; Pelet, J.M.; Ruda, V.M.; Foley, M.H.; Hu, J.K.; Kumari, S.; Crampton, J.; Baldeon, A.D.; et al. Sustained Antigen Availability during Germinal Center Initiation Enhances Antibody Responses to Vaccination. Proc Natl Acad Sci U S A 2016, 113, E6639–E6648. [Google Scholar] [CrossRef]
- Aung, A.; Cui, A.; Maiorino, L.; Amini, A.P.; Gregory, J.R.; Bukenya, M.; Zhang, Y.; Lee, H.; Cottrell, C.A.; Morgan, D.M.; et al. Low Protease Activity in B Cell Follicles Promotes Retention of Intact Antigens after Immunization. Science (1979) 2023, 379. [Google Scholar] [CrossRef]
- Wang, C.; Hart, M.; Chui, C.; Ajuogu, A.; Brian, I.J.; de Cassan, S.C.; Borrow, P.; Draper, S.J.; Douglas, A.D. Germinal Center B Cell and T Follicular Helper Cell Responses to Viral Vector and Protein-in-Adjuvant Vaccines. The Journal of Immunology 2016, 197, 1242–1251. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Mateus, J.; Coelho, C.H.; Dan, J.M.; Moderbacher, C.R.; Gálvez, R.I.; Cortes, F.H.; Grifoni, A.; Tarke, A.; Chang, J.; et al. Humoral and Cellular Immune Memory to Four COVID-19 Vaccines. Cell 2022, 185, 2434–2451.e17. [Google Scholar] [CrossRef] [PubMed]
- Nuñez, N.G.; Schmid, J.; Power, L.; Alberti, C.; Krishnarajah, S.; Kreutmair, S.; Unger, S.; Blanco, S.; Konigheim, B.; Marín, C.; et al. High-Dimensional Analysis of 16 SARS-CoV-2 Vaccine Combinations Reveals Lymphocyte Signatures Correlating with Immunogenicity. Nat Immunol 2023, 24, 941–954. [Google Scholar] [CrossRef] [PubMed]
- Islas-Vazquez, L.; Cruz-Aguilar, M.; Velazquez-Soto, H.; Jiménez-Corona, A.; Pérez-Tapia, S.M.; Jimenez-Martinez, M.C. Effector-Memory B-Lymphocytes and Follicular Helper T-Lymphocytes as Central Players in the Immune Response in Vaccinated and Nonvaccinated Populations against SARS-CoV-2. Vaccines (Basel) 2022, 10. [Google Scholar] [CrossRef] [PubMed]
- Jandl, C.; King, C. Cytokines in the Germinal Center Niche. Antibodies 2016, 5. [Google Scholar] [CrossRef] [PubMed]
- Yegorov, S.; Kadyrova, I.; Negmetzhanov, B.; Kolesnikova, Y.; Kolesnichenko, S.; Korshukov, I.; Baiken, Y.; Matkarimov, B.; Miller, M.S.; Hortelano, G.H.; et al. Sputnik-V Reactogenicity and Immunogenicity in the Blood and Mucosa: A Prospective Cohort Study. Sci Rep 2022, 12. [Google Scholar] [CrossRef]
- Jaspe, R.C.; Loureiro, C.L.; Sulbaran, Y.; Moros, Z.C.; D’angelo, P.; Hidalgo, M.; Rodríguez, L.; Alarcón, V.; Aguilar, M.; Sánchez, D.; et al. Description of a One-Year Succession of Variants of Interest and Concern of SARS-CoV-2 in Venezuela. Viruses 2022, 14. [Google Scholar] [CrossRef] [PubMed]
- Marklund, E.; Leach, S.; Axelsson, H.; Nystrom, K.; Norder, H.; Bemark, M.; Angeletti, D.; Lundgren, A.; Nilsson, S.; Andersson, L.M.; et al. Serum-IgG Responses to SARS-CoV-2 after Mild and Severe COVID-19 Infection and Analysis of IgG Non-Responders. PLoS One 2020, 15. [Google Scholar] [CrossRef] [PubMed]
- Blanco, S.; Salomé Konigheim, B.; Diaz, A.; Spinsanti, L.; Javier Aguilar, J.; Elisa Rivarola, M.; Beranek, M.; Collino, C.; MinSalCba working group; FCM-UNC working group; et al. Evaluation of the Gam-COVID-Vac and Vaccine-Induced Neutralizing Response against SARS-CoV-2 Lineage P.1 Variant in an Argentinean Cohort. Vaccine 2022, 40, 811–818. [Google Scholar] [CrossRef]
- Watson, O.J.; Barnsley, G.; Toor, J.; Hogan, A.B.; Winskill, P.; Ghani, A.C. Global Impact of the First Year of COVID-19 Vaccination: A Mathematical Modelling Study. Lancet Infect Dis 2022, 22, 1293–1302. [Google Scholar] [CrossRef]
- Lam, I.C.H.; Zhang, R.; Man, K.K.C.; Wong, C.K.H.; Chui, C.S.L.; Lai, F.T.T.; Li, X.; Chan, E.W.Y.; Lau, C.S.; Wong, I.C.K.; et al. Persistence in Risk and Effect of COVID-19 Vaccination on Long-Term Health Consequences after SARS-CoV-2 Infection. Nat Commun 2024, 15. [Google Scholar] [CrossRef] [PubMed]
- Wilks, S.H.; Mühlemann, B.; Shen, X.; Türeli, S.; LeGresley, E.B.; Netzl, A.; Caniza, M.A.; Chacaltana-Huarcaya, J.N.; Corman, V.M.; Daniell, X.; et al. Mapping SARS-CoV-2 Antigenic Relationships and Serological Responses. Science (1979) 2023, 382. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Reactivity to S protein of SARS-CoV-2 variants after 42 dpv. Differences between the reactivities of IgG antibodies from Sputnik-V vaccinated individuals towards S (ancestral), Sα (alpha), Sβ (beta), Sγ (gamma), Sδ (delta) and Sο (omicron) were determined by indirect ELISA (n = 85). The percentage of responders is shown at the bottom of the violin plots. Significant differences were assessed with Kruskal-Wallis with Dunn's post hoc test. The y-axis represents the ratio of each sample´s signal to the to the signal of the positive control, while the x-axis represents each SARS-CoV-2 variant.
Figure 1.
Reactivity to S protein of SARS-CoV-2 variants after 42 dpv. Differences between the reactivities of IgG antibodies from Sputnik-V vaccinated individuals towards S (ancestral), Sα (alpha), Sβ (beta), Sγ (gamma), Sδ (delta) and Sο (omicron) were determined by indirect ELISA (n = 85). The percentage of responders is shown at the bottom of the violin plots. Significant differences were assessed with Kruskal-Wallis with Dunn's post hoc test. The y-axis represents the ratio of each sample´s signal to the to the signal of the positive control, while the x-axis represents each SARS-CoV-2 variant.
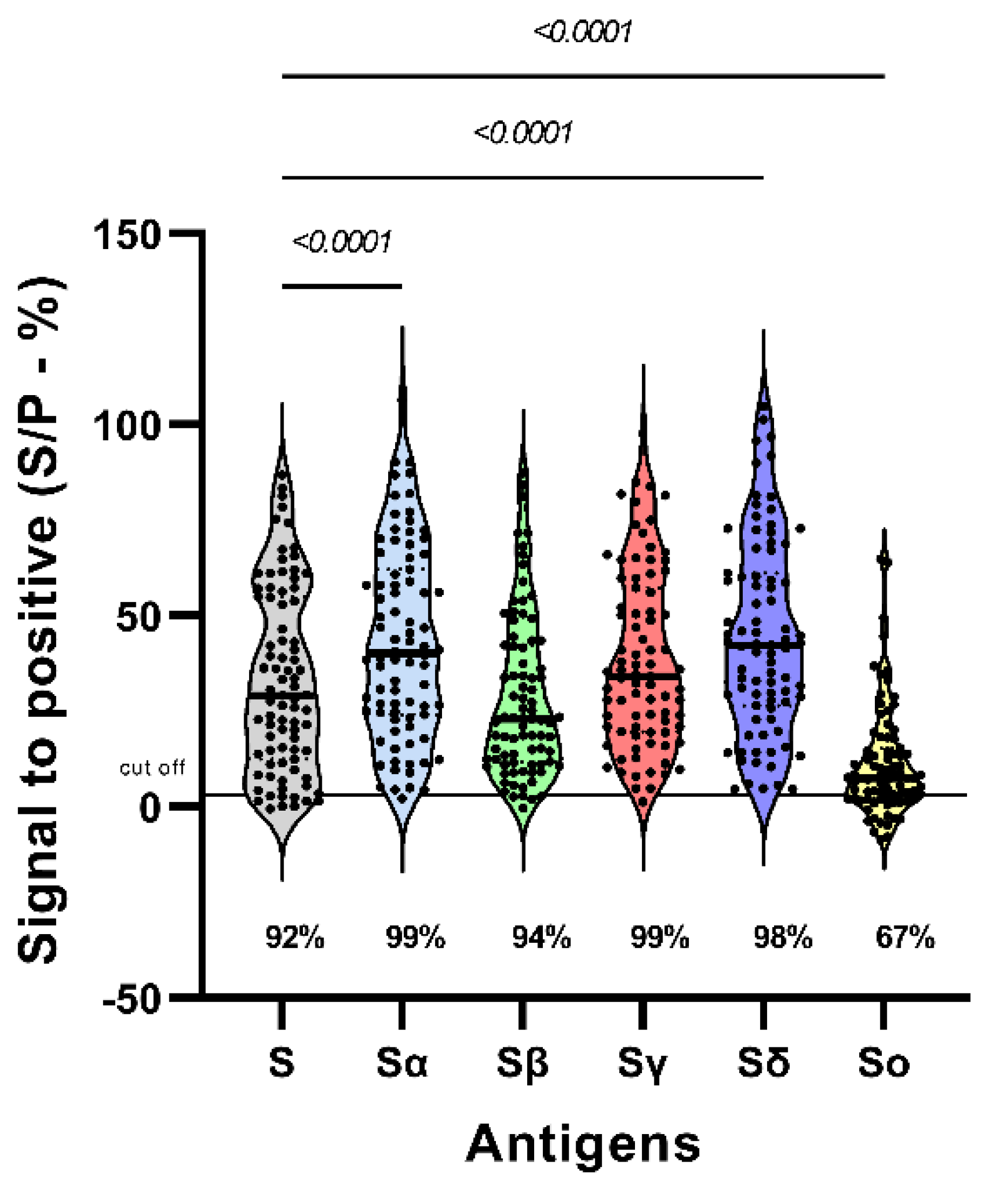
Figure 2.
Reactivity to S protein of SARS-CoV-2 variants after 42 and 180 dpv (n=18). The points on the graph represent the same set of samples tested for each antigen. Antigens with a prime symbol correspond to 180 dpv. The percentage of responders is shown below each violin plot. Wilcoxon test was performed to estimate significant differences of reactivity between both times.
Figure 2.
Reactivity to S protein of SARS-CoV-2 variants after 42 and 180 dpv (n=18). The points on the graph represent the same set of samples tested for each antigen. Antigens with a prime symbol correspond to 180 dpv. The percentage of responders is shown below each violin plot. Wilcoxon test was performed to estimate significant differences of reactivity between both times.
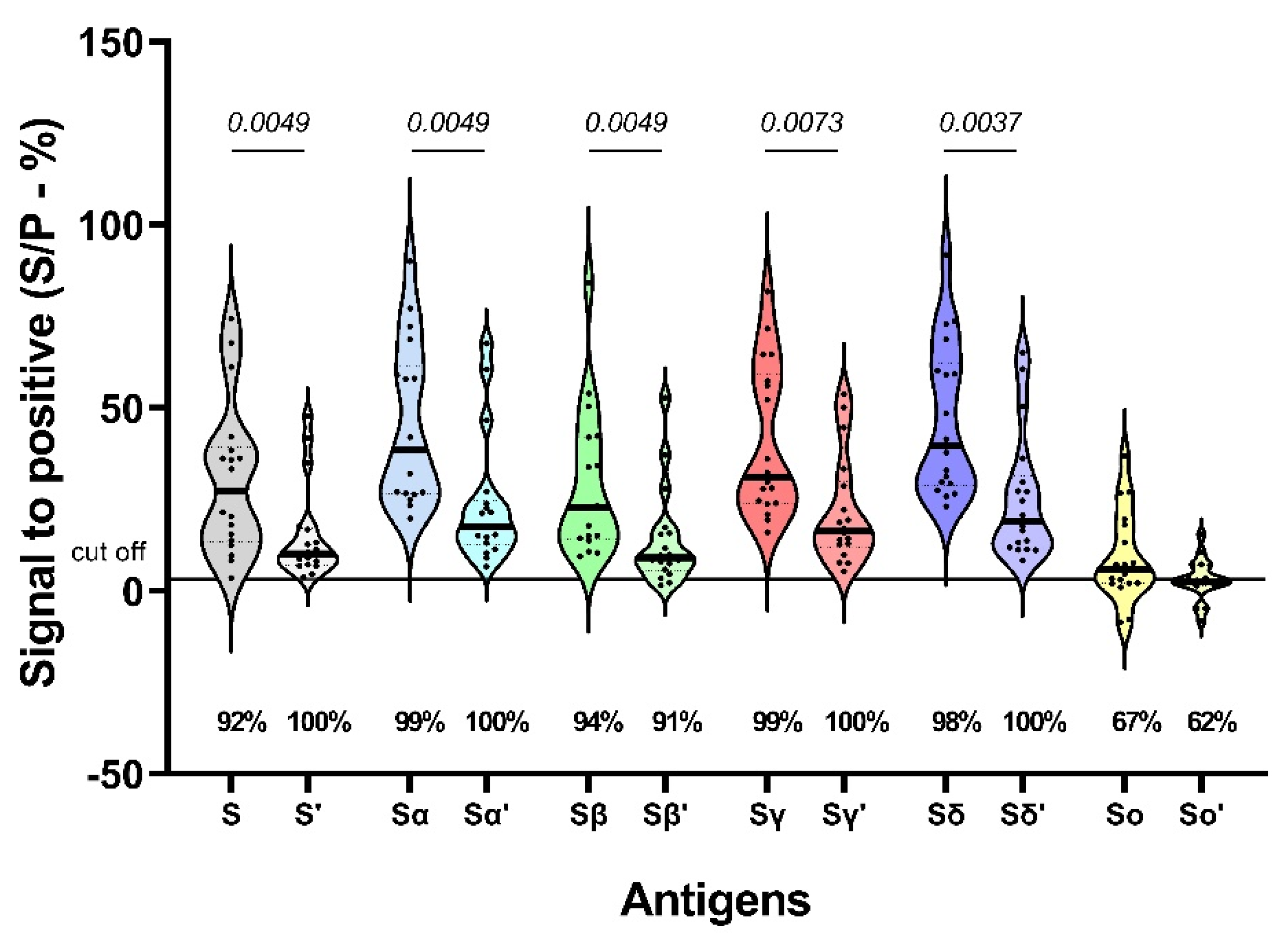
Figure 3.
Neutralization titers of sera from Sputnik V vaccinees 42 and 180 dpv against SARS-CoV-2 variants. (A) Sera with higher titers against Gamma VOC respect to the ancestral variant are highlighted in red, while those that did not show this behavior are highlighted in black (n = 18). (B) Sera with higher titers against the ancestral variant compared to the other variants (n = 10). (C) Samples that elicited a higher titer against gamma compared to the ancestral variant (n = 8). Each symbol represents an individual sample and indicates the titer required to neutralize 50% of lytic plaques, together with the standard error of the mean. Green stars indicate samples whose PRNT50 after 180 dpv was higher than 42 dpv. The numbers in brackets indicate the factor of increase or decrease in neutralization titer against each variant compared to the ancestral variant. The percentage of responders is given at the bottom of each bar. Greek letters with a prime symbol correspond to 180 dpv. Wilcoxon test was performed to estimate significant differences PRNT50 between variants and times. Although some points equal to 0 are not plotted, their value are included in the analysis.
Figure 3.
Neutralization titers of sera from Sputnik V vaccinees 42 and 180 dpv against SARS-CoV-2 variants. (A) Sera with higher titers against Gamma VOC respect to the ancestral variant are highlighted in red, while those that did not show this behavior are highlighted in black (n = 18). (B) Sera with higher titers against the ancestral variant compared to the other variants (n = 10). (C) Samples that elicited a higher titer against gamma compared to the ancestral variant (n = 8). Each symbol represents an individual sample and indicates the titer required to neutralize 50% of lytic plaques, together with the standard error of the mean. Green stars indicate samples whose PRNT50 after 180 dpv was higher than 42 dpv. The numbers in brackets indicate the factor of increase or decrease in neutralization titer against each variant compared to the ancestral variant. The percentage of responders is given at the bottom of each bar. Greek letters with a prime symbol correspond to 180 dpv. Wilcoxon test was performed to estimate significant differences PRNT50 between variants and times. Although some points equal to 0 are not plotted, their value are included in the analysis.
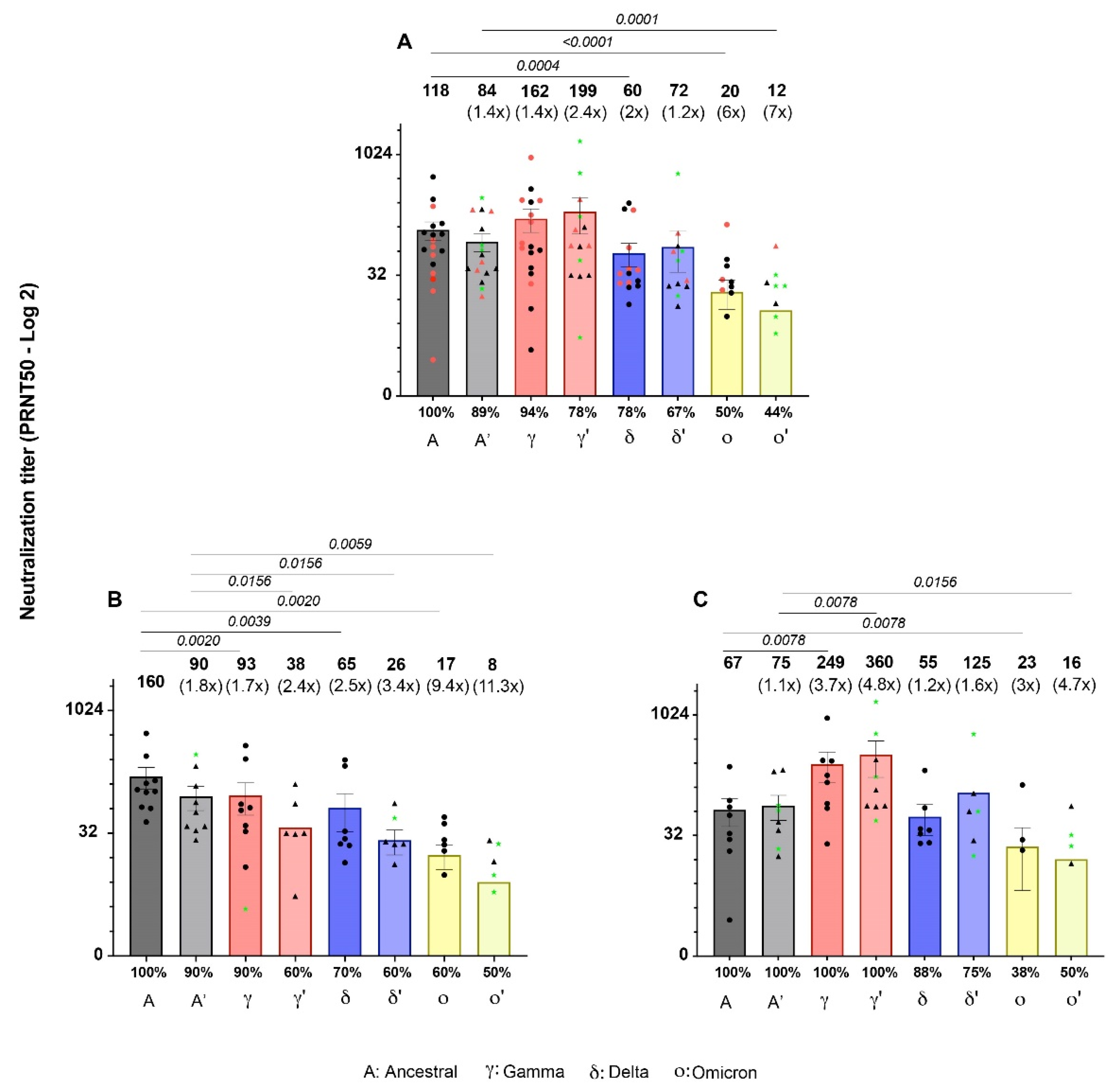
Figure 4.
Serology of low and high neutralizers of the gamma variant 42 vs 180 dpv. Differences between antibody reactivities are shown. (A) Reactivities against S protein of sera with higher neutralization titer to ancestral variant compared to gamma (n = 10). (B) Reactivities towards S protein of sera with higher titers against gamma are shown (n = 8). (C) Reactivities against RBD of higher neutralizers to ancestral variant compared to gamma (n = 10) and (D) higher neutralizers to gamma variant compared to ancestral (n = 8). Antigens with a prime symbol correspond to 180 dpv. Wilcoxon test was performed only to point out comparisons between ancestral and gamma reactivities.
Figure 4.
Serology of low and high neutralizers of the gamma variant 42 vs 180 dpv. Differences between antibody reactivities are shown. (A) Reactivities against S protein of sera with higher neutralization titer to ancestral variant compared to gamma (n = 10). (B) Reactivities towards S protein of sera with higher titers against gamma are shown (n = 8). (C) Reactivities against RBD of higher neutralizers to ancestral variant compared to gamma (n = 10) and (D) higher neutralizers to gamma variant compared to ancestral (n = 8). Antigens with a prime symbol correspond to 180 dpv. Wilcoxon test was performed only to point out comparisons between ancestral and gamma reactivities.

Figure 5.
Correlation matrix between ELISA reactivity, plaque reduction neutralization titer and age at 42 and 180 dpv. The Spearman correlation coefficient is shown in white (> 0.6) and black (< 0.6). The degree of association between these variables is represented by a heat map, where blue and red represent positive and negative correlations, respectively. All correlations are statistically significant, except for correlations with subject age. Black squares indicate correlations between reactivities and PRNT50. n = 18.
Figure 5.
Correlation matrix between ELISA reactivity, plaque reduction neutralization titer and age at 42 and 180 dpv. The Spearman correlation coefficient is shown in white (> 0.6) and black (< 0.6). The degree of association between these variables is represented by a heat map, where blue and red represent positive and negative correlations, respectively. All correlations are statistically significant, except for correlations with subject age. Black squares indicate correlations between reactivities and PRNT50. n = 18.
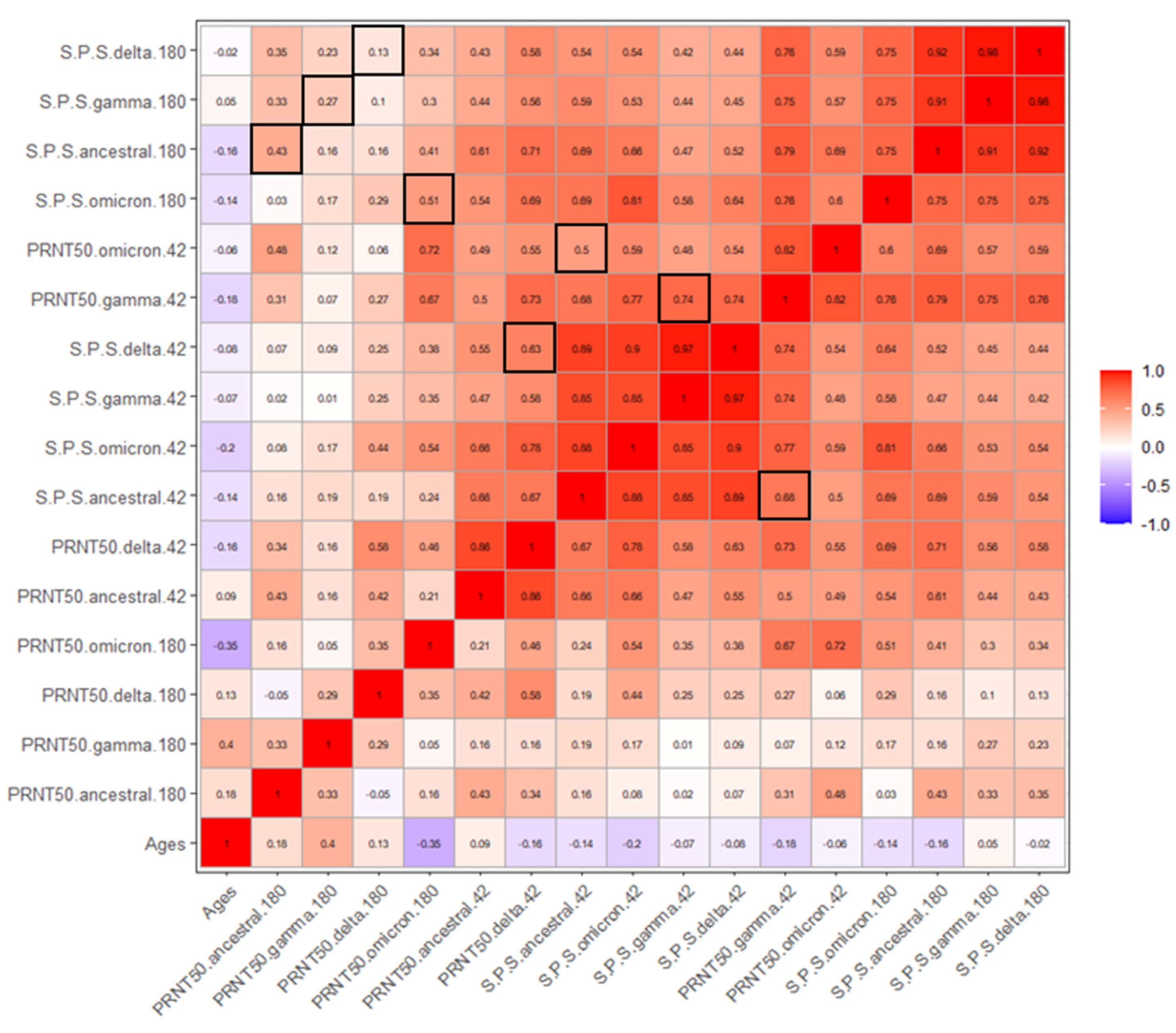
Figure 6.
Ascending Hierarchical Classification of the individuals. Two components denoting the percentage of variance explained Dim 1 (52.95%) and Dim 2 (32.87%). Dim 1 and 2 represents the two first components of all reactivities and PRNT50s evaluated. They help to explain the variance observed in the system and allow to make a classification of the samples based in their antibody response.Cluster 1 consists of low responders (n = 14). Sample 8: high responder for PRNT50 Gamma 180 dpv and PRNT50 Delta 180 dpv (descending order). Sample 12: high responder for PRNT50 Omicron 42 dpv, S/P S Beta 42 dpv, S/P S ancestral 42 dpv, S/P S Omicron 42 dpv and S/P S Delta 42 dpv (descending order). Samples 6 and 18: high responders for S/P S Omicron 180 dpv, PRNT50 ancestral 42 dpv, PRNT50 Delta 42 dpv, PRNT50 Gamma 42 dpv, S/P S ancestral 180 dpv and S/P S Beta 180 dpv (descending order).
Figure 6.
Ascending Hierarchical Classification of the individuals. Two components denoting the percentage of variance explained Dim 1 (52.95%) and Dim 2 (32.87%). Dim 1 and 2 represents the two first components of all reactivities and PRNT50s evaluated. They help to explain the variance observed in the system and allow to make a classification of the samples based in their antibody response.Cluster 1 consists of low responders (n = 14). Sample 8: high responder for PRNT50 Gamma 180 dpv and PRNT50 Delta 180 dpv (descending order). Sample 12: high responder for PRNT50 Omicron 42 dpv, S/P S Beta 42 dpv, S/P S ancestral 42 dpv, S/P S Omicron 42 dpv and S/P S Delta 42 dpv (descending order). Samples 6 and 18: high responders for S/P S Omicron 180 dpv, PRNT50 ancestral 42 dpv, PRNT50 Delta 42 dpv, PRNT50 Gamma 42 dpv, S/P S ancestral 180 dpv and S/P S Beta 180 dpv (descending order).
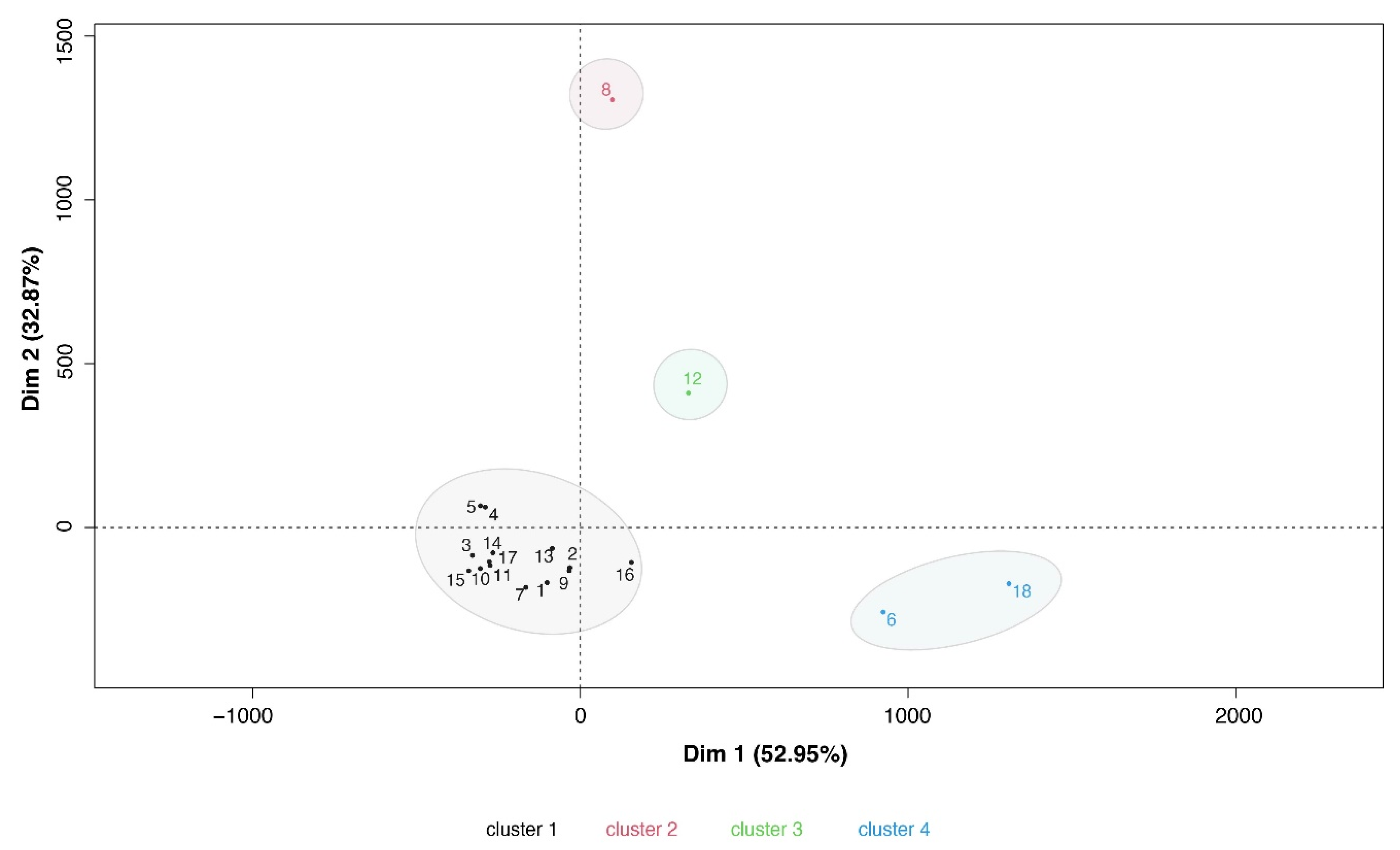

Table 1.
Number of PUBMED entries per number of countries where the main COVID-19 vaccines were approved.
Table 1.
Number of PUBMED entries per number of countries where the main COVID-19 vaccines were approved.
| Vaccine | Manufacturer | Country | PUBMED entries | Vaccine approved in countries (N) |
|---|---|---|---|---|
| BNT162b2 (Comirnaty) |
Pfizer – BioNTech | Germany | 14309 | 149 |
| ChAdOx1 nCoV-19 (AZD1222) | Astra Zeneca – Oxford University | Sweden | 5911 | 149 |
| Ad26.COV 2-S (Jcovden) | Janssen Biotech, Inc (Johnson & Johnson) | Belgium | 4088 | 113 |
| mRNA-1273 (Spikevax) | Moderna | USA | 4824 | 88 |
| CoronaVac | Sinovac | China | 1223 | 56 |
| BBIBP-CorV (Covilo) |
Sinopharm | China | 725 | 93 |
| NVX-CoV2373 (Nuvaxovid) | Novavax | USA | 989 | 40 |
| Gam-COVID-Vac (Sputnik V) | Gamaleya | Russia | 563 | 74 |
1A search query was performed in PubMed using the following words and operators: ((((COVID-19) OR (SARS-CoV-2)) AND (“name of the vaccine”)) OR (“name of the manufacturer”)) AND (vaccine). 2The number of countries was assessed as reported by Kudlay and Svistunov, and the COVID19 vaccine tracker team [16,17].
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



