
Submitted:
02 July 2024
Posted:
03 July 2024
You are already at the latest version
Abstract
Recent advancements in mass spectrometry have significantly enhanced our understanding of complex lipid profiles, opening new avenues for oncological diagnostics. This review highlights the importance of lipidomics in the comprehension of certain metabolic pathways and its potential for the detection and characterization of various cancers, in particular melanoma. Through detailed case studies, we demonstrate how lipidomic analysis has led to significant breakthroughs in the identification and understanding of cancer types and its potential for detecting unique biomarkers that are instrumental in its diagnosis. Additionally, the review addresses the technical challenges and future perspectives of these methodologies, including their potential expansion and refinement for clinical applications. The discussion underscores the critical role of lipidomic profiling in advancing cancer diagnostics, proposing a new paradigm in how we approach this devastating disease, with particular emphasis on its application in comparative oncology.
Keywords:
melanoma
; comparative oncology
; liquid biopsy
; mass spectrometry
; lipidomics
1. Introduction
Cancer is one of the major causes of demise in developed countries, only second to cardiovascular disease [1]. The types of malignancies most prevalent vary with country and year. However, breast cancer and melanoma are frequently found among the ones showing greatest incidence worldwide [2,3,4]. Furthermore, in the USA, it is predicted that they will become the major types by the year 2040 [5]. This roughly coincides with the prevalence of spontaneous malignancies in other mammals, such cats and dogs. Nevertheless, in the former species, lymphoma is the most common, while in the latter case, mastocytoma, lymphomas and hemangiosarcoma show very similar numbers to melanoma [6,7,8,9].
Early, accurate diagnosis is paramount to success in any medical intervention and so much so in cancer. In the case of Non-White individuals, delayed diagnosis of melanoma is more common than in Caucasians and associated with worse survival rates [10]; a similar situation may occur in the case of vulvar melanoma in women [11]. On the other hand, misdiagnose is also a source for intervention errors. Difficulties are known to differentiate melanoma from certain types of benign nevi on the basis of usual histopathological criteria [12]. Furthermore, the choice of solid biopsy method used, either shave, punch, or incisional, may alter the estimation of the depth of tumor invasion, and change the recommendations for surgical management in up to 18% of the cases [13].
The most common methods for detection and characterization of malignant lesions rely on surgical removal of a tissue biopsy and the use of immunohistochemical techniques. Thus, in general, a suspicious lesion can be broadly separated into benign and malignant, characterized initially as sarcoma or carcinoma and even further subtyped identifying a panel of different, complementary antigens [14,15]. While this is broadly the standard procedure, in the case of some tumor types, among them melanoma, variations occur and, thus, distinction between benign and malignant nevi does not follow the staining profile of any specific immunohistochemical marker, although Ki-67, pHH3 and p16 are proposed and sometimes used in that respect [12]. In addition to histopathology, imaging techniques based on radiology, ultrasound or magnetic resonance, among others, are also widely used, often prior to a biopsy [15]. Radiological detection of mammary cancer, for example, is long known for its convenience in routine checks, although this technique is cumbersome, potentially dangerous and may induce non-reattendance [16].
There are several aspects to take into account when judging the utility of a diagnostic technique. The ability to distinguish between malignant and normal tissue or benign lesions is first to mind. The accuracy, sensitivity and specificity of immunohistochemical techniques is difficult to quantify. Nevertheless, some studies have shown that pathologists with >10 years of experience confronted with the identification of putative melanomas were accurate in 80% of the cases presented to them, while the sensitivity was estimated to be 91%. However, it was also shown that those figures were heavily influenced by training, dropping to only 62% and 56%, respectively, in the case of less trained personnel, albeit they already had 1-2 years of experience [17]. Imaging procedures also show variable figures depending on the actual technique. In a typical study, sensitivity and specificity for human visually analyzed mammograms were found to be around 73.0% and 80.0%, respectively, while ultrasound figures were 100%, 80.4% respectively [18]. The tendency towards the introduction of artificial intelligence is strong in this field, but, to the best of our knowledge, sensitivity and specificity figures for machine-guided diagnostic are still just similar to those obtained by trained human radiologists to date [19].
In addition to reliability parameters, other aspects are important to assess the utility of a technology, like ease of acquisition, secondary effects and risks, cost-effectiveness or patient suffering, among others. This is obvious in the case of difficult-to-access tumors, like gliomas, but are also important in less evident others, like breast cancer. As stated above, discomfort associated to mammography tests may induce non-reattendance [16]. In the case of melanoma there is room for improvement in this area too. For example, partial biopsies are frequent and may lead to underestimation of Breslow thickness, while similarity to nevi can make testing based on solid biopsies challenging in individuals with a large number of suspicious lesions [20]. An alternative to this is the use of liquid biopsies. Urine and blood are easy to obtain using non or minimally invasive procedures and, in the case of blood, it is a tissue that is in intimate contact with nearly all others. This last characteristic makes blood plasma to transport a variety of molecules leaked or released from diseased organs, or other tissues, as a response to a health imbalance. It is the correlation in the changes in concentrations of these molecules, with respect to the levels found in healthy organism, what makes them useful biomarkers [21].
Cancer cells deviate from normal ones in many aspects, and these differences are vital to maintain a neoplastic phenotype. Among the hallmarks that have been identified as common in all tumors are important variations in the metabolism [22]. While the Warburg effect, i.e. lactic fermentation-based energy metabolism in the presence of normal tissue oxygen pressure, and enhanced glutaminolysis are long known to be part of these metabolic alterations [23,24], other, less apparent changes accompany them, such as those in amino acid and lipid metabolism [25].
Lipids are a heterogeneous category of molecules characterized by their hydrophobicity. Major groups of lipids contribute to membrane structure and function, such as phospholipids, sphingolipids and sterols, while others serve as energy storage, vitamins, enzyme co-factors, digestive surfactants or hormones. In human blood plasma, lipids are abundant, with total lipid levels above 500 mg/l [26,27]. Similar figures are found in canine plasma [28]. Major lipid types, in rough order of concentration, are phospholipids, cholesterol (either free or fatty acid esterified) and triglycerides [26,27,28]. Nevertheless, levels and species of several of these lipids may fluctuate according to the diet and time of day. For example, odd-chain fatty acid containing phospholipids in plasma may be found in association with a dairy meal [29]. Being hydrophobic molecules, lipids are not found in significant concentrations in solution in plasma, rather, they are present in colloidal form. In order to form colloids, they need to associate to specific lipid transport systems (LDL, HDL, chylomicrons) or form microvesicles. Variations exist in lipid profiles between the different types of lipoproteins, as a reflection of physiology. Thus, phospholipids are more abundant in HDL than in VLDL, while the reverse is true for triglycerides [27], in accordance with those lipoproteins belonging to two different lipid transport systems [30].
Tissue cells, either normal or tumoral, release microvesicles into the extracellular medium and, eventually, they reach the blood plasma. These vesicles are known as exosomes or extracellular vesicles (EV). They can be described as small, single-membrane, secreted organelles with diameters between 30 and 200 nm [31]. The subcellular origin of these microvesicles can be either the plasma membrane or, most often, endosomal structures, particularly the multivesicular body [31,32]. In the case of tumor-derived exosomes, these are known to be quite different from other exosomes and part of an intercellular crosstalk with multiple consequences. For example, they may induce changes in healthy tissue favoring metastatic colonization [33,34]. Being biomembrane-based secreted vesicular structures, they carry proteins, nucleic acids and several other molecules that can be used as cancer biomarkers [35]. Naturally, they can also be a source of lipids that can contribute to lipid biomarker signatures in plasma.
Finally, it is important to have in mind the consequences of the changes in lipid metabolism in cancer. For example, a constant creation of new membranes and cells, together with the increasing need to maintain activated signaling routes that rely on lipid rafts, make tumor cells avid for cholesterol. Uptake of this lipid through LDL receptor overexpression and increased internalization, leads, on the one hand, to a greater flux of LDL along the tumor cell endocytic pathway [36]. On the other hand, it leads to blood hypocholesterolemia, as observed in various types of cancer including hematopoietic, bowel, lung, prostate and head and neck cancers [37]. This situation is by no means particular for cholesterol, rather, it is common for many types of lipids, with natural variations among them [38]. Accordingly, plasma lipids are considered as a cancer specific biomarker source with diagnostic value for some time already [39].
2. Interplay Between Lipid Metabolism and Mitochondrial Function in Cancer
The metabolic signature of cancer cells encompasses alterations in glycolysis, mitochondrial respiration, and lipid and amino acid metabolism. Mitochondria play a crucial role in bioenergetic regulation, metabolism, and apoptosis [40,41]. Moreover, they are interconnected with lipid metabolism pathways, suggesting their involvement in metastatic progression and cancer phenotype, such as the carnitine shuttle system, lipoic acid biosynthesis, and sterol modifications (cholesterol) [42].
The lipid composition of mitochondrial membranes (MM) primarily comprises glycerophospholipids at 75-95%, of which 80% are phosphatidylethanolamines (PE) and phosphatidylcholines (PC), and 10-15% are cardiolipins (CL) [43,44]. CL accounts for 20% of the lipid mass in the inner mitochondrial membrane (IMM) [45,46], being essential for the electron transport chain and oxidative phosphorylation (OXPHOS), indispensable processes for bioenergetics [47]. Abnormal CL metabolism is associated with various pathologies, including cancer, affecting the structural and functional stability of mitochondria. Abnormal levels of CL interfere with mitochondrial metabolism by altering binding sites in OXPHOS complexes I, III, and IV, necessary for the stability of supercomplexes and the synthesis of acetyl-CoA, which is essential for the Krebs cycle, involved in metastasis and cell migration [46,48]. Additionally, CL participates in mitochondrial quality control through mitophagy. Mitochondrial dysfunction, resulting from increased oxidative stress due to OXPHOS, leads to cellular protection mechanisms such as apoptosis. In turn, increased OXPHOS leads to lipid oxidation, with the translocation of oxidized CL from the IMM to the outer mitochondrial membrane (OMM), where it is recognized by the mitophagy machinery [44,46,48]. This acts as a survival mechanism, promoting the elimination of damaged mitochondria and preventing apoptosis (Figure 1a).
Cholesterol, present in all cellular membranes and associated with glycosphingolipids in lipid rafts, constitutes approximately 3% of mitochondrial membranes, mainly in the IMM [49,50]. In cancer cells, there is an accumulation of mitochondrial cholesterol, correlated with tumor growth and malignancy [51]. This alters the fluidity of MM, affecting transmembrane proteins and increasing the production of reactive oxygen species (ROS) [49,51]. As a result, oxidative phosphorylation is promoted, changing energy metabolism and contributing to chemotherapy resistance by altering mitochondrial apoptosis [50] (Figure 1b).
Carnitine is a key cofactor in metabolism, regulating the acyl-CoA/CoA balance, modulating lipid biosynthesis and degradation, and gene expression. It participates in the synthesis of trimethylamine-N-oxide (TMAO), inflammatory processes, and fatty acid oxidation (FAO) in the mitochondria [52,53,54]. Carnitine facilitates the transfer of acyl groups across mitochondrial membranes for FAO and ATP production. In cancer, alterations in the expression or activity of carnitine transport are observed, especially in neoplasms with dysregulated fatty acid utilization [53]. Under hypoxia, cancer cells obtain energy from fatty acid oxidation, demonstrating carnitine's involvement in metabolic plasticity [53]. Furthermore, the rate-limiting enzyme in fatty acid oxidation (FAO) in the metabolic adaptation of cancer is carnitine palmitoyltransferase I (CPTI), overexpressed in cancer cells, promoting FAO and adaptation to metabolic stress [52,53]. This increases NADPH production, facilitating acetyl-CoA generation and providing redox power, counteracting oxidative stress, and promoting cell survival [53,54,55] (Figure 1c).
Lipoic acid (LA) is a mitochondrial cofactor with therapeutic effects, inhibiting tumor proliferation, migration, and invasion, acting as a metal chelator and antioxidant [42,56]. However, studies show that in colon cancer cell lines, LA administration increases OXPHOS, acting as a pro-oxidant and triggering apoptosis by increasing mitochondrial ROS in cancer cells [57,58]. Besides promoting apoptosis, LA modulates mitochondrial metabolism and circulating lipid levels, activating the MAPK signaling pathway and inhibiting mTOR, hindering cell proliferation [42,58] (Figure 1d).
To sum up, lipid metabolism plays a fundamental role in the regulation of mitochondrial function within the context of cancer. Alterations in the lipid composition of mitochondrial membranes, the handling of lipids such as cholesterol and phospholipids, and the modulation of key metabolic pathways such as fatty acid oxidation directly influence mitochondrial bioenergetics and cell signaling. These interactions not only affect the survival and proliferation of cancer cells but also their ability to resist treatments and adapt to adverse environments.
3. Lipid Profile in Melanoma
Lipids play crucial roles in cell biology, functioning as structural components of membranes, signaling molecules, and energy reservoirs. Among the various lipid classes, sphingolipids and glycerolipids are particularly significant in the development and progression of melanoma.
The sphingolipid family is involved in melanoma cell adhesion, metabolic plasticity, and aggressiveness [59]. Levels of S-1-P produced by downregulated acid ceramidase induce the loss of E-cadherin in melanoma cells, along with an increase in gangliosides, promoting epithelial-mesenchymal transition (EMT) processes and adhesion to the extracellular matrix, characterizing the aggressive phenotype of melanoma [59,60,61].
In melanoma cells, low activity of enzymes responsible for ceramide production has been reported, favoring the consumption of ceramides towards the production of sphingosine-1-phosphate and gangliosides [59,62,63]. In stage II patients and proliferative melanoma cells, increased hydrolysis of ceramide to sphingosine-1-phosphate has been observed [63]. Additionally, a poorer prognosis has been correlated with the downregulation of sphingomyelin synthase 1, which catalyzes the transformation of ceramide to sphingomyelin, in patients with metastasis. Lipid profiling of melanocytes, primary melanoma cells, and metastatic melanoma using mass spectrometry has reported a drastic reduction of SM species in melanoma cells compared to melanocytes [64].
Phospholipids encompass various lipid classes derived from phosphatidic acid (PA): phosphatidylethanolamine (PE), phosphatidylcholine (PC), phosphatidylinositol and its phosphorylated derivatives (PI, PIP), phosphatidylserine (PS), and phosphatidylglycerol (PG). Abnormalities in glycerophospholipid metabolism leading to increased levels of PC and PA favor cell survival by inhibiting apoptosis and inducing EMT and cellular invasion processes. In primary melanoma tumor tissue samples, a significant increase in total PC and PG was revealed compared to nevus melanocytes [65]. Furthermore, mass spectrometry analysis determined that PC, PG, PA, and PI species show increased signals in primary and metastatic melanoma cells [64].
4. Canine Models in Comparative Melanoma Research
The current challenge is to develop molecular tools and relevant models that enable early diagnosis of breast cancer. Traditional in vitro preclinical models, such as tumor cell lines, offer highly controlled conditions with reproducible results, facilitating validation, repetition, and optimization of experiments. In vivo models, including murine xenograft and transgenic models, are valuable for studying the complexity of human cancer, providing insights into cellular processes in neoplasms. These models offer advantages in sample management due to their high reproducibility in captivity, short life span, and certain similarities to humans. Specifically, the xenograft model allows for the study of human cell lines in a living system, resulting in tumors composed of human cancer cells and murine stromal cells. However, both preclinical models have limitations, such as the difficulty in replicating interactions with the microenvironment, the absence of a metastatic pattern, and the inability to fully integrate the host's immune response.
For the study of melanoma in murine models, key considerations include tumor heterogeneity in humans, replication difficulties in the laboratory, and structural dissimilarities of the skin and differential melanocyte locations between species (epidermis in humans and dermis in mice) [66,67]. Additionally, the low efficacy for pharmacological response studies in phase III is well-known [66]. Therefore, spontaneous tumor models are of great interest for studying the pharmacokinetics of innovative therapies in vivo, their effects on the tumor and the patient, and the interactions between tumor cells and the microenvironment [66,68,69].
Canine melanoma accounts for 7% of neoplasms [66,70], 14-45% of all oral neoplasms [66], and 9-20% of cutaneous tumors [66]. The most common locations include the mouth (62%), skin (27%), digits (6%), and eyes (1%), among others [71]. There is a predisposition in males for developing oral melanomas [72]. The following sections focus on the similarities between species in cutaneous and mucosal (oral) melanomas as comparative oncology models, given that each is more frequent and among the most aggressive in humans, respectively.
The primary risk factor for cutaneous melanomas in humans is UV radiation, with a higher risk in individuals with fair skin, or more specifically, those with very low melanin concentrations [73]. Contrary to expectations, the risk of cutaneous melanoma in dogs increases in breeds with dark skin tones (e.g., Schnauzers, Scottish Terriers, Poodles) [66,73]. Additionally, UV radiation is not a risk factor of the same magnitude as in humans, as fur acts as a protective factor [73].
In contrast to cutaneous melanoma, mucosal melanoma is one of the most common neoplasms in dogs, not induced by UV radiation. The most common locations are the oral cavity, lips, tongue, palate, anal region, and intranasal area [74]. Around 30% of melanoma cases in dogs and 50% of mucosal melanomas in humans are amelanotic, making diagnosis challenging due to the lack of melanin [75]. Both exhibit high aggressiveness and metastatic potential, primarily affecting lymph nodes and lungs [66,73,74]. This type of melanoma also has the worst prognosis in dogs, with a median survival of 3 months to 2 years, depending on the stage and treatment choice [66,73,74]. Another common feature in both species is resistance to chemotherapy and radiation. Clinical trials in dogs have shown that using human genes (human gp100) resulted in tumor regression in 18% of cases and increased survival to nearly a year [76].
In both species, a common library of single-nucleotide polymorphisms (SNPs) has been detected, with fewer than five mutations per megabase, reflecting UV-independent molecular events. Loss of heterozygosity in TP53, amplification in MDM2, mutations in c-KIT, NRAS, and disruptions in PTEN have been found in oral melanoma cases in both humans and dogs, highlighting the involvement of the MAPK and PI3K/Akt pathways in tumor formation and progression [74,77]. Additionally, histopathological similarities include pleomorphic cytomorphologies occurring in both species, such as epithelioid (polygonal) malignant melanocytes, spindloid malignant melanocytes, and small round blue cell malignant melanocytes [78].
Currently, the prognostic method for oral melanoma in dogs, according to the WHO, is based on tumor size and the presence of distal or regional metastasis. Furthermore, as in humans, prognostic parameters include nuclear atypia, mitotic count, presence of lymphatic invasion, and the Ki-67 index [74,79,80,81]. Therefore, there are no validated biomarkers capable of providing a reliable prognosis.
The relevance of using domestic animals for comparative oncology studies has increased over the past decade, primarily due to the rise of the One Health movement, but also because of the advantages they present over conventional in vivo models [67,82]. In 2005, the dog genome was sequenced, marking the beginning of comparative studies of various diseases [83,84]. It has been demonstrated that the canine genome has high homology with the human genome, indicating that the evolutionary lines of dogs and humans are more similar in terms of nucleotide divergence and reorganization than those of humans and rodents [83]. However, despite the genomic similarities, there are specific mutations present in human melanoma that are absent in canine melanoma, and vice versa. Furthermore, there is a lack of studies on canine lipidomics in melanoma. Given the critical role of lipid metabolism in cancer progression, invasion, and metastasis, this field warrants more extensive investigation.
5. Lipidomics and Machine Learning
In chemical terms, lipids are usually divided in classes, like phosphatidylcholines (PC) or sterol esters. For many purposes, like understanding overall biomembrane structure, this level of detail is enough. However, each variation in the molecule resulting from inclusion of a different fatty acid or an ether bond, for example, results in a different species that is chemically and physiologically different, with some species being more abundant than others. For instance, the most abundant phospholipid species in the plasma membrane tends to be 1-stearoyl-2-oleoyl-sn-phosphatidylcholine (abbreviated PC 18:0/18:1, or often simply, PC 36:1). However, its analogous plasmalogen, 1-O-1'-(Z)-octadecenyl-2-oleoyl-sn-glycero-3-phosphocholine (PC 36:1; O), is less abundant and shows patently different membrane behaviors [85]. Untargeted lipidomics is the individual identification and quantitation of the whole set of lipid species in a sample. This is usually attained using direct mass spectrometry approaches, such as MALDI-TOF or ESI [65,86,87] (Figure 2). Alternatively, lipid-class separation techniques, such as UHPLC, may be associated to MS to improve identification of isobaric species [64,88,89].
Needless to say, the vast number of lipid species that can be identified from complex samples, such as blood plasma, require solid, multivariate statistical processing to reach meaningful conclusions. It is out of the scope of this review to describe the plethora of different mathematical approaches at the hands of lipidomics researchers. Nevertheless, we will try to provide some simple descriptions when possible. The reader is referred to more authoritative works published elsewhere for both multivariate statistics and machine learning algorithms [90,91,92,93]. The reader is also advised that there exists a plethora of method modifications that can increase the difficulties for a precise identification of the actual methods used.
The present relatively easy use of computer-aided multivariate statistics makes the analysis of large sets of data amenable and, as a consequence, attractive. Not surprisingly, the first analysis with data obtained from lipidomic studies is usually done utilizing dimension reduction methods, such as Principal Component Analysis (PCA), Partial Least Squares (PLS), Maximum Margin Criterion (MMC), or their derivatives. This simplifies the analysis of a large number of variables (the levels of individual lipid species) obtained from a single experimental case and permits clustering by graphical means. In many studies, this is enough to show useful differences in lipid composition between healthy and pathological tissues. For example, in a hepatocellular cancer carcinoma, the exploratory analysis was done using PCA, but later, PLS-DA (Partial Least Square Discriminant Analysis, a variant based on PLS) and SIMCA (Soft Independent Modelling of Class Analogy) clustering models were constructed [94]. Similarly, OPLS-DA (Orthogonal PLS-DA) was used to successfully cluster healthy individual plasmas from that of patients suffering from pancreatic [95] or papillary thyroid cancer [96]. PLS also helped in the discovery of possible lipid biomarkers in melanoma using cell lines [97] and separated grades in meningioma biopsies [98]. In dogs, OPLS was found superior to PCA to distinguish mammary tumors [99]. Likewise, OPLS and PCA were used successfully in a more recent study to create models discriminating nevi from primary and metastatic human melanoma [64].
However, it has been the recent development of algorithms capable of deciding autonomously the tests and parameters to be used in mathematical modelling (machine learning, ML) that set a different ball in motion. A pubmed search using “machine learning”, “diagnosis” and “cancer” as keywords for papers published since January 2020 offered 8927 results in June 2024, but only 2821 in the previous twenty years (2000-2020). ML algorithms used in lipidomics are nearly exclusively supervised and with the aim of clustering. Supervised algorithms need the data sets to be divided into training and validating groups. The first group is fed to the chosen ML algorithm to obtain the parameters necessary to construct a second algorithm, a mathematical model, able to classify individual cases into different groups on the basis of their associated data. The second data group measures how successful the constructed algorithm is in its predictions when facing an unknown-type case. A list with several lipidomics studies published between 2021 and 2024, using different ML algorithms can be found in Table 1.
ML algorithms may be separated into linear and non-linear types. Linear algorithms assume at some stage, that the variation relationship between variables is due to a constant factor. Non-linear ones do not make that assumption. Linear algorithms, like Multiple Linear Regression (MLR) or Logistic Regression (LR) are mathematically simpler. LR is a dichotomic popular algorithm and has been successfully used when classifying gastric lesions using plasma biopsies [108,109] (Table 1). In the case of the 2021 study, the figures for specificity (true negative rate) and sensitivity (true positive rate) obtained were 93.8% and 95.0%, respectively. Moreover, the Receiver Operating Characteristic Area Under the Curve (ROC AUC, or diagnostic accuracy) reached a remarkable 94.4% [108]. Similar figures were observed for several of the models built using LR in the 2022 study too [109]. Other linear approaches have been successfully applied to renal carcinoma [103] and mammary cancer [89]. In this last case, solid biopsies were used to train ML Support Vector Machine (SVM) and LR algorithms with similar results; values for ROC AUC above 95% were obtained. It must be noted that SVM algorithms can be used under linear and non-linear assumptions, but the actual strategy is seldom stated in the manuscripts. Another case of successful use of SVM comes from a study on menangioma [100]. The authors were able to separate three different types of lesions with a diagnostic accuracy of 87%. Other non-linear algorithms are also widely used. The most common ones include k-Nearest Neighbor (KNN) and Naïve Bayes (NB), among others (e.g. [65,86,88,100,105]). In any case, it is difficult to predict which ML algorithm will best cluster the cases available from a study. For that reason, and due to their ease of implementation, it is usual to test several algorithms and opt for the best performing one. In several cases, the data provided similar results using a panel of ML algorithms, like in the cases of small cell lung cancer [101], and a model of ovarian cancer [88]. Although non-linear algorithms can be considered more general, they do not necessarily fit better in all cases. For example, in melanoma solid biopsies the linear ML model constructed using LR was found superior to non-linear models constructed using NB and KNN, achieving the former a perfect classification of the samples [65].
6. Perspectives
There are some aspects that open before cancer lipidomics. Some are known for some time, like the necessity for normalization and quality control of lipidomic data to ease interpretation [110]. In this respect, several efforts have been made and the lipidomics community is close to a consensus [111,112,113,114,115,116]. Related to this, shotgun mass spectrometry techniques are widely used for the identification of pathogens in clinical environments, both as an in-house analytical service or as a contract one [117]. Therefore, if shotgun lipidomic approaches are standardized, an opportunity exists to reach the same level of industry development as other omics, like genomics or proteomics. As an example of the existing gap, a search in LinkedIn in June 2024 showed 4400 companies using genomics in their description, but only three mentioned lipidomics.
Up to this point, most reports have made use of machine learning algorithms. The inclusion of more advanced artificial intelligence algorithms, like deep learning, is possibly the next step in this area [118,119,120]. Actually, some attempts have already been made in cancer, albeit not in lipidomics [121,122]. In any case, the present state of the art makes easy the discrimination of certain types of cancer from healthy individuals in a dichotomic way. This may suffice in the case of imaging lipidomics, like the already mentioned example in breast cancer [86], since, in these cases, the suspected cancer type is known and healthy tissue from the same individual may be available for a comparison. Possibly, a closer to implementation example for this is melanoma, since biopsies from lesions are easy to obtain, but their analysis still suffer from lack of a consensus antigenic marker [12] that can be filled by lipidomics imaging [65]. However, in more ambitious endeavors, like making use of liquid biopsies for oncological testing, it will be necessary to implement lipidomic strategies able to distinguish between several types of cancer and healthy individuals at the same time. To the best of the authors’ knowledge, no work has dealt with this issue yet and the question remains on whether lipidomics will be able to distinguish beyond healthy and oncological patients. Be that as it may, proposals exist for the combined use of multi-omics as a plausible solution to the limitations of single approach strategies [123]. On the other hand, even if limited to a dichotomic answer, an opportunity is clear for liquid biopsy-based lipidomics as a follow up technique, making post-intervention control tests easier, faster and less invasive.
Finally, a natural tendency is seen to employ the power of lipidomics to gain insight into mechanistical aspects in cancer. For example, alterations in metabolism related to prostate cancer were identified [124], and the actual influence of PIK3CA mutations on lipid metabolism clarified [125] by lipidomic approaches. Nonetheless, more work and efforts are needed to exploit the full potential of lipidomics in cancer research.
In conclusion, lipidomics, together with machine learning/artificial intelligence approaches, are a promising tool in cancer research and diagnostics. Breakthroughs are expected to occur in the immediate future.
References
- Curtin, S.C.; Tejada-Vera, B.; Bastian, B.A. Deaths: Leading Causes for 2021. National Vital Statistics Reports 2024, 73, 1–116. [Google Scholar]
- Zheng, R.; Zeng, H.; Zhang, S.; Chen, T.; Chen, W. National Estimates of Cancer Prevalence in China, 2011. Cancer Lett 2016, 370, 33–38. [Google Scholar] [CrossRef]
- Yao, C.; Billette, J.M. Short-Term Cancer Prevalence in Canada, 2018. Health Rep 2022, 33, 15–21. [Google Scholar] [CrossRef]
- De Angelis, R.; Demuru, E.; Baili, P.; Troussard, X.; Katalinic, A.; Chirlaque Lopez, M.D.; Innos, K.; Santaquilani, M.; Blum, M.; Ventura, L.; et al. Complete Cancer Prevalence in Europe in 2020 by Disease Duration and Country (EUROCARE-6): A Population-Based Study. Lancet Oncol 2024, 25, 293–307. [Google Scholar] [CrossRef]
- Rahib, L.; Wehner, M.R.; Matrisian, L.M.; Nead, K.T. Estimated Projection of US Cancer Incidence and Death to 2040. JAMA Netw Open 2021, 4. [Google Scholar] [CrossRef]
- Sleeckx, N.; de Rooster, H.; Veldhuis Kroeze, E.J.B.; van Ginneken, C.; van Brantegem, L. Canine Mammary Tumours, an Overview. Reprod Domest Anim 2011, 46, 1112–1131. [Google Scholar] [CrossRef]
- Misdorp, W.; Misdorp1, W. Veterinary Cancer Epidemiology. Veterinary Quarterly 1996, 18, 32–36. [Google Scholar] [CrossRef]
- Blackwood, L. Cats with Cancer: Where to Start. J Feline Med Surg 2013, 15, 366–377. [Google Scholar] [CrossRef]
- Grüntzig, K.; Graf, R.; Hässig, M.; Welle, M.; Meier, D.; Lott, G.; Erni, D.; Schenker, N.S.; Guscetti, F.; Boo, G.; et al. The Swiss Canine Cancer Registry: A Retrospective Study on the Occurrence of Tumours in Dogs in Switzerland from 1955 to 2008. J Comp Pathol 2015, 152, 161–171. [Google Scholar] [CrossRef]
- Brunsgaard, E.K.; Wu, Y.P.; Grossman, D. Melanoma in Skin of Color: Part I. Epidemiology and Clinical Presentation. J Am Acad Dermatol 2023, 89, 445–456. [Google Scholar] [CrossRef] [PubMed]
- Wohlmuth, C.; Wohlmuth-Wieser, I. Vulvar Melanoma: Molecular Characteristics, Diagnosis, Surgical Management, and Medical Treatment. Am J Clin Dermatol 2021, 22, 639–651. [Google Scholar] [CrossRef] [PubMed]
- Abbas, O.; Miller, D.D.; Bhawan, J. Cutaneous Malignant Melanoma: Update on Diagnostic and Prognostic Biomarkers. Am J Dermatopathol 2014, 36, 363–379. [Google Scholar] [CrossRef] [PubMed]
- Pavri, S.N.; Clune, J.; Ariyan, S.; Narayan, D. Malignant Melanoma: Beyond the Basics. Plast Reconstr Surg 2016, 138, 330e–340e. [Google Scholar] [CrossRef] [PubMed]
- de Matos, L.L.; Trufelli, D.C.; de Matos, M.G.L.; Pinhal, M.A. da S. Immunohistochemistry as an Important Tool in Biomarkers Detection and Clinical Practice. Biomark Insights 2010, 5, 9. [Google Scholar] [CrossRef] [PubMed]
- Goyal, L.; Hingmire, S.; Parikh, P.M. Newer Diagnostic Methods in Oncology. Med J Armed Forces India 2006, 62, 162. [Google Scholar] [CrossRef]
- Whelehan, P.; Evans, A.; Wells, M.; MacGillivray, S. The Effect of Mammography Pain on Repeat Participation in Breast Cancer Screening: A Systematic Review. Breast 2013, 22, 389–394. [Google Scholar] [CrossRef] [PubMed]
- Morton, C.A.; Mackie, R.M. Clinical Accuracy of the Diagnosis of Cutaneous Malignant Melanoma. Br J Dermatol 1998, 138, 283–287. [Google Scholar] [CrossRef] [PubMed]
- Badu-Peprah, A.; Adu-Sarkodie, Y. Accuracy of Clinical Diagnosis, Mammography and Ultrasonography in Preoperative Assessment of Breast Cancer. Ghana Med J 2018, 52, 133–139. [Google Scholar] [CrossRef] [PubMed]
- Schaffter, T.; Buist, D.S.M.; Lee, C.I.; Nikulin, Y.; Ribli, D.; Guan, Y.; Lotter, W.; Jie, Z.; Du, H.; Wang, S.; et al. Evaluation of Combined Artificial Intelligence and Radiologist Assessment to Interpret Screening Mammograms. JAMA Netw Open 2020, 3, E200265. [Google Scholar] [CrossRef]
- Marghoob, A.A.; Changchien, L.; DeFazio, J.; Dessio, W.C.; Malvehy, J.; Zalaudek, I.; Halpern, A.C.; Scope, A. The Most Common Challenges in Melanoma Diagnosis and How to Avoid Them. Australas J Dermatol 2009, 50, 1–13. [Google Scholar] [CrossRef]
- Poulet, G.; Massias, J.; Taly, V. Liquid Biopsy: General Concepts. Acta Cytol 2019, 63, 449–455. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D.; Weinberg, R.A. Hallmarks of Cancer: The next Generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Copeland, C.; Le, A. Glutamine Metabolism in Cancer. Adv Exp Med Biol 2021, 1311, 17–38. [Google Scholar] [CrossRef] [PubMed]
- Vaupel, P.; Schmidberger, H.; Mayer, A. The Warburg Effect: Essential Part of Metabolic Reprogramming and Central Contributor to Cancer Progression. Int J Radiat Biol 2019, 95, 912–919. [Google Scholar] [CrossRef] [PubMed]
- Schiliro, C.; Firestein, B.L. Mechanisms of Metabolic Reprogramming in Cancer Cells Supporting Enhanced Growth and Proliferation. Cells 2021, 10. [Google Scholar] [CrossRef] [PubMed]
- Dougherty, R.M.; Galli, C.; Ferro-Luzzi, A.; Iacono, J.M. Lipid and Phospholipid Fatty Acid Composition of Plasma, Red Blood Cells, and Platelets and How They Are Affected by Dietary Lipids: A Study of Normal Subjects from Italy, Finland, and the USA. Am J Clin Nutr 1987, 45, 443–455. [Google Scholar] [CrossRef] [PubMed]
- Halpern, Z.; Rubin, M.; Harach, G.; Grotto, I.; Mosor, A.; Dvir, A.; Lichtenberg, D.; Gilat, T. Bile and Plasma Lipid Composition in Non-Obese Normolipidemic Subjects with and without Cholesterol Gallstones. Liver 1993, 13, 246–252. [Google Scholar] [CrossRef]
- Maldonado, E.N.; Romero, J.R.; Ochoa, B.; Aveldao, M.I. Lipid and Fatty Acid Composition of Canine Lipoproteins. Comparative Biochemistry and Physiology - B Biochemistry and Molecular Biology 2001, 128, 719–729. [Google Scholar] [CrossRef]
- Meikle, P.J.; Barlow, C.K.; Mellett, N.A.; Mundra, P.A.; Bonham, M.P.; Larsen, A.; Cameron-Smith, D.; Sinclair, A.; Nestel, P.J.; Wong, G. Postprandial Plasma Phospholipids in Men Are Influenced by the Source of Dietary Fat. J Nutr 2015, 145, 2012–2018. [Google Scholar] [CrossRef]
- Feingold, K.R. Lipid and Lipoprotein Metabolism. Endocrinol Metab Clin North Am 2022, 51, 437–458. [Google Scholar] [CrossRef]
- Pegtel, D.M.; Gould, S.J. Exosomes. Annu Rev Biochem 2019, 88, 487–514. [Google Scholar] [CrossRef] [PubMed]
- Hessvik, N.P.; Llorente, A. Current Knowledge on Exosome Biogenesis and Release. Cell Mol Life Sci 2018, 75, 193–208. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, X.F. A Niche Role for Cancer Exosomes in Metastasis. Nat Cell Biol 2015, 17, 709–711. [Google Scholar] [CrossRef] [PubMed]
- Costa-Silva, B.; Aiello, N.M.; Ocean, A.J.; Singh, S.; Zhang, H.; Thakur, B.K.; Becker, A.; Hoshino, A.; Mark, M.T.; Molina, H.; et al. Pancreatic Cancer Exosomes Initiate Pre-Metastatic Niche Formation in the Liver. Nat Cell Biol 2015, 17, 816–826. [Google Scholar] [CrossRef] [PubMed]
- Yu, W.; Hurley, J.; Roberts, D.; Chakrabortty, S.K.; Enderle, D.; Noerholm, M.; Breakefield, X.O.; Skog, J.K. Exosome-Based Liquid Biopsies in Cancer: Opportunities and Challenges. Ann Oncol 2021, 32, 466–477. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, M.K.L.; Jose, J.; Wahba, M.; Bernaus-Esqué, M.; Hoy, A.J.; Enrich, C.; Rentero, C.; Grewal, T. Linking Late Endosomal Cholesterol with Cancer Progression and Anticancer Drug Resistance. Int J Mol Sci 2022, 23. [Google Scholar] [CrossRef] [PubMed]
- Munir, R.; Usman, H.; Hasnain, S.; Smans, K.; Kalbacher, H.; Zaidi, N. Atypical Plasma Lipid Profile in Cancer Patients: Cause or Consequence? Biochimie 2014, 102, 9–18. [Google Scholar] [CrossRef]
- Irshad, R.; Tabassum, S.; Husain, M. Aberrant Lipid Metabolism in Cancer: Current Status and Emerging Therapeutic Perspectives. Curr Top Med Chem 2023, 23, 1090–1103. [Google Scholar] [CrossRef] [PubMed]
- Yan, F.; Zhao, H.; Zeng, Y. Lipidomics: A Promising Cancer Biomarker. Clin Transl Med 2018, 7. [Google Scholar] [CrossRef]
- Di Gregorio, J.; Petricca, S.; Iorio, R.; Toniato, E.; Flati, V. Mitochondrial and Metabolic Alterations in Cancer Cells. Eur J Cell Biol 2022, 101. [Google Scholar] [CrossRef]
- Caino, M.C.; Altieri, D.C. Molecular Pathways: Mitochondrial Reprogramming in Tumor Progression and Therapy. Clin Cancer Res 2016, 22, 540–545. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, A.; Srivastava, P.; Mathur, S.; Abbas, S.; Rai, N.; Tiwari, S.; Tiwari, M.; Sharma, L. Lipid Metabolism and Mitochondria: Cross Talk in Cancer. Curr Drug Targets 2022, 23, 606–627. [Google Scholar] [CrossRef]
- Horvath, S.E.; Daum, G. Lipids of Mitochondria. Prog Lipid Res 2013, 52, 590–614. [Google Scholar] [CrossRef] [PubMed]
- Poulaki, A.; Giannouli, S. Mitochondrial Lipids: From Membrane Organization to Apoptotic Facilitation. Int J Mol Sci 2022, 23. [Google Scholar] [CrossRef]
- Paradies, G.; Paradies, V.; Ruggiero, F.M.; Petrosillo, G. Role of Cardiolipin in Mitochondrial Function and Dynamics in Health and Disease: Molecular and Pharmacological Aspects. Cells 2019, 8. [Google Scholar] [CrossRef]
- Ahmadpour, S.T.; Mahéo, K.; Servais, S.; Brisson, L.; Dumas, J.F. Cardiolipin, the Mitochondrial Signature Lipid: Implication in Cancer. Int J Mol Sci 2020, 21, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Paradies, G.; Paradies, V.; De Benedictis, V.; Ruggiero, F.M.; Petrosillo, G. Functional Role of Cardiolipin in Mitochondrial Bioenergetics. Biochim Biophys Acta 2014, 1837, 408–417. [Google Scholar] [CrossRef]
- Praharaj, P.P.; Naik, P.P.; Panigrahi, D.P.; Bhol, C.S.; Mahapatra, K.K.; Patra, S.; Sethi, G.; Bhutia, S.K. Intricate Role of Mitochondrial Lipid in Mitophagy and Mitochondrial Apoptosis: Its Implication in Cancer Therapeutics. Cell Mol Life Sci 2019, 76, 1641–1652. [Google Scholar] [CrossRef]
- Goicoechea, L.; Conde de la Rosa, L.; Torres, S.; García-Ruiz, C.; Fernández-Checa, J.C. Mitochondrial Cholesterol: Metabolism and Impact on Redox Biology and Disease. Redox Biol 2023, 61. [Google Scholar] [CrossRef]
- Ribas, V.; García-Ruiz, C.; Fernández-Checa, J.C. Mitochondria, Cholesterol and Cancer Cell Metabolism. Clin Transl Med 2016, 5. [Google Scholar] [CrossRef]
- Garcia-Ruiz, C.; Conde de la Rosa, L.; Ribas, V.; Fernandez-Checa, J.C. MITOCHONDRIAL CHOLESTEROL AND CANCER. Semin Cancer Biol 2021, 73, 76–85. [Google Scholar] [CrossRef] [PubMed]
- Qu, Q.; Zeng, F.; Liu, X.; Wang, Q.J.; Deng, F. Fatty Acid Oxidation and Carnitine Palmitoyltransferase I: Emerging Therapeutic Targets in Cancer. Cell Death Dis 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Melone, M.A.B.; Valentino, A.; Margarucci, S.; Galderisi, U.; Giordano, A.; Peluso, G. The Carnitine System and Cancer Metabolic Plasticity. Cell Death Dis 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Console, L.; Scalise, M.; Mazza, T.; Pochini, L.; Galluccio, M.; Giangregorio, N.; Tonazzi, A.; Indiveri, C. Carnitine Traffic in Cells. Link With Cancer. Front Cell Dev Biol 2020, 8. [Google Scholar] [CrossRef] [PubMed]
- Farahzadi, R.; Hejazi, M.S.; Molavi, O.; Pishgahzadeh, E.; Montazersaheb, S.; Jafari, S. Clinical Significance of Carnitine in the Treatment of Cancer: From Traffic to the Regulation. Oxid Med Cell Longev 2023, 2023. [Google Scholar] [CrossRef] [PubMed]
- Dörsam, B.; Fahrer, J. The Disulfide Compound α-Lipoic Acid and Its Derivatives: A Novel Class of Anticancer Agents Targeting Mitochondria. Cancer Lett 2016, 371, 12–19. [Google Scholar] [CrossRef] [PubMed]
- Wenzel, U.; Nickel, A.; Daniel, H. Alpha-Lipoic Acid Induces Apoptosis in Human Colon Cancer Cells by Increasing Mitochondrial Respiration with a Concomitant O2-*-Generation. Apoptosis 2005, 10, 359–368. [Google Scholar] [CrossRef]
- Bosso, M.; Haddad, D.; Al Madhoun, A.; Al-Mulla, F. Targeting the Metabolic Paradigms in Cancer and Diabetes. Biomedicines 2024, 12. [Google Scholar] [CrossRef]
- Carrié, L.; Virazels, M.; Dufau, C.; Montfort, A.; Levade, T.; Ségui, B.; Andrieu-Abadie, N. New Insights into the Role of Sphingolipid Metabolism in Melanoma. Cells 2020, 9, 1–29. [Google Scholar] [CrossRef]
- Yesmin, F.; Bhuiyan, R.H.; Ohmi, Y.; Yamamoto, S.; Kaneko, K.; Ohkawa, Y.; Zhang, P.; Hamamura, K.; Cheung, N.K. V.; Kotani, N.; et al. Ganglioside GD2 Enhances the Malignant Phenotypes of Melanoma Cells by Cooperating with Integrins. Int J Mol Sci 2021, 23. [Google Scholar] [CrossRef]
- Noujarède, J.; Carrié, L.; Garcia, V.; Grimont, M.; Eberhardt, A.; Mucher, E.; Genais, M.; Schreuder, A.; Carpentier, S.; Ségui, B.; et al. Sphingolipid Paracrine Signaling Impairs Keratinocyte Adhesion to Promote Melanoma Invasion. Cell Rep 2023, 42. [Google Scholar] [CrossRef] [PubMed]
- Portoukalian, J.; Zwingelstein, G.; Doré, J. -F Lipid Composition of Human Malignant Melanoma Tumors at Various Levels of Malignant Growth. Eur J Biochem 1979, 94, 19–23. [Google Scholar] [CrossRef]
- Realini, N.; Palese, F.; Pizzirani, D.; Pontis, S.; Basit, A.; Bach, A.; Ganesan, A.; Piomelli, D. Acid Ceramidase in Melanoma: EXPRESSION, LOCALIZATION, AND EFFECTS OF PHARMACOLOGICAL INHIBITION. J Biol Chem 2016, 291, 2422–2434. [Google Scholar] [CrossRef] [PubMed]
- Perez-Valle, A.; Abad-García, B.; Fresnedo, O.; Barreda-Gómez, G.; Aspichueta, P.; Asumendi, A.; Astigarraga, E.; Fernández, J.A.; Boyano, M.D.; Ochoa, B. A UHPLC-Mass Spectrometry View of Human Melanocytic Cells Uncovers Potential Lipid Biomarkers of Melanoma. Int J Mol Sci 2021, 22. [Google Scholar] [CrossRef] [PubMed]
- Huergo-Baños, C.; Velasco, V.; Garate, J.; Fernández, R.; Martín-Allende, J.; Zabalza, I.; Artola, J.L.; Martí, R.M.; Asumendi, A.; Astigarraga, E.; et al. Lipid Fingerprint-Based Histology Accurately Classifies Nevus, Primary Melanoma, and Metastatic Melanoma Samples. Int J Cancer 2024, 154, 712–722. [Google Scholar] [CrossRef]
- Prouteau, A.; André, C. Canine Melanomas as Models for Human Melanomas: Clinical, Histological, and Genetic Comparison. Genes (Basel) 2019, 10. [Google Scholar] [CrossRef]
- Schiffman, J.D.; Breen, M. Comparative Oncology: What Dogs and Other Species Can Teach Us about Humans with Cancer. Philos Trans R Soc Lond B Biol Sci 2015, 370. [Google Scholar] [CrossRef]
- Abdelmegeed, S.M.; Mohammed, S. Canine Mammary Tumors as a Model for Human Disease. Oncol Lett 2018, 15, 8195–8205. [Google Scholar] [CrossRef]
- Pinho, S.S.; Carvalho, S.; Cabral, J.; Reis, C.A.; Gärtner, F. Canine Tumors: A Spontaneous Animal Model of Human Carcinogenesis. Transl Res 2012, 159, 165–172. [Google Scholar] [CrossRef]
- Stevenson, V.B.; Klahn, S.; LeRoith, T.; Huckle, W.R. Canine Melanoma: A Review of Diagnostics and Comparative Mechanisms of Disease and Immunotolerance in the Era of the Immunotherapies. Front Vet Sci 2023, 9. [Google Scholar] [CrossRef]
- Gillard, M.; Cadieu, E.; De Brito, C.; Abadie, J.; Vergier, B.; Devauchelle, P.; Degorce, F.; Dréano, S.; Primot, A.; Dorso, L.; et al. Naturally Occurring Melanomas in Dogs as Models for Non-UV Pathways of Human Melanomas. Pigment Cell Melanoma Res 2014, 27, 90–102. [Google Scholar] [CrossRef]
- Gardner, H.L.; Fenger, J.M.; London, C.A. Dogs as a Model for Cancer. Annu Rev Anim Biosci 2016, 4, 199–222. [Google Scholar] [CrossRef] [PubMed]
- Nishiya, A.T.; Massoco, C.O.; Felizzola, C.R.; Perlmann, E.; Batschinski, K.; Tedardi, M.V.; Garcia, J.S.; Mendonça, P.P.; Teixeira, T.F.; Dagli, M.L.Z. Comparative Aspects of Canine Melanoma. Vet Sci 2016, 3. [Google Scholar] [CrossRef]
- Palma, S. Di; McConnell, A.; Verganti, S.; Starkey, M. Review on Canine Oral Melanoma: An Undervalued Authentic Genetic Model of Human Oral Melanoma? Vet Pathol 2021, 58, 881–889. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, T.F.; Gentile, L.B.; Da Silva, T.C.; Mennecier, G.; Chaible, L.M.; Cogliati, B.; Roman, M.A.L.; Gioso, M.A.; Dagli, M.L.Z. Cell Proliferation and Expression of Connexins Differ in Melanotic and Amelanotic Canine Oral Melanomas. Vet Res Commun 2014, 38, 29–38. [Google Scholar] [CrossRef] [PubMed]
- Alexander, A.N.; Huelsmeyer, M.K.; Mitzey, A.; Dubielzig, R.R.; Kurzman, I.D.; MacEwen, E.G.; Vail, D.M. Development of an Allogeneic Whole-Cell Tumor Vaccine Expressing Xenogeneic Gp100 and Its Implementation in a Phase II Clinical Trial in Canine Patients with Malignant Melanoma. Cancer Immunol Immunother 2006, 55, 433–442. [Google Scholar] [CrossRef] [PubMed]
- van der Weyden, L.; Brenn, T.; Patton, E.E.; Wood, G.A.; Adams, D.J. Spontaneously Occurring Melanoma in Animals and Their Relevance to Human Melanoma. J Pathol 2020, 252, 4–21. [Google Scholar] [CrossRef] [PubMed]
- Simpson, R.M.; Bastian, B.C.; Michael, H.T.; Webster, J.D.; Prasad, M.L.; Conway, C.M.; Prieto, V.M.; Gary, J.M.; Goldschmidt, M.H.; Esplin, D.G.; et al. Sporadic Naturally Occurring Melanoma in Dogs as a Preclinical Model for Human Melanoma. Pigment Cell Melanoma Res 2014, 27, 37–47. [Google Scholar] [CrossRef] [PubMed]
- Spangler, W.L.; Kass, P.H. The Histologic and Epidemiologic Bases for Prognostic Considerations in Canine Melanocytic Neoplasia. Vet Pathol 2006, 43, 136–149. [Google Scholar] [CrossRef]
- Shuman, A.G.; Light, E.; Olsen, S.H.; Pynnonen, M.A.; Taylor, J.M.G.; Johnson, T.M.; Bradford, C.R. Mucosal Melanoma of the Head and Neck: Predictors of Prognosis. Arch Otolaryngol Head Neck Surg 2011, 137, 331–337. [Google Scholar] [CrossRef]
- Millanta, F.; Fratini, F.; Corazza, M.; Castagnaro, M.; Zappulli, V.; Poli, A. Proliferation Activity in Oral and Cutaneous Canine Melanocytic Tumours: Correlation with Histological Parameters, Location, and Clinical Behaviour. Res Vet Sci 2002, 73, 45–51. [Google Scholar] [CrossRef]
- Oh, J.H.; Cho, J.Y. Comparative Oncology: Overcoming Human Cancer through Companion Animal Studies. Exp Mol Med 2023, 55, 725–734. [Google Scholar] [CrossRef] [PubMed]
- Ranieri, G.; Gadaleta, C.D.; Patruno, R.; Zizzo, N.; Daidone, M.G.; Hansson, M.G.; Paradiso, A.; Ribatti, D. A Model of Study for Human Cancer: Spontaneous Occurring Tumors in Dogs. Biological Features and Translation for New Anticancer Therapies. Crit Rev Oncol Hematol 2013, 88, 187–197. [Google Scholar] [CrossRef]
- K, L.-T.; CM, W.; TS, M.; EK, K.; DB, J.; M, K.; M, C.; JL, C.; EJ, K.; MC, Z.; et al. Genome Sequence, Comparative Analysis and Haplotype Structure of the Domestic Dog. Nature 2005, 438, 803–819. [Google Scholar] [CrossRef]
- Broniec, A.; Goto, M.; Matsuki, H. A Peculiar Phase Transition of Plasmalogen Bilayer Membrane under High Pressure. Langmuir 2009, 25, 11265–11268. [Google Scholar] [CrossRef]
- Aramaki, S.; Tsuge, S.; Islam, A.; Eto, F.; Sakamoto, T.; Oyama, S.; Li, W.; Zhang, C.; Yamaguchi, S.; Takatsuka, D.; et al. Lipidomics-Based Tissue Heterogeneity in Specimens of Luminal Breast Cancer Revealed by Clustering Analysis of Mass Spectrometry Imaging: A Preliminary Study. PLoS One 2023, 18. [Google Scholar] [CrossRef]
- Wu, Q.; Yu, J.; Zhang, M.; Xiong, Y.; Zhu, L.; Wei, B.; Wu, T.; Du, Y. Serum Lipidomic Profiling for Liver Cancer Screening Using Surface-Assisted Laser Desorption Ionization MS and Machine Learning. Talanta 2024, 268. [Google Scholar] [CrossRef] [PubMed]
- Bifarin, O.O.; Sah, S.; Gaul, D.A.; Moore, S.G.; Chen, R.; Palaniappan, M.; Kim, J.; Matzuk, M.M.; Fernández, F.M. Machine Learning Reveals Lipidome Remodeling Dynamics in a Mouse Model of Ovarian Cancer. J Proteome Res 2023, 22, 2092–2108. [Google Scholar] [CrossRef]
- Xiao, Y.; Ma, D.; Yang, Y.S.; Yang, F.; Ding, J.H.; Gong, Y.; Jiang, L.; Ge, L.P.; Wu, S.Y.; Yu, Q.; et al. Comprehensive Metabolomics Expands Precision Medicine for Triple-Negative Breast Cancer. Cell Res 2022, 32, 477. [Google Scholar] [CrossRef]
- Li, R.; Li, L.; Xu, Y.; Yang, J. Machine Learning Meets Omics: Applications and Perspectives. Brief Bioinform 2022, 23. [Google Scholar] [CrossRef]
- Cai, Z.; Poulos, R.C.; Liu, J.; Zhong, Q. IScience Machine Learning for Multi-Omics Data Integration in Cancer. iScience 2022, 25, 103798. [Google Scholar] [CrossRef] [PubMed]
- Arjmand, B.; Hamidpour, S.K.; Tayanloo-Beik, A.; Goodarzi, P.; Aghayan, H.R.; Adibi, H.; Larijani, B. Machine Learning: A New Prospect in Multi-Omics Data Analysis of Cancer. Front Genet 2022, 13, 824451. [Google Scholar] [CrossRef] [PubMed]
- Csala, A.; Zwinderman, A.H. Multivariate Statistical Methods for High-Dimensional Multiset Omics Data Analysis. Computational Biology 2019, 71–83. [Google Scholar] [CrossRef]
- Caponigro, V.; Tornesello, A.L.; Merciai, F.; La Gioia, D.; Salviati, E.; Basilicata, M.G.; Musella, S.; Izzo, F.; Megna, A.S.; Buonaguro, L.; et al. Integrated Plasma Metabolomics and Lipidomics Profiling Highlights Distinctive Signature of Hepatocellular Carcinoma in HCV Patients. J Transl Med 2023, 21, 1–15. [Google Scholar] [CrossRef]
- Wolrab, D.; Jirásko, R.; Cífková, E.; Höring, M.; Mei, D.; Chocholoušková, M.; Peterka, O.; Idkowiak, J.; Hrnčiarová, T.; Kuchař, L.; et al. Lipidomic Profiling of Human Serum Enables Detection of Pancreatic Cancer. Nat Commun 2022, 13. [Google Scholar] [CrossRef]
- Jiang, N.; Zhang, Z.; Chen, X.; Zhang, G.; Wang, Y.; Pan, L.; Yan, C.; Yang, G.; Zhao, L.; Han, J.; et al. Plasma Lipidomics Profiling Reveals Biomarkers for Papillary Thyroid Cancer Diagnosis. Front Cell Dev Biol 2021, 9. [Google Scholar] [CrossRef]
- Kim, H.Y.; Lee, H.; Kim, S.H.; Jin, H.; Bae, J.; Choi, H.K. Discovery of Potential Biomarkers in Human Melanoma Cells with Different Metastatic Potential by Metabolic and Lipidomic Profiling. Sci Rep 2017, 7. [Google Scholar] [CrossRef]
- Kurokawa, G.A.; Hamamoto Filho, P.T.; Delafiori, J.; Galvani, A.F.; de Oliveira, A.N.; Dias-Audibert, F.L.; Catharino, R.R.; Pardini, M.I.M.C.; Zanini, M.A.; Lima, E. de O.; et al. Differential Plasma Metabolites between High- and Low-Grade Meningioma Cases. Int J Mol Sci 2023, 24, 394. [Google Scholar] [CrossRef]
- Gallart-Ayala, H.; Courant, F.; Severe, S.; Antignac, J.P.; Morio, F.; Abadie, J.; Le Bizec, B. Versatile Lipid Profiling by Liquid Chromatography-High Resolution Mass Spectrometry Using All Ion Fragmentation and Polarity Switching. Preliminary Application for Serum Samples Phenotyping Related to Canine Mammary Cancer. Anal Chim Acta 2013, 796, 75–83. [Google Scholar] [CrossRef]
- Safari Yazd, H.; Bazargani, S.F.; Fitzpatrick, G.; Yost, R.A.; Kresak, J.; Garrett, T.J. Metabolomic and Lipidomic Characterization of Meningioma Grades Using LC-HRMS and Machine Learning. J Am Soc Mass Spectrom 2023, 34, 2187–2198. [Google Scholar] [CrossRef]
- Shang, X.; Zhang, C.; Kong, R.; Zhao, C.; Wang, H. Construction of a Diagnostic Model for Small Cell Lung Cancer Combining Metabolomics and Integrated Machine Learning. Oncologist 2024, 29, e392–e401. [Google Scholar] [CrossRef] [PubMed]
- Starodubtseva, N.L.; Tokareva, A.O.; Rodionov, V. V.; Brzhozovskiy, A.G.; Bugrova, A.E.; Chagovets, V. V.; Kometova, V. V.; Kukaev, E.N.; Soares, N.C.; Kovalev, G.I.; et al. Integrating Proteomics and Lipidomics for Evaluating the Risk of Breast Cancer Progression: A Pilot Study. Biomedicines 2023, 11. [Google Scholar] [CrossRef] [PubMed]
- Manzi, M.; Palazzo, M.; Knott, M.E.; Beauseroy, P.; Yankilevich, P.; Giménez, M.I.; Monge, M.E. Coupled Mass-Spectrometry-Based Lipidomics Machine Learning Approach for Early Detection of Clear Cell Renal Cell Carcinoma. J Proteome Res 2021, 20, 841–857. [Google Scholar] [CrossRef]
- Wang, G.; Yao, H.; Gong, Y.; Lu, Z.; Pang, R.; Li, Y.; Yuan, Y.; Song, H.; Liu, J.; Jin, Y.; et al. Metabolic Detection and Systems Analyses of Pancreatic Ductal Adenocarcinoma through Machine Learning, Lipidomics, and Multi-Omics. Sci Adv 2021, 7, 2724. [Google Scholar] [CrossRef]
- Yang, C.; Zhou, S.; Zhu, J.; Sheng, H.; Mao, W.; Fu, Z.; Chen, Z. Plasma Lipid-Based Machine Learning Models Provides a Potential Diagnostic Tool for Colorectal Cancer Patients. Clinica Chimica Acta 2022, 536, 191–199. [Google Scholar] [CrossRef]
- Krishnan, S.T.; Winkler, D.; Creek, D.; Anderson, D.; Kirana, C.; Maddern, G.J.; Fenix, K.; Hauben, E.; Rudd, D.; Voelcker, N.H. Staging of Colorectal Cancer Using Lipid Biomarkers and Machine Learning. Metabolomics 2023, 19. [Google Scholar] [CrossRef]
- Zhou, J.; Ji, N.; Wang, G.; Zhang, Y.; Song, H.; Yuan, Y.; Yang, C.; Jin, Y.; Zhang, Z.; Zhang, L.; et al. Metabolic Detection of Malignant Brain Gliomas through Plasma Lipidomic Analysis and Support Vector Machine-Based Machine Learning. EBioMedicine 2022, 81. [Google Scholar] [CrossRef] [PubMed]
- Saito, R.; Yoshimura, K.; Shoda, K.; Furuya, S.; Akaike, H.; Kawaguchi, Y.; Murata, T.; Ogata, K.; Iwano, T.; Takeda, S.; et al. Diagnostic Significance of Plasma Lipid Markers and Machine Learning-Based Algorithm for Gastric Cancer. Oncol Lett 2021, 21. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.C.; Wu, W.H.; Huang, S.; Li, Z.W.; Li, X.; Shui, G.H.; Lam, S.M.; Li, B.W.; Li, Z.X.; Zhang, Y.; et al. Plasma Lipids Signify the Progression of Precancerous Gastric Lesions to Gastric Cancer: A Prospective Targeted Lipidomics Study. Theranostics 2022, 12, 4671. [Google Scholar] [CrossRef]
- Kujala, M.; Nevalainen, J. A Case Study of Normalization, Missing Data and Variable Selection Methods in Lipidomics. Stat Med 2015, 34, 59–73. [Google Scholar] [CrossRef]
- Del Prete, E.; Campos, A.M.; Della Rocca, F.; Gallo, C.; Fontana, A.; Nuzzo, G.; Angelini, C. ADViSELipidomics: A Workflow for Analyzing Lipidomics Data. Bioinformatics 2022, 38, 5460–5462. [Google Scholar] [CrossRef]
- Ding, X.; Yang, F.; Chen, Y.; Xu, J.; He, J.; Zhang, R.; Abliz, Z. Norm ISWSVR: A Data Integration and Normalization Approach for Large-Scale Metabolomics. Anal Chem 2022, 94, 7500–7509. [Google Scholar] [CrossRef]
- Ulmer, C.Z.; Ragland, J.M.; Koelmel, J.P.; Heckert, A.; Jones, C.M.; Garrett, T.J.; Yost, R.A.; Bowden, J.A. LipidQC: Method Validation Tool for Visual Comparison to SRM 1950 Using NIST Interlaboratory Comparison Exercise Lipid Consensus Mean Estimate Values. Anal Chem 2017, 89, 13069–13073. [Google Scholar] [CrossRef]
- Köfeler, H.C.; Ahrends, R.; Baker, E.S.; Ekroos, K.; Han, X.; Hoffmann, N.; Holcapek, M.; Wenk, M.R.; Liebisch, G. Recommendations for Good Practice in Ms-Based Lipidomics. J Lipid Res 2021, 62. [Google Scholar] [CrossRef] [PubMed]
- O’Donnell, V.B.; Fitzgerald, G.A.; Murphy, R.C.; Liebisch, G.; Dennis, E.A.; Quehenberger, O.; Subramaniam, S.; Wakelam, M.J.O. Steps Toward Minimal Reporting Standards for Lipidomics Mass Spectrometry in Biomedical Research Publications. Circ Genom Precis Med 2020, 13, E003019. [Google Scholar] [CrossRef]
- Triebl, A.; Burla, B.; Selvalatchmanan, J.; Oh, J.; Tan, S.H.; Chan, M.Y.; Mellet, N.A.; Meikle, P.J.; Torta, F.; Wenk, M.R. Shared Reference Materials Harmonize Lipidomics across MS-Based Detection Platforms and Laboratories. J Lipid Res 2020, 61, 105–115. [Google Scholar] [CrossRef]
- Tsuchida, S.; Nakayama, T. MALDI-Based Mass Spectrometry in Clinical Testing: Focus on Bacterial Identification. Applied Sciences 2022, Vol. 12, Page 2814 2022, 12, 2814. [Google Scholar] [CrossRef]
- Poirion, O.B.; Jing, Z.; Chaudhary, K.; Huang, S.; Garmire, L.X. DeepProg: An Ensemble of Deep-Learning and Machine-Learning Models for Prognosis Prediction Using Multi-Omics Data. Genome Med 2021, 13. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Song, M.; Shen, H.; Hong, H.; Gong, P.; Deng, H.-W.; Zhang, C. Deep Learning Methods for Omics Data Imputation. Biology (Basel) 2023, 12. [Google Scholar] [CrossRef]
- Kang, M.; Ko, E.; Mersha, T.B. A Roadmap for Multi-Omics Data Integration Using Deep Learning. Brief Bioinform 2022, 23. [Google Scholar] [CrossRef]
- Albaradei, S.; Thafar, M.; Alsaedi, A.; Van Neste, C.; Gojobori, T.; Essack, M.; Gao, X. Machine Learning and Deep Learning Methods That Use Omics Data for Metastasis Prediction. Comput Struct Biotechnol J 2021, 19, 5008–5018. [Google Scholar] [CrossRef] [PubMed]
- Tran, K.A.; Kondrashova, O.; Bradley, A.; Williams, E.D.; Pearson, J. V.; Waddell, N. Deep Learning in Cancer Diagnosis, Prognosis and Treatment Selection. Genome Med 2021, 13. [Google Scholar] [CrossRef] [PubMed]
- Babu, M.; Snyder, M. Multi-Omics Profiling for Health. Mol Cell Proteomics 2023, 22, 100561. [Google Scholar] [CrossRef] [PubMed]
- Lima, A.R.; Carvalho, M.; Aveiro, S.S.; Melo, T.; Domingues, M.R.; Macedo-Silva, C.; Coimbra, N.; Jerónimo, C.; Henrique, R.; Bastos, M. de L.; et al. Comprehensive Metabolomics and Lipidomics Profiling of Prostate Cancer Tissue Reveals Metabolic Dysregulations Associated with Disease Development. J Proteome Res 2022, 21, 727–739. [Google Scholar] [CrossRef]
- Jung, J.H.; Yang, D.Q.; Song, H.; Wang, X.; Wu, X.; Kim, K.P.; Pandey, A.; Byeon, S.K. Characterization of Lipid Alterations by Oncogenic PIK3CA Mutations Using Untargeted Lipidomics in Breast Cancer. OMICS 2023, 27, 327–335. [Google Scholar] [CrossRef]
Figure 1.
Lipids and mitochondrial metabolism in cancer. (a) Cardiolipin involvement in mitochondrial quality control and induction of mitophagy (b) Increased cholesterol levels in tumor mitochondria and the regulation of membrane characteristics. (c) Carnitine in cancer; involvement in Warburg effect, Tricarboxylic acid (TCA) and β-oxidation pathways. (d) Regulation of mTOR through lipoic acid in cancer.
Figure 1.
Lipids and mitochondrial metabolism in cancer. (a) Cardiolipin involvement in mitochondrial quality control and induction of mitophagy (b) Increased cholesterol levels in tumor mitochondria and the regulation of membrane characteristics. (c) Carnitine in cancer; involvement in Warburg effect, Tricarboxylic acid (TCA) and β-oxidation pathways. (d) Regulation of mTOR through lipoic acid in cancer.
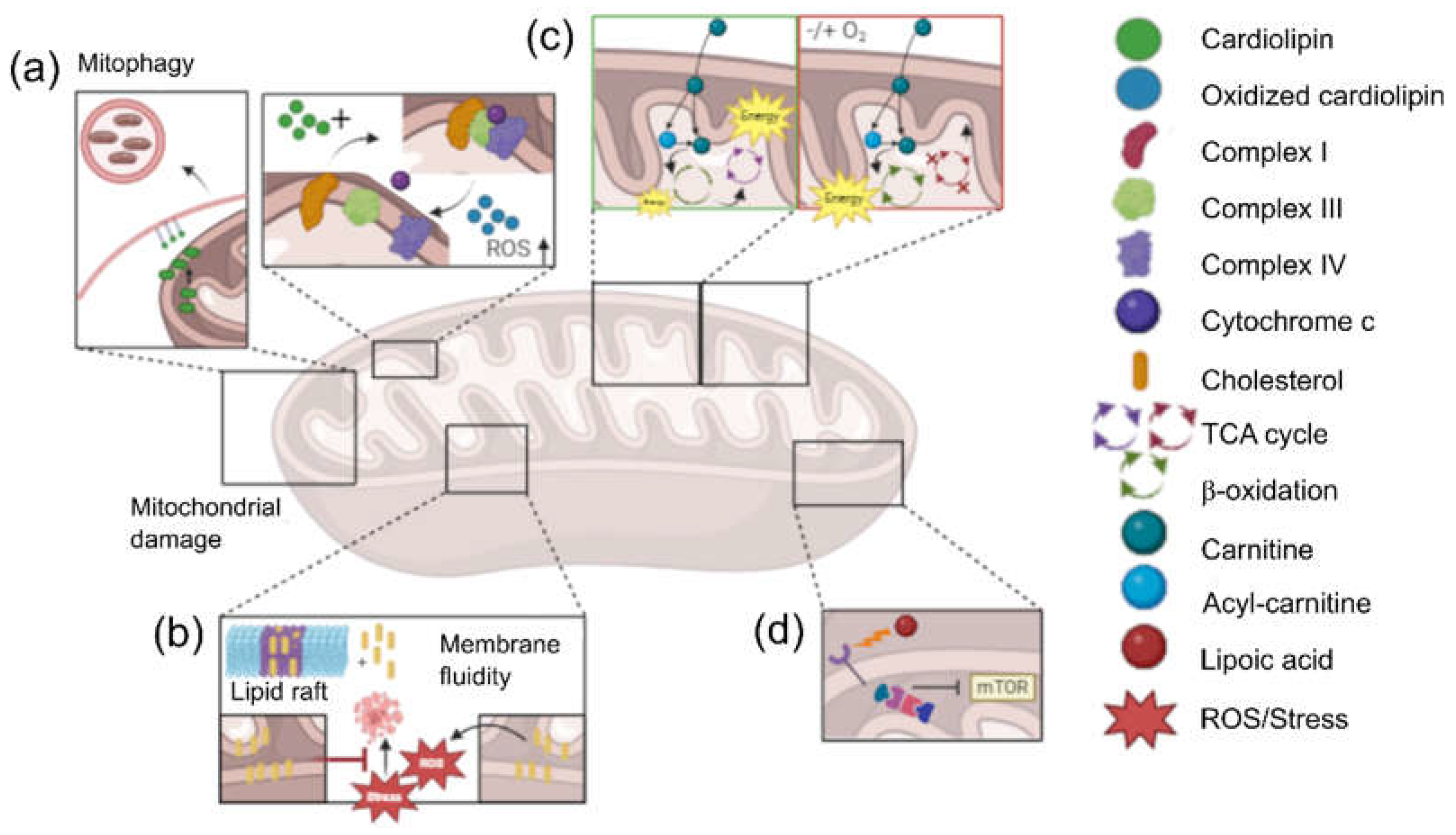
Figure 2.
Graphic representation of the process of using mass spectrometry for untargeted lipidomic analysis in the early diagnosis of cancer. (a) Sample collection. (b) Lipid profile analysis using mass spectrometry. (c) Data analysis using Machine Learning algorithms. (d) Early diagnosis of cancer.
Figure 2.
Graphic representation of the process of using mass spectrometry for untargeted lipidomic analysis in the early diagnosis of cancer. (a) Sample collection. (b) Lipid profile analysis using mass spectrometry. (c) Data analysis using Machine Learning algorithms. (d) Early diagnosis of cancer.
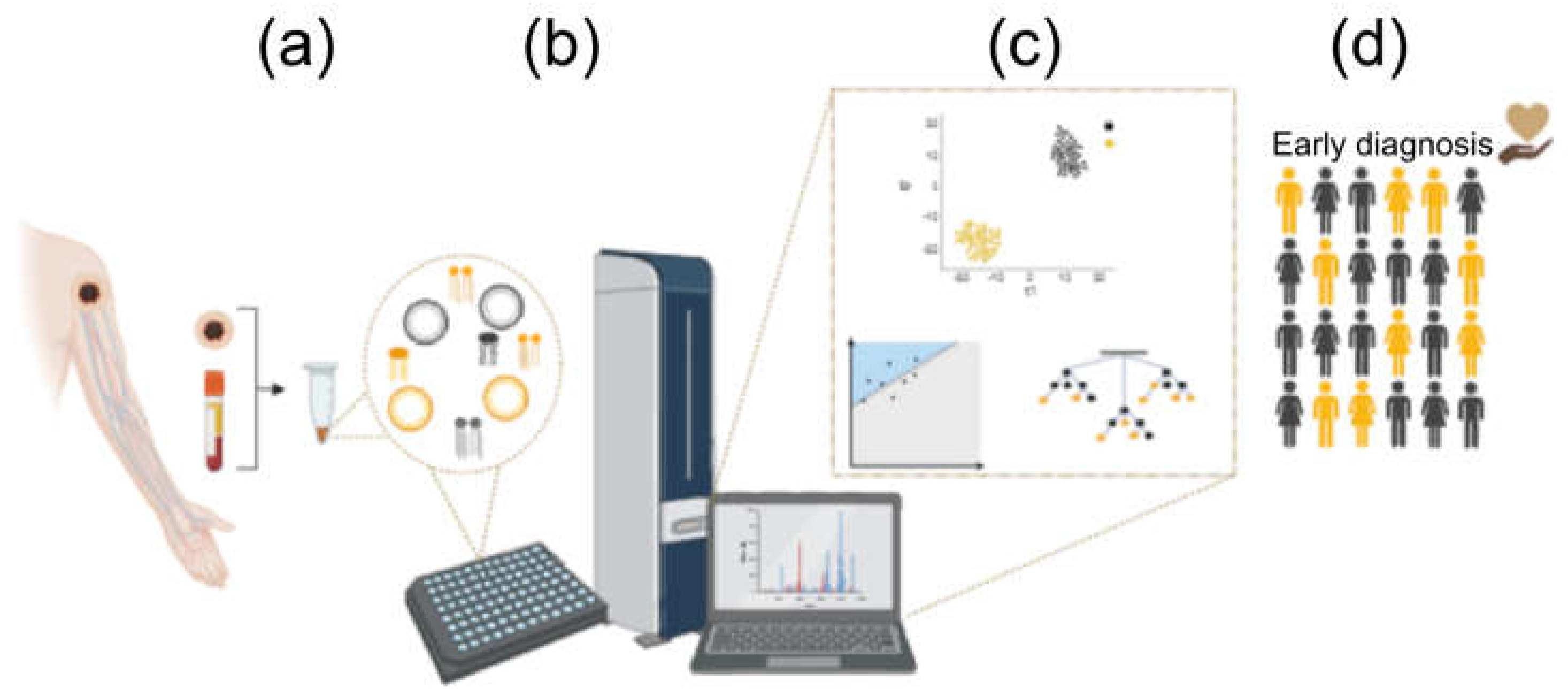

Table 1.
Lipidomics studies and Machine Learning algorithms used.
| Reference | Cancer Origin | Analyzed Tissue | Acquisition | ML Algorithms | Notes |
|---|---|---|---|---|---|
| [65] | Melanoma | Solid biopsy | MALDI-MS | LR, NB, SVM | Lipid imaging |
| [89] | Mammary | Solid biopsy | UHPLC-MS | LASSO, SVM | SRAA |
| [86] | Mammary | Solid biopsy | DESI | KNN | Lipid imaging |
| [100] | Meningioma | Solid biopsy | LC-HRMS | DT, KNN, LR, NB, RF, SVM | |
| [88] | Mouse ovarian | Solid biopsy | UHPLC-MS | LR, RF, KNN, SVM, VC | SRAA |
| [101] | Lung | Serum | LC-MS/MS | LR, RF, SVM | Panel of 8 metabolites, SRAA |
| [102] | Mammary | Serum | LC-MS | LR | Tumor metastatic potential |
| [87] | Liver | Serum | MALDI-MS | LDA, LR, MLP, RF, SVM | |
| [103] | Renal | Serum | UPLC-MS | LASSO-SVM | Coupled ML algorithms |
| [104] | Pancreas | Serum | MALDI-MS | SVM | |
| [105] | Colorectal | Plasma | LC-MS | KNN, PLS, RF, SVM | |
| [106] | Colorectal | Plasma | LC-MS | MLR-EM, BRANN | Tumor Stage Classification |
| [107] | Glioma | Plasma | HPLC-MS | SVM | |
| [108] | Gastric | Plasma | LC/ESI-MS | LR | |
| [109] | Gastric | Plasma | UHPLC-MS | LR |
* In bold, the algorithm showing best results. Abbreviations: BRANN, Bayesian Regularized Artificial Neural Networks; DT, Decision Tree; KNN, k-Nearest Neighbor; LASSO, Least Absolute Shrinkage and Selection Operator; LDA, Linear Discriminant Analysis; LR, Logistic Regression; ML, Machine Learning; MLP, Multilayer Perceptron; MLR-EM, Multiple Linear Regression with Expectation Maximization; NB, Naïve Bayes; RF, Random Forest; SRAA, Similar results for all algorithms; SVM, Support Vector Machine; VC, Voting Classifier.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



