Submitted:
04 July 2024
Posted:
05 July 2024
You are already at the latest version
Abstract
Leishmaniasis represents a severe global health problem. In the last decades, there have been significant challenges in controlling this disease due to the unavailability of licensed vaccines, the high toxicity of the available drugs, and an unrestrained surge of drug-resistant parasites, and human immunodeficiency virus (HIV)–Leishmania co-infections. Leishmania spp. preferentially invade macrophage lineage cells of vertebrates for replication after subverting cellular functions of humans and other mammals. These early events in host–parasite interactions are likely to influence the future course of the disease. Thus, there is a continuing need to discover a simple cellular model that reproduces the in vivo pathogenesis. Acanthamoeba spp. are non-mammalian phagocytic amoeba with remarkable similarity to the cellular and functional aspects of macrophages. We aimed to assess whether the similarity reported between macrophages and Acanthamoeba spp. is sufficient to reproduce the infectivity of Leishmania spp. Herein, we analyzed co-cultures of Acanthamoeba castellanii or Acanthamoeba polyphaga with Leishmania infantum, Leishmania amazonensis, Leishmania major, and Leishmania braziliensis. Light and fluorescence microscopy revealed that the flagellated promastigotes attach to the A. castellanii and/or A. polyphaga in a bipolar and or random manner, which initiates their uptake via pseudopods. Once inside the cells, the promastigotes undergo significant changes, which result in the obligatory amastigote-like intracellular form. There was a productive infection with a continuous increase in intracellular parasites. However, we frequently observed intracellular amastigotes in vacuoles, phagolysosomes, and the cytosol of Acanthamoeba spp. Our findings corroborate that Leishmania spp. infects Acanthamoeba spp. and replicates in them but does not cause their rapid degeneration or lysis. Overall, the evidence presented here confirms that Acanthamoeba spp. have all prerequisites and can help elucidate how Leishmania spp. infect mammalian cells. Future work exposing the mechanisms of these interactions should yield novel insights into how these pathogens exploit amoebae.
Keywords:
Acanthamoeba castellanii
; Acanthamoeba polyphaga
; Leishmania
; interaction host-parasite
Author Summary
Leishmaniasis represents a severe global health problem, and drug resistance is a growing concern. Leishmania spp. are obligate intracellular parasites that survive within cells of the vertebrate macrophage, modulating their activation. Understanding the multilayered relationship between metabolism and function of innate immune cells during infection has great therapeutic and preventive potential. Mammalian macrophages and Acanthamoeba spp. display similarities in the molecular mechanisms involved in directional motility, recognition, binding, engulfment, and the phagolysosomal processes, and they express similar receptors. Hence, we hypothesize that Acanthamoeba spp. represent a model that can be used to evaluate macrophage–pathogen interactions from the perspective of innate immunity. However, it has not yet been described whether Leishmania spp. can survive in Acanthamoeba spp. We found that Acanthamoeba spp. support Leishmania growth. Acanthamoeba spp. contain ancient pathogen recognition mechanisms, and Leishmania spp. could manipulate amoeboid functions to favor their survival and replication based on strategies of ancestral metabolic pathways. Our robust evidence highlights that amoebae could be used as a model to understand the biology and evolution of host–Leishmania interactions.
Introduction
Leishmaniasis is a term used to define a wide-ranging group of diseases caused by protozoa of the genus Leishmania [1,2]. These infections result in a broad spectrum of clinical manifestations that range from asymptomatic infection to localized, disseminated ulcers, sometimes with mucosal involvement (cutaneous leishmaniasis), or affect internal organs, such as the liver, spleen, and bone marrow (visceral leishmaniasis), which can be fatal if left untreated [3,4]. In recent years, leishmaniasis has been considered a neglected and emerging group of diseases. These diseases constitute a global grave public-health problem that leads to great suffering and high socio-cultural and economic costs [5]. There is no available vaccine and drug resistance is emerging [6]. Additionally, the available drugs have severe side effects, further complicating treatment, and cannot completely eliminate the pathogen [6]: The parasite remains in the body and can cause a relapse when there is immunosuppression [6,7]. According to Word Health Organization (WHO), over 1 billion people are at risk of infection in high-burden countries [8,9]. There are an estimated 700,000 to 1 million new cases each year, highlighting the importance of this public health problem [10].
In a vertebrate host, Leishmania parasites can evade the defense barriers of the host and even benefit from them [9]. Indubitably, macrophages are key cells to the body’s guards against pathogens [10]. However, Leishmania spp. have evolved to modulate vacuole biogenesis to generate a unique intravacuolar niche that is suitable for their proliferation inside macrophages [4]. In this niche, Leishmania differentiates into the amastigote form that inhabits digestive vacuoles, which fuse to macrophage lysosomes, generating phagolysosomes [3]. Elucidating the complex Leishmania–macrophage interactions is crucial to prevent and treat leishmaniasis [5]. To survive successfully and multiply within macrophages, Leishmania must undergo profound biochemical and morphological adaptations to enter a host cell, establish an infection, and evade or modulate the subsequent immune response. These abilities might have resulted from cellular and molecular adaptations to the ancestral phagocytosis process [15].
Mammalian macrophages and Acanthamoeba spp. display similarities in the molecular mechanisms involved in directional motility, recognition, binding, engulfment, and phagolysosome processing of bacteria and fungi [16]. They also display conserved pathogen-recognition mechanisms [16,17]. Acanthamoeba spp. are opportunistic protozoans, ubiquitously distributed in nature, that are mainly found in soil and water. In general, the life cycle of these amoeba consists of two stages, trophozoite and cyst. Trophozoites are the motile forms that mainly feeds on bacteria, algae, yeast, or small organic particles via phagocytosis or pinocytosis [16,18]. Trophozoites closely resemble the internal structure of a mammalian cell. They contain various organelles such as mitochondria, ribosomes, centrosomes, vacuoles, and the Golgi apparatus. Vacuoles are a critical component of trophozoites: they include the contractile vacuole, which expels water for osmotic regulation, lysosomes, digestive vacuoles, and many glycogen-containing vacuoles [18].Over the past several decades, Acanthamoeba spp. have gained increasing attention due to their ability to act as a host/reservoir for microbial pathogens, particularly their role in causing damage and sometimes fatal human and animal infections [16]. Recent studies suggest that Acanthamoeba–microbe adaptations are considered an evolutionary model of macrophage–pathogen interactions and acts as a “genetic melting pot” with cross-species exchange of genes [17].
Due to their remarkable cellular and functional similarities, authors have speculated that Acanthamoeba spp. and macrophages are evolutionarily related [15,18]. An exciting hypothesis is Acanthamoeba spp. may be an in vitro model to investigate pathogen–macrophage interactions [19].
Phagocytosis is a complex process that has been maintained during evolution [15,20]. Acanthamoeba spp. are well known for their exuberant motility and active pseudopodia when interact with bacteria, viruses, protists, and fungi [17,19]. Acanthamoeba spp. can harbor important human pathogens such as Legionella pneumophila, Aeromonas spp., Pseudomonas spp., Escherichia coli, and Mycobacterium spp., and others cultured from clinical specimens [19,21,22,23]. It is noteworthy that some of these pathogens can subvert the microbicidal activity and replicate inside Acanthamoeba spp. with mechanisms that are similar to those used for survival within macrophages [24,25,26]. While Acanthamoeba spp. and macrophages take up microbes via phagocytosis, whether these distinct hosts have an evolutionary or convergent evolution remains unknown. Moreover, not all the phagocytic pathways are totally conserved between amoebae and macrophages, the endocytic system evasion mechanisms such as inhibition of phagosome–lysosome fusion, are common to escape from either host [19].
Despite efforts to unravel the mechanisms of Leishmania pathogenicity and the risk of infection, and to develop novel treatments and vaccines against the parasite, there are still gaps in the state-of-the-art that need be explored. The innate immunity pathways are widely conserved in nature, and evaluation of Acanthamoeba–Leishmania interactions could represent an important model to understand the mechanisms of virulence and host immunity. We evaluated whether Acanthamoeba spp. are similar enough to macrophages to reproduce Leishmania spp. infectivity. Understanding these interactions will potentially reveal new mechanisms for investigating host–parasite interactions.
Material and Methods
Acanthamoeba stains and cultivation conditions
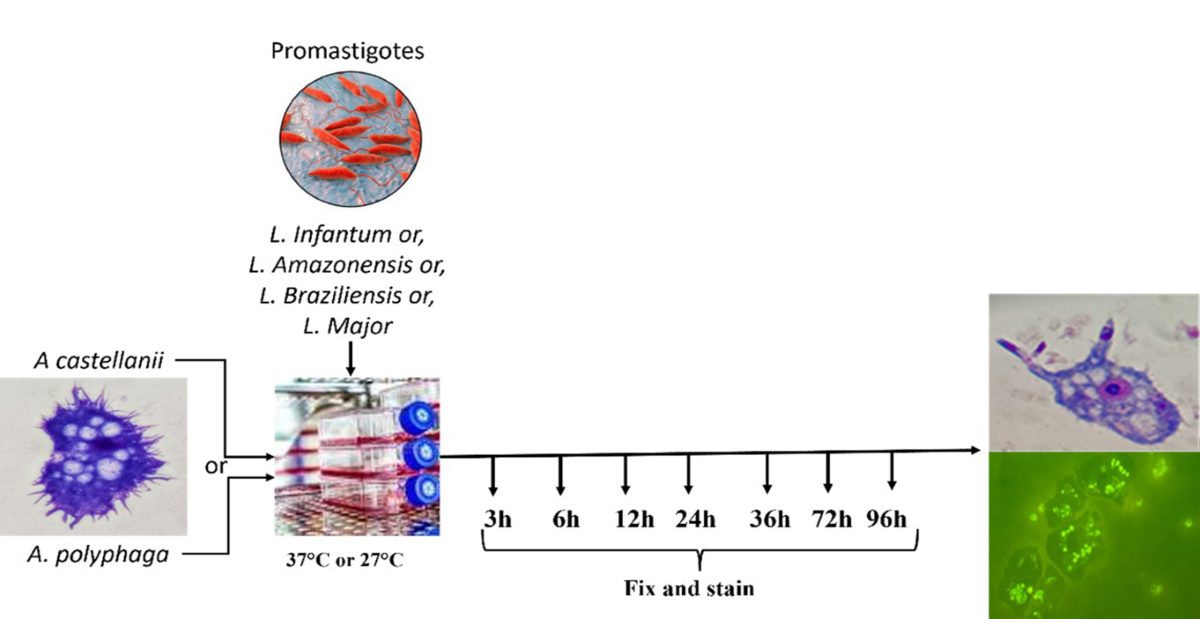
Two Acanthamoeba spp. strains were used in this study: an environmental strain Acanthamoeba castellanii designated as Neff (ATCC 30010), and Acanthamoeba polyphaga (ATCC 30431) that was isolated from corneal scrapings of a patient with acute ulceration of the right eye. The amoebae were grown in Peptone Yeast Glucose (PYG) medium containing 0.75% (w/v) proteose peptone, 0.75% (w/v) yeast extract, and 1.5% (w/v) glucose as a monolayer in 25 cm2 tissue culture flasks at 27°C. When cells formed a monolayer, the trophozoites were harvested, centrifuged (200 g for 5 min), washed three times with phosphate-buffered saline (PBS), and suspended in PYG broth or PBS. Cells were counted in a hemocytometer chamber, and the viability was assessed by the trypan blue exclusion assay. Prior to the experiments, the amoebae were grown in tissue culture flasks at 27°C without shaking, and the media refreshed after 17–20 h. In addition, the influence of culture media (Grace’s insect cell culture medium (Grace’s), liver infusion tryptose [LIT], and Schneider’s insect medium [Schneider’s], without supplementation with fetal calf serum [FCS]) on the growth and viability of Acanthamoeba spp. was assessed.
Parasite growth was monitored daily for 6 days with a Neubauer chamber. Viability was assessed by mobility and lack of staining after challenging with trypan blue. All results correspond to three independent assays that were run in duplicates. Before starting the experiments, parasites were maintained in each medium for a minimum of three passages.
Leishmania strains and cultivation conditions of promastigotes
Three causative species of cutaneous leishmaniasis, Leishmania amazonensis (MHOM/BR/PH8), Leishmania braziliensis (strain Thor, MCAN/BR/1998/619), and Leishmania major (MHOM/SU/1973/5-ASKH), and one causative specie of visceral leishmaniasis, Leishmania infantum (MHOM/BR/1974/PP75), were used in this study. Promastigotes were obtained from the Protozoa Culture Collection (COLPROT) and the Leishmania Collection (CLIOC) from Instituto Oswaldo Cruz – Fundação Oswaldo Cruz (IOC-FIOCRUZ) and cultured in Schneider’s medium (pH 7.2) supplemented with 20% fetal bovine serum (FBS), streptomycin (40 μg/mL), and penicillin (100 U/mL) at 27°C. The parasites were maintained by serial passaging twice per week. Before starting the experiments, promastigotes were sub-passaged to a fresh medium every 4 days to maintain the growth and viability of the parasites.
Co-culture of Leishmania spp. and Acanthamoeba spp.
For light microscopy analyses, A. castellanii and A. polyphaga were seeded in eight-well Nunc TM Lab-Tek TM II Chamber slides at 1 × 105 cells per well. Then, 1 × 106 promastigotes (L. braziliensis, L. amazonensis, L. infantum, and L. major) in the stationary growth phase of the respective culture media (1:10 ratio) were added to two wells each at 37°C or 27°C. After 3, 6, 12, 24, 36, 72 h, and 96 h, the supernatant from each chamber slide well was discarded and the top of the plate was removed. Non-adhered promastigotes were removed by three washing cycles with PBS (pH 7.2). The attached cells were fixed with methanol before being submitted to Panoptic fast staining. To evaluate infectivity, the slides were analyzed under an optical microscope (Olympus Optical) at 1000× total magnification using immersion oil. For fluorescence microscopy analyses, promastigotes in the mid-log growth phase were incubated with 0.1% fluorescein isothiocyanate (FITC) in a carbonate-bicarbonate buffer (pH 9.4; 9.5 mL of 0.2 M Na2CO3 mixed with 41.5 mL of 0.2 M NaHCO3) for 1 h at room temperature in the dark, based on a previously described method [27]. Finally, the pellet was resuspended in 1 mL of PBS. A. castellanii and A. polyphaga were seeded in eight-well Nunc TM Lab-Tek TM II Chamber slides at a density of 1 × 105 cells per well. Then, 1 × 106 FITC-labeled promastigotes forms (L. braziliensis, L. amazonensis, L. infantum, and L. major) in the stationary growth phase of the respective culture media (1:10 ratio) were added to two wells each, as described above. Each dried slide was fixed with absolute methanol and examined under a fluorescence microscope (Zeiss Axiolab) at 400× and 1000× total magnification using immersion oil.
The parasite release and differentiation assay
Twenty-four hours after infection, Acanthamoeba monolayers were washed and incubated with 0.01% sodium dodecyl sulfate (SDS; Sigma-Aldrich) for 10 min to allow lysis of Acanthamoeba and the release of amastigotes. Then, cultures were fed with five mL of Schneider's medium, supplemented with 10% FBS, and incubated at 27°C and for an additional 2 days.
Results
Before the infection experiments, Acanthamoeba spp. were assessed for growth potential in conventional culture media commonly used to isolate and maintain Leishmania spp. in vitro, namely Liver Infusion Typtose (LIT), Schneider’s medium, and Grace’s medium.
Acanthamoeba spp. were able to tolerate different growth conditions, without special nutritional requirements. Over six days, we counted the number of amoebae that grew in each culture medium (Figure 2).
Number of trophozoites in the different growth media: PYG, black, as the control media; LIT (yellow); Schneider’s (green), and Grace’s medium (red). Black arrowheads = the beginning of the logarithmic phase (BLP). The days of quantification are represented on the x-axis and the concentration of parasites per mL is expressed in a logarithmic scale on the y-axis.
At the control medium, trophozoite reached the beginning of the logarithmic phase (BLP) on day two, and a decrease in cell density was observed at six days over the course of the experiment. An initial population of 1.0 x 106 organisms/ml regularly gave rise to a maximal population > 1.0 X 107 organisms/ml within five days. A. castellanii showed slow growth over six days in LIT and Schneider’s media (1.3 ×106/mL). The number of parasites was low when compared with the classic PYG medium (1.8 × 107/mL at 5 days); however, morphological homogeneity and the trophozoites remained viable with a slightly increased cellular count at six days. It was easily observed live trophozoites extremely pleomorphic, moderately mobile, cytoplasmic acantopodia of varying size and shape, and a large round to oval nucleus with a prominent and centrally located nucleolus as well as abundant vacuoles of different size and content.
We assessed parasite viability with the trypan blue exclusion assay and by observing pseudopodia and morphology. The trypan blue exclusion assay showed a smooth reduction of viable parasites compared with the controls. Except for Grace’s medium, the other media tested showed a similar viability percentage. The estimated viability of Acanthamoeba was not less than 99%.
Acanthamoeba–Leishmania co-culture
We co-cultured L. braziliensis, L. amazonensis, L. infantum, and L. major with A. castellanii or A. polyphaga for up to 96 h. Light microscopical examination showed, after initial contact with the flagellum, promastigote forms attached to Acanthamoeba cells.
Formation of surface projections of the amoeba and protrusions engulfed the promastigotes from the flagellar tip toward the cell body (Figure 3A, B, D). We noted that long tubular pseudopods tightly encircled their flagellum (Figure 3C, E) or the posterior (non-flagellated) pole (Figure 3 C). Note the pseudopodial membrane that appears to form a tight grip on the promastigotes and to “flow” forward to engulf them. Engulfment of the adhering promastigotes occurred via funnel-like extensions of the Acanthamoeba surface extending along the radiating parasites (Fig 3C, F). Insertion of promastigote flagellum into the pseudopodium of an A. castellanii, indicative of a mode of entry with the flagellar end first.
Parasite ratio of 1:10. Protrusions engulfed the promastigotes from the flagellar tip toward the cell body and vice-versa (red arrowheads); K= kinetoplasts; N= nucleus; black arrowheads = flagella; * = numerous pine-like pseudopods (acanthopodia) gave the cell a spiny appearance. Scale bar 10 µm.
Once inside the cells, the promastigotes undergo significant changes, which result in the obligatory amastigote-like intracellular form. Acanthamoeba spp. engulfed promastigotes, permitted the intracellular conversion of the promastigotes to an amastigote-like state, and supported intracellular multiplication of L. braziliensis, L. amazonensis, L. infantum, and L. major (Figure 4). Cytosolic and intravascular intracellular localizations of the internalized parasite were observed. We found the parasites either membrane-bound within host cell–derived vacuoles (Figure 4A, B, C, D, E and 4H) or free in the cytosol (Figure 4F). The vacuoles showed either a tight-fitting (Figure 4E) or a loose membrane (Figure 4C and D) around the parasites.
A and B = Leishmania. amazonensis; C and D = Leishmania. braziliensis; E and F= Leishmania infantum and G and H= Leishmania major. N= nucleus of A. castellanii; black arrowheads indicate intracellular parasites inside vacuoles; * = the rounded/oval body; Scale bar 10 µm.
Moreover, we observed loose vacuoles harboring several parasites, with the parasites attached or not attached to the wall (Figure 5, black arrows), and the amastigote partially adheres to the PV membrane with the remaining portion being free in vacuolar (Figure 5A). The amastigote forms are observed free in the cytosol (Figure 5B, L, and I; yellow arrows). Figure 5A and G show parasites within the vacuole had two nuclei and two kinetoplasts probably ready to undergo cellular division. This process suggests that amastigotes may replicate within amoeba, supported by the demonstration of duplicated kinetoplasts, nucleus (Figure 5C), and flagella (Figure 5G). The duplication of these organelles is known to represent the initial event of the parasite's mitosis. Moreover, we also observed morphologically intact parasites (Figure 5F and M), and or the rounded/oval body (Figure 5D and N) and started losing elongated shape towards a more rounded one (Figure 5D and E.). Figure 5P and C suggest that replicated amastigote forms were released, after the host cells burst or, alternatively, in a synchronized event reminiscent of exocytosis (red arrows). This speculation is purely based on the light microscopy and needs further observations.
K= kinetoplast; N= nucleus; V= Vacuole; N= nucleus of the amoeba; black arrowheads indicate intracellular parasites inside vacuoles; Yellow arrowheads indicate amastigotes in the cytosol; red arrowheads indicate release of amastigotes; green arrowheads indicate amastigotes inside vacuoles; Scale bar 10 µm.
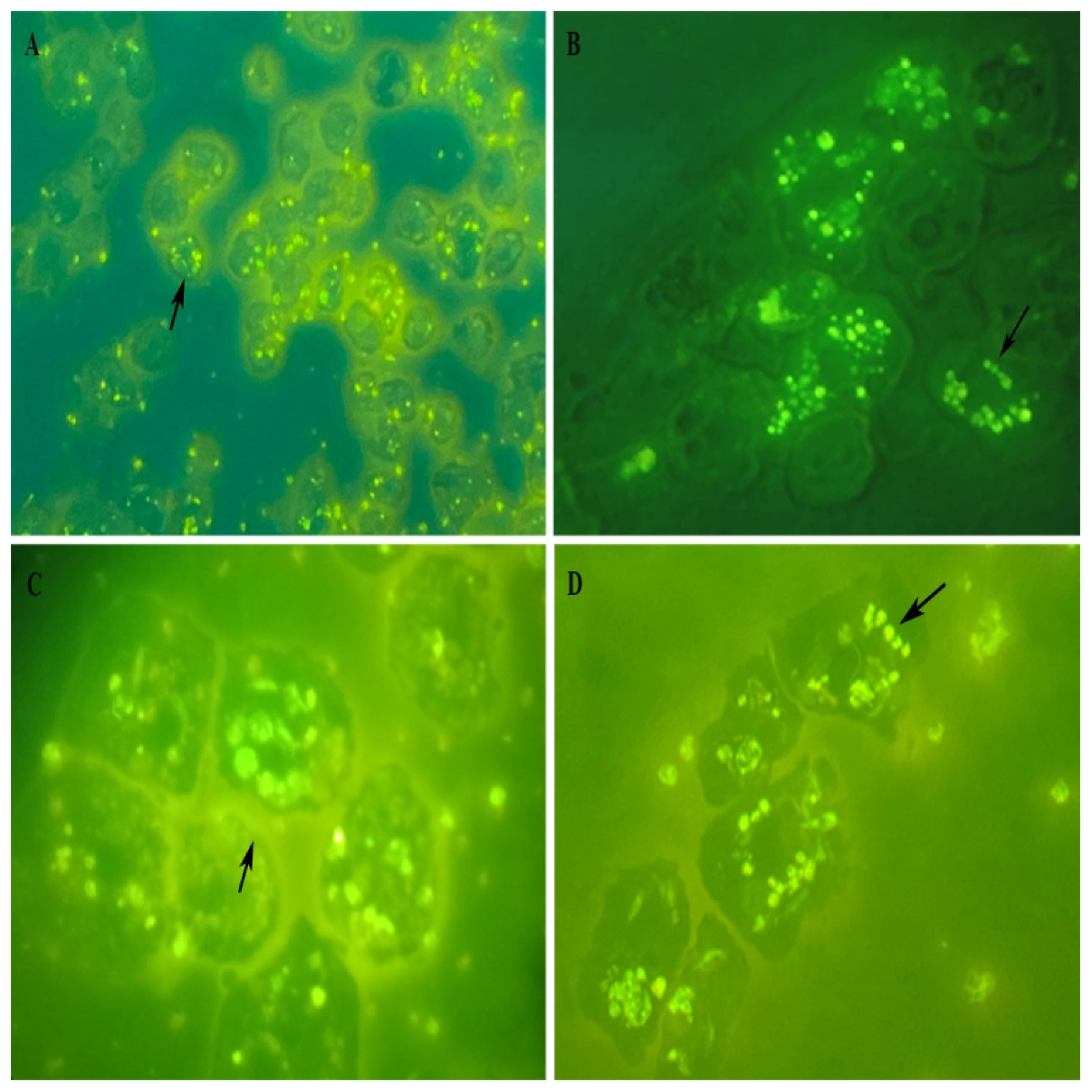
Some of those observations were further confirmed by fluorescence microscopy imaging of Acanthamoeba infected with FITC-labeled promastigotes that reinforced our observation on internalization of Leishmania promastigotes. Figure 6 shows promastigotes undergoing significant changes, intact parasites (Figure 6C and D), rounded/oval bodies free in the cytosol (Figure 6A), or in vacuole (Figure 6C and D), that result in the obligatory amastigote-like intracellular forms, and fluorescence microscopy images of parasites multiplied within vacuoles (Figure 6C and D). Taken together, these results show that Leishmania promastigotes internalize and multiply into host cells.
(A) Low magnification view of Leishmania-FITC-labeled promastigotes, magnification × 100; (B) The fluorescence signal showing the labeling within trophozoite, magnification × 400; (C) Internalized promastigote and amastigotes, showing the labeling in vacuole, magnification × 1000, enlargement, and (D) illustrates the high degree of multiplication revealed by the labeling of intravacuolar amastigotes (black arrows), magnification × 1000.
Although different from each other, all the four infection profiles indicated that the interval for Leishmania is between 24 and 48h, as infected cells and the number of parasites per cell decreased after this period and in extra-cellular increased the number of rounded parasites with long flagella.
After Acanthamoeba lysis with 0.01% SDS, the release of amastigote forms was observed in the medium. These intracellular amastigotes were viable, since numerous newly generated free promastigote forms could be seen in the culture medium after transfer of 48 and 72 h-cocultures for 4 to 6 additional days at 27°C (data not shown). All results correspond to three independent assays run in duplicate using Schneider’s medium at 27 or 37°C.
Discussion
This report represents the first description of the abilities of L. infantum, L. amazonensis, L. major, and L. braziliensis to grow and subvert the functions of A. castellanii and A. polyphaga. Like macrophages, intracellular infection of Acanthamoeba spp. by Leishmania promastigotes relies on a series of key events, including attachment, internalization, amastigote differentiation, and intracellular survival [26]. The use of a cell line to study infections in vitro is an essential approach to evaluate the distinct aspects of Leishmania biology as well as its prevention and treatment, which require a clear understanding of the molecular network between parasites and their hosts [27]. Understanding the interactions would allow us to combat the disease by developing new drugs and therapies or by identifying important parasite antigens that could be exploited to develop vaccines.
There have been several reports of Leishmania parasitizing cells other than macrophages [28,29]; however, it is recognized that in a vertebrate host, Leishmania spp. mainly resides within cells of the macrophage lineage [29]. Macrophages from different origins are amenable to successful Leishmania infection in vitro [30,31]. These cells have been a mainstay of leishmaniasis research for more than 40 years, although there are many knowledge gaps. The main challenge is to elucidate that mechanisms lead to the subversion of macrophages [32]. Designing and applying a physiologically robust in vitro assay is highly relevant for drug screening and evaluation of biochemistry, physiology, and metabolism.
Macrophages represent the major effector cells that phagocytose parasites within vertebrate hosts [29]. The interaction between the innate and adaptive immune response is a crucial factor in the process of differentiating macrophages into pro-inflammatory (M1) or anti-inflammatory (M2) phenotypes, leading to resistance or susceptibility to Leishmania infection. Besides, various immune cells, including T cells, play a significant role in modulating macrophage polarization by releasing cytokines that influence macrophage maturation and function. Other immune cells can also influence macrophage polarization in a T cell–independent manner [31,32]. The host’s immune responses and individual variations in parasite strains may be critical to determine the clinical outcome of leishmaniasis in a patient [30]. The influence of the innate immune response during Leishmania infection has been proposed to favor parasite survival, allowing it to fully establish itself within the mammalian host [29,32].
In the present study, Acanthamoeba spp. were similar to macrophages in that they were permissible to Leishmania spp., but they would have no influence on the adaptive immune response. Interestingly, Acanthamoeba spp. are well-characterized non-mammalian phagocytic cells with remarkable similarity to macrophages. Indeed, the similar traits and features of macrophages and Acanthamoeba spp.—including the ancient process of phagocytosis—provide evidence of a common ancestry [34,35] or convergent evolution. Mammalian macrophages and Acanthamoeba spp. display similarities in the molecular mechanisms involved in directional motility, recognition, binding, engulfment, and phagolysosome processes, and express common primitive receptors [16,17,36]. Like macrophages, some microorganisms have evolved strategies for efficient uptake by Acanthamoeba, and they are able to regulate phagosome maturation to make the phagosome more hospitable for growth and to avoid destruction [19].
Acanthamoeba spp. are considered hosts for bacteria, viruses, yeast, and protozoa [37,38]. Their genome includes several genes that encode pattern recognition receptors (e.g., lipopolysaccharide-binding protein/bactericidal permeability enhancing protein [LBP/BPI], C-lectin, C-type lectin, mannose-binding protein [MBP], D-galactoside/L-rhamnose-containing lectin SUEL domain, NB-ARC tetratricopeptide containing repeat-containing protein ERVR, and endogenous virus receptor) [39]. These receptors allow amoeba to interact with a vast array of species. Over the past several decades, Acanthamoeba spp. have received increasing attention due to their ability to act as a host/reservoir for microbes, including viruses (mimivirus, coxsackieviruses, adenoviruses, poliovirus, echovirus, enterovirus, vesicular stomatitis virus, etc.), bacteria (Aeromonas, Bacillus, Bartonella, Burkholderia, Campylobacter, Chlamydophila, Coxiella, E. coli, Flavobacterium, Helicobacter, Legionella, Listeria, Staphylococcus, Mycobacterium, Pasteurella, Prevotella, Porphyromonas, Pseudomonas, Rickettsia, Salmonella, Shigella, Vibrio, etc.), protists (Cryptosporidium and Toxoplasma gondii), and yeast (Cryptococcus, Blastomyces, Sporothrix, Histoplasma, Streptomyces, Exophiala, etc.) [16,17,40]. Moreover, several pathogens can survive within A. castellanii by using similar strategies as when they encounter macrophages [16,41,42,43,44,45]. A survey showed that the Dot/Icm type IV secretion system is required by L. pneumophila for intracellular proliferation within human macrophages and Acanthamoeba [44]. Moreover, L. pneumophila is taken up by Acanthamoeba by coiling phagocytosis but can impede phagosome–lysosome fusion. In turn, Vibrio cholerae can resist amoeboid phagosome killing by neutralizing proteolytic enzymes, changing the local pH, and counteracting reactive nitrogen and oxygen species that may otherwise kill internalized bacteria [46]. Likewise, invasion of A. castellanii or macrophages by Mycobacterium spp. shows notable similarities at the transcriptional and post-translational levels [47].
A. castellanii can also engulf fungi that are potential pathogens, such as Cryptococcus neoformans, Histoplasma capsulatum, Blastomyces dermatitides, Fusarium solani, and Sporothrix schenckii [48]. Mannose-binding lectins (MBLs) play a critical role in activating antifungal responses by immune cells [49] (Allen PG, Dawidowicz EA , 1990). A study has suggested that mannose-binding proteins identified in fungal recognition by both amoeboid and macrophage surfaces belong to the Conavalin A-like superfamily. Yet, the results reveal a limitation: a short sequence of 53 amino acids does not have enough resolution to predict protein folding accurately. [36]. However, data from the literature show us that macrophages and Acanthamoeba may share similar interaction mechanisms with fungal and bacteria pathogens, indicating a possible convergent evolution.
We found that L. braziliensis, L. Amazonensis, L. infantum and L. major could infect and grow within A. castellanii and A. polyphaga. These amoebae engulfed promastigotes, permitted the intracellular conversion of the promastigotes to an amastigote-like state, and supported their intracellular multiplication. There was a visible increase in the number of parasites per infected Acanthamoeba and in the total number of parasites per monolayer (although the number of parasites were not quantified). We evaluated attachment and infection primarily by inspecting fixed specimens from co-cultures of promastigotes and Acanthamoeba spp. A potential pitfall in this method is the possibility that some of the attached parasites might have been superimposed on the Acanthamoeba image and thus erroneously scored as phagocytosed. Despite this limitation, our findings provide novel insight into host–leishmania interactions.
A. castellanii showed slow growth over 6 days in LIT and Schneider’s medium but remained viable and the death phase was not detected. Schneider’s insect medium (Schneider’s) and Grace’s insect cell culture medium (Grace’s) are defined media and are indicated for the continuous in vitro cultivation of promastigote forms of Leishmania [51]. However, a study demonstrated that the supplementation of LIT culture medium with fetal calf serum is the most suitable strategy to cultivate L. infantum parasites enabling the maintenance of growth and infective parasites for research uses. The authors showed that promastigote forms have a better growth in LIT and Schneider’s with or without FCS when compared to that in Grace’s [52].
In nature, promastigotes may be inoculated at a cutaneous site where complement is not active. Acanthamoeba spp. are lower eukaryotes that utilize a diverse repertoire of predicted pattern recognition receptors, many with predicted orthologous functions in the innate immune systems of higher organisms [39]. Many studies have shown that the interaction between the innate and adaptive immune responses is a crucial factor leading to resistance or susceptibility to Leishmania infection [53]. It is conceivable that interfering with these processes represents a promising new strategy for antileishmanial intervention. In the present study, Leishmania spp. could subvert the functions of A. castellanii and A. polyphaga based on the innate immune response. Leishmania spp. could manipulate amoeboid functions to favor their survival and replication based on strategies of ancestral metabolic pathways. These novel findings indicate that Leishmania spp. and Acanthamoeba spp. co-cultures are a relatively inexpensive and easy-to-control model system for which multiple variables can be controlled. This cell model might provide new insights into how Leishmania infects cells and new therapeutic avenues to manipulate the innate immune response to protect cells from Leishmania infection.
Conclusion
We have shown that L. braziliensis, L. amazonensis, L. infantum, and L. major are amoeba-resistant flagellates and that A. castellanii and A. polyphaga can be useful as a model to evaluate interactions for Leishmania species. Our findings contribute to the knowledge about the molecular aspects of the Leishmania–Acanthamoeba interaction considering innate immunity. The identification of promising molecules involved during phagocytosis and subversion of Leishmania spp. in amoeba could be extrapolated to macrophages. Thus, this cellular model should contribute to the understanding of the yet-to-be-elucidated mechanisms that are important to combat leishmaniasis.
Funding
This study was supported by the Instituto Oswaldo Cruz intramural funding (PAEF II-IOC-23-FIO-18-2-53), and the Fundação Carlos Chagas Filho de Amparo à Pesquisa do Estado do Rio de Janeiro (FAPERJ) Project ID: Nº DO PROCESSO SEI-260003/002148/2021 - APQ1. All the funding received during this specific study had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Data Availability
All relevant data are within the paper and its Supporting Information files.
Author’s contribution
HLCS: Conception, design, carried out all the experiments and analyzed data, and draft writing. CM: Funding acquisition and writing-review. IC: assisted in immunofluorescence assay. VEV: assisted in writing-review and editing. GL and RBR: Assisted in the maintenance-culture. All authors read and approved the final manuscript.
Acknowledgments
The manuscript benefitted from the insightful comments of three anonymous reviewers.
Competing Interests
The authors have declared that no competing interests exist.
References
- Mann S, Frasca K, Scherrer S, et al. A review of Leishmaniasis: current knowledge and future directions. Curr Trop Med Rep. 2021; 8:121-132.
- Gupta AK, Das S, Kamran M, Ejazi SA, Ali N. The pathogenicity and virulence of Leishmania - interplay of virulence factors with host defenses. Virulence, 2022; 13(1):903-935.
- Torres-Guerrero E, Quintanilla-Cedillo MR, Ruiz-Esmenjaud J, Arenas R. Leishmaniasis: a review. F1000Res. 2017; 6: 750.
- Yadav, P.; Azam, M.; Ramesh, V.; Singh, R. Unusual Observations in Leishmaniasis—An Overview. Pathogens 2023; 12:297.
- Sunyoto T, Boelaert M, Meheus F. Understanding the economic impact of leishmaniasis on households in endemic countries: a systematic review. Expert Rev Anti Infect Ther. 2019; 17:57-69.
- Burza S, Croft SL, Boelaert M. Leishmaniasis. Lancet. 2018; 15;392(10151):951-970.
- Hefnawy, A., Berg, M., Dujardin, J.-C. & De Muylder, G. Exploiting knowledge on Leishmania drug resistance to support the quest for new drugs. Trends Parasitol.2017; 33,162–174.
- Lindoso, J. A. L., Cunha, M. A., Queiroz, I. T. & Moreira, C. H. V. Leishmaniasis–HIV coinfection: current challenges. HIV AIDS. 2016;8, 147-156.
- Sampaio Tavares Veras P, Castro-Gomes T, Perrone Bezerra de Menezes J. Elucidating the Complex Interrelationship on Early Interactions between Leishmania and Macrophages [Internet]. Macrophages - Celebrating 140 Years of Discovery. IntechOpen; 2022. pp. 1-21. [CrossRef]
- WHO. Leishmaniasis Fact Sheet. World Health Organ (2022). Available online at: https://www.who.int/news-room/fact-sheets/detail/leishmaniasis (accessed: July 15, 2023).
- Conceição-Silva F, Morgado FN. Leishmania Spp-Host Interaction: There Is Always an Onset, but Is There an End? Front Cell Infect Microbiol. 2019; 19 (9):330.
- Almeida, F.S.; Vanderley, S.E.R.; Comberlang, F.C.; Andrade, A.G.d.; Cavalcante-Silva, L.H.A.; Silva, E.d.S.; Palmeira, P.H.d.S.; Amaral, I.P.G.d.; Keesen, T.S.L. Leishmaniasis: Immune Cells Crosstalk in Macrophage Polarization. Trop. Med. Infect. Dis. 2023, 8, 276. [Google Scholar] [CrossRef] [PubMed]
- Alexander J, Vickerman K. Fusion of host cell secondary lysosomes with the parasitophorous vacuoles of Leishmania mexicana-infected macrophages. The Journal of Protozoology. 1975;22(4):502-508.
- Podinovskaia M, Descoteaux A. Leishmania and the macrophage: a multifaceted interaction. Future Microbiol. 2015;10(1):111-29.
- Bajgar A, Krejčová G. On the origin of the functional versatility of macrophages. Front Physiol. 2023; 23(14):1128984.
- Siddiqui, R., Khan, N.. Acanthamoeba is an evolutionary ancestor of macrophages: A myth or reality?. Experimental Parasitology. 2012; 130(2):95-97.
- Mungroo MR, Siddiqui R, Khan NA. War of the microbial world: Acanthamoeba spp. interactions with microorganisms. Folia Microbiol (Praha). 2021;66(5):689-699.
- Bowers, B., and Korn, E. D.. The fine structure of Acanthamoeba castellanii. I. The trophozoite. J. Cell Biol. 1968;39, 95-111.
- Allan J. Guimaraes, Kamilla Xavier Gomes, Juliana Reis Cortines, José Mauro Peralta, Regina H.Saramago Peralta. Acanthamoeba spp. as a universal host for pathogenic microorganisms: One bridge from environment to host virulence. Microbiological Research. 2016; 193:30-38.
- Boulais J, Trost M, Landry CR, Dieckmann R, Levy ED, Soldati T, Michnick SW, Thibault P, Desjardins M. Molecular characterization of the evolution of phagosomes. Mol Syst Biol. 2010; 19(6):423.
- Mella, C; Medina, G; Flores-Martin, S; Toledo, Z; Simaluiza, RJ; Pérez-Pérez, G; Fernández, H.Interaction between zoonotic bacteria and free living amoebas. A new angle of an epidemiological polyhedron of public health importance? Austral Journal of Veterinary Sciences. 2016; 48(1): 1-10.
- Sandstrom, G., Saeed, A., Abd, H., 2011. Acanthamoeba-bacteria: a model to study host interaction with human pathogens. Curr. Drug Targets. 2011; 12 (7):936–941.
- Segal, Gil, and Howard A. Shuman. "Legionella pneumophila utilizes the same genes to multiply within Acanthamoeba castellanii and human macrophages." Infection and immunity. 1999; 67(5): 2117-2124.
- Derengowski Lda, S., Paes, H. C., Albuquerque, P., Tavares, A. H., Fernandes, L., Silva-Pereira, I., et al. (2013). The transcriptional response of Cryptococcus neoformans to ingestion by Acanthamoeba castellanii and macrophages provides insights into the evolutionary adaptation to the mammalian host. Eukaryot. Cell. 2013; 12:761–774.
- Rayamajhee B, Subedi D, Peguda HK, Willcox MD, Henriquez FL, Carnt N. A Systematic Review of Intracellular Microorganisms within Acanthamoeba to Understand Potential Impact for Infection. Pathogens. 2021; 10(2):225. [CrossRef]
- Steenbergen, J.N., Shuman, H.A., Casadevall, A. Cryptococcus neoformans interactions with amoebae suggest an explanation for its virulence and intracellular pathogenic strategy in macrophages. Proc. Natl. Acad. Sci. U. S. A. 2001; 98(26):15245–15250.
- Roy S, Kumar GA, Jafurulla M, Mandal C, Chattopadhyay A. Integrity of the actin cytoskeleton of host macrophages is essential for Leishmania donovani infection. Biochim Biophys Acta. 2014;1838(8):2011-8.
- Valigurová, A.; Koláˇrová, I.Unrevealing the Mystery of Latent Leishmaniasis: What Cells Can Host Leishmania? Pathogens. 2023; 12, 246.
- Costa-da-Silva AC, Nascimento DO, Ferreira JRM, Guimarães-Pinto K, Freire-de-Lima L, Morrot A, Decote-Ricardo D, Filardy AA, Freire-de-Lima CG. Immune Responses in Leishmaniasis: An Overview. Trop Med Infect Dis. 2022; 31;7(4):54. [CrossRef]
- Yasmin, H.; Adhikary, A.; Al-Ahdal, M.N.; Roy, S.; Kishore, U. Host–Pathogen Interaction in Leishmaniasis: Immune Response and Vaccination Strategies. Immuno 2022; 2, 218-254.
- Tomiotto-Pellissier F, Bortoleti BTdS, Assolini JP, Gonçalves MD, Carloto ACM, Miranda-Sapla MM, Conchon-Costa I, Bordignon J and Pavanelli WR. Macrophage Polarization in Leishmaniasis: Broadening Horizons. Front. Immunol 2018; 9:2529.
- Lecoeur H, Prina E, Gutiérrez-Sanchez M, Späth GF. Going ballistic: Leishmania nuclear subversion of host cell plasticity. Trends Parasitol. 2022;38(3):205-216.
- Costa-da-Silva, A.C.; Nascimento, D.d.O.; Ferreira, J.R.M.; Guimarães-Pinto, K.; Freire-de-Lima, L.; Morrot, A.; Decote-Ricardo, D.; Filardy, A.A.; Freire-de-Lima, C.G. Immune Responses in Leishmaniasis: An Overview. Trop. Med. Infect. Dis. 2022; 7(4):54.
- Mills DB. The origin of phagocytosis in Earth history. Interface Focus. 2020; 6, 10(4):20200019.
- Bajgar A and Krejčová G. On the origin of the functional versatility of macrophages. Front. Physiol. 2023; 14:1128984.
- Ferreira MdS, Mendoza SR, Goncalves DdS, Rodrıguez-de la Noval C, Honorato L, Nimrichter L, Ramos LFC, Nogueira FCS, Domont GB, Peralta JM and.
- Guimarães AJ. Recognition of Cell Wall Mannosylated Components as a Conserved Feature for Fungal Entrance, Adaptation and Survival Within Trophozoites of Acanthamoeba castellanii and Murine Macrophages. Frontiers in Cellular and Infection Microbiology. 2022; 12: 858979.
- Casadevall A, Fu MS, Guimaraes AJ, Albuquerque P. The 'Amoeboid Predator-Fungal Animal Virulence' Hypothesis. J Fungi (Basel). 2019; 21, 5(1):10.
- Guimaraes A. J., Gomes K. X., Cortines J. R., Peralta J. M., Peralta R. H. S.. Acanthamoeba Spp. As a Universal Host for Pathogenic Microorganisms: One Bridge From Environment to Host Virulence. Microbiol. Res. 2016;193, 30–38.
- Clarke M, Lohan AJ, Liu B, Lagkouvardos I, Roy S, Zafar N, Bertelli C, Schilde C, Kianianmomeni A, Bürglin TR, Frech C, Turcotte B, Kopec KO, Synnott JM, Choo C, Paponov I, Finkler A, Heng Tan CS, Hutchins AP, Weinmeier T, Rattei T, Chu JS, Gimenez G, Irimia M, Rigden DJ, Fitzpatrick DA, Lorenzo-Morales J, Bateman A, Chiu CH, Tang P, Hegemann P, Fromm H, Raoult D, Greub G, Miranda-Saavedra D, Chen N, Nash P, Ginger ML, Horn M, Schaap P, Caler L, Loftus BJ. Genome of Acanthamoeba castellanii highlights extensive lateral gene transfer and early evolution of tyrosine kinase signaling. Genome Biol. 2013; 1;14(2):R11.
- Rayamajhee, B., Willcox, M. D. P., Henriquez, F. L., Petsoglou, C., Subedi, D., and Carnt, N. (2022). Acanthamoeba, an environmental phagocyte enhancing survival and transmission of human pathogens. Trends Parasitol. 2022; 38, 975–990.
- Khan, N.A., 2009. Acanthamoeba Biology and Pathogenesis, Caister Academic Press, ISBN 978-1-904455-43-1.
- Steenbergen J. N., Shuman H. A., Casadevall A. (2001). Cryptococcus Neoformans Interactions with Amoebae Suggest an Explanation for Its Virulence and Intracellular Pathogenic Strategy in Macrophages. Proc. Natl. Acad. Sci. U.S.A. 2001; 98, 15245–15250.
- Segal G, Shuman HA. Legionella pneumophila utilizes the same genes to multiply within Acanthamoeba castellanii and human macrophages. Infect Immun 1999; 67:2117–2124.
- Al-Khodor S, Price CT, Habyarimana F, Kalia A, Abu Kwaik Y. A Dot/Icm-translocated ankyrin protein of Legionella pneumophila is required for intracellular proliferation within human macrophages and protozoa. Mol Microbiol. 2008; Nov;70(4):908-23.
- Derengowski L., da S., Paes H. C., Albuquerque P., Tavares A. H. F. P., Fernandes L., et al.. The Transcriptional Response of Cryptococcus Neoformans to Ingestion by Acanthamoeba Castellanii and Macrophages Provides Insights Into the Evolutionary Adaptation to the Mammalian Host. Eukaryot Cell. 2013; 12, 761–774.
- Espinoza-Vergara G, Hoque MM, McDougald D, Noorian P. The Impact of Protozoan Predation on the Pathogenicity of Vibrio cholerae. Front Microbiol. 2020; 21(11):17.
- Danelishvili, L, Wu, M., Stang, B., Harriff, M., Cirillo, S.L., Cirillo, J.D., Bildfell, R., Arbogast, B., Bermudez, L.E.. Identification of Mycobacterium avium pathogenicity island important for macrophage and amoeba infection. Proceedings of the National Academy of Science USA. 2007; 104 (26):1038–11043.
- Nikolakopoulou, C., Willment, J. A., Brown, G. D.. C-Type Lectin Receptors in Antifungal Immunity. Adv. Exp. Med. Biol. 2020; 1204, 1–30.
- Allen, PG, Dawidowicz, EA. Phagocytosis in Acanthamoeba: I. A mannose receptor is responsible for the binding and phagocytosis of yeast. J Cell Physiol. 1990; 145(3):508–513. [CrossRef]
- Corsaro D. Acanthamoeba Mannose and Laminin Binding Proteins Variation across Species and Genotypes. Microorganisms. 2022; 31;10(11):2162. [CrossRef]
- Merlen T, Sereno D, Brajon N, Rostand F, Lemesre JL. Leishmania spp: completely defined medium without serum and macromolecules (CDM/LP) for the continuous in vitro cultivation of infective promastigote forms. Am J Trop Med Hyg. 1999; 60:41–50.
- Costa AFP, de Brito RCF, Carvalho LM, Cardoso JMO, Vieira PMA, Reis AB, Aguiar-Soares RDO, Roatt BM. Liver infusion tryptose (LIT): the best choice for growth, viability, and infectivity of Leishmania infantum parasites. Parasitol Res. 2020; 119(12):4185-4195.
- Dos Santos A.L, Carvalho-Kelly, LF.,Dick, CF, Meyer-Fernandes, JR. Innate immunomodulation to trypanosomatid parasite infections. Exp. Parasitol. 2016;167: 67-75.
Figure 2.
Effect of media on the growth of Acanthamoeba castellanii.
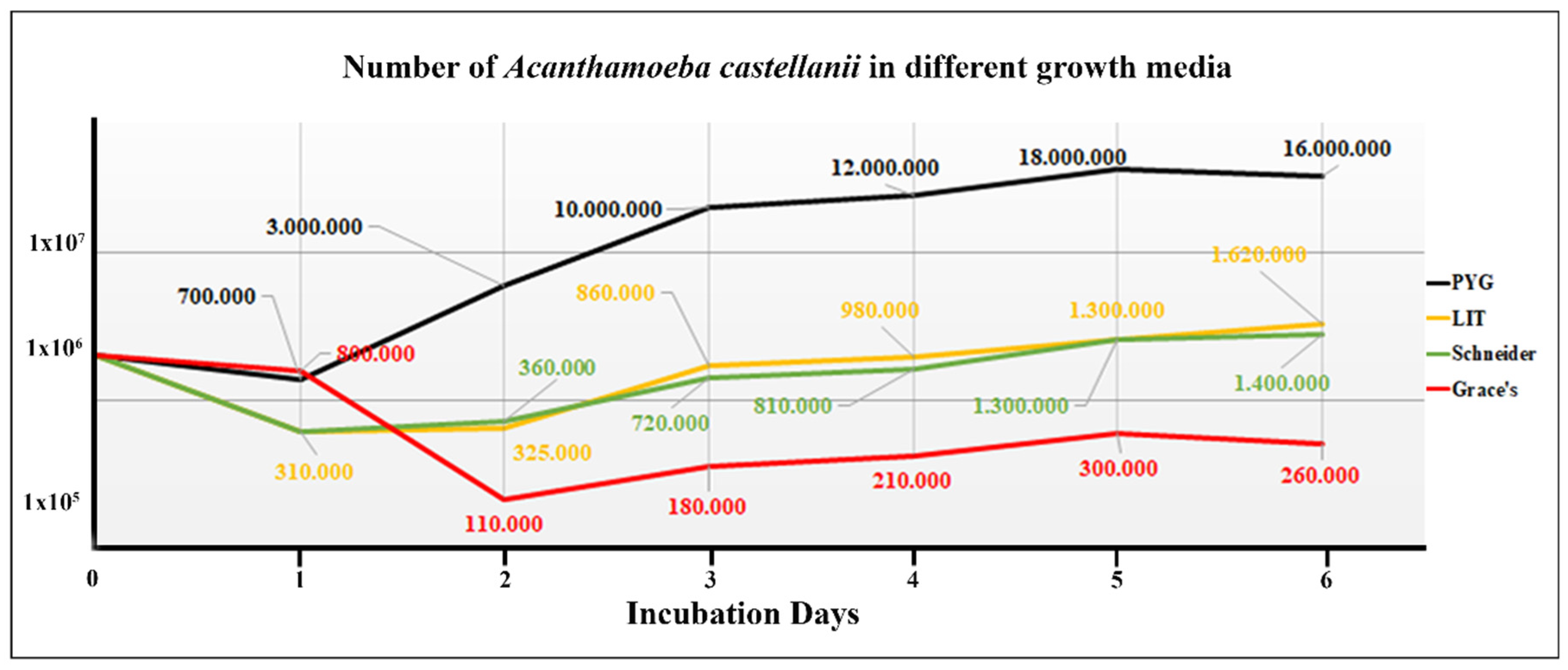
Figure 3.
Photomicrographs of Acanthamoeba castellanii infected with Leishmania spp. stained with Panoptic fast staining.
Figure 3.
Photomicrographs of Acanthamoeba castellanii infected with Leishmania spp. stained with Panoptic fast staining.
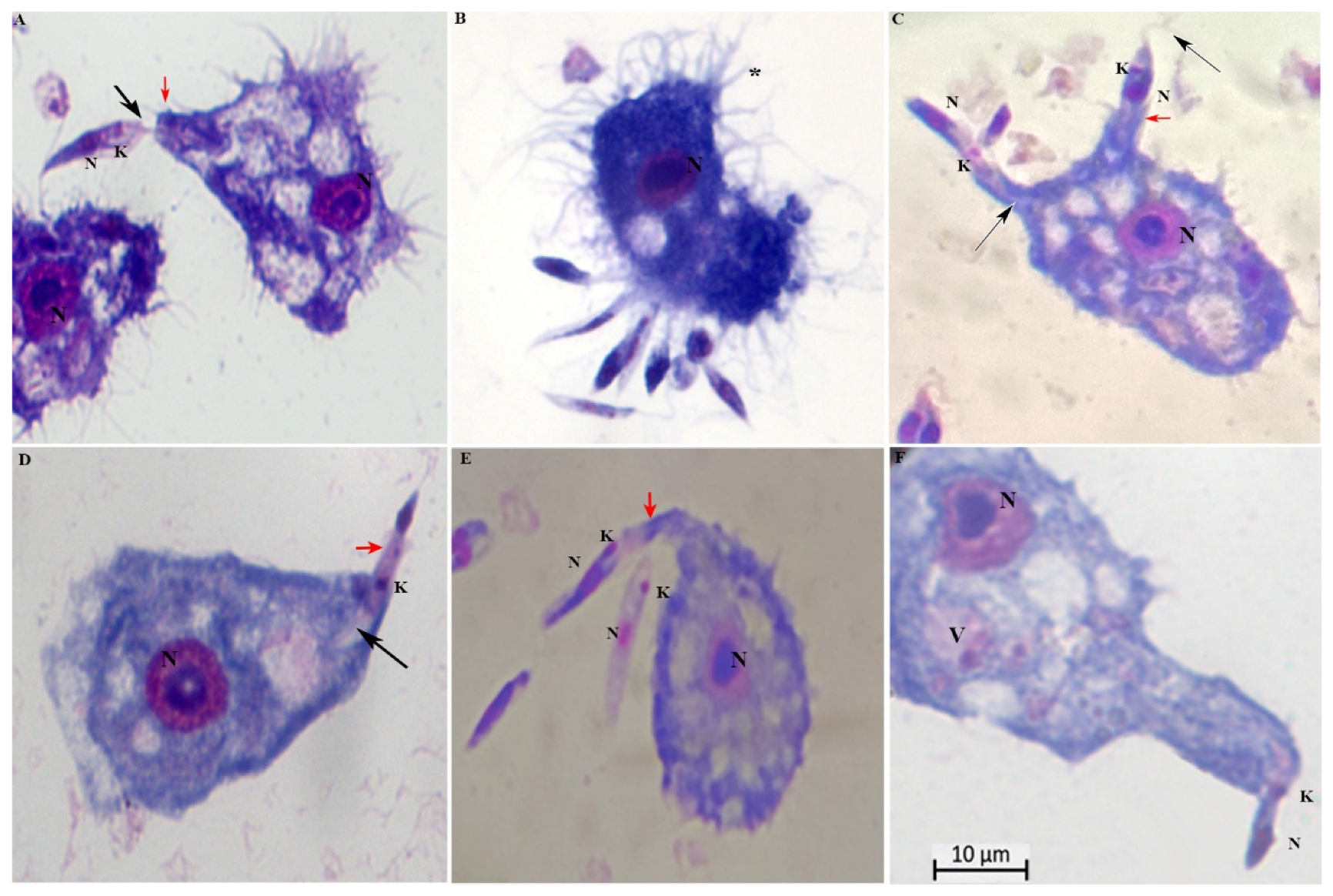
Figure 4.
Photomicrographs of Acanthamoeba castellanii infected with Leishmania spp., parasite ratio of :10 for 3 h, stained with Panoptic fast staining.
Figure 4.
Photomicrographs of Acanthamoeba castellanii infected with Leishmania spp., parasite ratio of :10 for 3 h, stained with Panoptic fast staining.
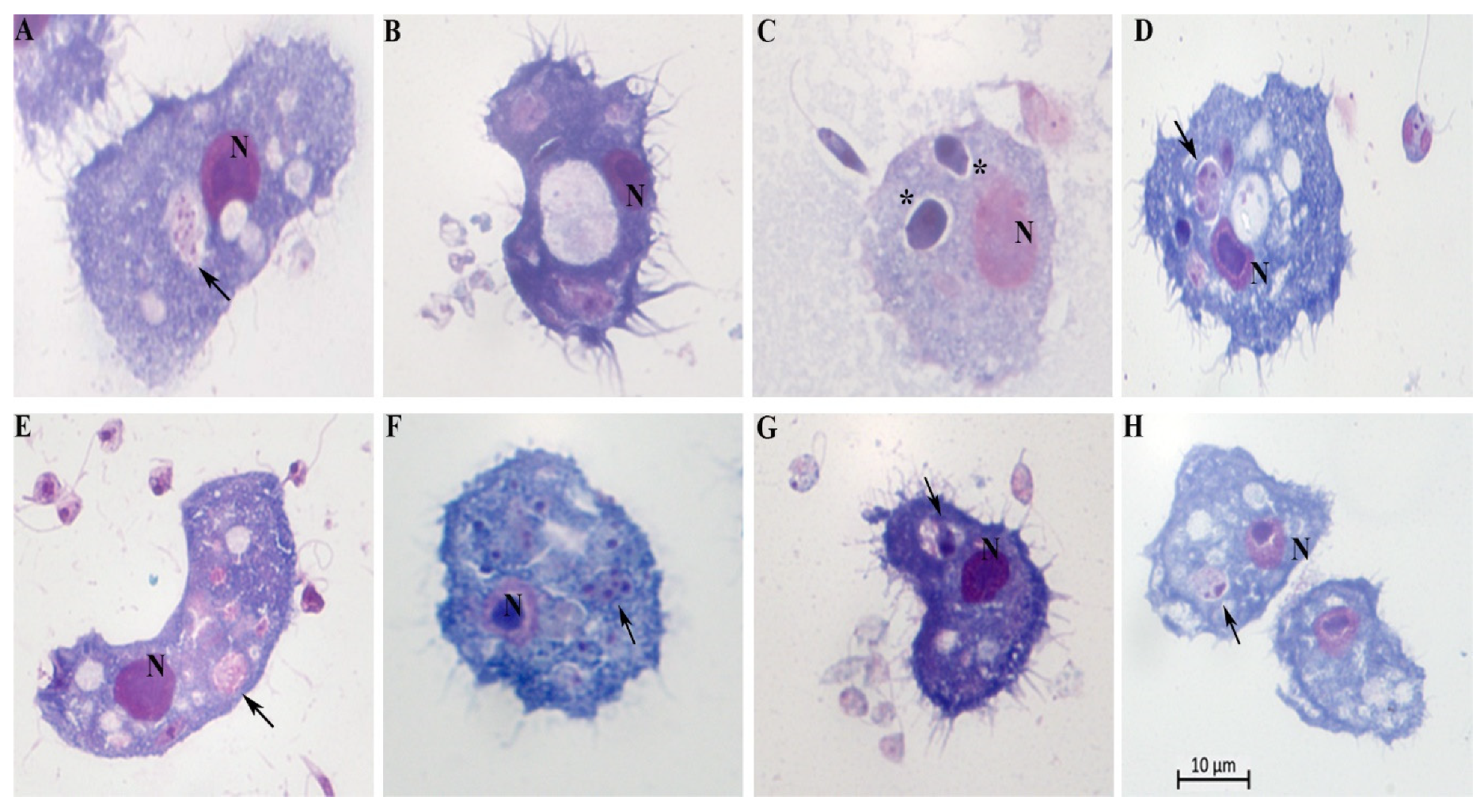
Figure 5.
Photomicrographs of Acanthamoeba castellanii infected with Leishmania spp., parasite ratio of 1:10 over different periods, stained with Panoptic fast staining.
Figure 5.
Photomicrographs of Acanthamoeba castellanii infected with Leishmania spp., parasite ratio of 1:10 over different periods, stained with Panoptic fast staining.
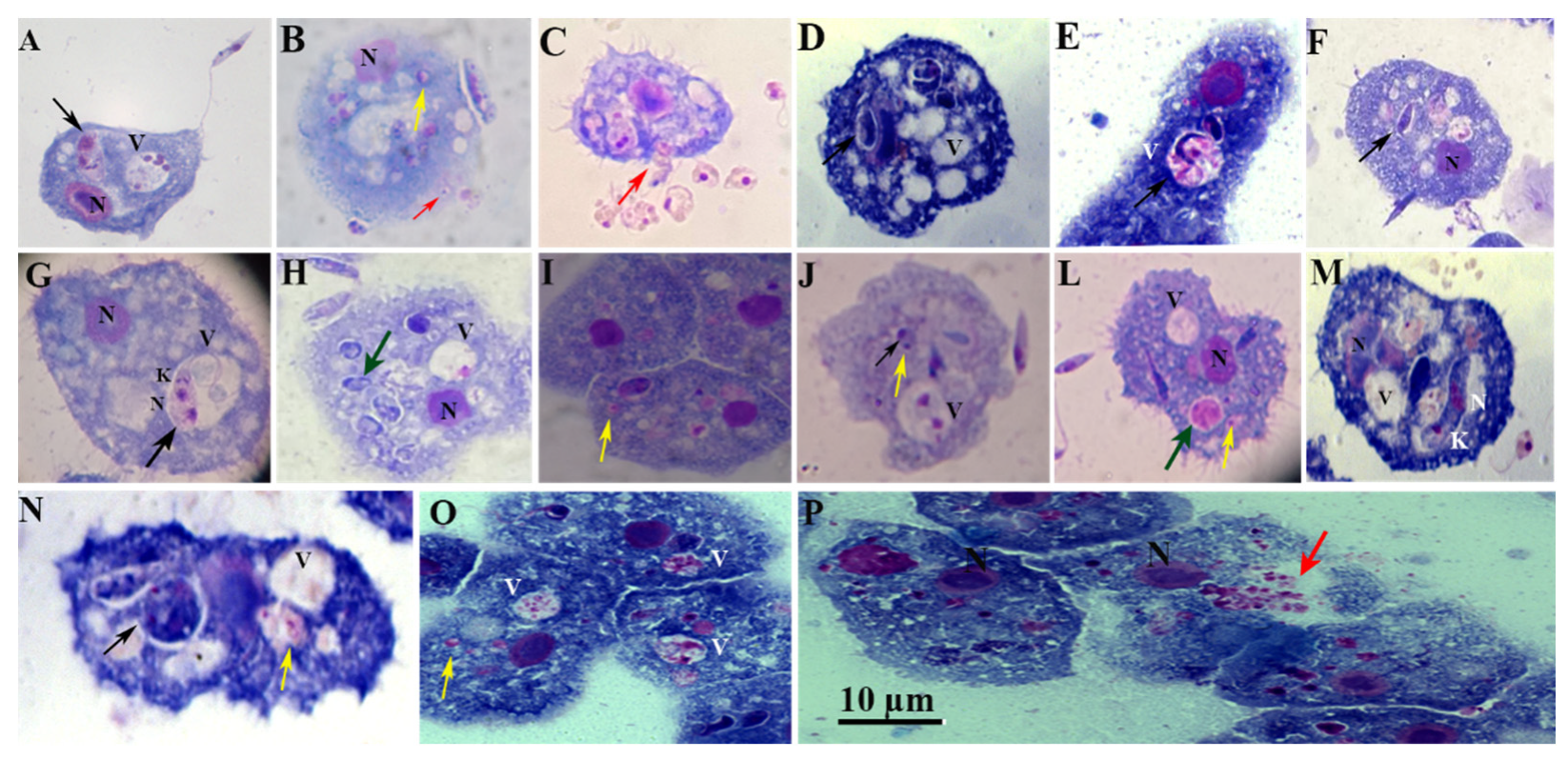
Figure 6.
Photomicrographs of Acanthamoeba castellanii infected with FITC-labeled promastigotes Leishmania spp., parasite ratio of 1:10 at 24 h of interactions.
Figure 6.
Photomicrographs of Acanthamoeba castellanii infected with FITC-labeled promastigotes Leishmania spp., parasite ratio of 1:10 at 24 h of interactions.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



