
Submitted:
06 July 2024
Posted:
08 July 2024
You are already at the latest version
Abstract
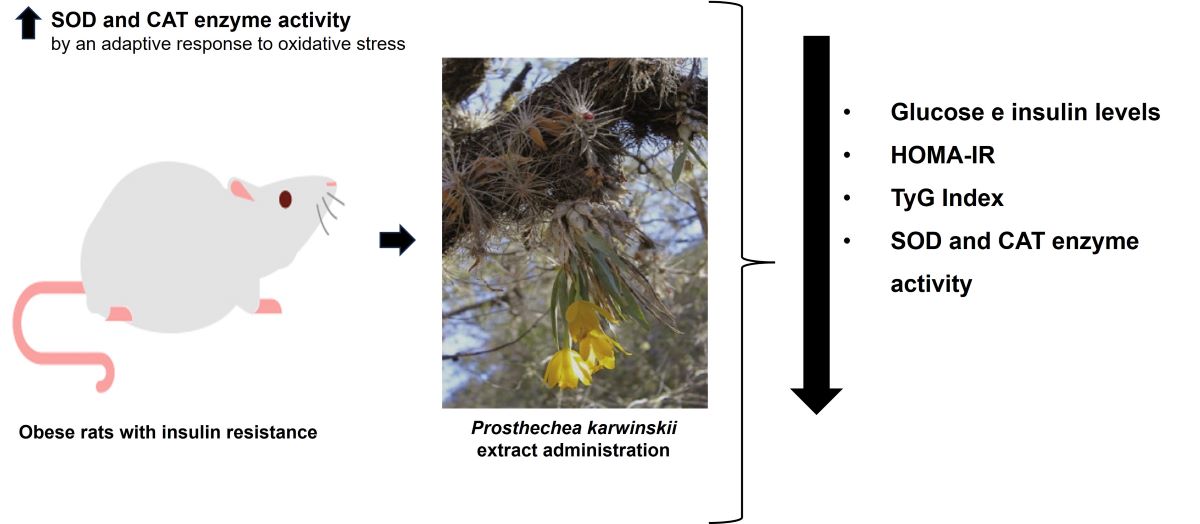
The present study evaluated the effect of Prosthechea karwinskii leaves extract on antioxidant enzymes superoxide dismutase (SOD) and catalase (CAT) in a model of obese rats with insulin resistance to know its nutraceutical potential on insulin resistance and oxidative stress. Obesity and insulin resistance were induced in rats by administering 40% sucrose in water. Once the model was established, four groups were evaluated (control, obese, obese administered with extract, and obese administered with metformin), and the indices of insulin resistance and the activity of antioxidant enzymes were measured. In addition, compounds in the extract were identified with UHPLC-ESI-qTOF-MS/MS. The glucose, insulin, triglycerides, and insulin resistance indicators (HOMA-IR and TyG) increased in obese rats. The activity of the antioxidant enzymes evaluated increased in the obese group, apparently as an adaptive response to oxidative stress to inhibit reactive oxygen species formed by hyperglycemia. Administration of P. karwinskii extract and metformin reduced glucose, insulin, triglyceride levels, insulin resistance indicators and antioxidant enzymes to values like those of the control group. This study shows the nutraceutical potential of P. karwinskii extract as an ingredient in the formulation of food supplements or functional foods to help treat diseases related to insulin resistance and oxidative stress.
Keywords:
reactive oxygen species
; HOMA-IR
; TyG index
; SOD
; CAT
1. Introduction
Insulin resistance (IR) is a metabolic disorder associated with obesity, where the sensitivity and cellular response to insulin in tissues is decreased, as well as glucose metabolism. IR is involved in the pathophysiology of metabolic disorders such as diabetes, dyslipidemia, non-alcoholic fatty liver disease, and cancer. Insulin resistance is characterized by elevated levels of reactive oxygen species (ROS), the origin of ROS being mainly mitochondrial, hyperglycemia can cause alterations of mitochondrial morphology and stimulate different mitochondrial enzymatic pathways, such as NADPH oxidase, NO synthase uncoupling, xanthine oxidase, and lipoxygenases, cyclooxygenases, and peroxidases. In turn, increased levels of oxidative stress with increased ROS production cause insulin resistance. Oxidative stress and insulin resistance are increased in obese individuals [1].
Oxidative stress participates in the pathogenesis of many chronic diseases, including IR and diabetes, and occurs due to an imbalance between an elevated ROS generation and/or poor antioxidant defense. Due to their antioxidant activity, medicinal plants are an alternative for the treatment of various diseases [2].
Prosthechea karwinskii (Mart.) J.M.H. Shaw is an endemic orchid of Mexico that is used in the Mixtec region of the state of Oaxaca to decorate church altars during Semana Santa celebrations, as well as to prepare various local dishes, and in traditional medicine [3]. The extract of its leaves decreases glucose levels [4] and IR [5] in rats with metabolic syndrome; and can inhibit ROS in an ex vivo model with peripheral blood mononuclear cells [6]. However, nothing has been reported on the effect of orchid extract on oxidative stress-related parameters in an in vivo model. Considering this background, this study aimed to evaluate the effect of P. karwinskii leaves extract on the antioxidant enzymes superoxide dismutase (SOD) and catalase (CAT) in a model of obese rats with insulin resistance to know its nutraceutical potential on insulin resistance and oxidative stress. In this way, it could be considered a potential ingredient in the formulation of food supplements and functional foods to help treat diseases related to insulin resistance and oxidative stress.
2. Materials and Methods
2.1. Materials, Reagents, and Kits
Commercial kits of Catalase assay Kit (707002, Cayman chemical), Superoxide Dismutase assay Kit (706002, Cayman chemical), Insulin assay Kit (589501, Cayman Chemical) was used, phosphate-buffered saline (PBS), distilled water, formic acid, acetonitrile, and other reagents were purchased from Sigma Aldrich Mexico, and the analytical kits for glucose and triglycerides were purchased from Spinreact Mexico.
2.2. Plant Material
The leaves of P. karwinskii used in this study were collected in 2017 in Zaachila Oaxaca (16º57' N latitude, 96°45' W longitude, 1490 m altitude). Species determination was performed by one of the authors (RS) and a backup specimen was deposited in the OAX herbarium (Solano 4037). In this study, Prosthechea is recognized as the accepted generic name of the orchid genus, in agreement with Pridgeon et al. [7], Soto et al. [8], Villaseñor [9], and Solano et al. [10].
2.3. Obtaining the Extract
The extract of P. karwinskii leaves was obtained by an ultrasound-assisted method following Barragan-Zarate et al. [6] (2020) in a 750 W ultrasound processor (VCX 750, Scientific SENSE). The solvent used was 50% ethanol in water with a sample: solvent ratio 1:18 (g: mL), and the extraction was performed at a temperature of 40°C for 20 min.
2.4. Compound Identification with UHPLC-ESI-qTOF-MS/MS
For the identification of plant compounds, the method described by Barragan-Zarate et al. [5] was followed. An ultra-high performance liquid chromatography system (UHPLC, Thermo Scientific, Ultimate 3000) combined with an Impact II mass spectrometer (Bruker) with electrospray ionization (ESI) and quadrupole time-of-flight (qTOF) was used. The column used was Thermo Scientific Acclaim 120 C18 (2.2 μm, 120 Å, 50 × 2.1 mm), and an elution gradient was used with mobile phases: A: 0.1% formic acid in water and B: acetonitrile, with a flow rate of 0.35 mL/min. The gradient system was 0% B (0–2 min), 1% B (2–3 min), 3% B (3–4 min), 32% B (4–5 min), 36% B (5–6 min), 40% B (6–8 min), 45% B (8–9 min), 80% B (9–11 min), 0% B (12–14). The mass spectrometer was operated in negative electrospray mode at 0.4 Bar (5.8 psi), in autoMSMS, with a mass range was 50–700 m/z. The data obtained were processed with DataAnalysis software. Compounds were identified by comparing their exact mass and fragmentation pattern with those collected from libraries and scientific articles.
2.5. Induction of Obesity
A total of 24 Wistar rats, 21 days old, were divided into 2 groups: the control group (CG, n=6) and the obese group (OG, n=18). The rats were fed ad libitum with standard food, the CG drank drinkable water, and the OG drank water with 40% sucrose to induce obesity, the induction period was 20 weeks as reported by Barragan-Zarate et al. [5]. The rats were maintained under normal conditions in the biotherium of the Escuela de Medicina from Universidad Cristóbal Colón, following the official Mexican standard for the care and handling of animals NOM-062-ZOO-1999 [11], as well as the international norms and standards for the care and use of laboratory animals.
2.6. Experimental Design
After the 20 weeks of obesity induction, the obese group (OG) was divided into 3 groups: OG received no treatment, PK was administered with P. karwinskii extract at a dose of 300 mg/kg, MET was administered with metformin at a dose of 200 mg/kg. The selection of the dose evaluated was based on a previous study [5], where the effective dose for controlling the metabolic syndrome parameters evaluated was 300 mg/kg. Treatments were administered with an oral feeding cannula. Rats in the OG, PK and MET groups continued to drink 40% sucrose water during the experimental diet. The comparison was made with the CG group (the group that did not induce obesity and did not receive any treatment). The experimental period was 4 weeks.
2.7. Parameters Evaluated
At the end of the experimental period, the rats were fasted for 12 hr and sacrificed. The weight of the rats was recorded, as well as the weight of the total adipose tissue. Blood was collected to determine glucose, insulin, triglycerides, and to calculate indicators of insulin resistance. The Homeostatic Model Assessment of Insulin Resistance (HOMA-IR) was calculated as described by Matthews et al. [12] with the following equation:
HOMA-IR = [fasting insulin (μIU/mL) × fasting glucose (mmol/L)]/ 22.5.
The triglyceride-glucose index (TyG) was calculated according to Simental-Mendía and Guerrero-Romero [13], with the following equation:
TyG = Ln [fasting triglycerides (mg/dL) × fasting glucose (mg/dL)]/ 2.
In addition, liver samples were taken and homogenized for the determination of the antioxidant enzymes SOD and CAT following the manufacturer's instructions.
2.8. Statistical Analysis
Results are reported as mean ± standard deviation (n=6). Analysis of variance (ANOVA) was performed, followed by Neuman's Keul test. Differences were considered statistically significant at P < 0.05.
3. Results
3.1. Compounds Identified with UHPLC-ESI-qTOF-MS/MS
Table 1 shows the information from the compounds identified in the extract of P. karwinskii with UHPLC-ESI-qTOF-MS/MS, which were quinic acid, malic acid, succinic acid, L-(-)-phenylalanine, guanosine, neochlorogenic acid, chlorogenic acid, rutin, kaempferol-3-O-ruthinoside, azelaic acid, pinelic acid, and embelin.
3.2. Effect of P. karwinskii Extract on Obesity, Insulin Resistance and Oxidative Stress Parameters
Table 2 shows the weight record of the rats at weeks 0 and 20 of induction (before the experimental diet), and at the end of the experiment, indices of insulin resistance at week 20, as well as the total adipose tissue of the rats at the end of the experimental diet, this information proves that the rats of the OG group were obese and presented insulin resistance before starting with the experimental diet.
Figure 1 shows the effect of P. karwinskii extract on glucose, insulin, and triglycerides, as well as on HOMA-IR and TyG insulin resistance indices. All these parameters evaluated presented higher values in the OG group compared to the CG. The administration of the extract, as well as metformin reduced glucose, triglycerides, and insulin levels compared to the OG group. As for IR, both indices showed a decrease in the PK and MET groups compared to the OG group. Table 3 shows the weekly liquid, and solid diet and caloric intake of each of the groups during the experimental diet period. Although there was no significant difference between the total caloric intake of the OG, PK, and MET groups at the end of week 4, there was a difference in the levels of glucose, triglycerides, insulin resistance indices, as well as body weight and adipose tissue, which makes evident the beneficial effect of the extract despite the same high sucrose diet.
Figure 2 shows the activity of the antioxidant enzymes SOD and CAT, which were increased in the OG group compared to the GC. However, in the PK and MET groups, the activity of these enzymes was downregulated to values that did not differ significantly from those of the GC.
4. Discussion
P. karwinskii leaves extract was able to decrease glucose levels and insulin resistance. The compounds identified in the extract that could be responsible for the effect of the extract on parameters related to glucose metabolism are chlorogenic acid [17] and embelin [18] which regulate glucose metabolism and decrease IR [17]; and rutin which decreases carbohydrate absorption, inhibits gluconeogenesis, increases tissue glucose uptake, increases insulin secretion by β-pancreatic cells and protects islets of Langerhans [19]. Given the relationship of IR with various metabolic disorders and the ability of P. karwinskii extract to attenuate it, it has the potential to help to treat diseases in which IR is implicated.
Hyperglycemia can cause altered mitochondrial morphology, increase cellular oxidation of glucose and production of NADH (Nicotinamide adenine dinucleotide, which contributes to oxidation in cellular processes), thereby increasing ROS generation [1]. ROS are produced in cells during normal aerobic metabolism, the generated ROS are removed by antioxidant enzymes, including SOD and CAT, which protect cells against oxidative stress by cellular detoxification of O2- and H2O2, excessive production of ROS and/or deficient antioxidant capacity can generate oxidative stress [2]. Hyperglycemia also activates different metabolic signaling pathways, triggering the activation of the antioxidant system to reduce ROS production and oxidative stress [1].
In the present study, the activity of the antioxidant enzymes evaluated increased in the obese group under oxidative stress compared to the control group. Although this behavior differs from the common behavior of antioxidant enzyme activity under oxidative stress, similar behavior has been reported in other investigations, as reported by Chen et al., [20] in their review on oxidative stress in nonalcoholic fatty liver disease. Or in the review by Barbosa et al. [21] on oxidative stress in hypertensive diseases of pregnancy, where, in some cases, an increase in the activity of the antioxidant enzymes CAT, SOD, and glutathione peroxidase was observed, suggesting a compensatory mechanism to the increase of ROS due to oxidative stress.
In the same way, the research of Zeng et al. [22], on the effect of cadmium stress on the antioxidant system of crabs, where showed that SOD activity increased in the cadmium-stressed group compared to the control group in both hepatopancreas and intestine, while in the case of CAT, its activity decreased in the Cd-stressed group in the hepatopancreas but increased in the intestine. So, they consider antioxidant enzymes as a non-specific adaptive mechanism that protects against oxidative damage and is the first line of antioxidant defense against oxidative stress.
According to Aouacheri et al. [23], whose results showed SOD activity in diabetic patients was increased compared to the control group, increased antioxidant enzyme activity stimulates the cellular ability to remove and limit damage caused by ROS. Acting as an adaptive response and a compensatory mechanism to detoxify harmful metabolites related to oxidative stress [23, 20]. Leghi et al. [24] also observed that SOD enzyme activity was increased in the group with Nonalcoholic steatohepatitis compared to those without, suggesting that the increased SOD activity was to counteract the overproduction of O2- radicals.
The increase in CAT activity could be in response to high SOD activity by the synergistic action of both enzymes [2], as it increases H2O2 production by SOD, which is subsequently catalyzed by CAT. The extract downregulates the activity of SOD and CAT enzymes that had increased in OG to values like those presented by CG, possibly through decrease of ROS increased by hyperglycemia and IR. In addition, a previous study had shown the ability of P. karwinskii leaves extract to inhibit ROS in cells [6]. This suggests that the compounds acted to reduce ROS, maintaining redox balance, without the physiological need to increase antioxidant enzyme activity. In other words, they can neutralize ROS and produce more stable radicals [25].
There are other investigations where the effect of the compounds evaluated have similarities to those reported in this research, such as that of Barbosa et al. [25], where they evaluated the effect of acai pulp in pregnant rats fed a high-fat diet, where the activity of antioxidant enzymes SOD and CAT were increased in the group under oxidative stress compared to the control group, and where similar to what happened with our extract, acai pulp modulated the activity of antioxidant enzymes and reduced the biomarkers of oxidative stress altered by the high-fat diet, thus improving the oxidative balance.
Something similar also occurred in okra plants, since when they were stressed with heavy metals such as cadmium, ROS production increased [26]. It was observed that the activity of antioxidant enzymes SOD and CAT increased in Cd-stressed plants compared to non-stressed and that receiving treatments with chelating agents such as malic acid and EDTA increased the resistance of plants to oxidative stress by decreasing H2O2 and the activity of SOD and CAT enzymes. Possibly something like what happens in plants occurred in our model, since according to Hussain et al. [27], they develop defensive mechanisms to minimize or protect themselves from the toxic effects of ROS, maintaining a balance between their synthesis and decomposition to preserve cellular redox hemostasis. Antioxidant enzyme activity is a key indicator of the antioxidant defense mechanism.
5. Conclusions
The activity of the antioxidant enzymes SOD and CAT was increased in the obese group as an adaptive response or compensatory mechanism to minimize the overproduction of ROS due to hyperglycemia and IR, which generated oxidative stress. Administration of the extract downregulated antioxidant enzyme activity to values like those of the control group, possibly by decreasing ROS formation by regulating glucose and IR levels, without having to increase antioxidant enzyme activity to reduce oxidative stress. This orchid has nutraceutical potential to treat conditions whose pathophysiology involves oxidative stress and/or insulin resistance and thus, can be considered a potential ingredient in the formulation of dietary supplements and functional foods to help treat diseases related to insulin resistance and oxidative stress.
Author Contributions
Conceptualization, G.S.B.Z., L.L.R. and A.A.A.; methodology, G.S.B.Z. and A.A.A.; writing—original draft preparation, G.S.B.Z.; writing—review and editing, L.L.R., A.A.A., R.S., and G.V.; visualization, G.S.B.Z., L.L.R., A.A.A., R.S., and G.V.; supervision, L.L.R., A.A.A., R.S., and G.V.; funding acquisition, L.L.R., A.A.A., and R.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by CONACYT (project 270428, postdoctoral grant 392827) and Instituto Politécnico Nacional (projects SIP-20161270, SIP-2016RE/50 and SIP-20220649).
Data Availability Statement
The original contributions presented in the study are included in the article, further inquiries can be directed to the corresponding author.
Acknowledgments
We thank the students who helped in the implementation of the animal model, and the organizers of the Semana Santa festivities in Zaachila Oaxaca for providing the plant material.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Andreadi, A.; Bellia, A.; Di Daniele, N.; Meloni, M.; Lauro, R.; Della-Morte, D.; Lauro, D. The Molecular Link between Oxidative Stress, Insulin Resistance, and Type 2 Diabetes: A Target for New Therapies against Cardiovascular Diseases. Curr. Opin. Pharmacol. 2022, 62, 85–96. [Google Scholar] [CrossRef] [PubMed]
- Hsouna, A. Ben; Dhibi, S.; Dhifi, W.; Saad, R. Ben; Brini, F.; Hfaidh, N.; Mnif, W. Essential Oil from Halophyte: Lobularia Maritima: Protective Effects against CCl4-Induced Hepatic Oxidative Damage in Rats and Inhibition of the Production of Proinflammatory Gene Expression by Lipopolysaccharide-Stimulated RAW 264.7 Macrophages. RSC Adv. 2019, 9, 36758–36770. [Google Scholar] [CrossRef] [PubMed]
- Cruz-Garcia, G.; Solano-Gomez, R.; Lagunez-Rivera, L. Documentation of the Medicinal Knowledge of Prosthechea Karwinskii in a Mixtec Community in Mexico. Rev. Bras. Farmacogn. 2014, 24, 153–158. [Google Scholar] [CrossRef]
- Rojas-Olivos, A.; Solano-Gómez, R.; Alexander-Aguilera, A.; Jiménez-Estrada, M.; Zilli-Hernández, S.; Lagunez-Rivera, L. Effect of Prosthechea Karwinskii (Orchidaceae) on Obesity and Dyslipidemia in Wistar Rats. Alex. J. Med. 2017. [CrossRef]
- Barragán-Zarate, G.S.; Alexander-Aguilera, A.; Lagunez-Rivera, L.; Solano, R.; Soto-Rodríguez, I. Bioactive Compounds from Prosthechea Karwinskii Decrease Obesity, Insulin Resistance, pro-Inflammatory Status, and Cardiovascular Risk in Wistar Rats with Metabolic Syndrome. J. Ethnopharmacol. 2021, 279. [Google Scholar] [CrossRef] [PubMed]
- Barragán-Zarate, G.S.; Lagunez-Rivera, L.; Solano, R.; Pineda-Peña, E.A.; Landa-Juárez, A.Y.; Chávez-Piña, A.E.; Carranza-Álvarez, C.; Hernández-Benavides, D.M. Prosthechea Karwinskii, an Orchid Used as Traditional Medicine, Exerts Anti-Inflammatory Activity and Inhibits ROS. J. Ethnopharmacol. 2020, 253. [Google Scholar] [CrossRef] [PubMed]
- Pridgeon, A. M., Cribb, P. J., Chase, M. W., Rasmussen, F. N. Genera Orchidacearum Volume 4: Epidendroideae (Part One). Oxford University Press, New York 2005.
- Soto, M. A., Hágsater, E., Jiménez, R., Salazar, G. A., Solano, R., Flores, R., Ruiz, I. Orchids of Mexico. Digital Catalogue. Herbario AMO, México City 2007.
- Villaseñor, J.L. Checklist of the native vascular plants of Mexico. Rev. Mex. De Biodivers. 2016, 87, 559–902. [Google Scholar] [CrossRef]
- Solano, R., Salazar-Chávez, G., Jiménez-Machorro, R., Hagsater, E., Cruz-García, G. Actualización del catálogo de autoridades taxonómicas de Orchidaceae de México. Instituto Politécnico Nacional. Centro Interdisciplinario de Investigación para el Desarrollo Integral Regional Unidad Oaxaca. Data base SNIB-CONABIO, Project KT005 (Mexico City) 2020.
- SAGARPA (Secretaría de Agricultura, Ganadería, Desarrollo Rural, Pesca y Alimentación), NORMA Oficial Mexicana NOM-062-ZOO-1999. Especificaciones técnicas para la producción, cuidado y uso de los animales de laboratorio 2001. Diario Oficial de la Federación. (accessed 13 March 2023).
- Matthews, D.R.; Hosker, J.P.; Rudenski, A.S.; Naylor, B.A.; Treacher, D.F.; Turner, R.C. Homeostasis Model Assessment: Insulin Resistance and β-Cell Function from Fasting Plasma Glucose and Insulin Concentrations in Man. Diabetologia 1985, 28, 412–419. [Google Scholar] [CrossRef] [PubMed]
- Simental-Mendía, L.E.; Guerrero-Romero, F. The Correct Formula for the Triglycerides and Glucose Index. Eur. J. Pediatr. 2020, 179, 1171. [Google Scholar] [CrossRef] [PubMed]
- De Souza, M.P.; Bataglion, G.A.; da Silva, F.M.A.; de Almeida, R.A.; Paz, W.H.P.; Nobre, T.A.; Marinho, J.V.N.; Salvador, M.J.; Fidelis, C.H.V.; Acho, L.D.R.; et al. Phenolic and Aroma Compositions of Pitomba Fruit (Talisia Esculenta Radlk.) Assessed by LC-MS/MS and HS-SPME/GC-MS. Food Res. Int. 2016, 83, 87–94. [Google Scholar] [CrossRef]
- Ma, T.; Lin, J.; Gan, A.; Sun, Y.; Sun, Y.; Wang, M.; Wan, M.; Yan, T.; Jia, Y. Qualitative and Quantitative Analysis of the Components in Flowers of Hemerocallis Citrina Baroni by UHPLC–Q-TOF-MS/MS and UHPLC–QQQ-MS/MS and Evaluation of Their Antioxidant Activities. J. Food Compos. Anal. 2023, 120, 105329. [Google Scholar] [CrossRef]
- Dahibhate, N.L.; Dwivedi, P.; Kumar, K. GC–MS and UHPLC-HRMS Based Metabolite Profiling of Bruguiera Gymnorhiza Reveals Key Bioactive Compounds. South Afr. J. Bot. 2022, 149, 1044–1048. [Google Scholar] [CrossRef]
- Ma, Y.; Gao, M.; Liu, D. Chlorogenic Acid Improves High Fat Diet-Induced Hepatic Steatosis and Insulin Resistance in Mice. Pharm. Res. 2015, 32, 1200–1209. [Google Scholar] [CrossRef] [PubMed]
- Durg, S.; Veerapur, V.P.; Neelima, S.; Dhadde, S.B. Antidiabetic Activity of Embelia Ribes, Embelin and Its Derivatives: A Systematic Review and Meta-Analysis. Biomed. Pharmacother. 2017, 86, 195–204. [Google Scholar] [CrossRef]
- Ghorbani, A. Biomedicine & Pharmacotherapy Mechanisms of Antidiabetic efects of Flavonoid Rutin. Biomed. Pharmacother. J. 2017, 96, 305–312. [Google Scholar] [CrossRef]
- Chen, Z.; Tian, R.; She, Z.; Cai, J.; Li, H. Role of Oxidative Stress in the Pathogenesis of Nonalcoholic Fatty Liver Disease. Free. Radic. Biol. Med. 2020, 152, 116–141. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, P.O.; Tanus-santos, J.E.; Cavalli, R.D.C.; Bengtsson, T.; Barbosa, P.O.; Tanus-santos, J.E.; Cavalli, R.D.C.; Bengtsson, T. The Nitrate-Nitrite-Nitric Oxide Pathway: Potential Role in Mitigating Oxidative Stress in Hypertensive Disorders of Pregnancy The Nitrate-Nitrite-Nitric Oxide Pathway: Potential Role in Mitigating Oxidative Stress in Hypertensive Disorders of Pregnancy. 2024. [CrossRef]
- Zeng, L.; Wang, Y.H.; Ai, C.X.; Zhang, B.; Zhang, H.; Liu, Z.M.; Yu, M.H.; Hu, B. Differential Effects of Oxytetracycline on Detoxification and Antioxidant Defense in the Hepatopancreas and Intestine of Chinese Mitten Crab under Cadmium Stress. Sci. Total Environ. 2024, 930, 172633. [Google Scholar] [CrossRef] [PubMed]
- Aouacheri, O.; Saka, S.; Krim, M.; Messaadia, A.; Maidi, I. The Investigation of the Oxidative Stress-Related Parameters in Type2 Diabetes Mellitus. Can. J. Diabetes 2015, 39, 44–49. [Google Scholar] [CrossRef] [PubMed]
- Leghi, G.E.; Domenici, F.A.; Vannucchi, H. Influence of Oxidative Stress and Obesity in Patients with Nonalcoholic Steatohepatitis. Arq. De Gastroenterol. 2015, 52, 228–233. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, P.O.; Souza, M.O.; Silva, M.P.S.; Santos, G.T.; Silva, M.E.; Bermano, G.; Freitas, R.N. Açaí (Euterpe Oleracea Martius) Supplementation Improves Oxidative Stress Biomarkers in Liver Tissue of Dams Fed a High-Fat Diet and Increases Antioxidant Enzymes’ Gene Expression in Offspring. Biomed. Pharmacother. 2021, 139, 111627. [Google Scholar] [CrossRef] [PubMed]
- Mousavi, A.; Pourakbar, L.; Siavash Moghaddam, S. Effects of Malic Acid and EDTA on Oxidative Stress and Antioxidant Enzymes of Okra (Abelmoschus Esculentus L.) Exposed to Cadmium Stress. Ecotoxicol. Environ. Saf. 2022, 248, 114320. [Google Scholar] [CrossRef] [PubMed]
- Hussain, I.; Siddique, A.; Ashraf, M.A.; Rasheed, R.; Ibrahim, M.; Iqbal, M.; Akbar, S.; Imran, M. Does Exogenous Application of Ascorbic Acid Modulate Growth, Photosynthetic Pigments and Oxidative Defense in Okra (Abelmoschus Esculentus (L.) Moench) under Lead Stress? Acta Physiol. Plant. 2017, 39, 1–13. [Google Scholar] [CrossRef]
- Londero, É.P.; Bressan, C.A.; Pês, T.S.; Saccol, E.M.H.; Baldisserotto, B.; Finamor, I.A.; Pavanato, M.A. Rutin-Added Diet Protects Silver Catfish Liver against Oxytetracycline-Induced Oxidative Stress and Apoptosis. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2021, 239, 108848. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Tang, C.; Zhang, H. Hepatoprotective Effects of Kaempferol 3-O-Rutinoside and Kaempferol 3-O-Glucoside from Carthamus Tinctorius L. on CCl4-Induced Oxidative Liver Injury in Mice. J. Food Drug Anal. 2015, 23, 310–317. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.; Sharma, A.K.; Sharma, M.C.; Gupta, R.S. Antioxidant Activity and Protection of Pancreatic b -Cells by Embelin in Streptozotocin-Induced Diabetes. J. Diabetes 2012, 4, 248–256. [Google Scholar] [CrossRef] [PubMed]
- Nonose, Y.; Pieper, L.Z.; da Silva, J.S.; Longoni, A.; Apel, R. V.; Meira-Martins, L.A.; Grings, M.; Leipnitz, G.; Souza, D.O.; de Assis, A.M. Guanosine Enhances Glutamate Uptake and Oxidation, Preventing Oxidative Stress in Mouse Hippocampal Slices Submitted to High Glutamate Levels. Brain Res. 2020, 1748, 147080. [Google Scholar] [CrossRef]
Figure 1.
Glucose A), insulin B) and C) triglycerides levels, and HOMA-IR D) and TyG E) insulin resistance indices. Values expressed as mean ± sd. *Indicates significant difference (P˂0.05) concerning CG. **Indicates significant difference (P˂0.05) concerning OG. CG (control group), OG (obese rats), PK (obese rats that received extract), MET (obese rats that received metformin).
Figure 1.
Glucose A), insulin B) and C) triglycerides levels, and HOMA-IR D) and TyG E) insulin resistance indices. Values expressed as mean ± sd. *Indicates significant difference (P˂0.05) concerning CG. **Indicates significant difference (P˂0.05) concerning OG. CG (control group), OG (obese rats), PK (obese rats that received extract), MET (obese rats that received metformin).
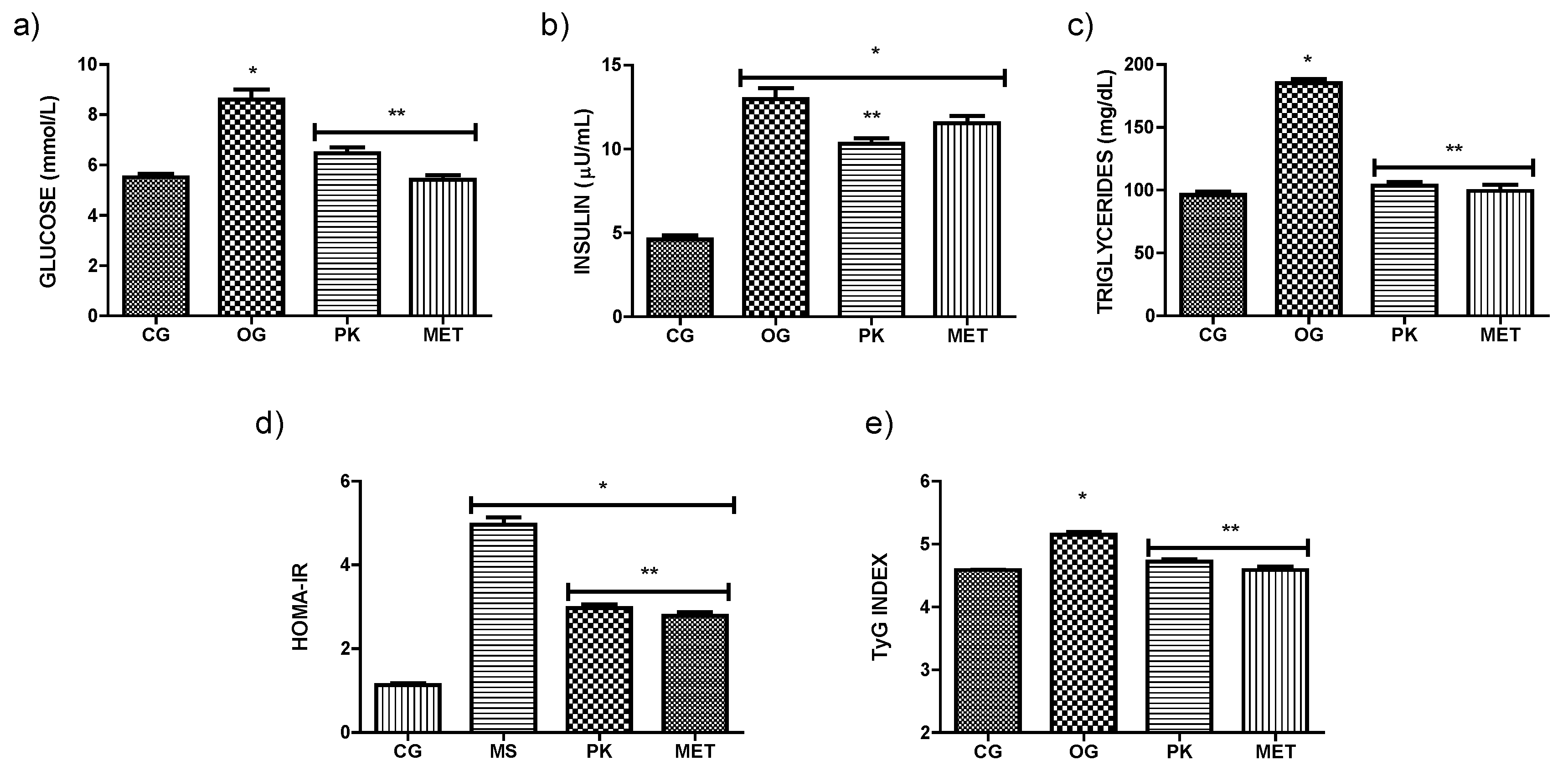
Figure 2.
Superoxide dismutase enzyme activity (SOD) A) and Catalase enzyme activity (CAT) B). Values expressed as mean ± sd. *Indicates significant difference (P˂0.05) with respect to GC. **Indicates significant difference (P˂0.05) with respect to OG. CG (control group), OG (obese rats), PK (obese rats that received extract), MET (obese rats that received metformin).
Figure 2.
Superoxide dismutase enzyme activity (SOD) A) and Catalase enzyme activity (CAT) B). Values expressed as mean ± sd. *Indicates significant difference (P˂0.05) with respect to GC. **Indicates significant difference (P˂0.05) with respect to OG. CG (control group), OG (obese rats), PK (obese rats that received extract), MET (obese rats that received metformin).
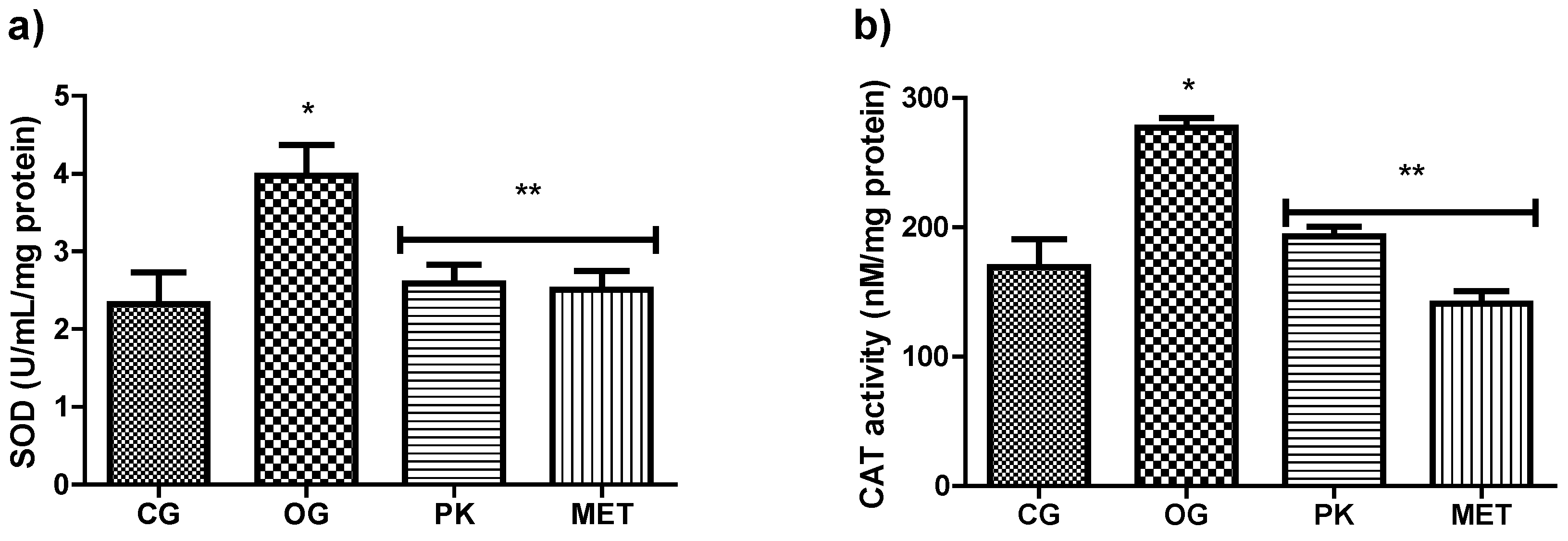

Table 1.
Chemical structures and information of compounds identified in leaves extract of Prosthechea karwinskii with UPLC-ESI-qTOF-MS/MS.
Table 1.
Chemical structures and information of compounds identified in leaves extract of Prosthechea karwinskii with UPLC-ESI-qTOF-MS/MS.
| Peak | RT (min) | m/z [M-H]- |
Error (ppm) | MS/MS fragments | Compound (Chemical formula) |
Type of compound | Relative yield (%) | Chemical structure |
|---|---|---|---|---|---|---|---|---|
| 1 | 0.7 | 191.0557 | 1.9 | 85.0293, 87.0078, 111.0443, 127.6945 | Quinic Acidbd (C7H12O6) |
Cyclitol, cyclic polyol, and cyclohexanecarboxylic acid | 22.21 | 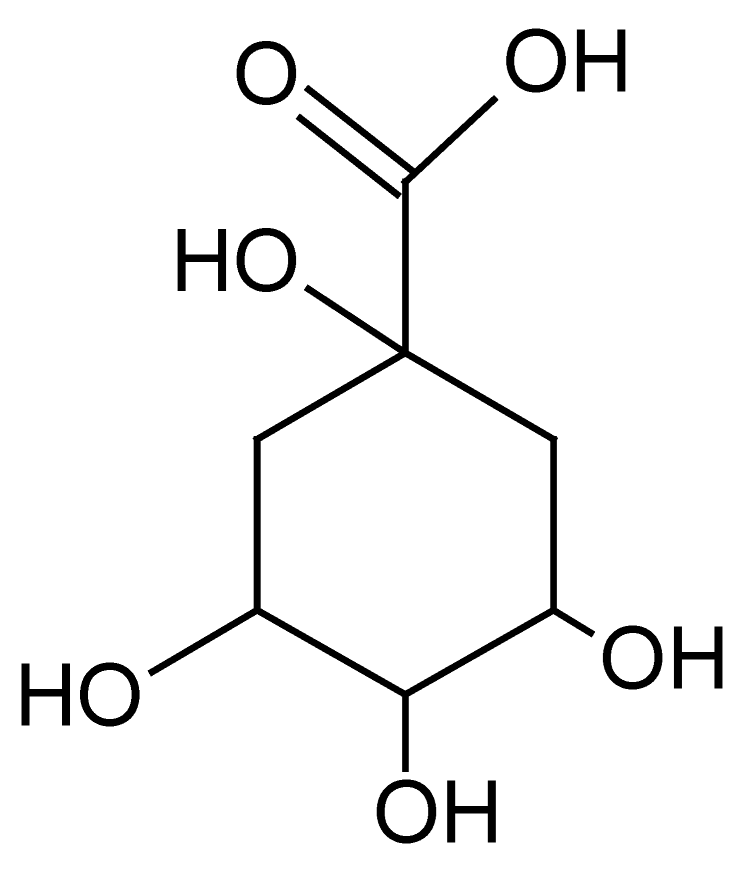 |
| 2 | 0.8 | 133.0140 | 1.8 | 115.0032 | Malic acidd (C4H6O5) |
Dicarboxylic organic acid | 5.71 | 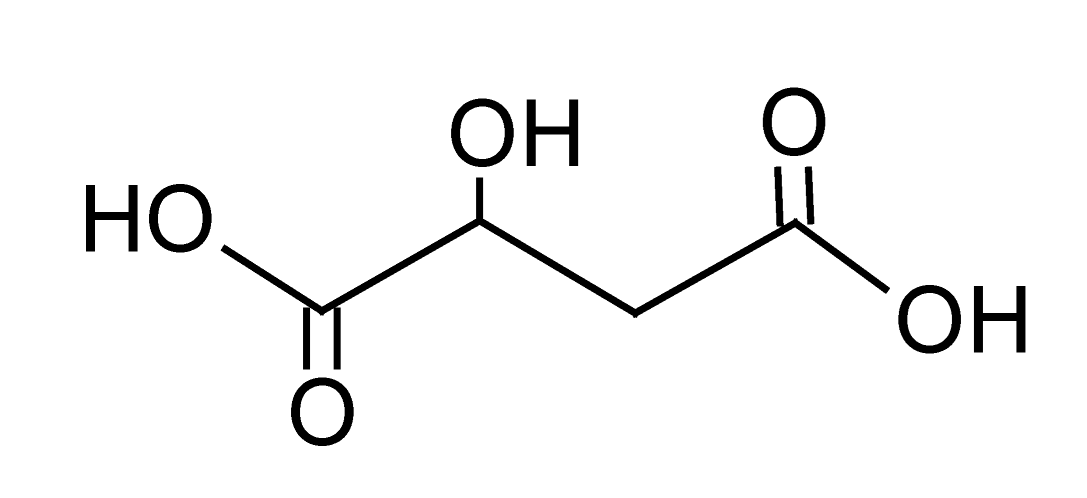 |
| 3 | 1.2 | 117.0191 | 2.3 | 73.0290, 99.0072 | Succinic acidd (C4H604) |
Dicarboxylic organic acid | 1.89 | 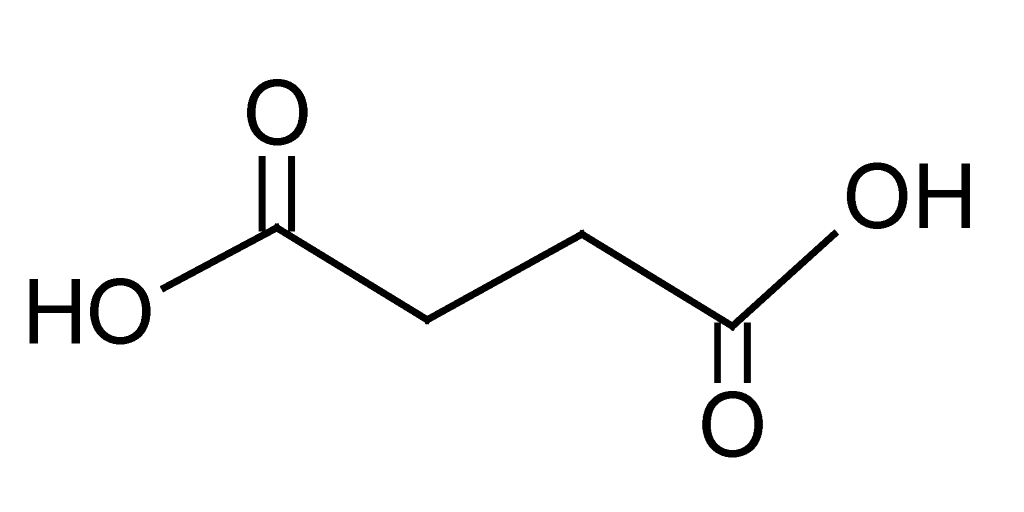 |
| 4 | 2.3 | 164.0712 | 3.2 | 72.0072, 103.0539, 147.0442 | L-(-)-Phenylalaninee (C9H11NO2) |
Amino acid | 0.53 | 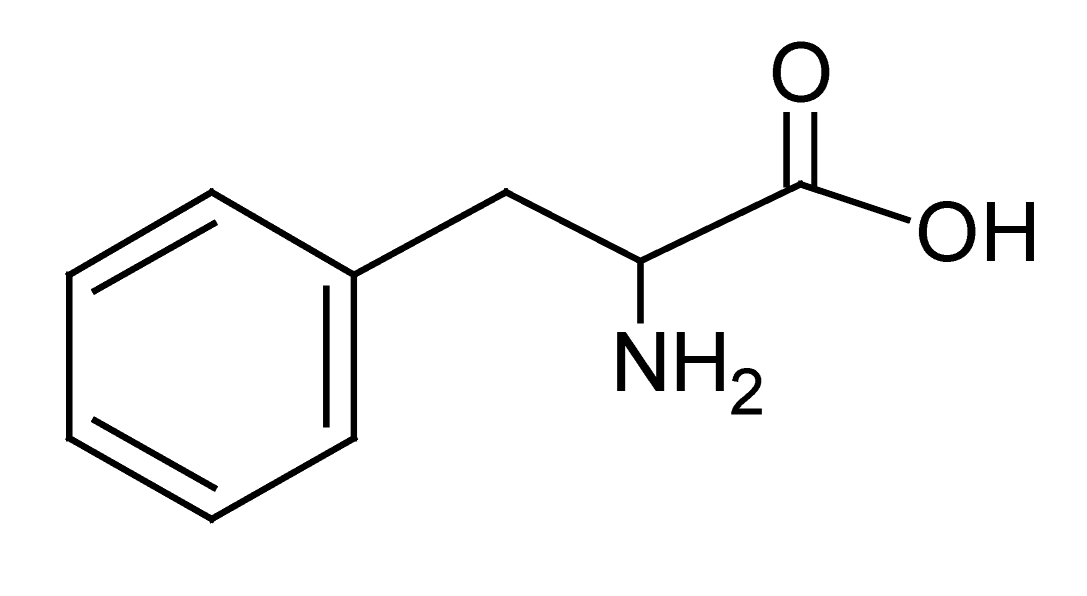 |
| 5 | 2.7 | 282.0833 | 3.9 | 108.5347, 133.0157, 150.0429 | Guanosinee (C10H13N5O5) |
Nucleoside | 0.46 | 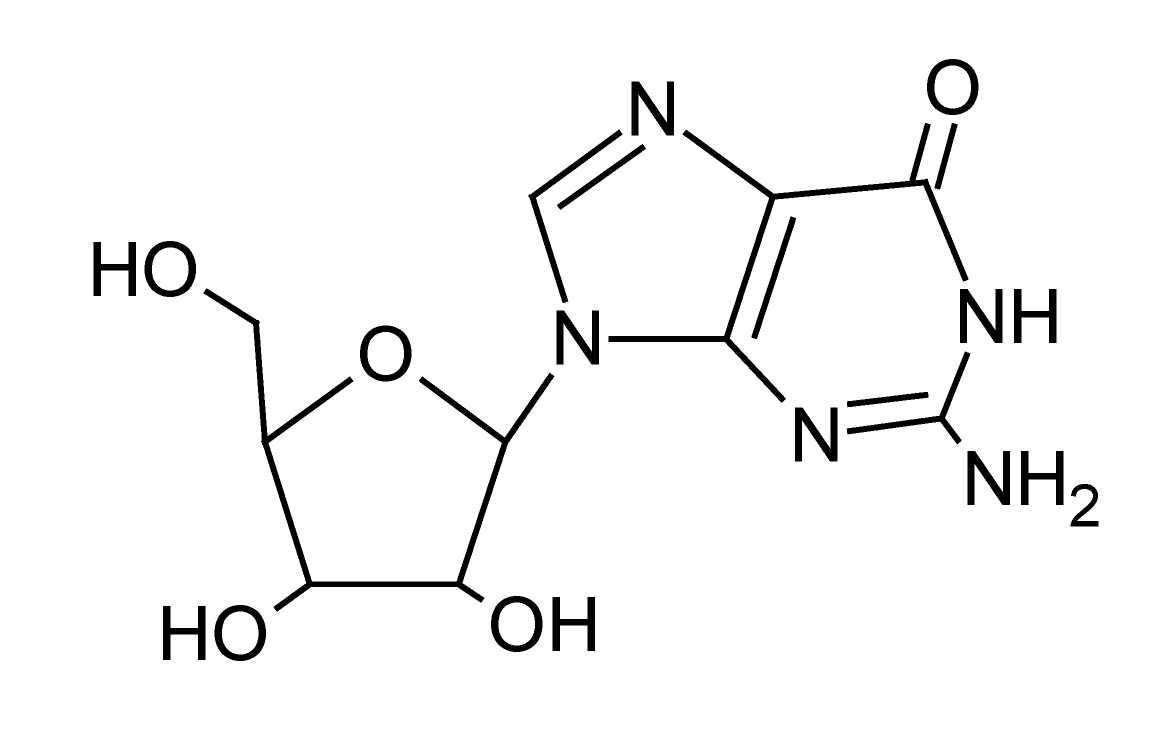 |
| 6 | 6.0 | 353.0867 | 3.1 | 179.0365, 191.0556 | Neochlorogenic acidb (C16H18O9) |
Caffeoylquinic acid, phenolic compound | 4.35 | 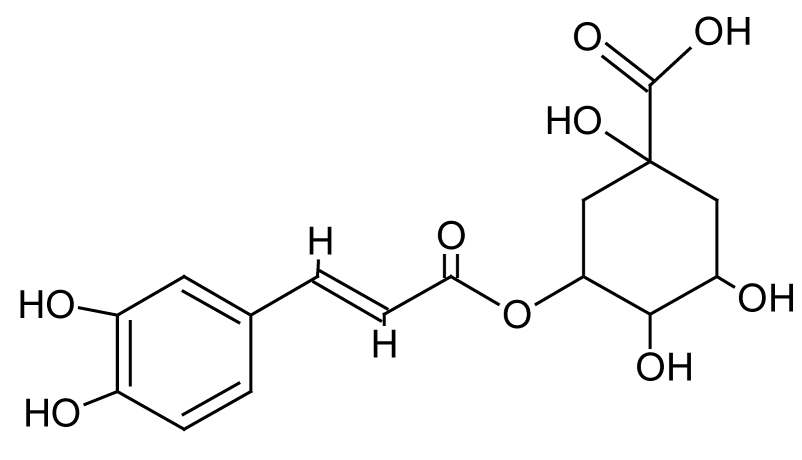 |
| 7 | 6.3 | 353.0866 | 3.4 | 173.0452, 179.0365, 191.0556 | Chlorogenic acida (C16H18O9) |
Caffeoylquinic acid, phenolic compound | 8.44 | 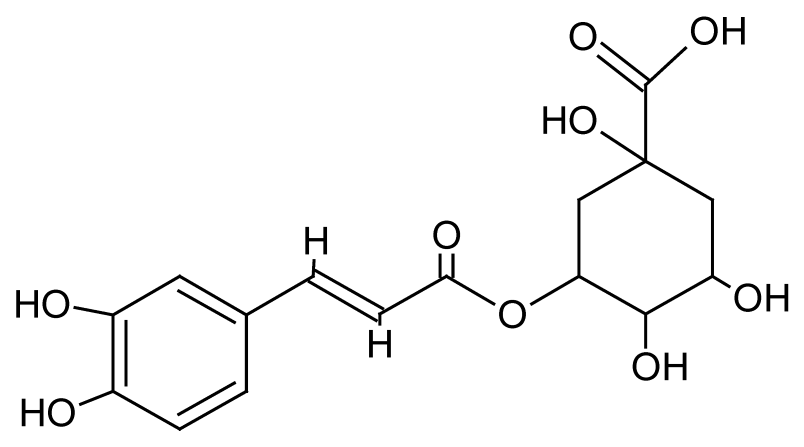 |
| 8 | 6.5 | 609.1438 | 3.8 | 300.0266, 301.0335 | Rutinabe (C27H30O16) |
Flavonoid glycoside | 15.52 | 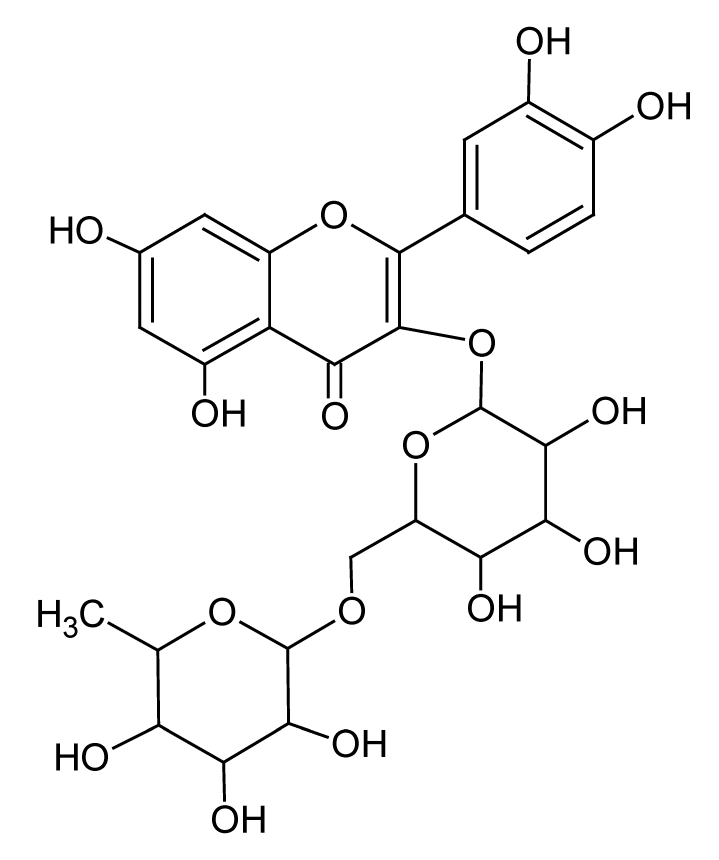 |
| 9 | 6.6 | 593.1489 | 3.9 | 284.0314, 285.0393 | Kaempferol-3-O-rutinosidee (C27H30O15) |
Flavonol glycoside | 25.30 | 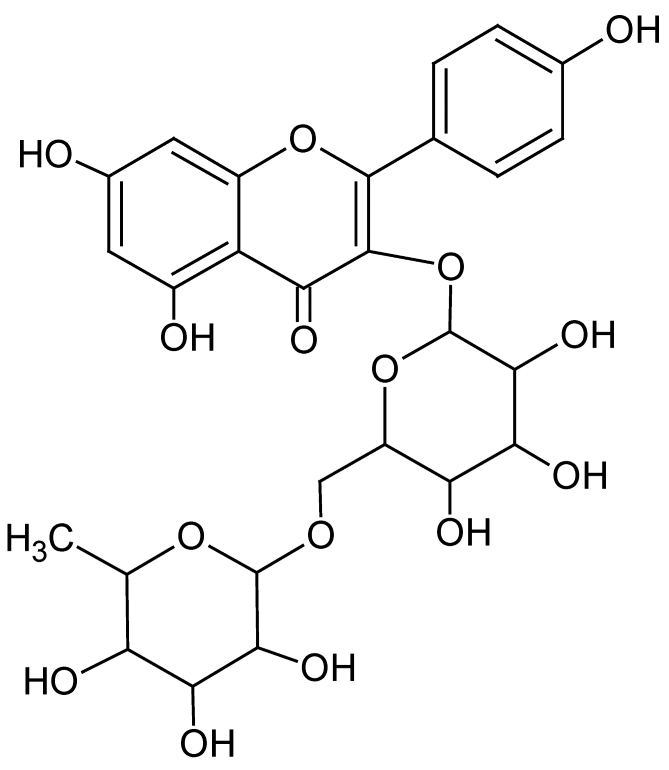 |
| 10 | 6.8 | 187.0970 | 3.1 | 97.0653, 125.0963, 169.0889 | Azelaic acidc (C9H1604) |
Dicarboxylic acid | 2.10 | 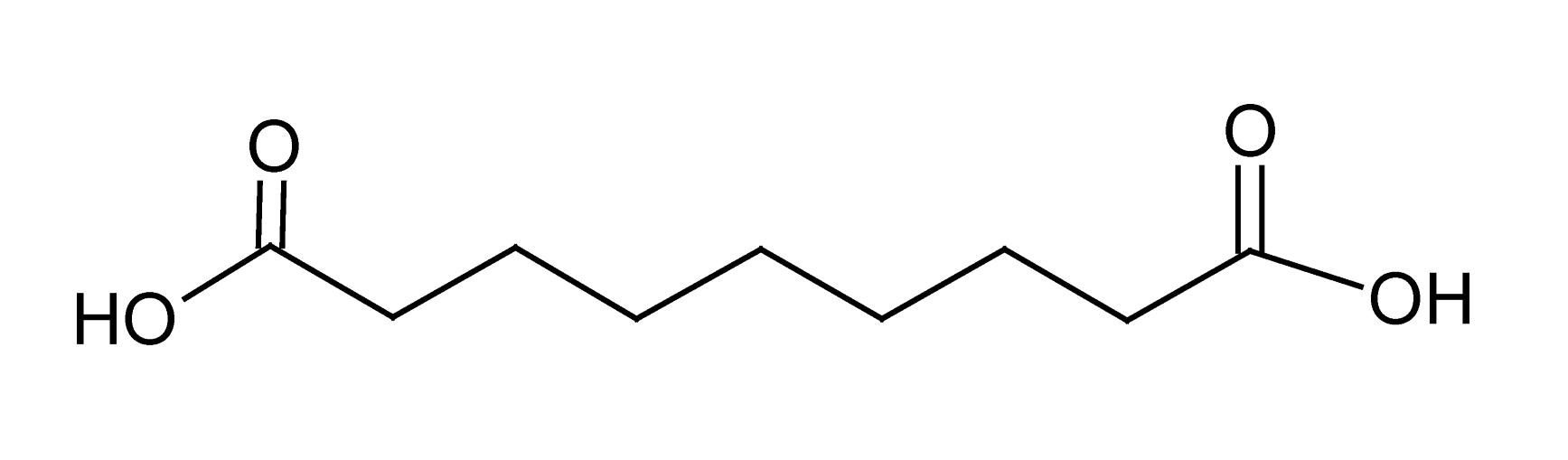 |
| 11 | 8.2 | 329.2321 | 3.9 | 171.1023, 229.1436 | Pinellic acidc (C18H34O5) |
Carboxylic acid | 1.09 | 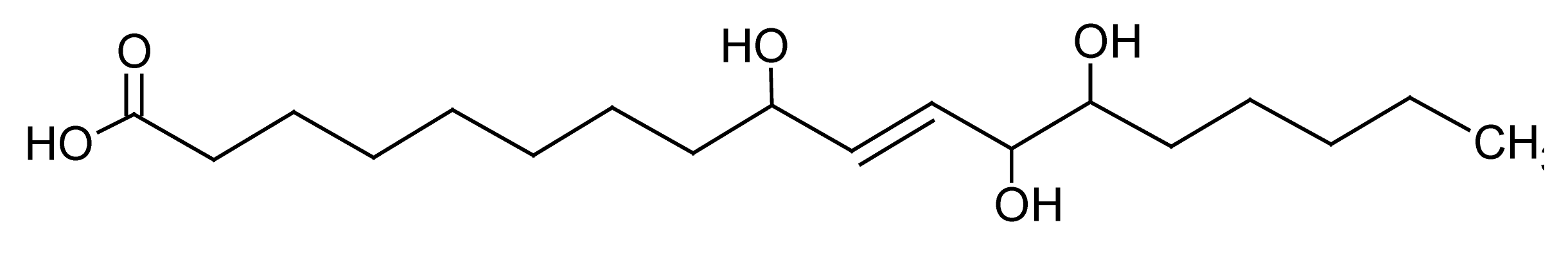 |
| 12 | 11.5 | 293.2112 | 3.6 | 275.2013, 235.1680, 223.1685 | Embelinc (C17H26O4) |
Para-benzoquinone | 8.60 | 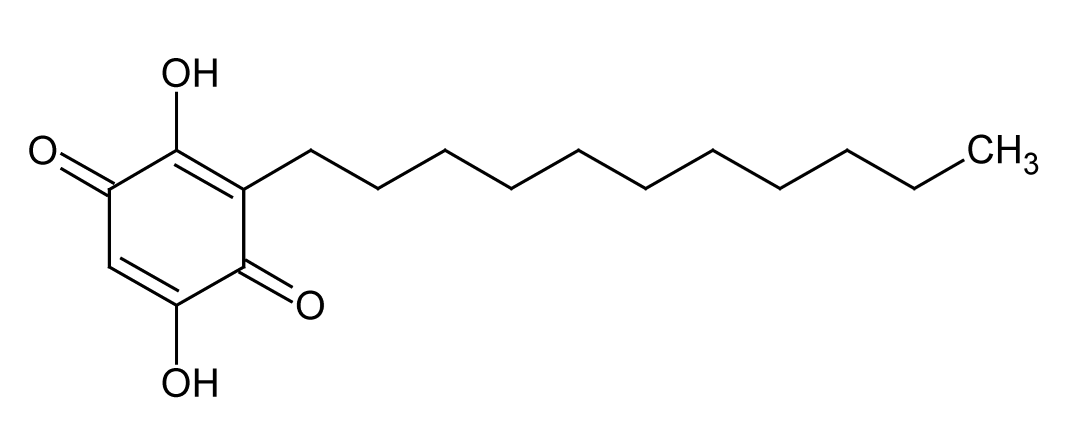 |
Table 2.
Body weight record in Wistar rats at week 0, 20 of obesity induction, and the end of the experiment, and total adipose tissue.
Table 2.
Body weight record in Wistar rats at week 0, 20 of obesity induction, and the end of the experiment, and total adipose tissue.
| CG | OG | PK | MET | |
|---|---|---|---|---|
| Weight at week 0 (g) | 74.3 ± 17.9 | 78.2 ± 15.5 | - | - |
| Weight at week 20 (g) | 368.5 ± 38.1 | 456.2 ± 42.6* | - | - |
| TyG index at week 20 | 7.73 ± 0.48 |
8.85 ± 0.53* |
- | - |
| HOMA-IR at week 20 | 1.73 ± 0.25 | 5.01 ± 0.47* | - | - |
| Weight at the end of the experiment (g) | 423.1 ± 30.2 | 527.0 ± 45.3* | 487.0 ± 42.8# | 489.0 ± 55.2# |
| Total adipose tissue at the end of the experiment (g) | 14.3 ± 3.1 | 37.92 ± 4.4* | 29.38 ± 3.5* ** | 33.35 ± 4.9* |
Values expressed as mean ± sd. (n=6). *P˂0.05 compared to CG, **P˂0.05 compared to OG, #does not differ significantly from either CG or OG. CG (control group), OG (obese rats), PK (obese rats that received extract), and MET (obese rats that received metformin).
Table 3.
Weekly record of weight, liquid and food consumption, as well as caloric intake of each of the experimental diet groups.
Table 3.
Weekly record of weight, liquid and food consumption, as well as caloric intake of each of the experimental diet groups.
| Week 1 | CG | OG | PK | MET |
|---|---|---|---|---|
| Weight (g) | 389.00±28.70a | 479.00±35.97b | 472.00±32.49b | 494.00±67.77b |
| Liquid consumption (mL/day) | 30.60±4.59a | 17.42±2.61b | 13.85±2.07bc | 9.85±1.47c |
| Liquid consumption (mL /day/100 g) | 7.86±0.58a | 3.63±0.27b | 2.93±0.20b | 1.99±0.27c |
| Equivalent in kcal in drinkable water | 0a | 13.58±1.02b | 10.96±0.75b | 7.44±1.02c |
| Feed consumption (g/day) | 24.28±3.64a | 17.57±2.63ab | 17.71±2.65bc | 18.42±2.76bc |
| Feed consumption (g/day/100 g) | 6.24±0.46a | 3.66±0.27b | 3.75±0.25b | 3.73±0.51b |
| Equivalent in kcal in feed | 19.35±1.42a | 11.37±0.85b | 11.63±0.80b | 11.56±1.58b |
| Total Kcal/day/100 g body weight | 19.35±1.42ab | 24.95±1.87b | 22.59±1.55b | 19.01±2.60ab |
| Week 2 | CG | OG | PK | MET |
| Weight (g) | 398.00 ± 25.21a | 482.00 ± 33.55b | 459.00 ± 35.41b | 482.00 ± 55.16b |
| Liquid consumption (mL/day) | 32.57 ± 4.88a | 28.14 ± 4.22a | 33.60 ± 5.04a | 30.77 ± 4.61a |
| Liquid consumption (mL /day/100 g) | 8.18 ± 1.10a | 5.83 ± 0.95b | 7.32 ± 0.38b | 6.38 ± 0.06b |
| Equivalent in kcal in drinkable water | 0a | 65.39 ± 4.55b | 76.80 ± 5.92bc | 75.08 ± 10.15bc |
| Feed consumption (g/day) | 27.28 ± 4.09a | 16.00 ± 2.4b | 11.85 ± 1.77b | 12.71 ± 1.90b |
| Feed consumption (g/day/100 g) | 6.85 ± 0.44a | 3.31 ± 0.23b | 2.58 ± 0.19b | 2.63 ± 0.35b |
| Equivalent in kcal in feed | 21.25 ± 1.36a | 10.29 ± 0.71b | 8.008 ± 0.61c | 8.177 ± 1.10c |
| Total Kcal/day/100 g body weight | 21.25 ± 1.36a | 75.68 ± 5.26b | 84.80 ± 6.54bc | 83.26 ± 11.25bc |
| Week 3 | CG | OG | PK | MET |
| Weight (g) | 402.00 ± 30.75a | 492.00 ± 37.76b | 468.00 ± 29.42b | 486.00 ± 54.94b |
| Liquid consumption (mL/day) | 30.42 ± 4.56a | 44.85 ± 6.72b | 46.28 ± 6.94b | 31.00 ± 4.650a |
| Liquid consumption (mL /day/100 g) | 7.56 ± 0.57a | 9.11 ± 0.69b | 9.89 ± 0.62b | 6.37 ± 0.90a |
| Equivalent in kcal in drinkable water | 0a | 102.1 ± 7.83b | 110.7 ± 6.96b | 71.44 ± 10.13c |
| Feed consumption (g/day) | 23.14 ± 3.47a | 12.14 ± 1.82b | 9.571 ± 1.43b | 9.571 ± 1.43b |
| Feed consumption (g/day/100 g) | 17.84 ± 1.36a | 7.65 ± 0.58b | 6.34 ± 0.39b | 6.10 ± 0.86b |
| Equivalent in kcal in feed | 5.75 ± 0.44a | 2.46 ± 0.18b | 2.04 ± 0.12b | 1.96 ± 0.27b |
| Total Kcal/day/100 g body weight | 17.84 ± 1.36a | 109.7 ± 8.42b | 117.10 ± 7.36b | 77.54 ± 11.00c |
| Week 4 | CG | OG | PK | MET |
| Weight (g) | 423.1 ± 30.20a | 527.0 ± 45.30b | 487.0 ± 42.80ab | 489.0 ± 55.20ab |
| Liquid consumption (mL/day) | 33.11 ± 4.96a | 48.91 ± 7.33b | 44.57 ± 6.68ab | 52.97 ± 8.54b |
| Liquid consumption (mL /day/100 g) | 7.82 ± 0.63a | 9.28 ± 0.75b | 9.15 ± 0.61b | 10.83 ± 1.61b |
| Equivalent in kcal in drinkable water | 0a | 103.93 ± 8.40b | 102.55 ± 6.85b | 121.32 ± 18.09b |
| Feed consumption (g/day) | 24.42 ± 3.66a | 11.71 ± 1.75b | 10.71 ± 1.60b | 9.57 ± 1.43b |
| Feed consumption (g/day/100 g) | 5.77 ± 0.46a | 2.22 ± 0.17b | 2.19 ± 0.14b | 1.96 ± 0.27b |
| Equivalent in kcal in feed | 17.89 ± 1.44a | 6.88 ± 0.55b | 6.81 ± 0.45b | 6.09 ± 0.84b |
| Total Kcal/day/100 g body weight | 17.89 ± 1.44a | 110.81 ± 8.95b | 109.36 ± 10.34bc | 127.41 ± 12.94c |
Values expressed as mean ± sd. (n=6). Different superscripts indicate significant difference. CG (control group), OG (obese rats), PK (obese rats that received extract), and MET (obese rats that received metformin).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



