
Submitted:
08 July 2024
Posted:
09 July 2024
You are already at the latest version
Abstract
K+ channels do play a role in cell shape changes observed during cell proliferation and apoptosis. Research suggested that the dynamics of aggregation of Aquaporin-4 (AQP4) into AQP4-OAPs isoforms can trigger cell shape changes in malignant glioma cells. Here, we investigated the relationship between AQP4 and some K+ channels in the malignant glioma U87 line. The U87 cells transfected with the human M1-AQP4 and M23-AQP4 isoforms were investigated for morphology, gene expression of KCNJ8, KCNJ11, ABCC8, ABCC9, KCNMA1, and Cyclin genes by RT-PCR, recording the whole-cell K+ ions currents by patch-clamp experiments. AQP4 aggregation into OAPs increases the plasma membrane functional expression of the Kir6.2 and SUR2 subunits of the KATP channels, and of the KCNMA1 of the BK channels in U87 cells leading to a large increase in inward and outward K+ ion currents. These changes were associated with changes in morphology with a decrease in cell volume in the U87 cells and an increase in the ER density. These U87 cells accumulate in the mitotic and G2 cell cycle. The KATP channel blocker zoledronic acid reduced cell proliferation in either M23 AQP4-OAPs and M1 AQP4-tetramers transfected cells leading to early and late apoptosis, respectively. BK channel sustains the efflux of K+ ions associated with the M23 AQP4-OAPs expression in the U87 cells, but it is downregulated in the M1 AQP4-tetramers cells; KATP channels are effective in the M1 AQP4-tetramers and M23 AQP4-OAPs cells. Zoledronic acid can be effective in targeting pathogenic M1 AQP4-tetramers cell phenotype inhibiting KATP channels and inducing early apoptosis.
Keywords:
Glioblastoma
; BK channels
; KATP channels
; Aquaporin-4
; M1-AQP4 (AQP4-tetramers forming isoform)
; M23-AQP4 (AQP4-OAPs forming isoform)
; Cell cycle.
1. Introduction
To date, several types of brain tumors are known, and they differ considerably in outcome. According to their malignancy, they are classified from grade I to grade IV. Glioblastoma Multiforme (GBM), one of the most common adult primary brain tumors arising from glial cells [1,2], exhibits the highest malignancy (grade IV) and median survival of approximately 15 months [3]. The management of glioblastomas poses a clinical challenge due to the ability of glioma cells to rapidly infiltrate healthy brain parenchyma, allowing the cancer to escape control through surgical resections and localized radiation therapy, and promoting recurrence in other brain regions [4]. Great attention is given to the importance of cerebral fluid homeostasis, as its alteration is a common feature of brain tumors [3,5]. In the last 15 years, a growing body of evidence has underscored the significance of splicing, dysregulated expression, and altered function of ion channels and Aquaporins (AQPs) in various cancer types. These molecular aberrations play crucial roles in multiple cancer processes, highlighting their potential as important therapeutic targets and biomarkers for cancer diagnosis and treatment [6]. Their pivotal role in determining the fate of these cells has led to the characterization of certain ion channels as oncogenic channels due to their impact on cancer development and/or progression [7]. Numerous studies conducted across various laboratories have consistently demonstrated that the manipulation of ion channels and/or AQPs can effectively impede the growth and migration of cancer cells. Investigations have reported the successful utilization of substances targeting different classes of AQPs and ion channels, such as the Ca2+-activated K+ channels of large (BK) and intermediate conductance (IKCa), and ATP-sensitive potassium channels (KATP), to inhibit cell migration, invasion, and metastasis in vitro and in vivo for diverse types of carcinomas [4,8,9].
To date, researchers have identified a minimum of 13 distinct isoforms of AQPs. These specialized channels are extensively expressed in diverse epithelial and endothelial cells within mammals and play a crucial role in modulating cell capacity in volume control and response to an environment osmotic change [10,11]. The most abundant AQP in the CNS is the glial water channel AQP4 expressed in astrocytes. Astrocytes exhibit a distinct polarized distribution of AQP4, with a prominent presence in the astrocytic feet that come into contact with blood vessels at the blood-brain barrier (BBB) and in those forming the glial limitans [12]. In these sites, under normal condition, AQP4 forms heterotetramers in the plasma membrane consisting mainly of the two different isoforms AQP4-OAP and AQP4-tetramer. A smaller amount of the recently discovered M23-AQP4 and M1-AQP4 is also expressed and shown to be important to lock AQP4 at astrocyte processes. The isoform ratio controls the aggregation of AQP4 into supramolecular structures called orthogonal particle arrays (AQP4-OAPs) [13]. Interaction with intracellular scaffold proteins, such as α-syntrophin, determines the formation and localization of OAPs in astrocytes [14,15,16]. It is interesting to note that these complexes also contain the inwardly rectifying potassium channel such as the ATP-sensitive K+ channel (KATP), and the large conductance calcium-activated K+ channels (BK), in particular the K+ ions are absorbed by the astrocytes and the water follows them osmotically through the AQP4 water channels to regulate the cell volume [17,18]. A pivotal player in preserving water and ion balance within the central nervous system, AQP4 has garnered attention for its vital role in brain edema.
Recently, researchers have uncovered its additional involvement in cell migration, further expanding its repertoire of functions. Notably, increased expression of AQP4 has been observed in glioblastoma multiforme (GBM) [19,20,21]. Simone and coworkers [13,22] demonstrated that the aggregation of AQP4 in OAPs influences the biology and the fate of glioma cells. In particular, M23 AQP4-OAPs triggered cell shape changes associated with alterations in the cytoskeleton of F -actin leading to glioma cell apoptosis. Conversely, expression of M1 AQP4-tetramers that are unable to aggregate into OAPs, increased the invasiveness, cell migration, and metalloproteinase-9 activity of glioma cells escaping apoptosis 13.
K+ ion channels regulating cell volume and resting potentials, play a role in cancer cells progression mechanisms [23,24,25]. For example, the overexpression of different potassium channel subunits, composing the BK and KATP channels, have been found in tumor tissues of different origin and especially in those of gliomas and other brain tumors (Table 1). Conducting an extensive search using PubMed on glioma/brain cancer-related articles of ABCC8, ABCC9, KCNJ11, KCNJ8, AQP4, and KCNMA1 genes has yielded interesting results. Apart from the well-established roles of AQP4 and K+ ion channels in diverse pathological conditions, the genes responsible for encoding AQPs and K+ ion channels exhibit differential expressions and mutations in different cancer types. This discovery highlights the potential significance of these genes in the context of cancer development and may offer valuable insights into their involvement in cancer-related processes [26] (Table 1).
The hypothesis that there is a relationship between the overexpression of the K+ channel and the generation and growth of malignant tumors has been confirmed in different studies as these channels are involved in cell proliferation, apoptosis and differentiation [27,28,29].

Table 1.
Glioma brain cancer-related articles of ABCC8, ABCC9, KCNJ11, KCNJ8, AQP4 and KCNMA1 genes in PubMed.
Table 1.
Glioma brain cancer-related articles of ABCC8, ABCC9, KCNJ11, KCNJ8, AQP4 and KCNMA1 genes in PubMed.
| Differential expression/ Mutation | Cancers | Information on analyzed samples | Consequences | PubMed code | |
|---|---|---|---|---|---|
| Up-regulation | Glioma | Tissue from 22 patients with newly diagnosed GBM | [30] | ||
| Up-regulation | Glioma | A total of 50 glioma samples, ranging in grades from II to IV, were collected from a cohort of 35 male and 15 female patients. | Cells exhibiting enhanced migration potential displayed notably high levels of AQP4 expression, indicating a potential association between AQP4 and glioma cell migration. | [31] | |
| Up-regulation | Brain tumors | The study involved analyzing 5 tumor samples from subependymomas located in the fourth ventricle, as well as subependymoma (SE) tumor samples found supratentorial with relation to the first to the third ventricle. | Increased AQP4 expression in malignant tumors, suggesting their involvement in edema formation, invasive growth and recurrent tumor formation but do not have a significant role in benign tumors. | [32] | |
| AQP4 | Up-regulation | GBM | The tumour samples from 14 patients with primary glioblastomas | Increased expression of AQP4, loss of polarity and alteration of the intra and extracellular matrix were found in the analysis of GBM samples and are more serious clinical signs of glioblastoma and the formation of cerebral edema. | [33] |
| Up-regulation | Brain tumors/ GBM | Tissue samples from brain tumors of 26 patients | The evident role of AQP4 in tumor malignancy suggests that targeted manipulation of this protein could potentially unlock a therapeutic avenue. | [34] | |
| Up-regulation (AQP4-tetramer and AQP4-OAP) | Brain tumors/ GBM | Tumor tissues obtained from a total of 22 patients diagnosed with astrocytoma of WHO grades II, III, and IV, and an additional patient diagnosed with glioblastoma multiforme (GBM), were included in the study. | Upregulation of AQP4-tetramers and upregulation of mRNA -AQP4-OAPs in all astrocytomas, but the AQP4-OAPs / AQP4-tetramers ratio differed from 1.14 to 1.5 in low-grade astrocytomas to 1.94 in glioblastomas. The possible impact on the development of new therapies. | [31] | |
| Up-regulation | Brain tumors/ GBM | Brain tumors and the corresponding adjacent tissues from 30 patients diagnosed with glioblastoma. | The overexpression of AQP4 was observed in both brain tumors and the adjacent tissues, and this heightened expression was found to be correlated with the extent of brain edema. | [35] | |
| down-regulation | Brain tumors/ GBM | A total of 16 tissue samples were collected from various regions within the tumoral core. | The presence of AQP4 alterations in GBMs appears to play a role in edema formation. Therefore, AQP4 could be viewed as a promising early biomarker for tracking GBM progression and also as a potential target for AQP4 modulation in therapeutic approaches. | [36] | |
| KATP | |||||
| Up-regulation of ABCC8 | Glioma | The information is based on the analysis of 1893 human glioma samples from four independent databases | Glioma chemosensitivity can be predicted by high ABCC8 mRNA expression, whereas low ABCC8 mRNA expression can serve as an indicator of glioma sensitivity to radiotherapy. | [37] | |
| Up-regulation of ABCC8 | Brain tumors | The information comes from the analysis of human tissue samples from 6 glioblastoma, 12 brain metastases, 11 medulloblastoma, 9 supratentorial ependymomas, and 8 posterior fossa ependymomas | SUR1 is a potential therapeutic target for reducing neuroinflammation in adult and pediatric brain tumors. Inhibition of SUR1 induces neuronal stabilization in glioblastoma, brain metastases and posterior fossa ependymoma, and edema reduction in medulloblastoma. | [38] | |
| Up-regulation of KCNJ8 and ABCC8 | Glioma | 20 human glioma biopsies | The Kir6.2 and SUR1 subunits of the KATP channel are involved in the proliferation of U87 and U251 glioma cells. The KATP channel inhibitors significantly reduced the growth curve. On the other hand, KATP channel agonists promoted the proliferation of U87 and U251 cells. | [39] | |
| BK | Up-regulation of KCNMA1 | Glioma | Biopsies from patients with malignant gliomas | The expression of BK channels has shown a positive correlation with tumor malignancy grades, indicating a significant role for the gBK channel in glioma biology. Utilizing BK channel agonists could potentially be advantageous for brain tumor patients, as they might enhance the delivery of anti-neoplastic agents to brain tumors. | [40] |
| Up-regulation of KCNMA1 | Brain tumors | Samples tissues from patients with malignant gliomas | [41] |
Drugs that specifically block K+ ion channels have shown anticancer effects by directly inhibiting tumor growth or by improving the efficacy of chemotherapy or cytotoxic drugs as a combined therapeutic strategy [23]. For instance, small molecules KATP channel inhibitor like glibenclamide that reduced cell proliferation in a variety of cells including leiomyoma and MDA-MB-231 cancer cells, were extensively investigated in cancer. More recently, a monoclonal antibody (Y4) targeting the cap domain of TASK-3 has garnered attention for its potential in inhibiting the growth of human lung cancer xenografts and reducing breast cancer metastasis in mice [23]. The first polyclonal antibody (BIL010t; Biocentre) targeting a non-functional form of P2X7 (nfP2X7) has reached the clinical trials for the treatment of basal cell carcinoma [23]. In addition, short peptides derived from venom are under investigation. On the other hand, several studies have shown the impact Ca2+ activated potassium channels (BK) on tumor cell proliferation and their association with the tumorigenesis process in patients and animals’ model [23,29].
In this study, we investigated the functional/expression changes of KATP and BK channel subunits by patch-clamp and RT-PCR experiments in U87 cells transfected with AQP4 isoforms enabling different aggregation states (AQP4 OAPs vs AQP4 tetramers) and compared to the non-transfected cells (WT). The aim has been to evaluate the relationship between the KATP and BK channels functional expression and the role of the AQP4 aggregation state in U87 cells.
KATP channel inhibitors like sulfonylureas drugs are known to induce apoptosis in different cell lines and tissues [42] with antiproliferative effects. The BK channel openers also induce apoptosis [43]. Given the expected role of the KATP and BK channels in glioblastoma, we also investigated the antiproliferative effects of zoledronic acid, a recently identified KATP channel blocker targeting either the Kir6.2/1 and SUR2 subunits in native cells [44,45,46] and in cell line transfected with KATP channels subunits [47]. This drug is capable to activate BK channels in MD-MBA 231 breast cell line with apoptosis [48], and it is a modulator of TRPV1 channel [49] and it is used in the treatment of the osteoporosis, bone metastasis and myeloma increasing the overall survival of patients with malignancy [50,51].
2. Materials and Methods
2.1. Cell Lines
The U87 MG (ATCC HTB-14) cell line, derived from a malignant glioma in a female patient using the explant technique, was procured and authenticated from the ATCC (www.lgcstandards-atcc.org) [52]. Experiments were conducted using cells from passages 1 to 20. Routine mycoplasma testing was carried out, either using MycoAlert Substrate (https://bioscience.lonza.com) or fluorescence staining with DAPI. The cells were cultured in DMEM/F12 supplemented with 10% FBS and 100 units of penicillin/streptomycin, in a 5% CO2 environment at 37˚C. To maintain exponential growth, the cells were sub cultured every 3 days and maintained at approximately 80% confluency until further experiments. Cell culture reagents were obtained from Euro clone (www.euroclonegroup.it).
2.2. Constructs and Transfection
Human M1M23I-AQP4 (referred to as AQP4-tetramers) and M23-AQP4 (referred to as AQP4-OAPs) coding sequences were cloned into pTarget vectors (A1410, www.Promega.com). For the experiments, the mutated form of M1-AQP4 (M23I) was used, which has been previously characterized and shown to exclusively produce AQP4-tetramers (PMID: 20007705). To prepare for transfection, cells were seeded in the antibiotic-free medium when they reached 70% confluence, 24 hours before the actual transfection. Lipofectamine 3000 (L3000015, www.thermofisher.com) was employed for the transient transfection, following the manufacturer's protocol, in the OptiMEM growth medium. After 24 hours of transfection, recordings of the potassium currents and immunofluorescence analysis were performed [22].
2.3. Drugs and Solutions
K+ channel modulators glyburide/glibenclamide (GLIB) cat. N° PHR1287, TEA, diazoxide (DIAZO), iberiotoxin (IbTX) cat. N° I5904 and BaCl2 cat. N° 449644 were purchased from Sigma (SIGMA Chemical Co., Milan, Italy). Zoledronic acid was prepared in our labs [47]. DMSO was observed to have no impact on the channel currents or cell viability [53].
2.4. Antibodies
For immunofluorescence analysis, the following antibodies were employed: rabbit polyclonal anti-AQP4 (H-80) (scbt, sc-20812, RRID: AB_2274338) at a dilution of 1:500, and donkey anti-rabbit Alexa Fluor 594–conjugated (Thermofisher, A21207, RRID: AB_141637) at a dilution of 1:1000. To stain F-Actin, Phalloidin Alexa Fluor 488–conjugated was used at a dilution of 1:500 (A12379, www.thermofisher.com).
2.5. Nuclear Staining
U87 cells were plated in a 96-well plate with a density of 3×103 cells per well and exposed to either zoledronic acid or diazoxide for 18 hours. After the incubation, the cells were fixed and processed for immunofluorescence as previously described. Alternatively, some cells were treated with 10 μg/mL of DAPI for 30 minutes at 37°C to stain the cell nuclei. Following the staining procedures, the cells were examined under an inverted fluorescence microscope. The nuclei were automatically detected and analyzed for their mean nuclear area using Fiji (Fiji, RRID:SCR_002285). For each experimental condition, nuclei from at least three fields were analyzed, and the experiments were independently repeated three times.
2.6. Immunofluorescence and Quantitative Analysis
U87 cells were fixed using 4% paraformaldehyde (PFA) for 15 minutes and then washed three times with PBS. To facilitate permeabilization, 0.3% Triton X-100 was applied. Following this, a 30-minute blocking step with 0.1% bovine albumin serum (BSA) was carried out at room temperature. Next, the cells were subjected to a 1-hour incubation with primary antibodies, followed by PBS washes containing BSA. For secondary antibody labeling, Alexa Fluor–conjugated secondary antibodies were used. Finally, the cells were mounted using a medium containing 50% glycerol, 1% n-propyl gallate in PBS, along with DAPI for nuclear staining. To visualize the immunostained cells, an epifluorescence photomicroscope equipped with 16x and 40x oil PL FL FLUOTAR objectives from Leica Microsystems (www.leica-microsystems.com) was utilized. Digital images were captured using a DMX1200 camera from Nikon (www.Nikon.it) and processed using LAS AF software, provided by Leica Microsystems GmbH (RRID:SCR_013673). The auto contrast function in Photoshop CS5 (RRID:SCR_014199) was applied to the entire images for image enhancement. For quantitative analysis following immunofluorescence, three to five different fields were chosen from each of the three independent experiments conducted on separate days. ImageJ or Fiji software (Fiji, RRID:SCR_002285) was used to accurately count the cells within each field, and the data were analyzed with GraphPad Prism 9 software (GraphPad Prism, RRID:SCR_002798).
2.7. Morphologic Analysis
Morphological analysis was carried out to assess the presence of two cell types: round-shaped cells and irregular-shaped cells exhibiting a star-like phenotype. The cells displaying these altered morphologies were distinctly identifiable from other cells in each field. Two independent investigators performed the analysis in a blinded manner. The quantification of cells with altered morphology was accomplished using Fiji software (Fiji, RRID:SCR_002285), and the data were analyzed using GraphPad Prism 9 software (GraphPad Prism, RRID:SCR_002798).
2.8. Cell Viability Assay
To examine the effect of K+ channel modulators on cell proliferation. Briefly, cell viability was assessed using the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) test. This assay is based on the principle that the reduction of MTT tetrazolium salt is primarily carried out by mitochondrial dehydrogenases. Following a 24-hour exposure or after specific incubation periods in the culture medium, an MTT stock solution (5 mg/mL in medium) was added to each dish at a volume equivalent to one-tenth of the original culture volume. The cells were then incubated for 2 hours at 37°C in a humidified CO2 environment. Subsequently, the supernatant was removed, and the cells were dissolved in 150 μl of DMSO-ethanol in the ratio 1:1., which is used to solubilize the formazan crystals formed as a result of the MTT reduction. Formazan formation was quantified using spectrophotometry, measuring the absorbance at 570 nm. The amount of formazan produced is proportional to the number of viable cells in the culture, allowing for the evaluation of cell viability under different experimental conditions.
2.9. Patch-Clamp Experiments
The whole-cell patch-clamp experiments for recorded membrane cur-rents were performed in asymmetrical K+ ion concentration in physio-logical conditions using pipettes with the resistance of 3–5 MΩ. The pipette solution contained 132 mM KCl (P3911, Sigma Chemical Co., Milan, Italy ), 1 mM ethylene glycol-bis (β-aminoethyl ether)-N, N,N′, N′-tetra acetic acid EGTA (E3889, Sigma Chemical Co., Milan, Italy), 10 mM NaCl (S9888, Sigma Chemical Co., Milan, Italy), 2 mM MgCl2 (M8266, Sigma Chemical Co., Milan, Italy), 10 mM HEPES (H3375, Sigma Chemical Co., Milan, Italy), 1 mM Na2ATP (A26209, Sigma Chemical Co., Milan, Italy), and 0.3 mM Na2GDP (51060, Sigma Chemical Co., Milan, Italy) (pH = 7.2). The bath solution contained 142 mM NaCl (S9888, Sigma Chemical Co., Milan, Italy), 2.8 mM KCl (P3911, Sigma Chemical Co., Milan, Italy ), 1 mM CaCl2 (C8106, Sigma Chemical Co., Milan, Italy ), 1 mM MgCl2 (M8266, Sigma Chemical Co., Milan, Italy), 11 mM glucose (D9434, Sigma Chemical Co., Milan, Italy), and 10 mM HEPES (H3375, Sigma Chemical Co., Milan, Italy)(pH = 7.4). CaCl2 was added to the pipette solutions to give a free Ca2+ ion concentration of 1.6 × 10−6 M in whole-cell experiments. The calculation of the free Ca2+ ion concentration in the pipette was performed using the MaxChelator software (Stanford University, Stanford, CA, USA). Drug actions on the K+ ion currents recorded during instantaneous I/V relationships were investigated by applying a depolarization protocol, in response to voltage pulses from −120 mV to +120 mV (Vm) in 20 mV steps. Currents were expressed as densities (pA/pF) to control cell size/capacitance differences. All experiments were per-formed as described in Scala et al. [54].
2.10. Polymerase Chain Reaction
Total RNA was isolated and purified from the U87 cells WT and U87 transfected by AQP4-OAP or AQP4-tetramer with Trizol reagent (Invitrogen, Thermo Fisher Scientific Inc., Waltham, MA, USA) and quantified using a spectrophotometer (ND-1000 Nano-Drop, Thermo Fisher Scientific Inc., Waltham, MA, USA). Real-time PCR was performed in triplicate using the Applied Biosystems Real-time PCR 7500 Fast system (United States). The mRNA expression of the genes was normalized to the best housekeeping gene β-actin (Actb) selected from glyceraldehyde-3-phosphate dehydrogenase (Gapdh), using Best Keeper and NorFinder software. TaqMan hydrolysis primer and probe gene expression assays were obtained from Life Technologies with the following assay KCNMA1 ID: Hs01119504_m1; KCNJ11 ID: Hs00265026_s1; KCNJ8 ID: Hs00958961_m1; ABCC8 ID: Hs01093752_m1; ABCC9 ID: Hs00245832_m1; TRPV1 ID: Hs00218912_m1; Cyclin A2 I; D: Hs00996788_m1; Cyclin B1 ID: Hs01030099_m1; Cyclin E ID: Hs01026536_m1; B-actin: ID: Hs01101944_s1. All gene expression experiments were conducted following the MIQE guidelines [55,56,57].
2.11. Statistical Analysis
Data were collected and analyzed using various software tools, including Excel software (Microsoft, RRID: SCR_016137), Clampfit 10.5 (Molecular Devices, RRID: SCR_011323), and SigmaPlot 10.0 (Systat Software, RRID: SCR_003210) [58]. The results are expressed as mean ± SEM unless stated otherwise. The number of replicates for each experimental dataset is mentioned in the figure description. For statistical analysis, one-way analysis of variance (ANOVA) followed by multiple comparison tests was applied, with a significance level of p < 0.05, unless otherwise specified. Additionally, the Student t-test was used to compare the significance between means. Statistical significance was considered when p < 0.05 unless explicitly stated otherwise.
3. Results
3.1. AQP4-OAP Expression in U87 Cells
After transfecting AQPs isoforms into U87 cells, we routinely conduct immunofluorescence assays on both the wild-type (WT) cells and the transfected cells to confirm the success of the experiment (Figure 1). Moreover, our knowledge about AQP4 expression in glioma cells is currently limited. In a study by McCoy et al. [59], the expression of AQP4 was investigated in various commonly used human glioma cell lines (D54, D65, STTG1, U87, U251) and several acute patient biopsies using PCR, Western blot, and immunocytochemistry. These findings were compared with non-malignant astrocytes and normal brain tissues. The researchers observed that all glioma patient biopsies expressed AQP4. However, when these cells were isolated and cultured as cell lines, they lost AQP4 protein expression. This phenomenon poses challenges in studying the endogenous expression of AQP4 in cultured cells and utilizing known inhibitors of AQP4, such as TGN-020. This issue has been commonly reported in studies involving glioma cell lines. In our previous research, we also found negative staining for AQP4 in U87 and U251 cell lines, as well as in GL95 primary cell cultures [13].
3.2. Whole-Cell Inward and Outward Macroscopic K+ Currents Recorded in U87WT Cells and Effects of the K+ Channel Modulators on Cell Proliferation
Large outward K+ currents were recorded in U87 cells (Figure 2A). To perform an electrophysiological characterization and identify K+ channels that are present endogenously in U87 cells and that may influence U87 cell behavior, including proliferation, we evaluated the effects of a variety of K+ channel blockers and an MTT assay was used to determine cell viability after drug treatment. The effects of TEA, a non-specific outward transient and voltage-dependent K+ channel blocker, of the glibenclamide (GLIB), a specific ATP K+ channel blocker, and the specific BK-channel blocker iberiotoxin (IbTX) were investigated. As reported in figure 2B, D, at 60 mv the TEA, IbTX exerted a current reduction of about -80% and -46 % respectively, GLIB, in the presence of low ATP content (1mM) in the pipette and at - 60 mV of voltage membrane, reduced the inward current by about -40% (Figure 2C,D). Regarding the effects on cell viability detected at 48 h, as shown in Figure 3, TEA, GLIB and IbTX reduced the number of U87 cells compared to the untreated control condition. However, the KATP channel agonist diazoxide (DIAZO) had no significant effect on cell proliferation. TEA, GLIB and IbTX at the highest concentrations reduced cell viability by approximately -60.14%, -65.4 % and -40.77 %, respectively, despite in our case high concentrations of these drugs were used especially for IbTX (Figure 3). This can be due to fact that IbTX is relatively impermeant and do not act on the mitochondrial BK channel. The data suggest of large outward and inward currents mostly sustained by BK channels and KATP channels, respectively, contributing to the cell proliferation in this cell. A contribution of other ion channels to the cell currents and proliferations cannot be excluded, for instance TRPV1 channel is expressed in these cells and can contribute to the outward currents despite with minor extend since all outward currents were inhibited by the unselective Kv/BK channel blocker TEA.
3.3. AQP4 Aggregation State Affects the TEA-K+ Sensitive Currents in U87 Glioma Cells
The transfection of U87 cells with the tetramer forming AQP4-tetramer or the OAP forming AQP4-OAP isoform, allowed to monitor the electrophysiological behavior of these cells, creating a system to study the impact of AQP4 aggregated (M23 AQP4-OAPs) and disaggregated pathogenic (M1 AQP4-tetramers) forms on electrical activity. The presence of AQP4 aggregation in AQP4-OAPs leads to a significant increase in both inward and outward K+ ion currents, with an increase of about 370% at -60 mV (Vm), and of 400% at 60 mV (Vm), respectively vs U87 cells not transfected (Figure 4A). Instead, the aggregation of AQP4-tetramers causes a significant increase of the inward K+ ion currents of about 76% at -60 mV, while the outward currents suffered a decrease of -52% at 60 mV vs U87 cells not transfected (Figure 4B, C). After transfection the currents were responsive to the the unselective blockers TEA and BaCl2 (Figure 4A, B, D). In U87/M23 cells the TEA sensitivity was observed at negative voltages where the BK should not be activated, the subunit may account for this effect in neuron [60]. BK channel phosporilatylation may account for BK channel openings increasing TEA-sensitive channel currents at negative membrane potentials. In addition, the openings of the intermediated calcium-activated K+ channel (IKCa), that is active at more negative potentials, contribute to the currents in our cells as it has been observed in U87-M GBM cells [60] where the BK and IKCa open in response to the hypotonic-induced activation of mechanosensitive channels with influx of Ca2+ ions. Moreover, in line with the large K+ ion currents recorded in the AQP4-OAPs, these cells were more repolarized vs the other cells with a resting potential, respectively, of -40 mV±12 and -10±5 mV in the AQP4-OAPs and AQP4-tetramers -transfected U87 cells, and -10±7 mV in the U87 cells not transfected cells.
3.4. AQP4 Aggregation State Changes the Expression Profile of KCNMA1, KCNJ11, ABCC8 and ABCC9 Genes in U87 Glioma Cells
AQP4 aggregation state similarly and significantly caused upregulation of the KCNJ11, ABCC8 and ABCC9 genes in U87 glioma cells (Figure 5). A different behavior concerns the KCNMA1 with AQP4 tetramers leading to a significant downregulation in line with the reduced outward current observed in these cells. On the contrary, AQP4-OAPs caused a statistically significant up-regulation of the KCNMA1 in U87 glioma cells in line with the elevated outward currents recorded in these cells. A not statistical enhancement of the expression of TRPV1 gene was found in the pathogenic AQP4-tetramers (Figure 5).
3.5. AQP4 and Kir6.2 are Involved in Glioma Apoptotic Fate
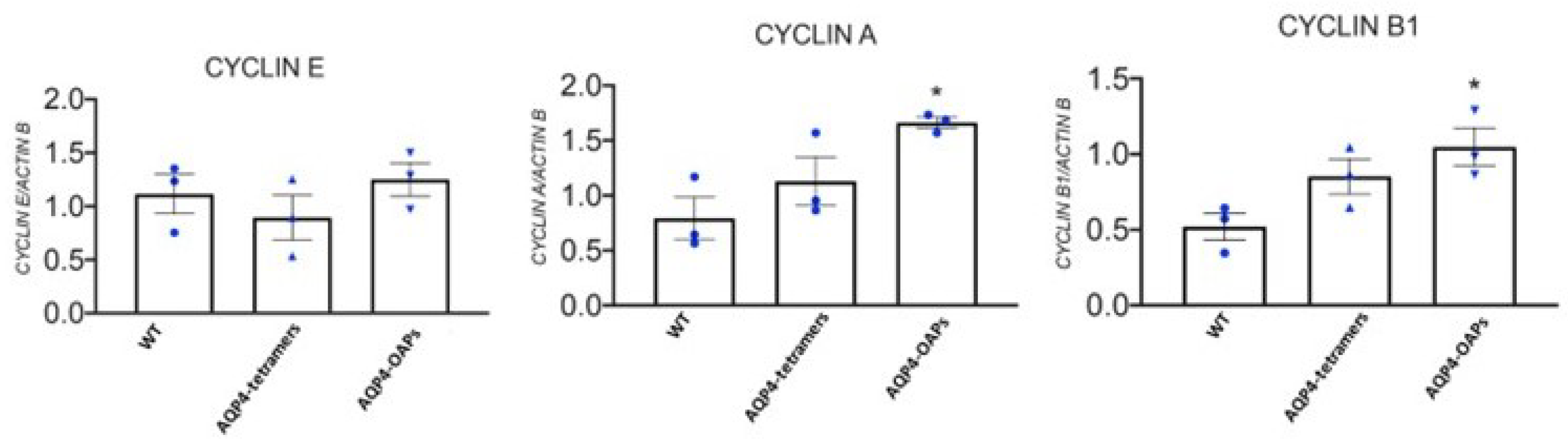
Due to the role of KATP channels in proliferation and apoptotic outcome in glioma, we tested the hypothesis that Kir6.2-SUR2 and AQP4 collaborate in glioma cell apoptotic fate. To this aim, AQP4-OAPs or AQP4-tetramers expressing glioma cells were treated with zoledronic acid, a Kir6.2-SUR2 K ATP channels inhibitor [47,61], and with KATP channel agonist diazoxide. The expression of AQP4 and F-actin was examined using immunofluorescence (Figure 6A). The results demonstrated that, upon treatment with zoledronic acid, a remarkable change in cell morphology was specifically observed in glioma cells expressing AQP4-OAPs, whereas no such effect was observed in glioma cells expressing AQP4-tetramers. Conversely, diazoxide treatment did not induce any noticeable effect. More specifically, the application of zoledronic acid transformed AQP4-OAPs expressing U87 cells from irregular-shaped to round-shaped, exhibiting characteristics consistent with apoptosis (Figure 6B). This observation aligns with Coffin's hypothesis. Notably, the round-shaped cells displayed membrane protrusions, known as "beads-on-a-string," which are typical apoptotic cell structures previously reported in stressed conditions [22]. Additionally, these cells exhibited membrane blebbing and condensed nuclei. The quantitative analysis (Figure 6B) further confirmed a significantly higher occurrence of round-shaped cells in glioma cells expressing human AQP4-OAPs following treatment with zoledronic acid, as compared to untreated cells. By analyzing nuclear condensation and DNA fragmentation we observed a SUR2A-specific enhancement of zoledronic acid-induced apoptosis (Figure 6C). These findings suggest that both AQP4 and Kir6.2 are involved in glioma apoptotic fate. RT PCR expression of the cyclin genes (E, A, B1) revealed that M23 AQP4-OAP transfection segregates the cells in the mitotic (M) and G2 states of the cycle. A tendency to segregation of the U87 cells in the mitotic (M) and G2 states is observed also in the M1 AQP4 tetramers transfected cells (Figure 7). Therefore, an abnormally enhancement of the functional/expression of the hslo/BK and Kir6.2-SUR2-1/KATP channel subunits were recorded in the AQP4-OAP transfected U087 cells that were repolarized and were associated with G2-M cycle, in comparison to the not transfected cells. The AQP4 tetramers transfected cells showed reduced KATP and BK channel currents, the downregulation of the hslo/BK channel subunit sensing the intracellular Ca2+ ions thereby favoring the influx of Ca2+ ions possibly carried by TRPV1 with cell depolarization.
4. Discussion
In the present work we showed that the AQP4 aggregation into the pathogenic AQP4-tetramer and AQP4-OAPs have differently affected the expression profile of KCNMA1, KCNJ11, ABCC8 and ABCC9 genes but not of the KCNJ8 gene in U87 glioma cell, evaluated by the RTPCR gene expression. In parallel, the endogenous whole cell currents, sustained by the BK and KATP channel subunits and recorded using patch-clamp experiments, were also affected on the same cells. In particular, the expression of the KCNMA1 gene, encoding for the hSlo pore forming subunit of the BK channels, increased by 4-fold in the AQP4-OAPs cells and the currents was enhanced by 8-fold in the physiological range of membrane potentials of -20 mV (Vm) and -10 mV (Vm) at which BK channels are operative vs not transfected cells. Conversely, the KCNMA1 expression decreased by 1-fold in the AQP4-tetramers cells in line with the reduced K+ ion currents observed in these cells. These data suggest of a significant role of the BK channels in mediating the efflux of K+ ion associated with AQP4 water efflux in U87 cells. Also, the expression of the KCNJ11 gene encoding for the pore forming subunit Kir6.2 enhanced by 2.5-fold in the AQP4-OAPs cells vs not transfected cells. The combined expression change of the KCNMA1 and KCNJ11 (about 6.5 folds) genes is in the same order of magnitude of the observed enhancement of the K+ ion whole cell currents in the AQP4-OAPs cells at the physiological range of membrane potentials of -20 mV and -10 mV (Vm) (about 8 folds), at which these ion channels are operative supporting the significant contribution of either KATP and BK channels to the K+ ion currents in the AQP4-OAPs cells.
According to the large K+ ion currents recorded in the AQP4-OAPs cells, these cells were significantly more repolarized vs not transfected and AQP4-tetramers transfected U87 cells. Cell depolarization is indeed associated to mitosis in the cell cycle [62,63,64]. All U87 cells investigated here were depolarized vs not cancer cells. The AQP4-OAPs cells show reduced volume and high density of ER, and apoptotic cell death vs the other cells suggesting of a relevant contribution of the BK and KATP channels and AQP4 to this phenomenon. The observed enhanced functional/expression of the KATP and BK channel subunits lead to cell repolarization in the AQP4-OAPs cells vs the not transfected and M1 AQP4-tetramers cells, and the AQP4-OAPs cells were more likely to segregate in the G2-M state indication of an active proliferative condition.
It should be noted that cell proliferation experiments were performed using the MTT assay that is metabolized at the mitochondrial level, and as potassium channels are present in the inner mitochondria membrane, their functionality affects cell viability.
Upregulation of KATP channel subunits SUR2, Kir6.2 and Kir6.1 has been recently associated with cancer in CNS and peripheral tissues [57]. The upregulation of the ABCC8/SUR1 subunit is associated with better prognosis in pancreatic cancers and in some brain tumors like glioma [37,59,65].
Stimulation of the KATP activity by growth factors, insulin [66,67], and by the KATP channel opener minoxidil induces cell proliferation and cancer progression in animals and humans [57]. Gain of function mutations (GOF) of the ABCC8 and KCNJ8 subunits are associated with Cantu ‘syndrome characterized by cardiovascular phenotypes in human and animals, and by cases of pituary adenoma in some family [68,69]. Inhibition of KATP channel activity induced by the apoptotic drug and ion channel modulator staurosporine [70,71], and the KATP channel blockers like sulfonylureas or by the pyruvate kinase antibodies instead induce apoptosis and antiproliferative effects [72,73,74]. In line with these data we found here the upregulation of the KATP channel subunits observed in the AQP4-tetramers and AQP4-OAPs cells and the enhancement of the relative ion channel currents supports the idea that KATP channels are involved in cell proliferation in either type of cells.
We found that inhibition of the KATP activity AQP4-OAPs by zoledronic acid that targets either the Kir6.1-2 and SUR2 subunits expressing cells, manages the cellular shape driving cells from irregular-shaped towards round-shaped and that phenomenon is linked to different stage of apoptosis. This drug is also a modulator of TRPV1 leading to inhibition of TRPV1 channel [75]. This mechanism may contribute to the inhibition of influx of Ca2+ ions in U87 cells. In our cells the TRPV1 is expressed but the observed currents were largely carried by BK and KATP channels since they were fully inhibited TEA and BaCl2. In addition, the possible contribution of other mechanisms such as the inhibition of farnesyl pyrophosphate synthase by zoledronic acid cannot be excluded to date. Zoledronic acid has been shown to act on different target cells inducing for instance cell proliferation of osteoblasts but reducing osteoclasts activity. The pro-survival and pro-differentiation effects of zoledronic acid in cancer and bone cells are observed between concentrations of 10−9 M to 10−6 M, while the proapoptotic effects at concentrations > 10−5 M [49,54] that were used in our experiments in the present work.
We can speculate that Kir6.2 upregulation in AQP4-OAPs expressing cells are helpful to glioma cells to escape from apoptosis and survive.
The hslo subunit and its splicing isoforms have a role in different pathophysiological conditions [76,77], they are well known regulators of cell proliferation and are targets of drugs affecting cell death [78,79], the upregulation of these ion channels in the presence of AQP4 regulating the efflux of K+ ions and water, instead selectivity drives the U87 cells to a less pathogenic AQP4-OAPs cells aggregation. These effects can be due to protein-protein interaction mechanisms or mediated by second messengers.
In previous work, the influx of Ca2+ ions mediated by the hypotonic-induced activation of mechanosensitive channels was found to be a key step for opening both the BK and the intermediate Ca2+ (IKCa) channels in normal U87-M GBM cells [80]. Here, these phenomena, the reduced reversal potentials of the K+ ion currents in our cells far from the equilibrium potentials for K+ ions, can be the result of cationic overactive components like the TRPV1 found in our work and the mechanosensitive channels previously described [80] that allow additional influx of calcium ions into the cells. In addition, the contribution of different sub cell GMB populations like neurosphere in the same culture also differing for the BK channel currents expression and response to inhibitors can contribute to BK channel currents recorded in our GBM cells [81].
5. Conclusions
BK channels sustain the efflux of K+ ions associated with the AQP4-OAPs expression in the U87 cells, KATP channels sustain the efflux in the M1 AQP4-tetramers and AQP4-OAPs cells. The KATP channel antagonist zoledronic acid can be effective in targeting pathogenic AQP4-tetramers cell phenotype inducing early apoptosis.
6. Patents
No patents are resulting from the work reported in this manuscript.
Supplementary Materials
N/A.
Author Contributions
Conceptualization, G.P. N., F.M., and D.T.; methodology, L.S., F.M., , M.A.; software, F.M.; validation, D.T.; formal analysis, F.M., L.S.; investigation, F.M.; resources, G.P.N.; data cura-tion, F.M., and L.S.; writing—original draft preparation, F.M., L.S.; writing—review and edit-ing, D.T.; visualization, F.M., L.S.; supervision, D.T.; project administration, G.P.N.; funding acquisition, G.P.N. All authors have read and agreed to the published version of the manu-script.” Please turn to the CRediT taxonomy for the term explanation. Authorship must be limited to those who have contributed to the work reported.
Funding
G.P.N. acknowledges funding from: 1) AstroDyn (FA9550-19-1-0370), AstroColl (FA9550-21-1-00352) and Stochastic Biophysical Interactions within Aquaporin-4 Assemblies (FA9550-20-1-0324) funded by AFOSR; 2) Marie Skłodowska-Curie Actions -ITN-2020 ASTROTECH (GA956325) funded by the European Commission; 3) HORIZON EUROPE SEEDS INTER-GLIO (S08I) funded by the University of Bari Aldo Moro; 4) NEXTGENERATIONEU (NGEU) funded by the Ministry of University and Research (MUR), National Recovery and Resilience Plan (NRRP), project MNESYS (PE0000006) – A Multiscale integrated approach to the study of the nervous system in health and disease (DD 1553, 11.10.2022); 5) NEXTGENERA-TIONEU (NGEU) funded by the Ministry of University and Research (MUR), National Re-covery and Resilience Plan (NRRP), project CN00000041 - National Center for Gene Therapy and Drugs based on RNA Technology (DD n.1035, 17.06.2022). The Italian Minister of Health have funded this work, Ricerca Corrente program 2022-2024 to L.S.
Institutional Review Board Statement
“Not applicable.”.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data are available from the authors on request.
Acknowledgments
The authors thank Prof. N Zizzo for his collaboration.
Conflicts of Interest
The authors declare no conflict of interest.
References
- A. Wawrzkiewicz-Jałowiecka, P. Trybek, B. Dworakowska, and Ł. Machura, “Multifractal Properties of BK Channel Currents in Human Glioblastoma Cells.,” J. Phys. Chem. B, vol. 124, no. 12, pp. 2382–2391, Mar. 2020. [CrossRef]
- K. Aldape et al., “Challenges to curing primary brain tumours.,” Nat. Rev. Clin. Oncol., vol. 16, no. 8, pp. 509–520, Aug. 2019. [CrossRef]
- L. Catacuzzeno, L. Sforna, V. Esposito, C. Limatola, and F. Franciolini, “Ion Channels in Glioma Malignancy BT - Transportome Malfunction in the Cancer Spectrum: Ion Transport in Tumor Biology,” C. Stock and L. A. Pardo, Eds. Cham: Springer International Publishing, 2021, pp. 223–267.
- A. Varricchio, S. A. Ramesh, and A. J. Yool, “Novel Ion Channel Targets and Drug Delivery Tools for Controlling Glioblastoma Cell Invasiveness,” International Journal of Molecular Sciences, vol. 22, no. 21. 2021. [CrossRef]
- M. C. Papadopoulos and S. Saadoun, “Key roles of aquaporins in tumor biology,” Biochim. Biophys. Acta - Biomembr., vol. 1848, no. 10, Part B, pp. 2576–2583, 2015. [CrossRef]
- J. Xia et al., “Ion channels or aquaporins as novel molecular targets in gastric cancer.,” Mol. Cancer, vol. 16, no. 1, p. 54, Mar. 2017. [CrossRef]
- S. F. Pedersen and C. Stock, “Ion channels and transporters in cancer: pathophysiology, regulation, and clinical potential.,” Cancer research, vol. 73, no. 6. United States, pp. 1658–1661, Mar. 2013. [CrossRef]
- K. L. Black et al., “Different effects of KCa and KATP agonists on brain tumor permeability between syngeneic and allogeneic rat models.,” Brain Res., vol. 1227, pp. 198–206, Aug. 2008. [CrossRef]
- N. S. Ningaraj, U. T. Sankpal, D. Khaitan, E. A. Meister, and T. Vats, “Activation of KATP channels increases anticancer drug delivery to brain tumors and survival.,” Eur. J. Pharmacol., vol. 602, no. 2–3, pp. 188–193, Jan. 2009. [CrossRef]
- R. E. Day et al., “Human aquaporins: regulators of transcellular water flow.,” Biochim. Biophys. Acta, vol. 1840, no. 5, pp. 1492–1506, May 2014. [CrossRef]
- F. Lang et al., “Functional significance of cell volume regulatory mechanisms.,” Physiol. Rev., vol. 78, no. 1, pp. 247–306, Jan. 1998. [CrossRef]
- E. A. Nagelhus, T. M. Mathiisen, and O. P. Ottersen, “Aquaporin-4 in the central nervous system: cellular and subcellular distribution and coexpression with KIR4.1.,” Neuroscience, vol. 129, no. 4, pp. 905–913, 2004. [CrossRef]
- L. Simone et al., “AQP4 Aggregation State Is a Determinant for Glioma Cell Fate,” Cancer Res., vol. 79, no. 9, pp. 2182–2194, May 2019. [CrossRef]
- M. Amiry-Moghaddam et al., “An alpha-syntrophin-dependent pool of AQP4 in astroglial end-feet confers bidirectional water flow between blood and brain.,” Proc. Natl. Acad. Sci. U. S. A., vol. 100, no. 4, pp. 2106–2111, Feb. 2003. [CrossRef]
- C. Palazzo et al., “AQP4ex is crucial for the anchoring of AQP4 at the astrocyte end-feet and for neuromyelitis optica antibody binding.,” Acta Neuropathol. Commun., vol. 7, no. 1, p. 51, Apr. 2019. [CrossRef]
- M. De Bellis et al., “Translational readthrough generates new astrocyte AQP4 isoforms that modulate supramolecular clustering, glial endfeet localization, and water transport.,” Glia, vol. 65, no. 5, pp. 790–803, May 2017. [CrossRef]
- H. Wolburg, S. Noell, P. Fallier-Becker, A. F. Mack, and K. Wolburg-Buchholz, “The disturbed blood-brain barrier in human glioblastoma.,” Mol. Aspects Med., vol. 33, no. 5–6, pp. 579–589, 2012. [CrossRef]
- F. Maqoud et al., “Cell Cycle Regulation by Ca (2+)-Activated K(+) (BK) Channels Modulators in SH-SY5Y Neuroblastoma Cells.,” Int. J. Mol. Sci., vol. 19, no. 8, Aug. 2018. [CrossRef]
- T. Ding et al., “Role of aquaporin-4 in the regulation of migration and invasion of human glioma cells.,” Int. J. Oncol., vol. 38, no. 6, pp. 1521–1531, Jun. 2011. [CrossRef]
- T. Ding et al., “Knockdown a water channel protein, aquaporin 4, induced glioblastoma cell apoptosis.,” PLoS One, vol. 8, no. 8, p. e66751, 2013. [CrossRef]
- P. Fallier-Becker, M. Nieser, U. Wenzel, R. Ritz, and S. Noell, “Is Upregulation of Aquaporin 4-M1 Isoform Responsible for the Loss of Typical Orthogonal Arrays of Particles in Astrocytomas?,” Int. J. Mol. Sci., vol. 17, no. 8, Jul. 2016. [CrossRef]
- L. Simone et al., “AQP4-dependent glioma cell features affect the phenotype of surrounding cells via extracellular vesicles,” Cell Biosci., vol. 12, no. 1, p. 150, 2022. [CrossRef]
- L. Zúñiga, A. Cayo, W. González, C. Vilos, and R. Zúñiga, “Potassium Channels as a Target for Cancer Therapy: Current Perspectives.,” OncoTargets and Therapy, vol. 15. pp. 783–797, 2022. [CrossRef]
- E. Bates, “Ion channels in development and cancer.,” Annu. Rev. Cell Dev. Biol., vol. 31, pp. 231–247, 2015. [CrossRef]
- F. Maqoud, R. Scala, M. Hoxha, B. Zappacosta, and D. Tricarico, “ATP-sensitive Potassium Channel Subunits in Neuroinflammation: Novel Drug Targets in Neurodegenerative Disorders.,” CNS Neurol. Disord. Drug Targets, vol. 21, no. 2, pp. 130–149, 2022. [CrossRef]
- J. S. Amberger and A. Hamosh, “Searching Online Mendelian Inheritance in Man (OMIM): A Knowledgebase of Human Genes and Genetic Phenotypes.,” Curr. Protoc. Bioinforma., vol. 58, pp. 1.2.1-1.2.12, Jun. 2017. [CrossRef]
- X. Huang and L. Y. Jan, “Targeting potassium channels in cancer.,” J. Cell Biol., vol. 206, no. 2, pp. 151–162, Jul. 2014. [CrossRef]
- H. Ouadid-Ahidouch and A. Ahidouch, “K+ channel expression in human breast cancer cells: involvement in cell cycle regulation and carcinogenesis.,” J. Membr. Biol., vol. 221, no. 1, pp. 1–6, Jan. 2008. [CrossRef]
- Z. Wang, “Roles of K+ channels in regulating tumour cell proliferation and apoptosis.,” Pflugers Arch., vol. 448, no. 3, pp. 274–286, Jun. 2004. [CrossRef]
- M. Levy et al., “Aquaporin-4 Expression Patterns in Glioblastoma Pre-Chemoradiation and at Time of Suspected Progression.,” Cancer Invest., vol. 37, no. 2, pp. 67–72, 2019. [CrossRef]
- W.-J. Zhao, W. Zhang, G.-L. Li, Y. Cui, Z.-F. Shi, and F. Yuan, “Differential expression of MMP-9 and AQP4 in human glioma samples.,” Folia Neuropathol., vol. 50, no. 2, pp. 176–186, 2012.
- S. Noell, P. Fallier-Becker, A. F. Mack, M. Hoffmeister, R. Beschorner, and R. Ritz, “Water Channels Aquaporin 4 and -1 Expression in Subependymoma Depends on the Localization of the Tumors.,” PLoS One, vol. 10, no. 6, p. e0131367, 2015. [CrossRef]
- S. Noell et al., “Dynamics of expression patterns of AQP4, dystroglycan, agrin and matrix metalloproteinases in human glioblastoma.,” Cell Tissue Res., vol. 347, no. 2, pp. 429–441, Feb. 2012. [CrossRef]
- E. J. Suero Molina et al., “Aquaporin-4 in glioma and metastatic tissues harboring 5-aminolevulinic acid-induced porphyrin fluorescence.,” Clin. Neurol. Neurosurg., vol. 115, no. 10, pp. 2075–2081, Oct. 2013. [CrossRef]
- K.-J. Mou et al., “[AQP4 expression in the brains of patients with glioblastoma and its association with brain edema].,” Sichuan da xue xue bao. Yi xue ban = J. Sichuan Univ. Med. Sci. Ed., vol. 40, no. 4, pp. 651–654, Jul. 2009.
- O. Valente et al., “Alteration of the translational readthrough isoform AQP4ex induces redistribution and downregulation of AQP4 in human glioblastoma.,” Cell. Mol. Life Sci., vol. 79, no. 3, p. 140, Feb. 2022. [CrossRef]
- K. Zhou et al., “ABCC8 mRNA expression is an independent prognostic factor for glioma and can predict chemosensitivity,” Sci. Rep., vol. 10, no. 1, p. 12682, 2020. [CrossRef]
- A. Trapani et al., “A novel injectable formulation of 6-fluoro-l-DOPA imaging agent for diagnosis of neuroendocrine tumors and Parkinson’s disease.,” Int. J. Pharm., vol. 519, no. 1–2, pp. 304–313, Mar. 2017. [CrossRef]
- L. Huang, B. Li, W. Li, H. Guo, and F. Zou, “ATP-sensitive potassium channels control glioma cells proliferation by regulating ERK activity,” Carcinogenesis, vol. 30, no. 5, pp. 737–744, May 2009. [CrossRef]
- X. Liu, Y. Chang, P. H. Reinhart, H. Sontheimer, and Y. Chang, “Cloning and characterization of glioma BK, a novel BK channel isoform highly expressed in human glioma cells.,” J. Neurosci. Off. J. Soc. Neurosci., vol. 22, no. 5, pp. 1840–1849, Mar. 2002. [CrossRef]
- N. S. Ningaraj, U. T. Sankpal, D. Khaitan, E. A. Meister, and T. S. Vats, “Modulation of KCa channels increases anticancer drug delivery to brain tumors and prolongs survival in xenograft model,” Cancer Biol. Ther., vol. 8, no. 20, pp. 1924–1933, 2009. [CrossRef]
- R. Zhang et al., “Different sulfonylureas induce the apoptosis of proximal tubular epithelial cell differently via closing K(ATP) channel.,” Mol. Med., vol. 24, no. 1, p. 47, Sep. 2018. [CrossRef]
- J. H. Han, O. S. Kwon, J. H. Chung, K. H. Cho, H. C. Eun, and K. H. Kim, “Effect of minoxidil on proliferation and apoptosis in dermal papilla cells of human hair follicle.,” J. Dermatol. Sci., vol. 34, no. 2, pp. 91–98, Apr. 2004. [CrossRef]
- R. Scala et al., “Zoledronic Acid Blocks Overactive Kir6.1/SUR2-Dependent K(ATP) Channels in Skeletal Muscle and Osteoblasts in a Murine Model of Cantú Syndrome.,” Cells, vol. 12, no. 6, Mar. 2023. [CrossRef]
- R. Scala et al., “Consequences of SUR2[A478V] Mutation in Skeletal Muscle of Murine Model of Cantu Syndrome.,” Cells, vol. 10, no. 7, Jul. 2021. [CrossRef]
- D. Tricarico, M. Barbieri, L. Antonio, P. Tortorella, F. Loiodice, and D. C. Camerino, “Dualistic actions of cromakalim and new potent 2H-1,4-benzoxazine derivatives on the native skeletal muscle K ATP channel.,” Br. J. Pharmacol., vol. 139, no. 2, pp. 255–262, May 2003. [CrossRef]
- F. Maqoud et al., “Zoledronic Acid as a Novel Dual Blocker of KIR6.1 / 2-SUR2 Subunits of ATP-Sensitive K + Channels: Role in the Adverse Drug Reactions,” pp. 1–17, 2021.
- Y.-G. Ma et al., “Activation of BKCa Channels in Zoledronic Acid-Induced Apoptosis of MDA-MB-231 Breast Cancer Cells,” PLoS One, vol. 7, no. 5, p. e37451, May 2012, [online]. Available: . [CrossRef]
- R. Scala et al., “Zoledronic Acid Modulation of TRPV1 Channel Currents in Osteoblast Cell Line and Native Rat and Mouse Bone Marrow-Derived Osteoblasts: Cell Proliferation and Mineralization Effect.,” Cancers (Basel)., vol. 11, no. 2, Feb. 2019. [CrossRef]
- T. J. Polascik and V. Mouraviev, “Zoledronic acid in the management of metastatic bone disease.,” Ther. Clin. Risk Manag., vol. 4, no. 1, pp. 261–268, Feb. 2008. [CrossRef]
- S. Pozzi and N. Raje, “The role of bisphosphonates in multiple myeloma: mechanisms, side effects, and the future.,” Oncologist, vol. 16, no. 5, pp. 651–662, 2011. [CrossRef]
- J. A. N. PONTÉN and E. H. MACINTYRE, “LONG TERM CULTURE OF NORMAL AND NEOPLASTIC HUMAN GLIA,” Acta Pathol. Microbiol. Scand., vol. 74, no. 4, pp. 465–486, Sep. 1968. [CrossRef]
- D. Tricarico et al., “Structural nucleotide analogs are potent activators/inhibitors of pancreatic beta cell KATP channels: an emerging mechanism supporting their use as antidiabetic drugs.,” J. Pharmacol. Exp. Ther., vol. 340, no. 2, pp. 266–276, Feb. 2012. [CrossRef]
- R. Scala et al., “Bisphosphonates Targeting Ion Channels and Musculoskeletal Effects.,” Front. Pharmacol., vol. 13, p. 837534, 2022. [CrossRef]
- R. Scala et al., “Pathophysiological Consequences of KATP Channel Overactivity and Pharmacological Response to Glibenclamide in Skeletal Muscle of a Murine Model of Cantù Syndrome,” Frontiers in Pharmacology , vol. 11. 2020, [Online]. Available: https://www.frontiersin.org/articles/10.3389/fphar.2020.604885.
- N. Zizzo et al., “Thymidine Phosphorylase Expression and Microvascular Density Correlation Analysis in Canine Mammary Tumor: Possible Prognostic Factor in Breast Cancer.,” Front. Vet. Sci., vol. 6, p. 368, 2019. [CrossRef]
- F. Maqoud et al., “Immunohistochemical, pharmacovigilance, and omics analyses reveal the involvement of ATP-sensitive K+ channel subunits in cancers: role in drug–disease interactions,” Frontiers in Pharmacology, vol. 14. 2023, [Online]. Available: https://www.frontiersin.org/articles/10.3389/fphar.2023.1115543.
- F. Maqoud, E. Vacca, and M. Tommaseo-Ponzetta, “From Morocco to Italy: How Women’s Bodies Reflect their Change of Residence.,” Coll. Antropol., vol. 40, no. 1, pp. 9–15, Apr. 2016.
- E. McCoy and H. Sontheimer, “Expression and function of water channels (aquaporins) in migrating malignant astrocytes.,” Glia, vol. 55, no. 10, pp. 1034–1043, Aug. 2007. [CrossRef]
- C. J. Lingle, P. L. Martinez-Espinosa, A. Yang-Hood, L. E. Boero, S. Payne, and D. Persic, et al.,"LRRC52 regulates BK channel function and localization in mouse cochlear inner hair cells", Proc Natl Acad Sci U S A., vol. 10, 116(37) pp. 18397-18403, Sept. 2019. Epub 2019 Aug 26. [CrossRef]
- S. R. Malwal et al., “Bisphosphonate-Generated ATP-Analogs Inhibit Cell Signaling Pathways,” J. Am. Chem. Soc., 2018. [CrossRef]
- L. Abdul Kadir, M. Stacey, and R. Barrett-Jolley, “Emerging Roles of the Membrane Potential: Action Beyond the Action Potential,” Frontiers in Physiology, vol. 9. 2018, [Online]. Available: https://www.frontiersin.org/articles/10.3389/fphys.2018.01661.
- R. Quadri et al., “Haspin regulates Ras localization to promote Cdc24-driven mitotic depolarization,” Cell Discov., vol. 6, no. 1, p. 42, 2020. [CrossRef]
- C. Baumgartner, “The world’s first digital cell twin in cancer electrophysiology: a digital revolution in cancer research” J. Exp. Clin. Cancer Res., vol. 41, no. 1, p. 298, 2022. [CrossRef]
- J. Ocampo-Garza, J. Griggs, and A. Tosti, “New drugs under investigation for the treatment of alopecias.,” Expert Opin. Investig. Drugs, vol. 28, no. 3, pp. 275–284, Mar. 2019. [CrossRef]
- D. Tricarico, L. Montanari, and D. Conte Camerino, “Involvement of 3Na+/2K+ ATP-ase and Pi-3 kinase in the response of skeletal muscle ATP-sensitive K+ channels to insulin.,” Neuromuscul. Disord., vol. 13, no. 9, pp. 712–719, Nov. 2003. [CrossRef]
- D. Tricarico, R. Mallamaci, M. Barbieri, and D. C. Camerino, “Modulation of ATP-Sensitive K+Channel by Insulin in Rat Skeletal Muscle Fibers,” Biochem. Biophys. Res. Commun., vol. 232, no. 2, pp. 536–539, 1997. [CrossRef]
- P. Marques et al., “Emergence of Pituitary Adenoma in a Child during Surveillance: Clinical Challenges and the Family Members’ View in an AIP Mutation-Positive Family,” Int. J. Endocrinol., vol. 2018, p. 8581626, 2018. [CrossRef]
- P. Marques et al., “Cantu syndrome with coexisting familial pituitary adenoma.,” Endocrine, vol. 59, no. 3, pp. 677–684, Mar. 2018. [CrossRef]
- D. Tricarico, D. Conte Camerino, S. Govoni, and S. H. Bryant, “Modulation of rat skeletal muscle chloride channels by activators and inhibitors of protein kinase C,” Pflügers Arch., vol. 418, no. 5, pp. 500–503, 1991. [CrossRef]
- A. Mele, G. M. Camerino, S. Calzolaro, M. Cannone, D. Conte, and D. Tricarico, “Dual response of the KATP channels to staurosporine: a novel role of SUR2B, SUR1 and Kir6.2 subunits in the regulation of the atrophy in different skeletal muscle phenotypes.,” Biochem. Pharmacol., vol. 91, no. 2, pp. 266–275, Sep. 2014. [CrossRef]
- D. Tricarico, M. Barbieri, and D. C. Camerino, “Taurine blocks ATP-sensitive potassium channels of rat skeletal muscle fibres interfering with the sulphonylurea receptor.,” Br. J. Pharmacol., vol. 130, no. 4, pp. 827–834, Jun. 2000. [CrossRef]
- D. Tricarico et al., “The biophysical and pharmacological characteristics of skeletal muscle ATP-sensitive K+ channels are modified in K+-depleted rat, an animal model of hypokalemic periodic paralysis.,” Mol. Pharmacol., vol. 54, no. 1, pp. 197–206, Jul. 1998. [CrossRef]
- A. Mele, M. Buttiglione, G. Cannone, F. Vitiello, D. C. Camerino, and D. Tricarico, “Opening/blocking actions of pyruvate kinase antibodies on neuronal and muscular KATP channels.,” Pharmacol. Res., vol. 66, no. 5, pp. 401–408, Nov. 2012. [CrossRef]
- K. Castillo et al., “The bisphosphonate zoledronic acid is a TRPV1 channel inhibitor,” Biophys. J., vol. 122, no. 3, p. 108a, Feb. 2023. [CrossRef]
- D. Tricarico, R. Capriulo, and D. C. Camerino, “Involvement of KCa2+ channels in the local abnormalities and hyperkalemia following the ischemia-reperfusion injury of rat skeletal muscle,” Neuromuscul. Disord., vol. 12, no. 3, pp. 258–265, 2002. [CrossRef]
- M. M. Dinardo, G. Camerino, A. Mele, R. Latorre, D. Conte Camerino, and D. Tricarico, “Splicing of the rSlo gene affects the molecular composition and drug response of Ca2+-activated K+ channels in skeletal muscle.,” PLoS One, vol. 7, no. 7, p. e40235, 2012. [CrossRef]
- D. Tricarico et al., “Acetazolamide prevents vacuolar myopathy in skeletal muscle of K (+) -depleted rats.,” Br. J. Pharmacol., vol. 154, no. 1, pp. 183–190, May 2008. [CrossRef]
- D. Tricarico et al., “Emerging role of calcium-activated potassium channel in the regulation of cell viability following potassium ions challenge in HEK293 cells and pharmacological modulation.,” PLoS One, vol. 8, no. 7, p. e69551, 2013. [CrossRef]
- A. Michelucci, L. Sforna, A. Di Battista, F. Franciolini, and L. Catacuzzeno, “Ca (2+) -activated K(+) channels regulate cell volume in human glioblastoma cells.,” J. Cell. Physiol., vol. 238, no. 9, pp. 2120–2134, Sep. 2023. [CrossRef]
- P. Rosa et al., “Overexpression of Large-Conductance Calcium-Activated Potassium Channels in Human Glioblastoma Stem-Like Cells and Their Role in Cell Migration.,” J. Cell. Physiol., vol. 232, no. 9, pp. 2478–2488, Sep. 2017. [CrossRef]
Figure 1.
Expression of AQP4 in U87 cells. A: Epifluorescence images of U87 WT and U87 expressing M1 AQP4-tetramers or M23 AQP4-OAPs. AQP4 staining is shown in red and DAPI in blue. Scale bar 20 μm. B: Immunoblot detection of AQP4 expression levels in U87 cells transfected with M1-AQP4 (AQP4-tetramers) and M23-AQP4 (AQP4-OAPs). GFAP and GAPDH were used to normalize for equal loading.
Figure 1.
Expression of AQP4 in U87 cells. A: Epifluorescence images of U87 WT and U87 expressing M1 AQP4-tetramers or M23 AQP4-OAPs. AQP4 staining is shown in red and DAPI in blue. Scale bar 20 μm. B: Immunoblot detection of AQP4 expression levels in U87 cells transfected with M1-AQP4 (AQP4-tetramers) and M23-AQP4 (AQP4-OAPs). GFAP and GAPDH were used to normalize for equal loading.
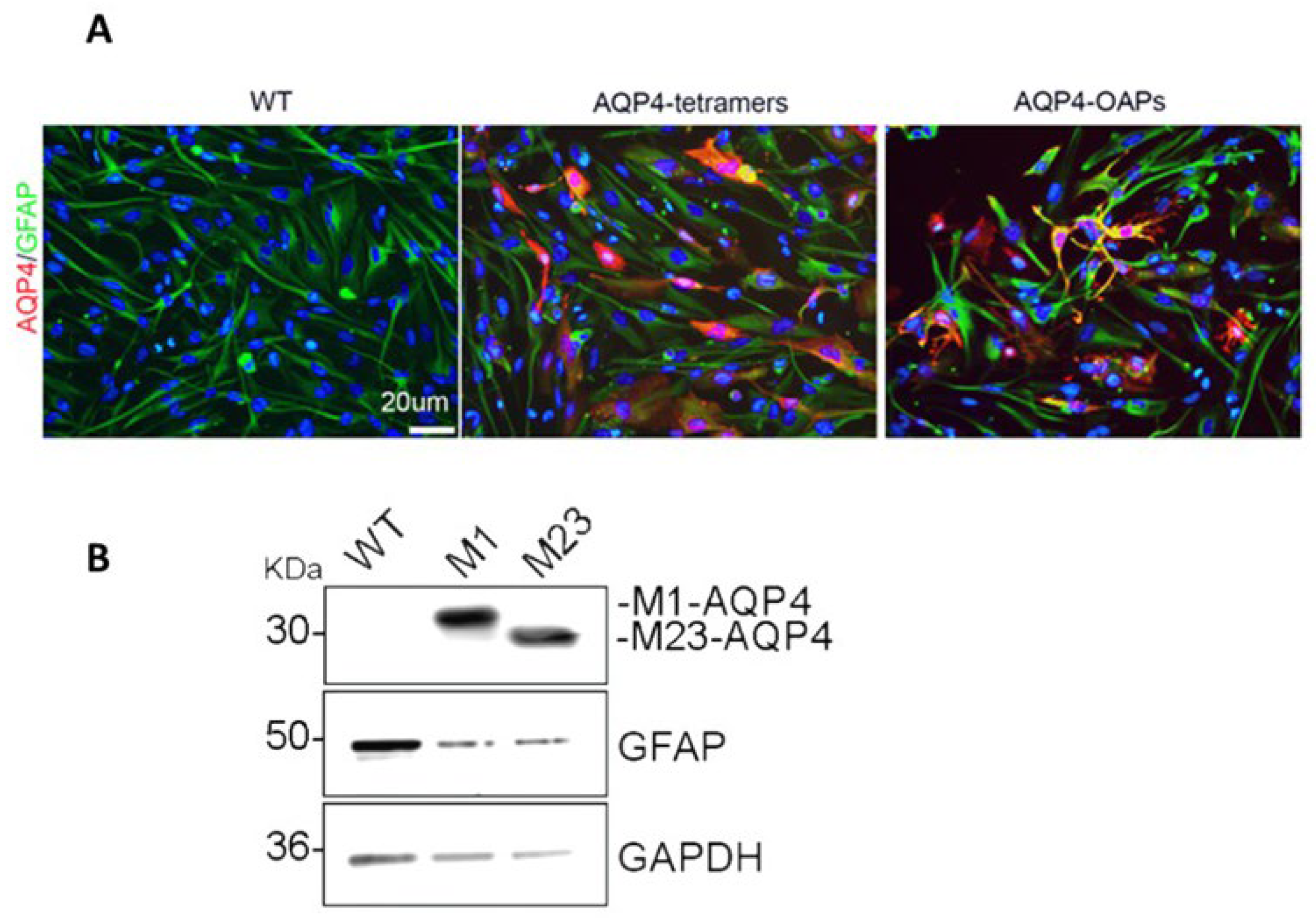
Figure 2.
Characterization of inward and outward macroscopic K+ ion currents recorded in U87 wt cells. The currents were recorded using a whole cell configuration under physiological concentration of K + ions in the bath and pipette and were obtained in response to voltage pulses from −120 to +120 mV in increments of 20 mV, starting at HP = −60 mV (Vm) and intracellular free Ca2+ ions (1.6x10-6M). (a) TEA (5 × 10−3 M) suppressed the outward K+ ion currents, TEA / Ba2+ (5 × 10−3 M) fully suppressed also the inward K+ ion currents. (b) The selective BK channel blocker IbTX (4 × 10-7 M) reduced the outward K+ ion currents that were fully reduced by TEA (5 × 10−3 M). (c) KATP currents recorded in U87wt cells in low intracellular ATP 1x10-3M. Glibenclamide (5x10-8 M) suppressed the inward currents suggesting the presence of KATP channels in the cells. Cells of the same size were selected for patch clamp experiments. Each point represented the mean ± SEM (N patches = 10–15). (d) Percentage of blocking of K+ ion currents in the presence of the antagonists with respect to the control condition at 60 mV and -60 mV (Vm).
Figure 2.
Characterization of inward and outward macroscopic K+ ion currents recorded in U87 wt cells. The currents were recorded using a whole cell configuration under physiological concentration of K + ions in the bath and pipette and were obtained in response to voltage pulses from −120 to +120 mV in increments of 20 mV, starting at HP = −60 mV (Vm) and intracellular free Ca2+ ions (1.6x10-6M). (a) TEA (5 × 10−3 M) suppressed the outward K+ ion currents, TEA / Ba2+ (5 × 10−3 M) fully suppressed also the inward K+ ion currents. (b) The selective BK channel blocker IbTX (4 × 10-7 M) reduced the outward K+ ion currents that were fully reduced by TEA (5 × 10−3 M). (c) KATP currents recorded in U87wt cells in low intracellular ATP 1x10-3M. Glibenclamide (5x10-8 M) suppressed the inward currents suggesting the presence of KATP channels in the cells. Cells of the same size were selected for patch clamp experiments. Each point represented the mean ± SEM (N patches = 10–15). (d) Percentage of blocking of K+ ion currents in the presence of the antagonists with respect to the control condition at 60 mV and -60 mV (Vm).
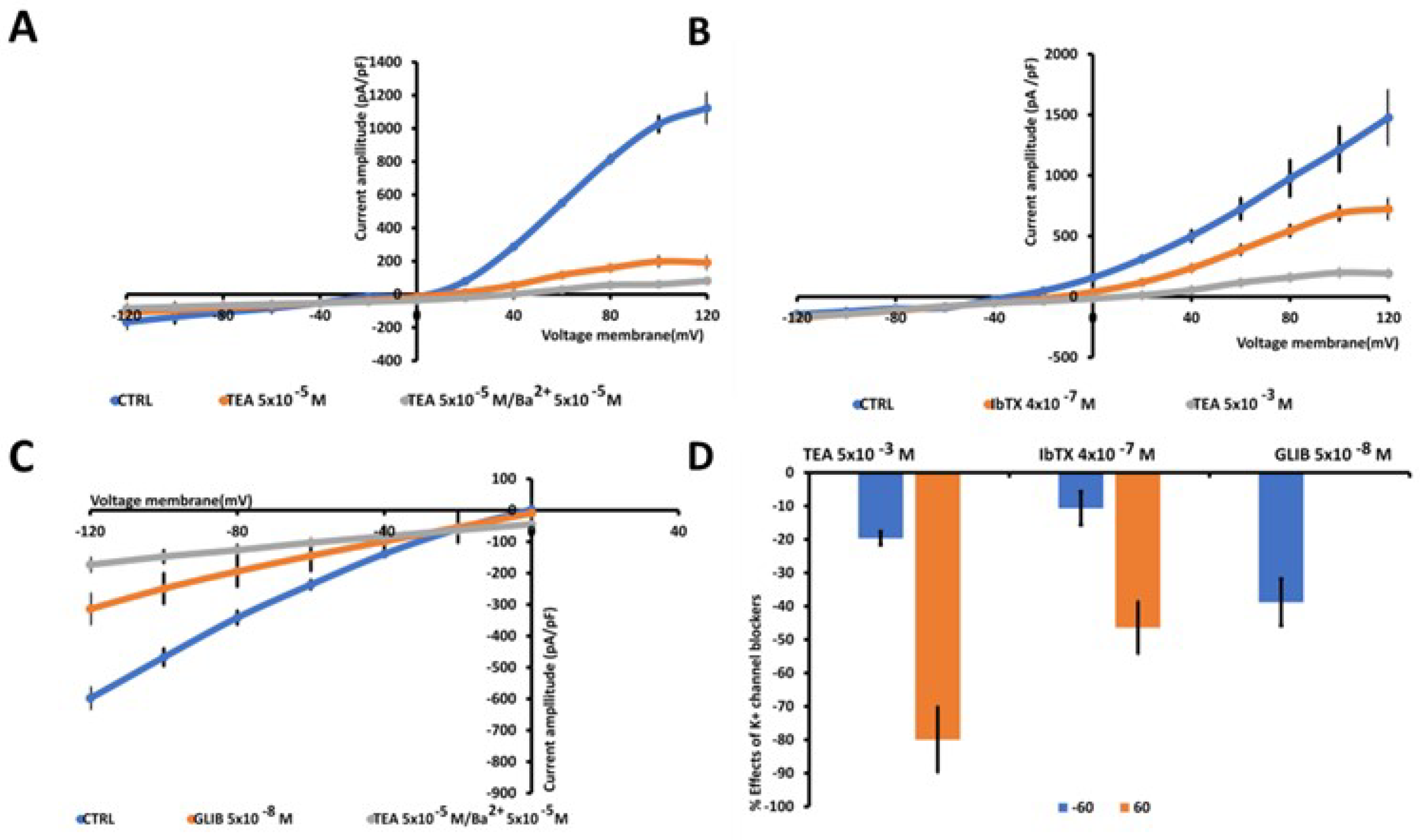
Figure 3.
Effects of antagonists and agonists of the K+ channels on the proliferation of U87wt cells. *P<0.05 and **P<0.01 when compared to the experimental group not treated after 48 hours of incubation. Each point represented the mean ± SEM.
Figure 3.
Effects of antagonists and agonists of the K+ channels on the proliferation of U87wt cells. *P<0.05 and **P<0.01 when compared to the experimental group not treated after 48 hours of incubation. Each point represented the mean ± SEM.
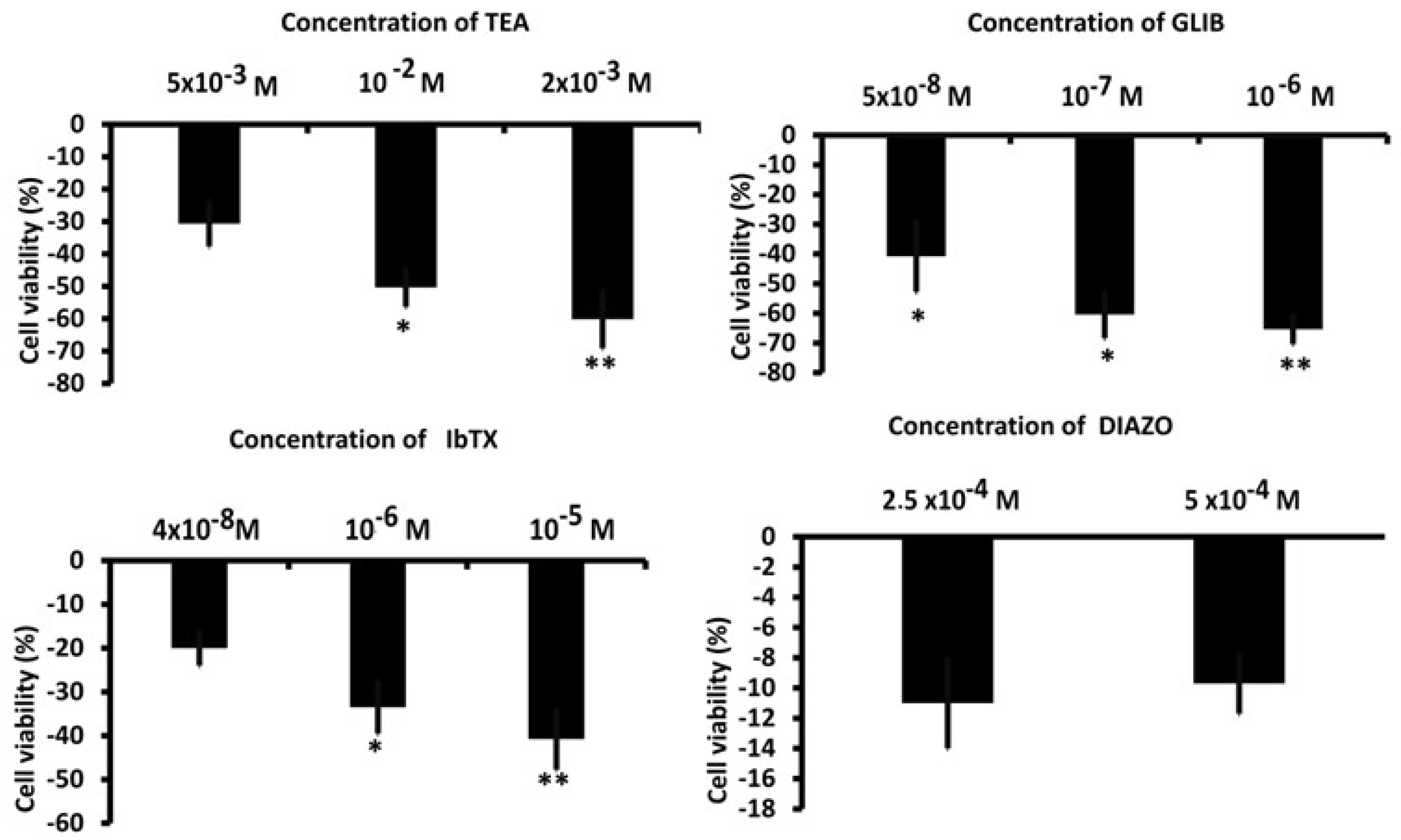
Figure 4.
Characterization of macroscopic inward and outward K+ ion currents recorded in U87 wt cells and after transfection with M1 AQP4-tetramer or M23 AQP4-OAP. Whole cell currents were recorded under physiological concentration of K+ ions in the bath and pipette and were obtained in response to voltage pulses from −120 to +120 mV in 20 mV increments, starting at HP = −60 mV (Vm ) (a) Macroscopic K+ ion currents recorded in AQP4-OAPs transfected U87 cells, the presence of AQP4-OAPs caused a significant increase in the inward and outward currents that were reduced by TEA and, TEA and BaCl2. (b) Macroscopic K+ ion currents recorded in AQP4-tetramer -transfected U87 cells, the presence of M1 AQP4-tetramers caused a significant increase of the inward currents, however the outward K+ ion currents were reduced in the amplitude. (c) The presence of M23 AQP4-OAPs caused a large increase of the currents at negative and positive membrane potentials vs not trasfected cell. Instead the presence of AQP4-tetramers led to an increase of the currents at negative membrane potentials, conversely the outward K+ ion currents decreased at positive membrane potentials. Data were pooled from N patches = 10–12. (d) Percentage of reduction of the K+ ion currents in the presence of the antagonist TEA with respect to the control condition at 60 mV and -60 mV (Vm).*P<0.05 when compared to the group U87 untransfected (wt). Each point represents the mean ± SEM.
Figure 4.
Characterization of macroscopic inward and outward K+ ion currents recorded in U87 wt cells and after transfection with M1 AQP4-tetramer or M23 AQP4-OAP. Whole cell currents were recorded under physiological concentration of K+ ions in the bath and pipette and were obtained in response to voltage pulses from −120 to +120 mV in 20 mV increments, starting at HP = −60 mV (Vm ) (a) Macroscopic K+ ion currents recorded in AQP4-OAPs transfected U87 cells, the presence of AQP4-OAPs caused a significant increase in the inward and outward currents that were reduced by TEA and, TEA and BaCl2. (b) Macroscopic K+ ion currents recorded in AQP4-tetramer -transfected U87 cells, the presence of M1 AQP4-tetramers caused a significant increase of the inward currents, however the outward K+ ion currents were reduced in the amplitude. (c) The presence of M23 AQP4-OAPs caused a large increase of the currents at negative and positive membrane potentials vs not trasfected cell. Instead the presence of AQP4-tetramers led to an increase of the currents at negative membrane potentials, conversely the outward K+ ion currents decreased at positive membrane potentials. Data were pooled from N patches = 10–12. (d) Percentage of reduction of the K+ ion currents in the presence of the antagonist TEA with respect to the control condition at 60 mV and -60 mV (Vm).*P<0.05 when compared to the group U87 untransfected (wt). Each point represents the mean ± SEM.
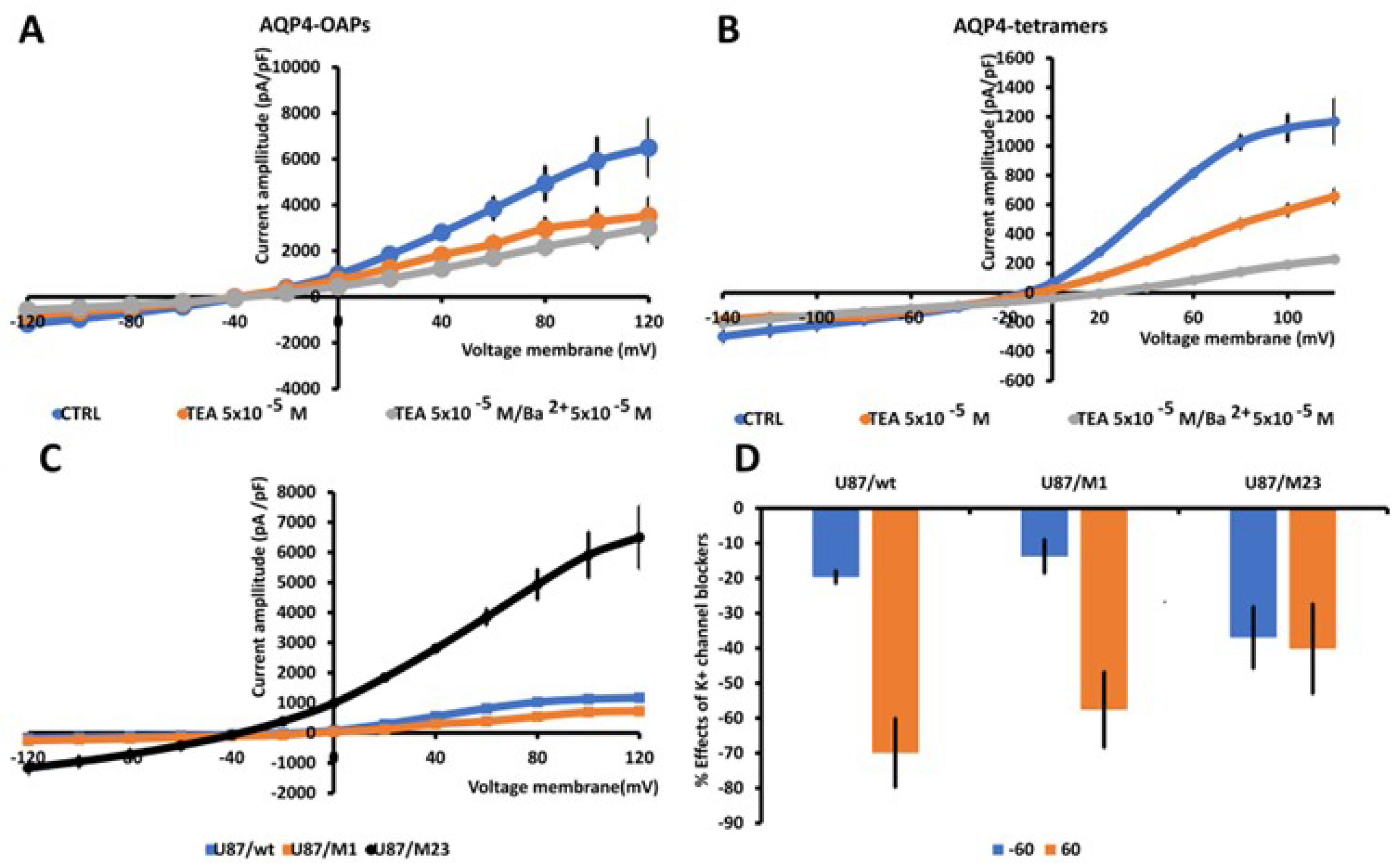
Figure 5.
Expression profile of KCNMA1, KCNJ11, KCNJ8, ABCC8, ABCC9 and TRPV1 genes in U87 glioma cells in the presence of the malignant M1 AQP4-tetramers and M23 AQP4-OAPs aggregation vs wt condition.
Figure 5.
Expression profile of KCNMA1, KCNJ11, KCNJ8, ABCC8, ABCC9 and TRPV1 genes in U87 glioma cells in the presence of the malignant M1 AQP4-tetramers and M23 AQP4-OAPs aggregation vs wt condition.
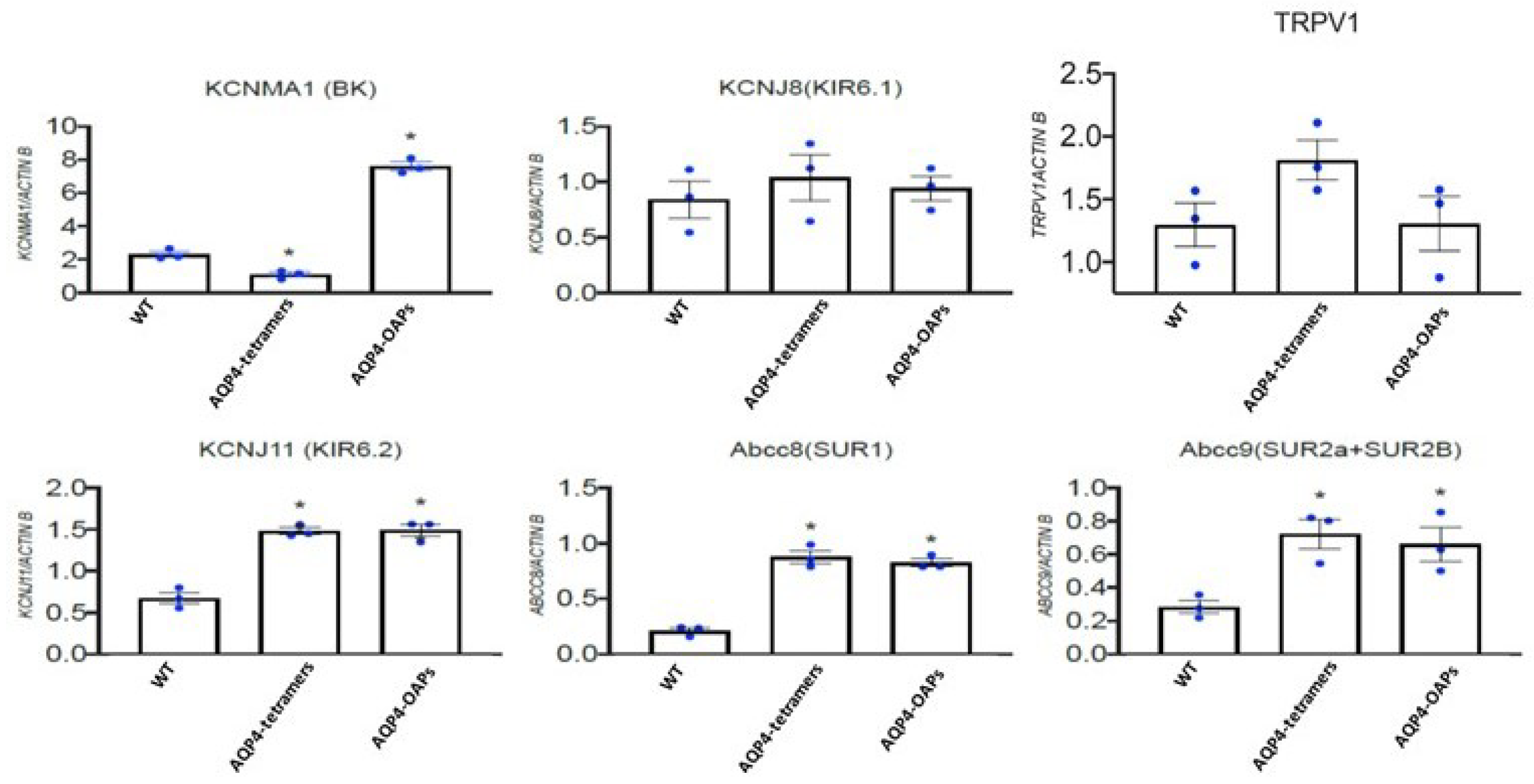
Figure 6.
Effect of pharmacological inhibition of the Kir6.2-SUR2 channel activity with zoledronic acid on AQP4 expressing U87 cells. (a) Epifluorescence images of U87 cells expressing AQP4-tetramers or AQP4-OAPs treated with diazoxide or zoledronic acid. AQP4 staining is shown in red and DAPI for nuclear staining in blue. Phalloidin (in green) was used to visualized F-actin. The white arrowheads indicate the round-shaped cells, the arrow indicate irregular-shaped cells and the blue arrowhead the apoptotic beads. Scale bar 100 μm. (b top) Drawing/Diagram showing the morphological change of U87 cells expressing AQP4-OAPs after pharmacological treatment with zoledronic acid according Coffin hypothesis and relative epifluorescence images. (b bottom) Epifluorescence images of U87 expressing AQP4-OAPs treated with zoledronic showing the irregular-shaped cell, the round-shaped cell and the apoptopodia. AQP4 staining is shown in red. Phalloidin (in green) was used to visualized F-actin. Scale bar 10 µm. Histobar showing the percentage of cells with round-shaped cells per fields calculated for U87, cells expressing and AQP4-OAPs after treatment with zoledronic acid (inhibitor) or control conditions (vehicle) or with diazoxide (agonist). Values are expressed as mean + SEM of percentage of cells with altered cell morphology out of the total number of transfected cells per field. **P < 0.005; (c top) Representative image of expressing AQP4-OAPs in control condition or after treated with zoledronic or diazoxide as indicated and stained with DAPI to visualize Nuclei. Scale bar 50 µm. (c bottom) Dot plot showing the analysis of % of condensed nuclei for field and the nuclear area of images in A. Values are expressed in µm2 and represent mean ± SEM. ****P < 0.0005, n=3, two-way ANOVA/Tukey’s tests.
Figure 6.
Effect of pharmacological inhibition of the Kir6.2-SUR2 channel activity with zoledronic acid on AQP4 expressing U87 cells. (a) Epifluorescence images of U87 cells expressing AQP4-tetramers or AQP4-OAPs treated with diazoxide or zoledronic acid. AQP4 staining is shown in red and DAPI for nuclear staining in blue. Phalloidin (in green) was used to visualized F-actin. The white arrowheads indicate the round-shaped cells, the arrow indicate irregular-shaped cells and the blue arrowhead the apoptotic beads. Scale bar 100 μm. (b top) Drawing/Diagram showing the morphological change of U87 cells expressing AQP4-OAPs after pharmacological treatment with zoledronic acid according Coffin hypothesis and relative epifluorescence images. (b bottom) Epifluorescence images of U87 expressing AQP4-OAPs treated with zoledronic showing the irregular-shaped cell, the round-shaped cell and the apoptopodia. AQP4 staining is shown in red. Phalloidin (in green) was used to visualized F-actin. Scale bar 10 µm. Histobar showing the percentage of cells with round-shaped cells per fields calculated for U87, cells expressing and AQP4-OAPs after treatment with zoledronic acid (inhibitor) or control conditions (vehicle) or with diazoxide (agonist). Values are expressed as mean + SEM of percentage of cells with altered cell morphology out of the total number of transfected cells per field. **P < 0.005; (c top) Representative image of expressing AQP4-OAPs in control condition or after treated with zoledronic or diazoxide as indicated and stained with DAPI to visualize Nuclei. Scale bar 50 µm. (c bottom) Dot plot showing the analysis of % of condensed nuclei for field and the nuclear area of images in A. Values are expressed in µm2 and represent mean ± SEM. ****P < 0.0005, n=3, two-way ANOVA/Tukey’s tests.
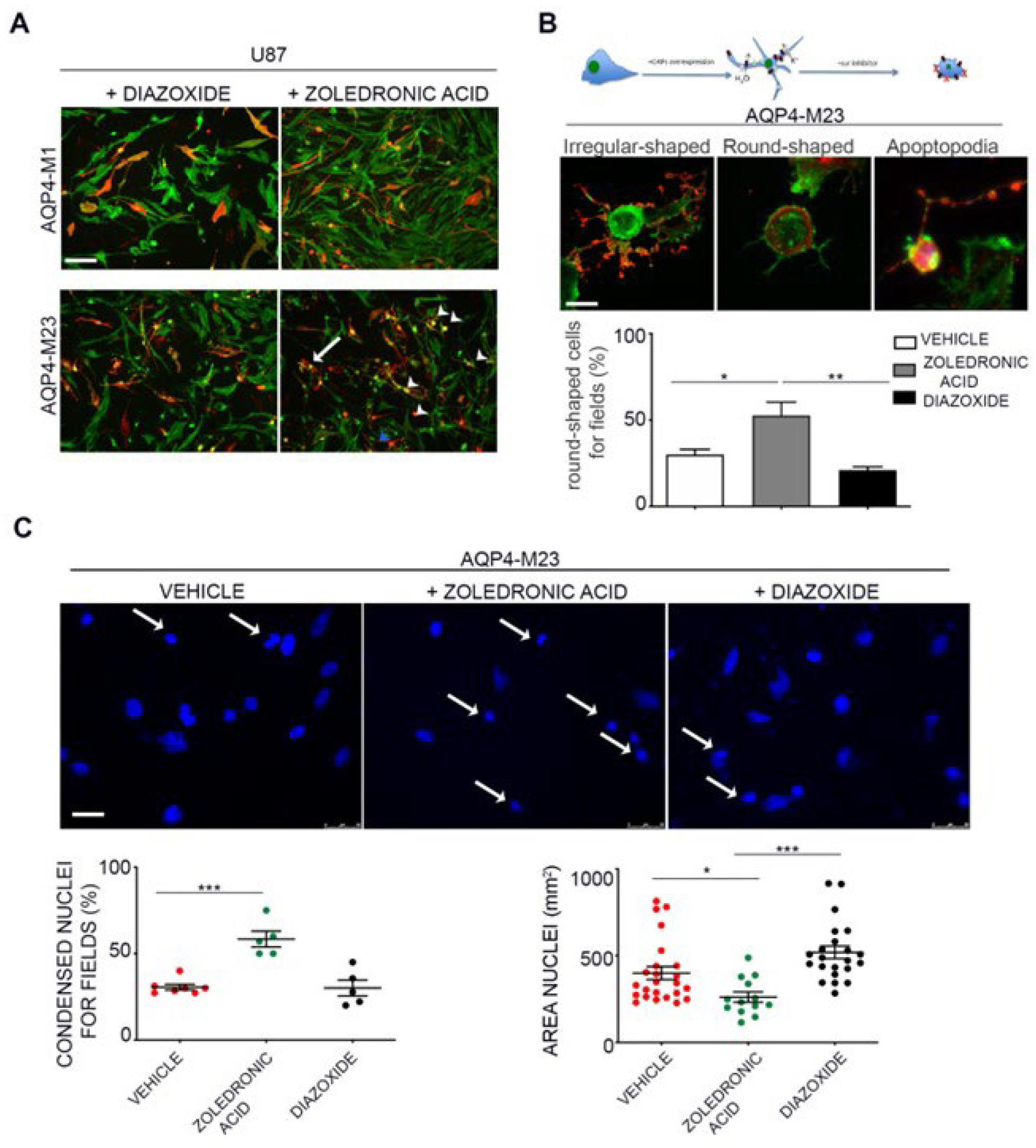
Figure 7.
RT PCR expression of the cyclin genes (E, A, B1) in the U87 cells following M23 AQP4-OAP transfection.
Figure 7.
RT PCR expression of the cyclin genes (E, A, B1) in the U87 cells following M23 AQP4-OAP transfection.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



