
Submitted:
08 July 2024
Posted:
09 July 2024
You are already at the latest version
Abstract
Recurrent Pregnancy Loss (RPL) affects 1-2% of women, and its triggering factors are unclear. Several studies have shown the vaginal, endometrial, and gut microbiota may play a role in RPL. A decrease in the quantity of Lactobacillus crispatus in local microbiota has been associated with an increase in local (vaginal and endometrial) inflammatory response and immune cell activation that leads to pregnancy loss. The inflammatory response may be triggered by gram-negative bacteria, lipopolysaccharides (LPS), viral infections, mycosis, or atypia (tumor growth). Bacterial structures and metabolites produced by microbiota could be involved in immune cell modulation and may be responsible for immune cell activation and molecular mimicry. Gut microbiota metabolic products may increase the amount of circulating pro-inflammatory lymphocytes, which, in turn, will migrate into vaginal or endometrial tissues. Local proinflammatory Th1 and Th17 subpopulations along with a decrease in Treg and tolerogenic NK cells are accountable for the increase in pregnancy loss. Local microbiota may modulate the local inflammatory response increasing pregnancy success. Analyzing local and gut microbiota may be necessary to characterize some RPL patients. Although oral supplementation of probiotics has not been shown to modify vaginal or endometrial microbiota, the metabolites produced by it may benefit patients. Lactobacillus crispatus transplantation into the vagina may enhance the required immune tolerogenic response to achieve a normal pregnancy. The effect of hormone stimulation and progesterone to maintain early pregnancy on microbiota has not been adequately studied, and more research is needed in this area. Well-designed clinical trials are needed to ascertain the benefit of microbiota modulation in RPL.
Keywords:
recurrent pregnancy loss (RPL)
; vaginal microbiota
; uterine microbiota
; gut microbiota
; probiotic supplementation
1. Introduction
Recurrent pregnancy loss (RPL) is defined as the prenancy loss before 20 weeks of gestation of two (US) or three (UK) consecutive clinical pregnancies. The loss must be documented by ultrasound or histopathology. This condition affects 1–2% of women of reproductive age [1,2,3] and is linked to genetic, anatomical, endocrine, autoimmune, and infectious factors [1,3]. Most cases of RPL lack clear etiology, and little is known about its associated factors [1,2,3].
There are two types of RPL: primary and secondary [3]. Primary RPL occurs in women who have never given birth to a live infant, while secondary RPL occurs in women who have given birth to a live infant. More research is required to define the differences between the two entities. It is generally assumed that the mechanisms of primary and secondary RPL differ, but this has not been proven.
Great efforts have been made to analyze vaginal and endometrial/uterine conditions in RPL [3]. The analysis of local tissue along with resident immune cells and the plausible role of infectious diseases in modifying local homeostasis has rendered exciting results [4,5,6]. Except for explainable problems with the spermatozoa, low amount, low migration, impaired function, and genetic mutations of the zygote/fetus, the rest mainly involve maternal physiological responses. In primary RPL, there is a need to understand the key elements that are involved in the increase of recurrent abortion; in secondary RPL, there is a need to know why, after a pregnancy, it has been impossible to achieve another normal pregnancy. Local factors may play a critical role in the process.
Microbiota are the diverse microbial communities that home in the human body. This comprises bacteria, archaea bacteria, fungi, viruses, and protists. These communities vary significantly in composition and function among different body sites and individuals. The changes in the microbiota (dysbiosis) are associated with various diseases [4]. Normal microbiota and the lack of inflammatory mediators due to bacterial, fungal, or viral infections or cancer are crucial to succeed in a normal pregnancy. Local microbiota plays a role in the defence of external pathogens and the modulation of local immune response [4,5,6,7]. Two key elements have been defined: the importance of zygote implantation and immune tolerogenic response to avoid fetal rejection [3,4,5,6,7]. In both permissive conditions, local microbiota plays a critical role against foreign pathogens, protecting tissues and promoting a vigilant response from local immune cells.
This brief overview will discuss the importance of genital and gut microbiota in RPL.
2. Immune Cells in the Female Reproductive Tract
Immune cells vary among different parts of the female reproductive tract [7,8]. In the vagina, contrary to expectation, T cells (CD4+ and CD8+ subpopulations, memory cells) predominate at around 50 % of the total leucocyte, NK cells are the second highest population, close to 20 %, the number of B cells is only 1 %, and the rest of the cells, approximately 30 %, are granulocytes and macrophages [7,8]. As a comparison, semen contains preferentially granulocytes and macrophages and a small amount of lymphocytes (~ 5 %) [9].
T cells (CD4+ and CD8+) are around 50 % of the leucocytes in the ecto and endocervix, with macrophages and NK cells representing around 12 % [7,8]. The rest are granulocytes and B lymphocytes. Finally, the cell type in the endometrium differs depending on the hormonal cycle. From the early follicular to the early secretory phase, the number of leucocytes (CD45+) in the endometrium remains low, but during the secretory phase, they increase about 5-fold [7]. Therefore, the total number of leukocytes peakes premenstrually. In the late secretory phase, NK cells (especially CD3-/CD56bright/CD16-) are predominant, compromising approximately 80% of CD45+ cells, while CD3+ T cells (predominantly CD8+) decrease to less than 10% [7]. The percentage of B cells is low in the proliferative and middle secretory phases and increases in the late secretory phase [7]. Macrophages, neutrophils, and eosinophils increase at the late secretory phase. Mast cells represent 1 -2% of the total endometrial cells during the menstrual cycle. T cells are high in the Fallopian tube, followed by granulocytes, NK cells, macrophages, and B cells [7]. In summary, in the endometrium, cells have high mobility depending on the hormonal cycle, which may change in case of infection. As expected, the immune cell response will differ according to the type (viral, bacterial, fungal), infection site, and the hormonal cycle stage [7,8].
It is also important to clarify that mucosal T cells (MAIT) and innate lymphoid cells will migrate to the reproductive tissues in the presence of infection [7,8,10,11,12,13,14]. Three types of innate lymphoid cells are precursors of Th1, Th2, and Th17/Th22, depending on the tissue milieu [11]. The role of immune cells, particularly uterine and decidual NK cells, has been reviewed before [13].
The role of interferon (IFN) signaling upon viral infection may condition endometrial local immune response [12]. Cell activation by Toll-like receptors or danger cell signals and inflammasome [10] activation negatively impacts RPL [11,12,13,14]. The production of local cytokines may partially explain the local response [16], and semen impacts local cytokine production [17].
3. Local (Vaginal and Uterine) Microbiota and RPL
Evidence suggests that vaginal and uterine microbiota may play a role in RPL [21,22,23,24,25,26,27]. The microbial population that lives in the vagina is composed of facultative and obligate anaerobes that form a symbiotic relationship with the host [15,22,23,24,25,26,27,28,29,30,31,32,33,34]. These bacteria maintain healthy vaginal tissue by promoting and supporting an ideal pH of ~4, producing hydrogen peroxide [31,32] and a proteinaceous outer protective layer [6,33,34]. Different species of Lactobacillus appear to be most prevalent among women [35,36], with L. crispatus being categorized as the most protective species and found in the highest proportions in healthy individuals who had successful pregnancies, and L. nervosa being the least protective. It is unclear whether this confers a negative effect [37,38]. Recurrent implantation failure (RIF) and RPL are associated with increased microbiome diversity and a loss of Lactobacillus dominance in the lower female reproductive system [25,29]. First-trimester miscarriage has been associated with reduced prevalence of Lactobacillus spp [39] and Lactobacillus spp. depleted vaginal microbiota was related to pro-inflammatory cytokine (IL-1β, IL-6, IL-8) levels most strongly in euploid miscarriage compared to viable term pregnancy [40]. On the other hand, Lactobacillus crispatus was less abundant in the endometrial samples of women with RPL compared with controls, and Gardnerella vaginalis was more abundant in the RPL group than in controls in both endometrial and vaginal samples [41]. Furthermore, Vomstein et al. [28] observed a lower abundance of Lactobacillaceae in the uterus of RPL and RIF patients at three points of their menstrual cycle. They found an increase in Proteobacteria in the RSA and RIF groups towards the end of the menstrual cycle. In this study, the RIF group exhibited a remarkably diverse composition, unlike the control and the RPL group [42]. In the same way, a relative dominance rate of Ureaplasma species in the endometrial microbiome was an independent risk factor for subsequent miscarriage with normal karyotype in a cohort of patients with a history of RPL [24]. Also, Proteobacteria and Firmicutes were significantly elevated in the endometrium of RPL patients in comparison with women requesting termination of normal pregnancy [43]. Further, the abundance of Bacteroides and Helicobacter in the vagina in an early embryonic arrest group was higher than that in the standard pregnancy-induced abortion group. Furthermore, the abundance of lactobacilli in the normal pregnancy-induced abortion group was higher than that in the embryonic arrest group. In this last group, the abundance of L. iners was significantly lower than that in the normal pregnancy group [44]. Therefore, women with an unbalanced population of bacteria, mainly composed of Gardnerella, Prevotella, Atopobium, Sneathia, Megasphaera, Delftia, Cutibacterium, Peptoniphilus, Anaerobacillus, and unknown bacteria, are at higher risk for premature birth and RPL [30,41,45,46,47]. In summary, Smith and Ravel [48] proposed a hierarchical clustering of the vaginal microbiota of reproductive-aged women into five distinct community state types (CST), four of which are dominated by Lactobacillus spp. (Lactobacillus crispatus (CST-I), L. iners (CST-III), L. gasseri (CST-II) or L. jensenii (CST-V)) and the fifth (CST-IV) is composed of a polymicrobial mixture of strict and facultative anaerobes, including species of the genera Atopobium, Megasphera, Mobiluncus, Prevotella, and other taxa in the order Clostridiales. CST I correlates with low obstetric-gynecological risk, and CST IV correlates mostly with vaginal discomfort and/or obstetric-gynecological diseases [49].
The microbiota is sensitive to environmental, hormonal, and dietary changes [48,49,50,51,52]. Microbiota plasticity, the ability to change the composition better to suit the host’s and bacteria’s needs, is implemented during the menstrual cycle [48,49,50]. In the genital tract of women of reproductive age, Prevotella spp. is linked to the proliferative phase of the menstrual cycle, whereas Sneathia spp. is related to the secretory phase [53]. On the other hand, Lactobacillus species are found in the highest numbers during the luteal phase and decrease during menstruation [50,54]. Interestingly, a prevalence of Prevotella spp. over Lactobacillus spp. and poor microbiota plasticity (little to no changes in bacterial population in response to different signals) are associated with an increased risk of infections and RPL [42].
Human leukocyte antigen genes (HLA) play a role in selecting the population of intestinal bacteria [55,56]; certain HLA haplotype carriers might be more susceptible to having a dysbiotic microbial population; in fact, women with the HLA-DQ2/DQ8 haplotype appear to have an altered microbiota [57].
A dysbiotic microbiota is less protective against sexually and non-sexually transmitted diseases such as HIV [58], syphilis, chlamydia, gonorrhea, Trichomonas [59,60], human papillomavirus (HPV) [61], herpes simplex virus (HSV) [62,63], pelvic inflammatory disease [64], aerobic vaginitis (AV) [65], bacterial vaginosis (BV) [20] and candidiasis [66]; all of which can negatively affect gestation [67,68,69,70,71] by causing inflammation and tissue destruction [72,73].
The composition of uterine endometrium microbiota (UEM) might predict pregnancy outcomes [24]. A dysbiotic UEM, consisting of Lactobacillus iners and Ureaplasma species, is associated with inflammatory conditions like chronic endometritis (CE) [26], and women with this condition are likely to be diagnosed with RPL, RIF, and infertility [74]. CE occurs when plasma cells and B lymphocytes migrate to the uterine endometrial stroma, creating permanent inflammation [75,76]. The administration of antibiotics has been shown to improve implantation outcomes [77] but has not improved miscarriage rates [78]. These results suggest that other bacteria may be essential to induce the tolerogenic response of the immune cells required for a normal pregnancy.
The gut microbiome might directly influence the vaginal and uterine microbiomes. Recently, Zhu et al. [31] provided evidence suggesting a connection between a dysbiotic gut microbiome and RPL. The gut microbiome, composed of symbiotic bacteria, is involved in metabolism, inflammation, and immunity [29]. To protect the host from infections, bacteria release factors and form a physical barrier by attaching to the intestinal wall [79]. They convert food particles into essential nutrients [80] and neurotransmitters and help modulate insulin response. The association between a dysbiotic gut microbiome and RPL is complex. Gut microbiota participates in numerous pathologies associated with higher incidences of RLP, such as chronic inflammation, vulnerability to infections, obesity, diabetes, and polycystic ovary syndrome (PCOS) [29]. The type of bacterial population in the gut microbiota is involved in local inflammatory responses due to abnormal production of cytokines [81].
A decrease in gut bacterial population diversity may result in the infiltration of gram-harmful bacteria and, consequently, the production of lipopolysaccharides (LPS). This endotoxin can enter the systemic circulation and promote the expression of pro-inflammatory genes [81,82]. LPS is also known to increase the production of reactive oxygen species (ROS), resulting in oxidative stress as well as insulin resistance (IR) [83,84]. Moreover, an abundance of non-protective bacterial species such as Prevotella [85] promotes inflammation that leads to immune cell migration and activation, which might induce the formation of autoantibodies [81] and excessive NK cell migration [86,87]. Autoimmune diseases such as Systemic Lupus Erythematosus (SLE), autoimmune thyroid disease (AITD), and celiac disease (CD) are all associated with higher incidences of RPL and a dysbiotic gut [57,86,87,88]. It is yet unclear whether treatment with the “beneficial” Lactobacillus species (L. crispatus) can improve autoimmunity and thus reduce the likelihood of fetal rejection [89]. A report has shown a direct association between regulated production and stimulation of NK cells and downregulation of pro-inflammatory cells, even in the placenta [90]. Interestingly, a higher prevalence of Gardnerella vaginalis and gram-negative bacteria has been associated with an increased number of circulating NK cells and recurrent miscarriage [91].
Changes in local microbiota not only affect RPL patients. In preeclampsia, an increase in Escherichia species was reported along with Rothia, Actinomyces, and Enterococcus, and a lower abundance of Coprococcus compared to pregnant women with normotension [92]. Changes may also occur in eclampsia and diabetic pregnancies. There are only a few reports relating RPL with metabolic syndrome after 40 years of age [93]. This association can be due to different events but may involve the gut microbiota. More research is required in this area.
4. Perspectives of Microbiota Modulation on RPL
The main problem in understanding the changes in local microbiota is the analysis of cultured samples. The molecular diagnosis assessment should aid in defining quantitative differences in the species encountered. In general, the changes in specific species of Lactobacillus and the increase in L. crispatus in women with successful pregnancy means that local microbiota changes may be necessary but not essential [94]. It is also difficult to ascertain the role of sperm microbiota in the process, the possible consequences of hormones like progesterone, and the relationship between different microbiotas, vaginal and endometrial, based on current knowledge.
Recent evidence indicates that diverse populations with a high proportion of Lactobacillus crispatus, are positively associated with lower incidences of infections, implantation failures, and RPL [15,23,95,96]. Efforts to modulate and improve the bacterial population in the vagina through the administration of antibiotics, boric acid, lactic acid, and estrogen have been unsuccessful long-term [97,98,99,100]. Treatments with vaginal and gut probiotics yielded mixed results [101,102,103,104]. Sex hormones may alter vaginal colonization, as shown in the mouse model [105], and using a low dose of estrogen in women with atrophic vaginitis [106]. Metronidazole treatment may also affect the process [107]. Vaginal microbiota transplants are a relatively new yet promising form of therapy and consist of transferring the entire vaginal microbiota of healthy women to patients [110,111,112]. Oral probiotic supplementation seems to aid vaginal microbiota {113}. A proof-of-concept case study where a woman successfully shifted her microbiota population after microbiota transplant from mostly Gardnerella spp. (90%) to 81.2% Lactobacillus crispatus and 9% Lactobacillus jensenii ameliorated her vaginal symptoms (vaginal irritation and discharge), and she was able to carry a healthy pregnancy to term after three pregnancy losses [114]. However, this topic and the therapies available are still relatively new, and there is room for improvement in the future.
Liu et al. [41] reported a critical drop in IFN-γ and IL-6 levels inside the uterine cavity of RPL women, suggesting a negative association between microbiota and immune response. Interestingly, this group [41] identified that imidazole propionic acid and 1,4-methyl imidazole acetic acid are associated with subsequent recurrent miscarriage. On the other hand, a reduction in butyrate-producing bacteria in the gut microbiota of RPL has been demonstrated, suggesting a link between immune vigilant responses and anti-inflammatory properties of the microbiota [41]. A report studied 20 fecal samples of patients with RPL, and antibody-related factors explained the conditions compared to 20 patients without antibodies [114]. Community richness and phylogenetic diversity in the antibody-positive group were higher than in the opposing group. Bacteroides were prevalent in the positive group, while in the opposing group, Bacteroides, Erysipelotrichaceae, Faecalibacterium, Enterococcus, Prevotella, Megasphaera, and Anaerostipes were abundant. Proteomic studies support the results and propose that cytoskeleton proteins may be crucial in the local disarrangements in endometrial tissue, which can lead to a lack of implantation [115]. In summary, well-defined clinical trials are required to identify the importance of gut microbiota and its possible therapeutic role in RPL.
Also, the association between gut bacterial dysbiosis and a Th1/Th17-mediated proinflammatory state in RPL has been demonstrated [116], uterine NK cells and macrophages [13]. Tolerogenic NK cells, dendritic cells, and macrophages [13], as well as a decreased amount of Treg [117}, seem responsible for the process.
Supplementary probiotics aid vaginal and endometrial microbiota [118,119]. Moreover, supplementary probiotic treatment benefits couples with RPL because of their capacity to improve aberrant spermatozoa antigenicity [104,120]. Tersigni and coworkers [121] reported that patients with celiac disease may benefit from oral probiotics by decreasing intestinal inflammation and increasing anti-inflammatory metabolites. There are still controversies in the field [122]. The number of well-designed clinical trials must be increased to ascertain the benefits of oral microbiota supplementation.
Omega-3 supplementation has been shown to increase fertility in mice [123] and is now being successfully used to enhance human fertility [124] and possibly reduce RLP [125]. However, in their review, Kello and Cho [126] mentioned that supplementation may only benefit patients with antiphospholipid syndrome. Consequently, it may assist a group of patients with RPL in which immune disorders may be involved. How oral supplementation of omega-3 may affect local microbiota is unknown; however, it can be postulated that a decrease in prostaglandin E2 production, as a product of an inflammatory response, may promote/support the production of other anti-inflammatory intermediates. The role of nutritional supplementation on RPL should be carefully studied [4,6,120,124].
Recently, it has been shown that gut microbiota metabolites affect circulating lymphocytes. There is an increased amount of Th1, Th17, and B cells, with a decrease in Treg and Breg cells [127]. Even though the report is only based on 12 samples, it raises an important question concerning oral supplementation. It should probably be assumed that RPL patients have a subclinical inflammatory condition that may be modulated pharmacologically.
It has been assumed that local prednisone treatment reduces local inflammatory response and consequently decreases abortion risk and increases pregnancy efficiency in IVF [128,129] and also the use of nutritional supplements [130]. Also, topic cytokines such as G-CSF [132] and interferon λ [133] can increase the possibility of normal pregnancy primarily based on the induction of tolerance at the tissue level. However, we know that critical scientific evidence of therapeutic local progesterone and cytokines local microbiota has not been documented.
5. Conclusions
Primary and secondary RPL are complex medical conditions involving 1-2 % of women of reproductive age. Despite numerous efforts to study the possible triggering factors, significant achievements have yet to be reported. Local (vaginal and endometrial) and gut microbiota and metabolites differ in RPL compared to normal pregnancies. There is evidence of cell migration and local tissue disarrangements in RPL. Protective microbiota prevents local inflammatory response and infections and protects endometrial tissue. Transplantation of microbiota and oral probiotics may help maintain vaginal microbiota.
Patients with RPL, especially those with immune disorders, may benefit from oral probiotic supplementation or microbiota transplantation. It is not clear whether antigen mimicry can play a role in RPL.
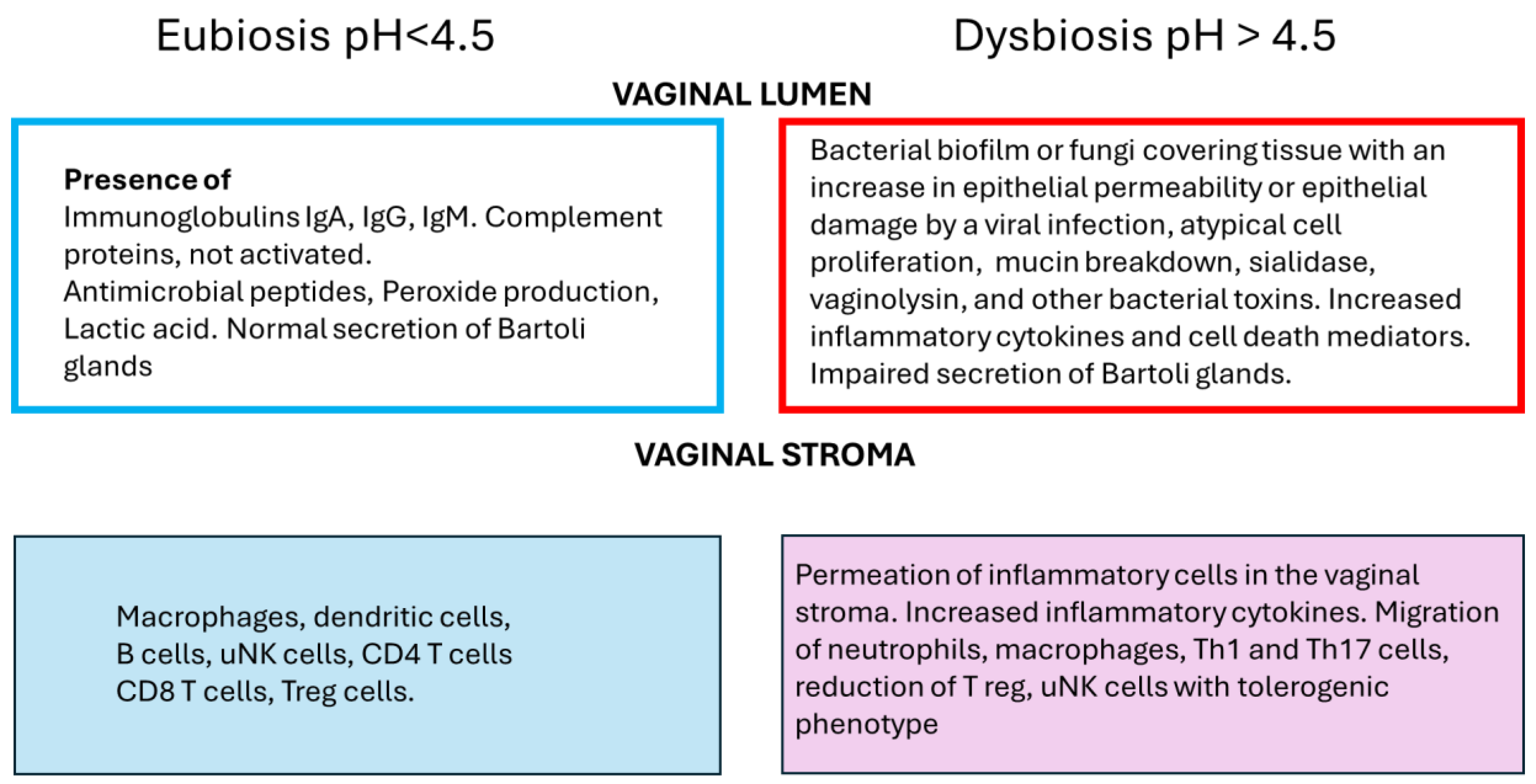
Figure 1 represents a summary of the events that occur in eubiosis, normal conditions, and dysbiosis in the vagina.
Well-planned clinical trials are required to provide evidence of the role of microbiota and microbiota transplantation in RPL. RPL patients may be at risk of developing medical conditions shortly after reproductive age; therefore, studies on RPL classification and triggers are urgently required.
Author Contributions
Conceptualization, CVDS, and JVG.; investigation, CVDS, JBDS, JVG; resources, MH.; writing—original draft preparation, CVDS, JVG.; writing—review and editing, CVDS, JVG, JBDS; project administration, MH.; funding acquisition, MH. All authors have read and agreed to the published version of the manuscript.
Funding
National Institute of Virology and Bacteriology [Program EXCELES, ID Project No. LX22NPO5103] - - Funded by the European Union - Next Generation EU from the Ministry of Education, Youth and Sports of the Czech Republic (MEYS)].
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Daya, S.; Stephenson, M.D. Frequency of factors associated with habitual abortion in 197 couples. Fertil. Steril. 1996, 66, 24–29. [CrossRef]
- Ford, H.B.; Schust, D.J. Recurrent Pregnancy Loss: Etiology, Diagnosis, and Therapy. Rev. Obstet. Gynecol. 2009, 2, 76–83.
- Pillarisetty, L.S.; Mahdy, H. Recurrent Pregnancy Loss. [Updated 2023 Aug 28]. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2024 Jan-. Available from: https://www.ncbi.nlm.nih.gov/books/NBK554460/ assessed June 4, 2024.
- Gao, H.; Liu, Q.; Wang, X.; Li, T.; Li, H.; Li, G.; Tan, L.; Chen, Y. Deciphering the role of female reproductive tract microbiome in reproductive health: a review. Front. Cell. Infect. Microbiol. 2024, 14, 1351540. [CrossRef]
- Agostinis, C.; Mangogna, A.; Bossi, F.; Ricci, G.; Kishore, U.; Bulla, R. Uterine Immunity and Microbiota: A Shifting Paradigm. Front. Immunol. 2019, 10, 2387. [CrossRef]
- Gao, X.; Louwers, Y.V.; Laven, J.S.E.; Schoenmakers, S. Clinical Relevance of Vaginal and Endometrial Microbiome Investigation in Women with Repeated Implantation Failure and Recurrent Pregnancy Loss. Int. J. Mol. Sci. 2024, 25, 622. [CrossRef]
- Lee, S.K.; Kim, C.J.; Kim, D.-J.; Kang, J.-H. Immune Cells in the Female Reproductive Tract. Immune Netw. 2015, 15, 16–26. [CrossRef]
- Dai, M.; Xu, Y.; Gong, G.; Zhang, Y. Roles of immune microenvironment in the female reproductive maintenance and regulation: novel insights into the crosstalk of immune cells. Front. Immunol. 2023, 14, 1109122. [CrossRef]
- Hedger, M.P. The Immunophysiology of Male Reproduction. Knobil and Neill’s Physiology of Reproduction. 2015:805–92. [CrossRef]
- Solders, M.; Gorchs, L.; Erkers, T.; Lundell, A.-C.; Nava, S.; Gidlöf, S.; Tiblad, E.; Magalhaes, I.; Kaipe, H. MAIT cells accumulate in placental intervillous space and display a highly cytotoxic phenotype upon bacterial stimulation. Sci. Rep. 2017, 7, 1–13. [CrossRef]
- Favaro, R.R.; Phillips, K.; Delaunay-Danguy, R.; Ujčič, K.; Markert, U.R. Emerging Concepts in Innate Lymphoid Cells, Memory, and Reproduction. Front. Immunol. 2022, 13, 824263. [CrossRef]
- Gibbs, A.; Leeansyah, E.; Introini, A.; Paquin-Proulx, D.; Hasselrot, K.; Andersson, E.; Broliden, K.; Sandberg, J.; Tjernlund, A. MAIT cells reside in the female genital mucosa and are biased towards IL-17 and IL-22 production in response to bacterial stimulation. Mucosal Immunol. 2017, 10, 35–45. [CrossRef]
- Garmendia, J.V.; De Sanctis, J.B. A Brief Analysis of Tissue-Resident NK Cells in Pregnancy and Endometrial Diseases: The Importance of Pharmacologic Modulation. Immuno 2021, 1, 174-193.
- Lund, J.M.; Hladik, F.; Prlic, M. Advances and challenges in studying the tissue-resident T cell compartment in the human female reproductive tract. Immunol. Rev. 2023, 316, 52–62. [CrossRef]
- Ali, S.; Majid, S.; Ali, N.; Taing, S.; Rehman, M.U.; Arafah, A. Cytokine imbalance at materno-embryonic interface as a potential immune mechanism for recurrent pregnancy loss. Int. Immunopharmacol. 2021, 90, 107118. [CrossRef]
- Prašnikar, E.; Kunej, T.; Gorenjak, M.; PotočNik, U.; Kovačič, B.; Knez, J. Transcriptomics of receptive endometrium in women with sonographic features of adenomyosis. Reprod. Biol. Endocrinol. 2022, 20, 1–16. [CrossRef]
- Jewanraj, J.; Ngcapu, S.; Osman, F.; Mtshali, A.; Singh, R.; Mansoor, L.E.; Karim, S.S.A.; Karim, Q.A.; Passmore, J.-A.S.; Liebenberg, L.J.P. The Impact of Semen Exposure on the Immune and Microbial Environments of the Female Genital Tract. Front. Reprod. Heal. 2020, 2, 566559. [CrossRef]
- Balci, C.N.; Acar, N. NLRP3 inflammasome pathway, the hidden balance in pregnancy: A comprehensive review. J. Reprod. Immunol. 2024, 161, 104173. [CrossRef]
- Koga, K.; Izumi, G.; Mor, G.; Fujii, T.; Osuga, Y. Toll-like Receptors at the Maternal-Fetal Interface in Normal Pregnancy and Pregnancy Complications. Am. J. Reprod. Immunol. 2014, 72, 192–205. [CrossRef]
- Benjelloun, F.; Quillay, H.; Cannou, C.; Marlin, R.; Madec, Y.; Fernandez, H.; Chrétien, F.; Le Grand, R.; Barré-Sinoussi, F.; Nugeyre, M.-T.; et al. Activation of Toll-Like Receptors Differentially Modulates Inflammation in the Human Reproductive Tract: Preliminary Findings. Front. Immunol. 2020, 11, 1655. [CrossRef]
- Demirezen, .; Isik, G.; Dönmez, H.; Beksaç, M. Bacterial vaginosis in association with spontaneous abortion and recurrent pregnancy losses. J. Cytol. 2016, 33, 135–140. [CrossRef]
- Benner, M.; Ferwerda, G.; Joosten, I.; van der Molen, R.G. How uterine microbiota might be responsible for a receptive, fertile endometrium. Hum. Reprod. Updat. 2018, 24, 393–415. [CrossRef]
- Fan, T.; Zhong, X.-M.; Wei, X.-C.; Miao, Z.-L.; Luo, S.-Y.; Cheng, H.; Xiao, Q. The alteration and potential relationship of vaginal microbiota and chemokines for unexplained recurrent spontaneous abortion. Medicine 2020, 99, e23558. [CrossRef]
- Liu, F.-T.; Yang, S.; Yang, Z.; Zhou, P.; Peng, T.; Yin, J.; Ye, Z.; Shan, H.; Yu, Y.; Li, R. An Altered Microbiota in the Lower and Upper Female Reproductive Tract of Women with Recurrent Spontaneous Abortion. Microbiol. Spectr. 2022, 10, e0046222. [CrossRef]
- Moreno, I.; Garcia-Grau, I.; Perez-Villaroya, D.; Gonzalez-Monfort, M.; Bahçeci, M.; Barrionuevo, M.J., et al. Endometrial microbiota composition is associated with reproductive outcome in infertile patients. Microbiome. 2022 Jan 4;10(1):1.
- Peuranpää, P.; Holster, T.; Saqib, S.; Kalliala, I.; Tiitinen, A.; Salonen, A.; Hautamäki, H. Female reproductive tract microbiota and recurrent pregnancy loss: a nested case- control study. Reprod. Biomed. Online 2022, 45, 1021–1031. [CrossRef]
- Shi, Y.; Yamada, H.; Sasagawa, Y.; Tanimura, K.; Deguchi, M. Uterine endometrium microbiota and pregnancy outcome in women with recurrent pregnancy loss. J. Reprod. Immunol. 2022, 152, 103653. [CrossRef]
- Vomstein, K.; Reider, S.; Böttcher, B.; Watschinger, C.; Kyvelidou, C.; Tilg, H.; Moschen, A.R.; Toth, B. Uterine microbiota plasticity during the menstrual cycle: Differences between healthy controls and patients with recurrent miscarriage or implantation failure. J. Reprod. Immunol. 2022, 151, 103634. [CrossRef]
- Caliskan, C.S.; Yurtcu, N.; Celik, S.; Sezer, O.; Kilic, S.S.; Cetin, A. Derangements of vaginal and cervical canal microbiota determined with real-time PCR in women with recurrent miscarriages. J. Obstet. Gynaecol. 2022, 42, 2105–2114. [CrossRef]
- Takimoto, K.; Yamada, H.; Shimada, S.; Fukushi, Y.; Wada, S. Chronic Endometritis and Uterine Endometrium Microbiota in Recurrent Implantation Failure and Recurrent Pregnancy Loss. Biomedicines 2023, 11, 2391. [CrossRef]
- Zhu, J.; Jin, J.; Qi, Q.; Li, L.; Zhou, J.; Cao, L.; Wang, L. The association of gut microbiome with recurrent pregnancy loss: A comprehensive review. Drug Discov. Ther. 2023, 17, 157–169. [CrossRef]
- Lebedeva, O.P.; Popov, V.N.; Syromyatnikov, M.Y.; Starkova, N.N.; Maslov, A.Y.; Kozarenko, O.N.; Gryaznova, M.V. Female reproductive tract microbiome and early miscarriages. APMIS 2023, 131, 61–76. [CrossRef]
- Mori, R.; Hayakawa, T.; Hirayama, M.; Ozawa, F.; Yoshihara, H.; Goto, S.; Kitaori, T.; Ozaki, Y.; Sugiura-Ogasawara, M. Cervicovaginal microbiome in patients with recurrent pregnancy loss. J. Reprod. Immunol. 2023, 157, 103944. [CrossRef]
- Odendaal, J.; Black, N.; Bennett, P.R.; Brosens, J.; Quenby, S.; A MacIntyre, D. The endometrial microbiota and early pregnancy loss. Hum. Reprod. 2024, 39, 638–646. [CrossRef]
- Klebanoff, S.J.; Hillier, S.L.; Eschenbach, D.A.; Waltersdorph, A.M. Control of the microbial flora of the vagina by H2O2-generating lactobacilli. J Infect Dis. 1991 Jul;164(1):94-100.
- Giannella, L.; Grelloni, C.; Quintili, D.; Fiorelli, A.; Montironi, R.; Alia, S.; Carpini, G.D.; Di Giuseppe, J.; Vignini, A.; Ciavattini, A. Microbiome Changes in Pregnancy Disorders. Antioxidants 2023, 12, 463. [CrossRef]
- Miko, E.; Barakonyi, A. The Role of Hydrogen-Peroxide (H2O2) Produced by Vaginal Microbiota in Female Reproductive Health. Antioxidants (Basel). 2023 May 6;12(5):1055.
- Sillanpää, J.; Martínez, B.; Antikainen, J.; Toba, T.; Kalkkinen, N.; Tankka, S.; Lounatmaa, K.; Keränen, J.; Höök, M.; Westerlund-Wikström, B.; et al. Characterization of the Collagen-Binding S-Layer Protein CbsA of Lactobacillus crispatus. J. Bacteriol. 2000, 182, 6440–6450. [CrossRef]
- Al-Memar, M.; Bobdiwala, S.; Fourie, H.; Mannino, R.; Lee, Y.; Smith, A.; Marchesi, J.; Timmerman, D.; Bourne, T.; Bennett, P.; et al. The association between vaginal bacterial composition and miscarriage: a nested case–control study. BJOG: Int. J. Obstet. Gynaecol. 2020, 127, 264–274. [CrossRef]
- Grewal, K.; Lee, Y.S.; Smith, A.; Brosens, J.J.; Bourne, T.; Al-Memar, M.; Kundu, S.; MacIntyre, D.A.; Bennett, P.R. Chromosomally normal miscarriage is associated with vaginal dysbiosis and local inflammation. BMC Med. 2022, 20, 1–15. [CrossRef]
- Liu, Y.; Chen, H.; Feng, L.; Zhang, J. Interactions between gut microbiota and metabolites modulate cytokine network imbalances in women with unexplained miscarriage. npj Biofilms Microbiomes 2021, 7, 1–12. [CrossRef]
- Ncib, K.; Bahia, W.; Leban, N.; Mahdhi, A.; Trifa, F.; Mzoughi, R.; Haddad, A.; Jabeur, C.; Donders, G. Microbial Diversity and Pathogenic Properties of Microbiota Associated with Aerobic Vaginitis in Women with Recurrent Pregnancy Loss. Diagnostics 2022, 12, 2444. [CrossRef]
- Shu, J.; Lin, S.; Wu, Y.; Zhu, J.; Gong, D.; Zou, X.; Zhu, H.; Gao, J. A potential role for the uterine microbiome in missed abortions. J Biol Regul Homeost Agents, 2022; 36, 1055–1063.
- Wang, L.; Chen, J.; He, L.; Liu, H.; Liu, Y.; Luan, Z.; Li, H.; Liu, W.; Luo, M. Association between the vaginal and uterine microbiota and the risk of early embryonic arrest. Front. Microbiol. 2023, 14. [CrossRef]
- Palomino, M.M.; Allievi, M.C.; Gordillo, T.B.; Bockor, S.S.; Martin, J.F.; Ruzal, S.M. Surface layer proteins in species of the family Lactobacillaceae. Microb. Biotechnol. 2023, 16, 1232–1249. [CrossRef]
- France, M.; Alizadeh, M.; Brown, S.; Ma, B.; Ravel, J. Towards a deeper understanding of the vaginal microbiota. Nat. Microbiol. 2022, 7, 367–378. [CrossRef]
- Mendes-Soares, H.; Suzuki, H.; Hickey, R.J.; Forney, L.J. Comparative Functional Genomics of Lactobacillus spp. Reveals Possible Mechanisms for Specialization of Vaginal Lactobacilli to Their Environment. J. Bacteriol. 2014, 196, 1458–1470. [CrossRef]
- Smith, S.B.; Ravel, J. The vaginal microbiota, host defence and reproductive physiology. J. Physiol. 2017, 595, 451–463. [CrossRef]
- Nelson, D.B.; Hanlon, A.; Nachamkin, I.; Haggerty, C.; Mastrogiannis, D.S.; Liu, C.; Fredricks, D.N. Early Pregnancy Changes in Bacterial Vaginosis-Associated Bacteria and Preterm Delivery. Paediatr. Périnat. Epidemiology 2014, 28, 88–96. [CrossRef]
- Brown, R.G.; Al-Memar, M.; Marchesi, J.R.; Lee, Y.S.; Smith, A.; Chan, D.; Lewis, H.; Kindinger, L.; Terzidou, V.; Bourne, T.; et al. Establishment of vaginal microbiota composition in early pregnancy and its association with subsequent preterm prelabor rupture of the fetal membranes. Transl. Res. 2019, 207, 30–43. [CrossRef]
- Elovitz, M.A.; Gajer, P.; Riis, V.; Brown, A.G.; Humphrys, M.S.; Holm, J.B.; Ravel, J. Cervicovaginal microbiota and local immune response modulate the risk of spontaneous preterm delivery. Nat. Commun. 2019, 10, 1–8. [CrossRef]
- Feehily, C.; Crosby, D.; Walsh, C.J.; Lawton, E.M.; Higgins, S.; McAuliffe, F.M.; Cotter, P.D. Shotgun sequencing of the vaginal microbiome reveals both a species and functional potential signature of preterm birth. npj Biofilms Microbiomes 2020, 6, 1–9. [CrossRef]
- Song, S.D.; Acharya, K.D.; Zhu, J.E.; Deveney, C.M.; Walther-Antonio, M.R.S.; Tetel, M.J.; Chia, N. Daily Vaginal Microbiota Fluctuations Associated with Natural Hormonal Cycle, Contraceptives, Diet, and Exercise. mSphere 2020, 5. [CrossRef]
- Toson, B.; Simon, C.; Moreno, I. The Endometrial Microbiome and Its Impact on Human Conception. Int. J. Mol. Sci. 2022, 23, 485. [CrossRef]
- Gholiof, M.; Luca, E.A.-D.; Wessels, J.M. The female reproductive tract microbiotas, inflammation, and gynecological conditions. Front. Reprod. Heal. 2022, 4, 963752. [CrossRef]
- Conlon, M.A.; Bird, A.R. The Impact of Diet and Lifestyle on Gut Microbiota and Human Health. Nutrients 2014, 7, 17–44. [CrossRef]
- Pelzer, E.S.; Willner, D.; Buttini, M.; Huygens, F. A role for the endometrial microbiome in dysfunctional menstrual bleeding. Antonie van Leeuwenhoek 2018, 111, 933–943. [CrossRef]
- Kadogami, D.; Nakaoka, Y.; Morimoto, Y. Use of a vaginal probiotic suppository and antibiotics to influence the composition of the endometrial microbiota. Reprod. Biol. 2020, 20, 307–314. [CrossRef]
- De Palma, G.; Capilla, A.; Nadal, I.; Nova, E.; Pozo, T.; Varea, V.; Polanco, I.; Castillejo, G.; López, A.; Garrote, J.; et al. Interplay Between Human Leukocyte Antigen Genes and the Microbial Colonization Process of the Newborn Intestine. Curr. Issues Mol. Biol. 2010, 12, 1–10. [CrossRef]
- Andeweg, S.P.; Keşmir, C.; Dutilh, B.E. Quantifying the Impact of Human Leukocyte Antigen on the Human Gut Microbiota. mSphere 2021, 6, e0047621. [CrossRef]
- Masucci, L.; D’ippolito, S.; De Maio, F.; Quaranta, G.; Mazzarella, R.; Bianco, D.M.; Castellani, R.; Inversetti, A.; Sanguinetti, M.; Gasbarrini, A.; et al. Celiac Disease Predisposition and Genital Tract Microbiota in Women Affected by Recurrent Pregnancy Loss. Nutrients 2023, 15, 221. [CrossRef]
- McClelland, R.S.; Lingappa, J.R.; Srinivasan, S.; Kinuthia, J.; John-Stewart, G.C.; Jaoko, W.; Richardson, B.A.; Yuhas, K.; Fiedler, T.L.; Mandaliya, K.N.; et al. Evaluation of the association between the concentrations of key vaginal bacteria and the increased risk of HIV acquisition in African women from five cohorts: a nested case-control study. Lancet Infect. Dis. 2018, 18, 554–564. [CrossRef]
- Brotman, R.M.; Klebanoff, M.A.; Nansel, T.R.; Yu, K.F.; Andrews, W.W.; Zhang, J.; Schwebke, J.R. Bacterial Vaginosis Assessed by Gram Stain and Diminished Colonization Resistance to Incident Gonococcal, Chlamydial, and Trichomonal Genital Infection. J. Infect. Dis. 2010, 202, 1907–1915. [CrossRef]
- Van Gerwen, O.T.; Muzny, C.A.; Marrazzo, J.M. Sexually transmitted infections and female reproductive health. Nat. Microbiol. 2022, 7, 1116–1126. [CrossRef]
- Zhao, F.; Chen, Y.; Gao, J.; Wu, M.; Li, C.; Wang, Z.; Huang, N.; Cui, L.; Du, M.; Ying, C. Characterization of Vaginal Microbiota in Women With Recurrent Spontaneous Abortion That Can Be Modified by Drug Treatment. Front. Cell. Infect. Microbiol. 2021, 11. [CrossRef]
- Brusselaers, N.; Shrestha, S.; van de Wijgert, J.; Verstraelen, H. Vaginal dysbiosis and the risk of human papillomavirus and cervical cancer: systematic review and meta-analysis. Am. J. Obstet. Gynecol. 2019, 221, 9–18.e8. [CrossRef]
- Cherpes, T.L.; Meyn, L.A.; Krohn, M.A.; Lurie, J.G.; Hillier, S.L. Association between Acquisition of Herpes Simplex Virus Type 2 in Women and Bacterial Vaginosis. Clin. Infect. Dis. 2003, 37, 319–325. [CrossRef]
- Shannon, B.; Gajer, P.; Yi, T.J.; Ma, B.; Humphrys, M.S.; Thomas-Pavanel, J.; Chieza, L.; Janakiram, P.; Saunders, M.; Tharao, W.; et al. Distinct Effects of the Cervicovaginal Microbiota and Herpes Simplex Type 2 Infection on Female Genital Tract Immunology. J. Infect. Dis. 2017, 215, 1366–1375. [CrossRef]
- Haggerty, C.L.; Ness, R.B.; Totten, P.A.; Farooq, F.; Tang, G.; Ko, D.B.; Hou, X.; Fiedler, T.L.B.; Srinivasan, S.; Astete, S.G.; et al. Presence and Concentrations of Select Bacterial Vaginosis-Associated Bacteria Are Associated With Increased Risk of Pelvic Inflammatory Disease. Sex. Transm. Dis. 2020, 47, 344–346. [CrossRef]
- Brown, S.E.B.; Schwartz, J.A.; Robinson, C.K.; O'Hanlon, D.E.; Bradford, L.L.; He, X.; Mark, K.S.; Bruno, V.M.; Ravel, J.; Brotman, R.M. The Vaginal Microbiota and Behavioral Factors Associated With Genital Candida albicans Detection in Reproductive-Age Women. Sex. Transm. Dis. 2019, 46, 753–758. [CrossRef]
- Chilaka, V.N.; Navti, O.B.; Al Beloushi, M.; Ahmed, B.; Konje, J.C. Human papillomavirus (HPV) in pregnancy – An update. Eur. J. Obstet. Gynecol. Reprod. Biol. 2021, 264, 340–348. [CrossRef]
- Tang, W.; Mao, J.; Li, K.T.; Walker, J.S.; Chou, R.; Fu, R.; Chen, W.; Darville, T.; Klausner, J.; Tucker, J.D. Pregnancy and fertility-related adverse outcomes associated with Chlamydia trachomatis infection: a global systematic review and meta-analysis. Sex Transm Infect. 2020 Aug;96(5):322-329.
- Tsai2, S.; Sun, M.Y.; Kuller, J.A.; Rhee, E.H.J.; Dotters-Katz, S. Syphilis in Pregnancy. Obstet Gynecol Surv. 2019 Sep;74(9):557-564.
- Zheng, N.; Guo, R.; Wang, J.; Zhou, W.; Ling, Z. Contribution of Lactobacillus iners to Vaginal Health and Diseases: A Systematic Review. Front. Cell. Infect. Microbiol. 2021, 11. [CrossRef]
- Lindsay, C.V.; A Potter, J.; A Grimshaw, A.; Abrahams, V.M.; Tong, M. Endometrial responses to bacterial and viral infection: a scoping review. Hum. Reprod. Updat. 2023, 29, 675–693. [CrossRef]
- Lennard, K.; Dabee, S.; Barnabas, S.L.; Havyarimana, E.; Blakney, A.; Jaumdally, S.Z.; Botha, G.; Mkhize, N.N.; Bekker, L.-G.; Lewis, D.A.; et al. Microbial Composition Predicts Genital Tract Inflammation and Persistent Bacterial Vaginosis in South African Adolescent Females. Infect. Immun. 2017, 86, e00410-17. [CrossRef]
- Zuend, C.F.; Lamont, A.; Noel-Romas, L.; Knodel, S.; Birse, K.; Kratzer, K.; McQueen, P.; Perner, M.; Ayele, H.; Mutch, S.; et al. Increased genital mucosal cytokines in Canadian women associate with higher antigen-presenting cells, inflammatory metabolites, epithelial barrier disruption, and the depletion of L. crispatus. Microbiome 2023, 11, 1–16. [CrossRef]
- Kimura, F.; Takebayashi, A.; Ishida, M.; Nakamura, A.; Kitazawa, J.; Morimune, A.; Hirata, K.; Takahashi, A.; Tsuji, S.; Takashima, A.; et al. Review: Chronic endometritis and its effect on reproduction. J. Obstet. Gynaecol. Res. 2019, 45, 951–960. [CrossRef]
- Song, D.; He, Y.; Wang, Y.; Liu, Z.; Xia, E.; Huang, X.; Xiao, Y.; Li, T.-C. Impact of antibiotic therapy on the rate of negative test results for chronic endometritis: a prospective randomized control trial. Fertil. Steril. 2021, 115, 1549–1556. [CrossRef]
- Freitas, A.C.; the VOGUE Research Group; Bocking, A.; Hill, J.E.; Money, D.M. Increased richness and diversity of the vaginal microbiota and spontaneous preterm birth. Microbiome 2018, 6, 1–15. [CrossRef]
- Fettweis, J.M.; Serrano, M.G.; Edwards, D.J.; Girerd, P.H.; Parikh, H.I.; Huang, B.; Arodz, T.J.; Edupuganti, L.; Glascock, A.L.; Xu, J.; et al. The vaginal microbiome and preterm birth. Nat. Med. 2019, 25, 1012–1021. [CrossRef]
- Peelen, M.J.; Luef, B.M.; Lamont, R.F.; de Milliano, I.; Jensen, J.S.; Limpens, J.; Hajenius, P.J.; Jørgensen, J.S.; Menon, R. The influence of the vaginal microbiota on preterm birth: A systematic review and recommendations for a minimum dataset for future research. Placenta 2019, 79, 30–39. [CrossRef]
- Ghosh, S.S.; Wang, J.; Yannie, P.J.; Ghosh, S. Intestinal Barrier Dysfunction, LPS Translocation, and Disease Development. J Endocr Soc. 2020 Feb 20;4(2):bvz039.
- Den Besten, G.; van Eunen, K.; Groen, A.K.; Venema, K.; Reijngoud, D.-J.; Bakker, B.M. The role of short-chain fatty acids in the interplay between diet, gut microbiota, and host energy metabolism. J. Lipid Res. 2013, 54, 2325–2340. [CrossRef]
- Charoensappakit, A.; Sae-Khow, K.; Leelahavanichkul, A. Gut Barrier Damage and Gut Translocation of Pathogen Molecules in Lupus, an Impact of Innate Immunity (Macrophages and Neutrophils) in Autoimmune Disease. Int. J. Mol. Sci. 2022, 23, 8223. [CrossRef]
- Tersigni, C.; D’Ippolito, S.; Di Nicuolo, F.; Marana, R.; Valenza, V.; Masciullo, V.; Scaldaferri, F.; Malatacca, F.; et al. Recurrent pregnancy loss is associated to leaky gut: a novel pathogenic model of endometrium inflammation? J Transl Med. 2018 Apr 17;16(1):102.
- Huang, L.; Thonusin, C.; Chattipakorn, N.; Chattipakorn, S.C. Impacts of gut microbiota on gestational diabetes mellitus: a comprehensive review. Eur. J. Nutr. 2021, 60, 2343–2360. [CrossRef]
- Belizário, J.E.; Faintuch, J.; Garay-Malpartida, M. Gut Microbiome Dysbiosis and Immunometabolism: New Frontiers for Treatment of Metabolic Diseases. Mediat. Inflamm. 2018, 2018, 1–12. [CrossRef]
- Larsen, J.M. The immune response toPrevotellabacteria in chronic inflammatory disease. Immunology 2017, 151, 363–374. [CrossRef]
- Russell, J.T.; Roesch, L.F.W.; Ördberg, M.; Ilonen, J.; Atkinson, M.A.; Schatz, D.A.; Triplett, E.W.; Ludvigsson, J. Genetic risk for autoimmunity is associated with distinct changes in the human gut microbiome. Nat. Commun. 2019, 10, 1–12. [CrossRef]
- Kalinkovich, A.; Livshits, G. A cross talk between dysbiosis and gut-associated immune system governs the development of inflammatory arthropathies. Semin. Arthritis Rheum. 2019, 49, 474–484. [CrossRef]
- Christiansen, O.B.; Steffensen, R.; Nielsen, H.S.; Varming, K. Multifactorial Etiology of Recurrent Miscarriage and Its Scientific and Clinical Implications. Gynecol. Obstet. Investig. 2008, 66, 257–267. [CrossRef]
- Esmaeili, S.; Mahmoudi, M.; Rezaieyazdi, Z.; Sahebari, M.; Tabasi, N.; Sahebkar, A.; Rastin, M. Generation of tolerogenic dendritic cells using Lactobacillus rhamnosus and Lactobacillus delbrueckii as tolerogenic probiotics. J. Cell. Biochem. 2018, 119, 7865–7872. [CrossRef]
- Kumar, M.; Murugesan, S.; Singh, P.; Saadaoui, M.; Elhag, D.A.; Terranegra, A.; Kabeer, B.S.A.; Marr, A.K.; Kino, T.; Brummaier, T.; et al. Vaginal Microbiota and Cytokine Levels Predict Preterm Delivery in Asian Women. Front. Cell. Infect. Microbiol. 2021, 11. [CrossRef]
- Rautava, S.; Collado, M.C.; Salminen, S.; Isolauri, E. Probiotics Modulate Host-Microbe Interaction in the Placenta and Fetal Gut: A Randomized, Double-Blind, Placebo-Controlled Trial. Neonatology 2012, 102, 178–184. [CrossRef]
- Kuon, R.; Togawa, R.; Vomstein, K.; Weber, M.; Goeggl, T.; Strowitzki, T.; Markert, U.; Zimmermann, S.; Daniel, V.; Dalpke, A.; et al. Higher prevalence of colonization with Gardnerella vaginalis and gram-negative anaerobes in patients with recurrent miscarriage and elevated peripheral natural killer cells. J. Reprod. Immunol. 2017, 120, 15–19. [CrossRef]
- Wu, Z.; Ge, M.; Liu, J.; Chen, X.; Cai, Z.; Huang, H. The gut microbiota composition and metabolites are different in women with hypertensive disorders of pregnancy and normotension: A pilot study. J. Obstet. Gynaecol. Res. 2023, 50, 334–341. [CrossRef]
- Bae, J.H.; Jung YM, Lee J, Shivakumar M, Park CW, Park JS, Jun JK, Kim D, Kim SK, Lee SM. Future risk of metabolic syndrome after recurrent pregnancy loss: a cohort study using UK Biobank. Fertil Steril. 2023 Dec;120(6):1227-1233.
- Reschini, M.; Benaglia, L.; Ceriotti, F.; Borroni, R.; Ferrari, S.; Castiglioni, M.; Guarneri, D.; Porcaro, L.; Vigano,’ P.; Somigliana, E.; Uceda Renteria, S. Endometrial microbiome: sampling, assessment, and possible impact on embryo implantation. Sci Rep. 2022 May 19;12(1):8467.
- Inversetti, A.; Zambella, E.; Guarano, A.; Dell’avanzo, M.; Di Simone, N. Endometrial Microbiota and Immune Tolerance in Pregnancy. Int. J. Mol. Sci. 2023, 24, 2995. [CrossRef]
- Reichman, O.; Akins, R.; Sobel, J.D. Boric Acid Addition to Suppressive Antimicrobial Therapy for Recurrent Bacterial Vaginosis. Sex. Transm. Dis. 2009, 36, 732–734. [CrossRef]
- Kitaya, K.; Matsubayashi, H.; Takaya, Y.; Nishiyama, R.; Yamaguchi, K.; Takeuchi, T.; Ishikawa, T. Live birth rate following oral antibiotic treatment for chronic endometritis in infertile women with repeated implantation failure. Am. J. Reprod. Immunol. 2017, 78. [CrossRef]
- Zeng, M.; Yang, L.; Mao, Y.; He, Y.; Li, M.; Liu, J.; Zhu, Q.; Chen, L.; Zhou, W. Preconception reproductive tract infections status and adverse pregnancy outcomes: a population-based retrospective cohort study. BMC Pregnancy Childbirth 2022, 22, 1–10. [CrossRef]
- Rafiee, M.; Sereshki, N.; Alipour, R.; Ahmadipanah, V.; Sarvar, D.P.; Wilkinson, D. The effect of probiotics on immunogenicity of spermatozoa in couples suffering from recurrent spontaneous abortion. BMC Immunol. 2022, 23, 1–8. [CrossRef]
- Rahman, N.; Mian, M.F.; Nazli, A.; Kaushic, C. Human vaginal microbiota colonization is regulated by female sex hormones in a mouse model. Front. Cell. Infect. Microbiol. 2023, 13, 1307451. [CrossRef]
- Shen, J.; Song, N.; Williams, C.J.; Brown, C.J.; Yan, Z.; Xu, C.; Forney, L.J. Effects of low dose estrogen therapy on the vaginal microbiomes of women with atrophic vaginitis. Sci. Rep. 2016, 6, 24380–24380. [CrossRef]
- Gustin, A.T.; Thurman, A.R.; Chandra, N.; Schifanella, L.; Alcaide, M.; Fichorova, R.; Doncel, G.F.; Gale, M. Jr; Klatt, N.R. Recurrent bacterial vaginosis following metronidazole treatment is associated with microbiota richness at diagnosis. Am J Obstet Gynecol. 2022 Feb;226(2):225.e1-225.e15.
- Plummer, E.L.; Bradshaw, C.S.; Doyle, M.; Fairley, C.K.; Murray, G.L.; Bateson, D.; Masson, L.; Slifirski, J.; Tachedjian, G.; Vodstrcil, L.A. Lactic acid-containing products for bacterial vaginosis and their impact on the vaginal microbiota: A systematic review. PLOS ONE 2021, 16, e0246953. [CrossRef]
- Oerlemans, E.F.M.; Bellen, G.; Claes, I.; Henkens, T.; Allonsius, C.N.; Wittouck, S.; Broek, M.F.L.v.D.; Wuyts, S.; Kiekens, F.; Donders, G.G.G.; et al. Impact of a lactobacilli-containing gel on vulvovaginal candidosis and the vaginal microbiome. Sci. Rep. 2020, 10, 1–10. [CrossRef]
- Tuniyazi, M.; Zhang, N. Possible Therapeutic Mechanisms and Future Perspectives of Vaginal Microbiota Transplantation. Microorganisms 2023, 11, 1427. [CrossRef]
- Meng, Y.; Sun, J.; Zhang, G. Vaginal microbiota transplantation is a truly opulent and promising edge: fully grasp its potential. Front. Cell. Infect. Microbiol. 2024, 14, 1280636. [CrossRef]
- Martinelli, S.; Nannini, G.; Cianchi, F.; Staderini, F.; Coratti, F.; Amedei, A. Microbiota Transplant and Gynecological Disorders: The Bridge between Present and Future Treatments. Microorganisms 2023, 11, 2407. [CrossRef]
- Lyra, A.; Ala-Jaakkola, R.; Yeung, N.; Datta, N.; Evans, K.; Hibberd, A.; Lehtinen, M.J.; Forssten, S.D.; Ibarra, A.; Pesonen, T.; et al. A Healthy Vaginal Microbiota Remains Stable during Oral Probiotic Supplementation: A Randomised Controlled Trial. Microorganisms 2023, 11, 499. [CrossRef]
- Wrønding, T.; Vomstein, K.; Bosma, E.F.; Mortensen, B.; Westh, H.; Heintz, J.E.; Mollerup, S.; Petersen, A.M.; Ensign, L.M.; DeLong, K.; et al. Antibiotic-free vaginal microbiota transplant with donor engraftment, dysbiosis resolution and live birth after recurrent pregnancy loss: a proof of concept case study. EClinicalMedicine 2023, 61, 102070. [CrossRef]
- Zhang, L.; Li, Q.; Su, Y.; Zhang, X.; Qu, J.; Liao, D.; Zou, Q.; Zou, H.; Liu, X.; Li, C.; et al. Proteomic profiling analysis of human endometrium in women with unexplained recurrent spontaneous abortion. J. Proteom. 2023, 288, 104996. [CrossRef]
- Cuadrado-Torroglosa, I.; García-Velasco, J.A.; Alecsandru, D. Maternal–Fetal Compatibility in Recurrent Pregnancy Loss. J. Clin. Med. 2024, 13, 2379. [CrossRef]
- Wang, W.; Zhou, X.; Zhang, Y.; Chen, Z.; Huang, J.; Zhang, X.; Kwak-Kim, J. The characteristics of antigenic specificity of memory regulatory t cells in women with unexplained recurrent pregnancy loss. J. Reprod. Immunol. 2022, 154, 103694. [CrossRef]
- Husain, S.; Allotey, J.; Drymoussi, Z.; Wilks, M.; Fernandez-Felix, B.; Whiley, A.; Dodds, J.; Thangaratinam, S.; McCourt, C.; Prosdocimi, E.; et al. Effects of oral probiotic supplements on vaginal microbiota during pregnancy: a randomised, double-blind, placebo-controlled trial with microbiome analysis. BJOG: Int. J. Obstet. Gynaecol. 2020, 127, 275–284. [CrossRef]
- Marcotte, H.; Larsson, P.G.; Andersen, K.K.; Zuo, F.; Mikkelsen, L.S.; Brandsborg, E.; Gray, G.; Laher, F.; Otwombe, K. An exploratory pilot study evaluating the supplementation of standard antibiotic therapy with probiotic lactobacilli in south African women with bacterial vaginosis. BMC Infect. Dis. 2019, 19, 1–15. [CrossRef]
- Giannella, L.; Grelloni, C.; Quintili, D.; Fiorelli, A.; Montironi, R.; Alia, S.; Carpini, G.D.; Di Giuseppe, J.; Vignini, A.; Ciavattini, A. Microbiome Changes in Pregnancy Disorders. Antioxidants 2023, 12, 463. [CrossRef]
- Tersigni, C.; Barbaro, G.; Castellani, R.; Onori, M.; Granieri, C.; Scambia, G.; Di Simone, N. Oral administration of Bifidobacterium longum ES1 reduces endometrial inflammation in women with recurrent pregnancy loss. Am. J. Reprod. Immunol. 2024, 91, e13804. [CrossRef]
- Vomstein, K.; Vomstein, K.; Krog, M.C.; Krog, M.C.; Wrønding, T.; Wrønding, T.; Nielsen, H.S.; Nielsen, H.S. The microbiome in recurrent pregnancy loss – A scoping review. J. Reprod. Immunol. 2024, 163, 104251. [CrossRef]
- Nnamonu, E.; Mgbenka, B.O.; Mbegbu, E.C. Impact of omega-3 fatty acids preconception intake on some fertility parameters and foetuses quality of female rats. 2020, 21, 115–119.
- Di Pierro, F.; Sinatra, F.; Cester, M.; Da Ros, L.; Pistolato, M.; Da Parè, V.; Fabbro, L.; Maccari, D.; Dotto, S. et al. Effect of L. crispatus M247 Administration on Pregnancy Outcomes in Women Undergoing IVF: A Controlled, Retrospective, Observational, and Open-Label Study. Microorganisms. 2023 Nov 17;11(11):2796.
- Mu, F.; Huo, H.; Wang, M.; Wang, F. Omega-3 fatty acid supplements and recurrent miscarriage: A perspective on potential mechanisms and clinical evidence. Food Sci. Nutr. 2023, 11, 4460–4471. [CrossRef]
- Kello, N.; Cho, Y.M. Natural supplements in antiphospholipid syndrome: A case for further study. Clin. Immunol. 2024, 258, 109848. [CrossRef]
- Li, Z.; Zheng, Y.; Zhang, M.; Wu, K.; Zhang, L.; Yao, Y.; Zheng, C. Gut microbiota-derived metabolites associate with circulating immune cell subsets in unexplained recurrent spontaneous abortion. Heliyon 2024, 10, e24571. [CrossRef]
- Kemp, M.; Newnham, J.; Challis, J.; Jobe, A.; Stock, S. The clinical use of corticosteroids in pregnancy. Hum. Reprod. Updat. 2016, 22, 240–259. [CrossRef]
- Giulini, S.; Grisendi, V.; Sighinolfi, G.; Di Vinci, P.; Tagliasacchi, D.; Botticelli, L.; La Marca, A.; Facchinetti, F. Chronic endometritis in recurrent implantation failure: Use of prednisone and IVF outcome. J. Reprod. Immunol. 2022, 153, 103673. [CrossRef]
- Hart, R.J. Nutritional supplements and IVF: an evidence-based approach. Reprod. Biomed. Online 2023, 48, 103770. [CrossRef]
- Piekarska, K.; Dratwa, M.; Radwan, P.; Radwan, M.; Bogunia-Kubik, K.; Nowak, I. Pro- and anti-inflammatory cytokines and growth factors in patients undergoing in vitro fertilization procedure treated with prednisone. Front. Immunol. 2023, 14, 1250488. [CrossRef]
- Su, Q.; Pan, Z.; Yin, R.; Li, X. The value of G-CSF in women experienced at least one implantation failure: a systematic review and meta-analysis. Front. Endocrinol. 2024, 15, 1370114. [CrossRef]
- Yao, K.; Sun, Y.; Ye, X.-Q.; Wu, Y. Interferon-λ contributes to endometrial receptivity. Reproduction 2023, 165, 569–582. [CrossRef]
Figure 1.
General overview of the differences between eubiosis and dysbiosis in the vagina. In the vaginal lumen, the expected protective effect of immunoglobulins, complement proteins, antimicrobial peptides, peroxide production, and lactic acid. In dysbiosis, the protective effect is lost, and the inflammatory response is due to bacterial proteins, increasing cell death inflammatory mediators. This increase in inflammatory mediators leads to a decrease in vaginal tolerogenic milieu which is the response to the reduction of annidation and increase of pregnancy loss.
Figure 1.
General overview of the differences between eubiosis and dysbiosis in the vagina. In the vaginal lumen, the expected protective effect of immunoglobulins, complement proteins, antimicrobial peptides, peroxide production, and lactic acid. In dysbiosis, the protective effect is lost, and the inflammatory response is due to bacterial proteins, increasing cell death inflammatory mediators. This increase in inflammatory mediators leads to a decrease in vaginal tolerogenic milieu which is the response to the reduction of annidation and increase of pregnancy loss.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



