
Submitted:
08 July 2024
Posted:
09 July 2024
You are already at the latest version
Abstract
Background: Glioblastoma (GBM) is a highly aggressive, invasive, and growth-factor-independent grade IV glioma. Survival following the diagnosis is generally poor, with a median survival of approximately 15 months, and it is considered the most aggressive and lethal central nervous system tumor. Conventional treatments based on surgery, chemotherapy, and radiation therapy only delay progression, and death is inevitable. Malignant glioma cells are resistant to traditional therapies, potentially due to a subpopulation of glioma stem cells that are invasive and capable of rapid regrowth. Methods: Systematic retrieval of information was performed on PubMed. Specified keywords were used in PubMed and the articles published in peer-reviewed scientific journals were associated with brain GBM cancer, sodium iodide symporter (NIS). Additionally, the words 'radionuclide therapy', 'radioiodine', 'iodine-131', 'molecular imaging', 'gene therapy', and 'translational imaging' were used. Other keywords such as ‘glioblastoma', 'targeted', 'theranostic', 'symporter', 'virus', 'solid tumor', 'combined therapy', 'pituitary', or 'plasmid' were also used in appropriate literature databases and search engines. 19 articles were found in this search on Mesenchymal Stem Cell Sodium Iodide Symporter and GBM. \ These articles were from the years 2000 to 2024. Appropriate studies were isolated, and important information from each of them was understood and entered into a database from which the information was used in this article. Results: For as long as they express functional NIS, mesenchymal stem cells systemically repress T and B cells; however, after that, they are largely replaced by mesenchymal stem cells generated from embryonic stem cells that are functionally competent. As a result of their natural capacity to identify malignancies, MSC are employed as tumor therapy vehicles. Because MSCs may be transplanted in several methods, they have been proposed as ideal vehicles for NIS gene transfer. Conclusion: Non-invasive imaging-based detection of glioma stem cells presents an alternate means to monitor the tumor and diagnose and evaluate recurrence. The sodium iodine symporter gene is a specific gene in a variety of human thyroid disease that functions to move iodine into the cell. In recent years, an increasing number of studies related to the sodium iodide symporter gene have been reported in a variety of tumors and as therapeutic vectors for imaging and therapy. Gene therapy and nuclear medicine therapy for GBM provide a new direction.
Keywords:
Glioblastoma
; targeted radionuclide therapy
; Neurosurgery
; brain tumor
; cancer
1. Introduction
Glioblastoma (GBM), also known as glioblastoma multiforme, is the most common glioma, which belongs to the aggressive and malignant type of brain tumor. At present, GBM has a low survival rate and the effective therapy is limited [1]. Histopathologically, GBM is composed of both the diffuse infiltration zone and the main tumor mass [1]. Conventional treatment such as surgical resection, adjuvant radiotherapy, and concomitant and adjuvant temozolomide chemotherapy has only limited effects [1,2]. The newly adjuvant photodynamic therapy proposed by our team to be combined with temozolomide chemotherapy also has few side effects [2]. However, the recurrence rate and the median survival period of patients with GBM are still significantly worse than other solid malignant tumors [2,3]. Research on using the sodium iodide symporter (NIS) gene to label cancer can open the door for GBM radionuclide theranostics, yet clinical applications are limited due to the limited labeling efficiency [4]. Glioma stem cells express the receptor for the glial cell-derived neurotrophic factor, and mutations can result in the glial cell-derived neurotrophic factor and signal transduction pathways constitutively active in GBM cells [5]. Due to the poor prognosis of patients with GBM, there is an urgent need to research effective new adjuvant treatments [1,5]. The lack of specific target expression in GBM stem cells effectively limits targeted therapy [6]. Non-invasive imaging-based detection of glioma stem cells presents an alternate means to monitor the tumor and diagnose and evaluate recurrence [6]. The sodium iodine symporter gene is a specific gene in a variety of human thyroid disease that functions to move iodine into the cell [7]. In recent years, an increasing number of studies related to the sodium iodide symporter gene have been reported in a variety of tumors and as therapeutic vectors for imaging and therapy. Gene therapy and nuclear medicine therapy for GBM provide a new direction [7]. Key therapeutic targets of GBM have relatively easily been found and are still being searched for. Radionucleotide therapy seems promising in the treatment of solid malignant tumors. As for the currently increasing number of studies on NIS radionucleotide therapy in vivo and in vitro, we aimed to focus on the therapy in GBM using the effective and feasible image-guided strategy to guide radiotherapy in order to develop a new clinical therapeutic strategy with societal impact. NIS is a transmembrane glycoprotein found primarily on the basolateral surface of the follicular cells in the thyroid, but also expressed in the parafollicular cells of the thyroid, lactating mammary glands, stomach, and a host of other tissues [8]. It has a highly typical and unique function of concentrating and incorporating iodine, which it usually does by pumping iodide into thyroid cells, where it is then oxidized and incorporated into the precursor molecule of the two thyroid hormones (T3 and T4) [8]. This iodine-concentrating capability of NIS is preserved in well-differentiated thyroid cancers and exploited in the form of radioiodine therapy after pre-treatment with radioactive iodine isotopes, particularly 131I, following resection of the diseased thyroid tissues [8]. It is the radioactivity created by the breakdown of 131I that removes the threat of any remaining cancerous thyroid tissues. The competitive inhibitors thiocyanate and perchlorate, as well as the Na+Ka+-ATPase inhibitor ouabain, can all block NIS-mediated iodide transport [9]. For the past 80 years, radioiodide has been utilized extensively in the treatment of differentiated thyroid cancer. This is due in part to the foundation of functional NIS expression. The “crossfire effect,” which is the radiation of accumulated radioisotopes in NIS-expressing cells on nearby non-expressing cells by particle decay, is linked to the cytoreductive effect of targeted NIS-mediated radioisotope treatment [10]. Beyond the application of NIS as a reporter gene for gene therapy with gene-directed enzyme/prodrug cancer therapy, in recent years, there has been great interest in applying NIS as a theranostic tool using radioiodine therapy, screening systems for the development of NIS-based radionuclide diagnostic and therapeutic strategies. Central to these undertakings, the overall goal is to build up sustaining concentrations of iodide and radioactive iodide that can continue to a level leading to radiation-induced lethal DNA double-strand breaks in tumor cells. With such a powerful built-in negative feedback loop, the function of the iodine concentration mechanism in turning on the expression of the hNIS transgene is necessary to cause tumor cell death [11].
2. Mesenchymal Stem Cells as Carriers of NIS Genes
Mesenchymal stem cells supply bone marrow stroma and have in vitro differentiation capacity into chondrocytes, osteoblasts, and adipocytes. As supporting cells in peripheral tissues, MSCs are closely related to their secretory functions in physiological homeostasis, tissue repair, and fibrosis. Additionally, MSCs exert immunoregulatory effects, improving multiple diseases, such as Graft Versus Host Disease (GVHD). MSCs have been immediately utilized after transplantation due to their low immunogenicity and the expression of MHC-class-I combination molecules, HLA-G, B7-H1, PD-L1, IDO, and prostaglandin E2, which can inhibit T cell and NK cell responses, particularly MHC-class-II negative peripheral blood MSCs, which can induce immune tolerance similar to fetal MSCs [12]. However, these molecules and cytokine-mediated immunoregulatory functions are unstable and are easily influenced by the cell microenvironment, such as inflammation and hypoxia [13]. Prior research found that affected tissues release more IL-1β, IL-6, and TNF-α when MSCs are transplanted. MSCs have a strong homing ability for inflammatory tissues and have abundant blood supply and rich nutrient sources [13]. After 6 weeks of injection, a small number of MSCs are undifferentiable and can be detected in the lungs, kidneys, and liver. However, they can only be detected by detection techniques, such as luciferase, PCR, qPCR, lentivirus-GFP, and cyclophosphamide-GFP, and fewer studies have used animal models of heart disease specifically [14]. In addition, MSCs possess a large, flat, spindle-shaped morphology, representing an adherence-independent characteristic of suspension culture, and are not sensitive to serum-containing medium or carrier contact. After a certain period of exponential growth, MSCs may enter the G0 phase, exhibiting cell senescence, cell aging, cell death, and G1/S blocking, thereby negatively impacting proliferation [15]. Finally, MSCs are recognized as the nuclei and cells by karyotyping techniques, which change the chromosomal number and affect cell differentiation potential, the secretion of cytokines, biological characteristics, and tumorigenicity [16].Bone marrow-derived mesenchymal stem cells and umbilical cord blood-derived mesenchymal stem cells are the only stem cell types that can be banked following plastic adherence and long-term expansion [15]. Mesenchymal stem cells systemically suppress T and B cells as long as they express functional NIS but thereafter are partially replaced by functionally competent embryonic stem cell-derived mesenchymal stem cells [16]. Co-transduction of NIS and thyroid peroxidase (TPO) for the non-immunogenic target-specific NIS-probe is another [16]. Specifically, mesenchymal stem cell attachment onto NIS-expressing primary non-thyroid cells may be the easiest if the NIS expression level is checked prior to initial stem cell isolation [16]. The potency by heterologous or allogeneic NIS-mesenchymal stem cells defined here should stimulate further in vivo NIS/gene delivery model development. In 2003, Herzog et al. proved that BM-derived MSCs could migrate to damaged brain tissues and transform into neurons, astrocytes, and oligodendrocytes [17]. The observation showed MSCs participated in neurodegenerative disease treatments. The BM-derived MSCs could also cure the chronic spinal cord injury in mouse. Intravenous transplantation of MSCs reduced astrogliosis, induced oligodendrocyte synthesis, and promoted serotonergic axon regeneration [17]. In 2003, Angelopoulou et al. investigated the property of BM-derived MSCs for expressing SP during the adipogenic differentiation [18]. This finding identifies the direct action of MSCs to angiogenesis. Furthermore, BM-derived MSCs take part in the skeletal muscle repairing process. After muscle injury in mice, the injected BM-derived MSCs repair the muscle and downregulate migration inhibitory factor. MSCs could also participate in the recovery from lung injury [19]. It created an antifibrogenic effect and restored cell proliferation of alveolar type II. Mesenchymal stem cells (MSCs) are self-renewing, pluripotent, and can differentiate into tissues like adipocytes, chondrocytes, osteoblasts, etc. Many kinds of MSCs could express normal tissue-specific proteins when seeded or transplanted in some kinds of tissues. Umbilical cord blood-derived MSCs are CD34-negative, CD45-negative, and HLA-DR-negative. Human umbilical cord blood-derived MSCs could differentiate into cells expressing liver-function proteins [20]. Human placental- or umbilical cord blood-derived MSCs are suitable for obtaining CB, and the process of collecting CB sample is noninvasive. It avoids donor morbidity and infection risks so that the safety issue is reduced. Moreover, it has little HLA identity, so transplantation of embryonic stem cells (ESCs) or MSCs from CB may reduce the side effects of immune rejection [21]. The characteristic of low immunogenicity and potent immunosuppression makes MSCs suppress the immune rejection, solve engraftment difficulties, and ameliorate the stem cell niche environment. Remarkably, the umbilical cord blood-derived MSCs sustain high paracrine activity with high inflammatory cytokine levels. Cell damage and cryopreservation strategies impact paracrine activity [22].
2.1. Origin and Differentiation Potential
MSCs are an adult and multipotent non-hematopoietic cell type. MSCs can differentiate across the mesodermal lineages, i.e., chondroblastic, adipogenic, osteoblastic, myoblastic, and tenoblastic lineages, and give rise to other connective tissue cells, i.e., fibroblasts, endothelial, and epidermal cells [23]. Additionally, MSCs differ from other stem cell types in that they are relatively easily procured from adults, induced to proliferate in culture, and employed in cell-based bioengineering strategies. These unique features have provided strong rationales for extensive empirical investigations and applications, and there has been a concomitant explosive interest in the cellular and molecular biology of MSCs en route to understanding their fundamental properties and designing useful translational strategies. MSCs that can undergo adipocyte, chondrocyte, osteoblast, and other lineage differentiation were initially isolated from rodent bone marrow by Friedenstein [24]. However, the MSCs exist not just in the bone marrow; they are scattered throughout various tissues, such as the subcutaneous tissue, dental pulp, kidney, amniotic fluid, cord blood, and adipose tissue. Among these resident MSC populations, adipose MSCs (ASCs) and bone marrow MSCs (BMSCs) have been intensively identified for their multi-lineage differentiation and powerful homeostasis [25,26]. Especially, ASCs are easier to obtain and their source is abundant in comparison with BMSCs, thus they are potential MSCs in medicine and research. Therefore, mesenchymal stem cells are considered to be effective target cells for the deep therapy of various diseases.
2.2. Immunomodulatory Properties
Human mesenchymal stem cells (hMSCs) are self-renewing progenitors best known for differentiation to mesenchymal lineage cells such as osteocytes, adipocytes, and chondrocytes. hMSCs can also differentiate to endodermic-like cells, for example hepatocytes, and neurogenic-like cells, for example neurons and glia; these cells can participate in tissue repair and regeneration mainly through paracrine effects. Surrounding the MSC and microenvironment in which they are located significantly affect their potential role in the immune response [27]. Endogenous MSCs are found in bone marrow, pericytes from various tissues, fibroblasts, and neural cells. Due to the low number of MSCs in human tissues, explantation techniques are used to expand these cells in in vitro cultures. In clinical trials, MSCs have been shown to be safe and hold particular promise for their immunophysiological properties in the treatment of various diseases [28].
MSCs-associated features include their apparent exclusion from the different types of lymphoid and myeloid cells; they are immunosuppressive. These facts, together with the property of suppressing proliferation of peripheral blood lymphocytes that do not produce lymphokines, can absorb or modulate immune responses [29]. Although these effects have been extensively studied in a large number of in vitro allogeneic and xenogeneic studies, MSC actions in vivo are less defined. In particular, labeling, ex vivo expansion, and reimplantation of MSCs showed that, a few days later, they were rejected in a reaction mediated by T-cells or NK cells. Lower immunosuppressive impacts in vivo were observed, although one working mechanism of MSC action includes their participation in the suppression of lymphopoiesis in the bone marrow [30]. Control of lymphopoiesis is achieved through withdrawal of IL-7 and CXCL12 support from the stem/progenitor cells. MSCs lack HLA class II expression. Even in conditions in which major histocompatibility complex class I expression was enhanced, few NK-activating molecules of MSCs were observed [31]. Most of the evidence is consistent with an immune escape mechanism. MSC immunosuppression was effective either in the presence or absence of a descending number of specific patient lymphoid cells. Furthermore, the suppressive effects of patient hMSCs were maintained in peripheral blood lymphocyte samples harvested after transplant, suggesting a resistance to tolerance factors [32,33]. The mechanism(s) that MSCs use to suppress immune responses are complex, but mainly involve localized effects via release of soluble factors. The immunosuppressive pattern of MSCs was also involved in the selective accumulation of these cells in some tumors [34]. Okolicsanyi and Griffiths assessed the isolation, expansion, and differentiation of WJ-MSCs and U-CMSCs isolated from different gestational age Wharton’s jelly (WJ) into osteoblasts, adipocytes, and neural progenitor cells and the hMSC cell surface markers, revealing that WJ-MSCs may be a potential stem cell therapy source for nervous system injury, and that the immunophenotypes of the expanded isolated U-CMSCs did not change with in vitro culturing [35]. These cells maintained an extraordinary growth rate, while the WJ-MSCs maintained their structural and functional features. Balakrishnan assessed the immunomodulatory properties of different sources of hMSCs priced the comparison based on the intensity of expression of immunomodulatory and cytokine receptors, and concluded that WJMSCs demonstrate intense anti-inflammatory and anti-apoptotic mechanisms, such as release of immunoregulatory and pro-angiogenic factors [36]. In summary, the results presented by Balakrishnan and Okolicsanyi & Griffiths provide evidence for the potential role of umbilical cords in using U-CMSCs for immune system regulation in different disorders.
2.3. Tumor-Tropic Migration
MSCs have the unique ability to migrate towards tumors. Studies have determined the migration ability of both mouse MSCs and human MSCs towards a variety of tumor types, both in vitro and in vivo [33]. As a consequence, MSCs were found to localize and engraft in various tumor models following transplantation into animals. Interestingly, MSCs possess the ability to specifically migrate towards sites of sterile inflammation and ischemia, and to migrate towards sites of allogeneic hematopoietic stem cell transplantation [37,38]. Although the natural physiological function of MSCs to migrate towards sites of injury and inflammation is beneficial in tissue repair, it has been suggested that MSCs could potentially enhance tumor metastasis through their migration tropism [39]. Some groups hypothesized that MSCs have anti-tumor abilities based on evidence showing that some cytokine-activated MSCs can inhibit the growth of various tumor type cells in vitro [40]. Filing of MSCs with some cytokines, such as interferon-c (IFN-c) can enhance the in vitro anti-tumor ability of MSCs. In addition, results from a number of in vivo studies clearly show that MSCs can inhibit tumor growth if they are coadministered with appropriate cytokines such as GM-CSF or IFN-c [41]. There is much controversial data on the interaction between MSCs and tumor cells: MSCs can enhance or inhibit tumor cell invasiveness in the presence or absence of tumor necrosis factor-a (TNF-a). The existing knowledge about the interaction between MSCs and different types of tumor cells imply that the design of MSC-based gene delivery as an “effector” therapy against systemic malignant tumors should take into account not only the tumor-tropism of MSCs, but also the effect of the tumor on the MSCs themselves, in particular, how MSCs are activated, reprogrammed or get sick when they meet tumor cells, and how these phenomena would synergistically impact on the ultimate gene therapy efficacy [42]. Due to their innate ability to locate tumors, mesenchymal stem cells (MSC) are used as vehicles for tumor treatment. Growth factors, chemokines, and inflammatory cytokines are all produced at higher levels in tumors, which encourages MSCs to actively recruit into the tumor microenvironment and aid in the formation of the tumor stroma [43]. Because MSCs are easily extracted, amplified, and transplanted across the allogenic barrier, they are highly suited for therapeutic uses. MSCs that have undergone genetic engineering hold promise as delivery systems for therapeutic genes like NIS. In early-stage human clinical studies, the use of modified MSCs to treat solid tumors is currently being investigated [44]. Preclinical research utilizing xenograft tumor mouse models has shown the potential of CMV (cytomegalovirus) promoter driven MSC-mediated NIS gene delivery, with successful selective NIS expression in tumors and metastases as well as a strong therapeutic response following [131I]NaI application [45]. MSC engineering has been exploited for cell-mediated gene therapy and for carrying therapeutic transgenes that protect the engrafting cells from the conditioning cytotoxic insult or that reinforce the functions of the stem/progenitor cells in the host recipients, promoting their engraftment, the differentiation processes, or conditioning a permissive local environment [46]. However, the beneficial effects have been obtained only in a subset of the genetically manipulated cells, and the strategies available are still very expensive and potentially associated with risk factors. Some laboratories have reported results with a more efficient MSC transduction, alongside with the use of different transduction systems such as commercial viral vectors or the production of viral vectors derived from in-house developed stable packaging cells [47,48]. Radioisotope therapy can be successfully carried out with cells expressing the sodium iodide symporter (pNIS). Mesenchymal stem cells (MSCs) have been put forward as desirable vehicles for NIS gene transfer since they can be transplanted in a variety of different ways [49]. They can contribute to viral clearing in the in vivo experiments and do not replicate in the target tissue, therefore providing tumor-specific pNIS expression [50]. This latter property can significantly reduce the potential toxicity when the NIS-targeted malignant cells are efficiently radiouptaken with gamma emitter radionucleotides.
3. Mechanism of NIS Gene Delivery
Large number of gene-transfected human MSCs could transmigrate into the intracranial lesions expressing the transferred NIS under cytokine enhancement, and the NIS-expressing cells could effectively concentrate radioiodine, therefore causing the accumulation of activity in vivo [51]. Moreover, the in vivo data of high retention time and high tumor to normal tissue ratio indicate the technique specificity and therapeutic potential. The manipulable MSCs pool easily obtained from humans is capable of reliably carrying and expressing the target gene, thus providing a feasible way for adoptive resist-tumor radiotherapy [52,53]. Research reported that human umbilical cord blood (hUCB) and human bone marrow (hBM)-derived mesenchymal stem cells (MSCs) were efficiently transfected with a plasmid vector expressing the marker sodium iodide symporter (pNIS) based on RRL-mediated gene transfer method [54,55]. The MSCs gene transfected were capable of migrating to basal ganglia of the infants. The NIS on the membrane could extract iodide from the media and then concentrate radioiodine in the cells [56,57]. Data showed that large numbers of administered gene transfected human BM-MSCs could transmigrate into the intracranial lesions and expressed the transferred human NIS gene under stimulations of granulocyte-macrophage colony stimulating factor (GM-CSF) or endostatin [58,59]. The labeled MSCs distributed in basal ganglia of the brains were assayed in the lymphoma-bearing mice by single photon emission computed tomography [60].
Advantages and Limitations
Multiple potential advantages have been proposed for the use of MSCs to deliver NIS. First, MSCs can be readily isolated from a patient’s bone marrow and expanded under GMP conditions, thus providing an autologous, relatively inexpensive source of cells that are well tolerated in most patients [61]. Second, the migratory and tropic properties of MSCs likely contribute to MSC distribution in areas of injury, including various premetastatic niches, and facilitates MSC extravasation through basement membranes and endothelial cell tight junctions to engraft into the target tissue, particularly following tissue-specific damage [62]. Despite the advantages of utilizing MSCs as a gene delivery vehicle, a number of important considerations also need to be addressed. Direct injection of radiolabeled MSCs into malignant tumors could reduce tumor burden by depositing what would be a therapeutic dose of beta radiation with the added advantage of localizing only in tumor, not in normal tissues [63]. At the recent NRVS Congress, an abstract by Lee et al. presented phase I data using directly injected Ad-TC-M1-MSCs into patients with metastatic breast cancer lesions and reported both safety and patients’ derived pain reduction [64]. MSCs have been found effective for several diseases in phase I/II clinical studies but have yet to show a convincingly large effect when tested in additional patients in Phase III trials. There are several obstacles to stem cells themselves that need to be overcome, such as the inability to drive differentiation into the appropriate lineage, poor survival of the implanted cells, and the inability of the implanted cells to cooperatively function on a large scale due to nonrepair cells [65]. In cancer treatment using stem cells as gene delivery vehicles, additional obstacles prevent successful treatments. Since tumors are heterogeneous, it is unlikely that a uniformly radiolabeled MSC population can uniformly target a solid mass [66]. Furthermore, MSCs are part of an immunosuppressive environment of the tumor that generates them—and efforts to use transformed MSCs that are not immunomodulatory improve cell survival and the effect of MSC. The application of endogenous MSCs can avoid an immunological response to allogeneic antigens [67]. Many cancer entities are MSC-homing, and MSCs have been demonstrated to be capable of carrying the NIS-transgene gene to a primary or metastatic cancer site to synthesize the NIS protein, which can trigger cancer cell death. MSCs also work as a stromal component that can resist forming radiation-induced immune cell populations and abrogating immune-based tumor rejection in postponed tats [68]. Although all of these characteristics make MSCs an optimal gene delivery vehicle, there are still some disadvantages. First, a suitable method for MSC transplantation and the timing of NIS-MSC transplantation for a better outcome have not been determined. Second, the final destination of NIS-MSCs and the dynamic changes at the molecular level of the target tumor-expressing NIS have not been analyzed [69]. Third, the probable tumor-forming risk resulting from the interaction between MSCs and the activated host’s microenvironment also remains unclear. In addition, MSCs can also be dedifferentiated to regain tumorigenic potential after being modified by the NIS gene, which can limit the applicability of MSCs as a gene delivery vehicle. Somatic cell dedifferentiation is concerning for future applications, in particular, the cell sources for reprogramming are used as carriers for gene therapy [70]. Taken together, based on these findings, we foresee that a combination therapy approach will be required to achieve effective antitumor therapy with the NIS gene to treat cancer.
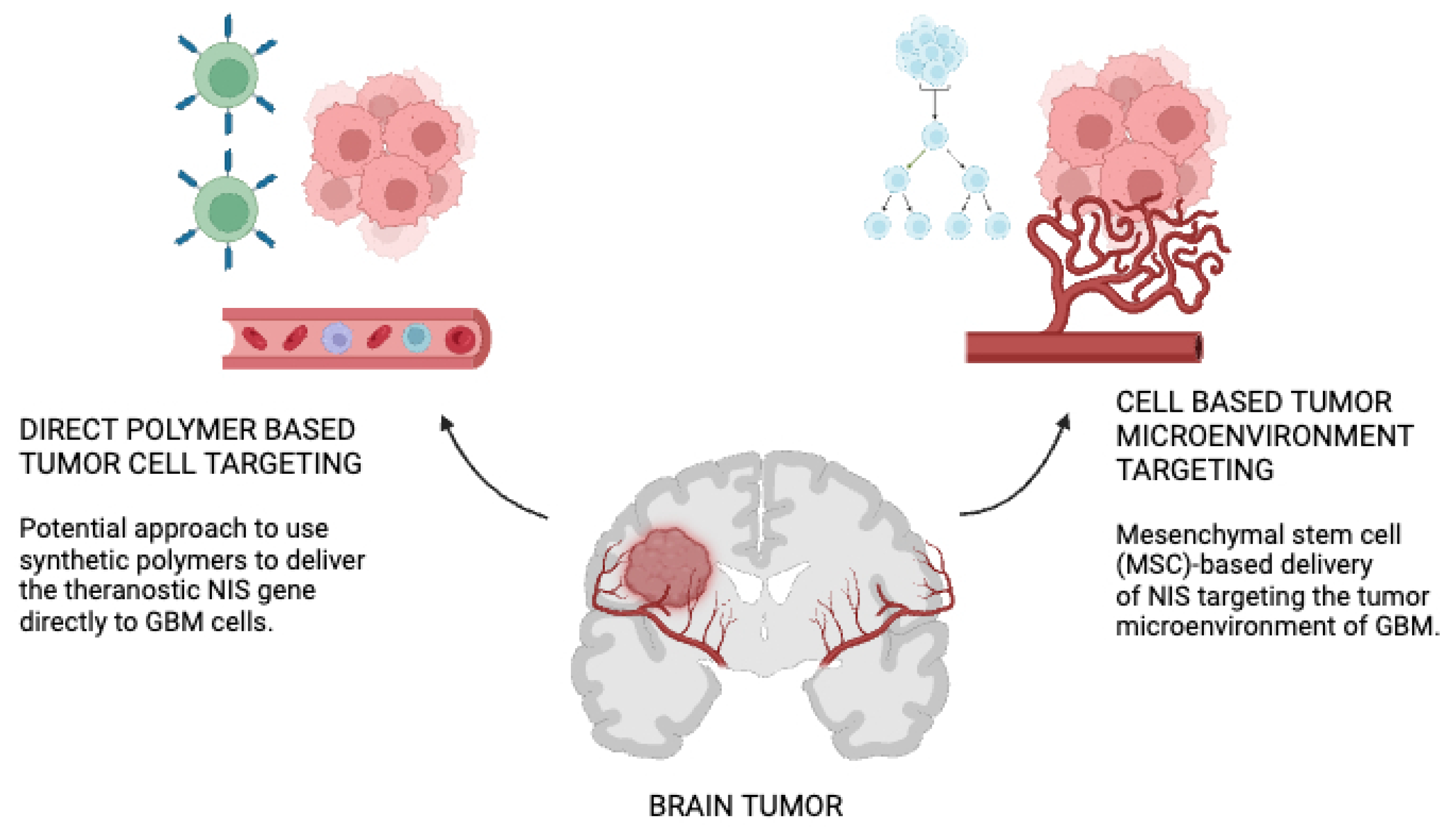
NIS gene delivery strategies to GBM has been depicted in Figure 1. On the left hand side is a potential approach to use synthetic polymers to deliver the theranostic NIS gene directly to GBM cells.The polymer backbone is functionalized with ligands (targeting domain) that have a high affinity to cell surface receptors that are overexpressed in GBM cells. Polymers are loaded with NIS pDNA. Following systemic administration of polymers, the pDNA is released to the GBM cells after binding of the polymer to the cell receptor [71]. On the right hand side is the Mesenchymal stem cell (MSC)-based delivery of NIS targeting the tumor microenvironment of GBM. MSCs can be easily isolated from patients from different tissue sources (e.g., bone marrow or adipose tissue) and genetically modified with the NIS gene under the control of tumor-stroma specific gene promoters. Engineered MSCs can be amplified in the laboratory and systemically administered back to the patient or over the allogenic barrier [72]. Tumor-secreted factors (e.g., inflammatory cytokines) promote direct migration and extravasation of MSCs to GBM where they become part of the tumor stroma. NIS expression is induced after promoter activation. Following successful NIS gene transfer using both delivery platforms, diagnostic and therapeutic application of radioactive NIS substrates can be applied. pDNA; plasmid DNA [72].
4. Image-Guided Approaches in Radionucleotide Therapy
The NIS, under the control of a modifying and specific promoter, can guide an enhanced and safer clinical translation potential with the capability for real-time noninvasive imaging that helps evaluate drug delivery, allowing for optimization and might even direct personalized therapy [73]. The use of the herpes simplex virus type 1 thymidine kinase (HSV1-tk) as reporter genes for PET imaging has been reported in preclinical models as a potential for selectivity and noninvasive evaluation of genetic immunotherapy for GBM [73]. Magnetic resonance imaging and other more advanced imaging techniques have also been explored to extract other information from NIS expression and gene therapy response in GBM as compared to NIS radionucleotide therapy. Each literature method has indications and limitations, the choice should rely on resources available and specific questions addressed.NIS is a potent theranostic gene that, as the preceding sections have shown, enables effective molecular therapy monitoring following radionuclide administration [74]. NIS possesses several attributes of an ideal reporter gene as well: This protein is found spontaneously in thyroid follicular cells and is neither immunogenic nor cytotoxic to cells. Since iodide buildup may only take place in active cells, cell viability is linked to functional NIS activity. The signal is concentrated due to the buildup of radiolabeled substrates caused by the active transport of substrates [75]. As a result, compared to a reporter that only binds its substrate stoichiometrically, the detection sensitivity is greater. Because NIS translocates different substrates, it may be used to localize NIS-positive cells using a variety of conventional nuclear medicine imaging methods. Gamma scintigraphy and single-photon emission computed tomography (SPECT) are made easier by the active transport of 123I, 125I, 131I, 99mTc, and 188Re. Furthermore, due to their beta decay, 131I and 188Re are useful radionuclides for medicinal purposes [76]. In clinical settings, planar scintigraphy or SPECT have been the primary tools for molecular imaging of NIS. On the other hand, PET imaging of functional NIS expression offers the possibility of enhanced sensitivity, resolution, and efficient quantitative analysis. The most well-known and often used positron emitter for NIS-mediated PET imaging in preclinical and clinical settings is 124I [77]. Recently, a new tracer for NIS-based PET imaging was developed. Due to the radiochemical and physical characteristics of 18F, 18F-Tetrafluoroborate (TFB) has been presented as a possible replacement for 124I. TFB offers a number of benefits for regular diagnostic use. [18F]Because of its shorter half-life (110 min vs. 100 h), branching ratio (97 vs. 23%), and most importantly, its lower positron energy (Emax; 0.634 vs. 2.14 MeV), TFB is superior than 124I in terms of producing PET images that are crisper and less blurred [78]. GBM is the most aggressive brain tumor in humans, and its current treatment strategies fall short of survival benefit. However, the recent advancements in molecular imaging allowed for improvements in the disease’s initial diagnosis and monitoring. The sodium iodide symporter (NIS) has been studied as a reporter gene to express NIS protein in GBM cells [79]. This results in NIS-expressing GBM to become a target for radionucleotide therapy using beta-emitting radionucleotides [80]. These radioisotopes, such as 131 iodine, have been shown to reduce GBM growth in preclinical and early clinical studies. But, maintaining an ideal drug delivery and therapeutic effect using radionucleotides in humans, and not only targeting NIS-expressing GBM cells but also the surrounding cells in the GBM microenvironment, need to be targetable with imaging or treatment [81].
5. Image-Guided NIS Radionucleotide Therapy in Glioblastoma
Image-guided radionuclide therapy (RNT) using the sodium iodide symporter (NIS) has demonstrated potential in preclinical animal models of GBM. However, there are only limited studies on its use in veterinary and human cancer patients to date. This narrative review considers the published work on NIS RNT in GBM cases and hopes to guide future investigator-initiated and hopefully some multicentered clinical studies [82]. These studies utilized more conventional intraoperative image-guided NIS targeted RNT than in other solid tumors due to logistic difficulties of radiation safety regulations and limitations in the NIS RNT agents applied, with concomitant radiation damage to normal brain tissue [83]. Novel therapeutic options for GBM are desperately needed, as it is a very complicated tumor that uses several pathways to avoid therapy. Due to its ability to block gene vectors and radiotracers, the blood-brain barrier (BBB) is one factor limiting the effectiveness of GBM therapy and detection. The small-sized radionuclides employed in NIS-mediated radionuclide imaging and treatment can diffuse into the tumor and pass through the blood-brain barrier without the need for complicated radiolabeling processes. Numerous preclinical investigations have exhibited the possible utility of NIS in glioma imaging and treatment [84]. A rat model with intracerebral F98 gliomas that had been retrovirally transduced with human NIS was utilized in a research by Cho et al. The scientists demonstrated the potential for non-invasive glioma imaging using [99mTc]pertechnetate- and [123I]NaI-scintigraphy, and they also demonstrated that rats receiving 131I treatment had longer survival times [85]. When the human glioma cell line U87 was injected into the right armpit of mice with xenografted tumors, it was transfected with a recombinant lentiviral vector carrying human NIS. Guo et al. [86] then reported imaging and therapeutic research using 188Re and 131I. According togamma camera imaging, in vivo imaging data demonstrated 188Re/131I accumulation in the NIS-containing tumors. Mice treated with 188Re or 131I saw an effective decrease in tumor volume when compared to untreated control mice. In another study, Opyrchal et al. [87] reported effective [123I]NaI or [99mTc]pertechnetate gamma camera or microSPECT/CT imaging of s.c. and orthotopic murine GBM xenografts following intratumoral infection with measles virus encoding NIS (MV-NIS) to induce NIS expression in brain tumor tissue. This study used one of the most extensively explored oncolytic viruses for NIS gene transfer. In both glioma scenarios, combined radiovirotherapy with MV-NIS and 131I produced better anticancer activity and survival than virotherapy alone. With MSCs designed to trigger NIS expression in response to IL-6 promoter activation, a novel tumor-targeted gene therapy strategy for GBM may be possible. This was studied by Kitzberger et al. [88] and found that When IL-6-NIS-MSCs were applied to tumors, there was an increase in radiotracer uptake by 18F-Tetrafluoroborate-PET/magnetic resonance imaging (MRI) as compared to animals who received wild-type MSCs. NIS protein expression in cancers was observed by ex vivo investigation of malignancies and non-target tissues. Following IL-6-NIS-MSC administration, 131I treatment dramatically slowed tumor development as measured by MRI and increased the median survival of GBM-bearing mice to 60% as compared to controls. In another recent study, they created dual-targeted NIS plasmid DNA complexes with targeting ligands for the EGFR and transferrin receptor (TfR), offering the possibility of active transport across the BBB and subsequent targeting of tumor cells. TfR- and EGFR-dependent transfection efficiency as well as NIS-specific iodide absorption of dual-targeted polyplexes were validated by in vitro125I transfection assays [89]. Employing 18F-labeled tetrafluoroborate (TFB) as a tracer, positron emission tomography (PET) imaging was used 48 hours after intravenous polyplex injection to evaluate in vivo gene transfer in mice harboring orthotopic U87 GBM xenografts. In comparison to animals treated with EGFR-mono-targeted polyplexes (0.33% ± 0.03% ID/mL) or TfR-mono-targeted polyplexes (0.27% ± 0.04% ID/mL), the tumoral 18F-TFB uptake of mice treated with dual-targeted polyplexes (0.56% ± 0.08% ID/mL) was considerably greater [90]. Application of 131I produced a better therapeutic impact of the dual-targeted therapy in therapy experiments, as evidenced by a notable delay in tumor development and an extended survival time.
6. Challenges and Future Directions Discussion on Image-Guided NIS Radionucleotide Therapy for Glioblastoma
In NIS gene therapy, the malignant cell should only acquire NIS expression that is both restricted to the tumor microenvironment compartment and configured to incorporate and concentrate the radioactive substrates, namely, radioiodine or other NIS substrates. Only self-destruct for this focalized area of NIS gene modification is desirable. Lastly, reduction in radiation exposure level and delaying long-term radiation effect on the remaining normal brain tissue constituent’s health by the NIS substrate after GBM is considered. The following challenges limiting the clinical development of NIS radionuclide therapy for GBM are discussed. Image-guided NIS radionuclide therapy for GBM is challenged by several factors herein summarized: lack of tumor-specific target, unwanted gene transfer to normal brain, confined gene expression within the tumor, specific overexpression level for NIS, the direct relationship of uptake level and efficacy, a minimized detrimental effect on the patient, and dose planning and prediction of therapeutic effect. Most NIS radionuclide therapy studies have adopted the strategy of radioiodide treatment following thyroidectomy or RAI refractory setting. In these patients, the level of RAI uptake in the tumors has been determined by the thyroglobulin test. However, several studies employed a fractionated dose schedule, and clinical progress was only detectable after 180 days. Moreover, it is desirable to repetitively administer RAI therapy to stop the progression of GBM during its repeated recurrences. In these settings, the novel tumor targeting principles for NIS radionuclide therapy are crucial.
7. Conclusion
While image-guided NIS radionuclide therapy has the potential to improve GBM treatment, several steps need to be addressed to translate the preclinical results into meaningful clinical practice. In all the studies, invasion using the ablation technique was carried out immediately prior to the administration of both NIS-hAMSC and 131I. The schedule does not reflect the actual benchmarks of patient care. Moreover, patients cannot be judged based on the metastasis characteristics of mouse models. They were supposed to eat before 131I administration in the report, but the actual treatment may not sustain when translated to the clinical application. The duration and intensity of the treatment need further optimization, and the present study cannot completely replace preexisting therapy methods. This marks a foundation for adjuvant or palliative therapy for poor prognosis gliomas, especially in elderly patients. With the continuous development of image-guided NIS radionuclide therapy, better efficacy can be achieved, and frontal lobe function damage can be avoided or minimized. The prevalence of GBM and the absence of standard effective treatments for the condition have contributed to research on possible treatment breakthroughs, among them the sodium ion symporter radionuclide therapy. In this narrative literature review, the process of opening the blood-brain barrier to establish T cell recruitment to the GBM niche before the administration of anti-PD-1 has the potential of providing a more efficient protocol for GBM treatment especially with the development of image-guided NIS radionuclide therapy. In all the preclinical studies reviewed, image-guided cell therapy led to greater survival benefit and, therefore, has the potential to be translated into techniques in glioma treatment.
Author Contributions
For research articles with several authors, a short paragraph specifying their individual contributions must be provided. The following statements should be used “Conceptualization, S.S. and B.L.; methodology, S.S.; software, S.S.; validation, S.S., and B.L.; formal analysis, S.S.; investigation, S.S.; resources, S.S.; data curation, S.S.; writing—original draft preparation, S.S.; writing—review and editing, S.S.; visualization, S.S.; supervision, B.L.; project administration, S.S.; funding acquisition, S.S. All authors have read and agreed to the published version of the manuscript.” Please turn to the CRediT taxonomy for the term explanation. Authorship must be limited to those who have contributed substantially to the work reported.
Funding
This research received no external funding
Institutional Review Board Statement
Not applicable
Informed Consent Statement
Not applicable
Data Availability Statement
We encourage all authors of articles published in MDPI journals to share their research data. In this section, please provide details regarding where data supporting reported results can be found, including links to publicly archived datasets analyzed or generated during the study. Where no new data were created, or where data is unavailable due to privacy or ethical restrictions, a statement is still required. Suggested Data Availability Statements are available in section “MDPI Research Data Policies” at https://www.mdpi.com/ethics.
Conflicts of Interest
The authors declare no conflicts of interest
References
- Shah, S. Novel Therapies in Glioblastoma Treatment: Review of Glioblastoma; Current Treatment Options; and Novel Oncolytic Viral Therapies. Med Sci (Basel). 2023, 12, 1. [Google Scholar] [CrossRef]
- Schaff, L.R.; Mellinghoff, I.K. Glioblastoma and Other Primary Brain Malignancies in Adults: A Review. JAMA. 2023, 329, 574–587. [Google Scholar] [CrossRef] [PubMed]
- Czarnywojtek, A.; Borowska, M.; Dyrka, K.; et al. Glioblastoma Multiforme: The Latest Diagnostics and Treatment Techniques. Pharmacology. 2023, 108, 423–431. [Google Scholar] [CrossRef]
- Le Rhun, E.; Preusser, M.; Roth, P.; et al. Molecular targeted therapy of glioblastoma. Cancer Treat Rev. 2019, 80, 101896. [Google Scholar] [CrossRef]
- Shah, S.; Mansour, H.M.; Aguilar, T.M.; Lucke-Wold, B. Advances in Anti-Cancer Drug Development: Metformin as Anti-Angiogenic Supplemental Treatment for Glioblastoma. Int J Mol Sci. 2024, 25, 5694. [Google Scholar] [CrossRef]
- Ohgaki, H.; Kleihues, P. The definition of primary and secondary glioblastoma. Clin Cancer Res. 2013, 19, 764–772. [Google Scholar] [CrossRef] [PubMed]
- Hamad, A.; Yusubalieva, G.M.; Baklaushev, V.P.; Chumakov, P.M.; Lipatova, A.V. Recent Developments in Glioblastoma Therapy: Oncolytic Viruses and Emerging Future Strategies. Viruses. 2023, 15, 547. [Google Scholar] [CrossRef]
- Oh, J.M.; Ahn, B.C. Molecular mechanisms of radioactive iodine refractoriness in differentiated thyroid cancer: Impaired sodium iodide symporter (NIS) expression owing to altered signaling pathway activity and intracellular localization of NIS. Theranostics. 2021, 11, 6251–6277. [Google Scholar] [CrossRef]
- Gong, Z.; Wei, M.; Vlantis, AC.; et al. Sodium-iodide symporter and its related solute carriers in thyroid cancer. J Endocrinol. 2024, 261, e230373. [Google Scholar] [CrossRef] [PubMed]
- Spitzweg, C.; Nelson, PJ.; Wagner, E.; et al. The sodium iodide symporter (NIS): novel applications for radionuclide imaging and treatment. Endocr Relat Cancer. 2021, 28, T193–T213. [Google Scholar] [CrossRef]
- Darrouzet, E.; Lindenthal, S.; Marcellin, D.; Pellequer, J.L.; Pourcher, T. The sodium/iodide symporter: state of the art of its molecular characterization. Biochim Biophys Acta. 2014, 1838 Pt B, 244–253. [Google Scholar] [CrossRef]
- Ding, D.C.; Shyu, W.C.; Lin, S.Z. Mesenchymal stem cells. Cell Transplant. 2011, 20, 5–14. [Google Scholar] [CrossRef] [PubMed]
- Lan, T.; Luo, M.; Wei, X. Mesenchymal stem/stromal cells in cancer therapy. J Hematol Oncol. 2021, 14, 195. [Google Scholar] [CrossRef]
- Mazini, L.; Rochette, L.; Admou, B.; Amal, S.; Malka, G. Hopes and Limits of Adipose-Derived Stem Cells (ADSCs) and Mesenchymal Stem Cells (MSCs) in Wound Healing. Int J Mol Sci. 2020, 21, 1306. [Google Scholar] [CrossRef]
- Naji, A.; Eitoku, M.; Favier, B.; Deschaseaux, F.; Rouas-Freiss, N.; Suganuma, N. Biological functions of mesenchymal stem cells and clinical implications. Cell Mol Life Sci. 2019, 76, 3323–3348. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Huang, Y.; Yang, L. Research progress in the use of mesenchymal stem cells and their derived exosomes in the treatment of osteoarthritis. Ageing Res Rev. 2022, 80, 101684. [Google Scholar] [CrossRef] [PubMed]
- Herzog, E.L.; Chai, L.; Krause, D.S. Plasticity of marrow-derived stem cells. Blood 2003, 102, 3483–3493. [Google Scholar] [CrossRef]
- Angelopoulou, M.; Novelli, E.; Grove, J.E.; et al. Cotransplantation of human mesenchymal stem cells enhances human myelopoiesis and megakaryocytopoiesis in NOD/SCID mice. Experimental Hematology 2003, 31, 413–420. [Google Scholar] [CrossRef] [PubMed]
- Matthay, M.A.; Goolaerts, A.; Howard, J.P.; Lee, J.W. Mesenchymal stem cells for acute lung injury: preclinical evidence. Crit Care Med. 2010, 38, S569–S573. [Google Scholar] [CrossRef]
- Yao, P.; Zhou, L.; Zhu, L.; Zhou, B.; Yu, Q. Mesenchymal Stem Cells: A Potential Therapeutic Strategy for Neurodegenerative Diseases. Eur Neurol. 2020, 83, 235–241. [Google Scholar] [CrossRef]
- Hua, Q.; Zhang, Y.; Li, H.; et al. Human umbilical cord blood-derived MSCs trans-differentiate into endometrial cells and regulate Th17/Treg balance through NF-κB signaling in rabbit intrauterine adhesions endometrium. Stem Cell Res Ther. 2022, 13, 301. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Wang, B.; Zhu, X.; et al. Human umbilical cord blood-derived MSCs exosome attenuate myocardial injury by inhibiting ferroptosis in acute myocardial infarction mice. Cell Biol Toxicol. 2021, 37, 51–64. [Google Scholar] [CrossRef]
- Fu, X.; Liu, G.; Halim, A.; Ju, Y.; Luo, Q.; Song, A.G. Mesenchymal Stem Cell Migration and Tissue Repair. Cells. 2019, 8, 784. [Google Scholar] [CrossRef] [PubMed]
- Friedenstein, A.J.; Chailakhyan, R.K.; Gerasimov, U.V. Bone marrow osteogenic stem cells: in vitro cultivation and transplantation in diffusion chambers. Cell Tissue Kinet. 1987, 20, 263–272. [Google Scholar] [CrossRef] [PubMed]
- Galipeau, J.; Sensébé, L. Mesenchymal Stromal Cells: Clinical Challenges and Therapeutic Opportunities. Cell Stem Cell. 2018, 22, 824–833. [Google Scholar] [CrossRef] [PubMed]
- Keshtkar, S.; Azarpira, N.; Ghahremani, M.H. Mesenchymal stem cell-derived extracellular vesicles: novel frontiers in regenerative medicine. Stem Cell Res Ther. 2018, 9, 63. [Google Scholar] [CrossRef] [PubMed]
- Ding, D.C.; Chang, Y.H.; Shyu, W.C.; Lin, S.Z. Human umbilical cord mesenchymal stem cells: a new era for stem cell therapy. Cell Transplant. 2015, 24, 339–347. [Google Scholar] [CrossRef] [PubMed]
- Samsonraj, R.M.; Raghunath, M.; Nurcombe, V.; Hui, J.H.; van Wijnen, A.J.; Cool, S.M. Concise Review: Multifaceted Characterization of Human Mesenchymal Stem Cells for Use in Regenerative Medicine. Stem Cells Transl Med. 2017, 6, 2173–2185. [Google Scholar] [CrossRef] [PubMed]
- Ankrum, J.A.; Ong, J.F.; Karp, J.M. Mesenchymal stem cells: immune evasive, not immune privileged. Nat Biotechnol. 2014, 32, 252–260. [Google Scholar] [CrossRef]
- Caplan, A.I. Mesenchymal Stem Cells: Time to Change the Name! . Stem Cells Transl Med. 2017, 6, 1445–1451. [Google Scholar] [CrossRef]
- Dzobo, K. Recent Trends in Multipotent Human Mesenchymal Stem/Stromal Cells: Learning from History and Advancing Clinical Applications. OMICS. 2021, 25, 342–357. [Google Scholar] [CrossRef] [PubMed]
- Al-Ghadban, S.; Bunnell, B.A. Adipose Tissue-Derived Stem Cells: Immunomodulatory Effects and Therapeutic Potential. Physiology (Bethesda). 2020, 35, 125–133. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Xia, M.; Gao, Y.; Chen, Y.; Xu, Y. Human umbilical cord mesenchymal stem cells: an overview of their potential in cell-based therapy. Expert Opin Biol Ther. 2015, 15, 1293–1306. [Google Scholar] [CrossRef] [PubMed]
- Zupan, J. Human Synovium-Derived Mesenchymal Stem Cells: Ex Vivo Analysis. Methods Mol Biol. 2019, 2045, 145–154. [Google Scholar] [CrossRef] [PubMed]
- Okolicsanyi, RK.; Camilleri, ET.; Oikari, LE.; et al. Human Mesenchymal Stem Cells Retain Multilineage Differentiation Capacity Including Neural Marker Expression after Extended In Vitro Expansion. PLoS One. 2015, 10, e0137255. [Google Scholar] [CrossRef] [PubMed]
- Balakrishna Pillai, A.; Mariappan, V.; JeanPierre, A.R.; Rao, S.R. Restoration of vascular endothelial integrity by mesenchymal stromal/stem cells in debilitating virus diseases. Hum Cell. 2022, 35, 1633–1639. [Google Scholar] [CrossRef] [PubMed]
- Dapkute, D.; Steponkiene, S.; Bulotiene, D.; Saulite, L.; Riekstina, U.; Rotomskis, R. Skin-derived mesenchymal stem cells as quantum dot vehicles to tumors. Int J Nanomedicine. 2017, 12, 8129–8142. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Feng, Y.; Xie, X.; et al. Engineered Mesenchymal Stem Cells as a Biotherapy Platform for Targeted Photodynamic Immunotherapy of Breast Cancer. Adv Healthc Mater. 2022, 11, e2101375. [Google Scholar] [CrossRef]
- Sadhukha, T.; O’Brien, T.D.; Prabha, S. Nano-engineered mesenchymal stem cells as targeted therapeutic carriers. J Control Release. 2014, 196, 243–251. [Google Scholar] [CrossRef] [PubMed]
- Xiao, J.; Zeng, L.; Ding, S.; et al. Tumor-Tropic Adipose-Derived Mesenchymal Stromal Cell Mediated Bi2 Se3 Nano-Radiosensitizers Delivery for Targeted Radiotherapy of Non-Small Cell Lung Cancer. Adv Healthc Mater. 2022, 11, e2200143. [Google Scholar] [CrossRef]
- Senst, C.; Nazari-Shafti, T.; Kruger, S.; et al. Prospective dual role of mesenchymal stem cells in breast tumor microenvironment. Breast Cancer Res Treat. 2013, 137, 69–79. [Google Scholar] [CrossRef]
- Golinelli, G.; Talami, R.; Frabetti, S.; et al. A 3D Platform to Investigate Dynamic Cell-to-Cell Interactions Between Tumor Cells and Mesenchymal Progenitors. Front Cell Dev Biol. 2022, 9, 767253. [Google Scholar] [CrossRef]
- Mishra, VK.; Shih, HH.; Parveen, F.; et al. Identifying the Therapeutic Significance of Mesenchymal Stem Cells. Cells. 2020, 9, 1145. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Chen, S.; Yang, P.; Cao, H.; Li, L. The role of mesenchymal stem cells in hematopoietic stem cell transplantation: prevention and treatment of graft-versus-host disease. Stem Cell Res Ther. 2019, 10, 182. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.; Huang, L.; Li, Y.; et al. Mesenchymal Stem Cells and Cancer: Clinical Challenges and Opportunities. Biomed Res Int. 2019, 2019, 2820853. [Google Scholar] [CrossRef] [PubMed]
- Munir, H.; Ward, L.S.C.; McGettrick, H.M. Mesenchymal Stem Cells as Endogenous Regulators of Inflammation. Adv Exp Med Biol. 2018, 1060, 73–98. [Google Scholar] [CrossRef] [PubMed]
- Ji, K.; Ding, L.; Chen, X.; et al. Mesenchymal Stem Cells Differentiation: Mitochondria Matter in Osteogenesis or Adipogenesis Direction. Curr Stem Cell Res Ther. 2020, 15, 602–606. [Google Scholar] [CrossRef]
- Miao, J.; Ren, Z.; Zhong, Z.; et al. Mesenchymal Stem Cells: Potential Therapeutic Prospect of Paracrine Pathways in Neonatal Infection. J Interferon Cytokine Res. 2021, 41, 365–374. [Google Scholar] [CrossRef]
- Selmi-Ruby, S.; Watrin, C.; Trouttet-Masson, S.; et al. The porcine sodium/iodide symporter gene exhibits an uncommon expression pattern related to the use of alternative splice sites not present in the human or murine species. Endocrinology. 2003, 144, 1074–1085. [Google Scholar] [CrossRef]
- Knoop, K.; Kolokythas, M.; Klutz, K.; et al. Image-guided, tumor stroma-targeted 131I therapy of hepatocellular cancer after systemic mesenchymal stem cell-mediated NIS gene delivery. Mol Ther. 2011, 19, 1704–1713. [Google Scholar] [CrossRef]
- Cho, J.Y. A transporter gene (sodium iodide symporter) for dual purposes in gene therapy: imaging and therapy. Curr Gene Ther. 2002, 2, 393–402. [Google Scholar] [CrossRef] [PubMed]
- Faivre, J.; Clerc, J.; Gérolami, R.; et al. Long-term radioiodine retention and regression of liver cancer after sodium iodide symporter gene transfer in wistar rats [published correction appears in Cancer Res. 2004 Dec 15;64, 9230]. Cancer Res. 2004, 64, 8045–8051. [Google Scholar] [CrossRef] [PubMed]
- Belmar-López, C.; Vassaux, G.; Medel-Martinez, A.; et al. Mesenchymal Stem Cells Delivery in Individuals with Different Pathologies: Multimodal Tracking, Safety and Future Applications. Int J Mol Sci. 2022, 23, 1682. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Liu, Y.Y.; Jhiang, S.M. Cell surface targeting accounts for the difference in iodide uptake activity between human Na+/I- symporter and rat Na+/I- symporter. J Clin Endocrinol Metab. 2005, 90, 6131–6140. [Google Scholar] [CrossRef] [PubMed]
- Yao, X.; Li, J.; Liu, J.; Liu, K. An Arabidopsis mitochondria-localized RRL protein mediates abscisic acid signal transduction through mitochondrial retrograde regulation involving ABI4. J Exp Bot. 2015, 66, 6431–6445. [Google Scholar] [CrossRef]
- Nowakowski, A.; Walczak, P.; Lukomska, B.; Janowski, M. Genetic Engineering of Mesenchymal Stem Cells to Induce Their Migration and Survival. Stem Cells Int. 2016, 2016, 4956063. [Google Scholar] [CrossRef] [PubMed]
- Gugjoo, M.B.; Sakeena, Q.; Wani, M.Y.; Abdel-Baset Ismail, A.; Ahmad, S.M.; Shah, R.A. Mesenchymal stem cells: A promising antimicrobial therapy in veterinary medicine. Microb Pathog. 2023, 182, 106234. [Google Scholar] [CrossRef] [PubMed]
- Siristatidis, C.; Vogiatzi, P.; Salamalekis, G.; et al. Granulocyte macrophage colony stimulating factor supplementation in culture media for subfertile women undergoing assisted reproduction technologies: a systematic review. Int J Endocrinol. 2013, 2013, 704967. [Google Scholar] [CrossRef] [PubMed]
- Bonaventura, G.; Incontro, S.; Iemmolo, R.; et al. Dental mesenchymal stem cells and neuro-regeneration: a focus on spinal cord injury. Cell Tissue Res. 2020, 379, 421–428. [Google Scholar] [CrossRef] [PubMed]
- Etchebehere, EC.; Cendes, F.; Lopes-Cendes, I.; et al. Brain single-photon emission computed tomography and magnetic resonance imaging in Machado-Joseph disease. Arch Neurol. 2001, 58, 1257–1263. [Google Scholar] [CrossRef]
- Hade, M.D.; Suire, C.N.; Suo, Z. Mesenchymal Stem Cell-Derived Exosomes: Applications in Regenerative Medicine. Cells. 2021, 10, 1959. [Google Scholar] [CrossRef] [PubMed]
- Nose, N.; Nogami, S.; Koshino, K.; et al. [18F]FDG-labelled stem cell PET imaging in different route of administrations and multiple animal species. Sci Rep. 2021, 11, 10896. [Google Scholar] [CrossRef]
- Lu, CH.; Chen, YA.; Ke, CC.; et al. Preclinical Characterization and In Vivo Imaging of 111In-Labeled Mesenchymal Stem Cell-Derived Extracellular Vesicles. Mol Imaging Biol. 2021, 23, 361–371. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Goh, E.L.K.; He, J.; et al. Emerging Intrinsic Therapeutic Targets for Metastatic Breast Cancer. Biology (Basel). 2023, 12, 697. [Google Scholar] [CrossRef] [PubMed]
- Zakrzewski, W.; Dobrzyński, M.; Szymonowicz, M.; Rybak, Z. Stem cells: past, present, and future. Stem Cell Res Ther. 2019, 10, 68. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.L.; Huang, T.; Wu, B.L.; He, W.X.; Liu, D. Stem cells in cancer therapy: opportunities and challenges. Oncotarget. 2017, 8, 75756–75766. [Google Scholar] [CrossRef] [PubMed]
- Rivera-Cruz, C.M.; Shearer, J.J.; Figueiredo Neto, M.; Figueiredo, M.L. The Immunomodulatory Effects of Mesenchymal Stem Cell Polarization within the Tumor Microenvironment Niche. Stem Cells Int. 2017, 2017, 4015039. [Google Scholar] [CrossRef] [PubMed]
- Szewc, M.; Radzikowska-Bűchner, E.; Wdowiak, P.; et al. MSCs as Tumor-Specific Vectors for the Delivery of Anticancer Agents-A Potential Therapeutic Strategy in Cancer Diseases: Perspectives for Quinazoline Derivatives. Int J Mol Sci. 2022, 23, 2745. [Google Scholar] [CrossRef]
- Attia, N.; Mashal, M.; Puras, G.; Pedraz, J.L. Mesenchymal Stem Cells as a Gene Delivery Tool: Promise, Problems, and Prospects. Pharmaceutics. 2021, 13, 843. [Google Scholar] [CrossRef]
- Cuiffo, B.G.; Karnoub, A.E. Mesenchymal stem cells in tumor development: emerging roles and concepts. Cell Adh Migr. 2012, 6, 220–230. [Google Scholar] [CrossRef]
- Rabha, B.; Bharadwaj, K.K.; Pati, S.; Choudhury, B.K.; Sarkar, T.; Kari, Z.A.; Edinur, H.A.; Baishya, D.; Atanase, L.I. Development of Polymer-Based Nanoformulations for Glioblastoma Brain Cancer Therapy and Diagnosis: An Update. Polymers. 2021, 13, 4114. [Google Scholar] [CrossRef] [PubMed]
- Yang, CT.; Lai, R.C.; Phua, V.J.X.; et al. Standard Radio-Iodine Labeling Protocols Impaired the Functional Integrity of Mesenchymal Stem/Stromal Cell Exosomes. Int J Mol Sci. 2024, 25, 3742. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Han, Y.; Zhao, G.; et al. Hypoxia-Responsive Lipid-Polymer Nanoparticle-Combined Imaging-Guided Surgery and Multitherapy Strategies for Glioma. ACS Appl Mater Interfaces. 2020, 12, 52319–52328. [Google Scholar] [CrossRef] [PubMed]
- Schug, C.; Gupta, A.; Urnauer, S.; et al. A Novel Approach for Image-Guided 131I Therapy of Pancreatic Ductal Adenocarcinoma Using Mesenchymal Stem Cell-Mediated NIS Gene Delivery. Mol Cancer Res. 2019, 17, 310–320. [Google Scholar] [CrossRef] [PubMed]
- Nicola, J.P.; Basquin, C.; Portulano, C.; Reyna-Neyra, A.; Paroder, M.; Carrasco, N. The Na+/I- symporter mediates active iodide uptake in the intestine. Am J Physiol Cell Physiol. 2009, 296, C654–C662. [Google Scholar] [CrossRef] [PubMed]
- National Research Council (US) and Institute of Medicine (US) Committee on the Mathematics and Physics of Emerging Dynamic Biomedical Imaging. Mathematics and Physics of Emerging Biomedical Imaging. Washington (DC): National Academies Press (US); 1996. Chapter 5, Single Photon Emission Computed Tomography. Available from: https://www.ncbi.nlm.nih.gov/books/NBK232492/.
- Penheiter, A.R.; Russell, S.J.; Carlson, S.K. The sodium iodide symporter (NIS) as an imaging reporter for gene, viral, and cell-based therapies. Curr Gene Ther. 2012, 12, 33–47. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; DeGrado, TR. [18F]Tetrafluoroborate ([18F]TFB) and its analogs for PET imaging of the sodium/iodide symporter. Theranostics. 2018, 8, 3918–3931. [Google Scholar] [CrossRef]
- Hanif, F.; Muzaffar, K.; Perveen, K.; Malhi, S.M.; Simjee, S.h.U. Glioblastoma Multiforme: A Review of its Epidemiology and Pathogenesis through Clinical Presentation and Treatment. Asian Pac J Cancer Prev. 2017, 18, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Gudkov, S.V.; Shilyagina, N.Y.; Vodeneev, V.A.; Zvyagin, A.V. Targeted Radionuclide Therapy of Human Tumors. Int J Mol Sci. 2015, 17, 33. [Google Scholar] [CrossRef]
- Shi, S.; Zhang, M.; Guo, R.; Miao, Y.; Li, B. Bone Marrow-Derived Mesenchymal Stem Cell-Mediated Dual-Gene Therapy for Glioblastoma. Hum Gene Ther. 2019, 30, 106–117. [Google Scholar] [CrossRef]
- Czarnywojtek, A.; Gut, P.; Borowska, M.; Dyrka, K.; Ruchała, M.; Ferlito, A. A NEW HYPOTHESIS IN THE TREATMENT OF RECURRENT GLIOBLASTOMA MULTIFORME (GBM). PART 1: INTRODUCTION. Pol Merkur Lekarski. 2023, 51, 430–432. [Google Scholar] [CrossRef] [PubMed]
- Czarnywojtek, A.; Gut, P.; Sowiński, J.; Ruchała, M.; Ferlito, A.; Dyrka, K. A NEW HYPOTHESIS IN THE TREATMENT OF RECURRENT GLIOBLASTOMA MULTIFORME (GBM). PART 2: IS THERE AN ALTERNATIVE THERAPY OPTION IN RECURRENT GM WHEN ALL STANDARD TREATMENTS HAVE BEEN EXHAUSTED? Pol Merkur Lekarski. 2023, 51, 433–435. [Google Scholar] [CrossRef] [PubMed]
- Yalamarty, S.S.K.; Filipczak, N.; Li, X.; et al. Mechanisms of Resistance and Current Treatment Options for Glioblastoma Multiforme (GBM). Cancers (Basel) 2023, 15, 2116. [Google Scholar] [CrossRef]
- Cho, JY.; Shen, DH.; Yang, W.; Williams, B.; Buckwalter, TL.; La Perle, KM.; et al. In vivo imaging and radioiodine therapy following sodium iodide symporter gene transfer in animal model of intracerebral gliomas. Gene Ther. 2002, 9, 1139–1145. [Google Scholar] [CrossRef]
- Guo, R.; Xi, Y.; Zhang, M.; Miao, Y.; Zhang, M.; Li, B. Human sodium iodide transporter gene-mediated imaging and therapy of mouse glioma, comparison between (188)Re and (131)I. Oncol Lett. 2018, 15, 3911–3917. [Google Scholar]
- Opyrchal, M.; Allen, C.; Iankov, I.; Aderca, I.; Schroeder, M.; Sarkaria, J.; et al. Effective radiovirotherapy for malignant gliomas by using oncolytic measles virus strains encoding the sodium iodide symporter (MV-NIS). Hum Gene Ther. 2012, 23, 419–427. [Google Scholar] [CrossRef] [PubMed]
- Kitzberger, C.; Shehzad, K.; Morath, V.; et al. Interleukin-6-controlled, mesenchymal stem cell-based sodium/iodide symporter gene therapy improves survival of glioblastoma-bearing mice. Mol Ther Oncolytics. 2023, 30, 238–253. [Google Scholar] [CrossRef]
- Spellerberg, R.; Benli-Hoppe, T.; Kitzberger, C.; et al. Dual EGFR- and TfR-targeted gene transfer for sodium iodide symporter gene therapy of glioblastoma. Mol Ther Oncolytics 2022, 27, 272–287. [Google Scholar] [CrossRef]
- Fu, Y.; Ong, LC.; Ranganath, SH.; et al. A Dual Tracer 18F-FCH/18F-FDG PET Imaging of an Orthotopic Brain Tumor Xenograft Model. PLoS One. 2016, 11, e0148123. [Google Scholar] [CrossRef]
Figure 1.
NIS gene delivery strategies to glioblastoma.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



