Submitted:
06 July 2024
Posted:
09 July 2024
You are already at the latest version
Abstract
Mango industry played important roles in the economy of tropical and subtropical regions in China. Embryo abortion, flesh rotten, pericarp stain and so on always result in heavy loss. Transgenic breeding is a potential way for solving these problems. However, dependable protocol for mango transgene has not been reported until now. In this research, we found that after mango embryogenic callus were infected by Agrobacterium, most callus necrotized and died. No seedling was differentiated from the survived callus. Contents of 106 compounds in mango callus infected by Agrobacterium were found significantly different from those in control. The top 7 up-regulated compounds were gallic acid, phosphoenolpyruvic acid, dihydrotricetin, ubiquinone, dihydrokaempferol, D-erythrose-4-phosphate and cyanidin. The expressions of ROS-related genes, gallic acid synthesis genes, and defense related genes in callus infected by Agrobacterium were significantly higher than those in control. The growth of Agrobacterium on culturing plate pipetting gallic acid was inhibited. Mango callus infected by Agrobacterium transformed with type III secretion system gene cluster (T3SS) and effector gene AvrPto isolated from Pseudomonas syringae pv. tomato strain DC3000 brown and necrotize slower than those infected by Agrobacterium transformed with net vector (NV). The contents of ROS in callus in-fected by Agrobacterium (T3SS-AvrPto) were less than those in callus infected by Agrobacterium (NV). The expressions of genes participated in immune responses in mango callus infected by Agrobacterium (T3SS-AvrPto) were also less than those in mango callus infected by Agrobacterium (NV). The transformation rate of mango callus infected with Agrobacterium (T3SS-AvrPto-gfp) was 1.6 %. No callus infected with Agrobacterium (NV-gfp) was transformed successfully. The efficiency of mango transformation can be improved by constructing Agrobacterium strains harboring type III secretion system gene cluster and its effector gene.
Keywords:
Mango callus
; Agrobacterium(T3SS) gene
; Reactive oxygen species(ROS)gene
; Gallic acid synthesis gene
1. Introduction
Mango (Mangifera indica L.) is the second largest tropical fruit in yield in world annually (Office of Tropical Crops in Southern Asian, Ministry of Agriculture, China). China is the second largest country for planting mango (Office of Tropical Crops in Southern Asian, Ministry of Agriculture, China). During mango planting, embryo abortion, flesh rotten, pericarp stain and so on always result in heavy loss [1,2,3,4,5]. Cultivating new varieties with various resistances can resolve these problems.
The conventional method for getting new mango varieties is cross breeding. A complete cycle of cross breeding for mango always takes about 20 years [6]. Transgene can solve problems above efficiently in short time and breeding efficiency can be improved significantly [7,8]. Pandey et al. (2022) reported that mango transgenic plants can be gotten by infecting apex meristematic tissue with Agrobacterium harboring target gene [9]. Two-week-old mango seedlings with emerging plumules were punctured 15-20 times with an insulin syringe at the apical meristem and incubated in AB minimal medium [9]. Apical meristem was infected with Agrobacterium transformed with vectors GFP::hptII in pCAMBIA1302 and GUS::nptII in pCAMBIA2301 individually [9]. 65.0% of GFP and 57.14% of GUS plants were transformed successfully [9]. Primary transformants demonstrated transgene integration at genomic and transcript level in some of the plants [9].
Adjei et al. (2023) optimized the conditions of the protoplast isolation and transformation system for mango [10]. GFP florescence was seen in the transformed protoplasts [10]. Target gene was successfully induced into the mango protoplast and that it has been transcribed and translated [10].
Stem apex meristematic tissue is highly differentiated tissues. When stem apex meristematic tissues were transformed, some cells were transformed and some failed. That always resulted in chimera [11,12]. When cell divided, transformation-failed cells always stand ahead of the competition and the divisions of the transformed cells stopped [13,14]. Foreign gene lost and experiment failed [13,14]. Experiment that Adjei et al. (2023) did was a transient expression and did not get transformed seedling [10]. Dependable method for mango transgene has not been reported until now.
Plant embryogenic callus is a loose and crisp tissue. When embryogenic callus was infected by Agrobacterium, plant cells had high chances being infected and transformed. Infecting callus using Agrobacterium harboring target gene is a conventional method for constructing transgene protocol [15,16,17]. However, in this research, we found that most mango callus necrotized and died after they were infected by Agrobacterium. No seedling was differentiated from the survived callus.
When infecting plant cells, Agrobacterium tumefaciens sends virulence proteins into host cells through the type IV secretion system (T4SS) [18]. Active plant defense relying on innate immune responses was triggered [19]. Some gram-negative plant pathogenic bacteria, such as Pseudomonas syringae pv. tomato, deliver bacterial proteins into eukaryotic hosts through type III secretion system (T3SS) [20,21]. They can suppress plant basal defense [20,21].
After being infected by Agrobacterium, mango callus brown in short time. Excessive ROS can injure and kill plant cells, resulting in callus browning and necrosis [22]. Excessive ROS is an indicator of immune response [23]. We speculated that immune responses were activated in mango callus cells when they were infected by Agrobacterium. Constructing Agrobacterium strain harboring T3SS and its effector gene might inhibit the responses and transgene efficiency of mango callus will be improved. To verify this speculation, key substances and genes participating in immune responses of mango callus against Agrobacterium were identified in this research. Agrobacterium harboring T3SS and its effector gene AvrPto isolated from Pseudomonas syringae pv. tomato strain DC3000 was constructed. Mango callus infected by Agrobacterium (Type3-AvrPto) brown and necrotize remarkably slower than those infected by Agrobacterium harboring net vector (NV). The transformation rate of mango callus infected with Agrobacterium (Type3-AvrPto) was about 1.6 %. No transformed callus was found in those infected with wild Agrobacterium or Agrobacterium (NV). Mango callus had less immune response against Agrobacterium (Type3-AvrPto) than against Agrobacterium (NV) or wild Agrobacterium. The efficiency of mango callus transformation can be improved by constructing Agrobacterium strains harboring T3SS and its effector gene.
2. Results
2.1. Phenotypic Characteristics of Mango Callus Infected by Agrobacterium
When callus was infected with Agrobacterium, most of them brown and necrotize within 90 hours. At the first 15 hours, callus browning and necrosis have not been observed. Since the second 15 hours, callus begin to brown. In the 3rd 15 hours, callus brown very fast. The climax of browning and necrosis happened at the 4th 15 hour. After callus was infected with Agrobacterium for 75 hours, most of them necrotized. Transferring callus onto fresh medium cannot alleviate browning and necrosis
2.2. Mango Callus Infected by Agrobacterium Contained More Reactive Oxygen Species (ROS) than Control
Excessive ROS injured and killed callus cells, resulting in callus browning and necrosis [22]. To study whether browning and necrosis of mango callus infected by Agrobacterium was due to ROS, ROS content in callus was measured. Results showed that hydrogen peroxide (H2O2) and superoxide anion (O2-) contents in mango callus infected were much higher than those in control. After callus was infected 15 hours, H2O2 contents in mango callus were similar. After that, H2O2 content in mango callus increased fast. After being infected 45 hours, H2O2 content reached to the maximum. After that, H2O2 content in callus decreased. The trend of O2- content in mango callus infected by Agrobacterium showed similar. The contents of H2O2, O2- and •OH were stable in callus uninfected at different time (data not shown). These demonstrated that after callus was infected by Agrobacterium, ROS contents in callus increased rapidly. The excessive ROS may kill the callus cells and caused browning and necrosis.
2.3. Gallic Acid Was up Regulated after Mango Callus Was Infected
When plants were infected by pathogen, innate immune responses in plants will be triggered [19]. Partial plant cells near the wound died and blocked pathogen cells growth [24,25,26]. To identify whether the browning and necrosis of mango callus infected was caused by immune responses, metabolome experiment was performed using mango callus infected by Agrobacterium for 45 hours and callus uninfected. 516 metabolites were detected in the samples. Contents of 106 compounds in callus infected by Agrobacterium were found significantly different from those in callus uninfected. Among them, 68 compounds were down-regulated and 38 were up-regulated. The top 7 up-regulated compounds were gallic acid, phosphoenolpyruvic acid, dihydrotricetin, ubiquinone, dihydrokaempferol, D-erythrose-4-phosphate and cyanidin respectively (Table 1). These compounds might have participated in immune responses in mango callus infected.
2.4. The Expressions of ROS-Related Genes and Gallic Acid Synthesis Genes Were up Regulated in Callus Infected by Agrobacterium
To further validate whether immune responses happened in mango callus infected by Agrobacterium transcriptome experiments were done using mango callus infected by Agrobacterium for 30 hours and that uninfected. Results showed that 1256 genes were up-regulated and 1528 were down-regulated using the criteria of |log2FC|>1 and FDR <0.05. Pathway was analyzed using Kyoto Encyclopedia of Genes and Genomes (KEGG) database. Up regulated genes were mainly grouped in phenylpropanoid biosynthesis, phenylalanine metabolism, and antioxidant biosynthesis. Down-regulated genes were significantly enriched in photosynthesis, nucleic acid metabolism, DNA replication, photosynthesis, base excision repair. The expressions of ROS-related genes, gallic acid synthesis genes, and defense related genes in callus infected by Agrobacterium were significantly higher than those in callus uninfected, indicating that immune responses in callus indeed have been activated by Agrobacterium cells (Table 2). Gallic acid synthesis genes participated in immune responses in mango callus infected.
2.5. Verification the Expressions of Genes Participating in Immune Responses in Mango Callus Infected by Agrobacterium
To verify the transcriptome experiments above, qRT-PCR was performed for the top 21 genes upregulated. Results showed that the expressions of these genes in callus infected by Agrobacterium measured through qRT-PCR were highly consistent with those through transcriptome (Table 3). These demonstrated that ROS related genes, gallic acid synthesis genes, and defense related genes were indeed the key genes participating in immune responses in mango callus infected by Agrobacterium. Knock out one or several of these genes or down regulation them may inhibit the immune responses in mango callus. The efficiencies of callus infection and transgene will be enhanced through this way.
2.6. Gallic Acid Can Inhibit the Growth of Agrobacterium
Both metabolome and transcriptome experiments showed that gallic acid participated in immune responses in mango callus infected by Agrobacterium. To further verify this conclusion, Agrobacterium was cultured on MS plate and gallic acid was pipetted in the center. Results showed that Agrobacterium did not grow in the plate containing gallic acid (Figure 4). With being cultured more time, more Agrobacterium cells accumulated in the plate without gallic acid (Figure 4). But they did not grow in the field containing gallic acid (Figure 4). These demonstrated that gallic acid can inhibit the growth of Agrobacterium. Failure of mango transgene might partially be due to gallic acid produced in callus infected by Agrobacterium.
2.7. Agrobacterium Harboring Type III Secretion Gene Cluster (T3SS) and Effector Gene AvrPto Had Higher Infection Efficiency than Control Agrobacterium
When Agrobacterium infected plant cells, effector proteins were expelled through secretion system [27]. The effector proteins adhered to plant cells and triggered immune responses [19,28]. The secretion system of Agrobacterium belongs to type IV [18]. Many plants had immune responses when infected by Agrobacterium [19]. The secretion system of Pseudomonas syringae pv. tomato belongs to type III [29]. Immune responses in wheat embryo or switchgrass callus infected by Agrobacterium transformed with T3SS and AvrPto isolated from Pseudomonas syringae pv. tomato strain DC3000 were significantly weaker than those in wheat embryo or switchgrass callus infected by control Agrobacterium [19]. Immune responses in mango callus might also be inhibited when Agrobacterium was replaced with Agrobacterium (T3SS-AvrPto). Transgene efficiency of mango callus may be enhanced through this way. To verify this speculation, T3SS and effector gene AvrPto isolated from Pseudomonas syringae pv. tomato strain DC3000 were transformed into Agrobacterium strain EHA105 and Agrobacterium (T3SS-AvrPto) was constructed. Mango callus was infected by Agrobacterium (Type3-AvrPto) transformed with gfp gene. Results showed that 1.6 % callus infected by Agrobacterium (Type3-AvrPto-gfp) had been transformed successfully (Figure 5). No callus infected with Agrobacterium (NV-gfp) showed GFP fluorescence. Agrobacterium harboring T3SS and AvrPto had higher infection efficiency than control Agrobacterium. Transgene efficiency of mango callus can be enhanced by constructing Agrobacterium strain harboring T3SS and AvrPto.
2.8. Mango Callus Infected by Agrobacterium (T3SS-AvrPto) Brown and Necrotize Slower than Those Infected by Control Agrobacterium
More callus infected by Agrobacterium (T3SS-AvrPto) was transformed than those infected by control Agrobacterium. This might be due to that less immune responses were triggered in callus infected by Agrobacterium (T3SS-AvrPto). To verify this speculation, the browning and necrosis of callus infected by Agrobacterium (T3SS-AvrPto) were observed. Results showed that callus infected by Agrobacterium (T3SS-AvrPto) brown and necrotize remarkably slower than those infected by Agrobacterium (NV) (Figure 6). Mango callus had less immune response against Agrobacterium (T3SS-AvrPto) than those against Agrobacterium (NV). Agrobacterium (T3SS-AvrPto) can escape being perceived by immune system in mango cells.
2.9. Mango Callus Infected by Agrobacterium (T3SS-AvrPto) Produced Less ROS than Those Infected by Agrobacterium (NV)
To identify whether mango callus infected by Agrobacterium (T3SS-AvrPto) brown and necrotize slower than Agrobacterium (NV) was due to that callus infected by Agrobacterium (T3SS-AvrPto) had weaker immune responses than those infected by Agrobacterium (NV), ROS in callus were determined. Results showed that ROS contents in mango callus infected by Agrobacterium (T3SS-AvrPto) was remarkably less than those in callus infected by Agrobacterium (NV) (Table 4). Mango callus infected by Agrobacterium (T3SS-AvrPto) did have weaker immune response than those infected by Agrobacterium (NV), which resulted that callus infected by Agrobacterium (T3SS-AvrPto) brown and necrotize slower than those infected by Agrobacterium (NV). That further demonstrated that more callus infected by Agrobacterium (T3SS-AvrPto) were transformed than those infected by Agrobacterium (NV) was due to that less immune responses were activated in callus infected by Agrobacterium (T3SS-AvrPto). Mango transgene efficiency was enhanced when callus was infected by Agrobacterium (T3SS-AvrPto).
2.10. QRT-PCR Analysis of the Genes Up-Regulated in Mango Callus Infected by Agrobacterium (T3SS-AvrPto)
To study the mechanism underlying immune responses activated in mango callus infected by Agrobacterium, the expressions of main genes up-regulated in immune responses in mango callus infected by Agrobacterium (T3SS-AvrPto) and those of them in callus infected by Agrobacterium (NV) were determined using QRT-PCR. Results showed that at each time point, the expressions of these genes in callus infected by Agrobacterium (T3SS-AvrPto) were less than those in callus infected by Agrobacterium (NV) (Table 5). This indicated that when callus was infected by Agrobacterium (T3SS-AvrPto), the expressions of the genes up regulated in immune responses were inhibited. Immune responses in callus infected by Agrobacterium (T3SS-AvrPto) were suppressed. This resulted that immune-responses activated in callus infected by Agrobacterium (T3SS-AvrPto) was weaker than those in callus infected by Agrobacterium (NV).
3. Discussion
Research found that DAHPS, DHQD, SHD participated in gallic acid biosynthesis [30]. We found that the expressions of these genes were up regulated after callus was infected by Agrobacterium. This might result that the synthesis of gallic acid were improved, as found in metabolome experiment in this research. Gallic acid is a common phenolic acid that is widely used as an antibacterial agent in the food industry [31]. It is recognized as a compound with broad-spectrum antibacterial and long-lasting sterilizing properties [32]. Our research showed that Agrobacterium did not grow on MS plate containing gallic acid. The growth of Agrobacterium cells can be inhibited by gallic acid. Failure of mango transgene might be partially due to that gallic acid produced in callus inhibited Agrobacterium growth.
Pattern-recognized receptors (PRRs) on plant cell membrane can perceive pathogens and activate immune system of plant cells [19,33]. Arabidopsis FLAGELIN-SENSITIVE2 (FLS2) was an identified PRR, which can perceive the 22 conserved amino acids in N terminal of bacterial flagellin (flg22) and interact with them [34,35]. After perceiving flg22, FLS2 combined with Brassinosteroid receptor-associated kinase 1 (BAK1) and formed complex, which activated the downstream mitogen-activated protein kinase 3 (MPK3) and mitogen-activated protein kinase 6 (MPK6) [36]. MPK3 and MPK6 induced the expressions of WRKY22, WRKY29, FRK1 (FLG22-induced receptor-like kinase 1) and other defense-related genes, which resulted that more ROS was produced [36]. High content of ROS induced plant cells necrotized and died [37]. Arabidopsis EF-Tu receptor (EFR) was also a PRR [38]. EFR can perceive Agrobacterium elongation factor Tu (EF-Tu) and activate plant immune response [39]. The polypeptide constituted by 18 amino acids located in N terminal of EF-Tu (elf18) can induce ROS being produced quickly, which induced the expressions of defense genes FRK1, NHL10 (NDR1/HIN1-like 10) and so on [40]. We found that the expressions of FLS2, MPK3, MPK6, WRKY22, NHL10 in mango callus infected by Agrobacterium were all higher than those in callus uninfected suggesting that when callus was infected by Agrobacterium, these proteins were synthesized rapidly, which further induced the expressions of the ROS-related genes. ROS was produced rapidly. Partial or all cells of callus were injured and killed by excessive ROS. Some infected callus brown and some necrotized (Figure 10). Gallic acid synthesis genes and defense-related genes together constituted the immune responses system in mango callus (Figure 6). This was different from that in Arabidopsis, which was only caused by defense-related genes. This might be the reason why the efficiency of Agrobacterium infection was low and mango transgene experiment was difficult.
The low efficiency of mango transgene was due to immune responses activated in callus. If the immune responses were suppressed, the infection and transformation efficiency might be improved significantly. The transformation efficiencies of both wheat embryos and switchgrass calli infected by Agrobacterium harboring T3SS and AvrPto isolated from Pseudomonas syringae pv. tomato strain DC3000 were significantly higher than those infected by Agrobacterium transformed with net vector [19]. We found that about 1.6 % callus infected with Agrobacterium (Type3-AvrPto-gfp) had been transformed successfully. No callus infected with Agrobacterium (NV-gfp) showed GFP fluorescence. Mango callus infected by Agrobacterium (T3SS-AvrPto) brown and necrotize remarkably slower than those infected by Agrobacterium (NV). ROS in mango callus infected by Agrobacterium (T3SS-AvrPto) was less than that in callus infected by Agrobacterium (NV). Less immune responses were activated in callus infected by Agrobacterium (T3SS-AvrPto) than those in callus infected by control Agrobacterium. Agrobacterium (T3SS-AvrPto) can escape being perceived by immune system in mango cells. Transgene efficiency of mango callus can be enhanced by constructing Agrobacterium strain transformed with T3SS and its effector gene.
4. Materials and Methods
4.1. Construction Agrobacterium Strains
DNA fragment containing P. syringae pv. tomato strain DC3000 T3SS gene cluster (GenBank accession number: AF232004.3), effector gene AvrPto (NC_004578.1) and its promoter (116 bp upstream of ATG) was synthesized with attB sequences at the two terminals. BP recombination reaction was done to insert the synthesized DNA into pENTRTM5’-TOPO (Invitrogen) using pENTRTM5’-TOPO Cloning Kit (Invitrogen). E. coli TOP10 was used to propagate the plasmid. Plasmids pENTRTM5’-TOPO-T3SS-AvrPto were extracted using GeneJET kit (Thermo Scientific). LR recombination reaction was performed to transfer T3SS-AvrPto in pENTRTM5’-TOPO-T3SS-AvrPto into pEarleyGate302 (BIOESN, Shanghai) using LR Clonase ™ Enzyme Mix (Invitrogen). E. coli TOP10 was used to propagate the plasmid. Plasmids pEarleyGate302-T3SS-AvrPto were extracted and transformed into Agrobacterium EHA105. Triparental mating [41] was used to mobilize pEarleyGate302-T3SS-AvrPto into A. tumefaciens strains. To construct Agrobacterium (T3SS-AvrPto-gfp), Agrobacterium (T3SS-AvrPto) was transformed with plasmid pBI221-GFP (HonorGene, Changsha, China) and the transformed Agrobacterium was screened on plate containing ampicillin.
4.2. Mango Embryogenic Callus Induction
Mango callus was induced according the published method and modified [42]. Fruitlet in cotyledon stage (45 days after flower) was collected. Contaminants on the surface were removed using brush. The fruit was rinsed for 30 minutes using tap water. It was transferred into 1% detergent and marinated for 20 minutes. The fruit was washed using flush water for 20 minutes. 75% ethanol was sprayed onto the surface. In the super clean bench, fruit was cut using scalpel and the seed was fetched. The seed was longitudinal cut. The embryonic matter was collected from the inner surface of the seed and cultured on medium M1 (B5 Mac elements, MS micro elements, 4% sucrose, 10% coconut water, 500 mg/L glutamine, 0.7% agar, 0.2% activated charcoal, 3.0 mg/L 2,4-D, 0.5 mg/L KT, 0.5 mg/L ZT, pH=5.8). The plates were put in dark 26 ℃ for one month. And then, the callus induced was transferred onto medium M2 (B5 Mac elements, MS micro elements, 4% sucrose, 10% coconut water, 500 mg/L glutamine, 0.7% agar, 0.2% activated charcoal, 1.0 mg/L 2,4-D, 0.5 mg/L KT, 0.5 mg/L GA3, pH=5.8). The plates were put in dark 26 ℃ and cultured for 1 month. Much embryogenic callus can be gotten.
4.3. Mango Callus Transformation
One milli gram of mango embryogenic callus was put on medium containing a series of hygromycin. The plates were put in dark and 25 ℃ for 1 month. The dose that callus cannot grow just was selected as the screening concentration. Agrobacterium cells transformed with pEarleyGate302-T3SS-AvrPto or pBI221-GFP was cultured overnight. The Agrobacterium culture (1.5 ml) was centrifugated at 500 rpm and the supernatant was discarded. The pellet was resuspended with AAM liquid medium containing 200 µmol/L acetosyringone and shaken at 100 rpm, 26 ℃ for 1 hour. Mango callus was put in AAM medium and cultured for 30 minutes. The liquid was discarded and the callus was put on sterilized filter paper in super clean bench for 60 minutes. And then, the callus was transferred onto medium M3 (B5 Mac elements, MS micro elements, 4% sucrose, 10% coconut water, 500 mg/L glutamine, 0.7% agar, 0.2% activated charcoal, 1.0 mg/L 2,4-D, 0.5 mg/L KT, 0.5 mg/L GA3, pH=5.8). The plates were put in dark 26 ℃ for 20 days and observed.
4.4. Metabolome and Transcriptome Experiments
Metabolome and transcriptome experiments were performed as described before [43]. Agrobacterium was cultured in 26 ℃ in dark for 2 days. The OD600 was adjusted to 0.4. 1.5 ml of Agrobacterium culture was collected and centrifugated at 500 rpm. The supernatant was discarded. The pellet was resuspended in 1 ml AAM medium containing 200 µmol/L acetosyringone. The mixture was transferred into 100 ml AAM medium containing 200 µmol/L acetosyringone and cultured for 1 hour in dark at 26 ℃. 10 g mango embryogenic callus was put in AAM medium and shaken at 500 rpm, 26 ℃ in dark for 30 minutes. The liquid was discarded. The callus was washed with sterilized water 10 times. The callus was stored in -80 ℃ until metabolome and transcriptome experiments were done.
4.5. QRT-PCR
QRT-PCR was done according to paper published [44]. NucleoSpin(R) 96 RNA core kit (Thermo Fisher) was used to extract mango callus RNA. RNA was transcribed into cDNA using Oligo(dT) 12-18 and SuperScript III Reverse Transcriptase (Invitrogen). QRT-PCR was done using CFX Real-time PCR system (Applied Biosystems) and KiCqStart SYBR Green qPCR ReadyMix (Catalog No. KCQS01, MilliporeSigma). QRT-PCR data were collected using software Bio-Rad CFX Manager Version 2.1.1022.0523. The relative expression was calculated using 2−ΔΔCT. Housekeeping gene tubulin (XM_044625649.1) was used as control. Genomic DNA in RNA samples were removed using TURBO DNase (Catalog No. AM1907, Invitrogen).
4.6. Confocal Observation
Green fluorescence in transformed callus was observed using confocal, as described in published paper [19]. Exciting light wave length was 488 nm. Scattered light wave length was 493-550 nm. The results were analyzed using software Leica Application Suite X (LAS X) software version 3.5.5.19976.
5.7. Determination of ROS Contents in Mitochondria Isolated from Mango Callus
Mitochondria were isolated and purified from mango callus according to the paper published [45]. Mitochondrial pellets were resuspended in buffer (20% glycerol, 0.1 M Tris, pH 7.0). Contents of H2O2 and O2- were measured according to the methods published [46,47]. •OH content was measured using the method described before [48]. 0.5 ml suspended mitochondrial buffer was added into 0.5 ml basic reaction system (120 mmol/L KCl, 0.4 mol/L NADH, 30 mmol/L DMSO, 40 mmol/L phosphate buffer, pH7.0). The mixture was incubated at 25 ℃ for 10 minutes. The production of formaldehyde was calculated from Nash reaction to determine the content of •OH [49].
Author Contributions
Conceptualization, Haiyan Shu, Shenghe Chang; formal analysis, Haiyan Shu, Zilhas Ahmed Jewel; investigation, Haiyan Shu, Luqiong He, Qing Wei, Rulin Zhan; resources, Shenghe Chang, Rulin Zhan; writing -original draft preparation, Haiyan Shu, Shenghe Chang; writing—review and editing, Zilhas Ahmed Jewel; supervision, Shenghe Chang; funding acquisition, Shenghe Chang, Rulin Zhan. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by National Key R&D Program Project (No. 2023YFD2300803).
Data Availability Statement
The original contributions presented in the study are included in the article; further inquiries can be directed to the corresponding authors.
References
- Rai AC, Halon E, Zemach H, Zviran T, Sisai I, Philosoph-Hadas S, Meir S, Cohen Y, Irihimovitch V. Characterization of two ethephon-induced IDA-like genes from mango, and elucidation of their involvement in regulating organ abscission. Genes (Basel), 2021, 12, 439. [CrossRef]
- Denisov Y, Glick S, Zviran T, Ish-Shalom M, Levin A, Faigenboim A, Cohen Y, Irihimovitch V. Distinct organ-specific and temporal expression profiles of auxin-related genes during mango fruitlet drop. Plant Physiol Biochem., 2017, 115:439-448. [CrossRef]
- Njie A, Zhang W, Dong X, Lu C, Pan X, Liu Q. Effect of melatonin on fruit quality via decay inhibition and enhancement of antioxidative enzyme activities and genes expression of two mango cultivars during cold storage. Foods, 2022, 11, 3209. [CrossRef]
- Zhou Y, Liu J, Zhuo Q, Zhang K, Yan J, Tang B, Wei X, Lin L, Liu K. Exogenous glutathione maintains the postharvest quality of mango fruit by modulating the ascorbate-glutathione cycle. PeerJ., 2023, 11: e15902. [CrossRef]
- Yan T, Hu C, Que Y, Song Y, Lu D, Gu J, Ren Y, He J. Chitosan coating enriched with biosynthetic CuO NPs: Effects on postharvest decay and quality of mango fruit. International Journal of Biological Macromolecules, 2023, 253:126668. [CrossRef]
- Bally ISE, Lu P, Johnson PR. Mango Breeding. Jain SM, Priyadarshan PM (eds. T: Breeding Plantation Tree Crops, 2009.
- Litz RE, Hormaza JI. Mangifera indica mango. In: Litz RE, Pliego-Alfaro F, Hormaza JI (eds) Biotechnology of fruit and nut crops, 2nd edn. 2020.
- Petri C, Litz RE, Singh SK, Hormaza JI. In vitro culture and genetic transformation in mango. C. Kole (ed.), The mango genome, compendium of plant genomes, Springer Nature Switzerland AG, 2021, pp 131-151.
- Pandey K, Karthik K, Singh SK, Vinod, Sreevathsa R, Srivastav M. Amenability of an Agrobacterium tumefaciens-mediated shoot apical meristem-targeted in planta transformation strategy in Mango (Mangifera indica L.). GM Crops & Food, 2022, 13, 342–354. [CrossRef]
- Adjei MO, Zhao H, Tao X, Yang L, Deng S, Li X, Mao X, Li S, Huang J, Luo R, Gao A, Ma J. Using a protoplast transformation system to enable functional studies in Mangifera indica L. International Journal of Molecular Sciences, 2023, 24: 11984. [CrossRef]
- Chen X, He S, Wang Z, Zhai Y, Guo W, Li X. Production of transgenic periclinal chimeras in pumpkin - a tool for revealing cell fates of L1 meristem. Plant biology, 2023, 26, 126–139. [CrossRef]
- Song X, Xie Y, Tian X, Wang N, Zhou Y, Xie Z, Ye J, Deng X. CRISPR/Cas9 editing characteristics of multiple transgenic generations in Fortunella hindsii, an early flowering mini-citrus. Scientia Horticulturae, 2023, 321: 112236. [CrossRef]
- Aida R, Sasaki K, Yoshioka S, Noda N. Distribution of cell layers in floral organs of chrysanthemum analyzed with periclinal chimeras carrying a transgene encoding fluorescent protein. Plant Cell Reports, 2020. [CrossRef]
- Raza G, Singh MB, Bhalla PL. Somatic embryogenesis and plant regeneration from commercial soybean cultivars. Plants, 2020, 9: 38. [CrossRef]
- Li M, Wang D, Long X, Hao Z, Lu Y, Zhou Y, Peng Y, Cheng T, Shi J, Chen J. Agrobacterium, 8: genetic transformation of embryogenic callus in a Liriodendron hybrid (L. Chinense × L. Tulipifera). Frontiers in Plant Science, 2022, 13, 2022. [CrossRef]
- Nakajima I, Endo M, Haji T, Moriguchi T, Yamamoto T. Embryogenic callus induction and Agrobacterium-mediated genetic transformation of ‘Shine Muscat’ grape. Plant Biotechnology, 2020, 37: 185-194. [CrossRef]
- Chen Z, Debernardi JM, Dubcovsky J, Gallavotti A. Recent advances in crop transformation technologies. Nat Plants, 2022, 8, 1343–1351.
- Christie, PJ. Type IV secretion: the Agrobacterium VirB/D4 and related conjugation systems. Biochim. Bophys. Acta, 2004, 1694: 219-234. [CrossRef]
- Raman V, Rojas CM, Vasudevan B, Dunning K, Kolape J, Oh S, Yun J, Yang L, Li G, Pant BD, Jiang Q, Mysore KS. Agrobacterium, 2: a type III secretion system delivers Pseudomonas effectors into plant cells to enhance transformation. Nature Communications, 2022, 13, 2022. [CrossRef]
- Büttner, D. Behind the lines–actions of bacterial type III effector proteins in plant cells. FEMS Microbiol. Rev., 2016, 40: 894-937. [CrossRef]
- Tang D, Wang G, Zhou J. Receptor kinases in plant-pathogen interactions: more than pattern recognition. Plant Cell, 2017, 29: 618-637.
- Zhang D, Ren L, Chen G, Zhang J, Reed BM, Shen X. ROS-induced oxidative stress and apoptosis-like event directly affect the cell viability of cryopreserved embryogenic callus in Agapanthus praecox. Plant Cell Rep, 2015, 34, 1499–513. [CrossRef]
- Agita A, Alsagaff MT. Inflammation, immunity, and hypertension. Acta Med Indones, 2017, 49, 158–165.
- Pitsili E, Phukan UJ, Coll NS. Cell death in plant immunity. Cold Spring Harb Perspect Biol 2020, 12: a036483. [CrossRef]
- Zavaliev R, Mohan R, Chen T, Dong X. Formation of NPR1 condensates promotes cell survival during plant immune response. Cell, 2020, 182, 1093–1108. [CrossRef]
- Peng T, Tao X, Xia Z, Hu S, Xue J, Zhu Q, Pan X, Zhang Q, Li S. Pathogen hijacks programmed cell death signaling by arginine ADPR-deacylization of caspases. Molecular Cell, 2022, 82: 1806-1820. [CrossRef]
- Yang F, Li G, Felix G, Albert M, Guo M. Engineered Agrobacterium improves transformation by mitigating plant immunity detection. New Phytologist, 2022, 237, 2493–2504. [CrossRef]
- Li X, Pan SQ. Agrobacterium, e: VirE2 protein into host cells via clathrin-mediated endocytosis. Science Advances, 2017, 3, 2017. [CrossRef]
- Huang HC, Schuurink R, Denny TP, Atkinson MM, Baker CJ, Yucel I, Hutcheson SW, Collmer A. Molecular cloning of a Pseudomonas syringae pv. syringae gene cluster that enables Pseudomonas fluorescens to elicit the hypersensitive response in tobacco plants. J. Bacteriol., 1988, 170, 4748–4756. [CrossRef]
- Muir RM, Ibanez AM, Uratsu SL, Ingham ES, Leslie CA, McGranahan GH, Batra N, Goyal S, Joseph J, Jemmis ED, Dandekar AM. Mechanism of gallic acid biosynthesis in bacteria (Escherichia coli) and walnut (Juglans regia). Plant Mol Biol, 2011, 75:555-565. [CrossRef]
- Zhu Y, Kun Lin K, Zhang X, Ma H, Yang L, Wei L, Yang L, Jiang M. Antibacterial effect of gallic acid in UV-C light treatment against Escherichia coli O157:H7 and the underlying mechanism. Food and Bioprocess Technology, 2023. [CrossRef]
- Ilie CI, Oprea E, Geana EI, Spoiala A, Buleandra M, Pircalabioru G, Badea IA, Ficai D, Andronescu E, Ficai A, Ditu LM. Bee pollen extracts: Chemical composition, antioxidant properties, and effect on the growth of selected probiotic and pathogenic bacteria. Antioxidants (Basel), 2022, 11(5). [CrossRef]
- Delplace F, Huard-Chauveau C, Berthome R, Roby D. Network organization of the plant immune system: from pathogen perception to robust defense induction. The Plant Journal, 2022, 109: 447-470. [CrossRef]
- Lovelace AH, Dorhmi S, Hulin MT, Li Y, Mansfield JW, Ma W. Effector identification in plant pathogens. Phytopathology, 2023, 113, 637–650. [CrossRef]
- Ngou BPM, Ding P, Jones JDG. Thirty years of resistance: Zig-zag through the plant immune system. The Plant Cell, 2022, 34: 1447-1478. [CrossRef]
- Liu T, Cao L, Cheng Y, Ji J, Wei Y, Wang C, Duan K. MKK4/5-MPK3/6 cascade regulates Agrobacterium-mediated transformation by modulating plant immunity in Arabidopsis. Frontiers in Plant Science, 2021, 12: 731690. [CrossRef]
- Cai J, Chen T, Wang Y, Qin G, Tian S. SlREM1 triggers cell death by activating an oxidative burst and other regulators. Plant Physiology, 2020, 183, 717–732. [CrossRef]
- Ding L, Li Y, Wu Y, Li T, Geng R, Cao J, Zhang W, Tan X. Plant disease resistance-related signaling pathways: recent progress and future prospects. International Journal of Molecular Sciences, 2022, 23: 16200. [CrossRef]
- Zhou JM, Zhang Y. Plant immunity: danger perception and signaling. Cell, 2020, 181: 978-989. [CrossRef]
- Tiwari M, Mishra AK, Chakrabarty D. Agrobacterium, 3: gene transfer: recent advancements and layered immunity in plants. Planta, 2022, 256, 2022. [CrossRef]
- Wise AA, Liu Z, Binns AN. in Agrobacterium Protocols (ed. Kan Wang) 43-54 (Humana Press, 2006).
- Xu W, Wu H, Luo C, Li L, Liang Q, Ma X, Yao Q, Gao Y, Wang S. Callus proliferation, early somatic embryogenesis and cytological observation of mango ‘Jinhuang’. Molecular Plant Breeding, 2019, 17, 3001–3008 (In Chinese language), (In Chinese language).
- Iqbal A, Qiang D, Xiangru W, Huiping G, Hengheng Z, Xiling Z, Meizhen S. Integrative physiological, transcriptome and metabolome analysis reveals the involvement of carbon and flavonoid biosynthesis in low phosphorus tolerance in cotton. Plant Physiology and Biochemistry, 2023, 196:302-317. [CrossRef]
- Huang and Hwang, Arabidopsis RETICULON-LIKE4 (RTNLB4) protein participates in Agrobacterium infection and VirB2 peptide-induced plant defense response. International Journal of Molecular Sciences, 2020, 21: 1722. [CrossRef]
- Vanlerberghe GC, McIntosh L. Lower growth temperature increases alternative pathway capacity and alternative oxidase protein in tobacco. Plant Physiol., 1992, 100: 115-119. [CrossRef]
- Patterson BD, Mackae EA, Ferguson IB. Estimation of hydrogen peroxide in plant extracts using Titanium. Anal. Biochem., 1984, 139: 487-492. [CrossRef]
- Wang AG, Luo GH. Quantitative relation between the reaction of hydroxylamine and superoxide anion radicals in plants. Plant Physiol. Commun., 1990, 6:55-57.
- Simontacchi M, Puntarulo S. Oxygen radical generation by isolated microsomes from soybean seedling. Plant Physiol., 1992, 100: 1263-1268. [CrossRef]
- Nash, T. The colorimetric estimation of formaldehyde by means of the Hantzsch reaction, Biochem. J., 1953, 55: 416-421. [CrossRef]
Figure 1.
Mango callus infected with Agrobacterium for different time.

Figure 2.
Reactive oxygen species produced in mango callus infected by Agrobacterium O2-, 0.1 nmol/(min mg protein); H2O2, μmol/mg protein; •OH, nmol/(min mg protein).
Figure 2.
Reactive oxygen species produced in mango callus infected by Agrobacterium O2-, 0.1 nmol/(min mg protein); H2O2, μmol/mg protein; •OH, nmol/(min mg protein).
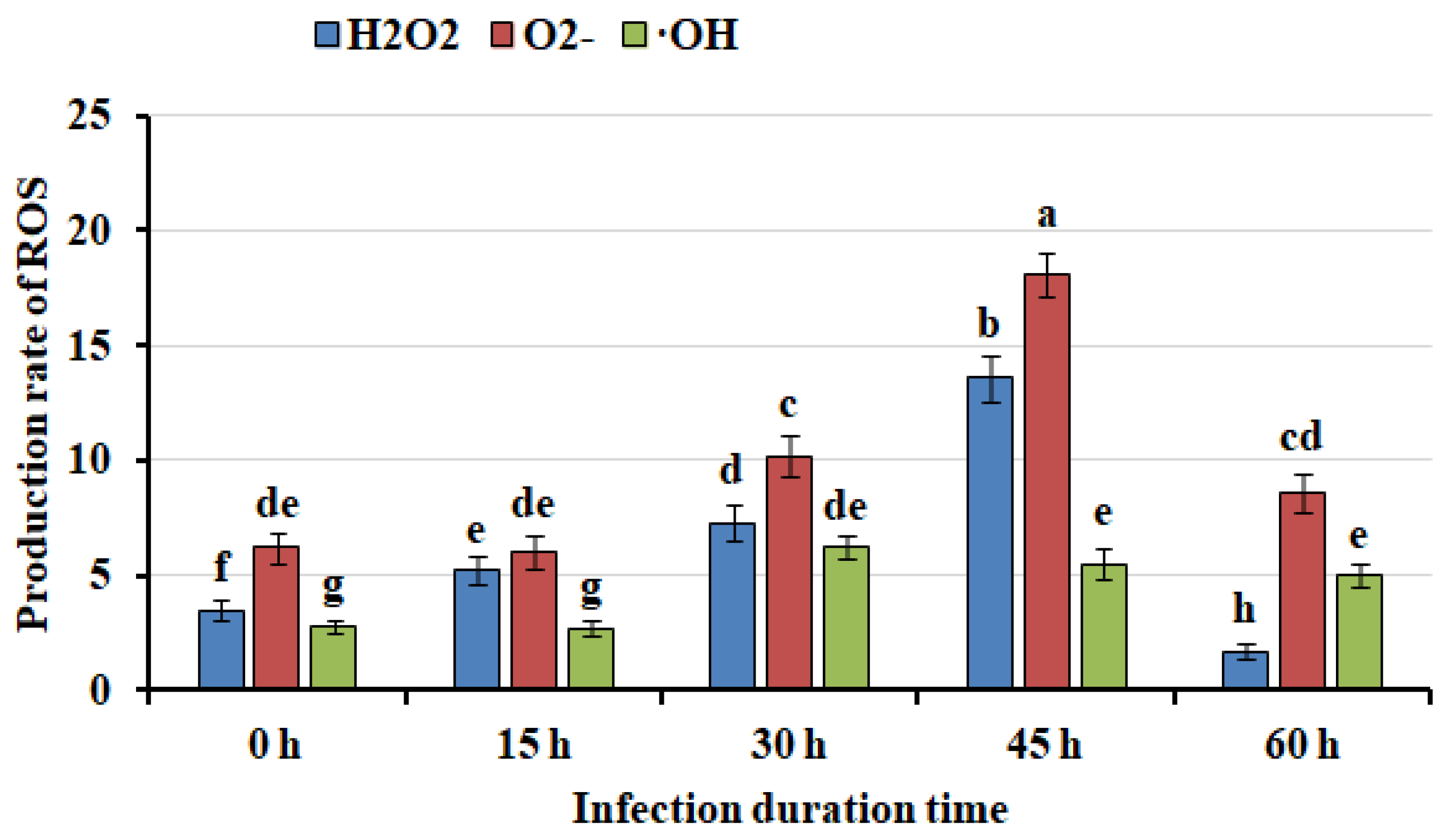
Figure 3.
Quantitative plot of differentially expressed metabolites in callus infected for 45 hours and uninfected.
Figure 3.
Quantitative plot of differentially expressed metabolites in callus infected for 45 hours and uninfected.
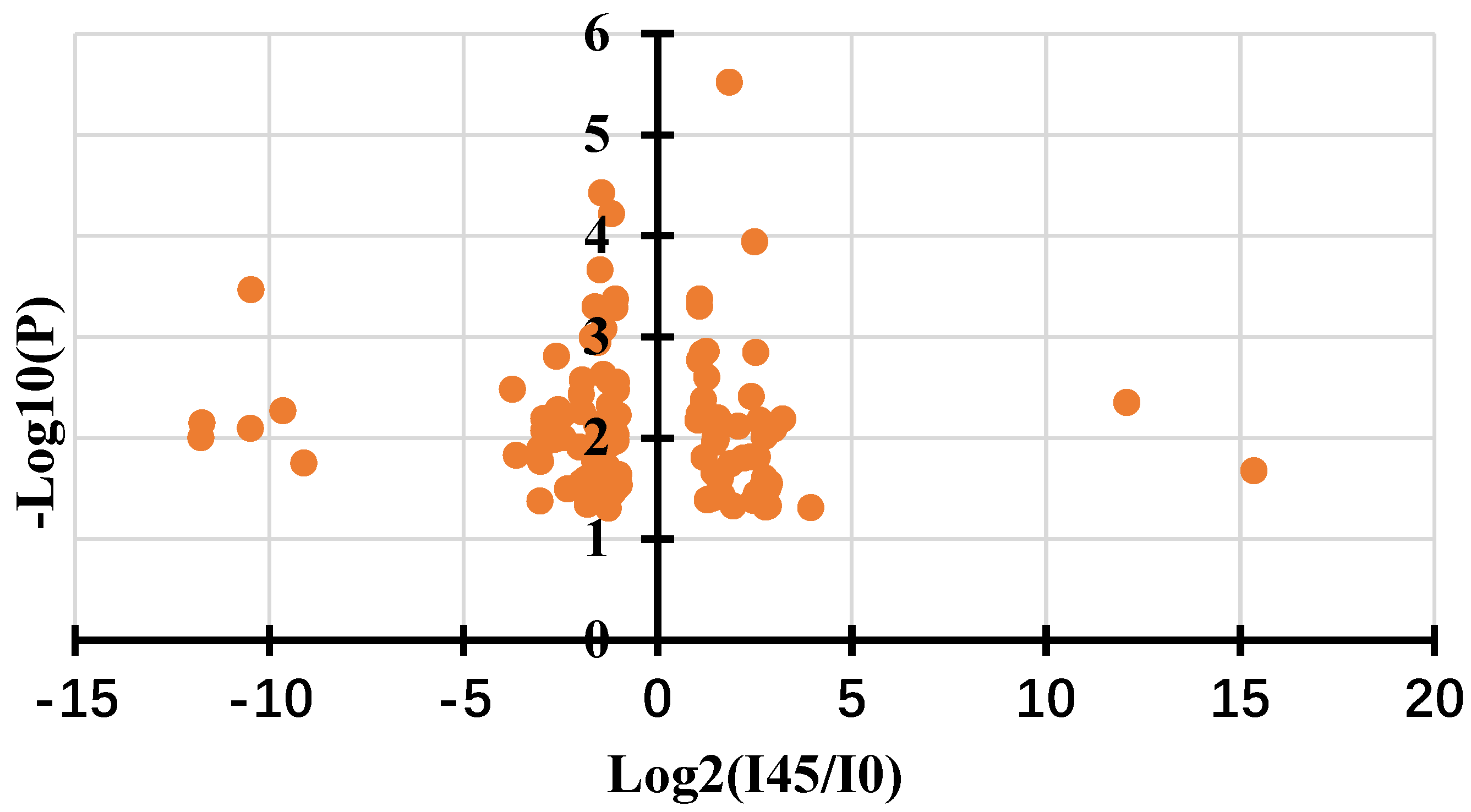
Figure 4.
Gallic acid inhibit the growth of Agrobacterium The top 4 plates showed inhibitory cycle of Agrobacterium by gallic acid. The bottom 4 plates showed Agrobacterium grown on medium without gallic acid.
Figure 4.
Gallic acid inhibit the growth of Agrobacterium The top 4 plates showed inhibitory cycle of Agrobacterium by gallic acid. The bottom 4 plates showed Agrobacterium grown on medium without gallic acid.
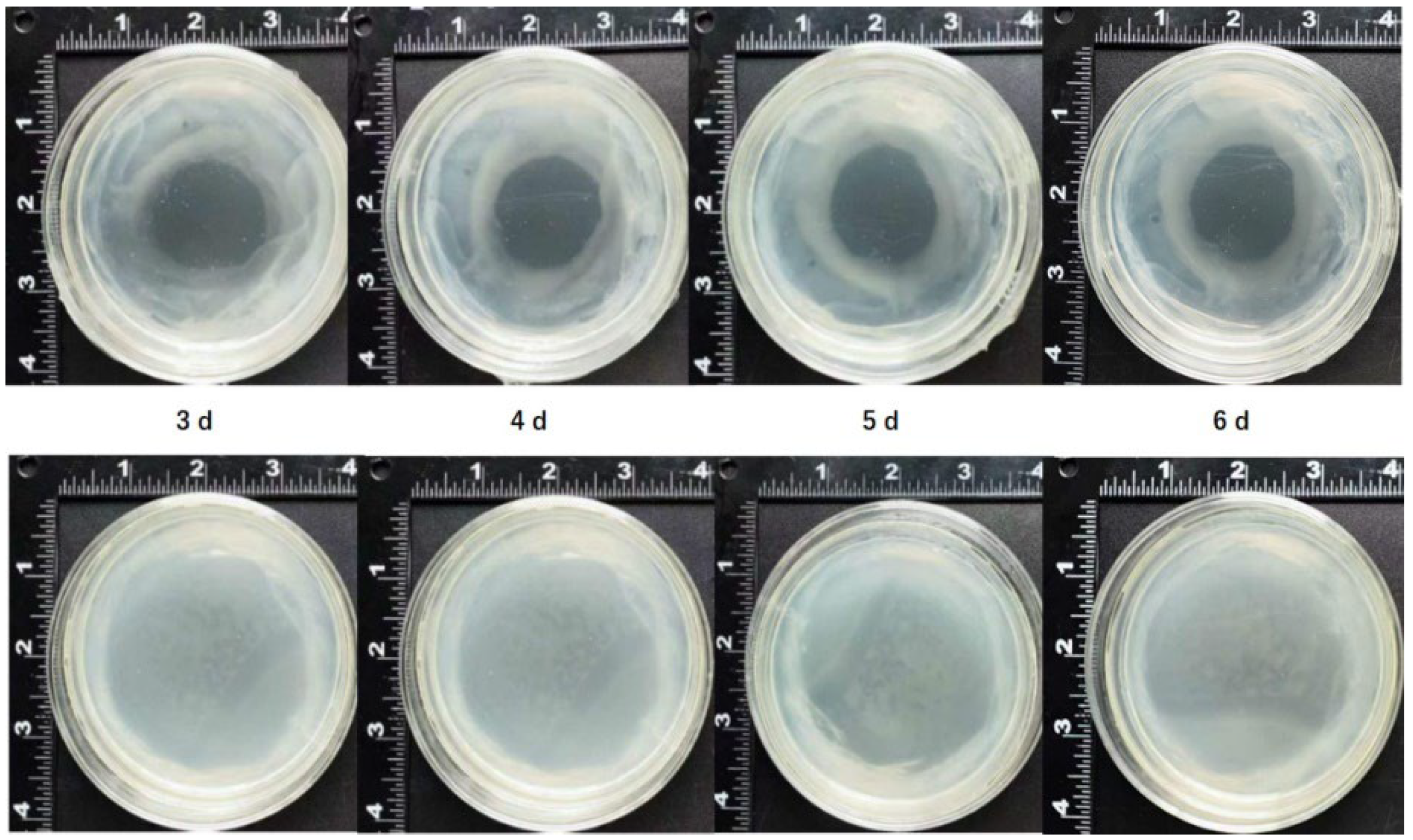
Figure 5.
GFP florescence observation after mango callus was infected by Agrobacterium (T3SS-AvrPto-gfp)
Figure 5.
GFP florescence observation after mango callus was infected by Agrobacterium (T3SS-AvrPto-gfp)
Figure 6.
The browning and necrosis of callus infected by Agrobacterium.
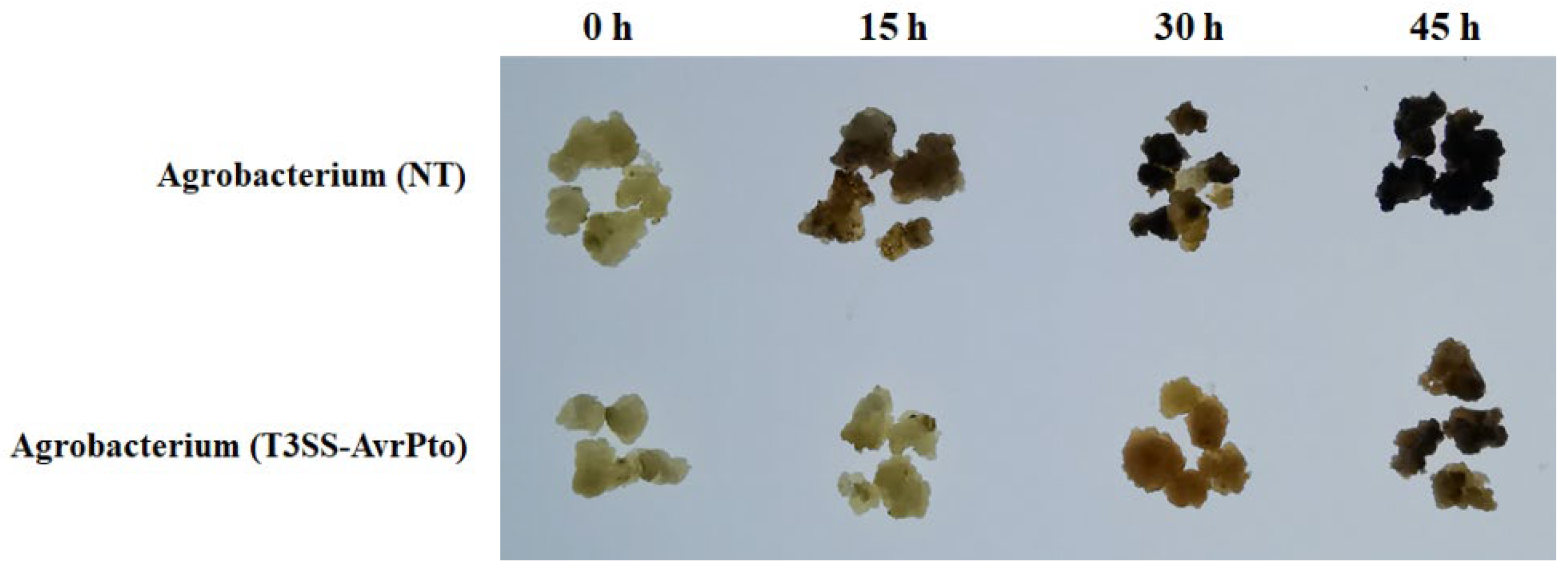
Figure 7.
The proposed mechanism underlying immune response of mango callus infected by Agrobacterium (The boldfaced arrow represented the expressions of the genes were up-regulated).
Figure 7.
The proposed mechanism underlying immune response of mango callus infected by Agrobacterium (The boldfaced arrow represented the expressions of the genes were up-regulated).


Table 1.
Main metabolites upregulated in mango callus infected by Agrobacterium.
| Index | compounds | Fold | VIP value |
| Gallic acid | Organic acid | 7.1 | 1.41 |
| Phosphoenolpyruvic acid | Organic acid | 2.9 | 1.22 |
| D-erythrose-4-phosphate | Phosphorylated derivatives of erythritol | 4.3 | 1.36 |
| Ubiquinone | Liposoluble quinone compound | 5.2 | 1.25 |
| Dihydrotricetin | Flavonoids | 2.3 | 1.26 |
| Dihydrokaempferol | Flavonoid | 1.8 | 1.19 |
| Cyanidin | Antioxidant | 2.5 | 1.28 |
Table 2.
Main genes up-regulated in callus infected by Agrobacterium for 45 hours.
| Query | Symbol | Log2(I45/I0) | Gene annotation | Subject | |
|---|---|---|---|---|---|
| LOC123211694 | DAHPS1 | 5.2 | Mangifera indica 3-oxoacyl-[acyl-carrier-protein] synthase | XM_044646917.1 | |
| LOC123214892 | DAHPS2 | 5.5 | Mangifera indica 3-oxoacyl-[acyl-carrier-protein] synthase II | XM_044641225.1 | |
| LOC123210632 | DHQD1 | 3 | Mangifera indica bifunctional 3-dehydroquinate dehydratase/shikimate dehydrogenase | XM_044644036.1 | |
| LOC123236405 | DHQD2 | 3.7 | Mangifera indica bifunctional 3-dehydroquinate dehydratase/shikimate dehydrogenase | XM_044644035.1 | |
| LOC123207068 | SHD1 | 2.4 | Mangifera indica bifunctional 3-dehydroquinate dehydratase/shikimate dehydrogenase | XM_044644037.1 | |
| LOC123215035 | SHD2 | 2.2 | Mangifera indica bifunctional 3-dehydroquinate dehydratase/shikimate dehydrogenase | XM_044644039.1 | |
| LOC123225839 | CAT1 | 2.9 | Mangifera indica catalase isozyme 1 | XM_044655447.1 | |
| LOC123228490 | CAT2 | 2.6 | Mangifera indica catalase isozyme 1 | XM_044655447.1 | |
| LOC123217301 | GOX1 | 1.7 | Mangifera indica peroxisomal (S)-2-hydroxy-acid oxidase-like | XM_044639087.1 | |
| LOC123227499 | GOX2 | 1.9 | Mangifera indica peroxisomal (S)-2-hydroxy-acid oxidase-like | XM_044618393.1 | |
| LOC123204838 | FOX1 | 3.4 | Mangifera indica FAD-linked sulfhydryl oxidase ERV1-like | XM_044652170.1 | |
| LOC123227490 | FOX2 | 3.8 | Mangifera indica FAD-linked sulfhydryl oxidase ERV1-like | XM_044625447.1 | |
| LOC123206396 | POD1 | 2.8 | Mangifera indica peroxidase A2-like | XM_044652117.1 | |
| LOC123208403 | POD2 | 2.5 | Mangifera indica lignin-forming anionic peroxidase-like | XM_044627935.1 | |
| LOC123217403 | NADO1 | 3.2 | Mangifera indica respiratory burst oxidase homolog protein D-like | XM_044606776.1 | |
| LOC123228596 | NADO2 | 3.6 | Mangifera indica respiratory burst oxidase homolog protein A-like | XM_044630849.1 | |
| LOC123217494 | FLS2-1 | 2.1 | Mangifera indica LRR receptor-like serine/threonine-protein kinase FLS2 | XM_044604788.1 | |
| LOC123227485 | MPK3-1 | 1.7 | Mangifera indica mitogen-activated protein kinase kinase 3-like | XM_044610886.1 | |
| LOC123210475 | MPK6-1 | 2.6 | Mangifera indica mitogen-activated protein kinase kinase 6 | XM_044607731.1 | |
| LOC123210574 | WRKY22-1 | 1.5 | Mangifera indica WRKY transcription factor 22-like | XM_044646893.1 | |
| LOC123226579 | NHL10-1 | 3.8 | Arabidopsis thaliana Late embryogenesis abundant (LEA) hydroxyproline-rich glycoprotein family (NDR1) | XM_044626069.1 |
Table 3.
Relative expressions of genes in mango callus infected by Agrobacterium using qRT-PCR.
| Gene | 0 h | 15 h | 30 h | 45 h | 60 h |
| MiDAHPS1 | 1.0±0.1 | 1.5±0.1 | 1.9±0.1 | 5.0±0.3 | 6.0±0.5 |
| MiDHQD1 | 0.6±0.1 | 0.6±0.1 | 0.8±0.1 | 1.9±0.2 | 2.0±0.2 |
| MiSHD1 | 1.5±0.1 | 1.7±0.1 | 1.9±0.2 | 3.6±0.2 | 3.8±0.3 |
| MiCAT1 | 0.8±0.1 | 0.8±0.1 | 1.2±0.1 | 1.9±0.2 | 1.8±0.2 |
| MiGOX1 | 2.3±0.2 | 2.5±0.2 | 2.8±0.2 | 3.9±0.3 | 3.5±0.3 |
| MiFOX1 | 1.6±0.1 | 1.7±0.1 | 2.1±0.2 | 5.6±0.3 | 5.0±0.3 |
| MiPOD1 | 3.1±0.2 | 3.2±0.2 | 3.5±0.2 | 7.3±0.5 | 6.8±0.5 |
| MiNADO1 | 1.2±0.1 | 1.2±0.1 | 1.8±0.1 | 3.6±0.2 | 3.2±0.2 |
| MiFLS2-1 | 0.8±0.1 | 0.8±0.1 | 1.1±0.1 | 1.5±0.1 | 3.9±0.2 |
| MiPK3-1 | 5.2±0.3 | 5.5±0.3 | 5.8±0.3 | 8.5±0.5 | 8.8±0.5 |
| MiPK6-1 | 3.3±0.2 | 3.5±0.2 | 3.6±0.3 | 7.2±0.5 | 7.5±0.5 |
| MiWRKY22-1 | 1.6±0.1 | 1.9±0.2 | 2.1±0.2 | 2.6±0.2 | 6.0±0.5 |
| MiNHL10-1 | 2.6±0.2 | 2.6±0.2 | 2.8±0.2 | 8.5±0.5 | 7.2±0.5 |
Table 4.
ROS contents in callus infected by Agrobacterium.
| Agrobacterium | Infection time (h) | H2O2 (μmol/mg protein) | O2- (0.1 nmol/(min mg protein)) | ·OH (nmol/(min mg protein)) |
| NV | 0 h | 3.5±0.5 | 6.2±0.6 | 2.8±0.2 |
| NV | 15 h | 5.2±0.6 | 6.0±0.5 | 2.7±0.2 |
| NV | 30 h | 13.6±1.3 | 18.1±1.5 | 5.5±0.4 |
| NV | 45 h | 7.3±0.8 | 10.2±1.0 | 6.2±0.5 |
| NV | 60 h | 1.7±0.3 | 8.6±0.7 | 5.0±0.4 |
| T3SS-AvrPto | 0 h | 3.6±0.4 | 6.1±0.5 | 2.9±0.2 |
| T3SS-AvrPto | 15 h | 3.9±0.5 | 6.3±0.6 | 2.7±0.2 |
| T3SS-AvrPto | 30 h | 4.5±0.5 | 6.6±0.5 | 3.0±0.2 |
| T3SS-AvrPto | 45 h | 4.7±0.5 | 6.9±0.6 | 3.5±0.3 |
| T3SS-AvrPto | 60 h | 5.0±0.6 | 7.6±0.6 | 4.3±0.3 |
Table 5.
Relative expressions of main genes up-regulated in callus infected by Agrobacterium.
| Gene | 0 h (NV) | 15 h (NV) | 30 h (NV) | 45 h (NV) | 60 h (NV) | 0 h (T) | 15 h (T) | 30 h (T) | 45 h (T) | 60 h (T) |
| MiDAHPS1 | 1.0±0.1 | 1.5±0.1 | 1.9±0.1 | 5.0±0.3 | 6.0±0.5 | 1.1±0.1 | 1.3±0.1 | 1.7±0.1 | 5.0±0.3 | 6.0±0.5 |
| MiDHQD1 | 0.6±0.1 | 0.6±0.1 | 0.8±0.1 | 1.9±0.2 | 2.0±0.2 | 0.5±0.1 | 0.6±0.1 | 0.8±0.1 | 1.9±0.2 | 2.0±0.2 |
| MiSHD1 | 1.5±0.1 | 1.7±0.1 | 1.9±0.2 | 3.6±0.2 | 3.8±0.3 | 1.6±0.1 | 1.7±0.1 | 1.9±0.2 | 3.1±0.2 | 3.8±0.3 |
| MiCAT1 | 0.8±0.1 | 0.8±0.1 | 1.2±0.1 | 1.9±0.2 | 1.8±0.2 | 0.7±0.1 | 0.8±0.1 | 1.0±0.1 | 1.9±0.2 | 1.8±0.2 |
| MiGOX1 | 2.3±0.2 | 2.5±0.2 | 2.8±0.2 | 3.9±0.3 | 3.5±0.3 | 2.2±0.2 | 2.5±0.2 | 2.8±0.2 | 3.9±0.3 | 3.5±0.3 |
| MiFOX1 | 1.6±0.1 | 1.7±0.1 | 2.1±0.2 | 5.6±0.3 | 5.0±0.3 | 1.5±0.1 | 1.7±0.1 | 2.1±0.2 | 3.6±0.3 | 5.0±0.3 |
| MiPOD1 | 3.1±0.2 | 3.2±0.2 | 3.5±0.2 | 7.3±0.5 | 6.8±0.5 | 3.0±0.2 | 3.2±0.2 | 3.5±0.2 | 5.6±0.3 | 7.7±0.5 |
| MiNADO1 | 1.2±0.1 | 1.2±0.1 | 1.8±0.1 | 3.6±0.2 | 3.2±0.2 | 1.3±0.1 | 1.2±0.1 | 1.8±0.1 | 3.6±0.2 | 3.2±0.2 |
| MiFLS2-1 | 0.8±0.1 | 0.8±0.1 | 1.1±0.1 | 1.5±0.1 | 3.9±0.2 | 0.9±0.1 | 0.8±0.1 | 1.1±0.1 | 1.5±0.1 | 3.9±0.2 |
| MiPK3-1 | 5.2±0.3 | 5.5±0.3 | 5.8±0.3 | 8.5±0.5 | 8.8±0.5 | 5.1±0.3 | 5.5±0.3 | 5.8±0.3 | 7.6±0.5 | 8.8±0.5 |
| MiPK6-1 | 3.3±0.2 | 3.5±0.2 | 3.6±0.3 | 7.2±0.5 | 7.5±0.5 | 3.2±0.2 | 3.5±0.2 | 3.6±0.3 | 5.2±0.5 | 7.5±0.5 |
| MiWRKY22-1 | 1.6±0.1 | 1.9±0.2 | 2.1±0.2 | 2.6±0.2 | 6.0±0.5 | 1.5±0.1 | 1.7±0.2 | 2.1±0.2 | 2.6±0.2 | 4.5±0.5 |
| MiNHL10-1 | 2.6±0.2 | 2.6±0.2 | 2.8±0.2 | 8.5±0.5 | 7.2±0.5 | 2.6±0.2 | 2.6±0.2 | 2.8±0.2 | 3.5±0.5 | 5.3±0.5 |
Note: T represented T3SS-AvrPto.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



