
Submitted:
06 July 2024
Posted:
09 July 2024
You are already at the latest version
Abstract
This study investigated the development of novel probiotic formulations specifically designed for rainbow trout aquaculture. The project evaluated the effects of two formulations containing potential probiotic strains isolated from rainbow trout itself. These effects were assessed on various parameters including growth performance, immune response, liver enzyme activity, gene expression related to immunity, and the composition of the bacterial gut microbiota in rainbow trout (Oncorhynchus mykiss). In a completely randomized experimental design, a total of 450 rainbow trout were randomly divided into 70-liter fiberglass tank. The study included six treatments of three replicates (n=18 tanks). Six dietary treatments were included (A) and (B) containing three strains of Pediococcus sp P15, Lactococcus lactis ml3, Weissella cibaria ml6 at two dosages (300 and 500 mg/kg), treatments (C) and (D) containing five strains of P. sp P15, L. lactis ml3, W. cibaria ml6, Enterococcus faecium E10 and Lactobacillus curvatus 13A at two dosages (300 and 500 mg/kg), a commercial probiotic (E) at 300 mg/kg, and a control group (F) without probiotics. Treatments of (A), (B) and (E) resulted in the highest final weights in compared to control group. Fish fed treatments (A) and (B) had the lowest FCR than other groups. Treatments (A) and (D) showed a significant increase in immunoglobulin M (IgM) concentration compared to other groups. The expression of tumor necrosis factor (TNF) was significantly lower in group (A) than the control. In contrast, the expression of interleukin 6 (IL6) and immunoglobulin T (IgT) was increased 4.4-fold and 41-fold, respectively, in group (A) compared to control group. Among the probiotic fed treatments, group (A) showed a significant higher number of lactic acid bacteria compared to other groups during the experiment. This study highlights the significant role of treatment (A) containing three strains of P. sp P15, L. lactis ml3, and W. cibaria ml6 at a dosage of 300 mg/kg as a potential probiotic product for the rainbow trout industry.
Keywords:
Rainbow trout
; Probiotics
; Growth performance
; Immunity
; Gut microbiota
; liver enzymes
; Blood parameters
1. Introduction
Aquaculture is a fast-growing industry that supplies human society with high-quality proteins [1]. Rainbow trout is one of the most economically important species in this industry. According to the latest global statistics for 2019, the global production of rainbow trout was 940,000 tonnes, an increase of 21% compared to 2015. As reported by the FAO in 2020, the production of rainbow trout in Iran doubled from 2010 to 2019. In 2019, Iran produced about 206.050 tonnes of rainbow trout, which accounts for 22% of the global production of this species [2]. The rainbow trout is among the most extensively researched species of fish. Their evolution includes a relatively recent event of tetraploidy, leading to a high frequency of gene duplication. This genetic variation could possibly enhance their ability to adapt [3]. In the field of aquaculture, infectious diseases caused by bacteria, viruses, and parasites present substantial challenges. Antibiotics are frequently used to treat and prevent bacterial infections in fish. However, excessive antibiotic usage can lead to the development of antibiotic-resistant bacteria, which poses risks to both fish and human health. Consequently, addressing this issue promptly is crucial to safeguard public health and promote sustainable aquaculture practices [1].
Probiotics are living microorganisms that can be beneficial for the health of fish if supplied in sufficient quantities. They are used in aquaculture as an alternative to antibiotics to promote growth, improve feed efficiency and strengthen the immune system of fish. The probiotics identified in aquatic animals are predominantly Bacillus spp, Pseudomonas spp, Micrococcus spp, Lactobacillus spp and Vibrio spp [4,5]. Probiotics are added to the food or water environment of fish in the form of single or multiple species. They are used to competitively prevent the growth of pathogens and improve the condition of the water. Probiotics work by colonizing the gut of fish and promoting the growth of beneficial bacteria that help maintain a healthy digestive system. They can also help prevent the growth of pathogenic bacteria by producing antimicrobial compounds. The preparation and use of specific probiotics in cold water farms can help to boost the immune system of fish, reduce economic losses due to microbial diseases, reduce incubation time by improving growth and food conversion ratio, and increase survival rate [6].
Building upon the established understanding that probiotic efficacy in fish is dependent to the strains, their viability, and water environment, this study aimed to identify novel probiotic strains specifically suited for rainbow trout (Oncorhynchus mykiss) aquaculture in Iran. We hypothesized that such strains could enhance fish health and productivity. Through a rigorous sampling and screening process, ten novel isolates exhibiting probiotic potential were identified. To assess their in vivo functionality, two formulations incorporating these strains were evaluated for their effects on rainbow trout growth performance, immune response, liver enzyme activity, immune-related gene expression, and the composition of the gut microbiota.
2. Materials and Methods
2.1. Isolation and Preparation of Microbial Suspension
Rainbow trout specimens obtained from various Iranian regions were dissected aseptically. Digestive tract contents were transferred to a sterile phosphate-buffered saline (PBS) solution to facilitate the recovery of bacteria adhering to solid particles. Approximately 10 g of homogenized samples were then suspended in 50 ml of sterile PBS and gently shaken to promote disassociation of microorganisms. Subsequently, 10 ml aliquots of the PBS suspension were inoculated into separate 190 ml liquid culture media containing de Man, Rogosa and Sharpe (MRS) and Brain Heart Infusion (BHI) broths (Ibresco, Iran) to enrich the resident bacterial population. The cultures were incubated at 37 °C under both aerobic and anaerobic conditions. Loopfuls of each broth culture were then streaked onto BHI and MRS agar plates (Ibresco, Iran) and incubated at 37 °C under both aerobic and anaerobic conditions. Finally, forty percent of colonies displaying distinct morphologies were selected for initial phenotypic characterization, including gram staining and catalase testing.
2.2. Investigating the Probiotic Properties
2.2.1. Hemolytic Activity
The hemolytic activity test was used to determine the ability of bacteria to lyse red blood cells. To perform the test, bacteria were cultured in a blood agar medium containing 5-10% (v/v) sterile defibrinated sheep blood according to the method of Yasmin et al. (2020). The plates were then incubated at 37 °C for 24-48 hours. Bacillus licheniformis 1525 with alpha-hemolytic activity was used as a positive control in this test [7].
2.2.2. Acid Test
To evaluate the acid tolerance of bacteria, a fresh bacterial culture with a starting turbidity of 1.5 × 108 colony forming units (CFU) per mL was inoculated at 10% (v/v) into MRS and BHI (Ibresco, Iran) liquid culture media adjusted to pH 3.0 with HCl. The cultures were then incubated at 37 °C under both aerobic and anaerobic conditions. Samples were collected from the cultures at zero and 3 hours after inoculation, and colony counts were performed using the dilution method on agar plates [8].
2.2.3. Bile Salts Test
To assess bile salt tolerance, a fresh bacterial culture with a turbidity of 0.5 McFarland standard (approximately 1.0 × 108 CFU/mL) was inoculated at 10% (v/v) into triplicate cultures of MRS and BHI broth (Ibresco, Iran) supplemented with 0.3% (w/v) bile salts (Sigma, USA) and uninoculated controls lacking bile salts. Cultures were incubated at 37 °C for 8 hours under aerobic conditions. Bacterial viability was then determined by measuring the optical density (OD) at 600 nm [9].
2.2.4. Antagonistic Assay
The ability of bacterial strains to inhibit the growth of pathogenic bacteria (antagonistic properties) is a crucial factor considered during strain selection. This study employed the well-diffusion assay, as described by [10], to evaluate this activity. Pathogenic bacterial isolates (Table 1) were first grown in nutrient broth (NB, Ibresco, Iran) with or without supplementation with blood, depending on the specific growth requirements. All cultures were incubated aerobically at either 37 °C or 30 °C (optimal for some pathogens) with agitation at 150 rpm for 24 hours. Subsequently, 100 µL of indicator bacterial cultures, standardized to a turbidity of 0.5 McFarland standard and in their logarithmic growth phase, were inoculated onto appropriate agar media specific for each pathogenic strain. Following surface inoculation and a 30-minute drying period, the plates were refrigerated. Four wells were then created in the agar using a sterile corn borer or punch. Supernatants (100 µL) from selected probiotic strains, previously cultured at 37 °C and 150 rpm for 48 hours, were dispensed into each well. The inoculated plates were then incubated for an additional 48 hours at 37 °C.
2.2.5. Antibiogram Assay
An antibiogram test is a standardized laboratory method used to determine the susceptibility of bacteria to various antibiotics. In this experiment, a fresh bacterial culture with a turbidity of 0.5 McFarland standard was prepared. Subsequently, 100 µL of the culture was inoculated onto Mueller-Hinton agar plates (Ibresco, Iran) using a sterile technique. Commercially available antibiotic discs containing ampicillin (10 µg), gentamicin (10 µg), streptomycin (10 µg), tetracycline (30 µg), erythromycin (15 µg), kanamycin (30 µg), vancomycin (30 µg), rifampin (5 µg), chloramphenicol (30 µg), penicillin (10 µg), ciprofloxacin (5 µg), clindamycin (2 µg), tylosin (30 µg), sulfamethoxazole/trimethoprim (30 µg), fosfomycin (200 µg), colistin (10 µg), and florfenicol (30 µg) (Padtan Teb Co, Iran) were placed onto the inoculated plates using sterile forceps, ensuring appropriate distance between each disc. The plates were then incubated for 24 hours at 37 °C under aerobic conditions. Following incubation, the presence or absence of inhibition zones surrounding each antibiotic disc was measured and interpreted according to the current Clinical and Laboratory Standards Institute (CLSI) guidelines to determine the susceptibility profile of the tested bacteria [11].
2.2.6. Attachment of Bacteria to the Epithelial Cells
Adherence of selected probiotic bacteria to Caco-2 cells was assessed using a gentamicin protection assay. Briefly, Caco-2 cells were seeded onto culture plates and allowed to grow to confluence in Dulbecco’s Modified Eagle Medium (DMEM, high glucose, Gibco-BRL) supplemented with antibiotics. The medium was then replaced with antibiotic-free DMEM, and the bacterial cultures were adjusted to a standardized density (OD 600nm = 0.5). Subsequently, 2 mL aliquots of the bacterial suspensions were added to each well containing the Caco-2 cell monolayer. The plates were incubated for 3 hours at 37 °C in a 5% CO2 incubator to allow bacterial adhesion. After incubation, non-adherent bacteria were removed by gentle aspiration of the supernatant. The monolayers were then washed twice with PBS to eliminate loosely bound bacteria. To fix and permeabilize the cells, methanol was added to the wells and incubated for 2 minutes. Following fixation, the cells were stained with a suitable viability stain (Giemsa) and visualized under a light microscope at 10x objective magnification in at least 10 random fields. The number of adherent bacteria associated with each cell was then quantified. Based on the established criteria (non-adherent: < 40 bacteria, adherent: 40-200 bacteria, highly adherent: > 200 bacteria) [12], the Caco-2 cell population was categorized for their level of bacterial adhesion.
2.2.7. Molecular Identification
The molecular identification of the bacteria was achieved using polymerase chain reaction (PCR). A specific region of the 16S ribosomal RNA (rRNA) gene was amplified with the universal primers 27F and 1492R. Following amplification, the resulting amplicons were separated by gel electrophoresis. The amplicons of interest were then sequenced using the Sanger sequencing method. The 16S rRNA gene sequences obtained from the selected probiotic strains were deposited in GenBank databases.
2.3. Farm Experiment
2.3.1. Diet Composition and Experimental Treatments
This study evaluated the effects of dietary probiotic supplementation on rainbow trout (Oncorhynchus mykiss). A commercially available rainbow trout feed (Beyza 21 Company, Iran) devoid of probiotics and antibiotics served as the basal diet. The feed contained 50-51% crude protein, 15% crude fat, ≤2% dietary fiber, ≤13% ash, 4700 kcal/kg digestible energy, and ≤10% moisture (as provided by the company). A completely randomized experimental design was employed, consisting of six treatments with three replicates each. Two probiotic formulations were prepared at varying dosages. Formulation 1 (treatments A and B) comprised Pediococcus sp P15, Lactococcus lactis ml3, and Weissella cibaria ml6. This formulation was administered at 300 mg/kg (treatment A) and 500 mg/kg (treatment B) of feed. Formulation 2 (treatments C and D) included a more complex blend of P. sp m15, L. lactis ml3, W. cibaria ml6, Enterococcus faecium E10, and Lactobacillus curvatus 13A. Similar to formulation 1, it was incorporated into the feed at 300 mg/kg (treatment C) and 500 mg/kg (treatment D). A commercially available multi-strain probiotic (300 mg/kg) served as the positive control (treatment E), while the basal diet without probiotics remained the negative control (treatment F). For probiotic incorporation, bacteria were subjected to freeze-drying with appropriate cryoprotectants (accepted paper). The viability of the resulting powder was monitored at regular intervals throughout a 12-month storage period. All formulations including the experimental and the commercial one contained 10⁹ CFU/gr bacteria. Probiotics were thoroughly mixed into the feed and stored at 4 °C until use. Fresh rations were prepared on a weekly basis.
2.3.2. Animal Experiment
A total of 450 juvenile rainbow trout (Oncorhynchus mykiss) with an average weight of 5 g and length of 7.5 cm were obtained from a commercial farm in Gilan Province, Iran. The fish were transported to the Fisheries Research Center of Bandar Anzali, Iran, for the experiment. Upon arrival, all fish underwent a thorough health examination to ensure they were free of bacterial, parasitic, and viral infections. Following a two-week acclimatization period to laboratory conditions in a single holding tank, the fish were randomly distributed among six treatment groups with three replicates per group (n = 18 tanks) of 70 L capacity each. During this acclimatization period, all fish were fed a basal diet obtained from Beyza 21 Co. at a daily ration equivalent to 3-5% of their body weight. Following acclimatization, fish in treatment groups received diets enriched with probiotics for 60 days, while the negative control group (treatment F) continued on the basal diet. Throughout the 60-day experiment, environmental parameters within the culture tanks were maintained at optimal levels for rainbow trout: temperature (12 °C ± 2.2 °C), dissolved oxygen (7.5 mg/L ± 0.1 mg/L), and pH (7.6 ± 0.4). These parameters were closely monitored.
2.3.3. Evaluation of Growth Parameters
To assess growth performance, five fish were randomly sampled from each treatment group at bi-weekly intervals (every 14 days) throughout the 60-day experiment. Fish were individually weighed and measured for length before careful return to their respective tanks. Following the sampling period, the following growth indices were calculated for fish reared under various dietary conditions: body weight, specific growth rate (SGR), condition factor (CF), and percentage weight gain. The specific growth rate was calculated using the following formula:
where T2-T1 represents the duration of the experiment in days.
The feed conversion ratio (FCR) was determined using the formula:
The condition factor (CF) was calculated using the formula:
2.3.4. Biochemical and Safety Analyses
Following the 60-day experimental period, six fish were randomly selected from each treatment group for blood collection. Blood samples (2 mL) were obtained via caudal venipuncture using a standardized procedure. Immediately after collection, samples were centrifuged at 3000 rpm for 10 minutes (Heraeus, Germany) to isolate plasma. The separated plasma samples were then stored at -21°C for subsequent analyses. Plasma samples were analyzed for a comprehensive assessment of biochemical and immunological parameters at Viromed Laboratory, Rasht, Gilan. Standardized methods and commercially available kits were used to measure the levels of various indices, including: Aspartate transaminase (AST), Alanine transaminase (ALT), Alkaline phosphatase (ALP), Lactate dehydrogenase (LDH), Immunoglobulin M (IgM), Lysozyme activity (LA), Total protein and Glucose.
2.3.5. Microbial Experiments
To investigate the composition of the gut microbiome at different time points, six fish per treatment group were randomly selected for gut sampling at 15, 30, 45, 60, and 75 days post-treatment initiation. These fish had been fed a basal diet supplemented with probiotics for 60 days, followed by a 15-day withdrawal period where probiotics were excluded from their diet. Following transportation to the laboratory, the fish were anesthetized using clove oil according to established protocols. Under sterile conditions, the abdominal cavity was disinfected with 70% ethanol, and the intestines were aseptically excised. The intestinal contents were carefully removed and rinsed three times with sterile physiological saline solution. Serial dilutions of the intestinal homogenate were prepared in sterile saline, and 0.1 mL aliquots from each dilution were plated onto MRS and TSA agar media (Ibresco, Iran) for bacterial enumeration. The plates were incubated at 37 °C for 48-72 hours under aerobic conditions. Colony forming units were then enumerated to determine bacterial abundance in the gut.
2.4. Real-Time Quantitative PCR (qPCR) Analysis
Gene expression level was measured for four immune-related genes using intestinal samples from treatments A and F. In the context of evaluating various treatments, treatment A was identified as the optimal intervention, whereas treatment F was designated as the control group for gene expression studies. Immediately after isolation, the samples were frozen in liquid nitrogen. Total RNA was extracted using Yekta Tajhiz Azma kit (Iran), and its quality and quantity were assessed by agarose gel electrophoresis and spectrophotometry (ND-2000, Nanodrop, USA), respectively. For cDNA synthesis, 3 µg RNA was subjected into the reverse transcription reaction using the AddScript cDNA Synthesis Kit (South Korea). The primers were designed using Oligo Primer 3 software as shown in Table 2.
A 10 μl real-time PCR reaction was prepared with 1 μl of cDNA, 0.3 μl (5 pmol) of reverse and forward primers, 5 μl of a 2X master mix amplicon containing SYBR Green (Amplicon, Denmark), and the remaining volume was made up with H2O. The thermal profile for all reactions consisted of an initial 15 min at 95 °C, followed by 40 cycles of 30s at 95 °C, 20s at 58 °C and 20s at 72 °C. The expression value was calculated using the following formula:
Here, ΔCt represents the change in cycle threshold, and ‘target’ refers to the gene of interest. In this equation, Etarget and Ereference are the efficiencies of the target and reference genes, respectively. The terms ΔCttarget (pool-sample) and ΔCtreference (pool-sample) represent the changes in cycle threshold for the target and reference genes, respectively, between the pool and the sample [13,14,15].
2.5. Data Analysis
After performing normality and homogeneity tests, the data set was analyzed using a one-way analysis of variance (ANOVA). The mean values were then compared using Tukey’s range test. The statistical significance of the observed differences was determined with a p-value of less than 0.05. The comparison of expression data was conducted using the student’s t-test to determine the significance of differences. Prior to this, the Shapiro-Wilk and Kolmogorov-Smirnov test was employed to assess the normality of the data distribution, ensuring the validity of parametric testing. Additionally, the Levene’s test was applied to evaluate the homogeneity of variances across groups, which is a prerequisite for the accurate application of the t-test. The entire analytical process was carried out using version 26.0 of the SPSS software.
3. Results
3.1. Isolation and Screening of Putative Probiotic Bacteria
During the initial screening phase, bacteria isolated from the gastrointestinal samples of rainbow trout exhibiting gram-positive staining and negative catalase activity were selected for further analysis. Next, isolates demonstrating negative hemolytic activity, indicating no detrimental effects on red blood cells, were evaluated for their tolerance to harsh gastrointestinal conditions. These strains were exposed to a low pH of 3, mimicking the acidic environment of the stomach, and 0.3% bile salts, simulating the intestinal environment. Isolates demonstrating high survival rates under these acidic and bile-rich conditions were considered to possess promising probiotic characteristics and were selected for further investigation. These putative probiotic strains are presented in Table 3.
Among the isolates, P15, ml3, ml6 and m6 showed superior competitiveness in inhibiting pathogenic bacteria. The antibiogram test showed analogous resistance patterns in all bacterial strains (Table 5). These results were then analyzed in accordance with the standards established by the CLSI.
Partial 16S rRNA gene sequencing was performed on the isolated bacteria to identify their taxonomic affiliations. Analysis of these sequences revealed that the isolates belonged to five distinct bacterial genera (Table 6).
Based on the above-mentioned isolates ’performance in various assays, five bacterial strains including P. sp P15, L. lactis m13, W. cibaria ml6, E. faecium E10, and L. curvatus 13A emerged as top candidates. These strains were chosen for further development and incorporated into two probiotic formulations for farm trials. The first formulation combined three of the selected strains of P. sp P15, L. lactis m13, and W. cibaria ml6. Each strain was included at a concentration of 10⁹ colony-forming units per gram of the formulation (CFU/g). The second formulation contained all five selected strains (including E. faecium E10 and L. curvatus 13A) at the same concentration of 10⁹ CFU/g per strain.
3.2.1. Physicochemical Indicators
The effects of different dietary treatments on rainbow trout were evaluated under identical conditions by examining the physicochemical properties of different tanks, including temperature, dissolved oxygen and pH. The results showed that the water temperature, dissolved oxygen and pH, was about 12 ± 3 ºC, 7.5 ± 0.1 mg/L, and 7.6 ± 0.4, respectively (Table 7). Throughout the experiment, the physicochemical factors in the different tanks showed no significant differences (p> 0.05).
3.2.2. Animal Experiment and Growth Performance
The influence of different probiotic-enriched diets on fish growth performance was investigated (Figure 1). No significant differences were observed in initial length, weight, or final length across treatments (p > 0.05) (Figure 1d). However, treatments A, B, and E resulted in significantly higher final weights (41.06 g, 37 g, and 38 g, respectively) compared to the control group (Figure 1e). These same treatments also displayed the lowest FCR among all groups (p < 0.05) (Figure 1a). In terms of SGR, fish from treatments A and B exhibited optimal growth with an average SGR of 3.5 (p < 0.05) (Figure 1b). Interestingly, no significant variations in CF, an indicator of fish plumpness, were observed between treatments (Figure 1c).
3.2.3. Hematological Parameters
Fish fed diets A and E exhibited the highest red blood cell (RBC) count at 1.5 × 10⁶ cells/mL compared to other groups (p < 0.05, Figure 2a). Mean MCH, mean MCV, mean corpuscular hemoglobin concentration (MCHC), and white blood cell (WBC) counts, including neutrophils and monocytes, were not significantly affected by the probiotic treatments (p > 0.05). Interestingly, treatment B displayed the lowest hematocrit (Hct) value among all groups. However, fish fed diets A, B, and E had the highest hemoglobin content (Hb) (p < 0.05). Furthermore, the percentage of lymphocytes was significantly higher in fish fed diets A and B compared to the other groups (83.9% vs. 78.9%, p < 0.05). Notably, eosinophil counts were the lowest in treatments A, B, C, and E compared to the control group, with treatments C and D showing intermediate values.
3.2.4. Biochemical and Immune Parameters
Fish fed diets A and D displayed a significant elevation in IgM levels compared to the control and other dietary probiotic groups (p < 0.05, Figure 3a). The mean IgM concentration in treatments A and D was 62.8 mg/dL, while the other probiotic groups exhibited intermediate values. Measuring the blood lysozyme concentration showed that the fish fed dietary treatments of A, D and E had a significant higher LA than other groups (p<0.05). The liver enzymes levels including ALP, LDH, ALT and AST were significantly higher in dietary treatments of E, F (555 u/l), E (2125 u/l) and A, B, C, F (15 u/l) and F (2125 u/l), respectively. The highest amount of total glucose was observed in treatments of A and F (114 mg/dl), while the highest level of total protein was observed in fish fed dietary treatments of B, C, D, E and F, in contrast to group A (3.7 u/l vs 2.94 u/l; p<0.05).
3.2.5. Intestinal Bacteria Measurement
As shown in Figure 4a, the number of the lactic acid bacteria in different treatments that fed with probiotic was significantly higher than the control group (treatment F) over the course of the experiment (p<0.05). Among the probiotic fed treatments, group A showed a significant higher number of lactic acid bacteria compared to other groups during the experiment, while there were no significant differences between other groups (p<0.05). Figure 4b presents the heterotrophic bacterial count in the fish’s intestines at various intervals - 15, 30, 45, and 60 days following the probiotics administration, and four weeks post the discontinuation of the probiotics. The experiment did not yield consistent results for the heterotrophic bacterial count. On the 15th and 30th days, the lowest count of heterotrophic bacteria was observed in treatments C and F, respectively, when compared to other treatments (p<0.05). However, on the 45th day, treatment A recorded the highest count of heterotrophic bacteria among the probiotic-fed treatments (p<0.05). Subsequently, on the 60th and 75th days, no significant differences were noted among the groups (p>0.05).
3.3. Relative mRNA Expression of Immune-Related Genes
In Figure 5a,d, IL-6 and IgT gene expression exhibited significant upregulation in group A compared to the control group (4.4-fold and 41-fold increase, respectively; p < 0.05). Conversely, TNF-α gene expression in group A displayed a significant downregulation (5-fold decrease) compared to the control (Figure 5b). No significant difference was observed in INF-γ gene expression between the two groups (Figure 5c).
4. Discussion
Probiotics are a promising alternative to promote the growth and health of various fish species. They are currently being investigated as a possible replacement for the overuse of antibiotics in aquaculture. These beneficial bacteria have been shown to significantly affect the health and growth performance of the host organism [16]. There is empirical evidence that the inclusion of probiotics in the feed of farmed fish increases disease resistance, reduce susceptibility to stress and enhance overall vitality [17,18]. In the current study, we conducted a screening of probiotic bacteria from the gastrointestinal tract of rainbow trout. In the initial phase of the experiment, the isolated strains were both Gram-positive and catalase-negative. Considering various factors such as bile and acid tolerance, hemolytic and antibacterial activity, attachment to caco2 cell line and antagonistic test, five strains of P. sp P15, L. lactis ml3, W. cibaria ml6, E. faecium E10 and L. curvatus 13A showed superior performance. The vast majority of the immunomodulatory probiotic bacteria are Gram-positive lactic acid bacteria. These Gram-positive bacteria are promising options for probiotics due to their beneficial properties and their ability to survive in the harsh conditions of the gastrointestinal tract [19,20]. Probiotic bacteria must be able to survive in an acidic environment. These bacteria have evolved several mechanisms to survive in an acidic environment. These mechanisms include changes in the pH of the environment, production of acid-neutralizing compounds, changes in membrane fatty acid composition, and activation of stress response systems [21]. In the context of aquaculture, especially in rainbow trout farms, the isolation of probiotics that can inhibit pathogens is of utmost importance. This is mainly due to the significant economic impact associated with disease outbreaks [22,23]. Therefore, the value of such probiotic strains cannot be overstated. Probiotics can prevent disease outbreaks by inhibiting the growth and colonization of pathogenic bacteria in the host organism. In addition, probiotics can improve the host’s immune response so that it is better able to fight off infections, resulting in better health and survival rates of rainbow trout populations.
The evaluation of growth parameters during the feeding experiment showed a significant improvement in the growth performance of rainbow trout. This improvement was particularly observed in fish fed diet containing P. sp P15, L. lactis m13 and W. cibaria ml6 (treatment A) compared to the control group. Although the final body weight of other probiotic treatments was also numerically increased. Previous studies have reported the beneficial effects of dietary supplementation with probiotic bacteria from the genera Weissella, Pediococcus and Lactococcus on rainbow trout [24,25,26,27]. Kahyani et al. (2021) showed that feeding rainbow trout with diet supplemented with Weissella confusa at the level of 3×107 CFU/g has been led to a significant improvement in growth performance and immunity compared to the control [25]. Another study by Yeganeh et al. (2021) reported the beneficial effects of L. lactis subsp. lactis PTCC 1403 on the feed efficiency, immune response, and resistance to Y. ruckeri infection in rainbow trout [27]. In this study, the observed increase in growth rate could possibly be attributed to the influence of lactic acid bacteria (LAB) in competitively excluding pathogenic microbes, producing digestive enzymes, providing nutrients or vitamins and improving intestinal structure [28].
Figure 2 shows the hematological parameters of rainbow trout fed with different treatments. Interestingly, an increase in the number of RBCs and hemoglobin was observed in rainbow trout fed with treatments A and E. Red blood cells in fish play an important role in the immune response. They are involved in the recognition of pathogens, the elimination of pathogens by binding microbial immune complexes, and the production of cytokines or specific signaling molecules in response to pathogens [29]. In accordance with the present results, previous studies have reported that probiotics can stimulate erythropoiesis, the process by which new RBCs are formed. Since hemoglobin is an important component of RBCs, this could lead to an increase in hemoglobin levels [30,31].
In fish species, immunoglobulin M (IgM) fulfils an important function in both the systemic and mucosal immune systems by protecting the internal systems and mucosal surfaces exposed to the environment [32]. The administration of probiotics has been observed to stimulate the immune system of fish, leading to an increase in the production of antibodies, including IgM [33,34]. Consistent with the literature, this research found a remarkable increase in IgM level in the fish subjected to probiotic treatments A and D. This increased IgM level may improve the fish’s ability to fight infections.
The present study was also designed to determine the effects of different probiotic formulations on the activity of certain liver enzymes in the fish intestine. The results showed that alkaline phosphatase (ALP) level was elevated in treatments of E and F compared to the other groups and lactate dehydrogenase (LDH) was profoundly higher in treatment E. Conversely, alanine transaminase (ALT) levels were lower in fish fed treatments of D and E and AST was higher in the control group than others. These results further support the idea that emphasizes the strain-specific effects of probiotics on liver enzyme activity in fish [34,35] and microbiome effects on different organs activity such as liver [36]. Previous studies have shown that two strains of Lactobacillus plantarum, 426951 and KC426951, improve the growth and immunity of rainbow trout. However, of the liver enzymes, only ALP was elevated in rainbow trout fed strain 426951. Alanine transaminase plays a crucial role in protein metabolism and serves as an important indicator of liver health in fish [37]. Alkaline phosphatase plays an important role in the immune system of fish. Alkaline phosphatase can dephosphorylate lipopolysaccharides originating from bacteria. This detoxification process can help prevent bacterial translocation and heal intestinal inflammation [38].
This study demonstrated a significant increase in gut lactic acid bacteria (LAB) counts in fish fed probiotic diets compared to the control group. Notably, treatment A displayed the highest LAB abundance. These findings align with previous research indicating that probiotics can enhance LAB colonization and modulate gut fish microbiome [39]. However, conflicting evidence exists. Another study reported a lack of probiotic L. lactis colonization in the host’s intestinal mucosa [40,41]. Establishing probiotic colonization within the gut depending to the individual and strains is complex and influenced by intricate molecular interactions, as acknowledged [41]. Interestingly, our results showed a persistent increase in LAB even two weeks after probiotic withdrawal (days 60-75). This suggests that all probiotic groups, particularly treatment A, may have the potential for transient and colonization of the intestinal mucosa and influence the gut microbiota composition. Furthermore, a numerical increase in gut heterotrophic bacteria was also observed in probiotic-treated rainbow trout, with treatment A again showing the most pronounced effect. Probiotics can modulate mucosal immunity in fish, potentially leading to the proliferation of specific bacterial populations, including heterotrophic bacteria, through mechanisms like stimulating granulocytes and lymphocytes (essential components of cell-mediated mucosal defense) [37]. Additionally, probiotics may produce compounds that inhibit pathogenic gut organisms, creating a more favorable environment for the growth of beneficial heterotrophic bacteria [42]. However, further investigation is needed to confirm these potential mechanisms in our specific context.
Probiotics have a multifaceted impact on the immune system. Our results indicated that while the gene expression level of Interferon-γ remained unchanged with the top selected probiotic diet (treatment A), there was a significant increase in the levels of IL-6 and IgT in the fish that consumed the same probiotic-infused diet. The influence of probiotics on immune gene expression can differ based on the probiotic strain used, the dosage administered, the duration of the treatment, and the fish species [43]. Immunoglobulin T (IgT) is a key player in the immune defense of fish. Beneficial bacteria such as probiotics boost immune response in fish, including the production of antibodies such as IgT. Probiotics administration can rise in the level of IgT and other antibodies in the fish gut mucus. This effect of probiotics is attributed to the’ ability of this bacteria to regulate the gut microbiota. Probiotics enhance the fish’s capacity to ward off pathogens, mitigate disease and stress, enhance water quality, and even promote growth and reproduction [13,44]. TNF-α is an inflammatory cytokine belonging to a group of cytokines that stimulate the acute phase response. IL-6 is often upregulated during inflammation, infection or probiotic ingestion and plays a key role in activating the immune response. Conversely, TNF-α, which plays a role in systemic inflammation, can be downregulated as a protective mechanism to attenuate excessive inflammation and potential tissue damage to the host organism [45,46].
5. Conclusion
Probiotics are essential in aquaculture as they provide numerous benefits, such as improving growth rate and strengthening the immune system, which improves the fish’s ability to resist disease. This project was undertaken to design two probiotic formulations each with two concentrations and evaluate their probiotic effects on the growth, enzyme activity and immune factors expression of rainbow trout. These experiments confirmed that feeding rainbow trout with different probiotic formulation enhanced the growth performance, liver enzymes level and modulate the immune systems. However, the results of this research support the idea that the beneficial effects of probiotic in host is strain specific and depends to the type of probiotic bacteria used in the formulation and the concentration of the used probiotic in feed as well. In the present study, dietary probiotic treatment containing three strains of P. sp P15, L. lactis m13 and W. cibaria ml6 showed significant better overall results in terms of fish performance and immune system and can therefore be recommended as a new potential probiotic formulation for the rainbow trout industry. It should be noted that the production of probiotics with three strains is easier and more economical from an industrial point of view than the production of products with several strains.
Author Contributions
Sahar Mahmoodian contributed to data curation, formal analysis, and original draft preparation. Amir Meimandipour contributed to conceptualization, funding acquisition, methodology, supervision, and review and editing. Monireh Faeed contributed to conceptualization, methodology, data curation, formal analysis, and original draft preparation. Mehdi Shamsara and Seyed Safa-Ali Fatemi contributed to methodology, supervision, review and editing. Javad Daghigh roohi, Mohades Ghasemi was responsible for fish feeding and took part in the sampling. Mahsa Chaharmahali contributed to data curation, review and editing.
Data Availability Statement
The data that support the findings of the present study are available from the corresponding author upon reasonable request.
Ethical Approval
All experiments and samplings were done based on the ethical recommendations in the guide for the care, protection, and use of laboratory animals approved by the Institutional Animal Care and Use Committee of the Iranian Fisheries Science Research Institute [47].
Acknowledgments
The current study has been financially supported by the National Institute of Genetic Engineering and Biotechnology (106T-I-951221).
Conflict of Interest
The authors declare that they have no conflict of interest.
References
- D’Agaro, E.; Gibertoni, P.; Esposito, S. “Recent Trends and Economic Aspects in the Rainbow Trout (Oncorhynchus mykiss) Sector”. Appl. Sci. 2022, 12, 8773. [Google Scholar] [CrossRef]
- FAO ed, “Contributing to food security and nutrition for all.”, Rome, 2016.
- Thorgaard, G.H.; Bailey, G.S.; Williams, D.; Buhler, D.R.; Kaattari, S.L.; Ristow, S.S.; Hansen, J.D.; Winton, J.R.; Bartholomew, J.L.; Nagler, J.J.; Walsh, P.J.; Vijayan, M.M.; Devlin, R.H.; Hardy, R.W.; Overturf, K.E.; Young, W.P.; Robison, B.D.; Rexroad, C.; Palti, Y. “Status and opportunities for genomics research with rainbow trout”. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2002, 133, 609–646. [Google Scholar] [CrossRef]
- Saettone, V.; Biasato, I.; Radice, E.; Schiavone, A.; Bergero, D.; Meineri, G. “State-of-the-Art of the Nutritional Alternatives to the Use of Antibiotics in Humans and Monogastric Animals”. Anim. Open Access J. MDPI. 2020, 10, 2199. [Google Scholar] [CrossRef] [PubMed]
- Monzón-Atienza, L.; Bravo, J.; Serradell, A.; Montero, D.; Gómez-Mercader, A.; Acosta, F. “Current Status of Probiotics in European Sea Bass Aquaculture as One Important Mediterranean and Atlantic Commercial Species: A Review”. Animals. 2023, 13, 2369. [Google Scholar] [CrossRef]
- Wuertz, S.; Schroeder, A.; Wanka, K.M. “Probiotics in Fish Nutrition—Long-Standing Household Remedy or Native Nutraceuticals?”. Water. 2021, 13, 1348. [Google Scholar] [CrossRef]
- Yasmin; Saeed, M. ; Khan, W.A.; Khaliq, A.; Chughtai, M.F.J.; Iqbal, R.; Tehseen, S.; Naz, S.; Liaqat, A.; Mehmood, T.; Ahsan, S.; Tanweer, S. “In Vitro Probiotic Potential and Safety Evaluation (Hemolytic, Cytotoxic Activity) of Bifidobacterium Strains Isolated from Raw Camel Milk.” Microorganisms. 2020, 8, 354.
- Yu, Z.; Zhang, X.; Li, S.; Li, C.; Li, D.; Yang, Z. “Evaluation of probiotic properties of Lactobacillus plantarum strains isolated from Chinese sauerkraut”. World J. Microbiol. Biotechnol. 2013, 29, 489–498. [Google Scholar] [CrossRef]
- Sato, S.T.A.; Marques, J.M.; da Luz de Freitas, A.; Progênio, R.C.S.; Nunes, M.R.T.; de Vasconcelos Massafra, J.M.; Moura, F.G.; Rogez, H. “Isolation and Genetic Identification of Endophytic Lactic Acid Bacteria From the Amazonian Açai Fruits: Probiotics Features of Selected Strains and Their Potential to Inhibit Pathogens.” Front. Microbiol. 2021, 11.
- Kavitha, M.; Raja, M.; Perumal, P. “Evaluation of probiotic potential of Bacillus spp. isolated from the digestive tract of freshwater fish Labeo calbasu (Hamilton, 1822).” Aquac. Rep. 2018, 11, 59–69. [Google Scholar]
- Hechard, Y.; Dherbomez, M.; Cenatiempo, Y.; Letellier, F. “Antagonism of lactic acid bacteria from goats’ milk against pathogenic strains assessed by the ‘sandwich method. ’” Lett. Appl. Microbiol. 1990, 11, 185–188. [Google Scholar] [CrossRef]
- Dimitrov, Z.; Gotova, I.; Chorbadjiyska, E. “In vitro characterization of the adhesive factors of selected probiotics to Caco-2 epithelium cell line”. Biotechnol. Biotechnol. Equip. 2014, 28, 1079–1083. [Google Scholar] [CrossRef]
- Pérez-Sánchez, T.; Balcázar, J.L.; Merrifield, D.L.; Carnevali, O.; Gioacchini, G.; De Blas, I.; Ruiz-Zarzuela, I. “Expression of immune-related genes in rainbow trout (Oncorhynchus mykiss) induced by probiotic bacteria during Lactococcus garvieae infection”. Fish Shellfish Immunol. 2011, 31, 196–201. [Google Scholar] [CrossRef] [PubMed]
- Evenhuis, J.P.; Cleveland, B.M. “Modulation of rainbow trout (Oncorhynchus mykiss) intestinal immune gene expression following bacterial challenge”. Vet. Immunol. Immunopathol. 2012, 146, 8–17. [Google Scholar] [CrossRef] [PubMed]
- Pfaffl, M.W. “A new mathematical model for relative quantification in real-time RT-PCR”. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Kong, Y.; Chang, X.; Feng, J.; Wang, X.; Hou, L.; Zhao, X.; Pei, C.; Kong, X. “Effects of two fish-derived probiotics on growth performance, innate immune response, intestinal health, and disease resistance of Procambarus clarkii”. Aquaculture. 2023, 562, 738765. [Google Scholar] [CrossRef]
- Mondal, S.; Debashri, M.; Mondal, T.; Malik, J. “Application of probiotic bacteria for the management of fish health in aquaculture.” Bacterial Fish Diseases. pp. 351–378 (2022).
- Hoseinifar, S.H.; Sun, Y.-Z.; Wang, A.; Zhou, Z. “Probiotics as Means of Diseases Control in Aquaculture, a Review of Current Knowledge and Future Perspectives.” Front. Microbiol. 2018, 9.
- Audisio, M.C. “Gram-Positive Bacteria with Probiotic Potential for the Apis mellifera L. Honey Bee: The Experience in the Northwest of Argentina.” Probiotics Antimicrob. Proteins. 2017, 9, 22–31. [Google Scholar] [PubMed]
- Chen, J.; Chen, X.; Ho, C.L. “Recent Development of Probiotic Bifidobacteria for Treating Human Diseases.” Front. Bioeng. Biotechnol. 2021, 9.
- Wang, J.; Chen, L. “Impact of a Novel Nano-Protectant on the Viability of Probiotic Bacterium Lactobacillus casei K17”. Foods. 2021, 10, 529. [Google Scholar] [CrossRef]
- Bălbărău; Ivanescu, L. M.; Martinescu, G.; Rîmbu, C.M.; Acatrinei, D.; Lazar, M.; Cocean, I.; Gurlui, S.; Cocean, A.; Miron, L. “Septicemic Outbreak in A Rainbow Trout Intensive Aquaculture System: Clinical Finds, Etiological Agents, and Predisposing Factors.” Life. 2023, 13, 2083.
- Daneshamouz, S.; Haghi, F.; Zeighami, H. “Detection and Identification of Bacterial Pathogens in Rainbow Trout (Oncorhynchus mykiss) Samples from Fish Farms in Iran”. Thalass. Int. J. Mar. Sci. 2020, 36, 133–141. [Google Scholar] [CrossRef]
- Al-Hisnawi; Rodiles, A. ; Rawling, M.D.; Castex, M.; Waines, P.; Gioacchini, G.; Carnevali, O.; Merrifield, D.L. “Dietary probiotic Pediococcus acidilactici MA18/5M modulates the intestinal microbiota and stimulates intestinal immunity in rainbow trout (Oncorhynchus mykiss).” J. World Aquac. Soc. 2019, 50, 1133–1151.
- Kahyani, F.; Pirali-Kheirabadi, E.; Shafiei, S.; Masouleh, A.S. “Effect of dietary supplementation of potential probiotic Weissella confusa on innate immunity, immune-related genes expression, intestinal microbiota and growth performance of rainbow trout (Oncorhynchus mykiss)”. Aquac. Nutr. 2021, 27, 1411–1420. [Google Scholar] [CrossRef]
- Martinez, M.P.; Pereyra, M.L.G.; Pena, G.A.; Poloni, V.; Juri, G.F.; Cavaglieri, L.R. “Pediococcus acidolactici and Pediococcus pentosaceus isolated from a rainbow trout ecosystem have probiotic and ABF1 adsorbing/degrading abilities in vitro”. Food Addit. Contam. Part Chem. Anal. Control Expo. Risk Assess. 2017, 34, 2118–2130. [Google Scholar] [CrossRef] [PubMed]
- Yeganeh, S.; Adel, M.; Nosratimovafagh, A.; Dawood, M.A.O. “The Effect of Lactococcus lactis subsp. lactis PTCC 1403 on the Growth Performance, Digestive Enzymes Activity, Antioxidative Status, Immune Response, and Disease Resistance of Rainbow Trout (Oncorhynchus mykiss).” Probiotics Antimicrob. Proteins. 2021, 13, 1723–1733. [Google Scholar] [PubMed]
- Pérez-Sánchez, T.; Ruiz-Zarzuela, I.; de Blas, I.; Balcázar, J.L. “Probiotics in aquaculture: a current assessment”. Rev. Aquac. 2014, 6, 133–146. [Google Scholar] [CrossRef]
- del M, M.; Coll, J.; Rimstad, E. “Editorial: The role of red blood cells in the immune response of fish.” Front. Immunol. 2022, 13.
- de C, D.; Tachibana, L.; Iwashita, M.K.P.; Nakandakare, I.B.; Romagosa, E.; Seriani, R.; Ranzani-Paiva, M.J.T. “Probiotic supplementation causes hematological changes and improves non-specific immunity in Brycon amazonicus.” Acta Sci. Biol. Sci. 2020, 42.
- Mohapatra, S.; Chakraborty, T.; Prusty, A.K.; PaniPrasad, K.; Mohanta, K.N. “Beneficial Effects of Dietary Probiotics Mixture on Hemato-Immunology and Cell Apoptosis of Labeo rohita Fingerlings Reared at Higher Water Temperatures”. PLOS ONE. 2014, 9, e100929. [Google Scholar] [CrossRef] [PubMed]
- Xia, H.; Lu, J.; Yang, P.; Chen, F.; Zhang, Y.; Liu, L.; Chen, Z.; Huang, R. “Research Progress in Molecular Biology of Fish Immunoglobulin M (IgM)”. Isr. J. Aquac. - Bamidgeh. 2023, 75, 1–7. [Google Scholar] [CrossRef]
- Kuebutornye, F.K.A.; Tang, J.; Cai, J.; Yu, H.; Wang, Z.; Abarike, E.D.; Lu, Y.; Li, Y.; Afriyie, G. “In vivo assessment of the probiotic potentials of three host-associated Bacillus species on growth performance, health status and disease resistance of Oreochromis niloticus against Streptococcus agalactiae”. Aquaculture. 2020, 527, 735440. [Google Scholar] [CrossRef]
- Soltani, M.; Pakzad, K.; Taheri-Mirghaed, A.; Mirzargar, S.; Shekarabi, S.P.H.; Yosefi, P.; Soleymani, N. “Dietary Application of the Probiotic Lactobacillus plantarum 426951 Enhances Immune Status and Growth of Rainbow Trout (Oncorhynchus mykiss) Vaccinated Against Yersinia ruckeri”. Probiotics Antimicrob. Proteins. 2019, 11, 207–219. [Google Scholar] [CrossRef]
- Soltani, M. “Effect of the probiotic, Lactobacillus plantarum on growth performance and haematological indices of rainbow trout (Oncorhynchus mykiss) immunized with bivalent streptococcosis / lactococcosis vaccine.” Iran. J. Fish. Sci. no. Online First, 2018.
- Singh, R.; Wang, Z.; Marques, C.; Min, R.; Zhang, B.; Kumar, S. “Alanine aminotransferase detection using TIT assisted four tapered fiber structure-based LSPR sensor: From healthcare to marine life”. Biosens. Bioelectron. 2023, 236, 115424. [Google Scholar] [CrossRef] [PubMed]
- Rader, B.A. “Alkaline Phosphatase, an Unconventional Immune Protein.” Front. Immunol. 2017, 8.
- Hasan, K.N.; Banerjee, G. “Recent studies on probiotics as beneficial mediator in aquaculture: a review”. J. Basic Appl. Zool. 2020, 81, 53. [Google Scholar] [CrossRef]
- Hasan; Rimoldi, S. ; Saroglia, G.; Terova, G. “Sustainable Fish Feeds with Insects and Probiotics Positively Affect Freshwater and Marine Fish Gut Microbiota.” Animals. 2023, 13, 1633.
- Knobloch, S.; Skírnisdóttir, S.; Dubois, M.; Kolypczuk, L.; Leroi, F.; Leeper, A.; Passerini, D.; Marteinsson, V.Þ. “Impact of Putative Probiotics on Growth, Behavior, and the Gut Microbiome of Farmed Arctic Char (Salvelinus alpinus).” Front. Microbiol. 2022, 13.
- Moroni, F.; Naya-Català, F.; Piazzon, M.C.; Rimoldi, S.; Calduch-Giner, J.; Giardini, A.; Martínez, I.; Brambilla, F.; Pérez-Sánchez, J.; Terova, G. “The Effects of Nisin-Producing Lactococcus lactis Strain Used as Probiotic on Gilthead Sea Bream (Sparus aurata) Growth, Gut Microbiota, and Transcriptional Response.” Front. Mar. Sci. 2021, 8.
- De, B.C.; Meena, D.K.; Behera, B.K.; Das, P.; Mohapatra, P.K.D.; Sharma, A.P. “Probiotics in fish and shellfish culture: immunomodulatory and ecophysiological responses”. Fish Physiol. Biochem. 2014, 40, 921–971. [Google Scholar]
- Dang, Y.; Sun, Y.; Zhou, Y.; Men, X.; Wang, B.; Li, B.; Ren, Y. “Effects of probiotics on growth, the toll-like receptor mediated immune response and susceptibility to Aeromonas salmonicida infection in rainbow trout Oncorhynchus mykiss”. Aquaculture. 2022, 561, 738668. [Google Scholar] [CrossRef]
- Hasan, K.N.; Banerjee, G. “Recent studies on probiotics as beneficial mediator in aquaculture: a review”. J. Basic Appl. Zool. 2020, 81, 53. [Google Scholar] [CrossRef]
- Yimin; Kohanawa, M. “A regulatory effect of the balance between TNF-alpha and IL-6 in the granulomatous and inflammatory response to Rhodococcus aurantiacus infection in mice.” J. Immunol. Baltim. Md 2006, 177, 642–650.
- Deichaite; Sears, T. J.; Sutton, L.; Rebibo, D.; Morgan, K.; Nelson, T.; Rose, B.; Tamayo, P.; Ferrara, N.; Asimakopoulos, F.; Carter, H. “Differential regulation of TNFα and IL-6 expression contributes to immune evasion in prostate cancer.” J. Transl. Med. 2022, 20, 527.
- Ahmadi-Noorbakhsh, S.; Ardakani, E.M.; Sadighi, J.; Aldavood, S.J.; Abbasi, M.F.; Farzad-Mohajeri, S.; Ghasemi, A.; Sharif-Paghaleh, E.; Hatami, Z.; Nikravanfard, N.; Gooshki, E.S. “Guideline for the Care and Use of Laboratory Animals in Iran”. Lab Anim. 2021, 50, 303–305. [Google Scholar] [CrossRef]
Figure 1.
Different growth factor of rainbow trout in treated and control groups a: Food conversion ratio, b: Specific growth rate, c: condition factor, d: length and e: weight. A: (Pediococcus sp P15, L. lactis ml3, W. cibaria ml6 at dosage (300 mg/kg)), B: (Pediococcus sp P15, L. lactis ml3, W. cibaria ml6 at dosage (500 mg/kg)), C: ( Pediococcus sp P15, L. lactis ml3, W. cibaria ml6, E. faecium E10 and L. curvatus 13A at dosage (300 mg/kg)); D: (Pediococcus sp P15, L. lactis ml3, W. cibaria ml6, E. faecium E10 and L. curvatus 13A at dosage (500 mg/kg)); E: (a commercial probiotic (E) at 300 mg/kg); F: (control group without probiotics).
Figure 1.
Different growth factor of rainbow trout in treated and control groups a: Food conversion ratio, b: Specific growth rate, c: condition factor, d: length and e: weight. A: (Pediococcus sp P15, L. lactis ml3, W. cibaria ml6 at dosage (300 mg/kg)), B: (Pediococcus sp P15, L. lactis ml3, W. cibaria ml6 at dosage (500 mg/kg)), C: ( Pediococcus sp P15, L. lactis ml3, W. cibaria ml6, E. faecium E10 and L. curvatus 13A at dosage (300 mg/kg)); D: (Pediococcus sp P15, L. lactis ml3, W. cibaria ml6, E. faecium E10 and L. curvatus 13A at dosage (500 mg/kg)); E: (a commercial probiotic (E) at 300 mg/kg); F: (control group without probiotics).
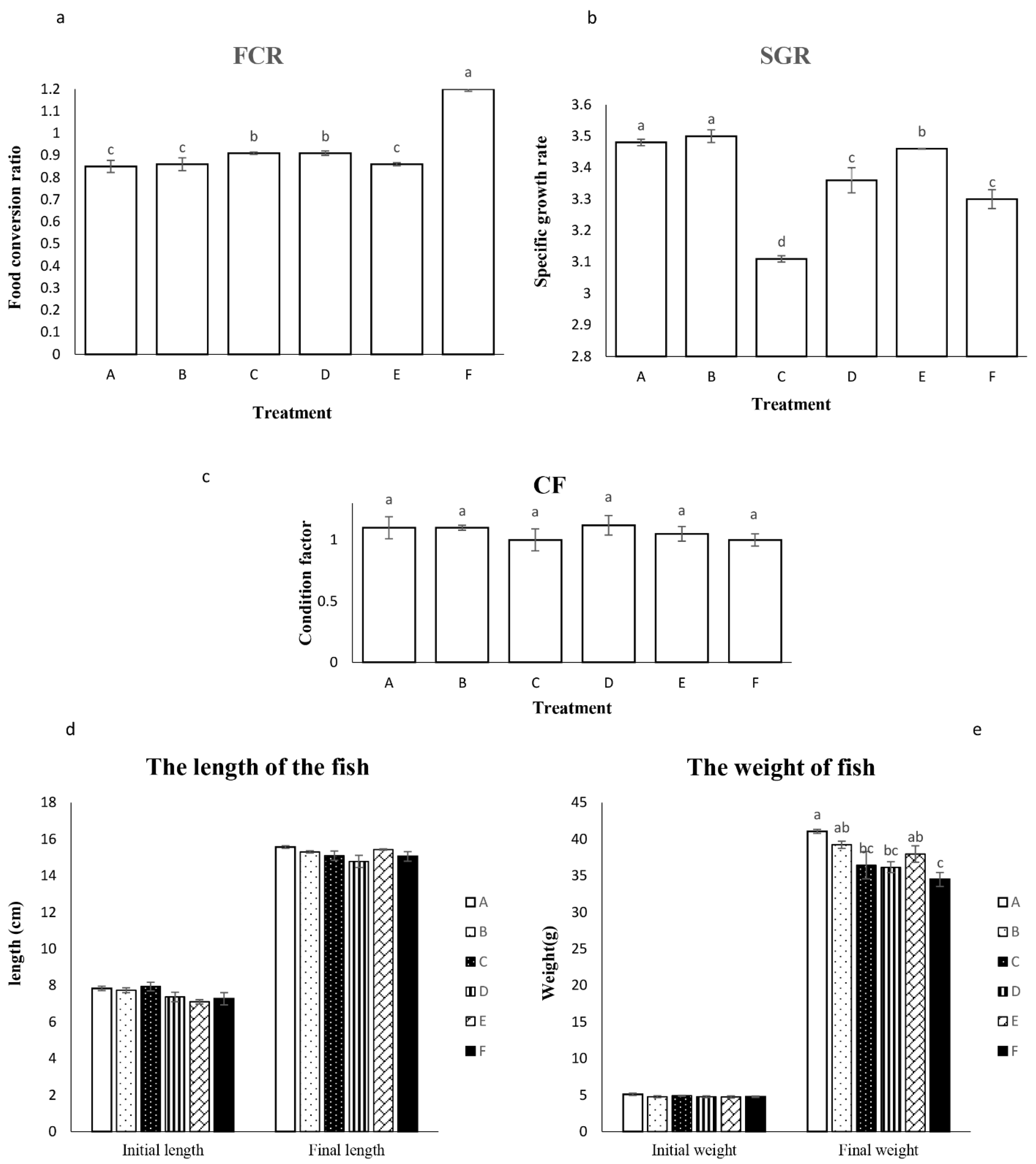
Figure 2.
Hematological parameter of rainbow trout in treated and control groups; a: Red blood cell, b: mean corpuscular hemoglobin, c: Mean Corpuscular Volume, d: mean corpuscular hemoglobin concentration, e: Hematocrit test, f: Hemoglobin test, g: White Blood Count, h: lymphocyte, i:Neutrophil, j:Eosinophil and k: Monocyte. A: (Pediococcus sp P15, L. lactis ml3, W. cibaria ml6 at dosage (300 mg/kg)), B: (Pediococcus sp P15, L. lactis ml3, W. cibaria ml6 at dosage (500 mg/kg)), C: ( Pediococcus sp P15, L. lactis ml3, W. cibaria ml6, E. faecium E10 and L. curvatus 13A at dosage (300 mg/kg)); D: (Pediococcus sp P15, L. lactis ml3, W. cibaria ml6, E. faecium E10 and L. curvatus 13A at dosage (500 mg/kg)); E: (a commercial probiotic (E) at 300 mg/kg); F: (control group without probiotics). Different letters in columns of each plot indicate a statistical difference in mean according to Duncan test (p < 0.05).
Figure 2.
Hematological parameter of rainbow trout in treated and control groups; a: Red blood cell, b: mean corpuscular hemoglobin, c: Mean Corpuscular Volume, d: mean corpuscular hemoglobin concentration, e: Hematocrit test, f: Hemoglobin test, g: White Blood Count, h: lymphocyte, i:Neutrophil, j:Eosinophil and k: Monocyte. A: (Pediococcus sp P15, L. lactis ml3, W. cibaria ml6 at dosage (300 mg/kg)), B: (Pediococcus sp P15, L. lactis ml3, W. cibaria ml6 at dosage (500 mg/kg)), C: ( Pediococcus sp P15, L. lactis ml3, W. cibaria ml6, E. faecium E10 and L. curvatus 13A at dosage (300 mg/kg)); D: (Pediococcus sp P15, L. lactis ml3, W. cibaria ml6, E. faecium E10 and L. curvatus 13A at dosage (500 mg/kg)); E: (a commercial probiotic (E) at 300 mg/kg); F: (control group without probiotics). Different letters in columns of each plot indicate a statistical difference in mean according to Duncan test (p < 0.05).
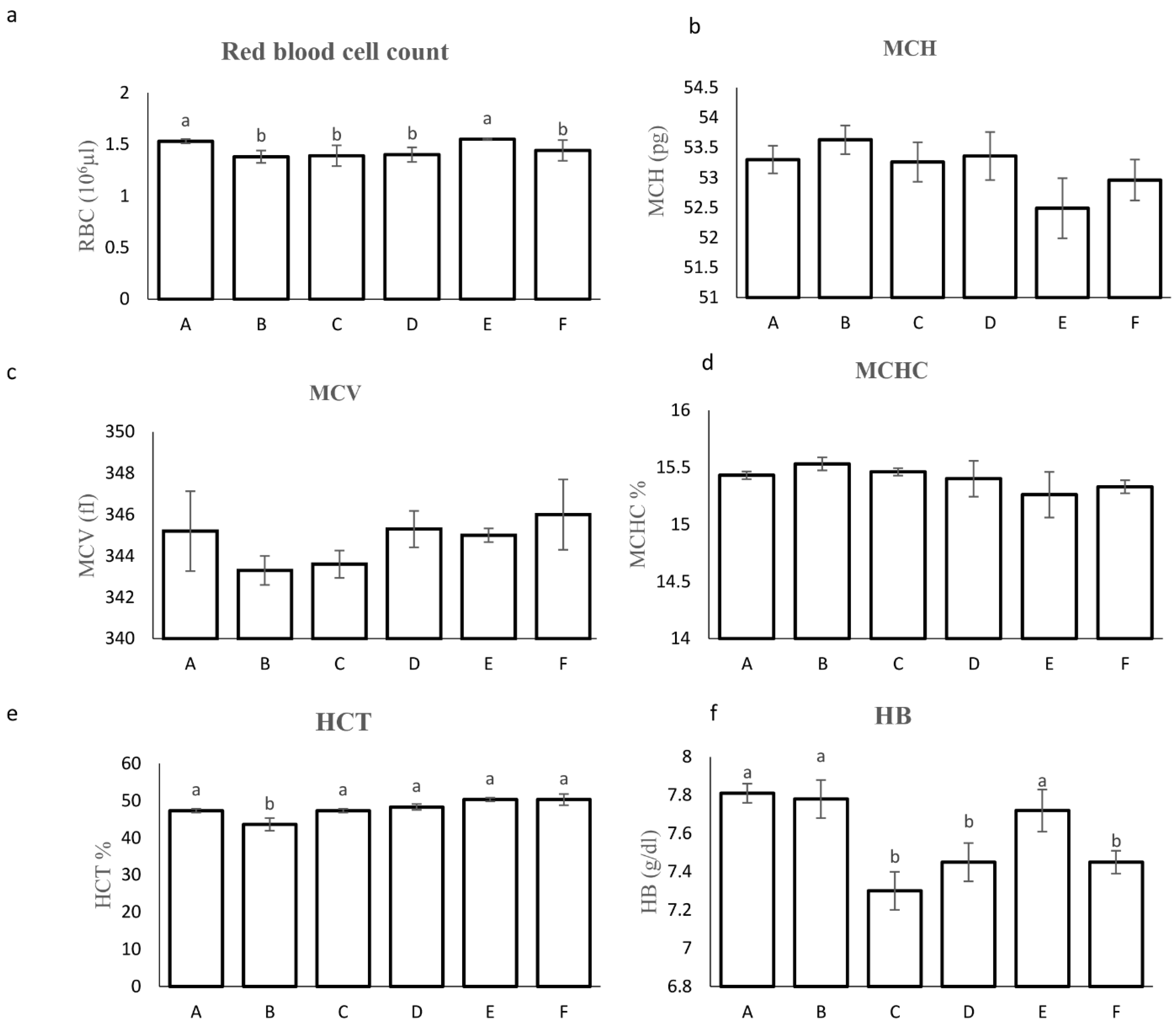
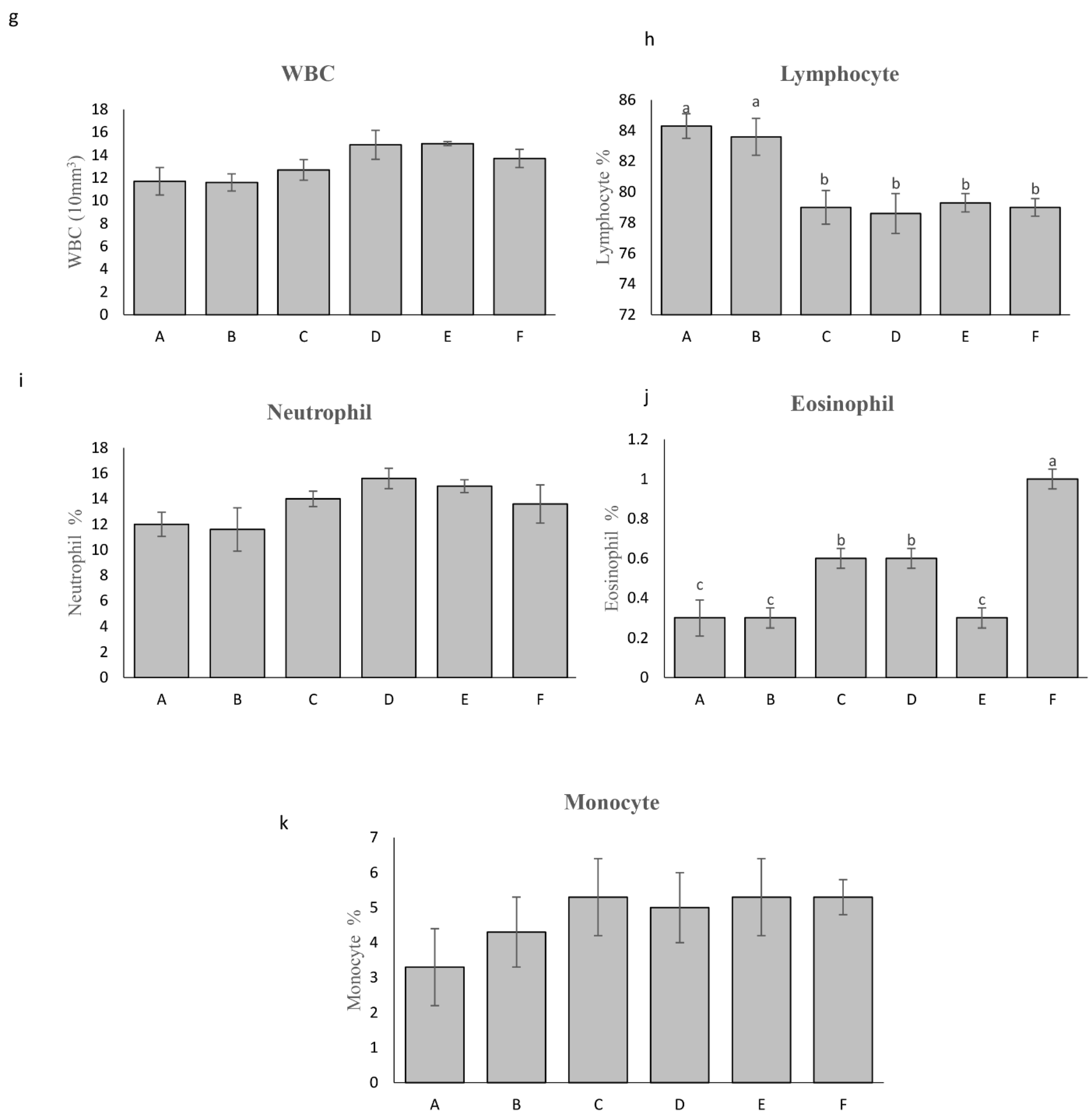
Figure 3.
Biochemical and immune factors in different dietary treatments a: Immunoglobulin M, b: Lysozyme, c: Lactate dehydrogenase, d: Alanine transaminase, e: Aspartate aminotransferase, f: Alkaline phosphatase, g: Total protein and h: Total glucose. A: (Pediococcus sp P15, L. lactis ml3, W. cibaria ml6 at dosage (300 mg/kg)), B: (Pediococcus sp P15, L. lactis ml3, W. cibaria ml6 at dosage (500 mg/kg)), C: ( Pediococcus sp P15, L. lactis ml3, W. cibaria ml6, E. faecium E10 and L. curvatus 13A at dosage (300 mg/kg)); D: (Pediococcus sp P15, L. lactis ml3, W. cibaria ml6, E. faecium E10 and L. curvatus 13A at dosage (500 mg/kg)); E: (a commercial probiotic (E) at 300 mg/kg); F: (control group without probiotics). Different letters in columns of each plot indicate a statistical difference in mean according to Duncan test (p < 0.05).
Figure 3.
Biochemical and immune factors in different dietary treatments a: Immunoglobulin M, b: Lysozyme, c: Lactate dehydrogenase, d: Alanine transaminase, e: Aspartate aminotransferase, f: Alkaline phosphatase, g: Total protein and h: Total glucose. A: (Pediococcus sp P15, L. lactis ml3, W. cibaria ml6 at dosage (300 mg/kg)), B: (Pediococcus sp P15, L. lactis ml3, W. cibaria ml6 at dosage (500 mg/kg)), C: ( Pediococcus sp P15, L. lactis ml3, W. cibaria ml6, E. faecium E10 and L. curvatus 13A at dosage (300 mg/kg)); D: (Pediococcus sp P15, L. lactis ml3, W. cibaria ml6, E. faecium E10 and L. curvatus 13A at dosage (500 mg/kg)); E: (a commercial probiotic (E) at 300 mg/kg); F: (control group without probiotics). Different letters in columns of each plot indicate a statistical difference in mean according to Duncan test (p < 0.05).
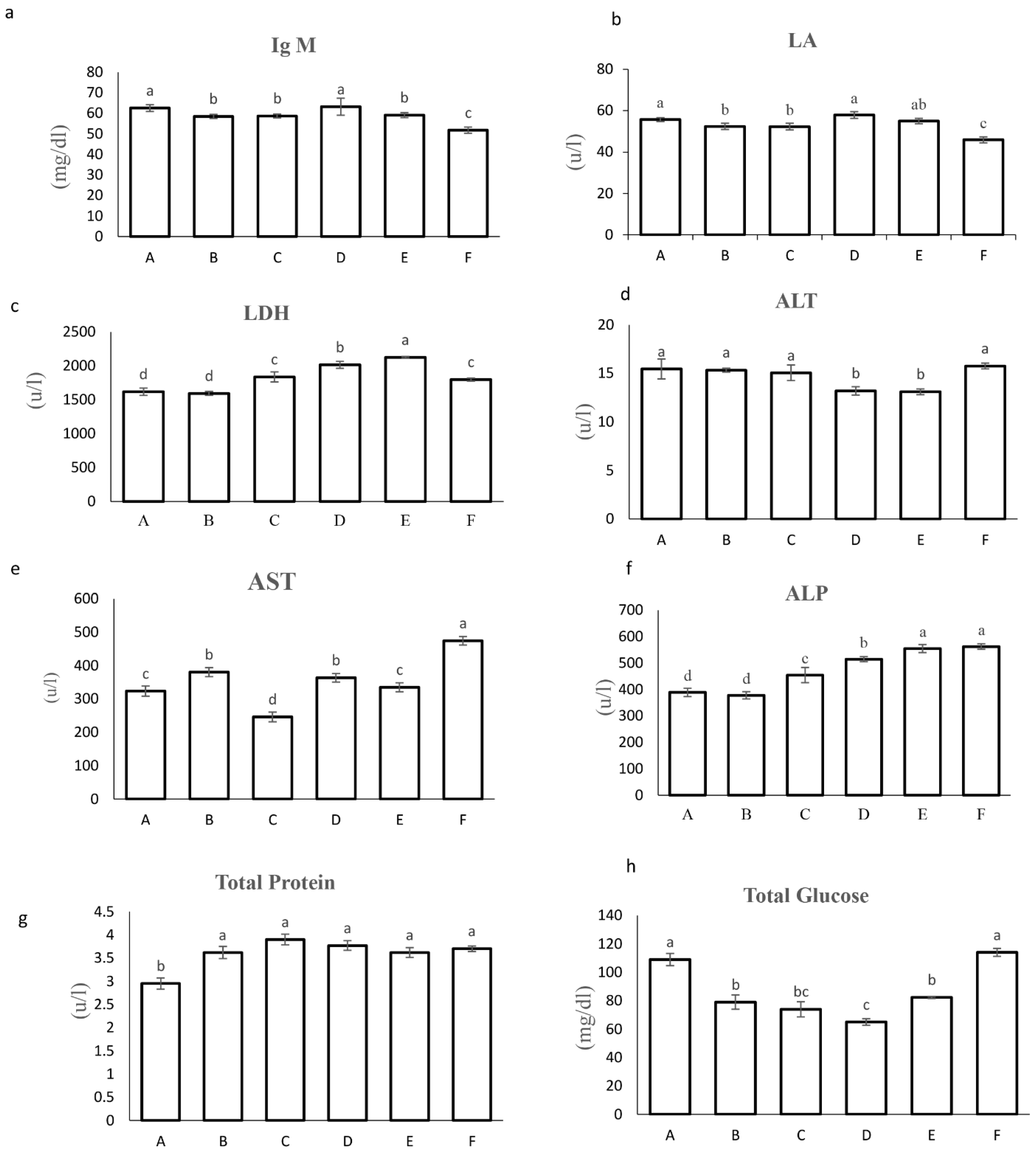
Figure 4.
Intestinal bacterial measurement a: measurement of lactic acid bacteria and b: number of Heterotrophic bacteria. 15, 30, 45, and 60 days following the probiotics administration, and 75 days (15 days post the discontinuation of the probiotics). Different letters in columns of each plot indicate a statistical difference in mean according to Duncan test (p < 0.05).
Figure 4.
Intestinal bacterial measurement a: measurement of lactic acid bacteria and b: number of Heterotrophic bacteria. 15, 30, 45, and 60 days following the probiotics administration, and 75 days (15 days post the discontinuation of the probiotics). Different letters in columns of each plot indicate a statistical difference in mean according to Duncan test (p < 0.05).
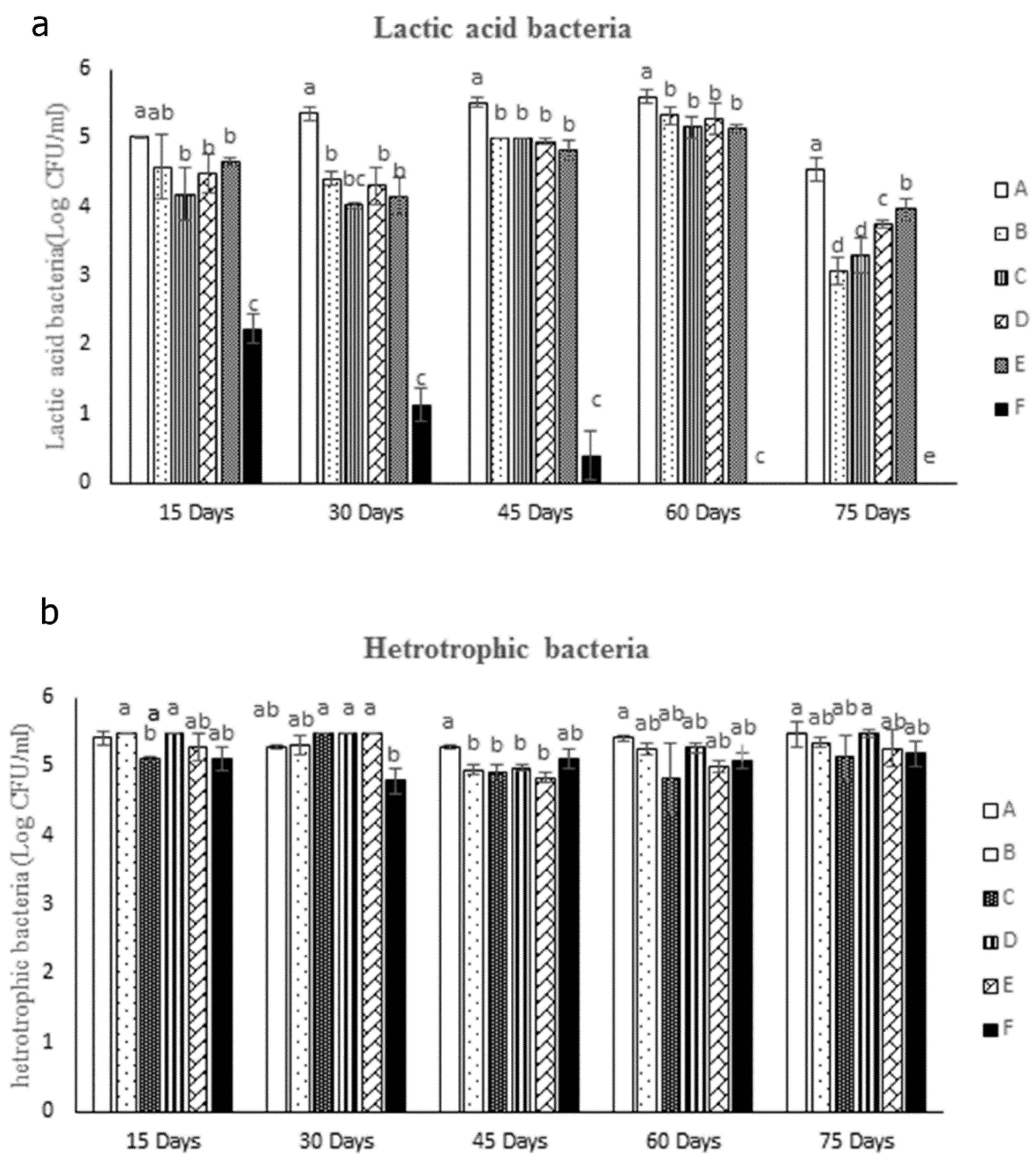
Figure 5.
Relative mRNA expression of immune-related genes, a: IL-6, b: TNF-α, c: INF-γ and d: IgT. A: (Pediococcus sp P15, L. lactis ml3, W. cibaria ml6 at dosage (300 mg/kg)), F: (control group without probiotics). Different letters in columns of each plot indicate a statistical difference in mean according to Duncan test (p < 0.05).
Figure 5.
Relative mRNA expression of immune-related genes, a: IL-6, b: TNF-α, c: INF-γ and d: IgT. A: (Pediococcus sp P15, L. lactis ml3, W. cibaria ml6 at dosage (300 mg/kg)), F: (control group without probiotics). Different letters in columns of each plot indicate a statistical difference in mean according to Duncan test (p < 0.05).
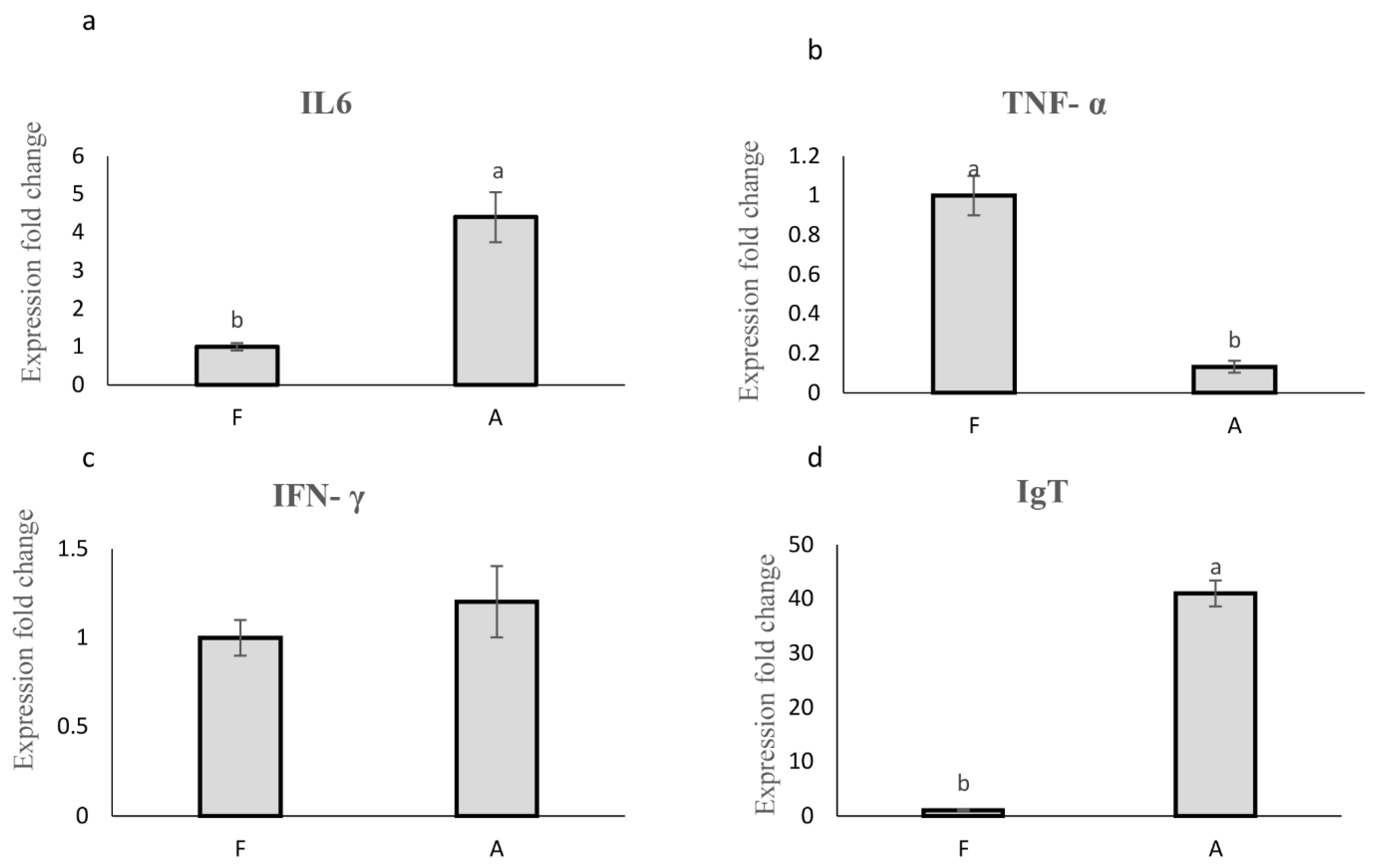

Table 1.
Pathogenic bacteria used in antagonistic assay.
| Pathogens | ATCC |
| Staphylococcus aureus | ATCC 25923 |
| Bacillus. Cereus | ATCC 29213 |
| Yersinia ruckeri | PTCC 1888 |
| Listeria Monocytogens | ATCC 13932 |
| Pseudomonas aeruginosa | ATCC27853 |
| Escherichia coli (E. coli) | ATCC 25922 |
| Candida albicans | ATCC 10231 |
| Enterococcus faecalis | ATCC 29219 |
Table 2.
The primers used in this study.
| gene | Accession number | Primer sequence | Tm | Product length in mRNA | Product length in DNA |
| IL6 | NM_001124657.1 | CGCTCGTGGTGTTAGTTAAGGG CGGGCTTCTGAAACTCCTCC |
60 60 |
201 | 858 |
| TNF |
NM_001124357 | TTATGTGCGGCAGCAGCC CCGTCATCCTTTCTCCACTGC |
61 60 |
221 | 843 |
| Ig T |
AY870265 | GTACTCTGACCATAGACCAGACA TCCTTCTTGGTGTCTTCCTC |
57 55 |
169 | 490 |
| INF |
NM_001160503.1 | TACCTGAGCTGAGGACACA TCCTGCGGTTGTCCTTCT |
57 58 |
153 | 1600 |
| Beta actin |
EZ908974 | CCTCAACCCCAAAGCCAACA CGGAGTCCATGACGATACC |
60 57 |
141 | 694 |
IL6, Interleukin 6; TNF, Tumour Necrosis Factor alpha; IgT, immunoglobulin; INF, Interferon gamma.
Table 3.
The putative probiotic bacterial strains identified in the final stage of the study.
| Tests | 5A | P3 | E10 | P20 | P37 | ml3 | ml6 | P15 | P12 | 13A | ml4 | m6 | P8 |
| Catalase | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Gram | + | + | + | + | + | + | + | + | + | + | + | + | + |
| Morphology | cocci | cocci | cocci | cocci | cocci | cocci | bacilli | cocci | cocci | bacilli | bacilli | bacilli | cocci |
| Hemolytic | - | - | - | - | - | - | - | - | - | - | - | - | - |
| pH3 time 0 | 2×107 | 2×107 | 2×107 | 2×107 | 2×107 | 2×107 | 2×107 | 2×107 | 2×107 | 2×107 | 2×107 | 2×107 | 2×107 |
| pH3 time 3 | 2×105 | 2×106 | 3×106 | 5×106 | 9×106 | 1×107 | 7.6×106 | 2×107 | 1.6×107 | 4×106 | 2×105 | 6×106 | 1.2×106 |
| 0.3% bile | ≥70 | ≥70 | ≥70 | ≥70 | ≥70 | ≥70 | ≥70 | ≥70 | ≥70 | ≥70 | ≥70 | ≥70 | ≥70 |
| Attachment | weak | good | weak | Very good |
Very good |
good | good | Very good |
good | good | weak | weak | good |
The antibacterial activity of the isolated strains against pathogens, is shown in Table 4.
Table 4.
The antibacterial activity of isolated strains against various bacteria.
| Strains | S. aureus | B. cereus | L. monocytogens | S. enterica | E. coli | En. faecalis |
P. aeruginosa |
Y. ruckeri |
C. albicans |
|---|---|---|---|---|---|---|---|---|---|
| P37 | >2 | >2 | >2 | >2 | >2 | >2 | >2 | - | >2 |
| P20 | >2 | >2 | >2 | - | >2 | >2 | >2 | >2 | - |
| 10A | >2 | >2 | >2 | >2 | - | - | - | >2 | - |
| 13A | - | - | >2 | - | - | >2 | >2 | - | - |
| 5A | - | - | >2 | >2 | - | >2 | - | >2 | - |
| P3 | >2 | >2 | >2 | - | - | >2 | - | - | - |
| ml6 | >2 | >2 | >2 | >2 | >2 | >2 | >2 | >2 | - |
| m6 | >2 | >2 | >2 | >2 | >2 | >2 | >2 | >2 | >2 |
| ml3 | >2 | >2 | >2 | >2 | >2 | >2 | >2 | >2 | - |
| P15 | >2 | >2 | >2 | >2 | >2 | >2 | >2 | >2 | >2 |
| P12 | >2 | >2 | >2 | - | - | >2 | - | >2 | - |
| P8 | >2 | >2 | >2 | - | - | >2 | - | >2 | - |
| ml4 | - | - | >2 | - | - | >2 | >2 | - | - |
Each number is the average of 3 replications. Values are in millimeters.
Table 5.
The antibiogram test result for putative probiotic bacterial strain.
| Antibiotics | Abbreviated name | 5A | P3 | E10 | P20 | P37 | ml3 | ml6 | P15 | P12 | 13A | ml4 | m6 | P8 |
| Clindamycin | CC | R | S | S | S | S | S | S | S | S | S | S | S | S |
| Tetracycline | TE | S | S | S | S | S | M | S | S | S | S | S | S | S |
| Ciprofloxacin | CP | S | R | R | R | R | R | R | R | R | R | R | R | R |
| Chloramphenicol | C | S | S | S | S | S | S | S | S | S | S | S | S | S |
| Penicillin | P | M | S | S | S | S | S | S | S | S | S | S | S | S |
| Erythromycin | E | R | S | S | S | S | S | S | S | S | S | S | S | S |
| Ampicillin | AM | S | S | S | S | S | S | S | S | S | S | S | S | S |
| Streptomycin | S | R | R | R | R | R | R | R | R | R | R | R | R | R |
| rifampin | RA | R | S | S | S | S | S | S | S | S | S | S | S | S |
| gentamicin | GM | S | R | R | R | R | S | S | S | R | R | R | S | R |
| fosfomycin | FO | S | R | R | R | R | R | R | R | R | R | R | R | R |
| Enrofloxacin | NFX | S | R | R | R | R | R | R | R | R | R | R | R | R |
| tylosin | TY | R | S | S | S | S | S | S | S | S | S | S | S | S |
| cholestin | CL | M | R | R | R | R | R | R | R | M | R | R | R | R |
| sultrim | SLT | M | R | R | R | R | R | R | R | R | R | R | R | R |
| Florfenicol | FF | M | S | S | S | S | S | S | S | S | S | S | S | S |
| Flumequine | FM | M | R | R | R | R | R | R | R | R | R | R | R | R |
R, Resistant; M, medium; S, sensitive.
Table 6.
Molecular identification of isolates based on partial 16S rRNA gene sequence information.
| Strain | Similarity (%) | Full Name | GenBank Accession Number |
| m6 | 100 | Weissella confusa m6 | MZ066823 |
| ml3 | 99.09 | Lactococcus lactis subsp. lactis strain ml3 | MN947246 |
| ml6 | 99.33 | Weissella cibaria strain ml6 | MN947228 |
| P12 | 99.44 | Pediococcus sp. strain P12 | MN120789 |
| P15 | 99.37 | Pediococcus sp. strain P15 | MN120790 |
| P8 | 99.51 | Pediococcus sp. strain P8 | MN093396 |
| P37 | 97.57 | Pediococcus acidilactici strain P37 | MK758075 |
| P20 | 98.71 | Pediococcus acidilactici strain P20 | MK757969 |
| E10 | 90 | Enterococcus faecium strain E10 | MK757968 |
| 13A | 99% | Lactobacillus curvatus13A | MK757915 |
Table 7.
Physicochemical indicators in tanks of different treatments.
| Indicators | A | B | C | D | E | F |
| Oxygen (mg/l) | 8.3±0.2 | 8.9±0.2 | 8.3±0.3 | 8.8±0.3 | 8.8±0.3 | 8.8±0.2 |
| pH | 7.96±0.5 | 8.05±0.6 | 7.95±0.5 | 8.02±0.7 | 7.96±0.5 | 7.91±0.7 |
| EC(μS/cm) | 521.3±7 | 519.3±8 | 498.3±9 | 504.2±7 | 494.6±8 | 498.5±6 |
| N-NO2(mg/l) | 0.15±0.08 | 0.18±0.06 | 0.2±0.05 | 0.24±0.05 | 0.2±0.04 | 0.29±0.08 |
| N-NH4(mg/l) | 0.42±0.08 | 0.49±0.04 | 0.46±0.05 | 0.48±0.03 | 0.45±0.05 | 0.44±0.08 |
| P-PO4(mg/l) | 0.194±0.006 | 0.198±0.004 | 0.190±0.008 | 0.194±0.006 | 0.190±0.009 | 0.181±0.006 |
There is no significant difference between the data.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



