
Submitted:
06 July 2024
Posted:
10 July 2024
You are already at the latest version
Abstract
Aedes aegypti and Aedes albopictus are responsible for transmitting major human arboviruses such as dengue, Zika, and Chikungunya, posing a global threat to public health. The lack of etio-logical treatments and efficient vaccines makes vector control strategies essential for reducing vector population density and interrupting pathogen transmission cycle. This study evaluated the impact of long-term pyriproxyfen exposure on the genetic structure and diversity of Ae. aegypti and Ae. albopictus mosquito populations. The study was conducted in Manaus, Amazonas, Bra-zil, where pyriproxyfen dissemination stations have been monitored since 2014 up to the present day. Double digest restriction-site associated DNA sequencing was performed, revealing that despite significant local population reductions, focal pyriproxyfen interventions have no signifi-cant impact on the population stratification of Ae. aegypti and Ae. albopictus in urban scenarios. The genetic structuring level of Ae. aegypti suggests it is more stratified and directly affected by PPF intervention, while for Ae. albopictus exhibits a more homogeneous and less structured popu-lation. The results suggest that although slight differences are observed among mosquitoes sub-populations, intervention focused on neighborhoods in a capital city is not efficient in terms of genetic structuring, indicating that larger-scale pyriproxyfen interventions should be considered for more effective urban mosquito control.
Keywords:
Aedes aegypti
; Aedes albopictus
; pyriproxyfen
; ddRADseq
; genomic surveillance
1. Introduction
Aedes aegypti and Ae. albopictus are among the most important mosquito species, responsible for transmitting major human arboviruses such as dengue, Zika, and Chikungunya, which makes them a global threat to public health [1,2]. Due to the lack of etiological treatments and efficient vaccines for those arboviruses, except for yellow fever and recently dengue, vector control strategies become essential for reducing vector population density, interrupting the pathogen transmission cycle, and reducing risks of epidemics [3,4].
Aedes spp. vectors are frequent targets of government and World Health Organization control campaigns. Although approximately 60% of investments in vector control between 2018-2030 will be in the development of new strategies and insecticides, current strategies still consist of adulticides in households and identification and elimination of breeding sites by health agents [5]. Even with the benefits of these strategies, such as drastic reduction in vector populations, the coverage rate of treated breeding sites is low, due to the biology of these vectors that skip-oviposit in inaccessible breeding sites [6].
A promising control strategy uses adult mosquitoes and dissemination stations (DSs) to self-disseminate potent larvicides, such as pyriproxyfen (PPF) [6,7]. This strategy aims to increase the rate of treated breeding sites, which impairs the life cycle of Aedes spp. and ultimately impacts arbovirus transmission [7,8,9]. Despite its effectiveness and being recommended in Brazil by the National Dengue Control Program (PNCD), there are few studies on the role of this control strategy on the structure and population genetics of these vectors [4,10].
Studies that correlate population genetics structure under the influence of insecticide use are scarce for Ae. aegypti and Ae. Albopictus [10,11]. Given the development of new DNA sequencing techniques and more robust molecular analyses, the scope of data obtained allows a more reliable understanding of consequent genetic stratification of vector populations, based on the detection of significant microvariations or single nucleotide polymorphism (SNP). Such data could be informative of how these insecticides may influence the population dynamics not only of Aedes spp. but also for other vectors relevant to public health [3,10,11,12]. Additionally, despite the large genome size of Ae. aegypti and Ae. albopictus, ddRADSeq effectively recovers nuclear genome markers, allowing us to obtain robust genomic data associated with this highly reliable technique while significantly reducing the per base sequencing cost [13,14,15].
In this study, we applied a high-throughput sequencing method to analyze the population genetic structure of Ae. aegypti and Ae. albopictus in monitored areas of Manaus, Amazonas. This approach allows us to assess the effects of PPF larvicide over a long-term exposure period, comparing control and experimental sites in an urban area. This information can contribute to control strategies planning and improve not only the genomic and epidemiological surveillance of these vectors but also the transmission scenario for the arboviruses they transmit.
2. Materials and Methods
2.1. Study Area and Mosquito Surveillance
Samples for this study were collected in Manaus, Amazonas, Brazil, a city covering approximately 11.401,092 km², with 2.063.689 inhabitants and 1.792.88 households (according to data from Brazilian Institute of Geography and Statistics – IBGE/2022). Control samples were collected in the neighborhoods of Adrianópolis (-3.09447, -60.00744) and Aleixo (-3.08840, -59.99265) – AdAl, starting in 2017 and still ongoing. Experimental samples were collected in the Glória neighborhood (-3.11948, -60.03449) during the same period. Dissemination stations were checked twice a month (Figure 1). Fifty collection points were randomly selected to set pyriproxyfen dissemination stations across the city to provide maximum coverage of the control and experimental sites. DSs were verified twice a month from August 2017 until the present year, with a pause in 2021 due to the COVID-19 pandemic. The juvenile and adult specimens of Ae. aegypti and Ae. albopictus were collected and morphologically identified. The collected samples were stored in 100% alcohol and placed in a fridge (4°C).
2.2. Sample Preparation and Sequencing
For DNA extraction, each specimen was extracted individually using the DNeasy Blood & Tissue kit (Qiagen, USA). DNA concentration and purity were assessed by fluorometric quantitation using the Thermo Fisher Scientific QuBit dsDNA HS Assay Kit. Both techniques were performed according to the manufacturer's recommendations.
DNA libraries were prepared, and samples were sequenced by double digest restriction-site associated DNA technique, in a 151-cycle single-read run and using EcoRI-MspI restriction enzymes, as described in Campos et al, 2017. Sequencing quality control analyses were performed using the FASTQC program [16] before and after reads filtering.
2.3. Species Identification
Sequencing data were aligned to Ae. aegypti and Ae. albopictus cytochrome oxidase subunit I (COI) reference sequence (available at KC913582.1 and NC_006817.1) using Burrows-Wheeler Aligner (BWA) software [17]. After alignment, individual COI consensus sequences were generated using SamTools software package [18]. BLASTn tool was used for multi-species identification using the individual COI consensus sequences [19]. Only the highest matching result from BLAST was used. Specimens were discarded if e-value > 1e-100, identities < 200, identity < 90% and the matching sequence was not identified as one of the species.
2.4. Variant Calling
Sequencing quality control was applied with mean quality filtering, trimming, minimum length filtering (80 bp), and adapter removal procedures using Trimmomatic [20]. All sequencing reads were aligned to the reference genome of Ae. aegypti (NC_035159.1) and Ae. albopictus (JAFDOQ000000000.1), both retrieved from the NCBI database by BWA program [17]. Variant calling was performed using Stacks v2.62 software [18]. with gstacks approach. The variant panel was exported in the VCF v4.2 format. SNPs were removed from the variant panel based on a minimum allele frequency (MAF) < 0.1 and at least 8 non-missing genotypes within each group.
2.5. Population and Structure Genetics Inferences
Statistical analyses were performed with PLINK 1.9 [21], and graphs and figures were generated using the GGPlot2 package for R in RStudio [22]. Pairwise FST, nucleotide diversity (θS and θπ) and Tajima's D were estimated using ARLEQUIN 3.5 [23]. The principal component analysis (PCA) was calculated based on pairwise genetic distance IBS (Identity-by-state matrix), while hierarchical clustering analysis was performed using pvclust v2.2-0 package for R [24].
3. Results
3.1. Data Collection and DNA Extraction
During the monthly checks from 2017 to 2020, which were interrupted due to the COVID-19 pandemic, 55,472 specimens of Ae. aegypti and 19,720 specimens of Ae. albopictus among juveniles and adults were collected. A total of 102 individuals were submitted to ddRAD sequencing, followed by species identification: 52 individuals from the control neighborhood, where PPF was not applied and 50 individuals from the experimental neighborhood, considering both vector species (Table 1).
3.2. Double Digest Restriction-Site Associated DNA Sequencing (ddRADseq) Performance
For Ae. aegypti, a total of 4,525,210 reads were properly mapped to the reference genome. The average sequencing depth was approximately 0.0032 ± 0.0583. After variant calling, the final dataset consisted of 1.443 SNPs distributed throughout the genome. The SNP average sequencing depth and quality (Phred Score) were 1.64 ± 1.79 and 23.11 ± 4.69, respectively. The SNP density was 0.00121 SNP/Kbp.
For Ae. albopictus, a total of 3.522.816 reads were properly mapped to the reference genome. The average sequencing depth was approximately 0.0026 ± 0.0037. After variant calling, the final dataset consisted of 645 SNPs distributed throughout the genome. The SNP average sequencing depth and quality (Phred Score) were 1.65 ± 1.29 and 23.12 ± 4.95, respectively. The SNP density was 0.00125 SNP/Kbp.
3.3. Statistical and Structural Analysis
For Ae. aegypti, significant Theta(Pi) molecular diversity estimates were observed for control and experimental groups (0.4 and 0.369, respectively). The estimates of Tajima's D of control and experimental groups were respectively 1.41 and 1.16 (p-value < 0.00454). Based on FIT estimate, no significant differences were observed for homozygosity (F ~0.84, p-value < 0.87).
The median pairwise IBS distance between samples within groups showed significant differences between control and experimental groups, 0.109 and 0.0631 (p-value < 2.2e-16) respectively. Significant stratification was observed between control and experimental groups, with a pairwise FST estimate of 0.015 (p-value < 0.037, number of permutations = 1000). Significant stratified sites were observed based on 70 SNPs (number of permutations = 1000).
PCA analysis showed SNP clustering for control and experimental groups (Figure 2A). The clustering is mainly represented by the first principal component which corresponds to the major explained variance (19,8%). The association test between control and experimental groups presented two significant SNPs (Figure 2B), adjacent to the gene region of "neuropeptide CCHamide-2 receptor". The SNPs were found at the positions (NC_035109.1:96061807:A:G) e (NC_035109.1:96061808:A:G).
For Ae. albopictus, Theta(Pi) molecular diversity estimates were 0.366 for both groups, showing that there is no significant molecular diversity between the control and experimental groups. The estimates of Tajima's D of control and experimental groups were 1.01 and 1.03 (p-value < 0.3518). Based on FIT estimate, significant differences were observed for homozygosity (control = 0.794, experimental = 0.824, p-value = 0.049). The median pairwise IBS distance between samples within groups was 0.113 for both groups, showing no significant difference between control and experimental groups (p-value = 0.4777). No significant stratification was observed between the control and experimental groups, with a pairwise FST estimate of 0.0013 (p-value < 0.409, number of permutations = 1000).
PCA analysis showed clustering for the SNP genotype matrix, as the figure below (Figure 3). The clustering is mainly represented by the first principal component which corresponds to the major explained variance by the model (21,0%).
4. Discussion
When we assessed the molecular diversity based on SNP typing for Ae. aegypti, a slight but significant difference was observed between the control and experimental groups, with approximately 7.8% nucleotide variation. This diversity was greater in the control group, which can be attributed to the absence of PPF intervention, which in turn could have resulted in no selective pressure on the local population and also a continuous migration flow from adjacent populations. It is noteworthy that the locations in this study correspond to neighborhoods in Manaus, making PPF intervention a local and/or focal strategy. A statistically significant difference was also observed between groups based on Tajima's D estimate, indicating that Ae. aegypti populations in both localities are in a state of equilibrium/neutrality, corroborating nucleotide diversity data.
On the other hand, no significant differences in homozygosity were observed comparing individuals across the entire population (Fit estimate). IBS estimate within groups was statistically significant, revealing that in the control group locality, there is a smaller genetic distance, as well as a greater genetic divergence between individuals from two localities. The average FST value for Ae. aegypti was significant and considerably high, even among populations sharing the same urban space. Additionally, the detection of 70 highly stratified and informative SNPs for clustering highlights a discrete pattern of stratification when comparing control and experimental groups. Despite the migratory flow of populations adjacent to these two groups, it is likely that distinct subpopulations exist in the analyzed localities.
PCA analysis (Figure 2A) demonstrated discrete clusters between control and experimental groups, but the most interesting data observed was the difference in dispersion of individuals within each group. The control group (AdAl) showed lower genetic distance among Ae. aegypti individuals compared to the experimental group (Glória), suggesting greater variability among individuals in intervention site. The closer the points, the greater genetic similarity between individuals was observed, suggesting genetic similarity and reduced genetic diversity within the experimental group. By analyzing dispersion, PC1 axis showed greater significance, explaining 19.8% of observed variations between the two groups. Although there is clustering in the control population, individuals are more dispersed, suggesting higher diversity. This finding corroborates the data from FST analysis, which, despite not showing a clear and abrupt clustering pattern, allows the identification of two groups with distinct nucleotide diversity.
The higher diversity of the control group can be attributed to two possible factors: 1- Control group has no contact with the PPF intervention, resulting in a high and constant population density between this locality and adjacent populations. Thus, even with migration flow, there is a balance in population density between control and neighboring populations. 2- In the experimental group where the intervention occurs, several individuals are locally eliminated while adjacent populations continue to migrate constantly to this site, causing a significantly greater impact on population dynamics with a reduction of the population.
The association test based on allelic frequency (Figure 2B) revealed that among 70 highly stratified SNPs, two SNPs are clearly highlighted, showing different fixation levels being found exclusively in individuals from the control population (AdAl). Those SNPs are not located inside gene regions, but rather in a region adjacent to neuropeptide receptor CCHamide-2. Recent studies have shown that this neuropeptide plays a fundamental role in the endocrine system of various arthropod species, such as Drosophila melanogaster, Diaphorina citri, and Acyrthosiphon pisum [25,26,27,28]. CCHa2-R allows endocrine cells in the midgut and peripheral adipose tissue to communicate with the central nervous system to ensure regulation of feeding, insulin production, lipid metabolism, development and growth, energy maintenance, and formation of appetitive and associative odor-sugar memory [29,30,31].
It is important to emphasize that although there are currently no studies of CCHa2-R neuropeptide in Aedes spp. mosquitoes, but given its function, the long-term exposure of these vectors to insecticides may influence the regulation of physiological processes related to sugar intake, as well as metabolic alterations that affect insecticide efficacy. Further investigations are necessary, which may suggest the development of control strategies using this receptor as a potential target. These data indicate that intervention significantly impacts population dynamics, leading to distinct subpopulations within both control and experimental groups of Ae. aegypti. Long-term exposure to PPF by the experimental group results in a bottleneck, highlighted by lower nucleotide diversity and higher average genetic distance between individuals, suggesting a drastic reduction in population density and incorporation of new individuals from adjacent populations. Besides that, PCA analysis suggests that the migration of new individuals into these locations contributes to different levels of diversity without changing overall population genetic neutrality. Although the control and experimental groups are not entirely distinct in terms of stratification, they represent populations with different and significant parameters of diversity and genetic dispersion, reflecting the effects observed in PPF intervention.
When analyzing Ae. albopictus populations, statistically significant differences were observed based on homozygosity for F statistics (Fit estimate). Despite being subtle, these differences indicate that the level of homozygosity is higher in individuals from the experimental group, suggesting that they are genetically similar (Figure 3). By comparing the two species, the level of homozygosity between Ae. aegypti and Ae. albopictus was quite similar. Although the average value of the fixation index (FST estimate) was 10 times lower for Ae. albopictus, this data was not statistically significant in the permutation test. No stratification between the two locations was observed, indicating that individuals in both groups are genetically similar regardless of the presence of PPF, suggesting the occurrence of a single population, characterizing a scenario of panmixia.
No significant differences were observed for Ae. albopictus based on molecular diversity, neutrality tests and also for IBS estimate within the same group, regardless of intervention status. Stratified SNP analyses and association tests were not conducted as no stratification was observed between the control and intervention populations. It is noteworthy that continuous migration flow from adjacent populations also occurs, as experimental design and study sites correspond to those analyzed for Ae. aegypti.
The scenario observed for Ae. albopictus contrasts with the population dynamics observed for Ae. aegypti. The control and experimental populations showed minor differences based on the analyzed genetic parameters, indicating a homogeneous population in both locations. These results reflect the biology of the vector itself, as Ae. albopictus is less domesticated and anthropophilic and, unlike Ae. aegypti, does not depend on high urban concentration to thrive[32,33]. Although being found in urban and peri-urban environments, the mosquito commonly inhabits forested and frontier areas, vegetations and plantations, primarily colonizing rural and wild environments, indicating a less defined population structure in urban settings compared to Ae. Aegypti [1,34,35]. Another important characteristic is flight dispersal: while Ae. aegypti has more limited flight dispersal (range of 100m-200m), Ae. albopictus has greater dispersal capacity, tending to fly longer distances (between 400-600m) which contributes to its geographic spread [36,37,38].
The results obtained so far indicate that focal intervention has a small or no impact on Ae. albopictus population found in urban areas in Manaus. Additionally, there are few studies regarding population dynamics of this vector in Brazil, due to its less importance as a local vector of arboviruses. Overall, focal intervention with PPF locally reduces the abundance of both vectors; however, in highly urbanized areas such as Manaus, vector population density is much higher, with a continuous migration flow among neighborhoods. Additionally, despite promoting a drastically reduced abundance of juvenile and adult mosquitoes, this intervention model presents challenges. As the selected neighborhoods designated as control and experimental groups are subjected to long-term intervention (since 2014) and at the same time, are separated by a short geographical distance (4.256 km apart in a straight line), observing significant genetic clustering on a microgeographic scale might be cumbersome.
Nevertheless, the population structure of Ae. aegypti correlates with its domestication and urbanization capacity, suggesting that it is more genetically structured and directly affected by PPF intervention. Ae. albopictus populations, however, colonize urban areas less frequently, which results in more homogeneous and less structured populations. Thus, PPF intervention in this model may not have the expected effect on genetic structure, besides interfering with clearer insights into the population dynamics of these vectors, as focal intervention does not affect neighboring populations that can rapidly colonize treated sites.
This study also provides insights on how sequencing can be a useful and valuable tool for assessing different intervention strategies, not only verifying evident reduction in population density and consequently transmission reduction of dengue and other arboviruses, but also how intervention affects genetic structuring and dynamics of vector populations. The data obtained so far is crucial regarding genomic and epidemiological surveillance, directly improving the planning of control activities and epidemiological surveillance of these vectors. Further investigations are needed to understand migration patterns, intervention on a larger geographic scale in urban settings, physical barriers and isolation of neighborhoods, as this is the first study associating Aedes spp. with PPF intervention on a microgeographic scale within an extensive urban area.
5. Conclusions
Our results indicate that focal intervention application has no significant impact on the population stratification of Ae. aegypti and Ae. albopictus in urban scenarios. In contrast, both species experience a drastic reduction in local populations, and the structuring level of Ae. aegypti suggests it is more genetically stratified and directly affected by PPF intervention. Meanwhile, Ae. albopictus, a more homogeneous and less structured population is observed, where focal intervention strategy did not lead to significant changes in terms of population genetics. Although slight differences are observed among Ae. aegypti subpopulations, it becomes clear that intervention based in this model focused on neighborhoods in a capital city, is not as efficient in terms of structuring as expected, indicating that intervention strategies with PPF on larger scales or other alternatives should be considered for more effective control of these vectors in urban environments.
Author Contributions
LFOL contributed to the conceptualization, formal analysis, investigation, methodology, validation, visualization, and writing the original draft preparation. MVNA contributed to the formal analysis, software, validation and visualization. JJCC contributed to the data curation, investigation and visualization. DPA contributed to the supervision and visualization. PEMR contributed to the project administration, resources and supervision. SLBL contributed to the conceptualization, data curation, funding acquisition, project administration and resources. All authors actively contributed to the writing – review and editing of this work. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Leônidas and Maria Deane Institute (Fiocruz Amazônia) in association with the municipal and state health departments.
Institutional Review Board Statement
Dr. Sérgio Luiz Bessa Luz is permanently licensed (27733-1) by Brazilian Institute of Environment and Natural Resources (IBAMA) to collect disease vectors. All procedures were conducted with residents' permission. Formal approval was not required for mosquito collection in urban environments. Ethical review and approval were waived for this study.
Informed Consent Statement
Not applicable.
Acknowledgments
We are grateful to Leônidas and Maria Deane Institute (Fiocruz Amazônia), Oswaldo Cruz Institute (Fiocruz) and Sao Paulo State University "Júlio de Mesquita Filho" (UNESP), who contributed to this work.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Kraemer, M.U.G; Sinka, M.E. The global distribution of the arbovirus vectors Aedes aegypti and Ae. albopictus. eLife 2015, 4:e08347. [CrossRef]
- Espinal, M.A.; Andrus, J.K. Emerging and Reemerging Aedes-Transmitted Arbovirus Infections in the Region of the Americas: Implications for Health Policy. Am J Public Health 2019, 109(3):387-392. [CrossRef]
- Evans, B.R.; Gloria-Soria, A. A multipurpose, high-throughput single-nucleotide polymorphism chip for the dengue and yellow fever mosquito, Aedes aegypti. G3 2015, 26-5(5):711-8. [CrossRef]
- BRASIL. Ministério da Saúde. Relatório da Reunião Internacional para Implementação de Alternativas para o Controle do Aedes aegypti no Brasil. Boletim Epidemiológico 2016, v. 47, n. 15:(2358-9450).
- ISGBLOBAL - Annual Report ISGlobal. Barcelona Institute for Global Health. Barcelona, 2018. Available online: https://www.isglobal.org. (accessed on 02.07.2024).
- Devine, G.J.; Zamora-Perea, E. Using adult mosquitoes to transfer insecticides to Aedes aegypti larval habitats. Proc Natl Acad Sci 2009. Jul-14;106(28):11530-4. [CrossRef]
- Garcia, K.K.S.; Versiani, H.S. Measuring mosquito control: adult-mosquito catches vs egg-trap data as endpoints of a cluster-randomized controlled trial of mosquito-disseminated pyriproxyfen. Parasit Vectors 2020, 14;13(1):352. [CrossRef]
- Abad-Franch, F.; Zamora-Perea, E. Mosquito-disseminated inseticide for citywide vector control and its potential to block arbovirus epidemics: entomological observations and modeling results from Amazonian Brazil, PLoS Med 2017, 17;14(1):e1002213. [CrossRef]
- Abad-Franch, F.; Zamora-Perea, E. Mosquito-disseminated pyriproxyfen yields high breeding-site coverage and boosts juvenile mosquito mortality at the neighborhood scale. PLoS Negl Trop Dis 2015, Apr 7;9(4):e0003702. [CrossRef]
- Leles, L.F.O. Alvarez, MVN. PLoS Negl Trop Dis 2024. DOI.
- Rahman, R.U.; Cosme, L.V. Insecticide resistance and genetic structure of Aedes aegypti populations from Rio de Janeiro State, Brazil. PLoS Negl Trop Dis. 2021, 15(2):e0008492. [CrossRef]
- Campos, M.; Conn, J.E. Microgeographical structure in the major Neotropical malaria vector Anopheles darlingi using microsatellites and SNP markers. Parasit Vectors 2017, Feb-13;10(1):76. [CrossRef]
- Matthews, B.J.; Dudchenko, O. Improved reference genome of Aedes aegypti informs arbovirus vector control. Nature 2018, 563(7732):501-507. [CrossRef]
- Boyle, J.H.; Rastas, P.M.A. A Linkage-Based Genome Assembly for the Mosquito Aedes albopictus and Identification of Chromosomal Regions Affecting Diapause. Insects 2021, 12(2):167. [CrossRef]
- Goodwin, S.; Mcpherson, J.D. Coming of age: ten years of next-generation sequencing technologies. Nature Reviews Genetics 2016, v. 17, n. 6, p. 333. [CrossRef]
- Andrews, S. FastQC: A Quality Control Tool for High Throughput Sequence Data. 2010. Available online: http://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 02.07.2024).
- Li, H. and Durbin, R. Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatics 2009. Jul-15;25(14):1754-60. [CrossRef]
- Li, H. A statistical framework for SNP calling, mutation discovery, association mapping and population genetical parameter estimation from sequencing data. Bioinformatics 2011. Nov-1;27(21):2987-93. [CrossRef]
- Altschul, S.F.; Gish, W. Basic local alignment search tool. J Mol Biol 1990. Oct-5;215(3):403-10. [CrossRef]
- Bolger, A.M.; Lohse, M. Trimmomatic: a flexible trimmer for Illumina sequence data. Bioinformatics 2014. Aug-1;30(15):2114-20. [CrossRef]
- Purcell, S.; Neale, B. PLINK: a tool set for whole-genome association and population-based linkage analyses. Am J Hum Genet 2007. Sep-81(3):559-75. [CrossRef]
- RSTUDIO TEAM. RStudio, Inc., Boston, MA., v. 42, p. 14, 2015. Available online: http://www.rstudio.com/ (accessed on 02.07.2024).
- Excoffier, L. and Lischer, H.E. Arlequin suite ver 3.5: a new series of programs to perform population genetics analyses under Linux and Windows. Mol Ecol Resour. 2010, May;10(3):564-7. [CrossRef]
- TEAM, R. Core. RA Lang Environ Stat Comput 2013, v. 55, p. 275-286, Available online: http://www.R-project.org/ (accessed on 02.07.2024).
- 2González Segarra, AJ.; Pontes, G. Hunger- and thirst-sensing neurons modulate a neuroendocrine network to coordinate sugar and water ingestion. eLife 2023, 12:RP88143. [CrossRef]
- Havula, E.; Ghazanfar, S. Genetic variation of macronutrient tolerance in Drosophila melanogaster. Nature Communications 2022, 13:1637. [CrossRef]
- Shahid, S.; Shi, Y. CCHamide2-receptor regulates feeding behavior in the pea aphid, Acyrthosiphon pisum .Science Direct 2021, Elsevier. Peptides 143. [CrossRef]
- Wang, Z.; Zhou, W. Characterization and Expression Profiling of Neuropeptides and G-Protein-Coupled Receptors (GPCRs) for Neuropeptides in the Asian Citrus Psyllid, Diaphorina citri (Hemiptera: Psyllidae). Int. J. Mol. Sci. 2018, 19, 3912. [CrossRef]
- Yamagata, N.; Imanishi, Y. Nutrient responding peptide hormone CCHamide-2 consolidates appetitive memory. Front. Behav. Neurosci. 2022, 16:986064. [CrossRef]
- Hobin, M.; Dorfman, K. The Drosophila microRNA bantam regulates excitability in adult mushroom body output neurons to promote early night sleep. iScience. Cell Press. 2022, 25, 104874. [CrossRef]
- Nagata, S.; Takei, Y. Chapter 84 - CCHamide: Handbook of Hormones, 2016, Academic Press, Pages 475-476.
- Bargielowski, I.E.; Lounibos, L.P. Evolution of resistance to satyrization through reproductive character displacement in populations of invasive dengue vectors. Proceedings of the National Academy of Sciences 2013, 110:2888–2892. [CrossRef]
- Honório, N.A.; Silva, W.D.C. Dispersal of Aedes aegypti and Aedes albopictus (Diptera: Culicidae) in an urban endemic dengue area in the State of Rio de Janeiro, Brazil. Memórias do Instituto Oswaldo Cruz 2003, 98:191–198. [CrossRef]
- Li, Y.; Kamara, F. Urbanization Increases Aedes albopictus Larval Habitats and Accelerates Mosquito Development and Survivorship. PLOS Neglected Tropical Diseases 2014, 8(11): e3301. [CrossRef]
- Consoli, R.A.G.B.; Oliveira, R.L. Principais mosquitos de importância sanitária no Brasil. Editora FIOCRUZ, Rio de Janeiro, Brasil, 1994, 228 p. Available from SciELO Books (http://books.scielo.org).
- Verdonschot, P.F.M. and Besse-Lototskaya, A.A. Flight distance of mosquitoes (Culicidae): A metadata analysis to support the management of barrier zones around rewetted and newly constructed wetlands, Limnologica 2014, Vol. 45, Pgs 69-79. [CrossRef]
- Guerra, C.A.; Reiner Jr, R.C. A global assembly of adult female mosquito mark-release-recapture data to inform the control of mosquito-borne pathogens. Parasites & Vectors 2014. [CrossRef]
- Harrington, L.C.; Scott, T.W. Dispersal of the dengue vector Aedes aegypti within and between rural communities. Am J Trop Med Hyg 2005. Feb;72(2):209-20. [PubMed]
Figure 1.
Study site: capital city of Manaus, Amazonas, Brazil. (A) Representation of collection points in AdAl neighborhood (control) and (B) Gloria neighborhood (experimental). The dots indicate the location of the ovitramps monitored for mosquito vectors.The map was created using the maps library on software QGIS.
Figure 1.
Study site: capital city of Manaus, Amazonas, Brazil. (A) Representation of collection points in AdAl neighborhood (control) and (B) Gloria neighborhood (experimental). The dots indicate the location of the ovitramps monitored for mosquito vectors.The map was created using the maps library on software QGIS.
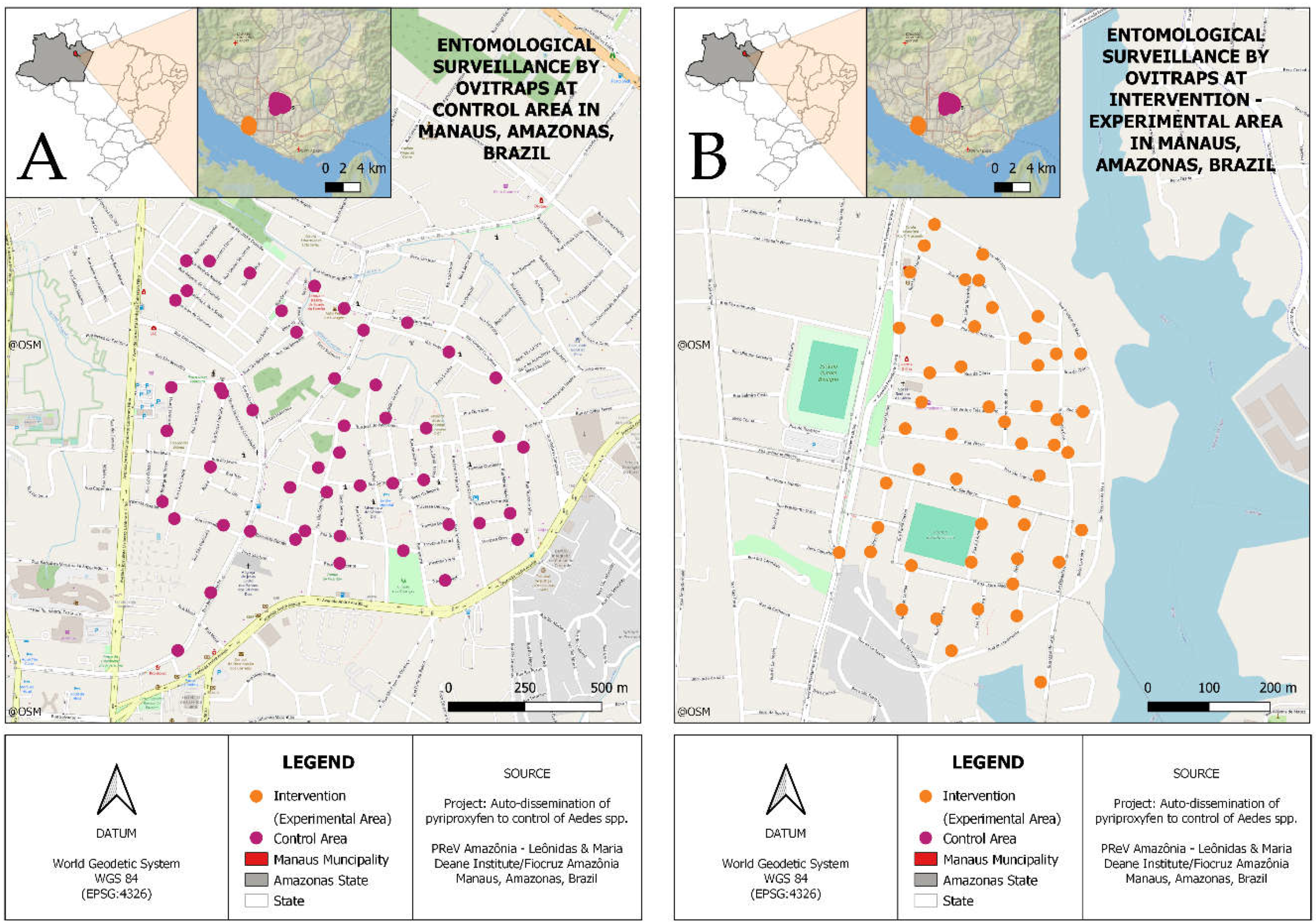
Figure 2.
(A) Principal Component Analysis Biplot for Ae. aegypti based on genotype matrix. Values between parentheses represent the variance explained by the respective principal component. (B) Manhattan plot of case/control allelic test (1Df Χ²). BONF: Bonferroni adjusted p-value.
Figure 2.
(A) Principal Component Analysis Biplot for Ae. aegypti based on genotype matrix. Values between parentheses represent the variance explained by the respective principal component. (B) Manhattan plot of case/control allelic test (1Df Χ²). BONF: Bonferroni adjusted p-value.
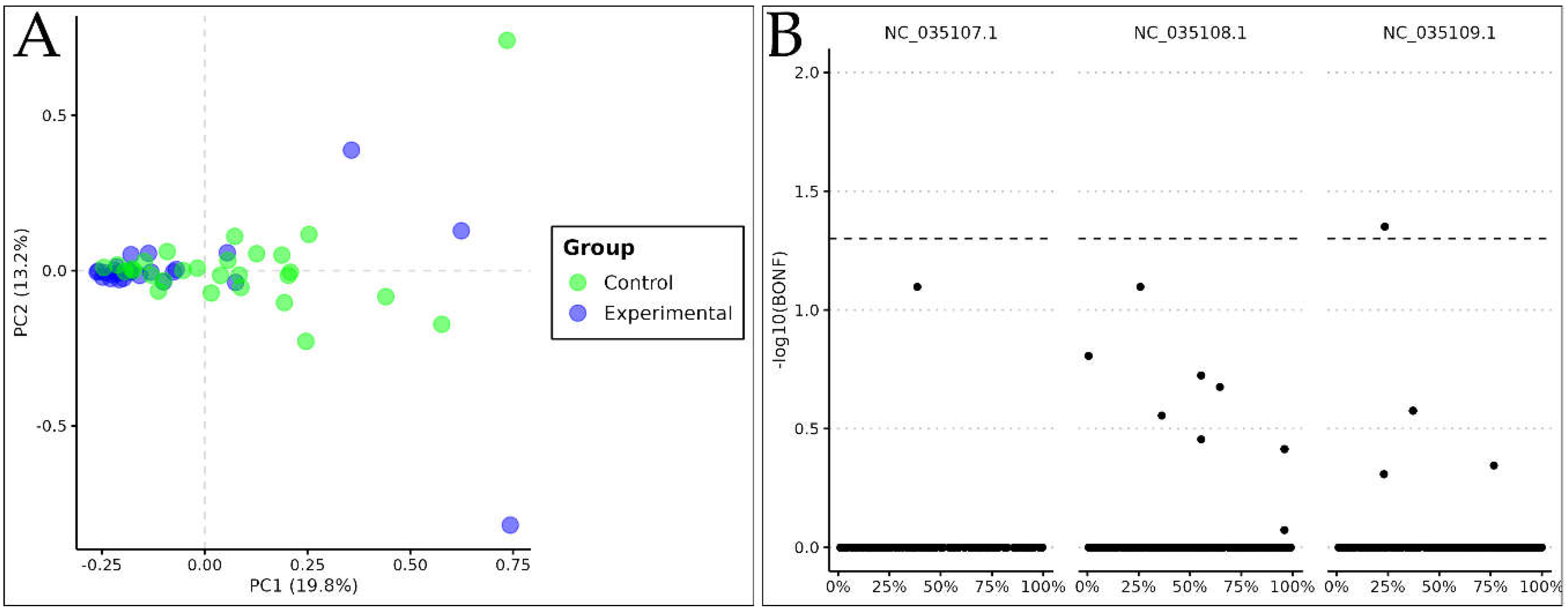
Figure 3.
Principal Component Analysis Biplot for Ae. albopictus based on genotype matrix. Values between parentheses represent the variance explained by the respective principal component.
Figure 3.
Principal Component Analysis Biplot for Ae. albopictus based on genotype matrix. Values between parentheses represent the variance explained by the respective principal component.
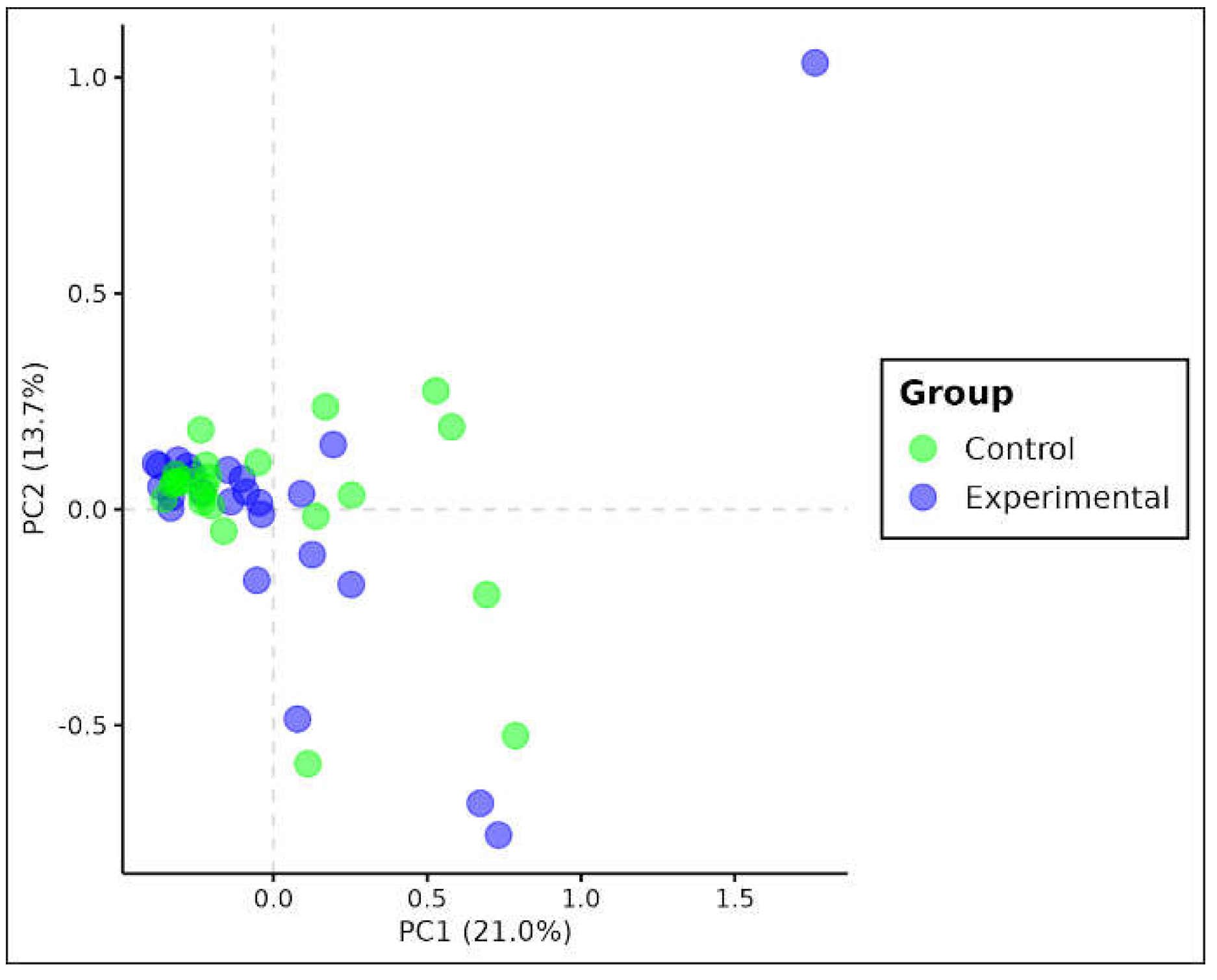

Table 1.
Individuals selected from each species for ddRAD sequencing.
| Ae. aegypti | Ae. albopictus | |||
|---|---|---|---|---|
| AdAl - Control | Total | Selected | Total | Selected |
| JAN/FEB – 2018 to 2020 | 28.311 | 28 | 16.820 | 23 |
| Glória - Experimental | Total | Selected | Total | Selected |
| JAN/FEB – 2018 to 2020 | 27.161 | 25 | 2.900 | 25 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



