
Submitted:
08 July 2024
Posted:
09 July 2024
You are already at the latest version
Abstract
The metabolites secreted by probiotics or released after their lysis are called postbiotics. They provide physiological benefits to the host, preventing the colonisation of pathogens by improving the intestinal environment for beneficial commensal bacteria, which reduces the incidence of digestive disorders and improve the immune system. The aim of this work was to evaluate the addition of postbiotics to dairy cow rations during the transition period on nutrient digestibility, composition, and milk yield. The effects of two postbiotics were evaluated in twelve Friesian cows from 30 days before calving to two months of lactation. The animals were randomly allocated to the treatments control (CT) and supplemented with postbiotics (PC and PR). Feeding was ad libitum with 60/40 of forage/concentrate ratio. Daily feed intake and milk production were recorded individually throughout the study. Two digestibility balances were performed, one before parturition and one after parturition. Colostrum was sampled at first milking and milk was sampled weekly. Data were analysed using a mixed mode in R software. The results suggest that postbiotic supplementation in late gestation and early lactation increase the voluntary intake of dry matter, especially in the PR treatment, with higher apparent total tract digestibility of dry matter, organic matter and neutral detergent fiber. Both treatments including postbiotics induced an increase in colostral immunoglobulin concentration. Milk production of cows receiving the PC treatment was the highest, with high fat and protein yields and a higher persistence of the production curve throughout the lactation.
Keywords:
postbiotic
; dairy cow
; voluntary intake
; digestibility
; immunoglobulin
; milk yield
1. Introduction
Ruminants have the ability to convert fiber-rich forage, which is indigestible to monogastrics, into a high-quality nutrient source. In grazing and mixed systems, the intake of ruminants relies mainly (about 90%) on forage: i.e. grass, silage and crop residues [1]. However, the high nutritional requirements of dairy cows during late gestation and early lactation fall within a period of low voluntary intake capacity. Thus, the supplementation with concentrate or improving forage digestibility is required to overcome this energy gap. Dairy cows are often fed energy-dense diets with high levels of starchy concentrates to meet all the animal's requirements. However, the rapid fermentation of starch by the rumen microbiome can reduce rumen pH and increase the risk of metabolic disorders [2].
The use of bacterial and yeast fermentation products as feed additives has been proposed to improve ruminal fermentation [3]. For example, yeast products containing Saccharomyces and Aspergillus can improve ruminal fermentation by increasing feed digestibility, whilst lactic acid producing bacteria can modulate the microbial balance in the gastrointestinal tract and improve feed efficiency. The beneficial health effects of probiotics lead to an increasing scientifical interest on the use of products derived from microbial metabolism. Probiotics are non-pathogenic living organisms which remain active in the gut and modify host's digestive microbiota when they are added to the diet in an adequate concentration and quantity [4]. However, the use of probiotics requires a cautious handling since they are living organisms which could lose their beneficial activity if they suffer adverse environmental conditions. They also carry potential risks such as infection and transfer of antibiotic resistance to pathogenic microbes, as well as excessive immune response, and even errors in dosage, formulation and route of administration [5]. The metabolites secreted by probiotics or released after their lysis also provide physiological benefits to the host and would avoid the risks associated with the administration of live bacteria. These soluble factors have been proposed as an alternative to the use of probiotics, been termed postbiotics [6], which exert a probiotic effect in the absence of live microbial cells [7]. Thus, the mode of action of postbiotics is expected to be similar to that of probiotics thanks to the presence of metabolites of probiotic bacteria. Postbiotics have a well-defined chemical structure (such as enzymes, peptides, teichoic acids, peptidoglycan-derived muropeptides, polysaccharides, cell surface proteins, and organic acids), safe dosage parameters and a long shelf life [8]. Postbiotics prevent the colonisation of pathogens by improving the intestinal environment for beneficial commensal bacteria [9]. The organic acids and bacteriocins in postbiotics reduce the pH of the gut, inhibiting the proliferation of opportunistic pathogens, and promoting the growth of beneficial bacteria that modulate the microbial balance and help maintain gut health [3]. In vivo studies suggest that they act as a biofilm that coats the intestinal surface facing the gut lumen, preventing the adhesion of pathogens [10]. Hence, postbiotics reduce the incidence of digestive disorders and improve the immune system [11]. In addition, the postbiotics have shown an increase of the fiber digestion in in vitro studies [12], which may help to improve the ruminal fermentation. Also, postbiotic supplementation in lactating goats improved fiber digestibility and energy efficiency for milk production [13].
Based on these findings, it was hypothesised that the addition of postbiotics to dairy cows' rations during the transition period could improve nutrient digestibility and increase productivity and milk yield. Therefore, the present study was designed to test the effects of including postbiotics in the diet of dairy cows on prepartum and postpartum feed digestibility and early lactation milk yield and composition of milk.
2. Materials and Methods
2.1. Ethics Statement
The trial was carried out at the SERIDA Demo-Farm located in Villaviciosa, Asturias, Spain (43°28'20.3"N 5°26'10.0"W) with the approval of the Regional Office for Animal Health and Animal Production (PROAE 18/2022), in accordance with the guidelines set out in the European Community Council Directive 2010/63/EU on the protection of animals used for scientific purposes.
2.2. Animals and Diets
Twelve Friesian dairy cows were selected for the experiment. The cows were six first calving heifers and six second calving cows, with a body weight of 621 kg ± 98.6 (mean ± standard deviation) and a body condition score (BCS) of 2.96 ± 0.235 (scale from 1 to 5). Feeding was ad libitum for both, pre- and postpartum, with total mixed rations (TMR) formulated according to NRC [14] with a 60/40 forage/concentrate ratio. In addition, all animals received a concentrate twice daily as an energy supplement: 3 kg of fresh matter daily before calving, and adjusted to the production level after calving: 3 kg/day as a base dose that was increased at a rate of 0.2 kg per kg of milk produced above the average milk yield of the herd, up to a maximum of 6 kg/day. Fresh water was continuously available for animals. The ingredients and chemical composition of the diets are listed in Table 1.
2.3. Experimental Design and Measurements
The feeding trial was arranged in a longitudinal, randomised block design with the animals allocated to 3 feeding groups (n = 4 per treatment) starting 45 days before the expected calving date and continuing for 60 days after calving. To minimise the differences among groups, cows were grouped by number of lactations (2 first and 2 second-lactation animals in each group), body weight and body condition score. Weight and body condition scores were recorded fortnightly. Two postbiotics supplied by Pentabiol S.L., (Noáin, Navarra, Spain) were tested. The postbiotics were added to the compound feedstuff at the time of manufacture. The supplementation per cow of both postbiotics was 8 g/d before calving and 15 g/d after calving. Each group of animals was assigned to an experimental treatment: Control (CT), Probisan-Ruminants-C® (PC), and Probisan-Ruminants® (PR). Daily TMR intake was recorded individually throughout the trial by a computerised electronic system integrated with a weighing pen, while daily concentrate intake was recorded by an automatic feeder attached to the milking system (DeLavalVMSTM, DeLaval International AB, Tumba, Sweden). The three TMR and the concentrate were sampled fortnightly throughout the experiment. Milk yield was recorded at both milkings at 07:00 h and 19:00 h. Colostrum was sampled at first milking and milk was sampled weekly. Morning and evening milk samples from each cow were mixed according to milk yield to produce a representative sample of 40 mL per day and cow, to which 0.13 mL of azidiol was added. Samples were kept refrigerated until analysis.
Two digestibility balances were made 15 days before expected calving and 30 days postpartum respectively. Two cows from each treatment, one from first and one from second calving, were randomly selected and placed in individual metabolic cages for 7 days to establish pre- and postpartum digestibility balances. The same cows were used in both digestibility balances. Animals were housed in metabolism cages throughout the digestibility balance to measure daily feed intake, milk yield, and faecal and urinary output. After two days of acclimatisation to the cages and for a period of 5 days, daily records of feed offered and refused, total faeces and urine output, and milk yield in the postpartum balance, were made for each cow to calculate the nutrient balance. An aliquot of 2 % of the total daily faecal output was collected from the collector box to provide a composite sample for each animal and period, and stored at –20 ⁰C until analysis. Total daily urine was collected through an external polyethylene vinyl acetate separator attached to the vulva with a biocompatible adhesive in plastic drums containing 1 L of a sulphuric acid solution (10 % v/v) to prevent microbial degradation and loss of volatile ammonium. After recording the weight and the specific gravity, a sample of 1 % of the daily urine production was collected and pooled on an animal basis to provide a composite sample for each animal and period, which was at –20 ⁰C until analysis. Cows were milked twice daily at 07:00 h and 19:00 h. Milk was collected from each animal on the second and fourth sampling days and milk samples were pooled and stored until analysis as described above.
2.4. Chemical Analyses
TMR samples were dried at 60 ⁰C for 24 hours and ground through a 0.75 mm sieve, and the concentrate samples were ground to 1 mm. Both samples were analysed by NIRS (Foss NIRSystem 5000, FOSS, Silver Spring, MD, USA) for dry matter (DM), ash, crude protein (CP), crude fiber (CF), ether extract (EE), starch, neutral detergent fiber (NDF), and acid detergent fiber (ADF). Ammonia N content of fresh faeces and urine was determined by the Kjeldahl method (Kjeltec 8400, FOSS Tecator, Hillerød, Denmark) after the alkalinisation of the samples with magnesium oxide (10 % v/v). The dry matter of faeces was determined by drying samples to a constant weight at 60 ⁰C, and the ash by combustion at 550 ⁰C (TGA-601 Thermogravimetric analyzer, Leco Corporation, St. Joseph, MI, USA). The concentration of NDF in faeces was determined by the Van Soest method [15]. The total N content of faeces and urine was determined by the Kjeldahl method, using Cu as the catalyst. Immunoglobulin of colostrum samples was analysed immediately after collection using an optical Brix refractometer (LLG-uniREFRACTO 5 pro, Lab Logistics Group GmbH, Meckenheim, Germany), then 0.13 mL of azidiol was added to 40 mL of colostrum and kept refrigerated until fat and protein contents were analysed by gravimetric method [16] and Kjeldahl method [17], respectively. Milk samples were analysed for fat, protein, lactose, solids-not-fat, and urea content by Fourier transform infrared (MilkoScan FT 6000, FOSS Tecator, Hillerød, Denmark).
2.5. Calculations and Statistical Analysis
Organic matter (OM) and Nitrogen-free extract (NFE) were calculated as:
The percentage of immediate principles is expressed on dry matter basis.
The net energy for lactation (NEmilk) of total mixed rations and concentrate was related to the concentration of metabolisable energy (ME) in the feedstuff using the NRC equation [14]:
Where ME was estimated as the average of the metabolizable energy calculated according to MAFF [18] and ADAS [19]:
The percentage of immediate principles is expressed on dry matter basis.
Apparent digestibility coefficients (ADC) of nutrients (on dry matter basis) were calculated for each animal as:
The concentration of immunoglobulin G (IgG) in colostrum was estimated according to the equation [20]:
Nitrogen conversion efficiency (NCE) was calculated as the ratio of daily milk protein yield to daily nitrogen intake.
Fat and protein corrected milk (FPCM) yield was calculated according to IDF [21]:
Statistical analyses were performed using the R statistical software [22]. For each variable tested, the mixed model included the fixed effects of treatment and trial day, and their interaction. Cow and number of lactations were considered as random effects. Day was considered as a repeated measure with cow as the subject. Multiple comparisons among treatments were evaluated using Duncan´s test. The significance level was set at p ≤ 0.05 and a trend was considered when 0.05 < p ≤ 0.10.
3. Results
Cows experienced the physiological weight gain at the end of gestation, as well as the typical decline in body weight and body condition during the transition to lactation (p < 0.05), although no differences among treatments were observed in these parameters. On average, cows started the trial with a body weight of 621 kg ± 98.6 and a body condition score of 2.96 ± 0.235, and reached calving with a body weight of 679 kg ± 85.5 and a body condition score of 3.21 ± 0.144. After 60 days in milk, the cows lost an average of 96 kg of body weight, whilst BCS decreased by 0.7 points. Cows calved normally without intervention, including one cow that gave birth to twins. All calves born were normal and healthy, with an average birth weight of 41.5 kg ± 4.24, with no differences among the experimental treatments.
Nutrient intake and digestibility before calving are shown in Table 2. Dry matter and organic matter intakes were higher in the PR treatment than in the CT treatment (p < 0.05), whereas the PC treatment showed intermediate values with no differences between the other treatments. Crude protein and neutral detergent fiber intakes were higher in both treatments supplemented with postbiotics than in the CT treatment (p < 0.05). The nutrient balance showed that the digestibility of dry matter and organic matter tended to be higher in both supplemented treatments than in the control treatment (p < 0.1). As a result, the intake of digestible organic matter was higher in both supplemented treatments compared to the control (5.84 kg/day vs. 6.38 and 6.58 kg/day for CT, PC, and PR respectively, p < 0.05). Crude protein digestibility was higher in the PC treatment compared to the control (p < 0.05), while the PR treatment showed intermediate values. Thus, the intake of digestible crude protein was higher in both postbiotic treatments compared to the control (0.87 kg/day vs. 1.08 and 0.99 kg/day for CT, PC, and PR respectively, p < 0.05). However, no differences in nitrogen balance were observed among treatments.
Table 3 shows nutrient intake and digestibility after calving. The intakes of dry matter, organic matter, and neutral detergent fiber were higher in the PR treatment than in the CT and PC treatments (p < 0.05). Crude protein intake was higher in the CT and PR treatments, with no differences between them. However, the PC treatment had a lower crude protein intake than the other treatments (p < 0.05).
The digestibility of dry matter, organic matter, and neutral detergent fiber was higher when the cows were supplemented with PR than in the CT and PC treatments (p < 0.05). As a result, the intake of digestible nutrients was also higher in PR treatment, especially the intake of digestible organic matter (13.40 kg/day vs. 10.89 and 10.39 kg/day for PR, PC, and CT respectively, p < 0.05) and neutral detergent fiber (5.23 kg/day vs. 3.42 kg/day for PR, and both PC and CT respectively, p < 0.05). No differences in crude protein digestibility and nitrogen balance were observed in the postpartum period among treatments.
The composition of colostrum is shown in Figure 1. Colostral protein concentration was significantly higher in the supplemented treatments than in the control (11.00 % vs. 13.44 % and 13.18 % for CT, PC, and PR respectively, p < 0.05), as well as the levels of immunoglobulins (52.46 g/L vs. 76.65 g/L and 76.61 g/L for CT, PC, and PR respectively, p < 0.01). In contrast, no differences in colostral fat concentrations were observed among treatments.
Table 4 shows the results of milk production and composition in the experimental treatments. The highest milk production was observed at the PC treatment, which was statistically different (p < 0.05) to the PR and the non-supplemented treatments.
Figure 2 shows the milk production for each of the experimental treatments up to day 60 postpartum of the experiment (a) and the entire lactation curve including data after the end of the study when all animals were fed with the same ration without postbiotic supplementation (b). Non-supplemented cows always had lower milk production than those receiving postbiotics. Both PC and PR treatments had similar milk yield up to the peak of lactation (around 35 days in milk). Thereafter, milk production decreased faster in the PR treatment than in the PC treatment, which was the key to the statistical differences between the postbiotic-supplemented treatments. Consequently, a higher persistence of the lactation curve was observed in the PC treatment than in the PR treatment throughout the lactation with a higher total milk yield. Milk protein concentration tended to be higher in the PC and PR treatments than the CT treatment (p < 0.1). The highest fat concentration occurred in the PC treatment, with a trend towards higher concentrations than in the CT and PR treatments (p < 0.1). Solids-not-fat were higher in milk from cows supplemented with PR than in the PC and CT treatments (p < 0.05). When the milk was corrected for fat and protein concentration, the highest milk yield was also observed in the PC treatment and was statistically different (p < 0.05) to the PR and the CT treatments. Consequently, milk performance was better in the PR treatment, with higher protein and fat yields, than in the PR and the CT treatments (p < 0.05). The efficiency of dietary nitrogen utilisation also tended (p < 0.1) to be better in cows supplemented with postbiotic PC.
4. Discussion
The effect of postbiotics on voluntary intake in dairy cows varies between studies. While some authors observed no effect of postbiotic supplementation during the peripartum period [23], others reported an increase in voluntary intake immediately before and after calving [24] or immediately after calving [25]. These authors speculate that postbiotic supplementation reduces inflammation and stress around parturition, which increases voluntary intake immediately afterwards. This is supported by the fact that serum cortisol, a biomarker of stress and pain [26], decreases in the days immediately following parturition [27], suggesting that postbiotic supplementation may reduce parturition stress. Previous studies have suggested that yeast-derived products may be used to a greater extent in stressed animals [28]. The different results reported in the literature have been attributed to high inter-animal variability [29], as they appear to depend on breed, animal type, age and physiological phase. For example, in Jersey-Holstein crossbred calves supplemented with postbiotics similar to those used in our study, an increase in dry matter intake was observed from weaning to ten weeks of age, while no differences were observed at later stages [30]. On the contrary, a decrease in dry matter intake was observed in adult Holstein cows supplemented with postbiotics after peak lactation [31], although other studies found no effect on voluntary intake [23]. Supplementation of dairy goats with postbiotics similar to those used in our study showed no differences in apparent nutrient digestibility, except for fiber digestibility [13]. However, higher dry matter intake and dry matter, protein, and fiber digestibilities were reported in lambs supplemented with another postbiotic [32]. A meta-analysis of supplementation with postbiotics derived from Saccharomyces cerevisiae showed that voluntary intake increased early in lactation, but decreased later, even when supplementation was maintained [33]. In our study, the PR treatment shows a higher voluntary intake than the PC and CT treatments, in parallel with a higher apparent digestibility of nutrients, especially after calving. The higher digestibility may be due to the fact that the higher dry matter intake during ruminal fermentation provides a higher amount of volatile fatty acids and nitrogen to the ruminal microbiome for microbial synthesis, which would consequently increase ruminal digestibility. Using Rusitec fermenters, it was observed that yeast hydrolysate increased the concentrations of ammonia nitrogen, propionic acid and butyric acid, and decreased the concentration of acetic acid and the acetic/propionic ratio [34]. These authors also observed an increase in microbial protein synthesis and bacterial diversity. In addition, other authors suggest that yeast metabolites would have effects on the rumen environment, including increasing pH and altering volatile fatty acids concentrations [35], as well as providing soluble growth factors [36] that stimulate the growth of cellulolytic bacteria [37], and consequently increase the extent of ruminal digestion of fiber and the rate of fiber transit improving dry matter intake. Therefore, it can be speculated that the postbiotic added with the PR treatment provides an improvement in the rumen environment, inducing increased nutrient degradability, and resulting in increased voluntary intake.
The concentration of colostral immunoglobulins was higher in the supplemented treatments than in the control, which was reflected in a higher concentration of protein, being positively correlated [38]. In any case, the concentration of immunoglobulins in colostrum was above 50 g IgG/L in all treatments, a concentration above which the colostrum is considered to be of high quality [39]. Calves are mainly protected by passively transferred immunity from colostrum [40]. Thus, a lower concentration of 50 g IgG/L of colostrum exposes the calf to increased susceptibility to disease in the first days of life, with negative consequences in the medium and long term. In monogastric animals, supplementation with prebiotics and probiotics increases the concentration of immunoglobulins in the colostrum of bitches [41,42] and sows [43,44]. In addition, the concentration of IgG in colostrum is influenced by the duration of the supplementation period [45]. To our knowledge, we have not found any studies reporting similar conclusions in ruminants. However, from the results of our work we can confirm that supplementing cows with postbiotics increases the concentration of immunoglobulins in colostrum. As a result, calves from cows supplemented with postbiotics during pregnancy may have higher levels of immunoglobulins.
The highest milk production was observed with the PC treatment in the current study, while the PR did not differ from the control treatment. The different results may be due to the different nature of the postbiotics evaluated. However, variable results can be found in the literature. By supplementing yeast as a probiotic, some authors have found improvements in milk production both in cows [46] and goats [47], while others have found no effect on production in early lactation [48]. Similarly, milk production responses are variable when yeast-derived postbiotics were used. Some studies described increases in milk production [27,49,50], whereas others did not observe any effect [24,51]. The response of dairy cows to probiotic supplementation, and therefore to postbiotics due to their similar mode of action, depends on the stage of lactation, the type of feed and the forage/concentrate ratio [52], as well as the type of probiotic [51]. In our case, both the physiological state of the cows and the ration were the same in all treatments, so the differences observed could be attributed to the different modes of action of the postbiotics evaluated. However, as these were commercial additives, we have no knowledge of the nature of the additives and the possible differences in how they work.
In the current study, the milk fat concentration of cows fed the PC treatment tended to be higher compared to the other treatments and the protein concentration also tended to be higher than the control treatment. This results, along with their higher milk yield observed for PC, resulted in significantly higher milk, protein and milk fat yields. These results are in agreement with other studies [23,53], who reported that cows supplemented with yeast culture as a probiotic had a higher milk fat content and a higher milk fat yield, although they did not observe a higher milk protein yield. However, other authors found no differences when supplementing with yeast-derived postbiotics [25]. The proportion of protein in the milk of cows supplemented with the PR treatment tended to be higher than in the control and PC treatments. This result, along with a non-significantly higher lactose concentration, resulted in a higher proportion of solids-not-fat of milk in PR compared to the control and PC treatments. Solids-not-fat of milk consist of proteins (mainly casein) carbohydrates (mainly lactose) minerals and vitamins, that add texture and creaminess to the milk [54], which are highly valued by consumers. Thus, the addition of postbiotic PR to the cows' ration could improve the organoleptic characteristics of the milk. In all treatments, urea values were always within the range considered optimal (210 and 320 mg/L), indicating that the rations had an optimal protein and energy intake [55].
5. Conclusions
The inclusion of postbiotics in dairy cows during the transition period increased the voluntary intake of dry matter, especially in the PR treatment, with higher apparent total tract digestibility of dry matter, organic matter and neutral detergent fiber. Both treatments including postbiotics induced an increase in colostral immunoglobulin concentration, which would favour calf health in the first weeks of life. Milk production of cows receiving the PC treatment was the highest, with high fat and protein yields and a higher persistence of the production curve throughout the lactation. Thus, the results of the current study suggest that postbiotic supplementation in late gestation and early lactation could be an effective strategy to improve colostrum immune quality and increase milk yield in dairy cows.
Author Contributions
Conceptualization, F.V.; methodology, F.V.; software, F.V. and M.M.-M.; validation, F.V. and A.M.-F.; formal analysis, F.V., M.M.-M. and A.M.-F.; investigation, F.V. and A.M.-F.; resources, F.V.; data curation, F.V., M.M.-M. and A.M.-F.; writing—original draft preparation, F.V.; writing—review and editing, F.V., M.C.-C., M.M.-M., J.G.-R and A.M.-F.; visualization, F.V., M.C.-C., M.M.-M., J.G.-R and A.M.-F.; supervision, F.V.; project administration, F.V.; funding acquisition, F.V. All authors have read and agreed to the published version of the manuscript.
Funding
Work financed by Sekuens: Asturian Agency for Science, Business Competitiveness and Innovation (former IDEPA) under project IDE/2021/000555, Open-Lab La Granja (FICYT IDI/2021/000200) and consolidated research group Nutrición y Sanidad Animal: NySA (FICYT IDI/2021/000102). María Campo-Celada is the recipient of a PhD grant PRE2021-100447 funded by MCIN/AEI/10.13039/501100011033 and by “ESF+”.
Institutional Review Board Statement
The study was conducted in accordance with the guidelines of the Research Ethics Committee of the University of Oviedo (PRONAE 18/2022).
Informed Consent Statement
Not applicable.
Data Availability Statement
All data used to support the findings of this study are included in the article.
Acknowledgments
The authors would like to thank the staff of SERIDA for their technical assistance. The authors would also like to thank Pentabiol S.L. for supplying the two postbiotics, CAPSA FOOD for supporting the realisation of this work, and Sociedad Asturiana de Servicios Agropecuarios, S.L. for producing the compound feedstuff containing the postbiotics.
Conflicts of Interest
The authors declare no real or perceived conflicts of interest.
References
- Mottet, A., de Haan, C., Falcucci, A., Tempio, G., Opio, C., Gerber, P. Livestock: On our plates or eating at our table? A new analysis of the feed/food debate. Glob. Food Sec. 2017, 14, 1-8. [CrossRef]
- Zebeli, Q., Metzler-Zebeli, B.U. Interplay between rumen digestive disorders and diet-induced inflammation in dairy cattle. Res. Vet. Sci. 2012, 93, 1099-1108. [CrossRef]
- Seo, J., Kim, S., Kim, M., Upadhaya, S., Kam, D., Ha, J. Direct-fed Microbials for Ruminant Animals. Asian-Aust. J. Anim. Sci. 2010, 23, 1657-1667. [CrossRef]
- Fijan, S. Microorganisms with claimed probiotic properties: An overview of recent literature. Int. J. Environ. Res. Public Health 2014, 5, 4745-4767. [CrossRef]
- Li, T., Teng, D., Mao, R., Hao, Y., Wang, X., Wang, J. A critical review of antibiotic resistance in probiotic bacteria. Food Res. Int. 2020, 136, 109571. [CrossRef]
- Salminen, S., Collado, M.C., Endo, A., Hill, C., Lebeer, S., Quigley, E.M.M., Sanders, M.E., Shamir, E., Swann, J.R., Szajewska, H., Vinderola, G. The International Scientific Association of Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of postbiotics. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 649–667. [CrossRef]
- Thanh, N.T., Loh, T.C., Foo, H.L., Hair-bejo, M., Azhar, B.K. Effects of feeding metabolite combinations produced by Lactobacillus plantarum on growth performance, faecal microbial population, small intestine villus height and faecal volatile fatty acids in broilers. Br. Poult. Sci. 2009, 50, 298–306. [CrossRef]
- Aguilar-Toalá, J.E., García-Varela, R., García, H.S., Mata-Haro, V., González-Córdova, A.F., Vallejo-Cordoba, B., Hernández-Mendoza, A. Postbiotics: An evolving term within the functional foods field. Trends Food Sci. Technol. 2018, 75, 105-114. [CrossRef]
- Izuddin, W.I., Loh, T.C., Foo, H.L., Samsudin, A.A., Humam, A.M. Postbiotic L. plantarum RG14 improves ruminal epithelium growth, immune status and upregulates the intestinal barrier function in post-weaning lambs. Sci. Rep. 2019, 9, 9938. [CrossRef]
- Kareem, K.Y., Ling, F.H., Chwen, L.T., Foong, O.M., Asmara, S.A. Inhibitory activity of postbiotic produced by strains of Lactobacillus plantarum using reconstituted media supplemented with inulin. Gut Pathog. 2014, 6, 23-29. [CrossRef]
- Rovai, M., Guifarro, L., Salama, A.A. Effects of long-term postbiotic supplementation on dairy heifer calves: Health status and wound healing after dehorning. J. Dairy Sci. 2019, 102, Suppl. 1, 221.
- Izuddin, W.I., Loh, T.C., Samsudin, A.A., Foo, H.L. In vitro study of postbiotics from Lactobacillus plantarum RG14 on rumen fermentation and microbial population. Rev. Bras. Zootecn. 2018, 47, e20170255.
- Fernández, C., Romero, T., Badiola, I., Díaz-Cano, J., Sanzol, G., Loor, J.J. Postbiotic yeast fermentation product supplementation to lactating goats increases the efficiency of milk production by enhancing fiber digestibility and ruminal propionate, and reduces energy losses in methane. J. Anim Sci. 2023, 101, skac370. [CrossRef]
- NRC Nutrient Requirements of Dairy Cattle, 7th ed.; National Academy Press: Washington, DC, USA, 2001.
- Van Soest, P.J., Robertson, J.B., Lewis, B.A. Methods of dietary fiber, neutral detergent fiber and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [CrossRef]
- ISO ISO 23318:2022 Milk, dried milk products and cream. Determination of fat content - Gravimetric method, Edition 1, 2022; International Organization for Standardization: Geneva, Switzerland, 2022.
- ISO ISO 8968-1:2014 IDF 20-1:2014 Milk and milk products- Determination of nitrogen content Part 1: Kjeldahl principle and crude protein calculation, Edition 2, 2014; International Organization for Standardization: Geneva, Switzerland, 2014.
- MAFF Energy allowances and feeding systems for ruminants. Reference Book 433, Ministry of Agriculture, Fisheries and Food: Reading, United Kingdom, 1984.
- ADAS Compound feed evaluation fir the ruminants. Technical Bulletin 85/21, Ministry of Agriculture, Fisheries and Food. Reading, United Kingdom, 1985.
- Quigley, J.D., Lago, A., Chapman, C., Erickson, P., Polo, J. Evaluation of the Brix refractometer to estimate immunoglobulin G concentration in bovine colostrum. J. Dairy Sci. 2013, 96, 1148-1155. [CrossRef]
- IDF A common carbon footprint approach for dairy. The IDF guide to standard lifecycle assessment methodology for the dairy sector. Bulletin IDF No. 479/2010. International Dairy Federation: Brussels, Belgium, 2015.
- R Core Team R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing: Vienna, Austria, 2024.
- Olagaray, K.E., Sivinski, S.E., Saylor, B.A., Mamedova, L.K., Sauls-Hiesterman, J.A., Yoon, I., Bradford, B.J. Effect of Saccharomyces cerevisiae fermentation product on feed intake parameters, lactation performance, and metabolism of transition dairy cattle. J. Dairy Sci. 2019, 102, 8092-8107. [CrossRef]
- Dann, H.M., Drackley, J.K., McCoy, G.C., Hutjens, M.F., Garrett, J.E. Effects of yeast culture (Saccharomyces cerevisiae) on prepartum intake and postpartum intake and milk production of Jersey cows. J. Dairy Sci. 2000, 83, 123-127. [CrossRef]
- Shi, W., Knoblock, C.E., Murphy, K.V., Bruinjé, T.C., Yoon, I., Ambrose, D.J., Oba, M. Effects of supplementing a Saccharomyces cerevisiae fermentation product during the periparturient period on performance of dairy cows fed fresh diets differing in starch content. J. Dairy Sci. 2019, 102, 3082-3096. [CrossRef]
- Gross, J.J., Wellnitz, O., Bruckmaier, R.M. Cortisol secretion in response to metabolic and inflammatory challenges in dairy cows. J. Anim Sci. 2015, 93, 3395–3401. [CrossRef]
- Zaworski, E.M., Shriver-Munsch, C.M., Fadden, N.A., Sanchez, W.K., Yoon, I., Bobe, G. Effects of feeding various dosages of Saccharomyces cerevisiae fermentation product in transition dairy cows. J. Dairy Sci. 2014, 97, 3081–3098. [CrossRef]
- Arambel, M.J., Kent, B.A. Effect of yeast culture on nutrient digestibility and milk yield response in early- to midlactation dairy cows. J. Dairy Sci. 1990, 73, 1560–1563. [CrossRef]
- Khalouei, H., Seranatne, V., Fehr, K., Guo, J., Yoon, I., Khafipour, E., Plaizier, J.C. Effects of Saccharomyces cerevisiae fermentation products and subacute ruminal acidosis on feed intake, fermentation, and nutrient digestibilities in lactating dairy cows. Can. J. Anim Sci. 2021, 101, 143-157. [CrossRef]
- Rovai, M., Guifarro, L., Anderson, J., Salama, A.A.K. Effects of long-term postbiotic supplementation on dairy heifer calves: Performance and metabolic indicators. In Proceedings of the ADSA Annual Meeting, Cincinnati, Ohio, USA, 23-26 June 2019.
- Shi, W., Knoblock, C.E., Yoon, I., Oba, M. Effects of supplementing a Saccharomyces cerevisiae fermentation product during the transition period on rumen fermentation of dairy cows fed fresh diets differing in starch content. J. Dairy Sci. 2019, 102, 9943-9955. [CrossRef]
- Izuddin, W.I., Loh, T.C., Samsudin, A.A., Foo, H.L., Humam, A.M., Shazali, N. Effect of postbiotic supplementation on growth performance, ruminal fermentation and microbial profile, blood metabolite and GHR, IGF-1 and MCT-1 gene expression in post-weaning lambs. BMC Vet. Res. 2019, 15, 315. [CrossRef]
- Poppy, G.D., Rabiee, A.R., Lean, I.J., Sanchez, W.K., Dorton, K.L., Morley, P.S. A meta-analysis of the effects of feeding yeast culture produced by anaerobic fermentation of Saccharomyces cerevisiae on milk production of lactating dairy cows. J. Dairy Sci. 2012, 95, 6027-6041. [CrossRef]
- Díaz, A., Ranilla, M.J., Saro, C., Tejido, M.L., Pérez-Quintana, M., Carro, M.D. Influence of increasing doses of a yeast hydrolyzate obtained from sugarcane processing on in vitro rumen fermentation of two different diets and bacterial diversity in batch cultures and Rusitec fermenters. Anim. Feed Sci. Technol. 2017, 232, 129-138. [CrossRef]
- Williams, P.E.V., Tait, C.A.G., Innes, G.M., Newbold, C.J. Effects of the inclusion of yeast culture (Saccharomyces cerevisiae plus growth medium) in the diet of dairy cows on milk yield and forage degradation and fermentation patterns in the rumen of steers. J. Anim Sci. 1991, 69, 3016–3026. [CrossRef]
- Callaway, E.S., Martin, S.A. Effects of a Saccharomyces cerevisiae culture on ruminal bacteria that utilize lactate and digest cellulose. J. Dairy Sci. 1997, 80, 2035–2044. [CrossRef]
- Wiedmeier, R.D., Arambel, M.J., Walters, J.L. Effects of yeast culture and Aspergillus oryzae fermentation extract on ruminal characteristics and nutrient digestibility. J. Dairy Sci. 1987, 70, 2063–2068. [CrossRef]
- Quigley III, J.D., Martin, K.R., Dowlen, H.H., Wallis, L.B., Lamar, K. Immunoglobulin concentration, specific gravity, and nitrogen fractions of colostrum from Jersey cattle. J. Dairy Sci. 1994, 77, 264-269. [CrossRef]
- Costa, A., Sneddon, N.W., Goi, A., Visentin, G., Mammi, L.M.E., Savarino, E.V., Zingone, F., Formigoni, A., Penasa, M., De Marchi, M. Bovine colostrum, a promising ingredient for humans and animals—Properties, processing technologies, and uses. J. Dairy Sci. 2023, 106, 5197–5217. [CrossRef]
- Galvao, K.N., Santos, J.E., Coscioni, A., Villasenor, M., Sischo, W.M., Berge, A.C. Effect of feeding live yeast products to calves with failure of passive transfer on performance and patterns of antibiotic resistance in fecal Escherichia coli. Reprod. Nutr. Dev. 2005, 45, 427–440. [CrossRef]
- Salmon, H. Immunophysiology of the mammary gland and transmission of immunity to the young. Reprod. Nutr. Dev. 2003, 43, 471–475. [CrossRef]
- Agazzi, A. The beneficial role of probiotics in monogastric animal nutrition and health. Dairy Vet. Anim. Res. 2015, 2, 116–132. [CrossRef]
- Zanello, G., Meurens, F., Serreau, D., Chevaleyre, C., Melo, S., Berri, M., D'Inca, R., Auclair, E., Salmon, H. Effects of dietary yeast strains on immunoglobulin in colostrum and milk of sows. Vet. Immunol. Immunopathol. .2013, 152, 20-27. [CrossRef]
- Wu, H., Xu, C., Wang, J., Hu, C., Ji, F., Xie, J., Yang, Y., Yu, X., Diao, X., Lv, R. Effects of Dietary Probiotics and Acidifiers on the Production Performance, Colostrum Components, Serum Antioxidant Activity and Hormone Levels, and Gene Expression in Mammary Tissue of Lactating Sows. Animals 2023, 13, 1536. [CrossRef]
- Alonge, S., Aiudi, G.G., Lacalandra, G.M., Leoci, R., Melandri, M. Pre and Probiotics to Increase the Immune Power of Colostrum in Dogs. Front. Vet. Sci. 2020, 7, 570414. [CrossRef]
- Ramsing, E.M., Davidson, J.A., French, P.D., Yoon, I., Keller, M., Peters-Fleckenstein, H. Effects of yeast culture on peripartum intake and milk production of primiparous and multiparous Holstein cows. Prof. Anim. Sci. 2009, 25, 487–495. [CrossRef]
- Stella, A.V., Paratte, R., Valnegri, L., Cigalino, G., Soncini, G., Chevaux, E., Dell’Orto, V., Savoini, G. Effect of administration of live Saccharomyces cerevisiae on milk production, milk composition, blood metabolites, and faecal flora in early lactating dairy goats. Small Rumin. Res. 2007, 67, 7–13. [CrossRef]
- Wohlt, J.E., Corcione, T.T., Zajac, P.K. Effects of yeast on feed intake and performance of cows fed diets based on corn silage during early lactation. J. Dairy Sci. 1998, 81, 1345–1352. [CrossRef]
- Acharya, S., Pretz, J.P., Yoon, I., Scott, M.F., Casper, D.P. Effects of Saccharomyces cerevisiae fermentation products on the lactational performance of mid-lactation dairy cows. Transl. Anim. Sci. 2017, 1, 221–228. [CrossRef]
- Dias, A.L.G., Freitas, J.A., Micai, B., Azevedo, R.A., Greco, L.F., Santos, J.E.P. Effects of supplementing yeast culture to diets differing in starch content on performance and feeding behavior of dairy cows. J. Dairy Sci. 2018, 101, 186–200. [CrossRef]
- Yuan, K., Liang, T., Muckey, M.B., Mendonça, L.G.D., Hulbert, L.E., Elrod, C.C., Bradford, B.J. Yeast product supplementation modulated feeding behavior and metabolism in transition dairy cows. J. Dairy Sci. 2015, 98, 532–540. [CrossRef]
- Piva, G., Belladonna, S., Fusconi, G., Sicbaldi, F. Effects of yeast on dairy cow performance, ruminal fermentation, blood components, and milk manufacturing properties. J. Dairy Sci. 1993, 76, 2717–2722. [CrossRef]
- White, R.A., Harrison, J.H., Yoon, I., Sanchez, W.K., Nicholson, N. Effect of yeast culture on efficiency of nutrient utilization for milk production and impact on fiber digestibility and fecal particle size. Prof. Anim. Sci. 2008, 24, 114-119. [CrossRef]
- O’Regan, J., Mulvihill, D. Preparation, characterization and selected functional properties of sodium caseinate–maltodextrin conjugates. Food Chem. 2009, 115, 1257–1267. [CrossRef]
- Kleefisch, M.T., Zebeli, Q., Humer, E., Gruber, L., Klevenhusen, F. Effects of feeding high-quality hay with graded amounts of concentrate on feed intake, performance and blood metabolites of cows in early lactation. Arch. Anim. Nutr. 2018, 72, 290–307. [CrossRef]
Figure 1.
Concentration of fat (%), protein (%), and immunoglobulin G (g/L) in colostrum from cows in each of the experimental treatments. CT: Control; PC: Probisan-Ruminants-C®; PR: Probisan-Ruminants®; Different letters indicate statistical differences between treatments.
Figure 1.
Concentration of fat (%), protein (%), and immunoglobulin G (g/L) in colostrum from cows in each of the experimental treatments. CT: Control; PC: Probisan-Ruminants-C®; PR: Probisan-Ruminants®; Different letters indicate statistical differences between treatments.
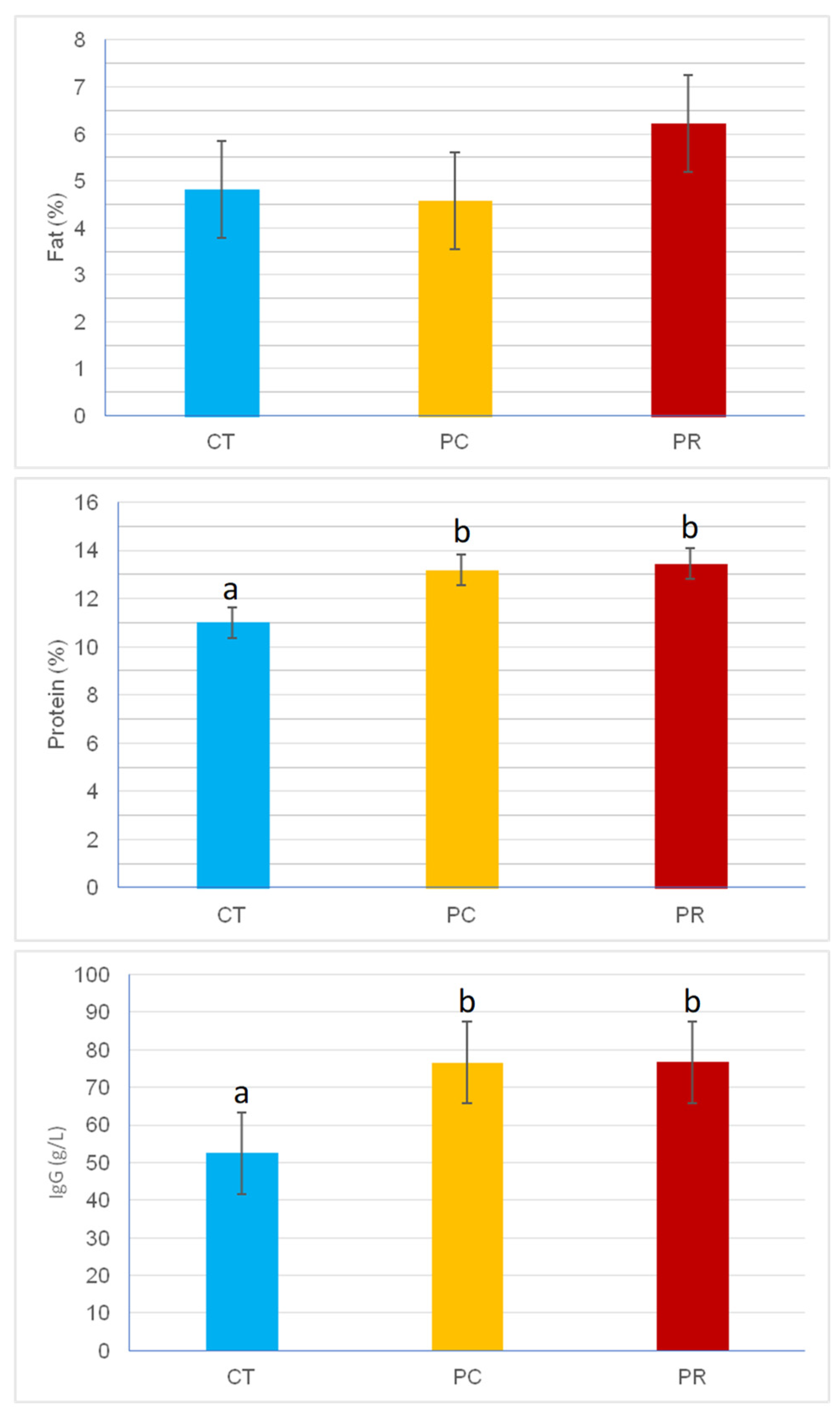
Figure 2.
Milk production with the experimental diets during the experimental postpartum period (a) and throughout the lactation after the end of the study when all animals were fed with the same ration without postbiotic supplementation (b). CT: Control; PC: Probisan-Ruminants-C®; PR: Probisan-Ruminants®.
Figure 2.
Milk production with the experimental diets during the experimental postpartum period (a) and throughout the lactation after the end of the study when all animals were fed with the same ration without postbiotic supplementation (b). CT: Control; PC: Probisan-Ruminants-C®; PR: Probisan-Ruminants®.
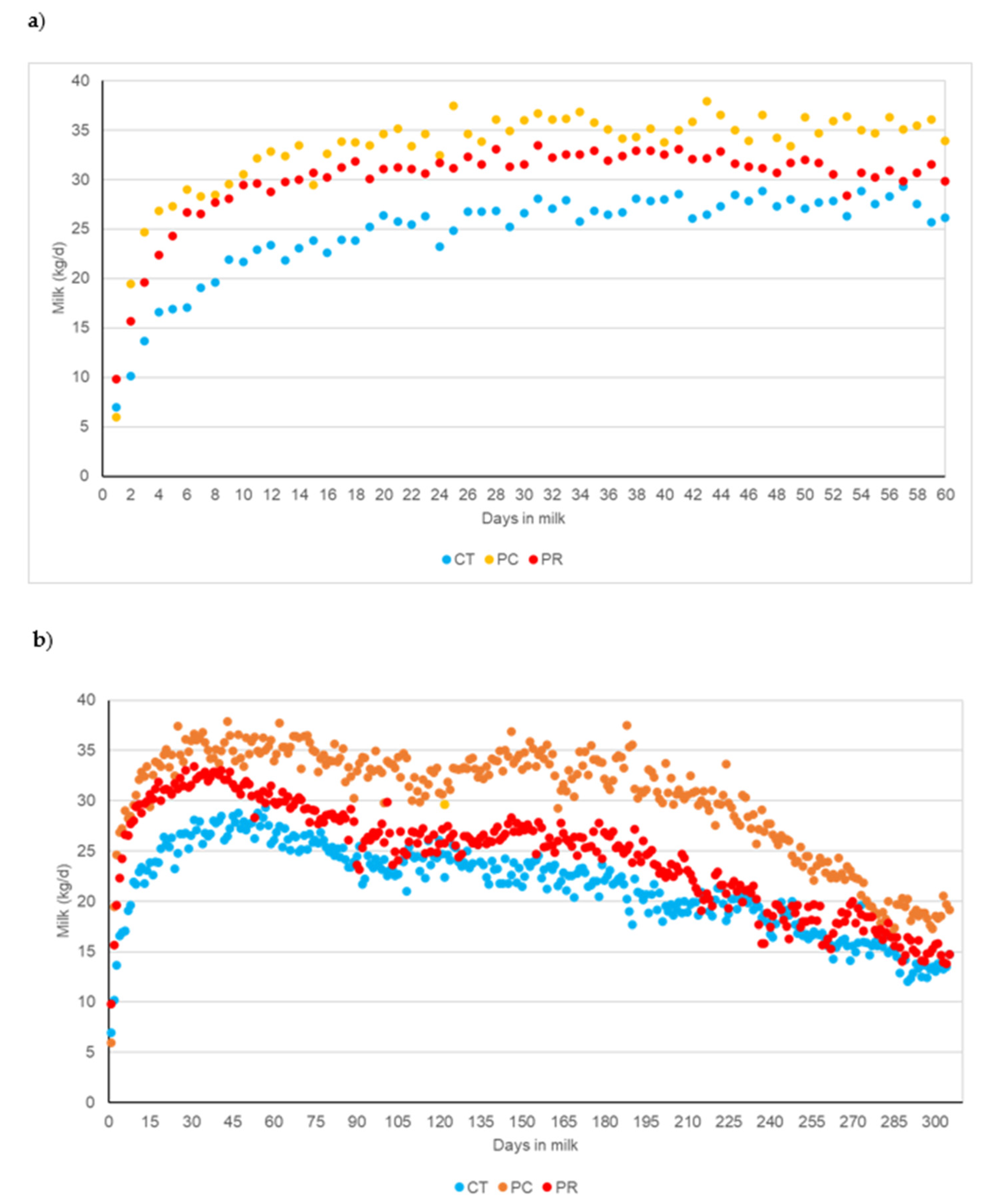

Table 1.
Ingredients and chemical composition of the pre and postpartum total mixed rations and the concentrate supplied.
Table 1.
Ingredients and chemical composition of the pre and postpartum total mixed rations and the concentrate supplied.
| Prepartum | Postpartum | Concentrate1 | |
|---|---|---|---|
| Diet composition (g/kg dry matter) | |||
| Grass silage | 411.6 | 468.1 | |
| Cereal straw | 121.4 | 61.2 | |
| Compound feedstuff1 | 467.0 | 470.7 | |
| Chemical composition (g/kg dry matter) | |||
| Dry matter (DM) | 469.8 | 430.1 | 887.1 |
| Organic matter (OM) | 903.9 | 902.6 | 913.6 |
| Crude protein (CP) | 131.0 | 139.6 | 228.2 |
| Crude fiber (CF) | 259.2 | 212.9 | 49.6 |
| Ether extract (EE) | 36.0 | 40.8 | 35.4 |
| Nitrogen-free extract (NFE) | 477.6 | 509.4 | 600.4 |
| Starch | 138.9 | 174.0 | 375.2 |
| Neutral detergent fiber (NDF) | 488.0 | 431.9 | 201.7 |
| Acid detergent fiber (ADF) | 306.3 | 271.9 | 85.6 |
| Net energy for lactation (Mcal/kg DM) | 1.44 | 1.52 | 1.89 |
1 Provided by Sociedad Asturiana de Servicios Agropecuarios, S.L. (Siero, Asturias, Spain).
Table 2.
Average nutrients intake and apparent digestibility of nutrients according to experimental treatments during the prepartum period.
Table 2.
Average nutrients intake and apparent digestibility of nutrients according to experimental treatments during the prepartum period.
| CT | PC | PR | rsd | p | |
|---|---|---|---|---|---|
| Intake (kg/day) | |||||
| Dry matter | 10.26b | 10.46ab | 10.97a | 0.498 | 0.027 |
| Organic matter | 9.28b | 9.50ab | 9.94a | 0.451 | 0.025 |
| Crude protein | 1.44b | 1.60a | 1.57a | 0.068 | 0.001 |
| Neutral detergent fiber | 4.37b | 4.71a | 4.72a | 0.233 | 0.009 |
| Digestibility (%) | |||||
| Dry matter | 62.00 | 66.91 | 64.92 | 4.030 | 0.091 |
| Organic matter | 62.87 | 67.59 | 66.03 | 3.889 | 0.088 |
| Crude protein | 60.16b | 68.20a | 63.01ab | 4.059 | 0.005 |
| Neutral detergent fiber | 50.38 | 52.30 | 52.12 | 5.532 | 0.773 |
| Nitrogen balance (%) | 45.21 | 54.64 | 48.77 | 7.253 | 0.344 |
CT: Control; PC: Probisan-Ruminants-C®; PR: Probisan-Ruminants®; rsd: residual standard deviation; p: significance, different letters in the same item indicate statistical differences between treatments.
Table 3.
Average nutrients intake and apparent digestibility of nutrients according to experimental treatments during the postpartum period.
Table 3.
Average nutrients intake and apparent digestibility of nutrients according to experimental treatments during the postpartum period.
| CT | PC | PR | rsd | p | |
|---|---|---|---|---|---|
| Intake (kg/day) | |||||
| Dry matter | 18.37b | 18.06b | 19.88a | 1.118 | 0.001 |
| Organic matter | 16.53b | 16.49b | 17.95a | 1.014 | 0.015 |
| Crude protein | 3.35a | 3.05b | 3.24a | 0.167 | 0.008 |
| Neutral detergent fiber | 5.90b | 6.29b | 7.30a | 0.435 | 0.001 |
| Digestibility (%) | |||||
| Dry matter | 64.00b | 64.76b | 73.44a | 7.406 | 0.036 |
| Organic matter | 65.90b | 66.36b | 74.68a | 6.952 | 0.035 |
| Crude protein | 55.02 | 55.56 | 63.56 | 8.921 | 0.131 |
| Neutral detergent fiber | 57.67b | 55.49b | 71.65a | 8.686 | 0.003 |
| Nitrogen balance (%) | 41.28 | 44.85 | 50.32 | 11.040 | 0.307 |
CT: Control; PC: Probisan-Ruminants-C®; PR: Probisan-Ruminants®; rsd: residual standard deviation; p: significance, different letters in the same item indicate statistical differences between treatments.
Table 4.
Milk production and milk composition in each experimental treatment.
| CT | PC | PR | rsd | p | |
|---|---|---|---|---|---|
| Milk (kg/d) | 29.61b | 33.73a | 30.06b | 6.908 | 0.044 |
| Fat (%) | 4.03 | 4.19 | 3.80 | 0.668 | 0.068 |
| Protein (%) | 3.10 | 3.22 | 3.27 | 0.307 | 0.083 |
| Solids-not-fat (%) | 8.73b | 8.68b | 8.99a | 0.413 | 0.007 |
| Lactose (%) | 4.83 | 4.79 | 5.08 | 0.648 | 0.148 |
| Urea (mg/kg) | 223 | 210 | 229 | 48.3 | 0.296 |
| Milk performance | |||||
| FPCM (kg/d) | 29.26b | 34.30a | 29.12b | 7.046 | 0.004 |
| kg fat/d | 1.16b | 1.42a | 1.13b | 0.336 | 0.002 |
| kg protein/d | 0.92b | 1.08a | 0.98b | 0.211 | 0.014 |
| Nitrogen eficiency (%) | 34.16 | 39.93 | 36.68 | 9.546 | 0.078 |
CT: Control; PC: Probisan-Ruminants-C®; PR: Probisan-Ruminants®; rsd: residual standard deviation; p: significance, different letters in the same item indicate statistical differences between treatments.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



