
Submitted:
09 July 2024
Posted:
10 July 2024
You are already at the latest version
Abstract
The Indian meal moth, Plodia interpunctella, poses a significant threat to global agricultural products. Although pheromone-based technologies show promise for P. interpunctella management, limitations like single-sex targeting necessitate exploring complementary strategies. Vegetable-based oils represent a potential alternative, but their efficacy, sex-specificity, and interaction with sex pheromones remain unclear. To answer these questions, we first examined attraction of P. interpunctella female and male adults to ten different vegetable oils, i.e., camellia, corn, olive, peanut, rapeseed, sesame, soybean, sunflower, walnut, and a blended oil (mixture of sunflower seed, rapeseed, soybean, peanut, and sesame oils), in the semi-field conditions. Sesame, olive, and blended oils demonstrated most attractive, capturing significantly more adults compared to other oils. We then evaluated effectiveness of these three attractive oils and their combinations with P. interpunctella sex pheromone in a grain warehouse. Traps baited with these oils captured significantly more females and males compared to control traps without attractants. However, sex pheromone addition did not improve male capture and significantly reduced female capture, suggesting an inhibitory effect. Our findings highlight the potential of blended, olive, or sesame oil trap alone as alternative monitoring and trapping tools for P. interpunctella, delivering novel insights into the development of efficient integrated pest management strategies for stored products.
Keywords:
attractant
; Plodia interpunctella
; sex pheromone
; storage pests
; vegetable oil
1. Introduction
Ensuring global food security is a paramount challenge requiring efficient storage and preservation practices. Stored product pests, particularly insects, cause significant quantitative and qualitative losses on stored grains, diminishing their economic value [1,2]. The Indian meal moth, Plodia interpunctella (Hübner) (Lepidoptera: Pyralidae), is a major polyphagous pest damaging stored grains and processed agricultural products worldwide [3], with reports of emergence even on the Antarctic Peninsula [4,5]. It has been reported that P. interpunctella causes damage to many stored products, including herbs, legumes, dried fruits, nuts, cereals, powdered milk, chocolate, birdseed, and dry pet food [6,7,8].
Infestations by P. interpunctella have substantial economic consequences due to direct product loss, pest control costs, and consumer complaints arising from product contamination. Larval feeding and the resulting production of silk and frass (insect excrement) are the primary causes of damage. This degrades product quality, promotes fungal growth, and ultimately leads to spoilage [9]. Contaminated products become intermixed with insect remains and waste products, significantly reducing their germination rate, nutritional value, and overall safety. This has translated to substantial economic losses in the United States, China, and many European countries [10,11]. Furthermore, P. interpunctella can potentially serve as a vector for various pathogens, including bacteria, viruses, fungi, protozoa, and helminths, and has been linked to human allergies such as asthma and skin diseases [12]. Its generalist feeding habits, diverse diet, and high capacity to damage food products have demonstrated its status as a globally recognised pest of significant concern [3,11].
Management of P. interpunctella infestations relies on various tactics, including irradiation [13], application of diatomaceous earth and fungal pathogens [14], insect growth regulators [15], and fogging [16]. Fumigation [17], introduction of parasitoid wasps [18], extreme temperature treatments [19,20], and pheromone-based approaches have also been employed [21]. The major components of sex pheromone of female P. interpunctella are (Z,E)-9,12-tetradecadienyl acetate (Z9,E12–14:OAc) and (Z,E)-9,12-tetradecadienol (Z9,E12–14:OH) [22,23,24], with the highest trapping efficacy at a 7:3 ratio at a 0.5 mg dose [25]. Pheromone-based technologies like monitoring traps, mass trapping, and mating disruption show promise for P. interpunctella management [26,27,28,29,30]. However, due to limitations like single-sex targeting and low catch rates [3,8,26], it is necessary to explore the integration of pheromones with other approaches, such as food-based attractants [31,32,33] to improve efficacy of pest management.
Insects may show specific behavioural responses to various olfactory cues from their social environments [34,35,36,37,38,39,40,41]. Food-derived volatile compounds can be particularly attractive to stored-product pest insects. For example, Storgard Oil, a blended vegetable-based oil with a proprietary composition [42], has been shown to broadly attract many stored product beetle species [42,43,44]. Studies also show that P. interpunctella can be attracted to traps baited with Storgard Oil [45], and prefers wheat kernels treated with walnut oil for oviposition [46]. These findings highlight the potential of vegetable-based oils as attractant lures for P. interpunctella. However, several questions remain unexplored: (1) Can a single, commercially available vegetable-based oil effectively attract both female and male P. interpunctella adults? (2) Would using blended oils or combining vegetable oils with sex pheromone enhance the attractancy? (3) Is there a sex-specific response to these vegetable oil attractants?
This study addresses these knowledge gaps by conducting two experiments under different environmental conditions. First, a semi-field experiment examined the attraction of P. interpunctella females and males to traps baited with 10 commercially available vegetable oils, i.e., camellia, corn, olive, peanut, rapeseed, sesame, soybean, sunflower, walnut, and a blended oil (mixture of sunflower seed, rapeseed, soybean, peanut, and sesame oils). Second, a field experiment explored the attractancy of traps baited with effective oils selected from the semi-field experiment, both alone and in combination with sex pheromone. Knowledge of this study delivers insights into the development of novel, environment-friendly, and cost-effective monitoring and trapping tools for the management of P. interpunctella.
2. Materials and Methods
2.1. Insects and Test Oils
We collected hundreds of P. interpunctella adults from the Zhoushan Reserve and Transit Grain Deport in Zhejiang Province, and reared the resultant larvae in transparent plastic boxes (26.0 cm length × 20.0 cm width × 12.5 cm height), with pinholes made in lids of boxes for ventilation. Each box housed 400 larvae on 100 g standard diet consisting of bran middlings, glycerol, and yeast with a 10:1:1 ratio [47]. We maintained the colony at 25 ± 1 °C and 60 ± 10% RH with a photoperiod of 16 h light and 8 h dark.
In this study, 10 commercially available vegetable oils (their corresponding suppliers) were used for experiments: Camellia (Lu Hua), corn (Jin Long Yu), olive (Olivolià), peanut (Lu Hua), rapeseed (Lu Hua), sesame (Chu Bang), soybean (Jin Long Yu), sunflower (Lu Hua), walnut (Dao Zi Shu Le), and a blended oil (Jin Long Yu, mixture of sunflower seed, rapeseed, soybean, peanut, and sesame oils).
2.2. Semi-Field Experiment
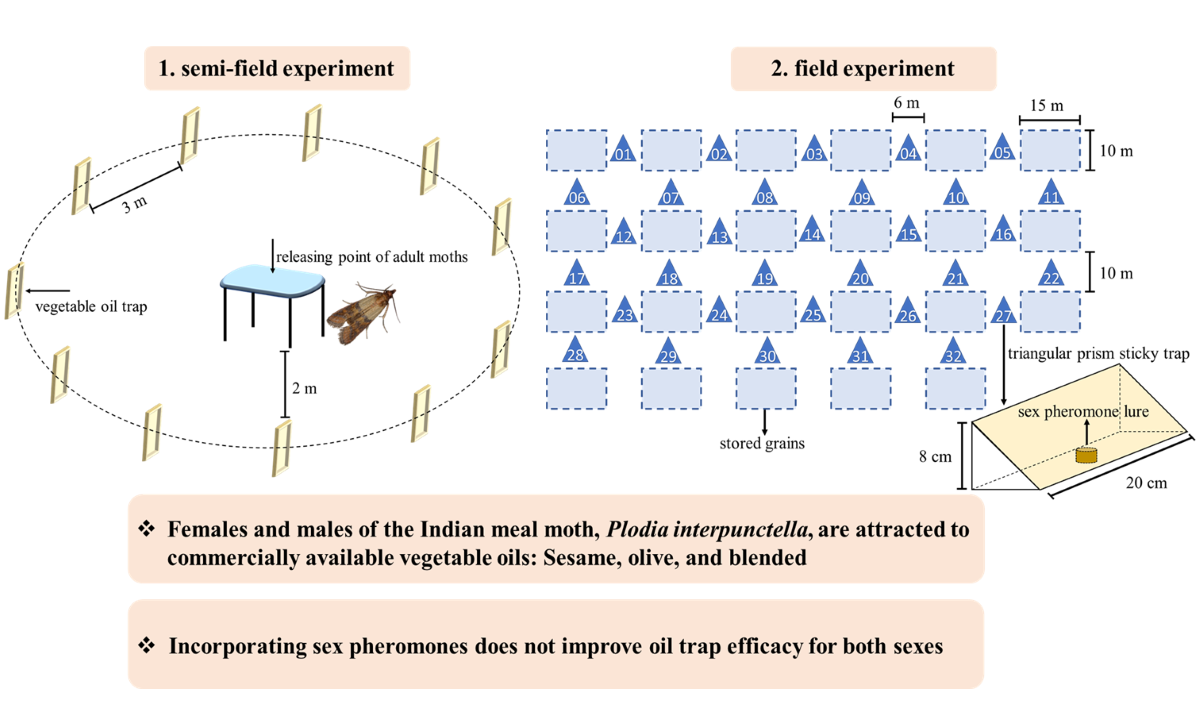
The semi-field experiment was conducted in July 2017 in a controlled environmental room (5.0 m length × 5.0 m width × 3.5 m height) located at the Zhoushan Reserve and Transit Grain Deport. The environmental conditions were the same as that for the laboratory colony. Thirty minutes before the scotophase, we prepared the oil baits by mixing 50 ml of each test oil with 75 ml of analytical grade n-Hexane. The oil baits were then applied to traps (i.e., insect sticky boards, 50.0 cm length × 20.0 cm width). The trap coated with 125 ml of hexane solvent only was used as control.
Eleven traps were numbered and randomly allocated along a loop shape with a radius of approximately 2 m from the moth releasing point (a platform) in the centre of the room (Figure 1). The traps and releasing point were 1.5 m above the floor, and the trap interval was approximately 3 meters between. Ten minutes before the scotophase, 30 female and 30 male adults (1–3 days old) randomly collected from the colony were placed on the releasing point. Moths were released soon after lights off and the room was secured. To eliminate the possible effect of trap location on the attractiveness, we rotated the traps clockwise every 8 hours over the subsequent 6 days, so that each trap occupied every position within the loop at least once. The number of trapped adult moths was recorded at the end of experiment and their sex was identified according to morphological characteristics [3]. We repeated the experiment three times, and the room was fully ventilated between each repetition to eliminate any accumulated odours.
2.3. Field Experiment
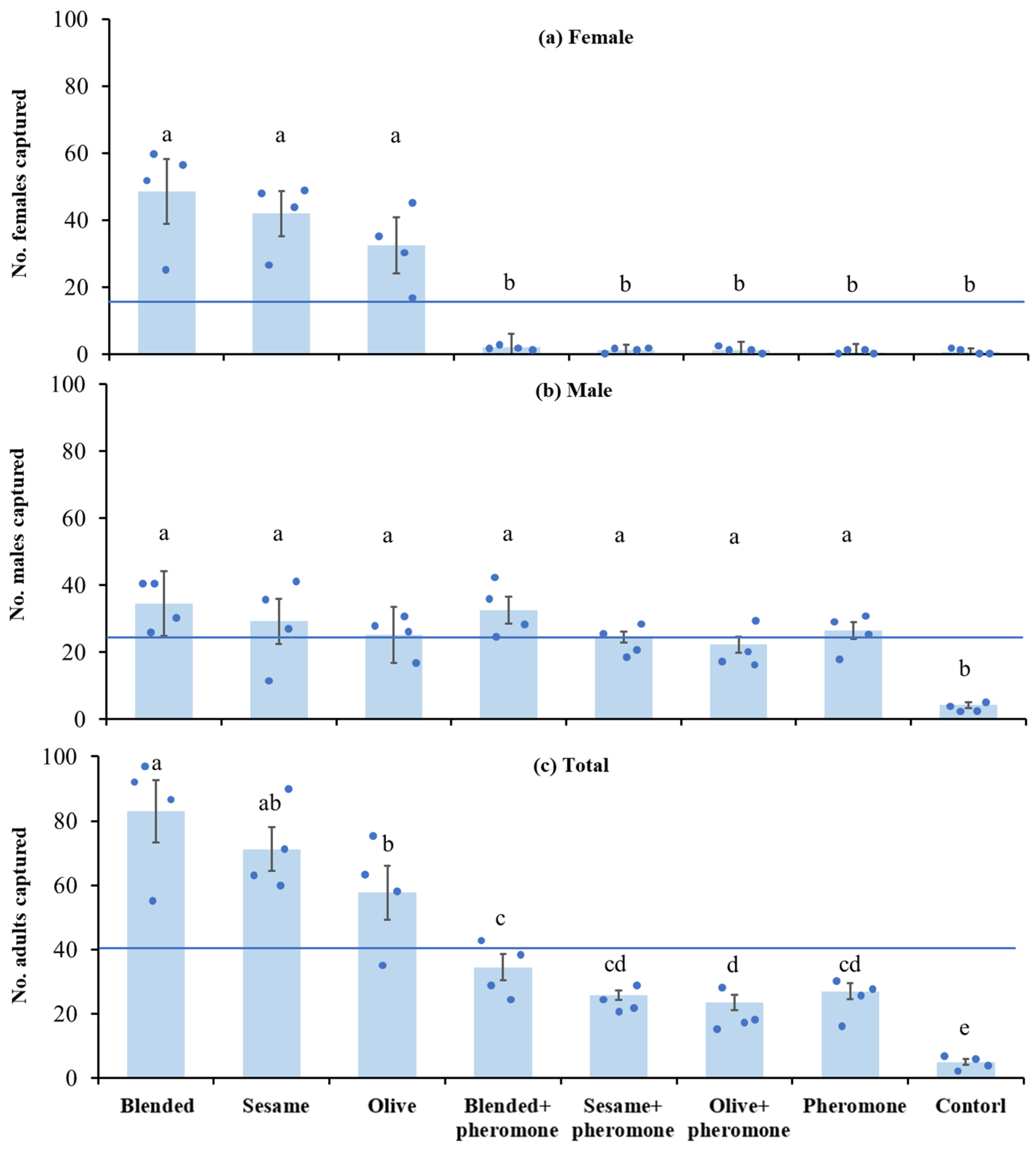
Basing on the high attractiveness of sesame, olive, and blended oils demonstrated in the semi-field experiment (see Figure 3), we conducted a field experiment to investigate potential additive effects of combining these oils with the P. interpunctella sex pheromone lure.
We prepared the oil baits as above and applied them to triangular prism sticky traps (20 cm long, side length 8 cm; Pherobio Technology Co., Ltd.; Figure 2). In the combination traps, a female sex pheromone lure containing a 0.5 mg mixture (3:7 ratio) of Z9,E12–14:OAc and Z9,E12-14:OH compounds [25] (Shin-Etsu Chemical Co., Ltd., Tokyo, Japan) was placed in the centre button of the trap. This resulted in 8 trap treatments: (1) olive oil only, (2) olive oil with sex pheromone, (3) sesame oil only, (4) sesame oil with sex pheromone, (5) blended oil only, (6) blended oil with sex pheromone, (7) sex pheromone only, and (8) control (no oil or sex pheromone). There were four replicates for each trap treatment with 32 traps in total.
This experiment was carried out in August 2017 within a P. interpunctella-infested warehouse (100.0 m length × 150.0 m width × 12.0 m height) at the Zhoushan Reserve and Transit Grain Deport. The warehouse stored various grain seeds and were ventilated based on the weather conditions. Thirty-two traps were randomly allocated to the arrangement as shown in Figure 2. Traps were 1.5 m above the floor and rotated in order every eight hours over twelve days to ensure each trap occupying each of the 32 positions for at least once. At the end of experiment, the number of trapped moths was recorded and females and males were identified.
2.4. Statistical Analysis
All data were analysed using SAS v.9.13 software (SAS Institute, Cary, NC, USA) with α = 0.05. The raw data from the semi-field experiment and natural-logarithm data [ln(x)] from the field experiment were normally distributed (Shapiro-Wilk test, UNIVARIATE procedure). An analysis of variance (ANOVA) (GLM procedure) was thus applied to compare the difference in number of moths captured between the different baits with a least significant difference (LSD) for multiple comparisons. A paired-t test (TTEST procedure) was used to compare the difference in number of moths captured between sexes for a given trap type in both semi-field and field experiments.
3. Results
3.1. Vegetable Oils Attractiveness to P. interpunctella in Semi-Field Conditions
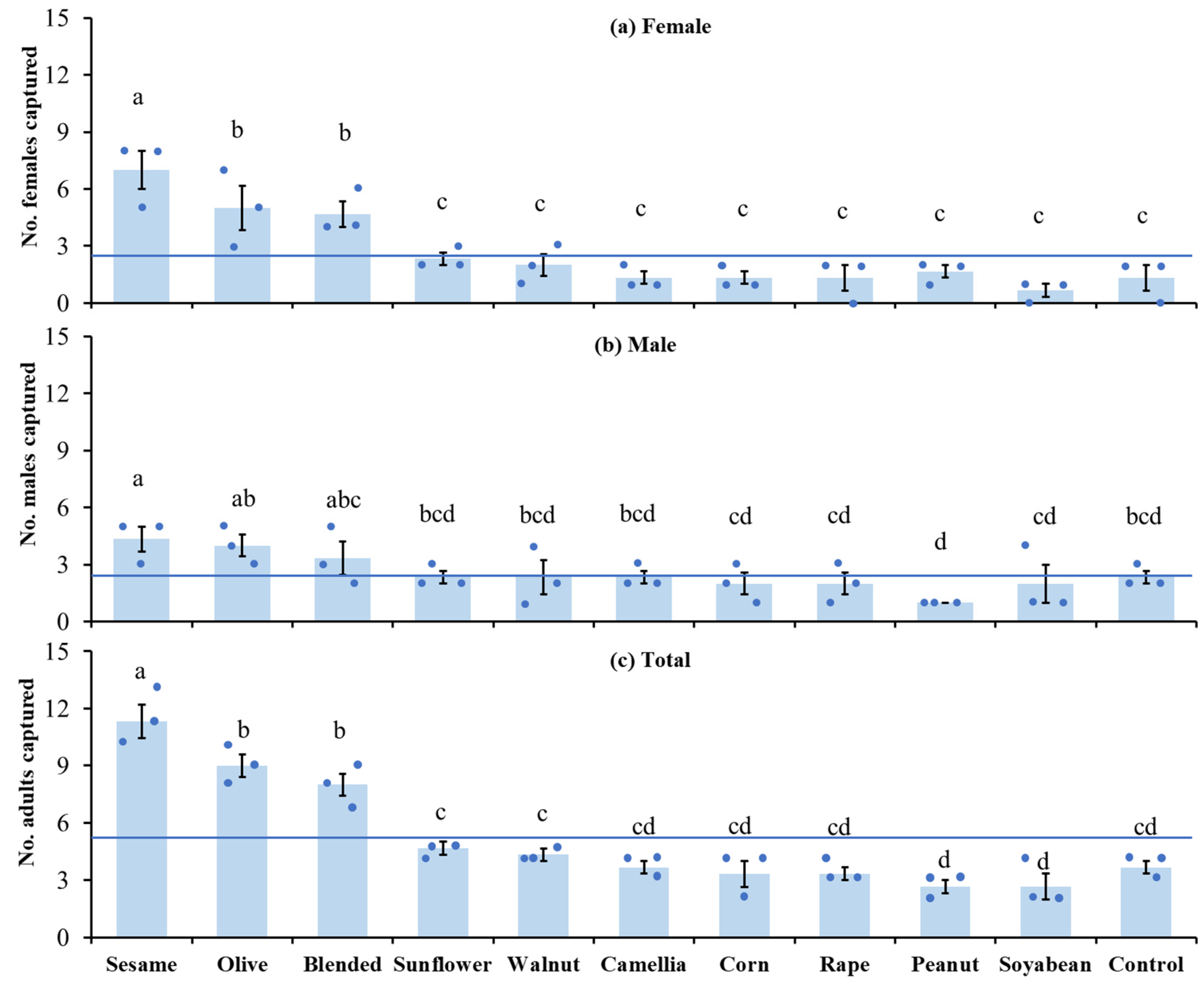
In the semi-field conditions, sesame oil trap captured significantly more female moths than did olive and blended oil traps, with significantly fewer females captured by the other oil and control traps (F10,22 = 9.86, p < 0.0001) (Figure 3a). The sesame, olive, and blended oil traps captured significant more male moths compared to peanut oil trap (F10,22 = 2.38, p = 0.0429) (Figure 3b). Overall, sesame oil trap had significantly higher efficacy in attracting adult moths than did olive and blended oil traps, and olive and blended oil traps captured significantly more adult moths than the sunflower and walnut oil traps, with significantly fewer adult moths captured by the peanut and soyabean oil traps (F10,22 = 31.32, p < 0.0001, Figure 3c). There was no significant difference in number of moths captured between sexes for all trap treatments (t3 = 0–1.73, p > 0.05 for all comparisons).
Figure 3.
The mean (± SE) number of Plodia interpunctella females (a), males (b), and adults (c) captured on sticky traps baited with 10 commercially available vegetable oils in semi-field conditions. Blended refers to the mixture of sunflower, rapeseed, soybean, peanut, and sesame oils. The horizontal line represents the mean number of trapped insects in all treatments. Each dot represents the capture number for a single replicate. Bars with different letters indicate statistically significant differences between oil treatments (LSD test: p < 0.05).
Figure 3.
The mean (± SE) number of Plodia interpunctella females (a), males (b), and adults (c) captured on sticky traps baited with 10 commercially available vegetable oils in semi-field conditions. Blended refers to the mixture of sunflower, rapeseed, soybean, peanut, and sesame oils. The horizontal line represents the mean number of trapped insects in all treatments. Each dot represents the capture number for a single replicate. Bars with different letters indicate statistically significant differences between oil treatments (LSD test: p < 0.05).
3.2. Combined Effects of Vegetable Oil and Sex Pheromone on Plodia interpunctella Captures in Field Conditions
In the field, blended, olive, and sesame oil traps captured significantly more female moths than all other traps (F7,24 = 55.59, p < 0.0001); however, pheromone alone or its combination with blended, olive, or sesame oil did not significantly increase the attraction to the females compared to the control (p > 0.05 for all comparisons) (Figure 4a). Blended, olive, sesame oil, and pheromone alone captured significantly more males than the control (F7,24 = 19.19, p < 0.0001); however, the combination of blended, olive, and sesame oil with pheromone did not significantly increase male captures compared to the blend, olive, sesame, or pheromone alone (p > 0.05 for all comparisons) (Figure 4b).
Regardless of the sex of moths, the total number of moths captured was significantly higher in blended, olive, and sesame oil traps; and although pheromone alone or its combination with blended, olive, or sesame oil did not significantly increase the number of moths captured, they captured significantly more moths than the control (F7,24 = 50.89, p < 0.0001) (Figure 4c). The blended, olive, or sesame oil traps trapped similar number of male and female moths (t3 = 1.29–2.70, p > 0.05 for all comparisons), while pheromone alone or its combination with blended, olive, or sesame oil traps captured significantly more males than females (t3 = 6.59–13.66, p < 0.05 for all comparisons) (Figure 4a,b).
4. Discussion
This study investigated the olfactory attraction of P. interpunctella adults to various commercially available vegetable oils under the semi-field and field conditions. Our results demonstrate that readily available olive, sesame, and blended oils (mixture of sunflower seed, rapeseed, soybean, peanut, and sesame oils) were attractive to both female and male P. interpunctella adults (Figure 3 and Figure 4). This is the first evidence on sex-specific attraction of P. interpunctella to vegetable oils, as previous work by Morrison et al. [45] only implied attraction of P. interpunctella to traps baited with Storgard Oil, a blended vegetable-based oil of undisclosed composition. The volatile compounds in vegetable oils, particularly certain fatty acids, may function as olfactory cues signaling potential food sources to stored product pests [46,48], thus showing attraction to P. interpunctella (in this study) and various stored-product pest beetles [42,43,44]. Such behavioural responses may enhance females’ likelihood of finding suitable oviposition sites for oviposition, and offspring feeding and development, ultimately improving reproductive fitness [46,49,50], as well as, increase the chance of males encountering females, thereby maximising mating opportunities and reproductive success [51,52]. Exploiting these innate responses of insect pests to food source cues, therefore, provides a foundation for the development of food-based attractant technology for stored product pest monitoring and control [53,54,55].
Combining pheromones with food cues (e.g., vegetable oils) is a common strategy to enhance the efficacy of pheromone-based insect pest monitoring and trapping [31,32,33]. This approach will enable the simultaneous attraction of both sexes. However, our results show that the addition of sex pheromones to the effective oil traps did not improve trapping efficacy for P. interpunctella (Figure 4c), but rather it significantly reduced the capture rate compared to that of the oil trap alone (Figure 4a). This unexpected outcome was probably attributed to the inhibitory effect of sex pheromone on female attraction. The observed repellent effect on females strongly suggests that P. interpunctella females were capable of detecting their sex pheromone, a phenomenon known as autodetection [56,57,58]. This autodetection behaviour may serve various functions, such as promoting resource conservation [59], increasing mating success [60] and progeny survival [61], and reducing resource competition among offspring [62] and possibility of predation [58,63]. The lack of enhanced male capture rate in traps baited with both vegetable oil and sex pheromone (Figure 4b) remains unclear and warrants further investigation. Based on these findings, we recommend that the blended, olive, or sesame oil trap alone rather than combining them with sex pheromone will be applied to manage P. interpunctella in storages for both practical and economic reasons.
Our findings regarding the attraction of sesame and blended oils to P. interpunctella differed between semi-field and field conditions. In the semi-field experiment, sesame oil traps captured significantly more adults than the blended oil ones (Figure 3c). However, in the field experiment, the blended oil traps captured a similar number of adults compared to sesame oil ones (Figure 4c), suggesting an increased efficacy of blended oil in field environments. Field environments are inherently more complex compared to controlled semi-field conditions. This complexity could modify the effects of specific volatile components in the attractant lures. The representation of these volatiles can also be altered by factors such as differences in experimental exposure time [38] and concentration [39]. Studies have shown that the mixture of odours present in an environment can significantly impact insects’ ability to detect a specific odour, because the background odour may alter the representation of individual odours [39,64,65]. Riffell et al. [66] demonstrate that in the tobacco hornworm Manduca sexta, background odours can influence the neuronal representation in the moth’s central olfactory system, leading to either excitation or inhibition of its ability to track the scent of the Datura wrightii flower bouquet.
Although P. interpunctella can infest a wide variety of stored goods, including the raw materials of the tested blended oil [6,7,8], olives and olive oil are not typically reported as susceptible commodities. Staff at the experimental site confirmed that the storage facilities were primarily used to store dry forms (seeds) of the blended oil components and their by-products, with minimal storage of olives or olive products. The attraction of P. interpunctella to olive oil could be due to its structural similarity to the oxygenated compounds found in other attractive oils (i.e., sesame and blended oils) or it represents a previously unknown attractant encountered in certain environments. Furthermore, our field experiment did not directly address the long-range orientation of P. interpunctella towards the test traps. Upon initiation of the experiment, moths were likely already present within or around the warehouse [10]. However, the significantly higher capture rate in traps baited with blended, sesame or olive oil alone compared to pheromone or control traps (Figure 4c) suggest that the volatile oil cues were perceivable at a long distance from the traps (Figure 2). Identifying the specific attractant compounds within these effective vegetable oils would be crucial for developing bisexual long-distance attractants for monitoring or trapping P. interpunctella in storages.
5. Conclusions
Our study highlights the potential of commercially available vegetable oils as attractants for both female and male adults of P. interpunctella. Identifying the specific attractant compounds within these oils will help improve food-based monitoring and trapping tools for this important stored-product pest. In addition, investigating the unexpected repellent effect of sex pheromones on females could develop novel management strategies that exploit autodetection behaviour for P. interpunctella control. Further research is needed to address the long-distance orientation of P. interpunctella towards vegetable-based attractants and the underlying mechanisms influencing the observed sex-specific responses. Overall, this study contributes valuable insights into the olfactory cues influencing P. interpunctella behaviour, providing novel knowledge for the development of safer and efficient integrated pest management strategies for pests of stored products.
Author Contributions
Conceptualization, G.Z. and J.D.; methodology, J.L., Z.Y. and J.D.; data curation, J.L.; formal analysis, J.L. and X.Z.H.; writing—original draft preparation, J.L.; writing—review and editing, X.Z.H., G.Z., M.G. and J.D. All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by the Zhejiang A&F University Research and Development Foundation (Grant No. 2024LFR057 and 2023LFR072), the Zhejiang Provincial Natural Science Foundation (Grant No. Y3100384), the “Pioneer” and “Leading Goose” R&D Program of Zhejiang (2022C02020), and the Major Science & Technology Task Tackling Program of Ningbo City (2022Z82).
Institutional Review Board Statement
Not applicable.
Data Availability Statement
Data are contained within the article.
Acknowledgments
We thank staff from the Zhoushan Reserve and Transit Grain Deport for their technical assistance.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design, data collection, analyses and interpretation, writing of the manuscript, or in the decision to publish the results.
References
- Arthur, F.H., Structural pest management for stored product insects. In Recent advances in stored product protection; Athanassiou, C.G., Arthur, F.H., Eds.; Springer: Berlin, Heidelberg, Germany, 2018; pp. 65‐81.
- Liang, J.; Zhang, X.; Zhang, Q.; Tang, Y.; Kong, W.; Zhang, J.; An, Y. Insect‐resistant activities of Elsholtzia ciliata essential oil and its main components carvone against Tribolium castaneum in different development stages. J. Stored Prod. Res. 2024, 107, 102317. [CrossRef]
- Mohandass, S.; Arthur, F.H.; Zhu, K.Y.; Throne, J.E. Biology and management of Plodia interpunctella (Lepidoptera: Pyralidae) in stored products. J. Stored Prod. Res. 2007, 43, 302–311. [Google Scholar] [CrossRef]
- Benitez, H.A.; Salinas, C.; Hernández, J.; Contador Mejías, T.; Kim, S.; Maturana, C.S.; Rebolledo, L.; Pérez, L.M.; Câmara, P.E.A.S.; Alves Ferreira, V.; Lobos, I.; Piñeiro, A.; Convey, P. An outsider on the Antarctic peninsula: A new record of the non-native moth Plodia interpunctella (Lepidoptera: Pyralidae). Ecol. Evol. 2024, 14, e10838. [Google Scholar] [CrossRef] [PubMed]
- Câmara, P.E.A.S.; Convey, P.; Ferreira, V.A.; Togni, P.H.B.; Pujol-Luz, J.R. First record of the Indian meal moth Plodia interpunctella (Lepidoptera: Pyralidae) at a research station in Antarctica. Antarct. Sci. 2022, 34, 361–364. [Google Scholar] [CrossRef]
- Razazzian, S.; Hassani, M.R.; Imani, S.; Shojai, M. Life table parameters of Plodia interpunctella (Lepidoptera: Pyralidae) on four commercial pistachio cultivars. J. Asia Pac. Entomol. 2015, 18, 55–59. [Google Scholar] [CrossRef]
- Jaafari-Behi, V.; Ziaee, M.; Kocheili, F.; Ali Hemmati, S.; Francikowski, J. Life-table parameters of Plodia interpunctella (Lepidoptera: Pyralidae) on different stored date palm fruits under laboratory conditions. J. Insect Sci. 2023, 23, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Pan, X.; Wang, C. Population dynamics of Indian meal moth in coffee bean warehouses in New Jersey. J. Stored Prod. Res. 2023, 100, 102044. [Google Scholar] [CrossRef]
- Rees, D.P. Insects of stored product. Csiro Publishing: London, UK, 2004.
- Adler, C.; Athanassiou, C.; Carvalho, M.O.; Emekci, M.; Gvozdenac, S.; Hamel, D.; Riudavets, J.; Stejskal, V.; Trdan, S.; Trematerra, P. Changes in the distribution and pest risk of stored product insects in Europe due to global warming: Need for pan-European pest monitoring and improved food-safety. J. Stored Prod. Res. 2022, 97, 101977. [Google Scholar] [CrossRef]
- Zhao, J.; Song, C.; Ma, L.; Yan, X.; Shi, J.; Hao, C. The impacts of climate change on the potential distribution of Plodia interpunctella (Hübner) (Lepidoptera: Pyralidae) in China. Insects 2022, 13, 636. [Google Scholar] [CrossRef]
- Binder, M.; Mahler, V.; Hayek, B.; Sperr, W.R.; Schöller, M.; Prozell, S.; Wiedermann, G.; Valent, P.; Valenta, R.; Duchêne, M. Molecular and immunological characterization of arginine kinase from the Indianmeal moth, Plodia interpunctella, a novel cross-reactive invertebrate pan-allergen. J. Immunol. 2001, 167, 5470–5477. [Google Scholar] [CrossRef]
- Hallman, G.J.; Phillips, T.W. Ionizing irradiation of adults of Angoumois grain moth (Lepidoptera: Gelechiidae) and Indianmeal moth (Lepidoptera: Pyralidae) to prevent reproduction, and implications for a generic irradiation treatment for insects. J. Econ. Entomol. 2008, 101, 1051–1056. [Google Scholar] [CrossRef] [PubMed]
- Sabbour, M.; Abd-El-Aziz, S.; Marwa, S. Efficacy of three entomopathogenic fungi alone or in combination with diatomaceous earth modifications for the control of three pyralid moths in stored grains. J. Plant Prot. Res. 2012, 52, 559–363. [Google Scholar] [CrossRef]
- Oh, H.W.; Yun, C.S.; Jeon, J.H.; Kim, J.A.; Park, D.S.; Ryu, H.W.; Oh, S.R.; Song, H.H.; Shin, Y.; Jung, C.S.; Shin, S.W. Conifer diterpene resin acids disrupt juvenile hormone-mediated endocrine regulation in the Indian meal moth Plodia interpunctella. J. Chem. Ecol. 2017, 43, 703–711. [Google Scholar] [CrossRef] [PubMed]
- Small, G.J. A comparison between the impact of sulfuryl fluoride and methyl bromide fumigations on stored-product insect populations in UK flour mills. J. Stored Prod. Res. 2007, 43, 410–416. [Google Scholar] [CrossRef]
- Jenson, E.A.; Arthur, F.H.; Nechols, J.R. Methoprene and synergized pyrethrins as aerosol treatments to control Plodia interpunctella (Hübner), the Indian meal moth (Lepidoptera: Pyralidae). J. Stored Prod. Res. 2010, 46, 103–110. [Google Scholar] [CrossRef]
- Trematerra, P.; Oliviero, A.; Savoldelli, S.; Schöller, M. Controlling infestation of a chocolate factory by Plodia interpunctella by combining mating disruption and the parasitoid Habrobracon hebetor. Insect Sci. 2017, 24, 503–510. [Google Scholar] [CrossRef] [PubMed]
- Arthur, F.H.; Phillips, T.W. Stored-product insect pest management and control. In Food Plant Sanitation; Hui, Y.H., Bruinsma, B.L., Gorham, J.R., Nip, W.K., Tong, P.S., Ventresca, P., Eds.; Marcel Dekker: New York, USA, 2003; pp. 341–358. [Google Scholar]
- Mason, L.J.; Strait, C.A., Stored product integrated pest management with extreme temperatures. In Temperature Sensitivity in Insects and Application in Integrated Pest Management; Hallman, G.J., Denlinger, D.L., Eds.; CRC Press: Boca Raton, USA, 2019; pp. 141–177. [CrossRef]
- Trematerra, P. Advances in the use of pheromones for stored-product protection. J. Pest Sci. 2012, 85, 285–299. [Google Scholar] [CrossRef]
- Kuwahara, Y.; Casida, J.E. Quantitative analysis of the sex pheromone of several phycitid moths by electron-capture gas chromatography. Agric. Biol. Chem. 1973, 37, 681–684. [Google Scholar] [CrossRef]
- Kuwahara, Y.; Kitamura, C.; Takashi, S.; Hara, H.; Ishii, S.; Fukami, H. Sex pheromone of the almond moth and the Indian meal moth: Cis-9,trans-12-tetradecadienyl acetate. Science 1971, 171, 801–802. [Google Scholar] [CrossRef]
- Sower, L.L.; Vick, K.W.; Tumlinson, J.H. (Z,E)-9,12-Tetradecadien-1-ol: A chemical released by female Plodia interpunctella that inhibits the sex pheromone response of male Cadra cautella. Environ. Entomol. 1974, 3, 120–122. [Google Scholar] [CrossRef]
- Yu, Z.; Deng, J.; Wang, Z.; Fu, W.; Zhou, G.; Lin, H.; Cao, Y.; Zheng, X.; Tang, J. Effect of different sex pheromone formulas, trap types and lure sources on captures of Plodia interpuntella. J. Chin. Cereals Oils. Assoc. 2018, 33, 6. [Google Scholar] [CrossRef]
- Trematerra, P.; Athanassiou, C.; Stejskal, V.; Sciarretta, A.; Kavallieratos, N.; Palyvos, N. Large-scale mating disruption of Ephestia spp. and Plodia interpunctella in Czech Republic, Greece and Italy. J. Appl. Entomol. 2011, 135, 749–762. [Google Scholar] [CrossRef]
- Campos, M.; Phillips, T.W. Attract-and-kill and other pheromone-based methods to suppress populations of the Indianmeal moth (Lepidoptera: Pyralidae). J. Econ. Entomol. 2014, 107, 473–480. [Google Scholar] [CrossRef] [PubMed]
- Fadamiro, H.Y.; Baker, T.C. Pheromone puffs suppress mating by Plodia interpunctella and Sitotroga cerealella in an infested corn store. Entomol. Exp. Appl. 2002, 102, 239–251. [Google Scholar] [CrossRef]
- Burks, C.S.; McLaughlin, J.R.; Miller, J.R.; Brandl, D.G. Mating disruption for control of Plodia interpunctella (Hübner) (Lepidoptera: Pyralidae) in dried beans. J. Stored Prod. Res. 2011, 47, 216–221. [Google Scholar] [CrossRef]
- Morrison III, W.R.; Scully, E.D.; Campbell, J.F. Towards developing areawide semiochemical-mediated, behaviorally-based integrated pest management programs for stored product insects. Pest Manag. Sci. 2021, 77, 2667–2682. [Google Scholar] [CrossRef] [PubMed]
- Athanassiou, C.G.; Kavallieratos, N.G.; Sciarretta, A.; Trematerra, P. Mating disruption of Ephestia kuehniella (Zeller) (Lepidoptera: Pyralidae) in a storage facility: Spatio-temporal distribution changed after long-term application. J. Stored Prod. Res. 2016, 67, 1–12. [Google Scholar] [CrossRef]
- James, D.G.; Moore, C.J.; Faulder, R.J. An improved coattractant for pheromone trapping of Carpophilus spp. (Coleoptera: Nitidulidae). Aust. J. Entomol. 1998, 37, 357–361. [Google Scholar] [CrossRef]
- Figueroa-Castro, P.; López-Martínez, V.; Silva-García, F.; González-Hernández, H. Food attractants to increase pheromone-baited trap performance for Scyphophorus acupunctatus (Coleoptera: Dryophthoridae) in mezcal maguey. Fla. Entomol. 2017, 100, 203–205. [Google Scholar] [CrossRef]
- Liu, J.; He, X.Z.; Wang, Q. Male larval experience of cues from adult rivals alters lifetime sperm investment patterns in a sperm heteromorphic moth, Ephestia kuehniella. Insect Sci. 2023, 30, 1773–1783. [Google Scholar] [CrossRef]
- Liu, J.; He, X.Z.; Zheng, X.L.; Zhang, Y.; Wang, Q. Juvenile socio-sexual experience determines lifetime sperm expenditure and adult survival in a polygamous moth, Ephestia kuehniella. Insect Sci. 2023, 30, 232–240. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; He, X.Z.; Zheng, X.L.; Zhang, Y.; Wang, Q. Larval social cues influence testicular investment in an insect. Curr. Zool. 2022, 68, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; He, X.Z.; Zheng, X.L.; Zhang, Y.; Wang, Q. Pupal cues increase sperm production but not testis size in an insect. Insects 2021, 12, 679. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Zhang, Y.; Zheng, X.L.; He, X.Z.; Wang, Q. Combined cues of male competition influence spermatozoal investment in a moth. Funct. Ecol. 2020, 34, 1223–1234. [Google Scholar] [CrossRef]
- Zhou, J.; Zhang, Z.; Liu, H.; Guo, M.; Deng, J. Inhibition effect of non-host plant volatile extracts on reproductive behaviors in the diamondback moth Plutella xylostella (Linnaeus). Insects 2024, 15, 227. [Google Scholar] [CrossRef]
- Deng, J.; Lan, C.; Zhou, J.; Yao, Y.; Yin, X.; Fu, K.; Ding, X.; Guo, W.; Liu, W.; Wang, N.; Wang, F. Analysis of sex pheromone production and field trapping of the Asian corn borer (Ostrinia furnacalis Guenée) in Xinjiang, China. J. Integr. Agric. 2023, 22, 1093–1103. [Google Scholar] [CrossRef]
- Deng, J.; Shen, Z.; Wang, F.; Liu, T.; Hong, W.; Fang, M.; Wo, L.; Chu, S. Enhancement of attraction to sex pheromone of Grapholita molesta (Busck) (Lepidoptera: Tortricidae) by structurally unrelated sex pheromone compounds of Conogethes punctiferalis (Guenée) (Lepidoptera: Crambidae). J. Asia Pac. Entomol. 2022, 25, 101859. [Google Scholar] [CrossRef]
- Doud, C.W.; Phillips, T.W. Responses of red flour beetle adults, Tribolium castaneum (Coleoptera: Tenebrionidae), and other stored product beetles to different pheromone trap designs. Insects 2020, 11, 733. [Google Scholar] [CrossRef] [PubMed]
- Athanassiou, C.G.; Kavallieratos, N.G.; Campbell, J.F. Effect of the presence of live or dead insects on subsequent captures of six stored-product beetle species: The relative species matters. J. Econ. Entomol. 2017, 110, 770–775. [Google Scholar] [CrossRef]
- Doud, C.W.; Cuperus, G.W.; Kenkel, P.; Payton, M.E.; Phillips, T.W. Trapping Tribolium castaneum (Coleoptera: Tenebrionidae) and other beetles in flourmills: Evaluating fumigation efficacy and estimating population density. Insects 2021, 12, 144. [Google Scholar] [CrossRef]
- Morrison, W.R., III; Agrafioti, P.; Domingue, M.J.; Scheff, D.S.; Lampiri, E.; Gourgouta, M.; Baliota, G.V.; Sakka, M.; Myers, S.W.; Athanassiou, C.G. Comparison of different traps and attractants in 3 food processing facilities in Greece on the capture of stored product insects. J. Econ. Entomol. 2023, 116, 1432–1446. [Google Scholar] [CrossRef] [PubMed]
- Nansen, C.; Phillips, T.W. Ovipositional responses of the Indianmeal moth, Plodia interpunctella (Hübner) (Lepidoptera: Pyralidae) to oils. Ann. Entomol. Soc. Am. 2003, 96, 524–531. [Google Scholar] [CrossRef]
- Cook, P.A.; Gage, M.J.G. Effects of risks of sperm competition on the numbers of eupyrene and apyrene sperm ejaculated by the moth Plodia interpunctella (Lepidoptera, Pyralidae). Behav. Ecol. Sociobiol. 1995, 36, 261–268. [Google Scholar] [CrossRef]
- Phelan, L. Evolution of sex pheromones and the role of asymmetric tracking. In Insect Chemical Ecology: An Evolutionary Approach; Roitberg B.D., Isman, M.B., Eds.; Chapman Hall, New York, USA, 1992; pp. 265–314.
- Yang, X.; Wang, M.; Gu, Y.; Han, W.; Li, X.; Li, X.; Zhong, Y.; Gao, J. The oviposition preference and offspring performance of Aethina tumida (Coleoptera: Nitidulidae). J. Econ. Entomol. 2024, 117, 696–704. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V.R.; Agavekar, G.; Agashe, D. Fitness landscapes reveal context-dependent benefits of oviposition behavior. Evolution 2022, 77, 550–561. [Google Scholar] [CrossRef] [PubMed]
- Fujiwara-Tsujii, N.; Yasui, H.; Wakamura, S. Population differences in male responses to chemical mating cues in the white-spotted longicorn beetle, Anoplophora malasiaca. Chemoecology 2013, 23, 113–120. [Google Scholar] [CrossRef]
- Nishida, Y.; Takagi, M. Male bull-headed shrikes use food caches to improve their condition-dependent song performance and pairing success. Anim. Behav. 2019, 152, 29–37. [Google Scholar] [CrossRef]
- Gregg, P.C.; Del Socorro, A.P.; Henderson, G.S. Development of a synthetic plant volatile-based attracticide for female noctuid moths. II. Bioassays of synthetic plant volatiles as attractants for the adults of the cotton bollworm, Helicoverpa armigera (Hübner) (Lepidoptera: Noctuidae). Aust. J. Entomol. 2010, 49, 21–30. [Google Scholar] [CrossRef]
- He, W.; Zhao, X.; Ali, A.; Ge, S.; Zhang, H.; He, L.; Wu, K. Population dynamics and reproductive developmental analysis of Helicoverpa armigera (Lepidoptera: Noctuidae) trapped using food attractants in the field. J. Econ. Entomol. 2021, 114, 1533–1541. [Google Scholar] [CrossRef]
- Ogawa, Y.; Miyake, T. How do rewardless Bletilla striata flowers attract pollinators to achieve pollination? Plant Syst. Evol. 2020, 306, 78. [Google Scholar] [CrossRef]
- Saad, A.D.; Scott, D.R. Repellency of pheromones released by females of Heliothis armigera and H. Zea to females of both species. Entomol. Exp. Appl. 1981, 30, 123–127. [Google Scholar] [CrossRef]
- Pérez-Aparicio, A.; Ammagarahalli, B.; Gemeno, C. A closer look at sex pheromone autodetection in the Oriental fruit moth. Sci. Rep. 2022, 12, 7019. [Google Scholar] [CrossRef] [PubMed]
- Holdcraft, R.; Rodriguez-Saona, C.; Stelinski, L.L. Pheromone autodetection: Evidence and implications. Insects 2016, 7, 17. [Google Scholar] [CrossRef] [PubMed]
- Yasui, H.; Fukaya, M.; Wakamura, S.; Akino, T.; Yasuda, T.; Kobayashi, A.; Arakaki, N. Aggregation of the black chafer Holotrichia loochooana loochooana (Sawada) (Coleoptera: Scarabaeidae): Function of female pheromone and possible adaptive significance. Appl. Entomol. Zool. 2007, 42, 507–515. [Google Scholar] [CrossRef]
- Pearson, G.A.; Dillery, S.; Meyer, J.R. Modeling intra-sexual competition in a sex pheromone system: how much can female movement affect female mating success? J. Theor. Biol. 2004, 231, 549–555. [Google Scholar] [CrossRef]
- Stelinski, L.; Rodriguez-Saona, C.; Meyer, W. Recognition of foreign oviposition-marking pheromone in a multi-trophic context. Sci. Nat. 2009, 96, 585–592. [Google Scholar] [CrossRef] [PubMed]
- Trematerra, P.; Battaini, F. Control of Ephestia kuehniella Zeller by mass-trapping. J. Appl. Entomol. 1987, 104, 336–340. [Google Scholar] [CrossRef]
- Walker, S.E.; Cade, W.H. A simulation model of the effects of frequency dependence, density dependence and parasitoid flies on the fitness of male field crickets. Ecol. Model. 2003, 169, 119–130. [Google Scholar] [CrossRef]
- Haverkamp, A.; Hansson, B.S.; Knaden, M. Combinatorial codes and labeled lines: How insects use olfactory cues to find and judge food, mates, and oviposition sites in complex environments. Front. Physiol. 2018, 9, 49. [Google Scholar] [CrossRef]
- Bruce, T.J.A.; Pickett, J.A. Perception of plant volatile blends by herbivorous insects – Finding the right mix. Phytochemistry 2011, 72, 1605–1611. [Google Scholar] [CrossRef]
- Riffell, J.A.; Shlizerman, E.; Sanders, E.; Abrell, L.; Medina, B.; Hinterwirth, A.J.; Kutz, J.N. Flower discrimination by pollinators in a dynamic chemical environment. Science 2014, 344, 1515–1518. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Arrangement of vegetable oil traps in the semi-field experiment.
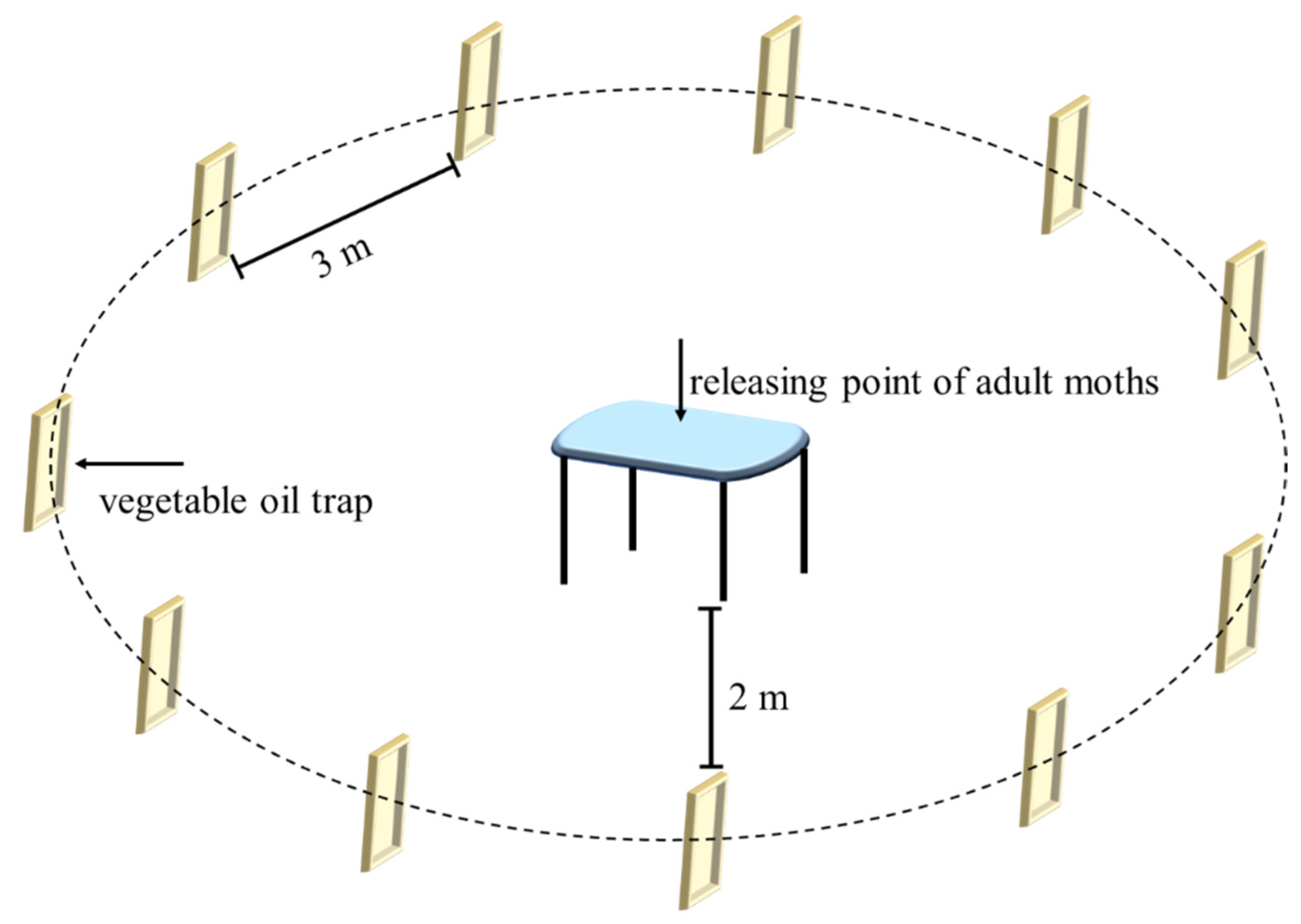
Figure 2.
Arrangement of vegetable oil and pheromone traps in the field experiment.
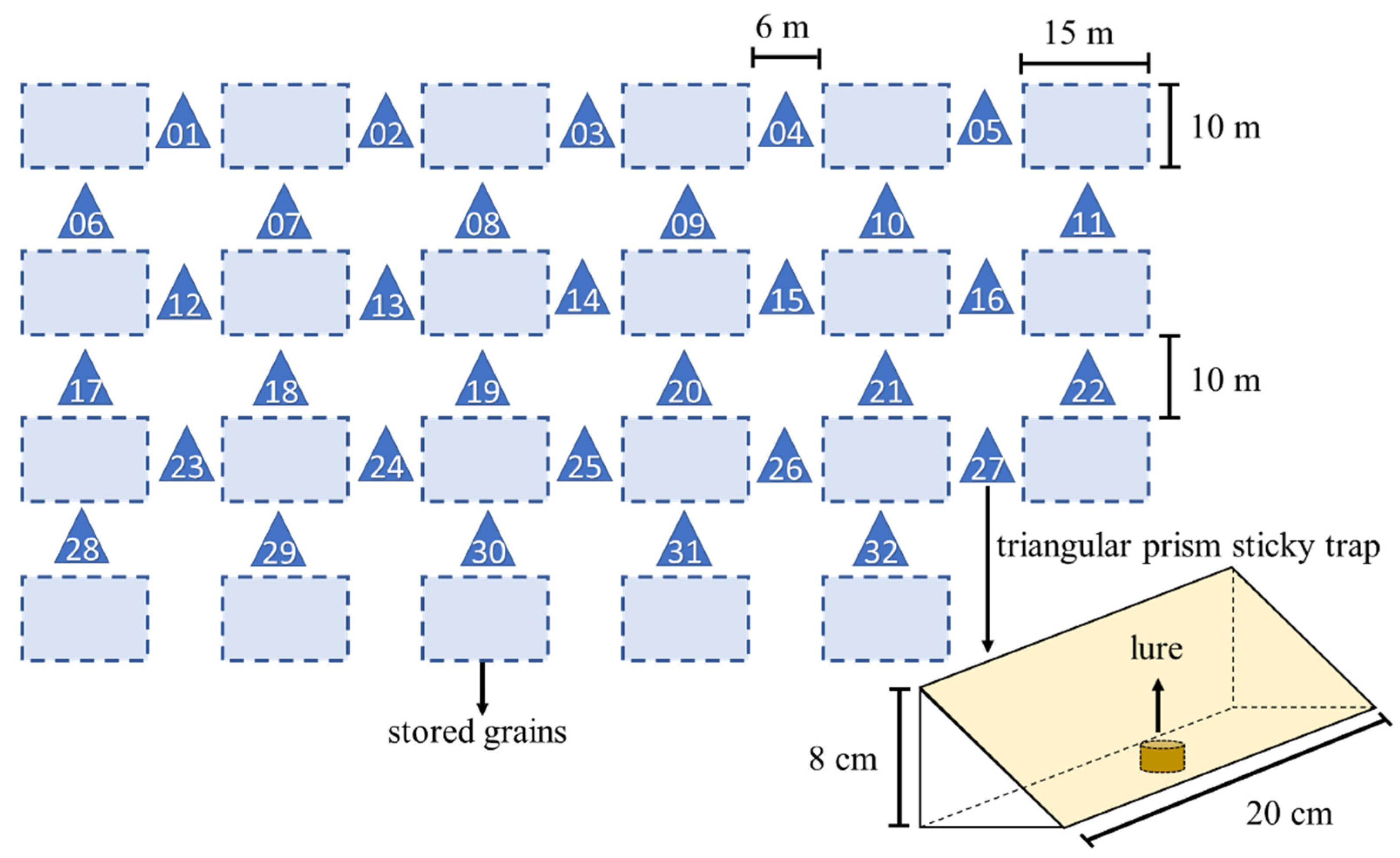
Figure 4.
The effect of vegetable oil and pheromone and their combination baits on the mean (± SE) number of Plodia interpunctella females (a), males (b), and all adults (c) captured on sticky traps in field conditions. Blended refers to mixture of sunflower, rapeseed, soybean, peanut, and sesame oils. The horizontal line represents the mean number of captured insects in all treatments. Each dot represents the capture number for a single replicate. Bars with different letters indicate statistically significant difference between traps (LSD test: p < 0.05).
Figure 4.
The effect of vegetable oil and pheromone and their combination baits on the mean (± SE) number of Plodia interpunctella females (a), males (b), and all adults (c) captured on sticky traps in field conditions. Blended refers to mixture of sunflower, rapeseed, soybean, peanut, and sesame oils. The horizontal line represents the mean number of captured insects in all treatments. Each dot represents the capture number for a single replicate. Bars with different letters indicate statistically significant difference between traps (LSD test: p < 0.05).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



