
Submitted:
10 July 2024
Posted:
11 July 2024
You are already at the latest version
Abstract
The fish fauna of Rio de Janeiro has been extensively studied, resulting in a comprehensive database of species collected over more than three centuries. This study aimed to identify fish species, their locations, and compile scattered information to aid in climate action and freshwater conservation prioritization and an evaluation of the sampling effort to date, as well as to identify patterns of diversity and distribution of freshwater ichthyofauna, delineate biogeographic units, and explore similarity relationships between areas. Analyzing data from nearly 25 ichthyological collections and literature on original species descriptions revealed 346 fish species: 172 freshwaters native, 22 allochthonous, and 152 marine species. The checklist includes updated species names. The sampling effort in Rio de Janeiro is high, especially in coastal lowlands. The findings indicate that inventory work is still needed in certain areas. Five bioregions of freshwater ichthyofauna were identified, along with six major areas of higher species richness. This biogeographic assessment underscores the diverse and distinctive freshwater fish fauna in the basins of Rio de Janeiro, with well-defined biogeographic units.
Keywords:
distribution
; endemism
; ichthyofauna
; sampling effort
; streams
1. Introduction
The state of Rio de Janeiro possesses unique characteristics of the Atlantic Forest Biome, comprising a diverse array of ecological niches shaped by proximity to the coast, varied relief, soil types, and rainfall regimes. These factors have driven the evolution of a rich biotic complex, encompassing both forest and aquatic systems. In the mountainous areas, the Rio Paraíba do Sul stands out to the west in its middle courses, embedded in the Serra da Mantiqueira, and lower course towards the coast. To the east, smaller coastal rivers descend the slopes of the Serra do Mar. The coastal region is distinguished by sandbanks, dunes, mangroves, swampy forests, ponds, and swamps. In the central and southern portions of Rio de Janeiro, bays occupy the coastal lowlands, notably Guanabara Bay, Sepetiba Bay, and Ilha Grande Bay. The remaining rivers and streams regulate water flow, ensure soil fertility, control the climate, and protect escarpments and mountain slopes. In the northern region, lake systems dominate the landscape, with many lagoons such as Maricá, Saquarema, Araruama, and Feia.
Recent knowledge about rivers and fish in the southeastern Atlantic Forest territory is extensive [1,2], yet there remain fewer contributions that evaluate the complete set of species for specific areas within the Atlantic Forest [3,4,5]. A comprehensive understanding of the species composition across the heterogeneous regional landscapes is still lacking. Thus, updating and disseminating knowledge about this crucial biodiversity set is a priority.
Collections as repositories stimulate curiosity about species diversity, leading to the pursuit of answers to new questions. The fish fauna of Rio de Janeiro has been extensively studied [6,7,8,9,10,11,12,13,14], resulting in the creation of a robust database of species in museums and university collections over more than three centuries. However, there is a gap of comprehensive information. Investigating the fish species inhabiting each region, consolidating scattered information, and making it easily accessible is now more indispensable than ever, particularly for disseminating and valuing freshwater ecosystems in the face of rapid climate change. This study highlights the rivers of Rio de Janeiro, identifies the respective basins in a territorial context, and evaluates the distribution patterns of their fish species.
2. Materials and Methods
Hydrographic region 08 of Brazil corresponds to an area adapted from the Ottobasins division by Pfafstetter [15,16]. This territorial division, named Southeastern Atlantic, comprises river basins flowing into the Atlantic Ocean. It is bordered to the north by the hydrographic basin of the Rio São Mateus in Espírito Santo state and to the south by the hydrographic basin of the Rio Ribeira do Iguape in Paraná state. The hydrographic region is bordered to the west by the hydrographic regions of the São Francisco and Paraná [17]. Within hydrographic region 08, each river basin is recognized by a number, with number 1 being the northernmost river, the Rio São Mateus. Accordingly, each river basin in Rio de Janeiro is identified by a number between 20 and 38, in parentheses, as detailed below.
2.1. Study Area
The Rio de Janeiro state is separated into nine hydrographic areas – a political division towards governance of its territory [18], as follows: RH-1- Ilha Grande Bay drainage; RH-2- Rio Guandu and transposition with Paraíba do Sul (08.22e); RH-3- Middle stretch of Rio Paraiba do Sul basin (08.22d); RH-4- Rio Piabanha and tributaries at Paraíba do Sul basin (08.22c); RH-5- Guanabara bay drainage; RH-6- Lake region and Rio São João; RH-7- Rio Dois Rios and tributaries at Paraíba do Sul basin (08.22b); RH-8- Rio Macaé and Rio das Ostras; RH-9- Lower Rio Paraíba do Sul and Itabapoana (08.22a) (Figure 1).
The river basins along the Rio de Janeiro territory were divided into 24 groups (Figure 2). Among those areas five belongs to the Rio Paraíba Sul basin, as stated above (RH-9, RH-7, RH-4, RH-3 and RH-2). Additionally 19 are coastal basins and microbasins: Itabapoana Basin (08.20- Figure 2- A), São Francisco de Itabapoana Watersheds (08.21- Figure 2- B), Lagoa Feia Watersheds (08.23- Figure 2- H), Jurubatiba Watersheds (08.24- Figure 2-I), Macaé River Basin (08.25- Figure 2-J), Rio das Ostras Watersheds (08.26- Figure 2-K), São João River Basin (08.27- Figure 2-L), Una River Basin and Búzios Watersheds (08.28- Figure 2-M), Araruama Lagoon Watersheds (08.29- Figure 2-N), Saquarema Watersheds (08.30- Figure 2-O), Maricá Watersheds (08.31- Figure 2-P), Niterói Watersheds (08.32 Figure 2-Q), Guanabara Bay Watersheds (08.33 Figure 2-R), Rio de Janeiro Watersheds (08.34- Figure 2-S), Sepetiba Watersheds (08.35 Figure 2-T), Mangaratiba Watersheds (08.36 Figure 2-U), Angra dos Reis Watersheds (08.37- Figure 2-V), Paraty Watersheds (08.38- Figure 2-W) and Cairuçu Watersheds (08.39- Figure 2-X).
2.2. Species Data
Available records in fish collections from the Rio de Janeiro were consulted and had the identification confirmed. Each of the sampling points at the area had coordinates estimated from the locations indicated in the records. Initially, a curatorial survey of data was carried out in the fish collections of the Museu Nacional (MNRJ) and fish collection at the Museu de Biologia Mello Leitão (MBML). In the same way, records available in the database – SpeciesLink – Centro de Referência em Informação Ambiental [Reference Center for Environmental Information – CRIA] were consulted, and additionally the literature of species descriptions through which the data available in the collections of the AMNH, CAS (including CAS-SU), DZSJRP, FMNH, INPA, MCP, MCZ, MNRJ, MTD, MZFS, MZUEL, MZUSP, NPM, NRM, UF, UFRGS, UFRN, UFRJ, UFMT, UMMZ, UMZC, UNT, USNM and ZUEC. were inventoried (Supplementary file- Table S1). These institutional acronyms follow [19]. The records in collections were geo referred and plotted on a map of the Rio de Janeiro state. A total of 13,327 lots collected on the continent were inventoried (Supplementary file- Table S2).
The taxonomic classification of fishes follows [20]. Taxonomic problems are commented under section results.
2.3. Geographic Data
In georeferencing, for correspondence of the coordinates of each point, the values reported in the respective database were first used. When a small discrepancy was found between the reported value and the reported location, this value was adjusted according to the indication of the location. In cases when there was a large discrepancy between the indicated coordinates and the location, or when these coordinates were not available, the coordinates were estimated considering the available location information.
The hydrographic maps were adapted for use in the TrackMaker software [21], starting with version 1:25,000 made available by Instituto Brasileiro de Geografia e Estatística (IBGE) [22]. In the suggested location, information was used according to the names assigned to the rivers in this version, complemented by the names available in the IBGE topographic maps of 1:50,000. Municipality areas were calculated based on IBGE data [23].
2.4. Sampling Coverage Assessment
To evaluate the quality of sampling in the state, the index of total lots (il) per 100 km² and the index of sampling points (ip) per 100 km² were calculated. These indices were calculated for each municipality, hydrographic region, and group of basins, and these results were compared with the index for the entire state [24]. The sampling quality was considered average when found in the range of about 30%, poor when significantly below 30%, and good when above 30% (for both ilq and ipq). We evaluated the quality of sampling for the nine hydrographic regions, for the twenty-nine river basin divisions and additionally by municipality.
2.5. Biogeographic and Diversity Patterns
We applied the constancy index [25], used to determine which species are constant on a time scale: C = n/N * 100, where n = number of collection points where the species were captured and N = total number of collection points. Based on the results obtained, each species was classified as: constant, when C > 50%; accessory, when C varies between 25% and 50%; and accidental when C < 25%.
To assess differences in the fish fauna present in each hydrographic region of the study area the diversity indices absolute richness, Shannon diversity, equitability, and dominance were calculated, using species abundance data for each region [26]. Additionally, we traced diversity profiles, using the Rényi series, to compare the diversity among hydrographic regions. In the Rényi series, the minimum value of α results in an index that is equal to species richness (equability is not considered), and very high values of α assess only equability and disregard species richness [27]. In other words, we can only say that one river basin region is more diverse than another if the curves do not intersect. These analyses were performed using the PAST software [28].
Since determining the total number of species in an area is virtually impossible, especially in regions with high species richness, estimators are useful for extrapolating the observed richness and attempting to estimate the total richness through an incomplete sample of a biological community [29]. Consequently, we employed diversity estimators to assess the completeness of the species sampling for the hydrographic regions: the Chao 1 index, a simple estimator of the absolute number of species in a community. It is based on the number of rare species within a sample [30,31]; the iChao index estimator, which brought greater precision to the evaluation of the results [32]; and the AC estimator, Abundance-based Coverage Estimator [33]. This method works with the abundance of rare species (i.e. low abundance) [33,34]. However, unlike the previous estimator, this method allows the researcher to determine the limits for which a species is considered rare. In general, species with an abundance of between 1 and 10 individuals are considered rare. The estimated richness may vary as the abundance threshold is raised or lowered, and unfortunately there are no defined biological criteria for choosing the best range. Lastly, we adopted the Squares index, a richness estimator [35], designed to be more accurate than Chao-1 when abundance distributions are uniform.
We exclusively considered records of native freshwater species for the following spatial assessments. A bioregionalization analysis was performed using the Infomap Bioregions 2 algorithm [36], to subdivide the state of Rio de Janeiro into smaller biogeographical units. This algorithm uses species distribution data, even in cases of inconsistent sampling efforts. It employs an adaptive resolution method that generates a bipartite network of species and grids, followed by a clustering analysis to create bioregions based on the presence of specific taxa. The following parameters were employed: cell size ranging from 1/4° to 1° and cell capacity ranging from 5 to 1000 samples, 500 trials. The remaining settings followed the program defaults. We performed a cluster analysis using the binary matrix resulting from the bioregionalization algorithm to verify the similarity in faunal compositions between the bioregions generated by the UPGMA algorithm, Jaccard's similarity coefficient, in the PAST software [28].
A species richness interpolation analysis was performed to map diversity patterns using the spline interpolation method, which smooths out potential sampling gaps by creating a continuous surface of data values, in the BioDinamica model [37] of the Dinamica-EGO software [38]. The following parameters were used: raster grid size = 0.03, smoothing factor = 10, minimum of one sample per hexagon, using a delimitation mask of the Rio de Janeiro state.
3. Results
3.1. Rio de Janeiro According to the Collections - Diversity in Numbers
Lots sampled. The records of 16,299 lots from the collections were analyzed. Of these, 13,327 lots (81.77%) were sampled in continental waters of the Rio de Janeiro state. In addition, 2,890 (17.73%) were in the oceanic area and 82 (0.50%) could not be evaluated in its respective municipality. Of the lots sampled in inland waters, 985 (7.4%) are of marine origin, 498 (3.7%) are allochthonous freshwater, 11,724 (88.0%) are native to freshwater and 120 (0.9%) were not identified at the species level (Table 1).
From a geographical point of view, from the records in continental waters, it was possible to georeference 13,327 lots (81.74%), leaving 86 lots (0.53%) for which it was only possible to identify them at the state level.
Sampling index. The average sampling index based on lots of freshwater fish recorded for the state of Rio de Janeiro was 30.5 lots per 100 km². The average index of sampling points on the continent was 3.7 sampling points per 100 km².
Regarding the hydrographic regions (Table 2), three were considered poorly sampled: RH-3 (Middle Paraíba do Sul) with il = 12.1 and ip = 1.9; RH-7 (Rio Dois Rios) with il = 15.8 and ip = 1.6, and RH-9 (lower Paraíba do Sul and Itabapoana) with il = 12.8 and ip = 1.4. Four regions appear with a good number of lots and sampling points, and two are within the state average (Table 2, Figure 3).
Twenty-four municipalities (Table A1) appear with a good number of lots, 14 are within the state average and 54 appear with a poor number of lots. Regarding sampling points, there are 25 with a good number of points, 15 are within the state average and 52 have a poor number of sites (Figure 4). Eight municipalities (Armação dos Búzios, Arraial do Cabo, Belford Roxo, Macuco, Miracema, Porciúncula, São João de Meriti and Varre Sai) do not have records deposited in ichthyological collections for freshwater species, and the first two have only records of marine species.
In the evaluation of the basin groups (Table 3), in relation to the number of lots, 12 have a good number of lots, four are within the state average and eight have a poor number of lots. Regarding the sampling points, 11 have a good number of sites, five are within an average number and eight have a poor quality of sampling points.
Temporal variation. The growth of the records deposited in ichthyological collections after the year 2000 was 50% greater than all previous years (Figure 5). The earliest records are from 1832. From 1980 onwards, the collections had a significantly higher growth than in the entire previous period, continuously increasing since the year 2000.
3.1.1. Taxonomic Diversity
Entire set of records. Considering the entire set of records (Supplementary file- Table S3), 346 species were documented for the state of Rio de Janeiro (27 orders, 80 families and 207 genera), of which 172 are native freshwater species, 22 are allochthonous freshwater species and 152 are marine (Table 4). The orders Siluriformes (93 spp., 26.9% of the total) and Characiformes (50 spp., 14.5%) have the highest species richness, followed by Acanthuriformes and Cyprinodontidormes (33 spp. each, 9.5%). The most representative families of the total (Figure 5B) were Loricariidae (35 spp., 10.1%), Trichomycteridae (30 spp., 8.7%), Characidae (25 spp., 7.2%) and Rivulidae (21 spp., 6.1%).
Native freshwater records. Regarding native freshwater species (Figure 6, Table 5, Table A2) (eight orders, 26 families, 31 subfamilies and 82 genera), the orders Siluriformes and Characiformes were the most representative, with 86 (54.7% of the total freshwater species) and 40 (29.1%) species, respectively, followed by Cyprinodontiformes (32 species., 19.2%). The most representative freshwater families are Loricariidae (36 species, 20.9%), Trichomycteridae (30 species, 17.4%), Characidae (25 species, 14.5%) and Rivulidae (21 spp., 12.2%). The most representative native freshwater fish genera were Trichomycterus (21 species), Characidium, and Deuterodon (7 species).
Constancy index. A total of 1,680 sampling points with different coordinates were identified in the state of Rio de Janeiro. No species was classified as constant and only Geophagus brasiliensis appeared as an accessory species, having been recorded in 497 points (29.58%). The others were all accidental, such as Poecilia vivipara with 268 sites (16.95%) and Phalloceros harpagos with 267 (15.89%). It is noticeably that 72 species were collected in one sampling point only.
Considering only the native freshwater fishes, 1,553 sampling points with georrefered coordinates were identified. In this scenario of native species, again only Geophagus brasiliensis again appeared as an accessory species. The others were classified as accidental.
Non-parametric estimation and diversity indices. All species richness estimators indicate that the species catalogued for the hydrographic regions RH-1, RH-2 and RH-6 are within the expected range (Table 6). The greatest variation between the species catalogued for a region and the estimated species is in RH-9, which estimates the possibility of an increase of up to 28.0% (ACE) in the species richness indicated by the records, and in RH-4 with up to 25.7% (iChao-1).
The highest species richness was in the Guandu hydrographic region (RH-2), with 97 species, followed by the Baía de Guanabara hydrographic region (RH-5), with 89 species. The lowest richness was in the Baía da Ilha Grande hydrographic region (RH-1) with 40 species, the Médio Rio Paraíba do Sul hydrographic region (RH-3) and the Macaé e das Ostras hydrographic region (RH-8), both with 67 species. The Shannon index indicates to a greater diversity of RH-5 (H = 3.528), followed by the Rio Dois Rios hydrographic region (RH-7) (H = 3.516), and the Lagos São João hydrographic region (RH-6) (H = 3.366). The lowest diversity was identified at the Baixo Paraíba do Sul e Itabapoana hydrographic region (RH-9) (H = 2.513). The dominance index was higher in RH-9 (D = 0.1549), RH-8 (D = 0.1214), and the Piabanha hydrographic region (RH-4) (D = 0.1038). In contrast, the equitability index indicates that RH-9, RH-8 and RH-4 are the less even communities, and RH-7 the most even one.
The Guandu region (RH-2) was the one with the highest richness of native freshwater species with 97 species, followed by the Baía de Guanabara (RH-5) with 89 species, Baixo Paraíba do Sul e Itabapoana (RH-9) with 81 species, and Lagos São João (RH-6) with 80 species, a result consonant with the special interpolation of species richness. The Baía da Ilha Grande region (RH-1), on the other hand, presented the lowest richness, with only 40 species. The highest values of the Shannon diversity index were reached in hydrographic regions RH-2, RH-3, RH-5, RH-6, RH-7, areas also identified as hotspots of ichthyofaunistic biodiversity in Rio de Janeiro (see Figure 9[MI1] ).
The diversity profiles of the hydrographic regions using the Rényi series indicate that the diversity of the Baía da Ilha Grande region (RH-5) is lesser than the diversity of the Guandu region (RH-2) only when considering indices more dependent on species richness, but their diversities are very close when using the indices more dependent on equability (Figure 7a).
Despite the great difference between the richness of the hydrographic regions from the extremes south and north of Rio de Janeiro, Baía da Ilha Grande (RH-1, 40 spp.) and lower Paraíba do Sul and Itabapoana (RH-9, 81 spp.), the graph with the diversity curves (Figure 7b) indicates that it is not possible to define which region has the greatest diversity.
The diversity profile of the hydrographic regions that include only the tributaries of the Rio Paraíba do Sul (Middle Paraíba do Sul, RH-3; Piabanha, RH-4; Rio Dois Rios, RH-7) (Figure 7c) show a greater diversity in a lower part of Rio Paraíba do Sul (RH-7), while in the highest parts (RH-3, RH4) the crossing of the curves suggests that these diversities are not comparable and will alternate depending on the index that is considered.
The hydrographic region Baía de Guanabara (RH-5) is more diverse than the hydrographic region Lagos São João (RH-6), which in turn is more diverse than the hydrographic region Macaé and das Ostras (RH-8, Figure 7d).
Although Rio de Janeiro is a widely collected territory within the Atlantic Forest, this sampling is not equal. The quality indices of the sampled lots are good for an area varying between 20% and 32% of the state (according to the division per municipalities, basins or hydrographic regions), between 11% and 18% of the state area presented average sampling quality, and between 55% and 60% of the state presented poor sampling. These results suggest that there are still under-sampled areas in the territory that may be the target of future studies and inventories.
3.2. Spatial Patterns of Distribution
Five bioregions have been delineated based on patterns of distribution of the freshwater fish fauna (Figure 8A). These areas are herein referred to as Lower Rio Paraíba do Sul bioregion, Guanabara-Guandu bioregion, Middle Rio Paraíba do Sul bioregion, Costa Verde bioregion and Lagos bioregion.
- Lower Rio Paraíba do Sul bioregion: The largest area in terms of territorial extension, this biogeographic unit is composed mainly of the lower Rio Paraíba do Sul basin and its tributaries, such as the Rio Muriaé, Rio Dois Rios and Rio Pomba, as well as independent basins such as the Rio Itabapoana (the geographical divide with the state of Espírito Santo), Rio Macaé and Rio São João, and coastal lagoon systems such as the Lagoa Feia. Species that delimit this bioregion include Listrura tetraradiata, Microglanis pleriqueater, and Trichomycterus puriventris, present in most of the drainages that compose this area. The bioregion has 114 species, of which 26 (22.8%) occur only in this area (e.g., Atlantirivulus janeiroensis, A. jurubatibensis, Bryconamericus tenuis, Characidium litorale, Delturus parahybae, Homodiaetus banguela, Ituglanis parahybae, Trichomycterus caipora, T. fuliginosus and T. vitalbrazili). Covering a large part of the basins of the state of Rio de Janeiro, many of the species present in these basins are common to several other drainages of the Atlantic Forest, such as fish species within the genera Deuterodon, Hypostomus and Trichomycterus.
- Guanabara-Guandu bioregion: It consists mainly of the basins that drain the Baía de Guanabara and Baía de Sepetiba, such as the Rio Caceribu, Rio Guapimirim, Rio Macacu, Rio Roncador, Rio Suruí, Rio Guandu and the Jacarepaguá lagoon system. Species that delimit this bioregion include Kryptolebias caudomarginatus, Listrura nematopteryx, and Notholebias minimus. Presents 99 species, 17 of them (17.2%) occurring only in this area (e.g., Atlantirivulus guanabarensis, Australoheros macacuensis, Characidium grajahuense, Homodiaetus passarelli, Kryptolebias brasiliensis, Leptopanchax opalescens, L. sanguineus, L. splendens, Microglanis nigripinnis, Trichomycterus giganteus, and T. potschi).
- Middle Rio Paraíba do Sul bioregion: Formed by tributaries of the middle course of the Rio Paraíba do Sul, such as the Rio Piraí, Rio Paquequer, Rio Preto, Rio Piabanha, and the Rio Paraíba do Sul. This bioregion is supported by the presence of species such as Parotocinclus bidentatus, Trichomycterus nigroauratus, and Trichomycterus florensis. The area presents 81 species, of which 11 (13.6%) occur only in this bioregion, especially species of the genus Trichomycterus, such as T. itatiayae, T. macrophthalmus, T. mariamole, and T. mirissumba.
- Costa Verde bioregion: This biogeographic unit consists of small basins that flow into the Baía da Ilha Grande, as the drainages of the Rio Mambucaba, Rio Perequê-Açu, Rio Taquari and Rio Parati-Mirim. Supported by the presence of species such as Characidium japuhybense, Hemipsilichthys nimius, and Neoplecostomus paraty. The bioregion presents 40 species, of which nine (22.5%) occur only in this area (e.g., Atlantirivulus lazzarotoi, A. simplicis, Listrura costai, and Phalloceros enneaktinos).
- Lagos bioregion: The smallest of the delimited areas, this bioregion consists of small drainages and lagoon systems that are part of the Região dos Lagos area, in the coast of Rio de Janeiro. It includes the drainages of the Rio Ubatiba, Rio Mato Grosso, coastal wetlands and lagoons of Maricá and Saquarema. The region is supported by the presence of species of rivulids such as Atlantirivulus maricensis, Leptopanchax citrinipinnis, Nematolebias papilliferus, and Notholebias fractifasciatus. It has 36 species, of which four (11.1%) only occur in that area.
Figure 8.
Bioregions of the freshwater icthyofauna within the territory of Rio de Janeiro (A) and Jaccard similarity among these areas (B).
Figure 8.
Bioregions of the freshwater icthyofauna within the territory of Rio de Janeiro (A) and Jaccard similarity among these areas (B).
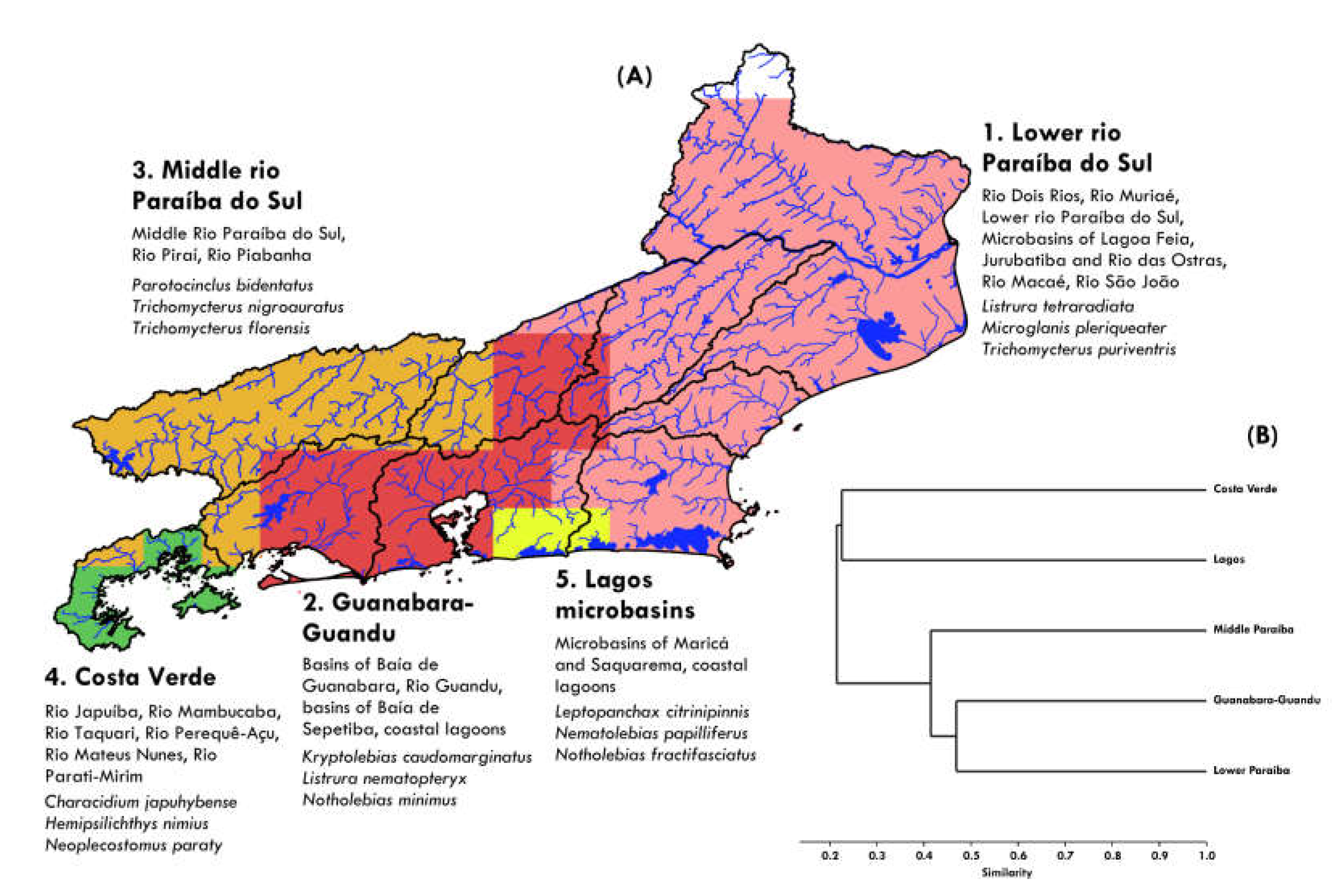
The cluster analysis (cophenetic correlation: 0.93) indicates that the Lower Paraíba do Sul and the Guanabara-Guandu bioregions have approximately 47% of ichthyofaunistic similarity, and both present about 40% of congruence with the Middle Paraíba do Sul region (Figure 8B). The Costa Verde and Lagos bioregions present greater dissimilarity with hydrographic networks of the state, showing less than 20% of global similarity.
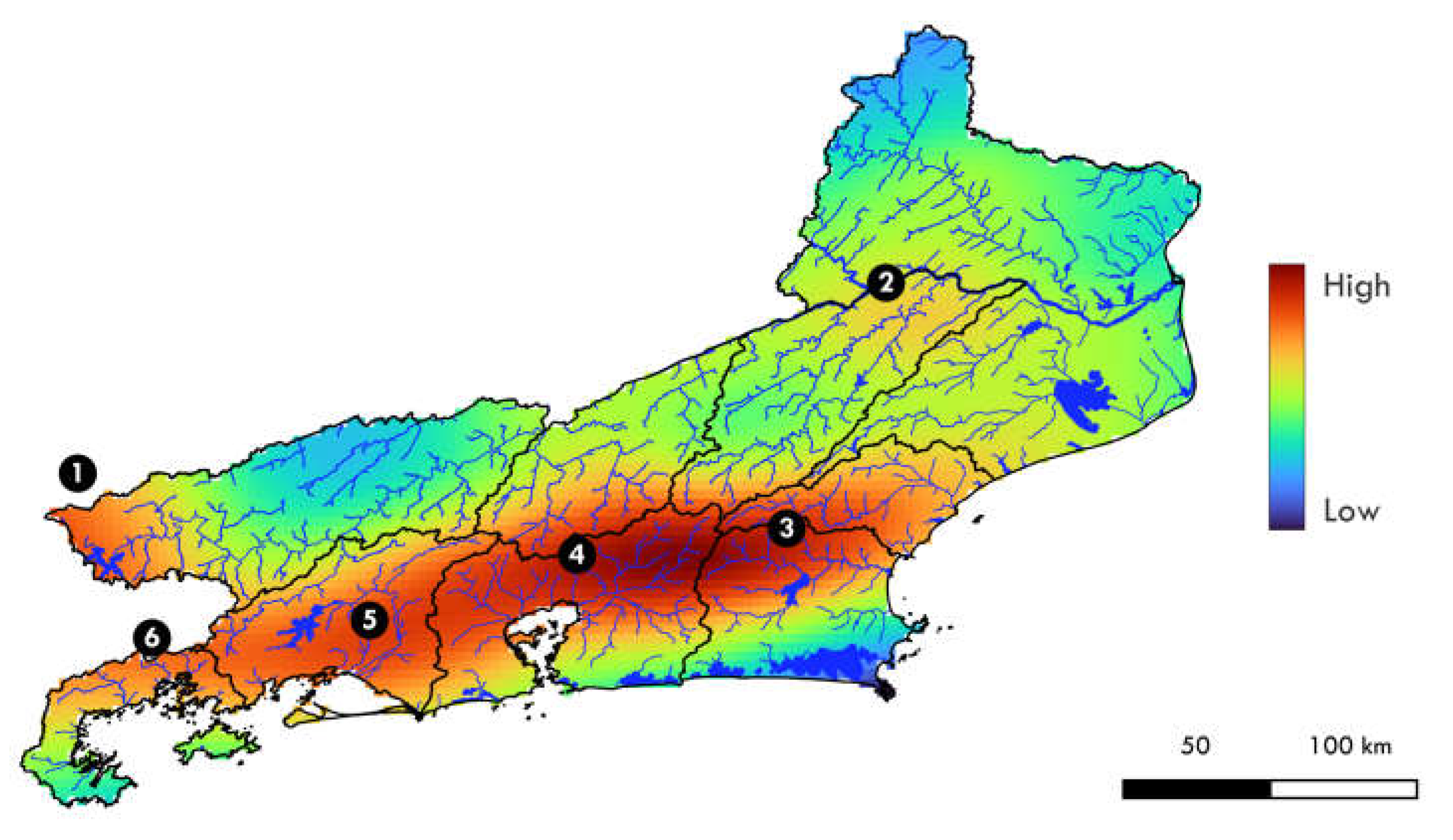
The interpolation of species richness shows six areas with the highest density of species in hydrographic regions of the state of Rio de Janeiro (Figure 9): (1) the middle Rio Paraíba do Sul, on the border with the state of São Paulo, in the vicinity of the Funil Reservoir, in the region of Itatiaia (RH-3); (2) the lower Rio Paraíba do Sul, in the region of the mouth of the Rio Dois Rios (RH-7); (3) the upper course of the Rio São João and Rio Macaé (RH-6 and RH-8); (4) several basins that flow into Baía de Guanabara (RH-5); (5) the Rio Guandu basin and adjacent basins (RH-2); and (6) Costa Verde basins (RH-1). In general, the region with the highest intensity of this index is specifically the sub-basin of the Rio Macacu and the Rio Guapimirim, followed by the upper Rio São João. In contrast, the regions with the lowest species richness were a stretch of the Middle Rio Paraíba do Sul, in the Rio das Flores basin; the upper course of the Rio Carangola and Rio Muriaé, in the Lower Rio Paraíba do Sul; and the region surrounding the lagoon system of Araruama and Cabo Frio.
Figure 9.
Interpolation of species richness within the state of Rio de Janeiro. The most relevant areas are highlighted with numbers 1-6.
Figure 9.
Interpolation of species richness within the state of Rio de Janeiro. The most relevant areas are highlighted with numbers 1-6.
4. Discussion
The perception that the rivers of the Atlantic Forest in Rio de Janeiro comprise sets of distinct areas of endemism for fish has been previously detailed by authors who have worked with the biome as a whole [39,40]. Subsequently, [41], in a global analysis of freshwater ecoregions, recognized three distinct ecoregions that include, in part, the territory of Rio de Janeiro: 329, Paraíba do Sul; 330, Ribeira de Iguape, and 352, Fluminense (see figure 1 in [41]). The division of Rio de Janeiro in bioregions of ichthyofaunal endemism recognized here roughly match the findings of [41]. The bioregions Lower Rio Paraíba do Sul and Middle Rio Paraíba do Sul partially adjusts to the ecoregion 329. The bioregion Costa Verde corresponds to part of the Ribeira do Iguape ecoregion. Additionally, the bioregions Guanabara-Guandu and Lagos almost corroborate the delineation of the Fluminense ecoregion [41].
The particularities of the relief and the configuration of the hydrographic basins in the Atlantic Forest biome in the territory of Rio de Janeiro include the Paraíba valley—depressed tectonic valley corridors along faults [42]—with rivers flowing between Serra do Mar and Serra da Mantiqueira Mountain ranges. Although the Rio Paraíba do Sul is the largest river system partially crossing the Rio de Janeiro territory, it was not recognized as the most diversified area for freshwater fishes. Possible reasons for this include the presence of numerous coastal drainages flowing from the Serra do Mar, as well as extensive areas of coastal lowlands with bays and lagoon systems. These diverse environments support a significant diversity of species. The mountainous nature with vertical escarpments and difficult access, that contributed to keep the mountain barrier impassable during the initial centuries of colonization [43], was also the feature that functioned as a geographical divide, isolating the Rio Paraíba do Sul from the coastal rivers. In the set of coastal lowlands, it is worth highlighting the special interest and diversity of freshwater species on the small river drainages at the Baía de Guanabara region. To mention its origin in a simplified way, Baía de Guanabara was geologically configured in the Cenozoic as a result of a tectonic depression, in a phenomenon known as block tipping, later filled by sediments derived from the erosion of adjacent mountain ranges, and in the Quaternary period drowned by the marine transgression that generated water mass of the bay [44]. Since the uplift of the mountains, the bay has been receiving sediments derived from the numerous rivers that drain into its interior. By its formation, the bay area holds a variety of living environments for freshwater forms, in different relief extracts. Additionally, the Sepetiba and Paraty bays were formed in a similar manner [42].
In the set of coastal lowlands, it is worth highlighting the special interest and diversity of freshwater species on the small river drainages at the Costa Verde bioregion. Despite corresponding to a small area of territory, the Costa Verde has a high endemism of species, with a unique set of stream fishes [14]. Being mostly fish of the first order, that is, intolerant of survival in brackish or salty waters, the process of isolation, dispersion and occupation of these species in the numerous small coastal streams has always been intriguing. The coastal drainages of the Costa Verde have particularities not observed in other areas, such as rivers flowing abruptly towards the coast, some of which are even devoid of coastal plain areas. Such topographic configuration of these small basins that flow from the mountains towards the sea has important implications for the aquatic biota, considering that isolated basins present similar fish fauna [45]. The coastal basins of the Atlantic Forest where the Serra do Mar is very close to the coast, such as those in the Costa Verde area, were influenced by climate change that caused oscillations in sea level during the Pleistocene. These transgressions and marine regressions generated isolation and connection between rivers [46]. The past connections among these coastal basins are hypothetized by [47] based on molecular evidence and paleodrainage reconstruction.
The orders Siluriformes and Characiformes were predominant in number of species, repeating a pattern commonly found in drainages of the Neotropical region and the Atlantic Forest biome [40,48], followed by Cyprinodontiformes. The predominance of these three orders is the result of characteristics that facilitate the occupation of species in different habitats and the great heterogeneity of environments available in the drainages of the state. This result is in line with several regional studies for Rio de Janeiro basins (e.g., [8,10,11,12,13,14,39,49].
Some species considered here have complex taxonomy, with phylogenies that need to be solved and probable species complexes, but we consider them as single taxa for analytic and inventory purposes. Rhamdia quelen has recently been redescribed and its distribution is now restricted to coastal basins from Rio de Janeiro south to the Rio Tubarão basin, in the state of Santa Catarina [50]. [51] describe seven subspecies of Gymnotus carapo for South American basins and suggest that the species appears to be absent from the coastal drainages of Northeastern and Southeastern Brazil, despite several records tentatively identified as G. carapo, G. aff. carapo or G. cf. carapo for these regions in museum collections and databases. Hoplias malabaricus and Synbranchus marmoratus most likely represent species complexes that require revision [52,53,54,55].
The greatest species richness in RH-2 (97 spp.) is associated with the fact that this hydrographic region encompassing coastal hydrographic systems is connected to the Rio Paraíba do Sul though links with the Rio Guandu system, which supplies water to the city of Rio de Janeiro. In contrast, the lowest species richness in RH-1 (40 spp.) is related to the geographic characteristics of this region, where the Serra do Mar is very close to the coast. The habitats are either characterized by steep rapids or estuarine plains.
5. Conclusions
This is the first time an evaluation of freshwater fishes in the Rio de Janeiro territory is accomplished with a geographical accuracy of data. We hope our results contribute for future research in the area and for conservation management measures of this complex and diverse territory. The distribution patterns of the fish species in the area corroborate previous studies of the Atlantic Forest and indicate that the recognized bioregions are in accordance with previous established biogeographic units.
The more sampled and diversified freshwater area —the vicinities of the Baía de Guanabara— may be due to the variety of environments in an area where rivers come from a slope mountainous area, the Serra dos Órgãos, with fast-flowing, clear water rivers and pebbles substrate, as well as swamps, slow flowing creeks and transitional environments such as mangroves in the coastal areas. Additionally, the proximity to the metropolitan area of Rio de Janeiro and its research centers historically facilitated the access. The less sampled regions in Rio de Janeiro corresponds to the middle reaches of Rio Paraíba do Sul at Rio das Flores, and also at its lower section, at traditional areas of coffee crops and nowadays mostly deforested, with river siltation and pollution.
Supplementary Materials
The following supporting information can be downloaded at: www.mdpi.com/xxx/s1, Table S1: Ichthyological collections with inventoried records from the state of Rio de Janeiro; Table S2: Records of fish species from the state of Rio de Janeiro listed for this study; Table S3: Inventory of all species recorded for the state of Rio de Janeiro.
Author Contributions
All authors designed the study; RFMP built the dataset and checked species distribution ranges. LMSS, FVG performed data curation and taxonomic validation. Biogeographical analysis was performed by FVG. Statistical analysis was performed by RFMP and FVG. All authors wrote the manuscript.
Funding
FGV was funded by CAPES (Coordenação de Aperfeiçoamento de Pessoal de Nível Superior, Finance Code 001, process no. 88887.512702/2020-00.
Data Availability Statement
Data is available in Supplementary Material Table S2.
Acknowledgments
We extend our gratitude to M. Gianeti, O. Oyakawa (MZUSP), M. R. Britto, (MNRJ), for providing collection records. We are grateful to M.R. Britto, P.A. Buckup (MNRJ) and J. P. da Silva (MBML) for the courtesy extended during the visit to the institutions. FVG is grateful to CAPES (Coordenação de Aperfeiçoamento de Pessoal de Nível Superior, Finance Code 001, process no. 88887.512702/2020-00) for his Ph.D. scholarship. This work received laboratory support from the Instituto Nossos Riachos (INR).
Conflicts of Interest
The authors declare no conflicts of interest.
Appendix A

Table A1.
Quality of sampling per municipality.
| Municipality | Area (km2) | Lots | Points | Il | Ilq | Ip | Ipq |
|---|---|---|---|---|---|---|---|
| Angra dos Reis | 813.21 | 541 | 56 | 66.5 | Good | 6.9 | Good |
| Aperibé | 94.54 | 2 | 2 | 2.1 | Poor | 2.1 | Poor |
| Araruama | 638.15 | 34 | 4 | 5.3 | Poor | 0.6 | Poor |
| Areal | 110.72 | 1 | 1 | 0.9 | Poor | 0.9 | Poor |
| Armação dos Búzios | 70.98 | 0 | 0 | 0.0 | Poor | 0.0 | Poor |
| Arraial do Cabo | 152.11 | 0 | 0 | 0.0 | Poor | 0.0 | Poor |
| Barra do Piraí | 584.61 | 41 | 8 | 7.0 | Poor | 1.4 | Poor |
| Barra Mansa | 547.13 | 40 | 7 | 7.3 | Poor | 1.3 | Poor |
| Belford Roxo | 78.99 | 0 | 0 | 0.0 | Poor | 0.0 | Poor |
| Bom Jardim | 382.43 | 19 | 5 | 5.0 | Poor | 1.3 | Poor |
| Bom Jesus do Itabapoana | 596.66 | 32 | 8 | 5.4 | Poor | 1.3 | Poor |
| Cabo Frio | 413.58 | 65 | 14 | 15.7 | Poor | 3.4 | Average |
| Cachoeiras de Macacu | 954.75 | 927 | 83 | 97.1 | Good | 8.7 | Good |
| Cambuci | 558.28 | 6 | 2 | 1.1 | Poor | 0.4 | Poor |
| Campos dos Goytacazes | 4 032.49 | 608 | 75 | 15.1 | Poor | 1.9 | Poor |
| Cantagalo | 747.21 | 48 | 6 | 6.4 | Poor | 0.8 | Poor |
| Carapebus | 304.89 | 293 | 20 | 96.1 | Good | 6.6 | Good |
| Cardoso Moreira | 522.60 | 24 | 3 | 4.6 | Poor | 0.6 | Poor |
| Carmo | 305.75 | 252 | 23 | 82.4 | Good | 7.5 | Good |
| Casimiro de Abreu | 462.92 | 343 | 32 | 74.1 | Good | 6.9 | Good |
| Comendador Levy Gasparian | 108.64 | 1 | 1 | 0.9 | Poor | 0.9 | Poor |
| Conceição de Macabu | 338.26 | 114 | 15 | 33.7 | Average | 4.4 | Average |
| Cordeiro | 113.05 | 8 | 1 | 7.1 | Poor | 0.9 | Poor |
| Duas Barras | 379.62 | 3 | 1 | 0.8 | Poor | 0.3 | Poor |
| Duque de Caxias | 467.32 | 233 | 38 | 49.9 | Good | 8.1 | Good |
| Engenheiro Paulo de Frontin | 139.38 | 31 | 4 | 22.2 | Average | 2.9 | Average |
| Guapimirim | 358.44 | 317 | 28 | 88.4 | Good | 7.8 | Good |
| Iguaba Grande | 50.98 | 7 | 2 | 13.7 | Poor | 3.9 | Average |
| Itaboraí | 429.96 | 127 | 10 | 29.5 | Average | 2.3 | Poor |
| Itaguaí | 282.61 | 167 | 16 | 59.1 | Good | 5.7 | Good |
| Italva | 291.19 | 18 | 2 | 6.2 | Poor | 0.7 | Poor |
| Itaocara | 433.18 | 118 | 8 | 27.2 | Average | 1.8 | Poor |
| Itaperuna | 1 106.69 | 319 | 16 | 28.8 | Average | 1.4 | Poor |
| Itatiaia | 241.04 | 230 | 38 | 95.4 | Good | 15.8 | Good |
| Japeri | 81.70 | 59 | 3 | 72.2 | Good | 3.7 | Average |
| Laje do Muriaé | 253.53 | 7 | 1 | 2.8 | Poor | 0.4 | Poor |
| Macaé | 1 216.99 | 886 | 82 | 72.8 | Good | 6.7 | Good |
| Macuco | 78.36 | 0 | 0 | 0.0 | Poor | 0.0 | Poor |
| Magé | 390.78 | 308 | 44 | 78.8 | Good | 11.3 | Good |
| Mangaratiba | 367.82 | 67 | 12 | 18.2 | Poor | 3.3 | Average |
| Maricá | 361.57 | 174 | 35 | 48.1 | Good | 9.7 | Good |
| Mendes | 95.32 | 15 | 2 | 15.7 | Poor | 2.1 | Poor |
| Mesquita | 41.17 | 1 | 1 | 2.4 | Poor | 2.4 | Poor |
| Miguel Pereira | 287.93 | 98 | 16 | 34.0 | Average | 5.6 | Good |
| Miracema | 303.27 | 0 | 0 | 0.0 | Poor | 0.0 | Poor |
| Natividade | 387.07 | 7 | 1 | 1.8 | Poor | 0.3 | Poor |
| Nilópolis | 19.39 | 1 | 1 | 5.2 | Poor | 5.2 | Good |
| Niterói | 133.76 | 35 | 8 | 26.2 | Average | 6.0 | Average |
| Nova Friburgo | 935.43 | 182 | 32 | 19.5 | Poor | 3.4 | Average |
| Nova Iguaçu | 520.58 | 479 | 58 | 92.0 | Good | 11.1 | Good |
| Paracambi | 190.95 | 24 | 3 | 12.6 | Poor | 1.6 | Poor |
| Paraíba do Sul | 571.12 | 15 | 4 | 2.6 | Poor | 0.7 | Poor |
| Paraty | 924.30 | 609 | 84 | 65.9 | Good | 9.1 | Good |
| Paty do Alferes | 314.34 | 8 | 2 | 2.5 | Poor | 0.6 | Poor |
| Petrópolis | 791.14 | 143 | 25 | 18.1 | Poor | 3.2 | Average |
| Pinheiral | 82.25 | 2 | 1 | 2.4 | Poor | 1.2 | Poor |
| Piraí | 490.26 | 120 | 17 | 24.5 | Average | 3.5 | Average |
| Porciúncula | 291.85 | 0 | 0 | 0.0 | Poor | 0.0 | Poor |
| Porto Real | 50.89 | 41 | 4 | 80.6 | Good | 7.9 | Good |
| Quatis | 284.83 | 51 | 8 | 17.9 | Poor | 2.8 | Poor |
| Queimados | 75.93 | 33 | 2 | 43.5 | Good | 2.6 | Average |
| Quissamã | 719.64 | 267 | 38 | 37.1 | Average | 5.3 | Good |
| Resende | 1 099.34 | 283 | 42 | 25.7 | Average | 3.8 | Average |
| Rio Bonito | 459.46 | 32 | 5 | 7.0 | Poor | 1.1 | Poor |
| Rio Claro | 846.80 | 719 | 74 | 84.9 | Good | 8.7 | Good |
| Rio das Flores | 478.78 | 10 | 2 | 2.1 | Poor | 0.4 | Poor |
| Rio das Ostras | 228.04 | 81 | 7 | 35.5 | Average | 3.1 | Average |
| Rio de Janeiro | 1 200.33 | 932 | 182 | 77.6 | Good | 15.2 | Good |
| Santa Maria Madalena | 810.96 | 84 | 19 | 10.4 | Poor | 2.3 | Poor |
| Santo Antônio de Pádua | 603.63 | 47 | 3 | 7.8 | Poor | 0.5 | Poor |
| São Fidélis | 1 034.83 | 308 | 16 | 29.8 | Average | 1.5 | Poor |
| São Francisco de Itabapoana | 1 118.04 | 212 | 18 | 19.0 | Poor | 1.6 | Poor |
| São Gonçalo | 248.16 | 2 | 1 | 0.8 | Poor | 0.4 | Poor |
| São João da Barra | 452.40 | 228 | 13 | 50.4 | Good | 2.9 | Average |
| São João de Meriti | 35.22 | 0 | 0 | 0.0 | Poor | 0.0 | Poor |
| São José de Ubá | 249.69 | 29 | 3 | 11.6 | Poor | 1.2 | Poor |
| São José do Vale do Rio Preto | 220.18 | 12 | 5 | 5.5 | Poor | 2.3 | Poor |
| São Pedro da Aldeia | 332.49 | 22 | 6 | 6.6 | Poor | 1.8 | Poor |
| São Sebastião do Alto | 397.21 | 108 | 9 | 27.2 | Average | 2.3 | Poor |
| Sapucaia | 540.67 | 98 | 14 | 18.1 | Poor | 2.6 | Average |
| Saquarema | 352.13 | 207 | 34 | 58.8 | Good | 9.7 | Good |
| Seropédica | 265.19 | 129 | 27 | 48.6 | Good | 10.2 | Good |
| Silva Jardim | 937.76 | 681 | 51 | 72.6 | Good | 5.4 | Good |
| Sumidouro | 413.41 | 18 | 3 | 4.4 | Poor | 0.7 | Poor |
| Tanguá | 143.01 | 8 | 2 | 5.6 | Poor | 1.4 | Poor |
| Teresópolis | 773.34 | 229 | 42 | 29.6 | Average | 5.4 | Good |
| Trajano de Moraes | 591.15 | 28 | 5 | 4.7 | Poor | 0.8 | Poor |
| Três Rios | 322.84 | 170 | 16 | 52.7 | Good | 5.0 | Good |
| Valença | 1 300.77 | 6 | 4 | 0.5 | Poor | 0.3 | Poor |
| Varre-Sai | 201.94 | 0 | 0 | 0.0 | Poor | 0.0 | Poor |
| Vassouras | 536.07 | 29 | 3 | 5.4 | Poor | 0.6 | Poor |
| Volta Redonda | 182.11 | 14 | 4 | 7.7 | Poor | 2.2 | Poor |
| Rio de Janeiro State | 43 750 | 13 327 | 1 623 | 30.5 | 3.7 |
Table A2.
Abundance of freshwater native fish species per hydrographic region.
| RH-1 | RH-2 | RH-3 | RH-4 | RH-5 | RH-6 | RH-7 | RH-8 | RH-9 | |
|---|---|---|---|---|---|---|---|---|---|
| Acentronichthys leptos | 109 | 20 | 0 | 0 | 171 | 17 | 36 | 5 | 7 |
| Ancistrus multispinis | 145 | 119 | 0 | 0 | 501 | 31 | 0 | 5 | 0 |
| Astyanax keronolepis | 1091 | 97 | 0 | 0 | 551 | 0 | 0 | 0 | 0 |
| Astyanax lacustris | 2 | 511 | 81 | 72 | 115 | 184 | 173 | 1950 | 1582 |
| Atlantirivulus janeiroensis | 0 | 0 | 0 | 0 | 0 | 11 | 0 | 0 | 0 |
| Atlantirivulus jurubatibensis | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 51 |
| Atlantirivulus guanabarensis | 0 | 0 | 0 | 0 | 73 | 0 | 0 | 0 | 0 |
| Atlantirivulus lazzarotoi | 17 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Atlantirivulus maricensis | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 |
| Atlantirivulus simplicis | 43 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Australoheros ipatinguensis | 0 | 6 | 112 | 99 | 0 | 23 | 32 | 0 | 257 |
| Australoheros oblongus | 0 | 18 | 4 | 0 | 89 | 49 | 0 | 126 | 47 |
| Awaous tajasica | 115 | 5 | 0 | 3 | 81 | 17 | 35 | 113 | 23 |
| Brachyhypopomus janeiroensis | 0 | 0 | 0 | 1 | 1 | 82 | 12 | 11 | 49 |
| Brycon insignis | 0 | 0 | 1 | 0 | 0 | 10 | 0 | 7 | 26 |
| Brycon opalinus | 0 | 30 | 20 | 8 | 0 | 0 | 14 | 0 | 1 |
| Bryconamericus microcephalus | 919 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Bryconamericus ornaticeps | 0 | 287 | 0 | 0 | 1900 | 0 | 0 | 0 | 0 |
| Bryconamericus tenuis | 0 | 0 | 0 | 20 | 0 | 128 | 88 | 287 | 40 |
| Callichthys callichthys | 0 | 47 | 12 | 3 | 198 | 19 | 11 | 56 | 165 |
| Characidium alipioi | 0 | 0 | 0 | 21 | 0 | 119 | 45 | 18 | 58 |
| Characidium grajahuense | 0 | 140 | 0 | 0 | 321 | 0 | 0 | 0 | 0 |
| Characidium interruptum | 0 | 14 | 0 | 0 | 249 | 236 | 0 | 38 | 152 |
| Characidium japuhybense | 869 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Characidium lauroi | 0 | 660 | 135 | 411 | 0 | 0 | 30 | 0 | 0 |
| Characidium litorale | 0 | 0 | 0 | 0 | 0 | 588 | 0 | 172 | 33 |
| Characidium vidali | 0 | 146 | 0 | 124 | 931 | 0 | 10 | 342 | 0 |
| Corydoras nattereri | 0 | 63 | 4 | 97 | 314 | 86 | 2 | 7 | 87 |
| Crenicichla lacustris | 0 | 19 | 36 | 7 | 0 | 43 | 60 | 71 | 597 |
| Crenicichla lepidota | 0 | 47 | 0 | 0 | 83 | 2 | 0 | 0 | 0 |
| Cyphocharax gilbert | 0 | 467 | 1 | 6 | 43 | 344 | 13 | 740 | 381 |
| Delturus parahybae | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 |
| Deuterodon giton | 0 | 141 | 154 | 312 | 0 | 2217 | 243 | 677 | 1954 |
| Deuterodon hastatus | 193 | 269 | 0 | 32 | 2620 | 0 | 0 | 0 | 0 |
| Deuterodon heterostomus | 0 | 4 | 42 | 0 | 2 | 50 | 0 | 1 | 1 |
| Deuterodon intermedius | 179 | 2574 | 463 | 187 | 0 | 0 | 151 | 0 | 35 |
| Deuterodon janeiroensis | 0 | 5745 | 0 | 7 | 1384 | 512 | 17 | 0 | 0 |
| Deuterodon luetkenii | 0 | 0 | 0 | 0 | 35 | 98 | 6 | 6284 | 1450 |
| Deuterodon taeniatus | 0 | 26 | 9 | 70 | 6 | 982 | 346 | 3072 | 865 |
| Dormitator maculatus | 11 | 13 | 0 | 0 | 8 | 1 | 0 | 3 | 80 |
| Eigenmannia virescens | 0 | 0 | 7 | 17 | 7 | 42 | 125 | 14 | 17 |
| Eleotris pisonis | 31 | 9 | 0 | 0 | 28 | 26 | 5 | 26 | 23 |
| Geophagus brasiliensis | 4092 | 983 | 241 | 346 | 1809 | 200 | 364 | 875 | 1114 |
| Glanidium melanopterum | 0 | 14 | 26 | 9 | 0 | 2 | 25 | 3 | 11 |
| Gymnotus carapo | 0 | 90 | 37 | 52 | 51 | 44 | 11 | 13 | 23 |
| Gymnotus pantherinus | 38 | 10 | 0 | 0 | 61 | 47 | 5 | 41 | 5 |
| Harttia carvalhoi | 0 | 88 | 11 | 24 | 0 | 0 | 0 | 0 | 23 |
| Harttia loricariformis | 0 | 32 | 13 | 52 | 0 | 0 | 55 | 0 | 7 |
| Hemipsilichthys gobio | 0 | 9 | 0 | 10 | 1 | 0 | 8 | 0 | 0 |
| Hemipsilichthys nimius | 145 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Hemipsilichthys papillatus | 0 | 7 | 1 | 0 | 0 | 0 | 0 | 0 | 0 |
| Hisonotus notatus | 0 | 205 | 4 | 3 | 689 | 24 | 31 | 0 | 113 |
| Hisonotus thayeri | 0 | 0 | 0 | 0 | 0 | 557 | 92 | 92 | 107 |
| Hollandichthys multifasciatus | 334 | 11 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Homodiaetus banguela | 0 | 0 | 0 | 0 | 0 | 9 | 0 | 0 | 0 |
| Homodiaetus passarellii | 0 | 2 | 0 | 0 | 48 | 0 | 0 | 0 | 0 |
| Hoplerythrinus unitaeniatus | 0 | 2 | 1 | 0 | 17 | 10 | 0 | 10 | 121 |
| Hoplias malabaricus | 14 | 45 | 13 | 20 | 104 | 83 | 6 | 142 | 163 |
| Hoplosternum littorale | 0 | 96 | 23 | 8 | 38 | 1 | 4 | 2 | 31 |
| Hyphessobrycon bifasciatus | 0 | 154 | 8 | 58 | 1070 | 192 | 11 | 5063 | 2894 |
| Hyphessobrycon boulengeri | 0 | 33 | 0 | 0 | 245 | 15 | 0 | 311 | 496 |
| Hyphessobrycon flammeus | 0 | 48 | 0 | 1 | 35 | 12 | 0 | 0 | 4 |
| Hypomasticus copelandii | 0 | 10 | 2 | 13 | 13 | 13 | 19 | 19 | 132 |
| Hypomasticus mormyrops | 0 | 10 | 14 | 7 | 0 | 0 | 17 | 1 | 3 |
| Hypomasticus thayeri | 0 | 0 | 0 | 3 | 0 | 0 | 0 | 0 | 21 |
| Hypostomus affinis | 0 | 0 | 3 | 0 | 5 | 98 | 1 | 65 | 0 |
| Hypostomus auroguttatus | 0 | 0 | 2 | 2 | 0 | 0 | 2 | 0 | 0 |
| Hypostomus luetkeni | 0 | 34 | 20 | 155 | 0 | 0 | 168 | 0 | 87 |
| Hypostomus punctatus | 0 | 95 | 0 | 0 | 244 | 2 | 0 | 0 | 0 |
| Hypostomus vermicularis | 0 | 0 | 52 | 4 | 0 | 0 | 2 | 0 | 4 |
| Imparfinis minutus | 0 | 61 | 59 | 4 | 0 | 0 | 6 | 0 | 7 |
| Imparfinis piperatus | 0 | 4 | 4 | 0 | 4 | 0 | 0 | 0 | 0 |
| Ituglanis parahybae | 0 | 0 | 0 | 0 | 0 | 10 | 0 | 0 | 3 |
| Jenynsia darwini | 0 | 0 | 0 | 0 | 7 | 8 | 0 | 160 | 1516 |
| Jenynsia lineata | 0 | 0 | 0 | 0 | 318 | 65 | 0 | 0 | 0 |
| Knodus moenkhausii | 0 | 0 | 0 | 267 | 0 | 0 | 10 | 0 | 29 |
| Kronichthys heylandi | 664 | 167 | 0 | 1 | 151 | 4 | 0 | 0 | 0 |
| Kryptolebias brasiliensis | 0 | 54 | 0 | 0 | 164 | 16 | 0 | 0 | 0 |
| Kryptolebias caudomarginatus | 0 | 166 | 0 | 0 | 56 | 0 | 0 | 0 | 0 |
| Kryptolebias gracilis | 0 | 0 | 0 | 0 | 0 | 28 | 0 | 11 | 0 |
| Kryptolebias ocellatus | 0 | 86 | 1 | 0 | 0 | 2 | 0 | 0 | 0 |
| Leptolebias marmoratus | 0 | 0 | 0 | 0 | 10 | 0 | 0 | 0 | 0 |
| Leptopanchax citrinipinnis | 184 | 0 | 0 | 0 | 0 | ||||
| Leptopanchax opalescens | 0 | 7 | 0 | 0 | 55 | 0 | 0 | 0 | 0 |
| Leptopanchax sanguineus | 0 | 0 | 0 | 0 | 15 | 0 | 0 | 0 | 0 |
| Leptopanchax splendens | 0 | 0 | 0 | 0 | 50 | 0 | 0 | 0 | 0 |
| Listrura costai | 15 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Listrura nematopteryx | 0 | 0 | 0 | 0 | 156 | 0 | 0 | 0 | 0 |
| Listrura tetraradiata | 0 | 0 | 0 | 0 | 0 | 100 | 0 | 0 | 1 |
| Loricariichthys castaneus | 0 | 89 | 15 | 1 | 19 | 317 | 19 | 1 | 122 |
| Megaleporinus conirostris | 0 | 3 | 0 | 4 | 0 | 0 | 23 | 0 | 37 |
| Microcambeva barbata | 0 | 0 | 0 | 0 | 0 | 71 | 0 | 4 | 0 |
| Microcambeva bendego | 0 | 0 | 0 | 0 | 7 | 0 | 0 | 0 | 0 |
| Microglanis nigripinnis | 0 | 0 | 0 | 0 | 21 | 0 | 0 | 0 | 0 |
| Microglanis parahybae | 0 | 31 | 0 | 0 | 0 | 0 | 44 | 0 | 61 |
| Microglanis pleriqueater | 0 | 0 | 0 | 0 | 0 | 55 | 0 | 10 | 0 |
| Mimagoniates microlepis | 808 | 255 | 0 | 637 | 1328 | 501 | 0 | 30 | 33 |
| Nematolebias papilliferus | 0 | 0 | 0 | 0 | 28 | 0 | 0 | 0 | 0 |
| Nematolebias whitei | 0 | 0 | 0 | 0 | 17 | 291 | 0 | 9 | 0 |
| Neoplecostomus microps | 0 | 825 | 225 | 135 | 82 | 0 | 118 | 219 | 1 |
| Neoplecostomus paraty | 163 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Neoplecostomus variipictus | 0 | 0 | 0 | 55 | 0 | 0 | 80 | 0 | 0 |
| Notholebias cruzi | 0 | 0 | 0 | 0 | 29 | 32 | 0 | 0 | 0 |
| Notholebias fractifasciatus | 0 | 0 | 0 | 0 | 20 | 2 | 0 | 0 | 0 |
| Notholebias minimus | 0 | 181 | 0 | 0 | 8 | 0 | 0 | 0 | 0 |
| Oligosarcus hepsetus | 10 | 346 | 94 | 80 | 64 | 24 | 92 | 154 | 249 |
| Ophthalmolebias constanciae | 0 | 0 | 0 | 0 | 0 | 47 | 0 | 65 | 0 |
| Otocinclus affinis | 0 | 6 | 1 | 14 | 26 | 3 | 1 | 1 | 13 |
| Otothyris lophophanes | 2 | 24 | 4 | 3 | 45 | 135 | 3 | 69 | 41 |
| Pachyurus adspersus | 0 | 3 | 9 | 1 | 0 | 0 | 7 | 0 | 29 |
| Paragenidens grandoculis | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 |
| Pareiorhaphis garbei | 0 | 0 | 0 | 0 | 232 | 13 | 22 | 54 | 0 |
| Pareiorhina brachyrhyncha | 0 | 0 | 160 | 0 | 0 | 0 | 0 | 0 | 0 |
| Pareiorhina rudolphi | 77 | 553 | 672 | 0 | 0 | 0 | 0 | 0 | 0 |
| Parotocinclus bidentatus | 0 | 0 | 22 | 1 | 0 | 0 | 0 | 0 | 0 |
| Parotocinclus fluminense | 0 | 0 | 0 | 0 | 0 | 515 | 0 | 0 | 0 |
| Parotocinclus maculicauda | 0 | 390 | 0 | 0 | 940 | 36 | 4 | 179 | 0 |
| Parotocinclus muriaensis | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 12 |
| Phalloceros anisophallos | 3391 | 1144 | 0 | 0 | 101 | 0 | 0 | 0 | 0 |
| Phalloceros enneaktinos | 701 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Phalloceros harpagos | 468 | 3200 | 276 | 1295 | 773 | 887 | 247 | 3306 | 1148 |
| Phalloceros leptokeras | 946 | 57 | 121 | 2216 | 1419 | 56 | 0 | 0 | 0 |
| Phalloceros tupinamba | 0 | 200 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Phalloptychus januarius | 0 | 33 | 0 | 0 | 1076 | 133 | 0 | 0 | 11100 |
| Phallotorynus fasciolatus | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 47 |
| Piabina argentea | 0 | 0 | 0 | 0 | 0 | 0 | 10 | 0 | 0 |
| Pimelodella lateristriga | 101 | 58 | 2 | 25 | 224 | 201 | 81 | 167 | 236 |
| Pimelodus maculatus | 0 | 11 | 57 | 11 | 0 | 0 | 8 | 0 | 13 |
| Poecilia vivipara | 120 | 378 | 3 | 58 | 2278 | 298 | 130 | 1660 | 10094 |
| Pogonopoma parahybae | 0 | 0 | 3 | 1 | 0 | 0 | 18 | 0 | 0 |
| Prochilodus lineatus | 0 | 2 | 4 | 0 | 0 | 0 | 13 | 6 | 233 |
| Prochilodus vimboides | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 11 | 170 |
| Psalidodon parahybae | 0 | 138 | 179 | 105 | 82 | 192 | 401 | 9 | 1395 |
| Pseudotothyris janeirensis | 0 | 147 | 0 | 0 | 29 | 9 | 0 | 29 | 18 |
| Rhamdia quelen | 43 | 138 | 24 | 29 | 188 | 26 | 24 | 53 | 55 |
| Rhamdioglanis frenatus | 55 | 72 | 0 | 0 | 48 | 0 | 3 | 0 | 0 |
| Rhamdioglanis transfasciatus | 0 | 1 | 0 | 0 | 63 | 108 | 1 | 131 | 0 |
| Rineloricaria nigricauda | 0 | 553 | 309 | 381 | 0 | 0 | 4 | 0 | 4 |
| Rineloricaria nudipectoris | 0 | 22 | 0 | 263 | 1249 | 62 | 0 | 145 | 0 |
| Rineloricaria steindachneri | 0 | 3 | 0 | 0 | 0 | 0 | 107 | 0 | 23 |
| Rineloricaria zawadzki | 40 | 227 | 14 | 0 | 95 | 0 | 0 | 0 | 0 |
| Schizolecis guentheri | 2592 | 622 | 0 | 20 | 1394 | 123 | 42 | 513 | 0 |
| Scleromystax barbatus | 431 | 147 | 0 | 134 | 732 | 144 | 37 | 51 | 7 |
| Scleromystax prionotos | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 11 | 7 |
| Spintherobolus broccae | 0 | 10 | 0 | 0 | 158 | 39 | 0 | 17 | 0 |
| Steindachneridion parahybae | 0 | 0 | 11 | 4 | 0 | 0 | 1 | 0 | 0 |
| Synbranchus marmoratus | 3 | 7 | 11 | 2 | 56 | 11 | 4 | 10 | 7 |
| Taunayia bifasciata | 2 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 |
| Trachelyopterus striatulus | 0 | 0 | 0 | 0 | 5 | 0 | 7 | 0 | 0 |
| Trichogenes longipinnis | 627 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Trachelyopterus striatulus | 0 | 31 | 1 | 4 | 0 | 23 | 0 | 71 | 78 |
| Trichomycterus albinotatus | 0 | 0 | 94 | 0 | 0 | 0 | 0 | 0 | 0 |
| Trichomycterus auroguttatus | 0 | 11 | 142 | 0 | 0 | 0 | 0 | 0 | 0 |
| Trichomycterus caipora | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 110 | 97 |
| Trichomycterus claudiae | 0 | 83 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Trichomycterus florensis | 0 | 0 | 16 | 16 | 0 | 0 | 0 | 0 | 0 |
| Trichomycterus fuliginosus | 0 | 0 | 0 | 0 | 0 | 0 | 58 | 0 | 0 |
| Trichomycterus goeldii | 0 | 0 | 0 | 179 | 0 | 0 | 27 | 0 | 0 |
| Trichomycterus giganteus | 0 | 201 | 0 | 0 | 153 | 0 | 0 | 0 | 0 |
| Trichomycterus itatiayae | 0 | 0 | 197 | 0 | 0 | 0 | 0 | 0 | 0 |
| Trichomycterus jacupiranga | 452 | 30 | 0 | 0 | 77 | 0 | 0 | 0 | 0 |
| Trichomycterus largoperculatus | 0 | 0 | 0 | 33 | 0 | 0 | 28 | 0 | 1 |
| Trichomycterus macrophthalmus | 0 | 142 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Trichomycterus mariamole | 0 | 13 | 82 | 0 | 0 | 0 | 0 | 0 | 0 |
| Trichomycterus mirissumba | 0 | 0 | 90 | 0 | 0 | 0 | 0 | 0 | 0 |
| Trichomycterus nigricans | 0 | 0 | 0 | 0 | 972 | 16 | 0 | 0 | 0 |
| Trichomycterus nigroauratus | 0 | 215 | 24 | 0 | 0 | 0 | 0 | 0 | 0 |
| Trichomycterus paquequerense | 0 | 0 | 0 | 1012 | 80 | 0 | 0 | 0 | 0 |
| Trichomycterus potschi | 0 | 134 | 102 | 0 | 0 | 0 | 0 | 0 | 0 |
| Trichomycterus puriventris | 0 | 0 | 0 | 0 | 0 | 0 | 228 | 0 | 3 |
| Trichomycterus vitalbrazili | 0 | 0 | 0 | 0 | 0 | 0 | 60 | 0 | 0 |
References
- Thomaz, A.T.; Malabarba, L.R.; Bonatto, S.L.; Knowles, L.L. Testing the effect of palaeodrainages versus habitat stability on genetic divergence in riverine systems: Study of a Neotropical fish of the Brazilian coastal Atlantic Forest. J Biogeogr 2015, 42, 2389–2401. [Google Scholar] [CrossRef]
- Dagosta, F.C.P.; de Pinna, M.C.C. A history of the biogeography of Amazonian fishes. Neotrop Ichthyol 2018, 16, e180023. [Google Scholar] [CrossRef]
- Sarmento-Soares, L.M.; Martins-Pinheiro, R.F. A fauna de peixes nas bacias do sul do Espírito Santo, Brasil. Sitientibus. Série Ciências Biológicas 2013, 13, 1–37. [CrossRef]
- Camelier, P.; Zanata, A.M. Biogeography of freshwater fishes from the Northeastern Mata Atlântica freshwater ecoregion: Distribution, endemism, and area relationships. Neotrop Ichthyol 2014, 12, 683–698. [CrossRef]
- Vieira-Guimarães, F.; Sarmento-Soares, L.M.; Nobre, D.M.; Neiva, D.C.; Silva, J.P.; Martins-Pinheiro, R.F. Biogeographic patterns of the freshwater fishes from the state of Espírito Santo, eastern Brazil. Stud Neotrop Fauna Environ 2023, 2023, 1–20. [CrossRef]
- Bizerril, C.R.S.F.; Primo, P.B.S. Peixes de Águas Interiores do Estado do Rio de Janeiro. FEMAR—SEMADS: Rio de Janeiro, Brazil, 2001; 417 p.
- Gomes, J.R. Levantamento da ictiofauna do Maciço da Pedra Branca e arredores, Rio de Janeiro, estado do Rio de Janeiro. Arq Mus Nac Rio J 2006, 64, 309–320.
- Mazzoni, R.; Verani-Fenerich, N.; Caramaschi, E.P.; Iglesias-Rios, R. Stream-Dwelling Fish Communities from an Atlantic Rain Forest Drainage. Braz Arch Biol Technol 2006, 49, 249–256. [CrossRef]
- Di Dario, F.; Petry, A.C.; Pereira, M.M.S.; Mincarone, M.M.; Agostinho, L.S.; Camara, E.M.; Caramaschi, E.P.; Britto, M.R. An update on the fish composition (Teleostei) of the coastal lagoons of the Restinga de Jurubatiba National Park and the Imboassica Lagoon, northern Rio de Janeiro State. Acta Limnol Bras 2013, 25, 257–278.
- Buckup, P.A.; Britto, M.R.; Souza-Lima, R.; Pascoli, J.C.; Villa-Verde, L.; Ferraro, G.A.; Salgado, F.L.K.; Gomes, J.R. Guia de identificação das espécies de peixes da bacia do Rio das Pedras. The Nature Conservancy: Rio de Janeiro, Brazil, 2014; 79 p.
- Camilo, G.S.; Terra, B.; Araújo, F.G. Ichthyofauna from the Parque Nacional da Serra dos Órgãos and its surrounding areas, Rio de Janeiro state, Brazil. Check List 2015, 11, 1696. [CrossRef]
- De Brito, V.; Buckup, P.A. The fish fauna of the upper Piraí drainage, a transposed mountain river system in southeastern, Brazil. Check List 2019, 15, 235–247. [CrossRef]
- Guimarães, F.V.; Souza, T.M.; Rodrigues, R.R.; Souza-Lima, R. Composition and distribution of fishes from the Perequê-Açu river basin, Paraty, Rio de Janeiro, Southeastern Brazil. Biota Neotrop 2021, 21, e20201096. [CrossRef]
- Dopazo, M.; Souto-Santos, I.C.A.; Britto, M.R.; Moreira, C.R.; Buckup, P.A. The freshwater fishes from the Costa Verde Fluminense region of southeastern Brazil. Biota Neotrop 2023, 23, e20221422. [CrossRef]
- Pfafstetter, O. Classificação de bacias hidrográficas – Metodologia de classificação. Departamento Nacional de Obras de Saneamento (DNOS): Rio de Janeiro, Brazil, 1989. Unpublished manuscript.
- Sarmento-Soares, L.M.; Martins-Pinheiro, R.F.; Rodrigues, L.N. Peixes do rio Doce segundo as coleções. Bol Soc Brasil Ictiologia 2017, 123, 9-25.
- Agência Nacional de Águas (ANA). Topologia hídrica: Método de construção e modelagem da base hidrográfica para suporte à gestão de recursos hídricos: V. 1.11. Agência Nacional de Águas, Superintendência de Gestão da Informação: Brasília, Brazil, 2006.
- Conselho Estadual de Recursos Hídricos do Rio de Janeiro (CERHI-RJ). Resolução nº 107 de 22 de maio de 2013, Conselho Estadual de Recursos Hídricos: Rio de Janeiro, Brazil, 2013.
- Sabaj-Pérez, M.H. Codes for natural history collections in ichthyology and herpetology. Ichthyol Herpetol 2020, 108, 1–76. [CrossRef]
- ricke, R.; Eschmeyer, W.N.; Van Der Laan, R. Eschmeyer’s Catalog of Fishes: Genera, Species, References. Available online: http://researcharchive.calacademy.org/research/ichthyology/catalog/fishcatmain.asp (Accessed 15 May 2024). Genera, Species.
- Ferreira Júnior, O. GPS Software for mapping and innovative solutions for asset tracking: GPS Trackmaker Pro v. 5.1. Geo Studio Tecnologia Ltda: Belo Horizonte, Brazil, 2012.
- Instituto Brasileiro de Geografia e Estatística (IBGE). Base cartográfica vetorial contínua do estado do Rio de Janeiro na escala 1:25.000 - BC25_RJ. Available online: https://www.ibge.gov.br/geociencias/downloads-geociencias.html?caminho=cartas_e_mapas/bases_cartograficas_continuas/bc250/versao2023/ (Accessed on 01 June 2024).
- Instituto Brasileiro de Geografia e Estatística (IBGE). Área territorial - Brasil, Grandes Regiões, Unidades da Federação e Municípios. Available online: https://geoftp.ibge.gov.br/organizacao_do_territorio/estrutura_territorial/areas_territoriais/2022/AR_BR_RG_UF_RGINT_MES_MIC_MUN_2022.xls (Accessed on 01 June 2024).
- Silva,T.N.; Sarmento-Soares, L.M.; Martins-Pinheiro, R.F.; Santos, A.C.A. (2021): Composition and distribution of the fish fauna in the Rio Jacuípe, northernmost tributary of the Rio Paraguaçu basin, Bahia, Brazil, Studies on Neotropical Fauna and Environment, 2021. [CrossRef]
- Dajoz, R. Ecologia Geral. 4th ed.; Vozes: Petrópolis, Brazil, 1983; 472 p.
- Harper, D.A.T. Numerical palaeobiology: Computer- based modelling and analysis of fossils and their distributions. John Wiley & Sons: Chichester, USA, 1999; 468 p.
- Tóthmérész, B. Comparison of different methods for diversity ordering. J Veg Sci 1995, 6, 283–290. [CrossRef]
- Hammer, O; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol Eletron 2001, 4, 1–9.
- Walther, B.A.; Moore, J.L. The concepts of bias, precision and accuracy, and their use in testing the performance of species richness estimators, with a literature review of estimator performance. Ecography 2005, 28, 815–829. [CrossRef]
- Chao, A. Nonparametric estimation of the number of classes in a population. Scand J Stat 1984, 11, 265–270.
- Chao, A. Estimating the population size for capture-recapture data with unequal catchability. Biometrics 1987, 43, 783–791. [CrossRef]
- Chiu, C.H.; Wang, Y.T.; Walther, B.A.; Chao, A. An improved nonparametric lower bound of species richness via a modified Good-Turing frequency formula. Biometrics 2014, 70, 671–682. [CrossRef]
- Chao, A.; Lee, S-M. Estimating the number of classes via sample coverage. J Am Stat Assoc 1992, 87, 210–217. [CrossRef]
- Chao, A.; Hwang, W-H; Chen, Y-C.; Kuo C-Y. Estimating the number of shared species in two communities. Stat Sinica 2000, 10, 227–246.
- Alroy, J. Limits to species richness in terrestrial communities. Ecol Lett 2018, 21, 1781–1789. [CrossRef]
- Edler, D.; Guedes, T.; Zizka, A.; Rosvall, M.; Antonelli, A. Infomap bioregions: Interactive mapping of biogeographical regions from species distributions. Syst Biol 2017, 66, 197–204. [CrossRef]
- Oliveira, U; Soares-Filho, B., Leitão, R.F.M.; Rodrigues, H.O. BioDinamica: A toolkit for analyses of biodiversity and biogeography on the Dinamica-EGO modelling platform. PeerJ 2019, 7, e7213. [CrossRef]
- Ferreira, B.M.; Soares-Filho, B S.; Pereira, F.M.Q. The Dinamica EGO virtual machine. Sci Comput Program 2019, 173, 3–20. [CrossRef]
- Bizerril, C.R.S.F. A Ictiofauna da Bacia do Rio Paraíba do Sul. Biodiversidade e Padrões Biogeográficos. Braz Arch Biol Technol 1999, 42, 1–17. [CrossRef]
- Menezes, N.A.; Weitzman, S.H.; Oyakawa, O.T.; Lima, F.C.; Castro, R.M.C.; Weitzman, M.J. Peixes de água doce da Mata Atlântica. Neotrópica: São Paulo, Brazil, 2007; 407 p.
- Abell, R.; Thieme, M.L.; Revenga, C.; Bryer, M.; Kottelat, M.; Bogutskaya, N.; Coad, B.; Mandrak, N.; Balderas, S.C.; Bussing, W.; Stiassny, M.L.J.; Skelton, P.; Allen, G.R.; Unmack, P.; Naseka, A.; Ng, R.; Sindorf, N.; Robertson, J.; Armijo, E.; Higgins, J.V.; Heibel, T.J.; Wikramanayake, E.; Olson, D.; López, H.L.; Reis, R.E.; Lundberg, J.G.; Pérez, M.H.S.; Petry, P. Freshwater ecoregions of the world: A new map of biogeographic units for freshwater biodiversity conservation. BioScience 2008, 58, 403–414. [CrossRef]
- Zalán, P.V.; Oliveira, J.A.B. Origem e evolução estrutural do Sistema de Riftes Cenozóicos do Sudeste do Brasil. Bol Geoci Petrobras 2005, 13, 269–300.
- Lamego, A.R. O homem e a Guanabara, 2nd ed. IBGE- Conselho Nacional de Geografia: Rio de Janeiro, Brazil, 1964; 408 p.
- Silva, C.G.; Reis, A.T.; Goiana, L.T.C.; Ferrari, A.L. A história do preenchimento sedimentar da baía de Guanabara através da geofísica. In Baía de Guanabara: Um ambiente em transformação; Fonseca, E.M.; Neto, J.A.B.; Pompermayer, F.C.L., Eds.; Ape’Ku: Rio de Janeiro, Brazil, 2021; pp. 47–72.
- Buckup, P.A. The Eastern Brazilian Shield. In Historical Biogeography of Neotropical Freshwater Fishes; Albert, J.S.; Reis, R.E., Eds.; University of California Press, Los Angeles, USA, 2011; pp. 203–210. [CrossRef]
- Pio, N.L.; Carvalho, T.P. Evidence on the paleodrainage connectivity during Pleistocene: Phylogeography of a hypoptopomatine endemic to southeastern Brazilian coastal drainages. Neotrop Ichthyol 2021, 19, e200128. [CrossRef]
- Lima, S.M.Q.; Vasconcellos, A.V.; Berbel-Filho, W.M.; Lazoski, C.; Russo, C.A.; Sazima, I.; Solé-Cava, A.M. Effects of Pleistocene climatic and geomorphological changes on the population structure of the restricted-range catfish Trichogenes longipinnis (Siluriformes: Trichomycteridae). Syst Biodivers 2017, 14, 155–170. [CrossRef]
- Lowe-McConnell, R.H. Estudos ecológicos de comunidades de peixes tropicais. EDUSP: São Paulo, Brazil, 1999; 536 p.
- Souza-Lima, R.; Miranda, J.C.; Portugal, A.S. Ictiofauna do Rio Aldeia, São Gonçalo. In Estudos Ambientais em Regiões Metropolitanas: São Gonçalo; Santos, M.G., Ed; Ed. UERJ: Rio de Janeiro, Brazil, 2012; pp. 115–134.
- Angrizani, R.C.; Malabarba, L.R. Genetic diversity and species delimitation in Rhamdia (Siluriformes: Heptapteridae) in South America, with a redescription of R. quelen (Quoy & Gaimard, 1824). Zootaxa 2020, 4801, 85–104. [CrossRef]
- Craig, J.M; Crampton, W.G.R.; Albert, J.S. Revision of the polytypic electric fish Gymnotus carapo (Gymnotiformes, Teleostei), with descriptions of seven subspecies. Zootaxa 2017, 4318, 401–438. [CrossRef]
- Dergam, J.A., Suzuki, H.I., Shibatta, O.A., Duboc, L.F., Júlio, H.F., Jr., Giuliano-Caetano, L.; Black, W.C., IV. Molecular biogeography of the neotropical fish Hoplias malabaricus (Erythrinidae: Characiformes) in the Iguaçu, Tibagi and Paraná Rivers. Genet Mol Biol 1998, 21, 493–496. [CrossRef]
- Torres, R.A.; Roper, J.J.; Foresti, F.; Oliveira, C. Surprising genomic diversity in the Neotropical fish Synbranchus marmoratus (Teleostei: Synbranchidae): How many species? Neotrop Ichthyol 2005, 3, 277–284. [CrossRef]
- Bertaco, V.A.; Ferrer, J.; Carvalho, F.R.; Malabarba, L.R. Inventory of the freshwater fishes from a densely collected area in South America – a case study of the current knowledge of Neotropical fish diversity. Zootaxa 2016, 4138, 401–440. [CrossRef]
- Guimarães, K.L.A.; Rosso, J.J.; González-Castro, M.; Souza, M.F.B.; Díaz de Astarloa, J.M.; Rodrigues, L.R.R. A new species of Hoplias malabaricus species complex (Characiformes: Erythrinidae) from the Crepori River, Amazon basin, Brazil. J Fish Biol 2022, 100, 425–443. [CrossRef]
Figure 1.
The nine hydrographic regions along Rio de Janeiro. RH-1- Ilha Grande Bay drainage; RH-2- Rio Guandu and transposition with Paraíba do Sul (08.22e); RH-3- Middle stretch of Rio Paraiba do Sul basin (08.22d); RH-4- Rio Piabanha and tributaries at Paraíba do Sul basin (08.22c); RH-5- Guanabara bay drainage; RH-6- Lake region and Rio São João; RH-7- Rio Dois Rios and tributaries at Paraíba do Sul basin (08.22b); RH-8- Rio Macaé and Rio das Ostras; RH-9- Lower Rio Paraíba do Sul and Itabapoana (08.22a).
Figure 1.
The nine hydrographic regions along Rio de Janeiro. RH-1- Ilha Grande Bay drainage; RH-2- Rio Guandu and transposition with Paraíba do Sul (08.22e); RH-3- Middle stretch of Rio Paraiba do Sul basin (08.22d); RH-4- Rio Piabanha and tributaries at Paraíba do Sul basin (08.22c); RH-5- Guanabara bay drainage; RH-6- Lake region and Rio São João; RH-7- Rio Dois Rios and tributaries at Paraíba do Sul basin (08.22b); RH-8- Rio Macaé and Rio das Ostras; RH-9- Lower Rio Paraíba do Sul and Itabapoana (08.22a).
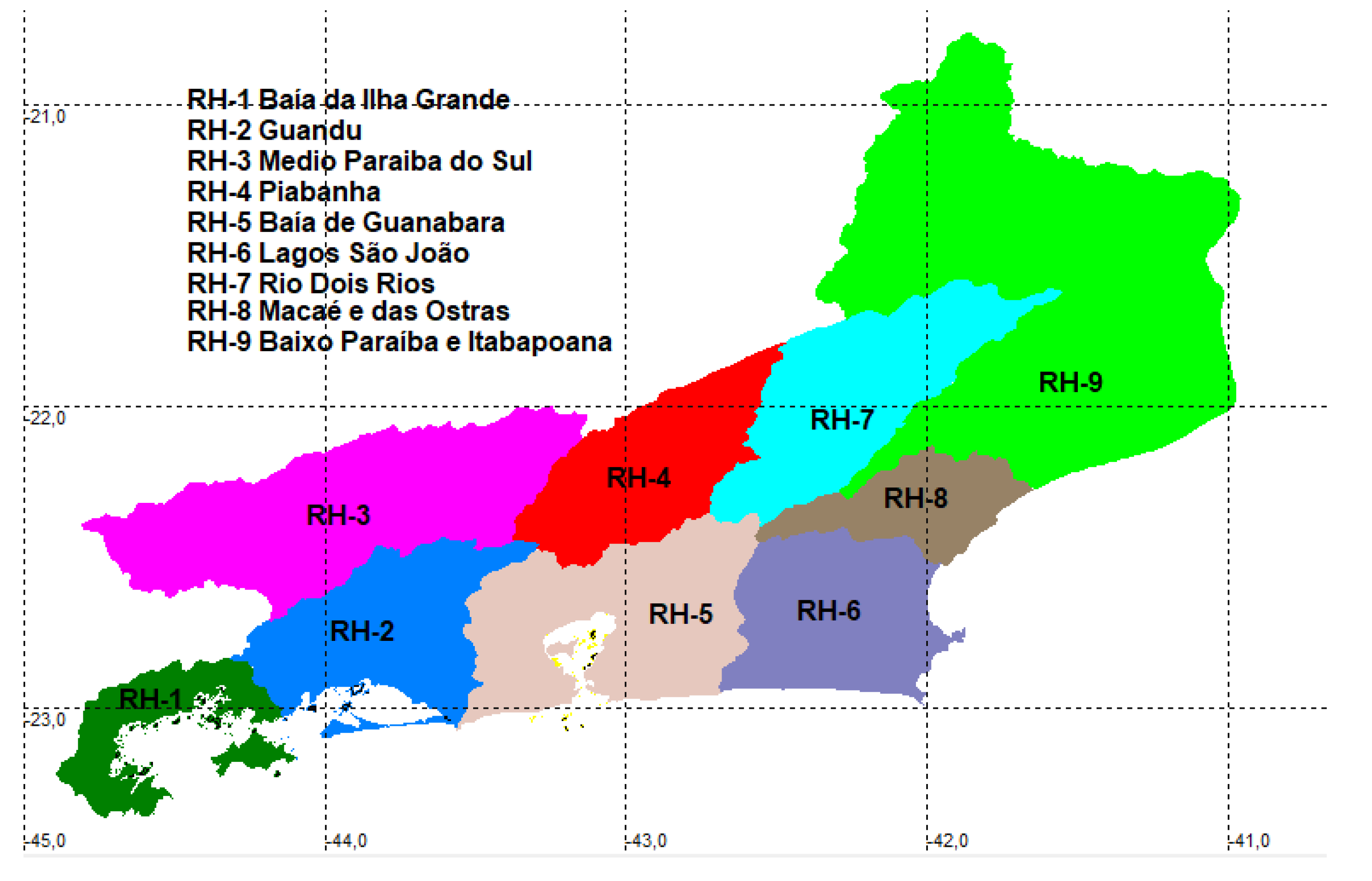
Figure 2.
The twenty-nine river basin divisions along Rio de Janeiro. Coastal drainages: Itabapoana Basin (08.20- Figure 2- A), São Francisco de Itabapoana Watersheds (08.21- Figure 2- B)- , Lagoa Feia Watersheds (08.23- Figure 2- H), Jurubatiba Watersheds (08.24- Figure 2- I), Macaé River Basin (08.25- Figure 2- J), Rio das Ostras Watersheds (08.26- Figure 2- K), São João River Basin (08.27- Figure 2- L), Una River Basin and Búzios Watersheds (08.28- Figure 2- M), Araruama Lagoon Watersheds (08.29- Figure 2- N), Saquarema Watersheds (08.30- Figure 2- O), Maricá Watersheds (08.31), Niterói Watersheds (08.32), Guanabara Bay Watersheds (08.33), Rio de Janeiro Watersheds (08.34), Sepetiba Watersheds (08.35), Mangaratiba Watersheds (08.36), Angra dos Reis Watersheds (08.37), Paraty Watersheds (08.38) and Cairuçu Watersheds (08.39). Rio Paraíba do Sul sections (08.22a to 08.22e).
Figure 2.
The twenty-nine river basin divisions along Rio de Janeiro. Coastal drainages: Itabapoana Basin (08.20- Figure 2- A), São Francisco de Itabapoana Watersheds (08.21- Figure 2- B)- , Lagoa Feia Watersheds (08.23- Figure 2- H), Jurubatiba Watersheds (08.24- Figure 2- I), Macaé River Basin (08.25- Figure 2- J), Rio das Ostras Watersheds (08.26- Figure 2- K), São João River Basin (08.27- Figure 2- L), Una River Basin and Búzios Watersheds (08.28- Figure 2- M), Araruama Lagoon Watersheds (08.29- Figure 2- N), Saquarema Watersheds (08.30- Figure 2- O), Maricá Watersheds (08.31), Niterói Watersheds (08.32), Guanabara Bay Watersheds (08.33), Rio de Janeiro Watersheds (08.34), Sepetiba Watersheds (08.35), Mangaratiba Watersheds (08.36), Angra dos Reis Watersheds (08.37), Paraty Watersheds (08.38) and Cairuçu Watersheds (08.39). Rio Paraíba do Sul sections (08.22a to 08.22e).
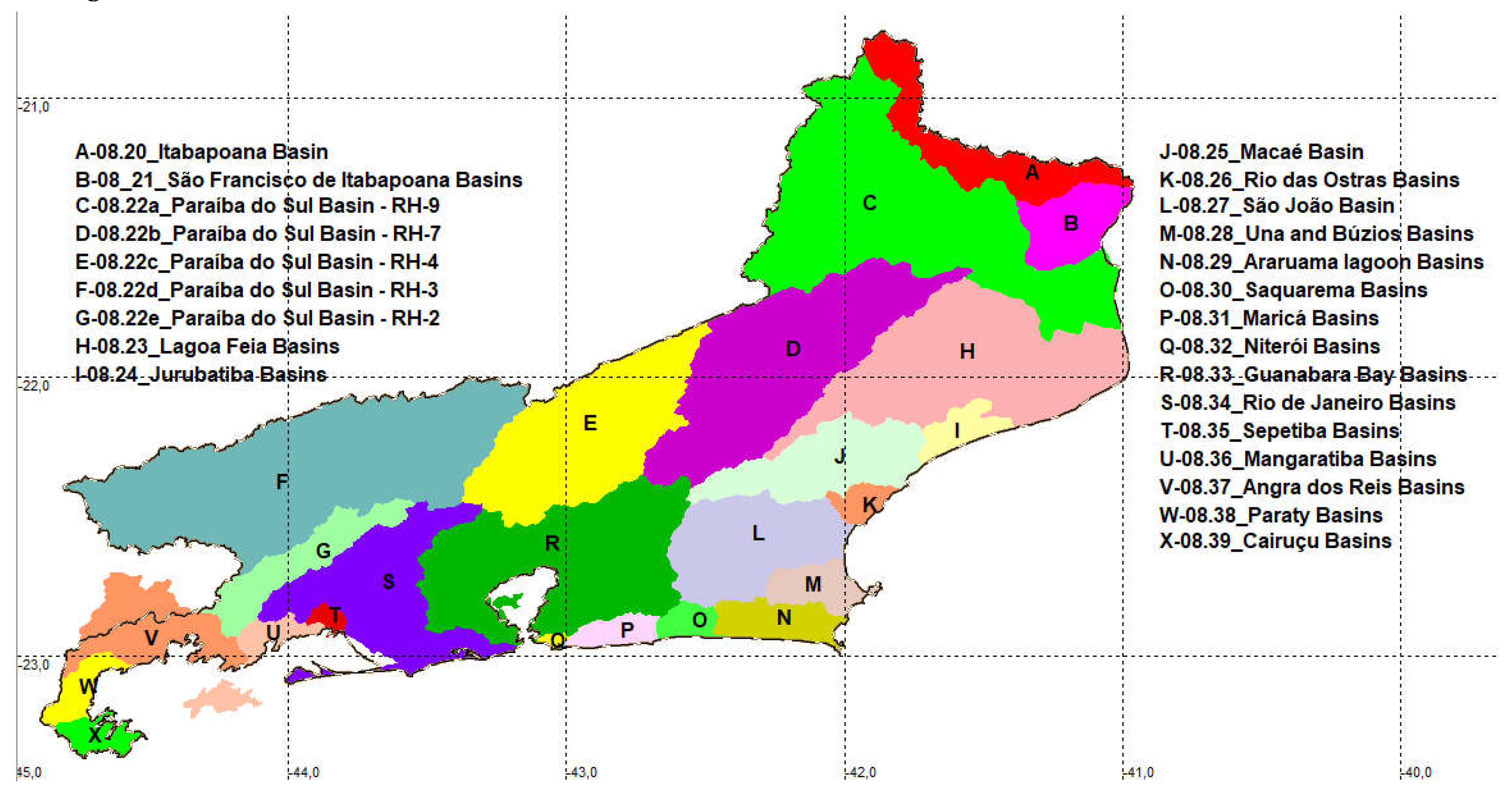
Figure 3.
Quality of sampling per hydrographic regions RH1 to RH-9. Green = good quality of both indices; yellow = average quality of both indices; red = poor quality of both indices.
Figure 3.
Quality of sampling per hydrographic regions RH1 to RH-9. Green = good quality of both indices; yellow = average quality of both indices; red = poor quality of both indices.
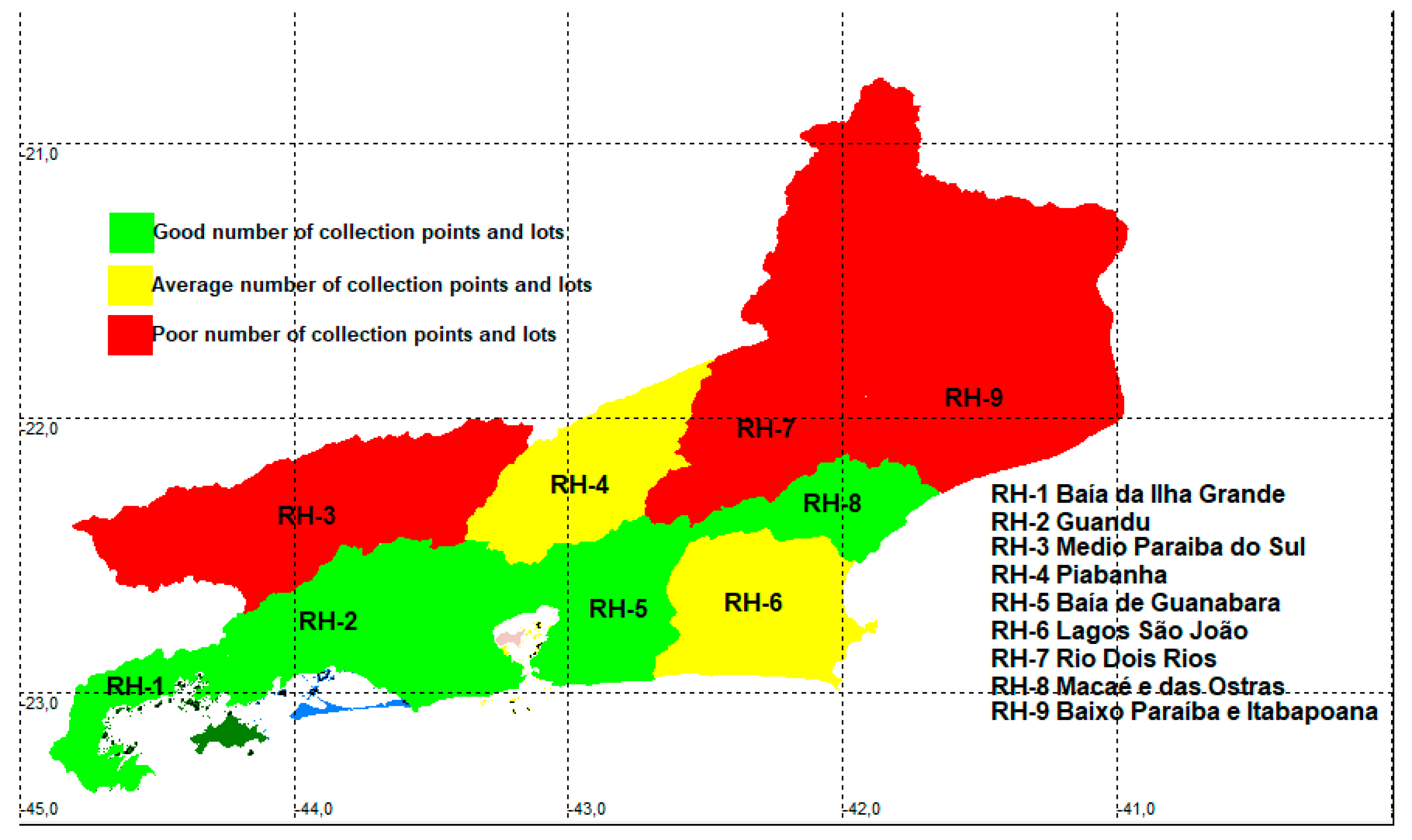
Figure 4.
Quality of sampling per municipality. Green = good quality of both indices; yellow = average quality of both indices; red = poor quality of both indices; orange = average number of collection points, good number of lots; pink = good number of collection points, average number of lots; blue = poor number of collection points, average number of lots; gray = good number of collection points, poor number of lots; black = average number of collection points, poor number of lots.
Figure 4.
Quality of sampling per municipality. Green = good quality of both indices; yellow = average quality of both indices; red = poor quality of both indices; orange = average number of collection points, good number of lots; pink = good number of collection points, average number of lots; blue = poor number of collection points, average number of lots; gray = good number of collection points, poor number of lots; black = average number of collection points, poor number of lots.
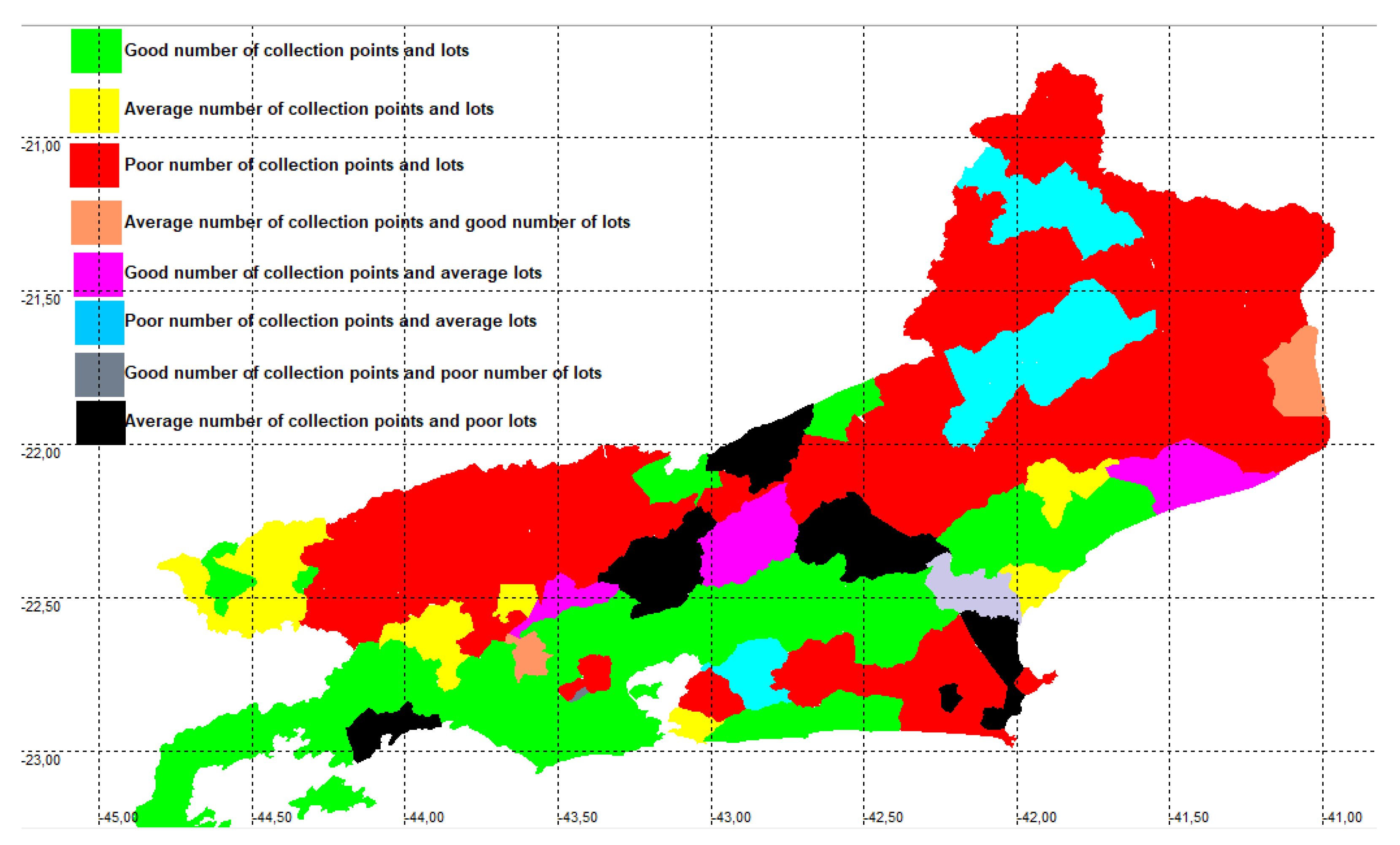
Figure 5.
Temporal variation in the cataloging of fish records in museum collections.
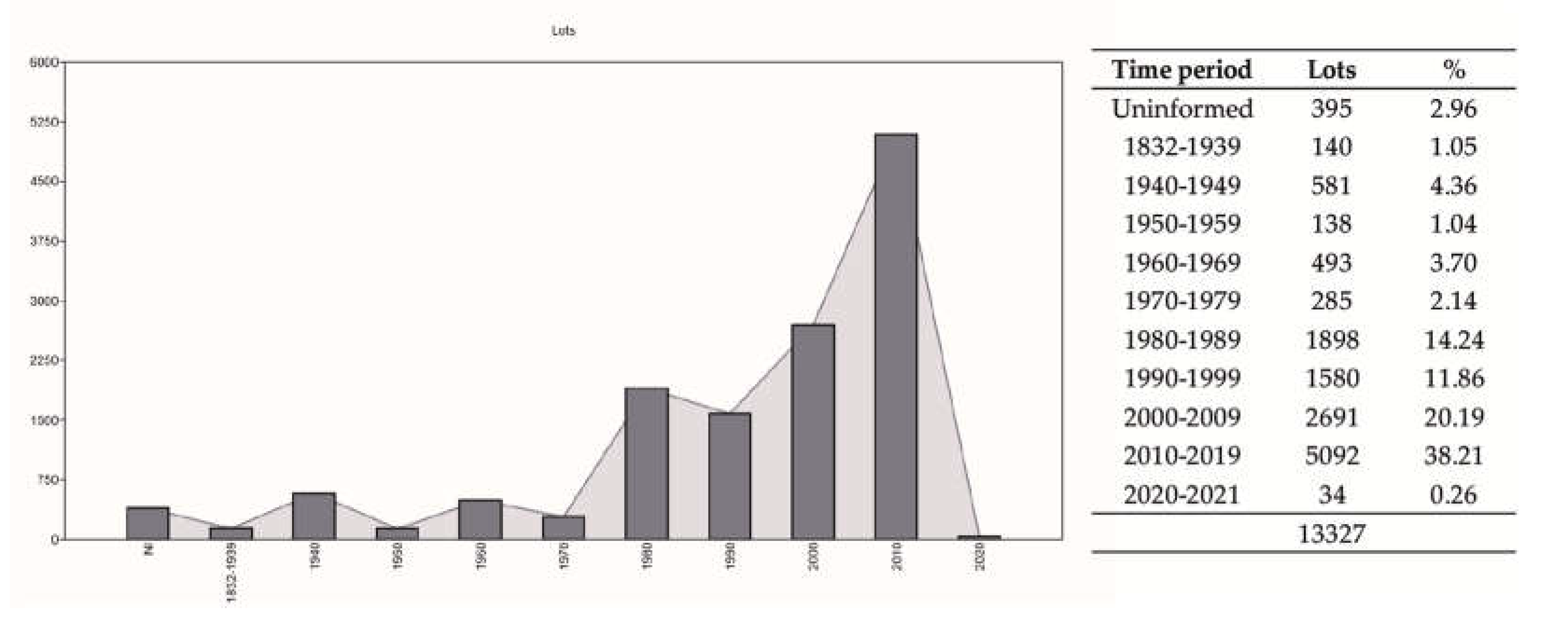
Figure 6.
Taxonomic representativeness of fish orders and families for all species (A, B) and freshwater native species (C, D) within the territory of Rio de Janeiro.
Figure 6.
Taxonomic representativeness of fish orders and families for all species (A, B) and freshwater native species (C, D) within the territory of Rio de Janeiro.
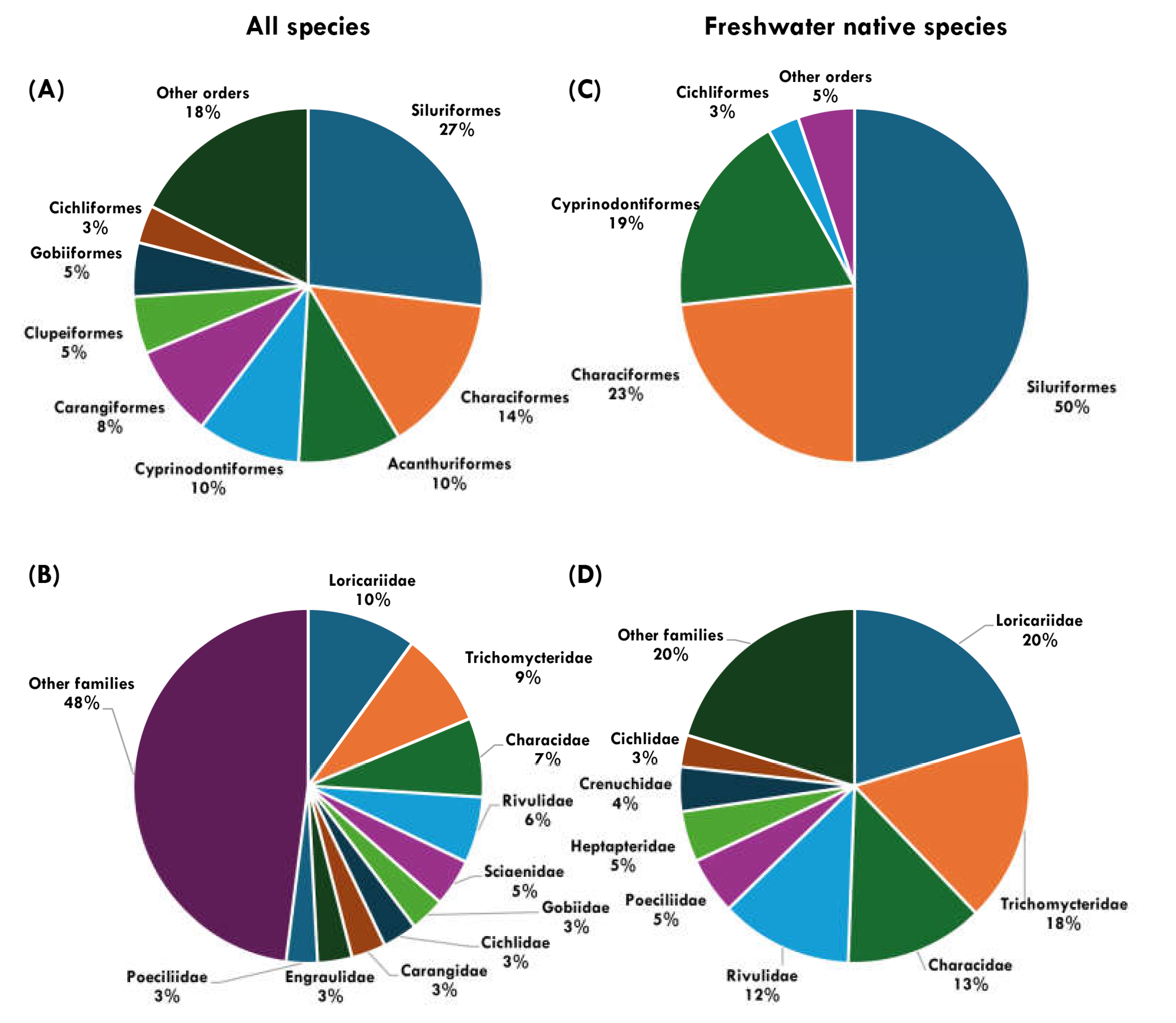
Figure 7.
Diversity profile using the Rényi series representing the fish fauna documented in the museum collections for the hydrographic regions of Rio de Janeiro. A. Diversity profile between RH-1 and RH-2. B. Profile between RH-1 and RH-9. C. Profile between RH-3, RH-4 and RH-7. D. Profile between RH-5, RH-6 and RH-8.
Figure 7.
Diversity profile using the Rényi series representing the fish fauna documented in the museum collections for the hydrographic regions of Rio de Janeiro. A. Diversity profile between RH-1 and RH-2. B. Profile between RH-1 and RH-9. C. Profile between RH-3, RH-4 and RH-7. D. Profile between RH-5, RH-6 and RH-8.
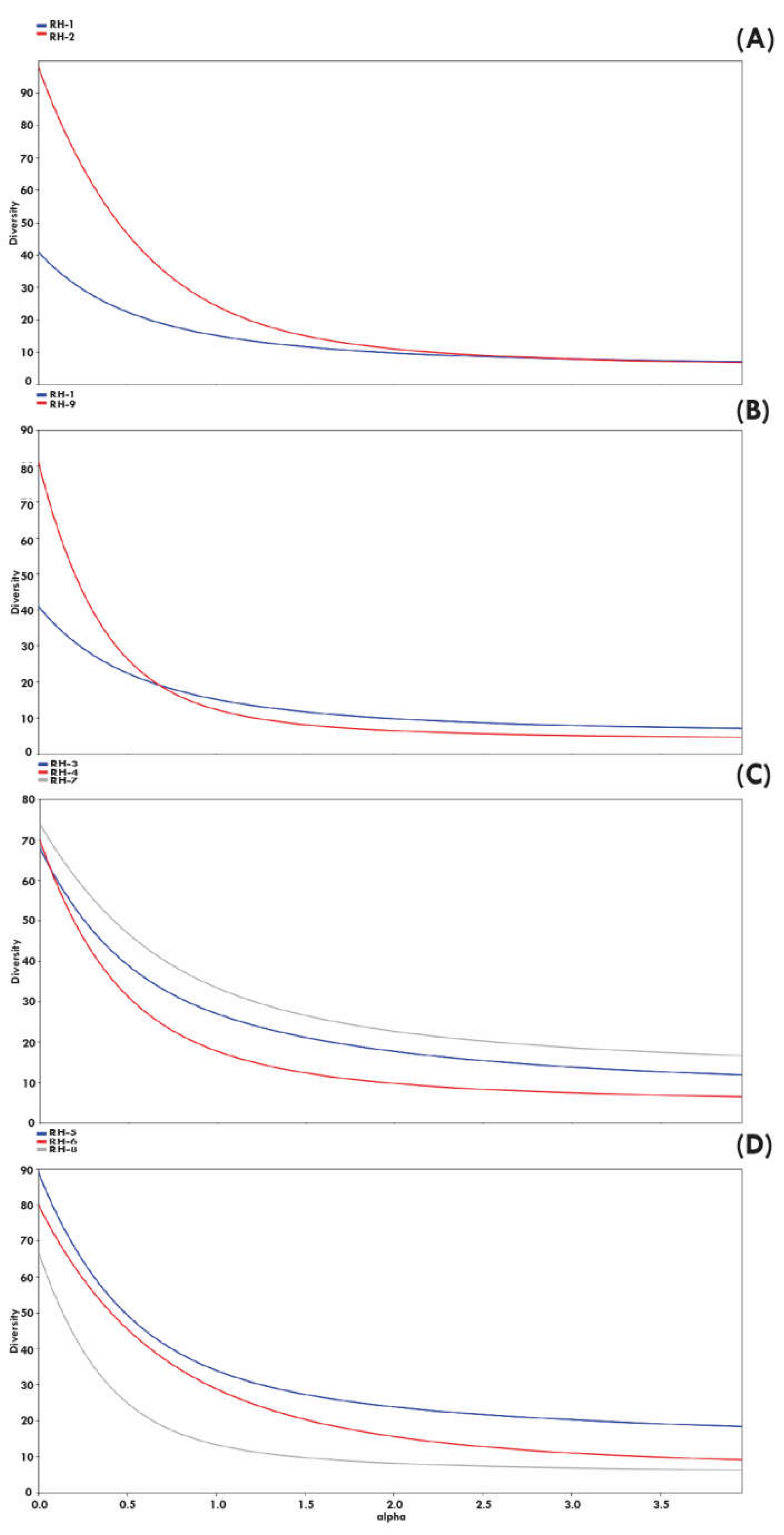
Table 1.
Lots sampled in the state of Rio de Janeiro.
| Ocean | Continental | Not identified | % | |
|---|---|---|---|---|
| Marine origin | 2,890 | 985 | ----- | 7.39% |
| Allochthonous freshwater | ----- | 498 | ----- | 3.74% |
| Native freshwater | ----- | 11,724 | ----- | 87.97% |
| Not identified at the species level | ----- | 120 | ----- | 0.90% |
| Not identified at the locality level | ----- | ----- | 82 | |
| Total | 2,890 | 13,327 | 82 | 16,299 |
| % | 17.73% | 81.77% | 0.50% |
Table 2.
Quality of sampling per hydrographic region.
| Code | Hydrographic region | Area (km2) | Lots | Points | il | ilq | ip | ipq |
|---|---|---|---|---|---|---|---|---|
| RH-1 | Baía da Ilha Grande | 1,919.1 | 1,150 | 140 | 59.9 | Good | 7.3 | Good |
| RH-2 | Guandu | 4,087.8 | 1,789 | 234 | 43.8 | Good | 5.7 | Good |
| RH-3 | Middle Paraíba do Sul | 7,114.1 | 860 | 133 | 12.1 | Poor | 1.9 | Poor |
| RH-4 | Piabanha | 3,831.4 | 816 | 121 | 21.3 | Average | 3.2 | Average |
| RH-5 | Baía de Guanabara | 696.0 | 3,190 | 428 | 458.3 | Good | 61.5 | Good |
| RH-6 | Lagos São João | 4,030.2 | 1,353 | 143 | 33.6 | Average | 3.5 | Average |
| RH-7 | Rio Dois Rios | 4,940.9 | 780 | 81 | 15.8 | Poor | 1.6 | Poor |
| RH-8 | Macaé and das Ostras | 2,226.9 | 1,474 | 133 | 66.2 | Good | 6.0 | Good |
| RH-9 | Lower Paraíba do Sul and Itabapoana | 14,904.0 | 1,915 | 210 | 12.8 | Poor | 1.4 | Poor |
| Rio de Janeiro State | 43,750 | 13,327 | 1.623 | 30.5 | 3.7 |
Table 3.
Sample quality per basin groups.
| Code | Basin groups | Area (km2) | Lots | Points | Il | Ilq | Ip | Ipq |
|---|---|---|---|---|---|---|---|---|
| 08.20 | Itabapoana | 1,523.4 | 122 | 19 | 8.0 | Poor | 1.2 | Poor |
| 08.21 | São Francisco do Itabapoana | 971.3 | 38 | 6 | 3.9 | Poor | 0.6 | Poor |
| 08.22a | Rio Paraíba do Sul - RH-9 | 6,321.4 | 1012 | 63 | 16.0 | Poor | 1.0 | Poor |
| 08.22b | Rio Paraíba do Sul - RH-7 | 4,468.6 | 780 | 81 | 17.5 | Poor | 1.8 | Poor |
| 08.22c | Rio Paraíba do Sul - RH-4 | 3,469.0 | 816 | 121 | 23.5 | Average | 3.5 | Average |
| 08.22d | Rio Paraíba do Sul - RH-3 | 6,430.8 | 860 | 133 | 13.4 | Poor | 2.1 | Poor |
| 08.22e | Rio Paraíba do Sul - RH-2 | 1,014.1 | 611 | 68 | 60.3 | Good | 6.7 | Good |
| 08.23 | Lagoa Feia | 4,310.8 | 512 | 93 | 11.9 | Poor | 2.2 | Poor |
| 08.24 | Jurubatiba | 410.1 | 707 | 62 | 172.4 | Good | 15.1 | Good |
| 08.25 | Rio Macaé | 1,706.4 | 894 | 92 | 52.4 | Good | 5.4 | Good |
| 08.26 | Rio das Ostras | 249.4 | 104 | 8 | 41.7 | Good | 3.2 | Average |
| 08.27 | Rio São João | 2,155.6 | 1052 | 86 | 48.8 | Good | 4.0 | Average |
| 08.28 | Rio Una and Búzios | 541.7 | 11 | 4 | 2.0 | Poor | 0.7 | Poor |
| 08.29 | Lagoa de Araruama | 677.5 | 67 | 16 | 9.9 | Poor | 2.4 | Poor |
| 08.30 | Saquarema | 265.2 | 210 | 35 | 79.2 | Good | 13.2 | Good |
| 08.31 | Maricá | 349.5 | 167 | 33 | 47.8 | Good | 9.4 | Good |
| 08.32 | Niterói | 51.5 | 23 | 6 | 44.7 | Good | 11.7 | Good |
| 08.33 | Baía de Guanabara | 4,073.8 | 2412 | 269 | 59.2 | Good | 6.6 | Good |
| 08.34 | Rio de Janeiro | 2,636.3 | 1673 | 269 | 63.5 | Good | 10.2 | Good |
| 08.35 | Sepetiba | 107.0 | 39 | 7 | 36.4 | Average | 6.5 | Good |
| 08.36 | Mangaratiba | 289.8 | 67 | 12 | 23.1 | Average | 4.1 | Average |
| 08.37 | Angra dos Reis | 1,028.2 | 557 | 59 | 54.2 | Average | 5.7 | Average |
| 08.38 | Paraty | 376.9 | 301 | 48 | 79.9 | Good | 12.7 | Good |
| 08.38 | Cairuçu | 322.2 | 292 | 33 | 90.6 | Good | 10.2 | Good |
| Rio de Janeiro State | 43,750 | 13,327 | 1,623 | 30.5 | 3.7 |
Table 4.
Number of fish species sampled in inland waters in the state of Rio de Janeiro.
| Environment/Origin | S | % |
|---|---|---|
| Native freshwater | 172 | 49.28% |
| Allochthonous freshwater | 22 | 6.30% |
| Marine origin | 152 | 43.55% |
| Not identified at the species level | 3 | 0.86% |
| Total | 349 |
Table 5.
Inventory of native freshwater species recorded for Rio de Janeiro state.
| Order | Family | Subfamily | Suggested species | Author |
|---|---|---|---|---|
| Gymnotiformes | Sternopygidae | Eigenmannia virescens | (Valenciennes, 1836) | |
| Gymnotidae | Gymnotinae | Gymnotus carapo | Linnaeus, 1758 | |
| Gymnotus pantherinus | (Steindachner, 1908) | |||
| Hypopomidae | Brachyhypopomus janeiroensis | (Costa & Campos-da-Paz, 1992) | ||
| Characiformes | Crenuchidae | Characidiinae | Characidium alipioi | Travassos, 1955 |
| Characidium grajahuense | Travassos, 1944 | |||
| Characidium interruptum | Pellegrin, 1909 | |||
| Characidium japuhybense | Travassos, 1949 | |||
| Characidium lauroi | Travassos, 1949 | |||
| Characidium litorale | Leitão & Buckup, 2014 | |||
| Characidium vidali | Travassos, 1967 | |||
| Erythrinidae | Hoplerythrinus unitaeniatus | (Spix & Agassiz, 1829) | ||
| Hoplias malabaricus | (Bloch, 1794) | |||
| Anostomidae | Hypomasticus copelandii | (Steindachner, 1875) | ||
| Hypomasticus mormyrops | (Steindachner, 1875) | |||
| Hypomasticus thayeri | (Borodin, 1929) | |||
| Megaleporinus conirostris | (Steindachner, 1875) | |||
| Curimatidae | Cyphocharax gilbert | (Quoy & Gaimard, 1824) | ||
| Prochilodontidae | Prochilodus lineatus | (Valenciennes, 1837) | ||
| Prochilodus vimboides | Kner, 1859 | |||
| Bryconidae | Bryconinae | Brycon insignis | Steindachner, 1877 | |
| Brycon opalinus | (Cuvier, 1819) | |||
| Characidae | Stethaprioninae | Astyanax keronolepis | Silva, Malabarba & Malabarba, 2019 | |
| Astyanax lacustris | (Lütken, 1875) | |||
| Deuterodon giton | (Eigenmann, 1908) | |||
| Deuterodon hastatus | (Myers, 1928) | |||
| Deuterodon heterostomus | (Eigenmann, 1911) | |||
| Deuterodon intermedius | (Eigenmann, 1908) | |||
| Deuterodon janeiroensis | (Eigenmann, 1908) | |||
| Deuterodon luetkenii | (Boulenger, 1887) | |||
| Deuterodon taeniatus | (Jenyns, 1842) | |||
| Hollandichthys multifasciatus | (Eigenmann & Norris, 1900) | |||
| Hyphessobrycon bifasciatus | Ellis, 1911 | |||
| Hyphessobrycon boulengeri | (Eigenmann, 1907) | |||
| Hyphessobrycon flammeus | Myers, 1924 | |||
| Oligosarcus hepsetus | (Cuvier, 1829) | |||
| Psalidodon parahybae | (Eigenmann, 1908) | |||
| Spintherobolinae | Spintherobolus broccae | Myers, 1925 | ||
| Stevardiinae | Bryconamericus microcephalus | (Miranda Ribeiro, 1908) | ||
| Bryconamericus ornaticeps | Bizerril & Perez-Neto, 1995 | |||
| Bryconamericus tenuis | Bizerril & Auraujo, 1992 | |||
| Knodus moenkhausii | (Eigenmann & Kennedy, 1903) | |||
| Mimagoniates microlepis | (Steindachner, 1877) | |||
| Piabina argentea | Reinhardt, 1867 | |||
| Siluriformes | Trichomycteridae | Trichogeninae | Trichogenes longipinnis | Britski & Ortega, 1983 |
| Trichomycterinae | Trichomycterus albinotatus | Costa, 1992 | ||
| Ituglanis parahybae | (Eigenmann, 1918) | |||
| Trichomycterus auroguttatus | Costa, 1992 | |||
| Trichomycterus caipora | Lima, Lazzarotto & Costa, 2008 | |||
| Trichomycterus claudiae | Barbosa & Costa, 2010 | |||
| Trichomycterus florensis | (Miranda-Ribeiro, 1943) | |||
| Trichomycterus fuliginosus | Barbosa & Costa, 2010 | |||
| Trichomycterus giganteus | Lima & Costa, 2004 | |||
| Trichomycterus goeldii | Boulenger, 1896 | |||
| Trichomycterus itatiayae | Miranda Ribeiro, 1906 | |||
| Trichomycterus jacupiranga | Wosiacki & Oyakawa, 2005 | |||
| Trichomycterus largoperculatus | Costa & Katz, 2022 | |||
| Trichomycterus macrophthalmus | Barbosa & Costa, 2012 | |||
| Trichomycterus mariamole | Barbosa & Costa, 2010 | |||
| Trichomycterus mirissumba | Costa, 1992 | |||
| Trichomycterus nigricans | Valenciennes, 1832 | |||
| Trichomycterus nigroauratus | Barbosa & Costa, 2008 | |||
| Trichomycterus paquequerense | (Miranda Ribeiro, 1943) | |||
| Trichomycterus potschi | Barbosa & Costa, 2003 | |||
| Trichomycterus puriventris | Barbosa & Costa, 2012 | |||
| Trichomycterus travassosi | (Miranda Ribeiro, 1949 | |||
| Trichomycterus vitalbrazili | Vilardo, Katz & Costa, 2020 | |||
| Microcambevinae | Listrura costai | Villa-Verde, Lazzarotto & Lima, 2012 | ||
| Listrura nematopteryx | de Pinna, 1988 | |||
| Listrura tetraradiata | Landim & Costa, 2002 | |||
| Microcambeva barbata | Costa & Bockmann, 1994 | |||
| Microcambeva bendego | Medeiros, Moreira, de Pinna & Lima, 2020 | |||
| Stegophilinae | Homodiaetus banguela | Koch, 2002 | ||
| Homodiaetus passarellii | (Miranda Ribeiro, 1944 | |||
| Callichthyidae | Callichthyinae | Callichthys callichthys | (Linnaeus, 1758) | |
| Hoplosternum littorale | (Hancock, 1828) | |||
| Corydoradinae | Corydoras nattereri | Steindachner, 1876 | ||
| Scleromystax barbatus | (Quoy & Gaimard, 1824) | |||
| Scleromystax prionotos | (Nijssen & Isbrücker, 1980) | |||
| Loricariidae | Delturinae | Delturus parahybae | Eigenmann & Eigenmann, 1889 | |
| Hemipsilichthys gobio | (Lütken, 1874) | |||
| Hemipsilichthys nimius | Pereira, Reis, Souza & Lazzarotto, 2003 | |||
| Hemipsilichthys papillatus | Pereira, Oliveira & Oyakawa, 2000 | |||
| Rhinelepinae | Pogonopoma parahybae | (Steindachner, 1877) | ||
| Loricariinae | Harttia carvalhoi | Miranda Ribeiro, 1939 | ||
| Harttia loricariformis | Steindachner, 1877 | |||
| Loricariichthys castaneus | (Castelnau, 1855) | |||
| Rineloricaria nigricauda | (Regan, 1904) | |||
| Rineloricaria nudipectoris | Mejia, Ferrar & Buckup, 2023 | |||
| Rineloricaria steindachneri | (Regan, 1904) | |||
| Rineloricaria zawadzki | Silva, Costa & Oliveira, 2022 | |||
| Hypoptopomatinae | Hisonotus notatus | Eigenmann & Eigenmann, 1889 | ||
| Hisonotus thayeri | Martins & Langeani, 2016 | |||
| Kronichthys heylandi | (Boulenger, 1900) | |||
| Neoplecostomus microps | (Steindachner, 1877) | |||
| Neoplecostomus paraty | Cherobim, Lazzarotto & Langeani, 2016 | |||
| Neoplecostomus variipictus | Bizerril, 1995 | |||
| Otocinclus affinis | Steindachner, 1877 | |||
| Otothyris lophophanes | (Eigenmann & Eigenmann, 1889) | |||
| Pareiorhaphis garbei | (Ihering, 1911) | |||
| Pareiorhina brachyrhyncha | Chamon, Aranda & Buckup, 2005 | |||
| Pareiorhina rudolphi | (Miranda Ribeiro, 1911) | |||
| Parotocinclus bidentatus | Gauger & Buckup, 2005 | |||
| Parotocinclus fluminense | Roxo, Melo, Silva & Oliveira, 2017 | |||
| Parotocinclus maculicauda | (Steindachner, 1877) | |||
| Parotocinclus muriaensis | Gauger & Buckup, 2005 | |||
| Pseudotothyris janeirensis | Britski & Garavello, 1984 | |||
| Schizolecis guentheri | (Miranda Ribeiro, 1918) | |||
| Hypostominae | Ancistrus multispinis | (Regan, 1912) | ||
| Hypostomus affinis | (Steindachner, 1877) | |||
| Hypostomus auroguttatus | Kner, 1854 | |||
| Hypostomus luetkeni | (Steindachner, 1877) | |||
| Hypostomus punctatus | Valenciennes, 1840 | |||
| Hypostomus vermicularis | (Eigenmann & Eigenmann, 1888) | |||
| Auchenipteridae | Centromochlinae | Glanidium melanopterum | Miranda Ribeiro, 1918 | |
| Auchenipterinae | Trachelyopterus striatulus | (Steindachner, 1877) | ||
| Heptapteridae | Rhamdiinae | Pimelodella lateristriga | (Lichtenstein, 1823) | |
| Rhamdia quelen | (Quoy & Gaimard, 1824) | |||
| Heptapterinae | Acentronichthys leptos | Eigenmann & Eigenmann, 1889 | ||
| Imparfinis minutus | (Lütken, 1874) | |||
| Imparfinis piperatus | Eigenmann & Norris, 1900 | |||
| Rhamdioglanis frenatus | Ihering, 1907 | |||
| Rhamdioglanis transfasciatus | Miranda Ribeiro, 1908 | |||
| Taunayia bifasciata | (Eigenmann & Norris, 1900) | |||
| Pimelodidae | Pimelodus maculatus | Lacepède, 1803 | ||
| Steindachneridion parahybae | (Steindachner, 1877) | |||
| Pseudopimelodidae | Microglanis nigripinnis | Bizerril & Perez-Neto, 1992 | ||
| Microglanis parahybae | (Steindachner, 1880) | |||
| Microglanis pleriqueater | Mattos, Ottoni & Barbosa, 2013 | |||
| Ariidae | Ariinae | Paragenidens grandoculis | (Steindachner, 1877) | |
| Gobiiformes | Eleotridae | Eleotrinae | Dormitator maculatus | (Bloch, 1792) |
| Eleotris pisonis | (Gmelin, 1789) | |||
| Oxudercidae | Gobionellinae | Awaous tajasica | (Lichtenstein, 1822) | |
| Synbranchiformes | Synbranchidae | Synbranchus marmoratus | Bloch, 1795 | |
| Cyprinodontiformes | Rivulidae | Rivulinae | Atlantirivulus guanabarensis | Costa, 2014 |
| Atlantirivulus janeiroensis | (Costa, 1991) | |||
| Atlantirivulus jurubatibensis | (Costa, 2008) | |||
| Atlantirivulus lazzarotoi | (Costa, 2007) | |||
| Atlantirivulus maricensis | Costa, 2014 | |||
| Atlantirivulus simplicis | (Costa, 2004) | |||
| Kryptolebiatinae | Kryptolebias brasiliensis | (Valenciennes, 1821) | ||
| Kryptolebias caudomarginatus | (Seegers, 1984) | |||
| Kryptolebias gracilis | Costa, 2007 | |||
| Kryptolebias ocellatus | (Hensel, 1868) | |||
| Cynolebiinae | Leptolebias marmoratus | (Ladiges, 1934) | ||
| Leptopanchax citrinipinnis | (Costa, Lacerda & Tanizaki, 1988) | |||
| Leptopanchax opalescens | (Myers, 1942) | |||
| Leptopanchax sanguineus | Costa, 2019 | |||
| Leptopanchax splendens | (Myers, 1942) | |||
| Nematolebias papilliferus | Costa, 2002 | |||
| Nematolebias whitei | (Myers, 1942) | |||
| Notholebias cruzi | (Costa, 1988) | |||
| Notholebias fractifasciatus | (Costa, 1988) | |||
| Notholebias minimus | (Myers, 1942) | |||
| Ophthalmolebias constanciae | (Myers, 1942) | |||
| Poeciliidae | Poeciliinae | Phalloceros anisophallos | Lucinda, 2008 | |
| Phalloceros enneaktinos | Lucinda, 2008 | |||
| Phalloceros harpagos | Lucinda, 2008 | |||
| Phalloceros leptokeras | Lucinda, 2008 | |||
| Phalloceros tupinamba | Lucinda, 2008 | |||
| Phalloptychus januarius | (Hensel, 1868) | |||
| Phallotorynus fasciolatus | Henn, 1916 | |||
| Poecilia reticulata | Peters, 1859 | |||
| Poecilia vivipara | Bloch & Schneider, 1801 | |||
| Anablepidae | Anablepinae | Jenynsia darwini | Amorim, 2018 | |
| Jenynsia lineata | (Jenyns, 1842) | |||
| Cichliformes | Cichlidae | Cichlinae | Australoheros ipatinguensis | Ottoni & Costa, 200 |
| Australoheros oblongus | (Castelnau 1855) | |||
| Crenicichla lacustris | (Castelnau, 1855) | |||
| Crenicichla lepidota | Heckel, 1840 | |||
| Geophagus brasiliensis | (Quoy & Gaimard, 1824) | |||
| Acanthuriformes | Sciaenidae | Pachyurus adspersus | Steindachner, 1879 |
Table 6.
Parametric and non-parametric diversity estimation indices per hydrographic region in the state of Rio de Janeiro.
Table 6.
Parametric and non-parametric diversity estimation indices per hydrographic region in the state of Rio de Janeiro.
| RH-1 | RH-2 | RH-3 | RH-4 | RH-5 | RH-6 | RH-7 | RH-8 | RH-9 | |
|---|---|---|---|---|---|---|---|---|---|
| Taxa_S | 40 | 97 | 67 | 69 | 89 | 80 | 74 | 67 | 81 |
| Individuals | 20058 | 24736 | 4581 | 9256 | 29430 | 11829 | 4243 | 27928 | 40232 |
| Dominance_D | 0.1032 | 0.0915 | 0.05703 | 0.1038 | 0.04186 | 0.06386 | 0.04392 | 0.1214 | 0.1549 |
| Shannon_H | 2.7 | 3.176 | 3.284 | 2.858 | 3.528 | 3.366 | 3.516 | 2.595 | 2.513 |
| Equitability_J | 0.732 | 0.6944 | 0.7811 | 0.675 | 0.786 | 0.7682 | 0.8169 | 0.6172 | 0.5719 |
| Chao-1 | 40 | 97.17 | 74 | 78.33 | 90.5 | 80.17 | 75.5 | 70 | 102 |
| iChao-1 | 40 | 97.17 | 78.57 | 86.75 | 93.5 | 80.4 | 77.07 | 76.5 | 103.7 |
| ACE | 40 | 97.3 | 73.45 | 74.2 | 90.16 | 81.2 | 75.69 | 68.96 | 85.34 |
| Squares | 40 | 97.09 | 70.66 | 75.65 | 89.38 | 80.26 | 74.71 | 68.94 | 88.59 |
| Variation of species richness for the non-parametric estimation indices | |||||||||
| RH-1 | RH-2 | RH-3 | RH-4 | RH-5 | RH-6 | RH-7 | RH-8 | RH-9 | |
| Chao-1 | 0.00% | 0.18% | 10.45% | 13.52% | 1.69% | 0.21% | 2.03% | 4.48% | 25.93% |
| iChao-1 | 0.00% | 0.18% | 17.27% | 25.72% | 5.06% | 0.50% | 4.15% | 14.18% | 28.02% |
| ACE | 0.00% | 0.31% | 9.63% | 7.54% | 1.30% | 1.50% | 2.28% | 2.93% | 5.36% |
| Squares | 0.00% | 0.09% | 5.46% | 9.64% | 0.43% | 0.32% | 0.96% | 2.90% | 9.37% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



