
Submitted:
10 July 2024
Posted:
11 July 2024
You are already at the latest version
Abstract
Autism spectrum disorder (ASD) is a complex neurodevelopmental disorder, with a highly variable expression of phenotypes (restricted interest or activity and repetitive behavior in communication and social interactions), genes (mutation), markers (alteration of transcription) and pathways. Loss of function of the CC2D1A gene appears to primarily affect the brain, leading to a range of behavioral problems in humans. In our study published in 2020; we found that the expressions of miR-19a-3p, miR-361-5p, miR-150-5p, miR-3613-3p, miR-126-3p and miR-499a-5p were downregulated in the serum samples of autistic patients and their families. Here, acquired non-Mendelian hereditary character in a genetically defined mouse model of autism (Cc2d1a +/-) correlates with the transcriptional alteration of five miRNAs. We seek to test the hypothesis that miRNA levels are varying by changes in RNA/DNA structure during development, thereby creating transcription alteration. Behavioral tests were conducted on the offspring of Cc2d1a +/- mutant and control mice, such as novel object, social interaction, marble burying and tail suspension behavior. Two RNA fractions were isolated from mouse hippocampal tissues and sperm cells by standard TRIzol extraction: total RNA and fraction of RNA bound to DNA in the form of a DNA/RNA hybrid (R-loop). The expression levels of miR-19a-3p, miR-361-5p, miR-150-5p, miR-126-3p and miR-499a-5p were investigated by quantitative real-time RT-PCR. We report differences in the distribution of five miRNAs in the hippocampus between male and female mice in RNA fractions, particularly in colonies of Cc2d1a +/- mice. Furthermore, the number of miRNAs engaged in the DNA/RNA hybrid fraction is generally higher in the mutant pedigree than in the control group. On the other hand, in sperm both fractions are at lower levels than in controls. R-loops contribute to the physiology and pathology of organisms including human disease. Here we report a variation in five miRNA levels between gender and tissue. Our results suggest that the transcription levels of these five miRNAs are directly regulated by their RNA.
Keywords:
Cc2d1a
; Autism
; microRNA
; transcripts
; DNA/RNA hybrid
; non-Mendelian heredity
1. Introduction
Autism spectrum disorder (ASD) is one of the most common neurodevelopmental disorders, affecting approximately 1 in 36 children [1]. ASD affects 4 times more boys than girls and appears early, with an average age of diagnosis before 3 years [2]. Diagnosis of ASD often reveals various problems such as gastrointestinal system abnormalities, epilepsy, intellectual disability (ID), sleep and motor difficulties [3,4]. These disorders result from complex interactions between environmental and genetics factors, with a heritability ranging from 40 to 80% [5]. ASD risk factors strongly influence synapse connectivity [6,7].
A large homozygous deletion has been reported in the CC2D1A gene, and loss of gene is associated with non-syndromic (ID) in human patients, suggesting that CC2D1A functions in the brain. CC2D1A protein is a transcription factor and suppressor of the HTR1A gene. In our previous study, the expression levels of the CC2D1A transcripts (q-PCR) were increased in ASD patients without mutation in autism linked known genes compared to controls [8]. Mice lacking Cc2d1a develop normally during the uterine period, but die immediately after birth due to swallowing and respiratory deficiencies. Tissues specific (hypothalamus) Cc2d1a in conditional knock-out mice exhibit learning, memory and social deficits, hyperactivity, repetitive behaviors and anxiety that are key features of autism and ID [9,10]. These results confirm that CC2D1A is one of the new candidate gene for ASD. Heterozygous Cc2d1a mouse model showed sex-specific differences and non-Mendelian heritable changes are transferred to offspring, even those with a wild-type genotype [9-12].
Several studies revealed various RNA population in sperm [13-15], but only recently was reported about the possible presence of DNA-RNA hybrids (R-loops). It has been shown that a significant portion of RNA is actually made up of complexes similar to the R-loop, in the form of three-chain structures consisting of an RNA-DNA hybrid and a single strand of DNA [16-18]. Since the discovery of R-loops, one of the most abundant non-B-DNA structures in mammalian genomes [19], various types of biological functions and consequences have been characterized, including their regulatory potential [20]. During spermatogenesis the histone-protamine transition ensures the integrity of paternal DNA in the sperm head. An incomplete transition from histone to protamine leaves residual histones-associated genomic regions, leading to an open chromatin state and DNA-RNA interactions [21]. Similarly, R-loops bring together canonical histone modifications, capable of influencing DNA-protein-interactions, and reconstructing a specific spatial landscape [19]. Additionally, the three-stranded nature of R-loops affects the broader chromatin conformation of the sperm genome and may therefore play a locally antagonistic role to protamine tight packaging.
MicroRNAs (miRNAs) are small non-coding RNA molecules consisting of about 22 nucleotides that act as post-transcriptional regulators of gene expression [22,23]. In mammalian cells, miRNAs were discovered to create double strand RNA by base pairing with complementary sequences within messenger RNA (mRNA) molecules, which often results in inactivation of transcripts through translational suppression [24,25]. In a previous study we conducted, the expression levels of six miRNAs (miR-19a-3p, miR-361-5p, miR-3613-3p, miR-15a-5p, miR-126-3p and miR-499a-5p) were downregulated in blood serum levels of patients with autism [9]. Based on this study, we attempt to investigate the down regulation of these six miRNAs in autism. In this study; our objective was to discover the molecular differences that participate in the down regulations of the expression profile of five of six miRNAs (miR-19a-3p, miR-361-5p, miR-150a-5p, miR-126-3p and miR-499a-5p) compared to the controls (miR-3613-3p, does not exist in mouse genome). Here, we fractionated cellular RNAs using two different isolation method; extraction of total RNA with the standard TRIzol method and isolation of the DNA-associated RNA fraction as DNA/RNA hybrid (R-loop) from blood, hippocampal and sperm tissues of Cc2d1a (+/-) and control mice. By comparing the levels of these miRNAs, we found differences in the fraction of RNA attached to DNA between sexes and tissues, suggesting that these changes would affect the expression levels. We suggest that miRNAs involved in the R-loop harbor memory in the development of autism.
2. Material and Methods
2.1. Preparing the Animal Groups
In the project, heterozygous (+/-) Cc2d1a mice with Balb-C background and Balb-C control mice were used. Mice kept in a 12-hour light and 12-hour dark light period so that their circadian rhythms were not disrupted. In the experimental group, Cc2d1a (+/-) were mated with normal Balb-C mice. Before starting the experiments, the mice were allowed to acclimatize to the new environment for at least one week. Heterozygote and wild type genotypes were identified by PCR according to the instructions with the 3 different oligonucleotides (Sener et al., 2023). 10 male and 10 female (+/-) genotype mice were selected from the nail tissues of each animal in the resulting F1 generation. When these heterozygous and normal genotyped mice were two months old behavioral tests: tail suspension, marble and novel object behavior were carried out. The control group consisted of wild-type Balb-C mice. 10 male and 10 female animals were used. This study was approved by the Animal Ethics Committee of Erciyes University (09.09.2020, 20/124).
2.2. Behavior Tests
All tests were performed under normal lighting in the light phase. We started with 10 males and females in each cohort. To minimize possible changes, all behavioral tests were performed by the same investigator from the Transgenic unit. All animal behavior was filmed, tracked, and analyzed with EthoVision video tracking systems. The tail suspension test (TST) shows behavioral despair in a stressful situation. The TST was performed in a quiet laboratory. The animal’s immobility time was recorded to determine the depression status [26]. The animal was suspended above the ground by its tail with a tape for 6 min and recorded with a video. Total immobility time and immobility latency for each one-minute block were scored. The time from the start of the test until the mice ceased struggling was recorded as latency. The total time mice spent motionless for at least 2 s was recorded as total immobility time [12]. The social interaction test was conducted to measure sociability and as described in our previous study [9]. The test mouse was introduced centrally to initiate habituation for 10 min while blocking access to the side compartments. The accumulated time and number of visits to the interaction zone were measured to quantify the mice’s social behavior. The Novel Object Recognition (NOR) task is a well-established test in various animal models and has been performed to assess cognitive ability. The NOR task consisted of habituation, training, and probe testing during 5 days of the experiment. In the first trial, we used (first-day acquisition) animals exposed to two similar objects (small orange boxes) in a chamber for 5 min. During the second trial (second-day retention), mice were again exposed to two different objects for 5 min, including a familiar object from the first trial and a novel object (blue box). During each test, the boxes and objects were cleaned with 70% ethanol. The time spent exploring the novel object and the time spent exploring the familiar object were analyzed by researchers blinded to the experiment. The marble burying test is used to describe anxiety The experiment was carried out under dim lighting in a quiet room to reduce the influence of anxiety on behavior. Standard glass marbles were washed with mild laboratory detergent, rinsed with distilled-deionized water and dried, then spaced out evenly in five rows of four marbles on top of the bedding. The mice were left in the cage with the marbles for 30 minutes. Test recording began immediately after the animal was placed in the cage, as far away from the marbles as possible. Marbles were counted and scored buried if two-thirds of their surface was covered with bedding [26].
2.3. Tissue Collection
After the behavioral experiment, mice with Cc2d1a (+/+) and Cc2d1a (+/-) genotypes were euthanized by cervical dislocation. Sperm and hippocampal tissues were collected. RNA samples with two different methodologies were obtained from these tissues (total RNA isolation and hybrid DNA/RNA isolation). Total RNA was extracted from sperm and hippocampus of Cc2d1a heterozygous mice with the TRIzol method and the DNA/RNA hybrid fraction was isolated as hybrid DNA/RNA (R-loop) with a manual kit [17].
2.4. Total RNA and Hybrid DNA/RNA Isolation from Hippocampal Tissue
The hippocampus is divided in half and transferred to TRIzol by adding 200 µl of chloroform. The aqueous phase is transferred to a new tube. The isopropanol is added proportionally to the aqueous phase. After centrifugation, the supernatant is discarded. 75% ethanol is added to the pellet. After centrifugation, the supernatant is discarded. 50 µl of nuclease-free water are added for dilution. The Zymo Research kit (Catalog No: D7003) was used for hybrid DNA/RNA isolation. Isolation was performed from hippocampal tissue according to the manufacturer’s protocol. Alternatively, for high concentrations of DNA and RNA, an elution of ≥50 µl was used. Tissues can be mechanically homogenized for optimal extraction efficiency. The samples were stored at -80ºC after isolation.
2.5. RNA and Hybrid DNA/RNA Isolation from Sperm Tissue
Sperm is extracted from the canal taken from mice. Half of the treated spermatozoa are introduced into a falcon tube for total RNA isolation. The other half of the treated sperm is introduced into another falcon tube for hybrid DNA/RNA isolation. After centrifugation, the supernatant is transferred to a new falcon. The DTT and TRIzol mixture was added and kept on ice for 5 minutes. Then 200 µl of chloroform was added. Isopropanol, equal to its own volume, was added to the aqueous phase and 1ml of 70% ethanol was added. The supernatant is discarded and 50 µl of nuclease-free water is used for dilution. The Zymo Research kit (Catalog No: D7003) was used for hybrid DNA/RNA isolation. Isolation was performed from sperm according to the manufacturer’s protocol. The samples were stored at -80ºC after isolation.
2.6. Complementary DNA (cDNA) Synthesis and miRNA Profiling
The amount of cDNA was determined with the Takara cDNA synthesis kit. The reaction mixture was prepared to make cDNA, with 3.75 µL of RNA, 5 µL of mRQ buffer and 1.25µL of mRQ enzyme kit to make a total volume of 10 µL. The reverse transcription step was performed at 37◦C for 60 min, followed by 5 min at 85◦C. The cDNA was stored at 4◦C until use. The expression levels of 5 miRNA (miR-19a-3p, miR-361-5p, miR-150-5p, miR-126 and miR-499a-5p) were measured using a chain reaction by real-time quantitative polymerase (qRT-PCR). The TAKARA SYBR® Green PCR Kit was used, as well as specific primers. U6 was used as a reference gene. After the initial denaturation at 95◦C for 15 min, the qPCR cycles were as follows: 40 cycles of denaturation at 94oC for 15 s, annealing at 55oC for 30 s, and 70oC for 30 s, reading of plate. Finally, the PCR was completed after 15 seconds at 60◦C with Roche LightCycler. The relative expression levels of miRNAs were normalized with U6 using the delta delta CT (2-ΔΔ CT) method.
2.7. Statistical Analysis
Data were presented as means ± SEM. The Shapiro-Wilk test was first applied to confirm the normality of the data. When data followed a normal distribution, The Student’s t-test (unpaired) was used to compare two groups. One-way analysis of variance (ANOVA) with Tukey’s post-hoc test for comparisons between multiple groups. P-values less than 0.05 were considered statistically significant. The GraphPad Prism program (version 8) was used to evaluate the data and plot the graphs.
3. Results
3.1. Behavioral Gender Differences Are greater in Autism
Two groups of offspring are produced by mating Cc2d1a+/- with controls partner (G1) or with a Cc2d1a+/- partner (G2) see Materials and Methods. Behaviors are clearly different in the offspring of Cc2d1a+/- offspring in both groups (G1 and G2) compared to controls.
Differences were found between groups by gender (Figure 1). Immobility time differed when the control group and the two Cc2d1a groups (G1 and G2) were compared (Figure 1). Immobility time was increased in G1 and G2+/+ and +/- genotypes compared to the control. The number of buried beads differed significantly between the control group and the Cc2d1a groups (p=0.0158). The number of buried marbles in the G1 and G2+/+ and +/- genotypes was compared to the control. According to the social interaction test, males and females spent more time in the empty cage, which was statistically significant (p<0.0001; Figure 1E).
In the novel object test, the distance traveled differed significantly between the control and Cc2d1a groups (p=0.0163, Figure 1F). The time spent with the novel object and the familiar object also differed. The distance traveled in the G1 and G2+/+ and +/- genotypes and compared to the control was statistically significant (Figure 1G). Differences were also found between groups by gender (Figure 1H, 1I).
3.2. Sex-Dependent Distribution of Two Free or R-loop miRNA Fractions in Autism
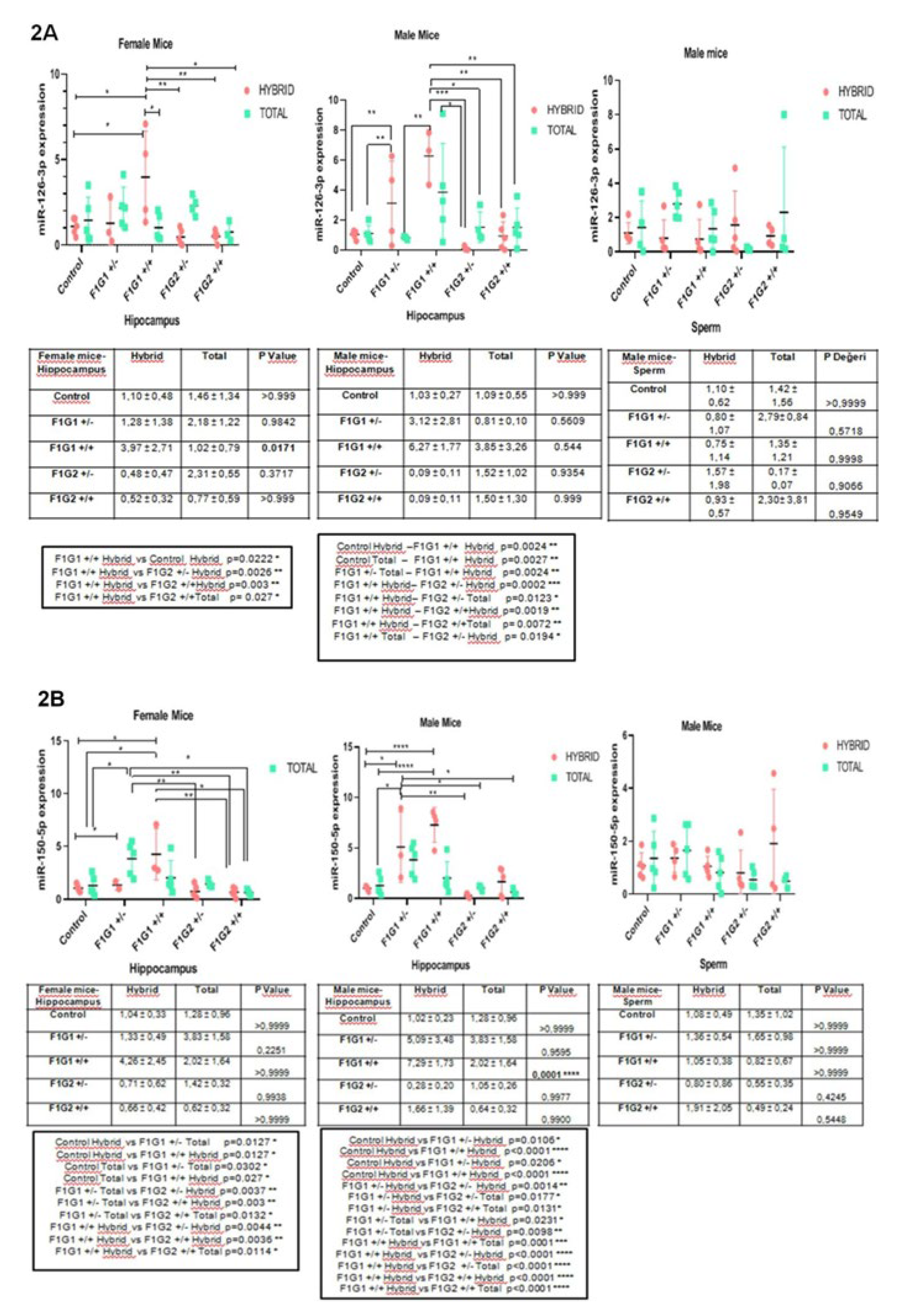
In Figure 2 shows the results of miRNAs levels in the hippocampus (male and female) and in the spermatozoa cells in all groups (G1, G2 and controls see above and Materials and Methods). Results indicate tissue-and sex dependent changes in the distribution of five miRNAs as fractions of the total relative to RNA engaged in R-loops. The levels of differences are specific to a given miRNAs and tissues.
The expression profiles of miR-126-3p are shown in Figure 2A. Differences are observed in the hippocampus in both sexes. MiR-126-3p was higher in the total RNA fraction in the hippocampus of female mice, the expression of miR-126-3p is higher and significantly different in males in G1 and G2 in the hybrid fraction of RNA. In sperm, the total RNA fraction was higher in G1 and G2 (Figure 2A).
MiR-150-5p expression profiles are shown in Figure 2B. Higher expression of miR-150-5p is observed in female hippocampus in G1 and G2 in the total (free) RNA fraction group. While a decrease is observed in all groups except G2+/- in the male hippocampus (p<0.0001). In sperm, there was a decrease in the (free) RNA fraction of the total group compared to the hybrid, with the exception of F1G1 +/- (Figure 2B).
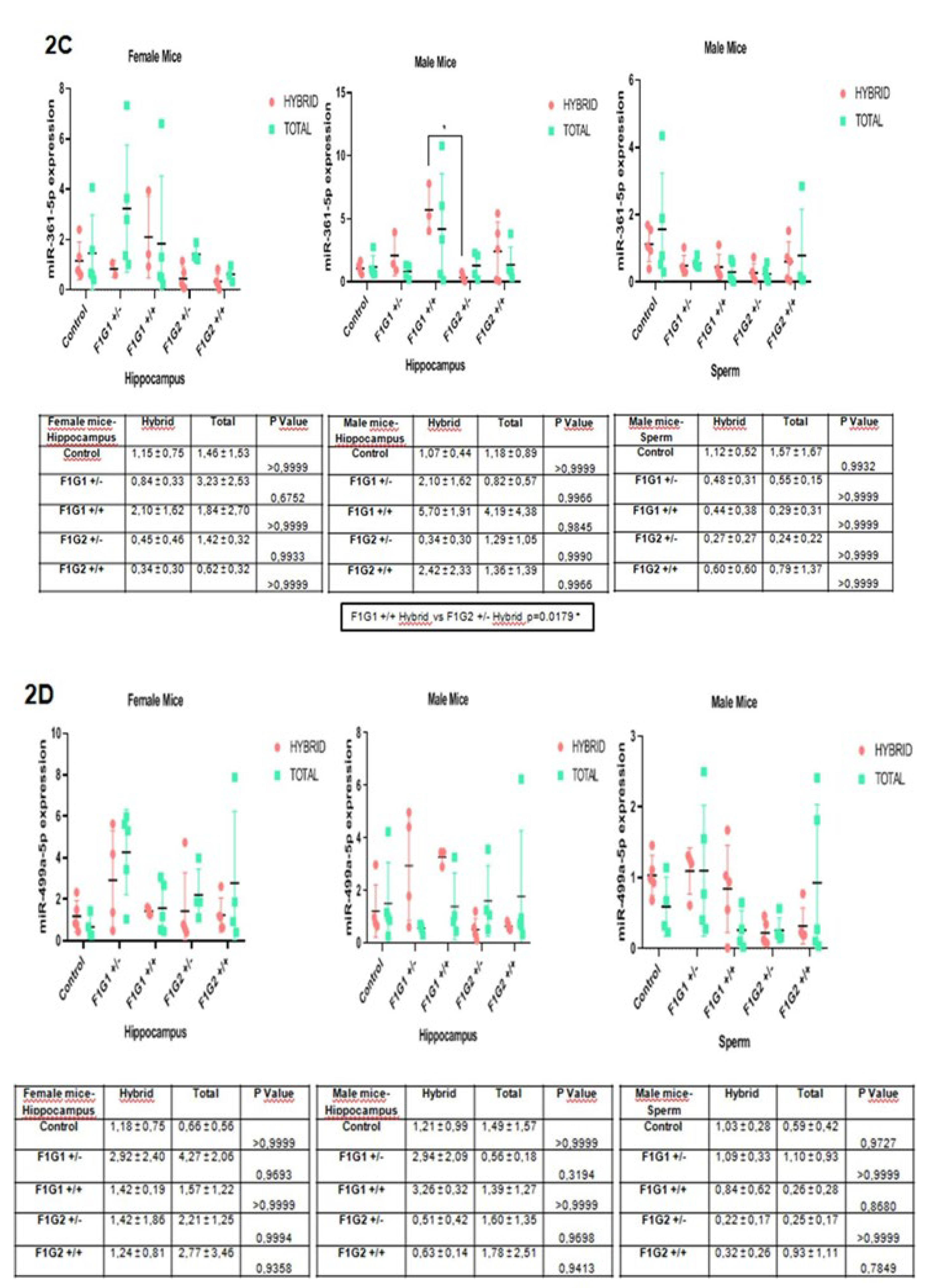
MiR-361-5p expression profiles are shown in Figure 2C. There was a decrease in F1G1 +/+ in the female hippocampus, an increase in all other groups overall, and this difference was not significant compared to the hybrid (Figure 2C). In sperm, it increased in F1G1 +/- and F1G2 +/+, (Figure 2C).
MiR-499-5p expression profiles are shown in Figure 2D. There is an increase in miRNA levels in the female hippocampus in all groups, and this difference is not significant compared to the RNA fraction of in the hybrid. There was a decrease in G1 and an increase in G2 in the male hippocampus, different from that in females. In sperm, there was a decrease in F1G1 +/- and an increase in G2 which was not significant (Figure 2D).
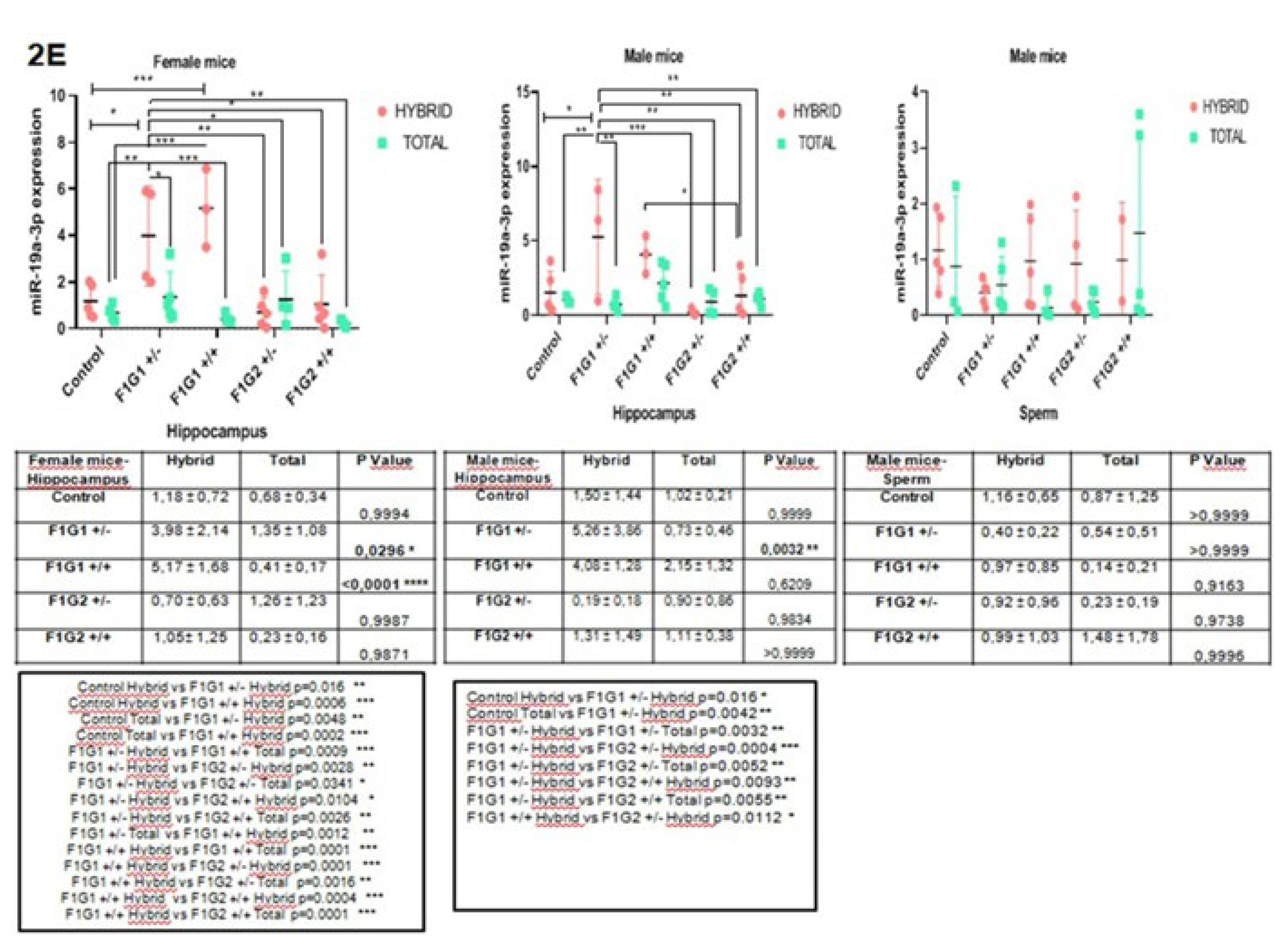
The expression profiles of miR-19a-3p are shown in Figure 2E. In the female hippocampus, there is an increase in the level of miR-19a-3p transcripts in F1G1+/- and a decrease in all other groups. The decreases observed in the F1G1+/- and F1G1+/+ groups compared to the hybrid RNA fractions and the free total fraction, this result was significant (p=0.03, p<0.0001). There is an increase in F1G2+/- animals in the male hippocampus, a decrease in the fraction of total (free) RNA in all other groups, and this difference was not significant compared to the hybrid, with the exception of F1G1+/- (p=0.0032). In sperm, it increased in F1G1+/- and F1G2+/+, and in other groups there was a decrease (Figure 2E).
4. Discussion
Neuronal development encompasses different stages from the embryo throughout neuronal life, primarily including the establishment and maintenance of transcription levels in mature adult brain cells [27]. However, the molecular mechanisms of gene expression adjustments are still largely unknown, particularly those subtle changes related to behavioral biology. In Cc2d1a heterozygous mice, we track Mendelian and non-Mendelian inheritance of autism both of which induce variations in miRNAs levels. Here we hypothesis of its molecular signature in memory control of gene expression by testing hybrid formation between miRNA and DNA. These results are based on monitoring the behavior and expression level of five autism-related miRNAs in heterozygous and wild-type offspring. The hybrid RNA fractions attached to the DNA (R-loop) are compared. The quantity of these miRNAs varies by sex in the hippocampus compared to controls. In addition, in sperm their levels are reduced compared to the hippocampus and controls. These results constitute the first direct demonstration of (a) the differential localization of miRNA DNA/RNA hybrids in mouse brain cell models, (b) its involvement in mouse behavior, and (c) sex differences along lineages.
Three key observations support these findings:
First, alterations in six miRNAs have previously been reported to be indicative of behavioral changes in autism [9]. Additionally, microinjection of miRNAs into fertilized eggs alters their levels in the generated mice. Each of these miRNAs affects part of the complex phenotypes associated with autism [28,29]. In our current experiments, we show the differential amount of the same miRNAs in the total (free) RNA fraction and/or the DNA/RNA hybrids fraction in heterozygous Cc2d1a model neuronal and germ cells.
We suggest that miRNA engaged in DNA/RNA hybrids may regulate their own transcriptional profiles in cells including neural cells. The formation of DNA/RNA hybrids with miRNA indicates the possibility of self-controlled expression. Alteration of DNA secondary structures causes a higher proportion of damage during cell divisions, leading to an expansion of cellular damage [30]. It was also revealed that inappropriate DNA/RNA hybrids could affect gene expression [31]. Increased levels of miRNA engaged in DNA/RNA hybrids may have the ability to alter miRNA expression levels and, at the same time, subsequently significantly affect the levels of target transcripts.
Second, the results indicate that miRNA expression is affected in tissues compared to controls and is more pronounced in a sex-dependent manner in genetically modified Cc2d1a heterozygous mice. The prevalence of boys among ASD patients are striking. None of these miRNAs are located on the X chromosome. Other genetic protections could therefore be responsible for the rarity of autism in girls. However, our results here show variation in behavior and miRNAs quantity in both sexes compared to controls, but still with differences between males and females in the mouse model. Our previous studies showed that the levels of six miRNAs, expressed in blood, hippocampus, testes, and sperm differed in Cc2d1a heterozygous tissues [9]. In this regard, we reported that sustained alteration of six miRNAs in Cc2d1a heterozygous cells resulted in subsequent behavioral impairment and sex-specific partial autophagy [9,12].
Third, previous studies have shown that miRNAs are highly expressed in the CNS during embryonic development and in the peripheral nervous system [32]. Here we show that miRNA expression is highly variable in regions where adult neurogenesis (hippocampus) continually occurs. We also demonstrate the presence of miRNAs in the purified DNA/RNA hybrid fractions. These results strongly suggest the possibility of a role for miRNA in the structure of DNA/RNA hybrids in the physiological functions of the CNS. Recently, the involvement of DNA/RNA hybrids in trinucleotide repeat neurologic diseases and the establishment of repressive histone modifications has been clearly reported [33-35]. However, direct evidence between R-loops and changes in miRNA expression required further study. Targeted deletions in vitro and in vivo are necessary to establish the details of the involvement of DNA/RNA hybrids.
These three key observations support the conclusion that miRNA is not only a post-transcriptional regulator, but also a key regulator of its own expression. Although we have not established the effects of miRNAs concentration on cellular memory, it seems interesting that a single short sequence can regulate both pre- and post-expression regulation [28]. A precedent for this phenomenon is observed in the case of TERRAs (Telomeric Repeat-containing RNA), present in the form of free RNA molecules in the nucleus but also in the form of DNA/RNA hybrids at the end of each chromosome. TERRA promotes telomere elongation, by associating with DNA, but also promotes telomerase activity [31,36]. In light of the many recent reports relating to the application of R-loops to disorders, it will be essential to understand in detail the roles of different miRNAs sequences in their own expression. In this regard, we propose that miRNA engaged in R-loop sensing provide a novel strategy to detect variations in gene expression for neurological changes. The expression levels of miRNAs are essential for tissue-specific identity. We know that miRNAs participate in the transcriptional regulation of several genes, including neuronal differentiation genes [37]. Impairment due to miRNAs tuning, triggered by binding of target RNAs to the enhancer region, is suggested to cause differentiation or division of precursor cells. For example, the concerted action of miRNAs and their localization would be important for inducing decisions regarding neural fate. miRNAs are involved in morphogenesis, patterning, and their proliferation-promoting roles are essential for cell maintenance and expansion of progenitor pools.
Author Contributions
Conceptualization: MR; Methodology: EFS, ST, HD, RT; Validation: EFS, HD; Formal analysis and Data Curation: HD, RT, EFS; Funding Acquisition: EFS; Writing—original draft: MR, ST, HD and EFS; Writing—review and editing: MR and EFS. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by the Erciyes University Research Fund (project number: TSA-2021-10499).
Institutional Review Board Statement
Animal use, care, and experimental procedures in this study were approved by Erciyes University Animal Ethics Committee (09.09.2020, 20/124).
Informed Consent Statement
Not applicable.
Data Availability Statement
The original contributions presented in the study are included in the article, further inquiries can be directed to the corresponding authors.
Acknowledgments
We thank to the Transgenic Unit of GENKOK.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Wormwood, K.L.; Charette, L.; Ryan, J.P.; Darie, C.C.; Woods, A.G. A Proteomics Investigation of Salivary Profiles as Potential Biomarkers for Autism Spectrum Disorder (ASD). Protein J 2023, 42(5), 607–620. [Google Scholar] [CrossRef] [PubMed]
- Dawson, G.; Rieder, A.D.; Johnson, MH. Prediction of autism in infants: progress and challenges. Lancet Neurol 2023, 22(3), 244–254. [Google Scholar] [CrossRef] [PubMed]
- Karaca, M.; Tahtasakal, R.; Dana, H.; Sahin, M.; Pirencioglu, S.N.; Tughan, E.; Dal, F.; Demirci, E.; Sener, E.F. Decreased levels of alpha synuclein in families with autism spectrum disorder and relationship between the disease severity. Brain Res 2023, 1814, 148410. [Google Scholar] [CrossRef] [PubMed]
- Sener, E.F.; Taheri, S.; Sahin, M.C.; Bayramov, K.K.; Marasli, M.K.; Zararsiz, G.; Mehmetbeyoglu, E.; Oztop, D.B.; Canpolat, M.; Canatan, H.; Ozkul, Y. Altered Global mRNA Expressions of Pain and Aggression Related Genes in the Blood of Children with Autism Spectrum Disorders. J Mol Neurosci 2019, 67(1), 89–96. [Google Scholar] [CrossRef] [PubMed]
- Ghafouri-Fard, S.; Pourtavakoli, A.; Hussen, B.M.; Taheri, M.; Ayatollahi, S.A. A Review on the Role of Genetic Mutations in the Autism Spectrum Disorder. Mol Neurobiol 2023, 60(9), 5256–5272. [Google Scholar] [CrossRef] [PubMed]
- Moseley, R.L.; Hitchiner, R.; Kirkby, J.A. Self-reported sex diferences in high-functioning adults with autism: A meta-analysis. Molecular Autism 2018, 9, 33. [Google Scholar] [CrossRef] [PubMed]
- Apte, M.; Kumar, A. Correlation of mutated gene and signalling pathways in ASD. IBRO Neurosci Rep 2023, 14, 384–392. [Google Scholar] [CrossRef] [PubMed]
- Sener, E.F.; Canatan, H.; Ozkul, Y. Recent Advances in Autism Spectrum Disorders: Applications of Whole Exome Sequencing Technology. Psyc Invest 2016, 13(3), 255–64. [Google Scholar] [CrossRef] [PubMed]
- Ozkul, Y.; Taheri, S.; Bayram, K.K.; Sener, E.F.; Mehmetbeyoğlu, E.; Oztop, D.B.; et al. A heritable profile of six miRNAs in autistic patients and mouse models. Scientific Reports 2020, 10(1), 9011. [Google Scholar] [CrossRef] [PubMed]
- Dana, H.; Bayram, K.K.; Delibaşı, N.; Tahtasakal, R.; Bayram, R.; Hamurcu, Z.; Sener, E.F. Disregulation of Autophagy in the Transgenerational Cc2d1a Mouse Model of Autism. Neuromolecular Med 2020, 22(2), 239–249. [Google Scholar] [CrossRef] [PubMed]
- Zamarbide, M.; Mossa, A.; Muñoz-Llancao, P.; Wilkinson, M.K.; Pond, H.L.; Oaks, A.W.; Manzini, M.C. Male-Specific cAMP Signaling in the Hippocampus Controls Spatial Memory Deficits in a Mouse Model of Autism and Intellectual Disability. Biol Psychiatry 2019, 85(9), 760–768. [Google Scholar] [CrossRef] [PubMed]
- Sener, E.F.; Dana, H.; Tahtasakal, R.; Hamurcu, Z.; Taheri, S.; Delibasi, N.; Mehmetbeyoglu, E.; Sukranli, Z.Y.; Dal, F.; Tufan, E.; Oflamaz, A.O.; Doganyigit, Z.; Ozkul, Y.; Rassoulzadegan, M. Heterozygous Cc2d1a mice show sex-dependent changes in the Beclin-1/p62 ratio with impaired prefrontal cortex and hippocampal autophagy. Prog Neuropsychopharmacol Biol Psychiatry 2023, 125, 110764. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, L.B.; Li, B.; Zhao, Q.; Wei, W.; Leighton, L.J.; Bredy, T.W.; Pang, T.Y.; Hannan, A.J. Chronically high stress hormone levels dysregulate sperm long noncoding RNAs and their embryonic microinjection alters development and affective behaviours. Mol Psychiatry 2024, 29(3), 590–601. [Google Scholar] [CrossRef] [PubMed]
- Leir, S.H.; Paranjapye, A.; Harris, A. Functional genomics of the human epididymis: Further characterization of efferent ducts and model systems by single-cell RNA sequencing analysis. Andrology 2024, 12(5), 991–1000. [Google Scholar] [CrossRef] [PubMed]
- Sendler, E.; Johnson, G.D.; Mao, S.; Goodrich, R.J.; Diamond, M.P. Stability, delivery and functions of human sperm RNAs at fertilization. Nucleic Acids Res 2013, 41, 4104–4117. [Google Scholar] [CrossRef] [PubMed]
- Rassoulzadegan, M.; Sharifi-Zarchi, A.; Kianmehr, L. DNA-RNA Hybrid (R-Loop): From a Unified Picture of the Mammalian Telomere to the Genome-Wide Profile. Cells 2021, 10(6), 1556. [Google Scholar] [CrossRef]
- Kianmehr, L.; Khazali, H.; Rajabi-Maham, H.; Sharifi-Zarchi, A.; Cuzin, F.; Rassoulzadegan, M. Genome-Wide Distribution of Nascent Transcripts in Sperm DNA, Products of a Late Wave of General Transcription. Cells 2019, 8(10), 1196. [Google Scholar] [CrossRef] [PubMed]
- Satir-Basaran, G.; Kianmehr, L.; Mehmetbeyoglu, E.; Korkmaz Bayram, K.; Memis, M.; Yilmaz, Z.; Tufan, E.; Taheri, S.; Kelestimur, F.; Rassoulzadegan, M. Mouse Paternal RNAs Initiate a Pattern of Metabolic Disorders in a Line-Dependent Manner. Front Genet 2022, 13, 839841. [Google Scholar] [CrossRef] [PubMed]
- Al-Hadid, Q.; Yang, Y. R-loop: an emerging regulator of chromatin dynamics. Acta Biochim Biophys Sin (Shanghai) 2016, 48(7), 623–31. [Google Scholar] [CrossRef] [PubMed]
- Scheuren, M.; Möhner, J; Zischler, H. R-loop landscape in mature human sperm: Regulatory and evolutionary implications. Front Genet 2023, 14, 1069871. [Google Scholar] [CrossRef] [PubMed]
- Patankar, A.; Gajbhiye, R.; Surve, S.; Parte, P. Epigenetic landscape of testis specific histone H2B variant and its influence on sperm function. Clin Epigenetics 2021, 13(1), 101. [Google Scholar] [CrossRef] [PubMed]
- Narayanan, R.; Schratt, G. miRNA regulation of social and anxiety-related behaviour. Cell Mol Life Sci 2020, 77(21), 4347–4364. [Google Scholar] [CrossRef] [PubMed]
- Stott, J.; Wright, T.; Holmes, J.; Wilson, J.; Griffiths-Jones, S.; Foster, D.; Wright, B. A systematic review of non-coding RNA genes with differential expression profiles associated with autism spectrum disorders. PLoS One 2023, 18(6), e0287131. [Google Scholar] [CrossRef] [PubMed]
- Vishnoi, A.; Rani, S. MiRNA Biogenesis and Regulation of Diseases: An Overview. Methods Mol Biol 2017, 1509, 1–10. [Google Scholar] [PubMed]
- Schepici, G.; Cavalli, E.; Bramanti, P.; Mazzon, E. Autism Spectrum Disorder and miRNA: An Overview of Experimental Models. Brain Sci 2019, 9(10), 265. [Google Scholar] [CrossRef]
- Kondaurova, E.M.; Belokopytova, I.I.; Kulikova, E.A.; Khotskin, N.V.; Ilchibaeva, T.V.; Tsybko, A.S.; Popova, N.K.; Naumenko, V.S. On the role of serotonin 5-HT1A receptor in autistic-like behavior: сross talk of 5-HT and BDNF systems. Behav Brain Res 2023, 438, 114168. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Nardelli, J. Cellular and molecular introduction to brain development. Neurobiol Dis 2016, 92(Pt A), 3–17. [Google Scholar] [CrossRef]
- Yilmaz Sukranli, Z.; Korkmaz Bayram, K.; Mehmetbeyoglu, E.; Doganyigit, Z.; Beyaz, F.; Sener, E.F.; Taheri, S.; Ozkul, Y.; Rassoulzadegan, M. Trans Species RNA Activity: Sperm RNA of the Father of an Autistic Child Programs Glial Cells and Behavioral Disorders in Mice. Biomolecules 2024, 14(2), 201. [Google Scholar] [CrossRef] [PubMed]
- Tufan, E.; Taheri, S.; Karaca, Z.; Mehmetbeyoglu, E.; Yilmaz Sukranli, Z.; Korkmaz Bayram, K.; Ulutabanca, H.; Tanrıverdi, F.; Unluhizarci, K.; Rassoulzadegan, M.; Kelestimur, F. Alterations in Serum miR-126-3p Levels over Time: A Marker of Pituitary Insufficiency following Head Trauma. Neuroendocrinology 2024, 114(4), 315–330. [Google Scholar] [CrossRef] [PubMed]
- García-Muse, T.; Aguilera, A. R Loops: From Physiological to Pathological Roles. Cell 2019, 179(3), 604–618. [Google Scholar] [CrossRef] [PubMed]
- Petermann, E.; Lan, L.; Zou, L. Sources, resolution and physiological relevance of R-loops and RNA-DNA hybrids. Nat Rev Mol Cell Biol 2022, 23(8), 521–540. [Google Scholar] [CrossRef] [PubMed]
- Cao, D.D.; Li, L.; Chan, W.Y. MicroRNAs: Key Regulators in the Central Nervous System and Their Implication in Neurological Diseases. Int J Mol Sci 2016, 17(6), 842. [Google Scholar] [CrossRef] [PubMed]
- Colak, D.; Zaninovic, N.; Cohen, M.S.; Rosenwaks, Z.; Yang, W.Y.; Gerhardt, J.; Disney, M.D.; Jaffrey, S.R. Promoter-bound trinucleotide repeat mRNA drives epigenetic silencing in fragile X syndrome. Science 2014, 343(6174), 1002–5. [Google Scholar] [CrossRef] [PubMed]
- Groh, M.; Gromak, N. Out of balance: R-loops in human disease. PLoS Genet 2014, 10(9), e1004630. [Google Scholar] [CrossRef] [PubMed]
- Skourti-Stathaki, K.; Kamieniarz-Gdula, K.; Proudfoot, N.J. R-loops induce repressive chromatin marks over mammalian gene terminators. Nature 2014, 516(7531), 436–9. [Google Scholar] [CrossRef] [PubMed]
- Azzalin, C.M. TERRA and the alternative lengthening of telomeres: a dangerous affair. FEBS Lett 2024, Mar 6. [CrossRef]
- Suster, I., Feng, Y. Multifaceted Regulation of MicroRNA Biogenesis: Essential Roles and Functional Integration in Neuronal and Glial Development. Int J Mol Sci 2021, 22(13), 6765.
- Komatsu, S.; Kitai, H.; Suzuki, H.I. Network Regulation of microRNA Biogenesis and Target Interaction. Cells 2023, 12(2), 306. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Tail suspension test for parent and F1 generation. A. Test results for parents (n=10, each sex). B. Test results for F1 generation (n=10, each sex). C. Marble burying test for parents (n=10, each sex). D. Test results for F1 generation (n=10, each sex). E. Social interaction test for parents and F1 generation (n=10, each sex). F. Novel object recognition test for parents (n=10, each sex). Distance was measured as cm. G. Test results for F1 generation (n=10, each sex). H. Novel object recognition test for female mice (n=10, each sex). Distance was measured as cm. I. Novel object recognition test for male mice (n=10, each sex). *p<0.05, **p<0.01, ***p<0.001, **** p<0.0001.
Figure 1.
Tail suspension test for parent and F1 generation. A. Test results for parents (n=10, each sex). B. Test results for F1 generation (n=10, each sex). C. Marble burying test for parents (n=10, each sex). D. Test results for F1 generation (n=10, each sex). E. Social interaction test for parents and F1 generation (n=10, each sex). F. Novel object recognition test for parents (n=10, each sex). Distance was measured as cm. G. Test results for F1 generation (n=10, each sex). H. Novel object recognition test for female mice (n=10, each sex). Distance was measured as cm. I. Novel object recognition test for male mice (n=10, each sex). *p<0.05, **p<0.01, ***p<0.001, **** p<0.0001.
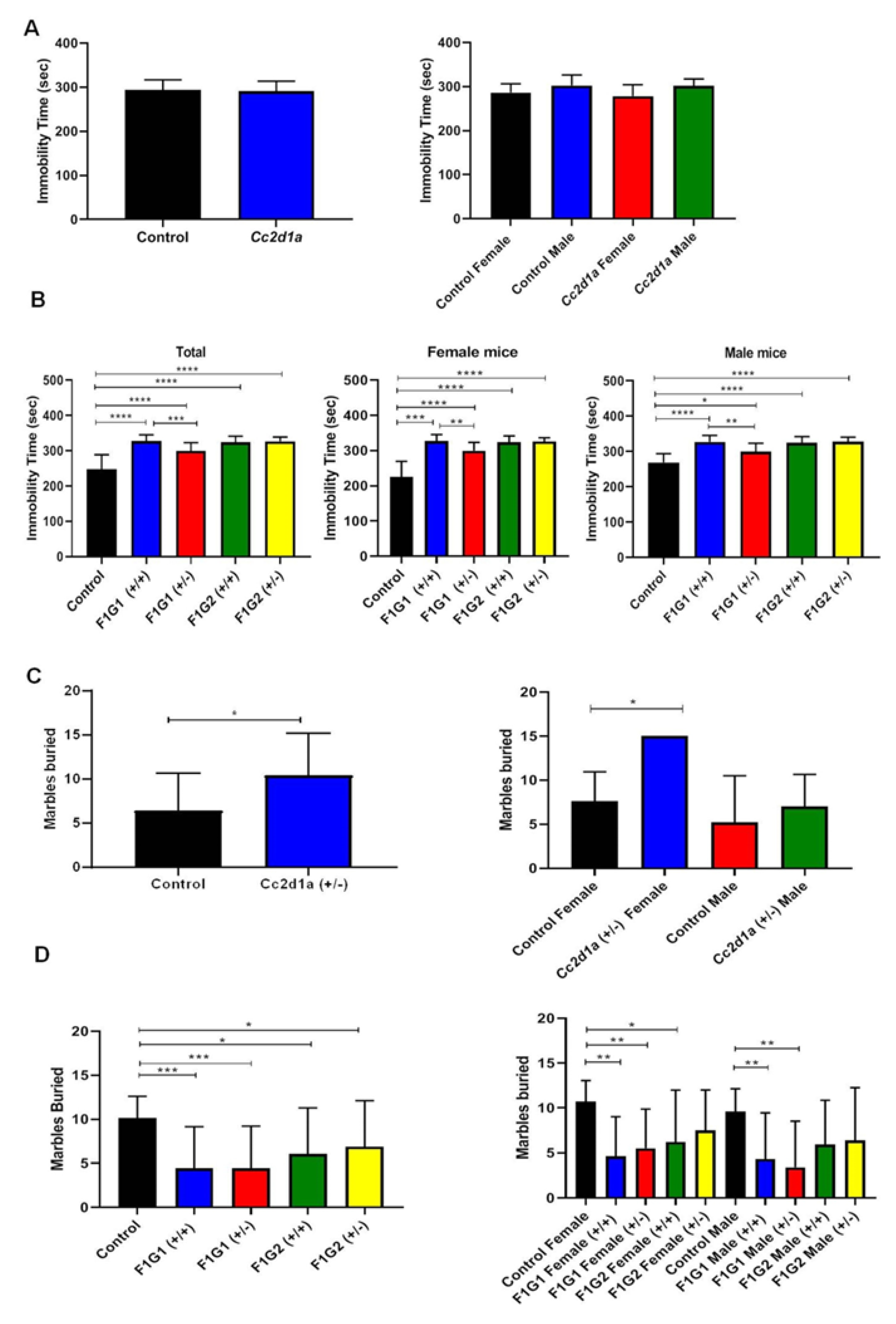
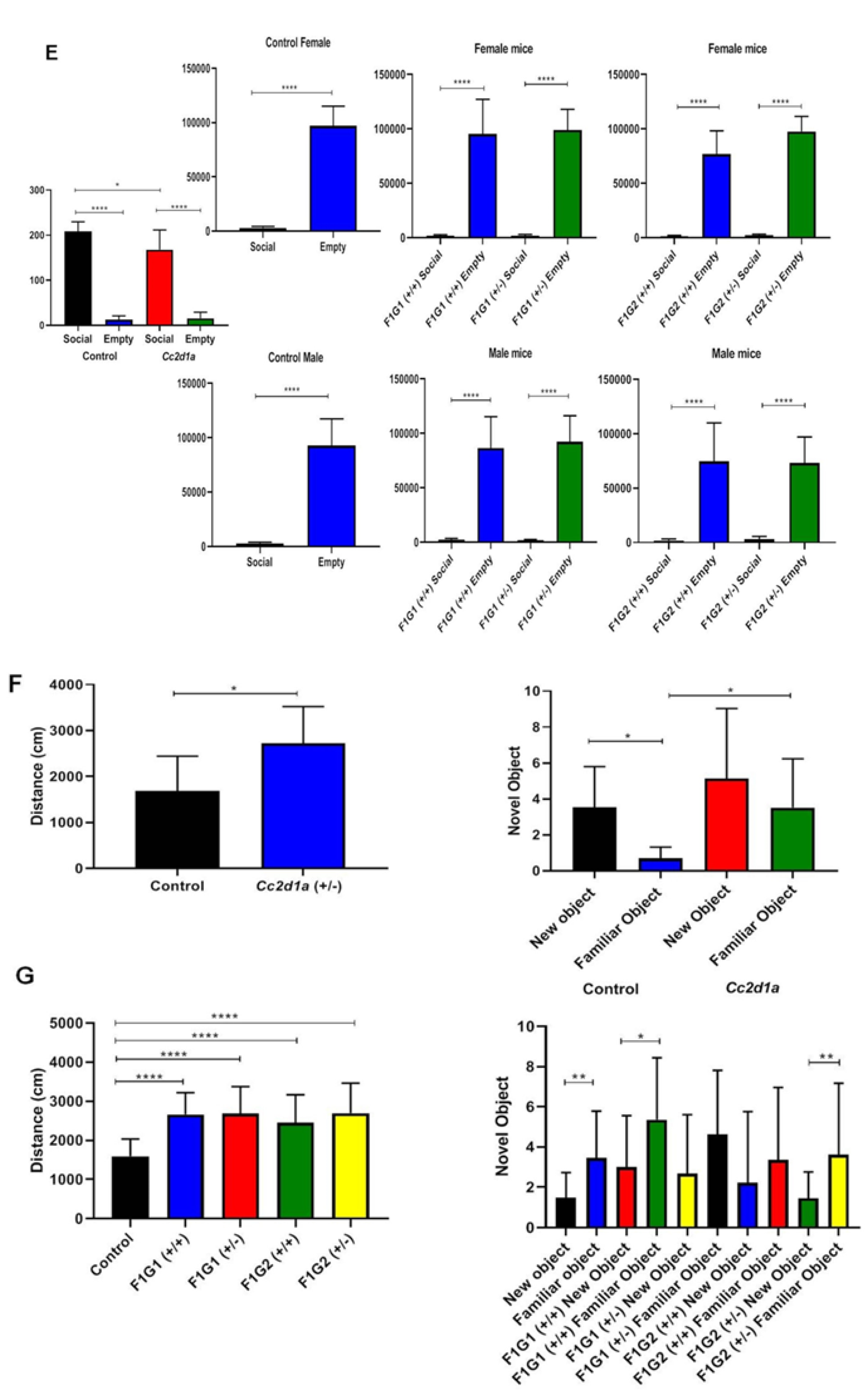
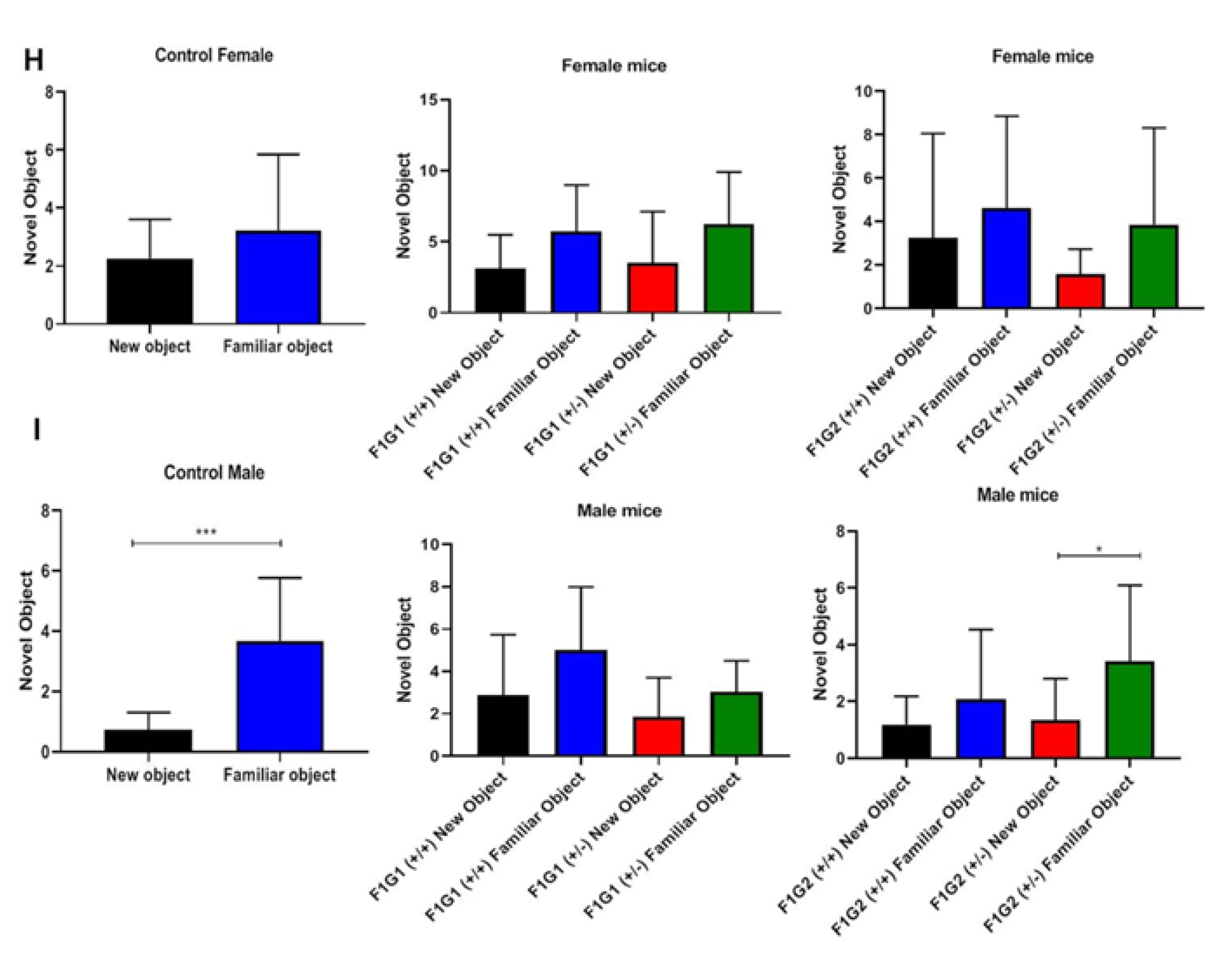
Figure 2.
A. miR-126-3p expression in hybrid and total groups. Graphs were shown for female hippocampus, male hippocampus and male sperm. Data were expressed in mean±standart deviation (n=5, each group). 2B. miR-150-5p expression in hybrid and total groups. Graphs were shown for female hippocampus, male hippocampus and male sperm. Data were expressed in mean±standart deviation (n=5, each group). 2C. miR-361-5p expression in hybrid and total groups. Graphs were shown for female hippocampus, male hippocampus and male sperm. Data were expressed in mean±standart deviation (n=5, each group). 2D. miR-499a-5p exppression in hybrid and total groups. Graphs were shown for female hippocampus, male hippocampus and male sperm. Data were expressed in mean±standart deviation (n=5, each group). 2E. miR-19a-3p expression in hybrid and total groups. Graphs were shown for female hippocampus, male hippocampus and male sperm. Data were expressed in mean±standart deviation (n=5, each group). * p<0.05, **p<0.01, ***p<0.001, ****p<0.0001.
Figure 2.
A. miR-126-3p expression in hybrid and total groups. Graphs were shown for female hippocampus, male hippocampus and male sperm. Data were expressed in mean±standart deviation (n=5, each group). 2B. miR-150-5p expression in hybrid and total groups. Graphs were shown for female hippocampus, male hippocampus and male sperm. Data were expressed in mean±standart deviation (n=5, each group). 2C. miR-361-5p expression in hybrid and total groups. Graphs were shown for female hippocampus, male hippocampus and male sperm. Data were expressed in mean±standart deviation (n=5, each group). 2D. miR-499a-5p exppression in hybrid and total groups. Graphs were shown for female hippocampus, male hippocampus and male sperm. Data were expressed in mean±standart deviation (n=5, each group). 2E. miR-19a-3p expression in hybrid and total groups. Graphs were shown for female hippocampus, male hippocampus and male sperm. Data were expressed in mean±standart deviation (n=5, each group). * p<0.05, **p<0.01, ***p<0.001, ****p<0.0001.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



