
Submitted:
10 July 2024
Posted:
11 July 2024
You are already at the latest version
Abstract
Vitronectin is a glycoprotein, present in plasma and extracellular matrix, implicated in cell migration. The high amount of vitronectin found in neuroblastoma biopsies was associated with poor prognosis. However, there are almost no studies of vitronectin levels in neuroblastoma cell culture media nor in plasma of neuroblastoma patients. Vitronectin concentration in culture media of four neuroblastoma cell lines grown in monolayer and in 3D-models, and in plasma of 114 neuroblastoma patients, was quantified using ELISA. Three of the neuroblastoma cell lines secreted vitronectin to culture media when cultured in monolayer and 3D-models. Vitronectin release was higher by neuroblastoma cells cultured in 3D-models than in monolayer, and was still great when cells grown in 3D-scaffolds with cross-linked vitronectin. Vitronectin secretion occurred independently of cell numbers in cultures. Its concentration in plasma of neuroblastoma patients ranged between 52.4 and 870µg/mL (median, 218µg/mL). A ROC curve was used to establish a cutoff of 361µg/mL, above which patients over 18-months-old had worse prognosis (p=0.0018). Vitronectin could be considered a new plasma prognostic biomarker in neuroblastoma, and warrants confirmation in collaborative studies. Drugs inhibiting vitronectin interaction with cells and/or extracellular matrix could represent a significant improvement in survival for neuroblastoma patients.
Keywords:
Pediatric cancer
; liquid biopsy
; precision medicine
; hydrogels
; extracellular matrix
1. Introduction
Vitronectin (VN), also known as serum spreading factor, is a multifunctional glycoprotein present in plasma and extracellular matrix (ECM) [1]. It contains multiple cell receptor binding sites including integrins, urokinase-type plasminogen activator receptor (uPAR) and plasminogen activator inhibitor-1 (PAI-1) [2]. VN seems to anchor to ECM fibers and proteoglycans, leading to transitory ECM element–cell junctions, cell adhesion and migration [3,4,5]. Besides its role in spreading and metastasis, VN is involved in other key steps of cancer such as apoptosis, inflammation, vascular permeability and vascular endothelial growth factor-induced angiogenesis [6,7,8]. Its association with tumor aggressiveness has been described in several cancers [6,9,10,11,12,13,14,15], including neuroblastoma (NB) [5].
NB originates from the neural crest in the sympathetic nervous system and is one of the most common pediatric solid tumors [16]. The risk of progression in NB patients is defined by several clinical, biological and genetic features [17], and the current survival rate of high risk (HR) NB is under 50% [18]. In previous studies, we detected an elevated amount of VN in cytoplasm of malignant neuroblasts and adjacent to them applying immunohistochemistry (IHC) on NB biopsies, which was related to poor outcome of HR-NB [5]. Moreover, we observed that this VN secreted to ECM form tracks, postulating that VN could participate in tumor cell migration [19]. In subsequent studies, our experimental models (orthotopic xenograft VN knock-out mice, and two 3D models: hydrogels (HGs) of methacrylated gelatin plus increasing concentrations of methacrylated alginate, and HGs of polyethylene glycol with and without cross-linked VN) supported the role of this glycoprotein in NB cell dynamics, aggressiveness, and in the clonal selection of segmental chromosomal aberrations (SCAs) [20,21,22,23].
Besides the role of VN in tumor ECM, its presence in blood plasma and in other types of liquid biopsies (ascites and cerebrospinal fluid) has also been studied in several tumor types. Specifically, elevated concentrations of circulating VN have been observed in adult patients with glioma, melanoma, breast, ovarian, endometrium cancer, and also in pediatric patients with Hodgkin lymphoma and acute lymphoblastic leukemia [10,12,13,14,24,25,26,27,28,29,30] VN in plasma is particularly understudied in pediatric solid tumors, including NB [31]. These tumors represent a great clinical challenge due to their aggressiveness and high mortality, and in many cases, to the difficulty of obtaining appropriate biopsies, given the young age of some patients and the high frequency of intratumor heterogeneity [32].
In this study, we detected VN secretion to culture media of four aggressive NB cell lines grown in monolayer and in HGs composed of gelatin-tyramine (GTA) and silk fibroin (sf) with and without cross-linked VN. In view of the results obtained in vitro, VN concentration was determined in 114 plasma samples from NB patients with different clinic and molecular tumor features.
2. Results
2.1. Higher VN Release in 3D Models
We studied the VN levels in culture media of four NB cell lines (Figure 1A) grown in 2D until reaching confluence and in HGs for two and three weeks. VN release in control HGs (no cells) with cross-linked VN was null at both culture times. However, all but one cell line secreted VN to culture media when grown in 2D and in 3D models. SK-N-BE(2) and SH-SY5Y cell lines secreted VN in all growth conditions. PDX2 [33] showed VN secretion when cultured in 2D and in HGs after three weeks of culture. We were unable to detect VN in culture media of PDX1 [33] in any of the culture conditions. Aside from the abovementioned negative cases, VN concentration was significantly higher in media of the 3D models (median = 11.1 ng/mL/million cells) than in monolayer (median = 1.5 ng/mL/million cells) cultures (p = 0.0052, Figure 1B).
2.2. VN Secretion Independent of Cell Numbers
Pearson r correlation showed that VN concentration secretion to culture media per million of cells and the estimated number of cells in HGs by digital analysis were independent variants. SK-N-BE(2) presented a marked reduction in VN secretion to the culture media after three weeks of culture (mean = 19.9 ng/mL/million cells) compared to two weeks (mean = 10.1 ng/mL/million cells), despite the greater number of cells estimated in the HGs with longer time. The opposite was observed for the SH-SY5Y (mean = 14.2 ng/mL at 3 weeks vs. 7 ng/mL at 2 weeks) and PDX2 (mean = 12.1 ng/mL at 3 weeks vs. 0 ng/mL at 2 weeks) cell lines, which presented an increase in VN secretion to media after three weeks of culture, not related to greater cell growth (Figure 1C). SK-N-BE(2) and SH-SY5Y cell lines showed a larger secretion of VN per million cells when cultured in HGs without added VN (mean = 16.3 ng/mL and 12.1 ng/mL, respectively) compared to HGs with added VN (mean = 13.5 ng/mL and 7.8 ng/mL, respectively) at both two and three weeks of culture, despite the increasing trend in cell growth observed in the HGs with added VN, which was more notable in the HGs with three weeks of culture(Figure 1C).
2.3. Clinical and Genetic Characteristics of NB Patients Included in the Study
Having detected VN in the culture media of the studied aggressive NB cell lines, we next sought to analyze whether the same would occur in the plasma of NB patients. A total of 114 NB patients aged between 0–296 months at time of diagnosis (median = 16) were included in this study, of whom 52 were older than 18 months and 54 showed metastasis at diagnosis. Homogeneous MYCN amplification was detected in 20 and heterogeneous amplification in four NB. Typical SCAs (typSCAs) of NB were detected in 60 cases, two tumors presented atypical SCAs (atypSCAs) and 33 NB showed numeric chromosomal aberrations (NCAs). After 5–204 months of follow up (median = 74), 81 patients remained alive (Figure 2).
2.4. Discriminatory Power of Plasma VN Levels for NB Patient Outcomes
VN levels in the plasma of 33 NB patients who died were significantly higher than those obtained in 81 patients who survived (median = 308 µg/mL vs. 205 µg/mL respectively, p = 0.0009, Figure S1). In line with the prognosis power of age in NB patients, however, this factor showed a pronounced association with death; only four of the 62 patients younger than 18 months died after the abovementioned follow-up, without significant differences in their plasma VN concentration (Figure 3A and B). Plasma VN levels in NB patients older than 18 months who died (N = 29) were significantly higher than in surviving ones [N = 23] (median = 324 µg/mL vs. 215 µg/mL respectively, p = 0.0074, Figure 3A and B). As plasma VN levels had outcome discriminatory power in those patients, a ROC curve was represented to identify the optimal cutoff according to survival (Figure 3C). The ROC curve was significant (p = 0.0026) with AUC = 0.75 (95% CI = 0.61 – 0.88) and a cutoff of 361 µg/mL showed the best likelihood ratio (10.31). The mean VN concentration previously described in plasma of healthy controls with similar ages was 367.7 µL/mL [34], so the cutoff is also almost over this value (Figure S2).
Kaplan-Meier curves for overall survival (OS) and event free survival (EFS) reflected the significantly worse outcome of NB patients older than 18 months with VN plasma levels over the cutoff compared with patients with VN levels under these values (p = 0.0018 and p = 0.0027, respectively) [Figure 4A and B]. Five-year OS was 7% for patients with VN levels over the cutoff and 53% for those under, and five-year EFS were 7% and 45%, respectively.
2.5. Plasma VN Level Association with Stage and Genetic Features of NB
Besides the association of plasma VN levels with outcomes in NB patients older than 18 months, we also studied the potential link of VN concentrations with certain clinical and genetic factors (Table 1). We detected an enrichment of patients with metastatic disease at diagnosis and VN levels over the cutoff (p = 0.021), and also of NB with SCAs (p = 0.014), specifically with the typical loss of 11q (-11q, p = 0.006).
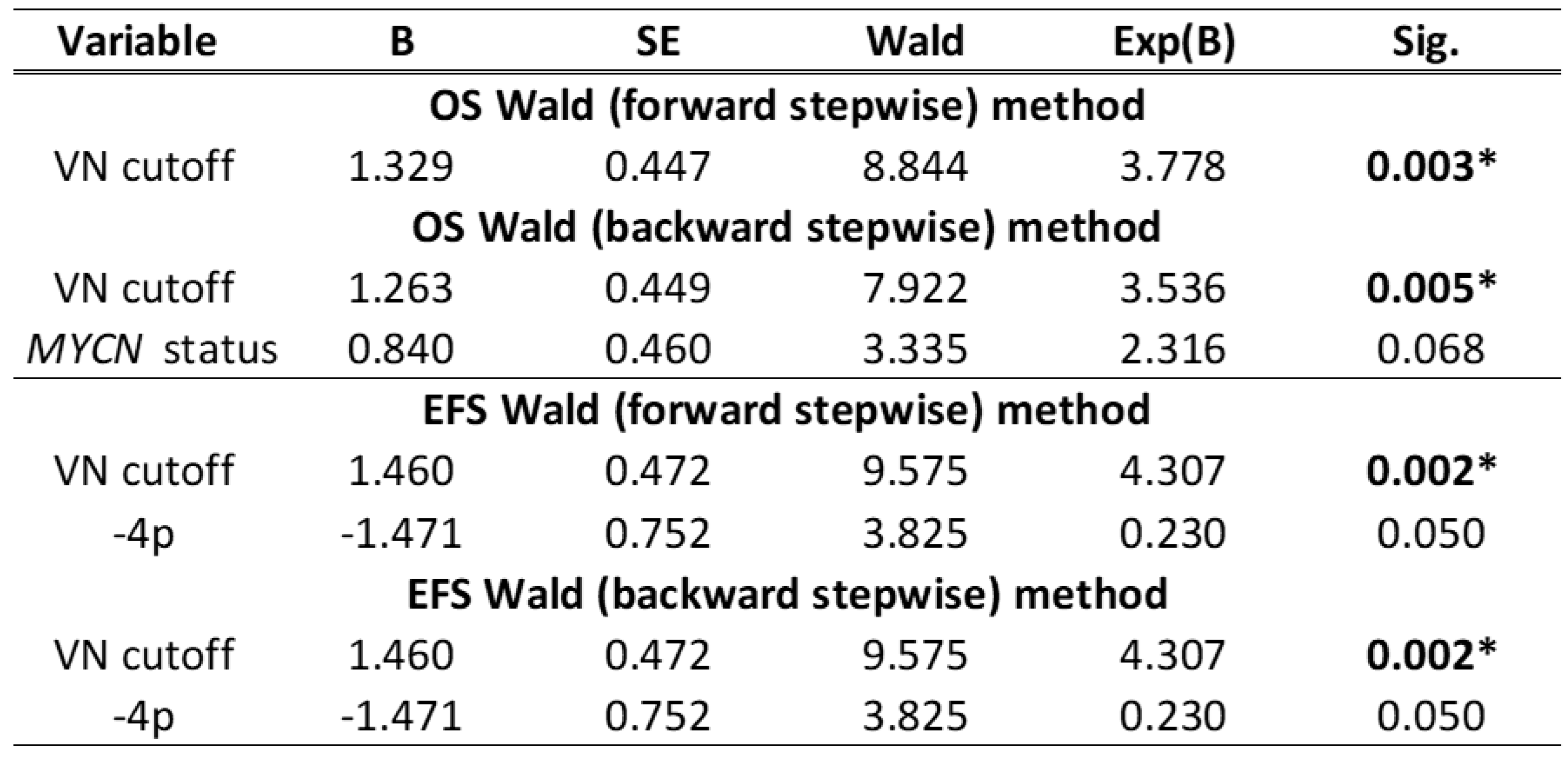
Cox regression using stepwise forward and backward Wald methods proved that the cutoff of VN level of 361 µg/mL had a significant outcome predictive value, independent of other clinical and genetic factors (Table 2).
3. Discussion
The search for new biomarkers that improve and speed up patient diagnosis, better classify them into risk groups and define new therapeutic strategies is one of the great challenges of oncological research in recent years. Circulating biomarkers have the added advantage of being non-invasive compared to biomarkers analyzed on tumor biopsies. Circulating biomarkers currently used in NB diagnosis and prognosis include urine catecholamines, serum lactate dehydrogenase, ferritin and neuron-specific enolase [35]. Circulating tumor cells (e.g. cell markers such as GD2), specific genetic alterations in cell-free DNA (e.g. ALK), and mRNAs (e.g. TH, PHOX2B and DCX), and more recently, hypermethylation (e.g. RASSF1A) [36] and extracellular vesicles [35], also seem of interest for defining prognosis and detecting early relapses and minimal residual disease. Currently, however, these biomarkers are sought not only at the level of malignant cells, but also in the tumor microenvironment, which as well as its important role in tumor progression and metastasis, represents a whole universe of therapeutic targets [37,38]. In previous studies, we proposed VN as a tissue biomarker based on its high expression by IHC in NB biopsies and several experimental models [5,19,20,21,22,23]. In this work we go one step further to study whether the cells in our new 3D models secret VN to culture media and whether secretion can be detected in the plasma of NB patients.
VN release from NB cells to culture media was higher in 3D models than in monolayer cultures. This reflects the importance of using biomimetic models which reflect not only cell–cell interactions, but also cell–scaffold communication. In this sense, despite the we linked high concentrations of VN to the HGs, cells continued to secrete their own VN at considerable levels (although somewhat lower than those of HGs without added VN). A previous study with glioma cells showed an increase in cell migration when cells were cultured with complete human serum, and a reduction of motility when the VN of those serums was depleted [29]. Similar effects were described in other publications with several cancer cells, and the authors postulated that VN present in plasma is a key migration-inducing factor and more potent chemoattractant than already known pro-metastatic chemokines or growth factors [30,39]. Moreover, VN glycosylation patterns can impact its ability to interact with ligands and induce advantageous conformational changes, as has been shown in human hepatocellular carcinoma-derived plasmas in comparison to healthy controls, and in ovarian cancer ascites [40,41]. Curiously, no detect relationship was found between number of cells and amount of VN secretion in our NB models. Hao et al. described high VN levels in serum of breast cancer patients compared with healthy controls, and also pointed to a negative correlation between tumor size and serum VN levels. Their hypothesis was that this could be due to a greater secretion of proteases (such as MMP2) which degrade the VN upon tumor progression [42]. Whether this could apply in our 3D models (and to what extent), or whether this reduction in VN secretion could be due to cellular activity focused on other cellular functions, to differences in synthetized VN retention between cells or in scaffolds, or to reduced viability caused by cell overgrowth, are also interesting questions for future studies.
High VN levels in plasma have been previously described in glioma, melanoma, hepatic, ovary, endometrium and breast cancer patients associated to poor prognosis [10,12,13,14,26,27,28,29,30]. Children and adolescents with Hodgkin lymphoma who relapsed and pediatric patients of acute lymphoblastic leukemia also showed increased levels in plasma of this glycoprotein [24,25]. Nonetheless, to our best knowledge only one publication has studied VN plasma levels in pediatric patients with solid tumors, without obtaining significant differences with healthy controls. Only eight plasmas from eight children with NB were included in that publication: one from an initial stage patient and seven from children in advanced stages of illness [31]. The small number of plasmas precluded studying the association of VN levels with tumor aggressiveness and prognosis. Our study includes 114 plasmas from NB patients, a not inconsiderable figure taking into account the small number of annual cases of this rare of cancer type. We observed that high plasma VN levels had prognostic value independently from clinical and genetic factors of NB applying Cox regression. These findings substantialize plasma VN as a potential circulating prognostic biomarker for NB in patients aged over 18 months. However, mean plasma VN values obtained in NB patients were lower than those reported in healthy controls [34] of the same age (269.5 µg/mL vs. 367.6 µg/mL, respectively; Supp. Figure 2). As the ELISA kits used for VN detection were different, there may be discrepancies between the values obtained, as has been observed in studies comparing healthy control levels [26,34,43].
High VN expression was observed by our group in ECM of HR-NB biopsies [5] with significant branching in VN patterns, which seemed to form tracks enabling malignant cells to reach blood vessels [19]. VN could intervene in cell adhesion, proteinase secretion and tissue remodeling to achieve a more favorable ECM leading to tumor progression, migration and survival. In this context, VN has been shown to activate phosphorylation of p42/44 MAPK and AKT in cell lines, and VN serum has been correlated with PI3K and AKT levels [13,30]. Moreover, VN is considered to play an important role in the regulation of endothelial permeability and VEGF signaling, promoting angiogenesis and vascular permeability [7]. Once in the bloodstream and in other fluids such as lymph, ascites and cerebrospinal fluid, VN could promote circulating tumor cells to disseminate and adhere to distal organs [42,44]. As VN can be produced in the peritoneal cavity, liver, and bones [39,45,46], these could represent suitable premetastatic niches for forming new NB metastatic tumors.
This study and our previous research with patient biopsies and other experimental models all suggest that VN (expressed by tumor cells and detected in plasma) could be closely linked to tumor aggressiveness and poor prognosis in NB patients, appearing to play a role in different steps of cancer progression. Collaborative studies including more plasmas from patients and healthy controls must be conducted for VN levels to be considered alongside classical prognosis factors in therapeutic decision-making, which would represent a step forward in precision medicine. In combination with classic therapies, drugs such as Cilengitide [47,48,49,50] (which binds to integrins and acts as a competitive inhibitor of their join to VN), and fibrinogen analogs (which seem to quench VN chemoattraction [30,39]) could bring a significant improvement in survival of patients with NB and other cancers.
4. Materials and Methods
4.1. 2D Cell Cultures
SK-N-BE(2), SH-SY5Y NB cell lines were acquired from American Type Culture Collection (ATCC, Masassas, VA, USA). Both cell lines were expanded in IMDM medium (Gibco, Life Technologies, Waltham, MA, USA) supplemented with 10% FBS, 1% insulin/transferrin and 1% Penicillin/streptomycin at 37 °C in 5% CO2 atmosphere. Patient-derived xenograft 1 (PDX1) and PDX2 cells were previously established as described in [33]. Both cell lines were expanded in DMEM - high glucose GlutaMAX™ medium, supplemented with 1% B-27™ without Vitamin A (Gibco, Life Technologies, Waltham, MA, USA), 20 ng/mL of EGF and FGF, and 1% Penicillin/streptomycin. Culture medium was replaced every 2-3 days.
4.2. 3D Hydrogel Construction
3D hydrogels were based on previous work [51]. Briefly, sf (Sigma Aldrich, Merck, USA) was mixed with previously synthetized and lyophilized GTA (Sigma Aldrich, Merck, USA) in unsupplemented IMDM cell culture medium to obtain 4% w/v solutions. The ratio of sf:GTA was 75:25. VN (PrepoTech, USA) resuspended in dPBS was added to a half of the solution, with a final concentration of 0.4 mg/mL. dPBS was added in the remaining solution to construct hydrogels without VN. Per each hydrogel, 1.25x105 of commercial NB cells or 2.5x105 of PDX cells previously cultured in 2D were resuspended in the solution prior to the gelification. To start the polymerization reaction, we added horseradish peroxidase (20 U/mL) and hydrogen peroxide (0.01%). Aliquots of 60 μL per hydrogel were placed in the center of each well in a 24-well plate and were kept at 37ºC for 1 hour. Subsequently, 2 mL of supplemented culture medium was added and replaced and collected for ELISA every three days. We kept the 3D models for two and three weeks.
4.3. Patients and Samples
Blood plasmas from 114 NB patients diagnosed between 2003 and 2023 were included in this study. Peripheral blood samples were collected at the time of primary tumor biopsies. All samples were sent to the Spanish Reference Centre for NB Molecular and Pathological studies (Department of Pathology, University of Valencia-INCLIVA) and stored in our biobank (reference B.0000339 29/01/2015). Histopathological data of the NBs were provided by the reference group pathologist. Clinical data were provided by the attending pediatric oncologist when possible, or by the Reference Centre for NB Clinical Studies. These included outcome data of EFS, defined as length of time from diagnosis to any progression, death or to the date of last contact; and OS, defined as length of time from diagnosis until death or last medical check-up in surviving patients. Genetic data were compiled from our internal database NeuPAT [52]. This study was approved by the Research Ethics Committee of the Clinic Hospital of Valencia (No. 2020/025, Act: 372, 30/09/2021). Participants or their family members/legal guardians provided written informed consent for the research studies performed in our laboratory.
4.4. Plasma VN Determination
Peripheral blood samples of NB patients were collected in sterile EDTA tubes. Samples were centrifuged at 2000g for 10 min to separate plasma. Plasmas were stored at − 80 °C until assayed. VN levels were measured using colorimetric enzyme-linked immunosorbent assay (Novus Biologicals, USA) according to the manufacturer's instructions. To bring VN levels within the detection ranges of the kit, plasmas of patients were diluted between 1:1000 and 1:5000 before assay. For VN level determination in media of 2D cell cultures, 8 mL of medium from confluent T75 cell culture flasks were centrifugated at 1200 rpm for 5 min to remove cellular debris. Next, media were concentrated 16 times with SpeedVac™. The number of cells of 2D cultures were determined by a BioRad TC20 Automated Cell Counter after mixing with trypan blue. For 3D cell cultures, mediums from two HGs (4mL) were collected, centrifugated as before and concentrated eight times with SpeedVac™. After medium collection, HGs were formalin-fixed and paraffine-embedded, and a 3µm section was stained with hematoxylin and eosin. Slides were scanned with Ventana iScan, Roche. The number of cells per section was determined with StarDist extension for QuPath and scaled to the total volume of HGs (28 mm3). The results were multiplied by the dilution or concentration factor, and those from culture media were also scaled to the number of cells (per millions) in the cultures.
4.5. Statistical Analysis
Statistical analysis was done using GraphPad Prism 8 (Graphpad Software, Boston, USA) and SPSS 28.0 (SPSS Inc., Chicago, IL, USA). The Kolmogorov-Smirnov test was carried out to investigate data distribution. Paired and non-paired T-tests and Spearman correlation tests were performed for intergroup comparisons as appropriate. The receiver operating characteristic (ROC) curve was performed to assess discriminatory ability with an estimated area under curve (AUC). A suitable cutoff value was selected from ROC curves to obtain the optimal sensitivity and specificity. Survival curves were made using the Kaplan-Meier method, and Cox regression analysis was employed to assess OS- and EFS-related predictors (Wald forward and backward stepwise methods).
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figure S1: VN levels in plasma of all NB patients; Figure S2: VN levels in plasma of NB patients stratified by age.
Author Contributions
AL-C, IV-M and SG-A carried out the in vitro experiments and the culture media collection. JH-A and RP collected blood samples of some NB patients. AL-C prepared plasma samples. DA-L and NE-C performed the ELISAs. AC provided pediatric clinical data. AL-C wrote the original draft of the manuscript. SN carried out histopathological analyses, reviewed the paper and suggested draft changes. RN elaborated the concept and designed the study, acquired funding, reviewed the paper and suggested draft changes. All authors read and approved the final manuscript.
Funding
This study was supported by FIS (Institute of Health Carlos III, Madrid/ERDF, PI20/01107) and CIBERONC (CB16/12/00484). FNB (Fundación Neuroblastoma) and Fundación CRIS (2023/188) fund AL-C contract. The funders had no involvement in the research process or the preparation and submission of the article.
Institutional Review Board Statement
This study was approved by the Research Ethics Committee of the Clinic Hospital of Valencia (No. 2020/025, Act: 372, 30/09/2021).
Informed Consent Statement
Participants or their family members/legal guardians provided written informed consent for histological and molecular studies performed in our laboratory.
Data Availability Statement
The datasets used and analyzed in the current study are available from the corresponding author on reasonable request.
Acknowledgments
The authors thank the Spanish Society of Paediatric Haemato-Oncology (SEHOP) and Desireé Ramal from University and Polytechnic Hospital la Fe de Valencia for patient data management. We also thank Ruth Collado, Ruth Panadero, Laura Martínez, Marta Gómez, Marta Valls, and Karina Parra-Haro, who are/were part of our Translational Research Group in Pediatric Solid Tumors of INCLIVA for her technical support, the oncopediatricians, for collaborating with us in obtaining plasma from NB patients, Kathryn Davies for English correction, and we especially thank the NB patients and legal guardians for participating in this study.
Conflicts of Interest
The authors declare that they have no competing interests.
References
- Schvartz, I.; Seger, D.; Shaltiel, S. Vitronectin. Int. J. Biochem. Cell Biol. 1999, 31, 539–544. [Google Scholar] [CrossRef] [PubMed]
- Waltz, D.A.; Natkin, L.R.; Fujita, R.M.; Wei, Y.; Chapman, H.A. Plasmin and plasminogen activator inhibitor type 1 promote cellular motility by regulating the interaction between the urokinase receptor and vitronectin. J. Clin. Invest. 1997, 100, 58–67. [Google Scholar] [CrossRef] [PubMed]
- Madsen, C.D.; Ferraris, G.M.S.; Andolfo, A.; Cunningham, O.; Sidenius, N. uPAR-induced cell adhesion and migration: Vitronectin provides the key. J. Cell Biol. 2007, 177, 927–939. [Google Scholar] [CrossRef] [PubMed]
- Leavesley, D.I.; Kashyap, A.S.; Croll, T.; Sivaramakrishnan, M.; Shokoohmand, A.; Hollier, B.G.; Upton, Z. Vitronectin-Master controller or micromanager? IUBMB Life 2013, 65, n/a-n/a. [Google Scholar] [CrossRef] [PubMed]
- Burgos-Panadero, R.; Noguera, I.; Cañete, A.; Navarro, S.; Noguera, R. Vitronectin as a molecular player of the tumor microenvironment in neuroblastoma. BMC Cancer 2019, 19. [Google Scholar] [CrossRef] [PubMed]
- Uhm, J.H.; Dooley, N.P.; Kyritsis, A.P.; Rao, J.S.; Gladson, C.L.; H U, N.-O.J. Vitronectin, a Glioma-derived Extracellular Matrix Protein, Protects Tumor Cells from Apoptotic Death 1. Clin Cancer Res. 1999, 5, 1587–1594. [Google Scholar] [PubMed]
- Li, R.; Ren, M.; Chen, N.; Luo, M.; Zhang, Z.; Wu, J. Vitronectin Increases Vascular Permeability by Promoting VE-Cadherin Internalization at Cell Junctions. PLoS One 2012, 7, e37195. [Google Scholar] [CrossRef] [PubMed]
- Reilly, J.T.; Nash, G. Vitronectin (serum spreading factor): its localisation in normal and fibrotic tissue. J Clin Pathol 1988, 41, 1269–1272. [Google Scholar] [CrossRef] [PubMed]
- Heyman, L.; Leroy-Dudal, J.; Fernandes, J.; Seyer, D.; Dutoit, S.; Carreiras, F. Mesothelial vitronectin stimulates migration of ovarian cancer cells. Cell Biol. Int. 2010, 34, 493–502. [Google Scholar] [CrossRef]
- Radwan, A.F.; Ismael, O.E.; Fawzy, A.; El-Mesallamy, H.O. Evaluation of Serum Integrin αvβ3 & Vitronectin in the Early Diagnosis of Breast Cancer. Clin. Lab. 2019, 65, 1185–1193. [Google Scholar] [CrossRef]
- Edwards, S.; Lalor, P.F.; Tuncer, C.; Adams, D.H. Vitronectin in human hepatic tumours contributes to the recruitment of lymphocytes in an avb3-independent manner. Br. J. Cancer 2006, 95, 1545–1554. [Google Scholar] [CrossRef] [PubMed]
- Ciereszko, A.; Dietrich, M.A.; Słowińska, M.; Nynca, J.; Ciborowski, M.; Kisluk, J.; Michalska-Falkowska, A.; Reszec, J.; Sierko, E.; Nikliński, J. Identification of protein changes in the blood plasma of lung cancer patients subjected to chemotherapy using a 2D-DIGE approach. PLoS One 2019, 14. [Google Scholar] [CrossRef] [PubMed]
- Bera, A.; Subramanian, M.; Karaian, J.; Eklund, M.; Radhakrishnan, S.; Gana, N.; Rothwell, S.; Pollard, H.; Hu, H.; Shriver, C.D.; et al. Functional role of vitronectin in breast cancer. PLoS One 2020, 15. [Google Scholar] [CrossRef] [PubMed]
- Tas, F.; Karabulut, S.; Bilgin, E.; Tastekin, D.; Duranyildiz, D. Clinical significance of serum fibronectin and vitronectin levels in melanoma patients. Melanoma Res. 2014, 24, 475–479. [Google Scholar] [CrossRef] [PubMed]
- Stahl, A.; Mueller, B.M. Melanoma cell migration on vitronectin: regulation by components of the plasminogen activation system. Int. J. Cancer 1997, 71, 116–122. [Google Scholar] [CrossRef]
- Cheung, N.-K. V; Dyer, M.A. Neuroblastoma: developmental biology, cancer genomics and immunotherapy. Nat Rev Cancer 2013, 13, 397–411. [Google Scholar] [CrossRef] [PubMed]
- Cohn, S.L.; Pearson, A.D.J.; London, W.B.; Monclair, T.; Ambros, P.F.; Brodeur, G.M.; Faldum, A.; Hero, B.; Iehara, T.; Machin, D.; et al. The International Neuroblastoma Risk Group (INRG) classification system: An INRG task force report. J. Clin. Oncol. 2009, 27, 289–297. [Google Scholar] [CrossRef]
- Von Stedingk, K.; De Preter, K.; Vandesompele, J.; Noguera, R.; Øra, I.; Koster, J.; Versteeg, R.; Påhlman, S.; Lindgren, D.; Axelson, H. Individual patient risk stratification of high-risk neuroblastomas using a two-gene score suited for clinical use. Int. J. Cancer 2015, 137, 868–877. [Google Scholar] [CrossRef]
- Vicente-Munuera, P.; Burgos-Panadero, R.; Noguera, I.; Navarro, S.; Noguera, R.; Escudero, L.M. The topology of vitronectin: A complementary feature for neuroblastoma risk classification based on computer-aided detection. Int. J. Cancer 2020, 146, 553–565. [Google Scholar] [CrossRef]
- López-Carrasco, A.; Martín-Vañó, S.; Burgos-Panadero, R.; Monferrer, E.; Berbegall, A.P.; Fernández-Blanco, B.; Navarro, S.; Noguera, R. Impact of extracellular matrix stiffness on genomic heterogeneity in MYCN-amplified neuroblastoma cell line. J. Exp. Clin. Cancer Res. 2020, 39. [Google Scholar] [CrossRef]
- Monferrer, E.; Sanegre, S.; Martín-Vañó, S.; García-Lizarribar, A.; Burgos-Panadero, R.; López-Carrasco, A.; Navarro, S.; Samitier, J.; Noguera, R. Digital image analysis applied to tumor cell proliferation, aggressiveness, and migration-related protein synthesis in neuroblastoma 3d models. Int. J. Mol. Sci. 2020, 21, 1–17. [Google Scholar] [CrossRef]
- Monferrer, E.; Martín-Vañó, S.; Carretero, A.; García-Lizarribar, A.; Burgos-Panadero, R.; Navarro, S.; Samitier, J.; Noguera, R. A three-dimensional bioprinted model to evaluate the effect of stiffness on neuroblastoma cell cluster dynamics and behavior. Sci. Rep. 2020, 10, 6370. [Google Scholar] [CrossRef]
- Monferrer, E.; Dobre, O.; Trujillo, S.; González Oliva, M.A.; Trubert-Paneli, A.; Acevedo-León, D.; Noguera, R.; Salmeron-Sanchez, M. Vitronectin-based hydrogels recapitulate neuroblastoma growth conditions. Front. cell Dev. Biol. 2022, 10. [Google Scholar] [CrossRef] [PubMed]
- Repetto, O.; Caggiari, L.; De Zorzi, M.; Elia, C.; Mussolin, L.; Buffardi, S.; Pillon, M.; Muggeo, P.; Casini, T.; Steffan, A.; et al. Quantitative Plasma Proteomics to Identify Candidate Biomarkers of Relapse in Pediatric/Adolescent Hodgkin Lymphoma. Int. J. Mol. Sci. 2022, 23. [Google Scholar] [CrossRef]
- Braoudaki, M.; Lambrou, G.I.; Vougas, K.; Karamolegou, K.; Tsangaris, G.T.; Tzortzatou-Stathopoulou, F. Protein biomarkers distinguish between high- and low-risk pediatric acute lymphoblastic leukemia in a tissue specific manner. J. Hematol. Oncol. 2013, 6, 1–20. [Google Scholar] [CrossRef]
- Kadowaki, M.; Sangai, T.; Nagashima, T.; Sakakibara, M.; Yoshitomi, H.; Takano, S.; Sogawa, K.; Umemura, H.; Fushimi, K.; Nakatani, Y.; et al. Identification of vitronectin as a novel serum marker for early breast cancer detection using a new proteomic approach. J. Cancer Res. Clin. Oncol. 2011, 137, 1105–1115. [Google Scholar] [CrossRef] [PubMed]
- Turan, T.; Torun, M.; Atalay, F.; Gönenç, A.; Turk, J. Endometrial ve Ovaryum Kanserinde Tanısal Değer Yönünden Serum Biyobelirteci Olarak Vitronektin, Solubl Epitel-Kaderin ve TGF-β1’in Değerlendirilmesi Assessment of Vitronectin, Soluble Epithelial-Cadherin and TGF-β1 as a Serum Biomarker with Predictive Value for Endometrial and Ovarian Cancers. Pharm Sci 2017, 14, 141–147. [Google Scholar] [CrossRef]
- Ortega-Martínez, I.; Gardeazabal, J.; Erramuzpe, A.; Sanchez-Diez, A.; Cortés, J.; García-Vázquez, M.D.; Pérez-Yarza, G.; Izu, R.; Luís Díaz-Ramón, J.; de la Fuente, I.M.; et al. Vitronectin and dermcidin serum levels predict the metastatic progression of AJCC I–II early-stage melanoma. Int. J. Cancer 2016, 139, 1598–1607. [Google Scholar] [CrossRef]
- Fukushima, Y.; Tamura, M.; Nakagawa, H.; Itoh, K. Induction of glioma cell migration by vitronectin in human serum and cerebrospinal fluid. J. Neurosurg. 2007, 107, 578–585. [Google Scholar] [CrossRef]
- Schneider, G.; Suszynska, M.; Kakar, S.; Ratajczak, M.Z. Vitronectin in the ascites of human ovarian carcinoma acts as a potent chemoattractant for ovarian carcinoma: Implication for metastasis by cancer stem cells. J. cancer stem cell Res. 2016, 4, 1. [Google Scholar] [CrossRef]
- Tugcu, D.; Devecioglu, O.; Unuvar, A.; Ekmekci, H.; Ekmekci, O.B.; Anak, S.; Ozturk, G.; Akcay, A.; Aydogan, G. Plasma Levels of Plasminogen Activator Inhibitor Type 1 and Vitronectin in Children With Cancer. Clin. Appl. Thromb. Hemost. 2016, 22, 28–33. [Google Scholar] [CrossRef] [PubMed]
- López-Carrasco, A.; Berbegall, A.P.; Martín-Vañó, S.; Blanquer-Maceiras, M.; Castel, V.; Navarro, S.; Noguera, R. Intra-Tumour Genetic Heterogeneity and Prognosis in High-Risk Neuroblastoma. Cancers (Basel). 2021, 13. [Google Scholar] [CrossRef] [PubMed]
- Braekeveldt, N.; Wigerup, C.; Gisselsson, D.; Mohlin, S.; Merselius, M.; Beckman, S.; Jonson, T.; Börjesson, A.; Backman, T.; Tadeo, I.; et al. Neuroblastoma patient-derived orthotopic xenografts retain metastatic patterns and geno- and phenotypes of patient tumours. Int. J. Cancer 2015, 136, E252–E261. [Google Scholar] [CrossRef] [PubMed]
- Newall, F.; Johnston, L.; Ignjatovic, V.; Summerhayes, R.; Monagle, P. Age-related plasma reference ranges for two heparin-binding proteins – vitronectin and platelet factor 4. Int. J. Lab. Hematol. 2009, 31, 683–687. [Google Scholar] [CrossRef] [PubMed]
- Trigg, R.M.; Shaw, J.A.; Turner, S.D. Opportunities and challenges of circulating biomarkers in neuroblastoma. Open Biol. 2019, 9. [Google Scholar] [CrossRef] [PubMed]
- Zogchel, L.M.J. van; Wezel, E.M. van; Wijk, J. van; Stutterheim, J.; Bruins, W.S.C.; Zappeij-Kannegieter, L.; Slager, T.J.E.; Schumacher-Kuckelkorn, R.; Verly, I.R.N.; Schoot, C.E. van der; et al. Hypermethylated RASSF1A as Circulating Tumor DNA Marker for Disease Monitoring in Neuroblastoma. JCO Precis Oncol. 2020, 4, PO.19.00261. [CrossRef]
- Filipe, E.C.; Chitty, J.L.; Cox, T.R. Charting the unexplored extracellular matrix in cancer. Int. J. Exp. Pathol. 2018, 99, 58–76. [Google Scholar] [CrossRef] [PubMed]
- Hahn, W.C.; Bader, J.S.; Braun, T.P.; Califano, A.; Clemons, P.A.; Druker, B.J.; Ewald, A.J.; Fu, H.; Jagu, S.; Kemp, C.J.; et al. An expanded universe of cancer targets. Cell 2021, 184, 1142–1155. [Google Scholar] [CrossRef] [PubMed]
- Schneider, G.; Bryndza, E.; Poniewierska-Baran, A.; Serwin, K.; Suszynska, M.; Sellers, Z.P.; Merchant, M.L.; Kaliappan, A.; Ratajczak, J.; Kucia, M.; et al. Evidence that vitronectin is a potent migration-enhancing factor for cancer cells chaperoned by fibrinogen: a novel view of the metastasis of cancer cells to low-fibrinogen lymphatics and body cavities. 7. Oncotarget, 2016; 7, 69829–69843. [Google Scholar] [CrossRef]
- Lee, H.-J.; Cha, H.-J.; Lim, J.-S.; Lee, S.H.; Song, S.Y.; Kim, H.; Hancock, W.S.; Yoo, J.S.; Paik, Y.-K. Abundance-Ratio-Based Semiquantitative Analysis of Site-Specific N-Linked Glycopeptides Present in the Plasma of Hepatocellular Carcinoma Patients. Proteome Res. 2014, 13, 2328–2338. [Google Scholar] [CrossRef] [PubMed]
- Benachour, H. ; Leroy-Dudal, J; Agniel, R; Wilson, J; Briand, M; Carreiras, F; Gallet, O. Vitronectin (Vn) glycosylation patterned by lectin affinity assays-A potent glycoproteomic tool to discriminate plasma Vn from cancer ascites Vn. J Mol Recognit, 2018; 31, e2690. [Google Scholar] [CrossRef]
- Hao, W.; Zhang, X.; Xiu, B.; Yang, X.; Hu, S.; Liu, Z.; Duan, C.; Jin, S.; Ying, X.; Zhao, Y.; et al. Vitronectin: a promising breast cancer serum biomarker for early diagnosis of breast cancer in patients. Tumor Biol. 2016, 37, 8909–8916. [Google Scholar] [CrossRef]
- Chen, M.H.; Lu, C.; Sun, J.; Chen, X.D.; Dai, J.X.; Cai, J.Y.; Chen, X.L. Diagnostic and prognostic value of serum vitronectin levels in human glioma. J. Neurol. Sci. 2016, 371, 54–59. [Google Scholar] [CrossRef]
- Gladson, C.L.; Cheresht, D.A. Glioblastoma Expression of Vitronectin and the av/83 Integrin. Adhesion Mechanism for Transformed Glial Cells Key words: glioblastoma * invasion * vitronectin * avβ3 integrin. J. Clin. Invest 1991, 88, 1924–1932. [Google Scholar] [CrossRef] [PubMed]
- Mikuła-Pietrasik, J.; Uruski, P.; Tykarski, A. ; Książek, · Krzysztof The peritoneal “soil” for a cancerous “seed”: a comprehensive review of the pathogenesis of intraperitoneal cancer metastases. Cell. Mol. Life Sci 2018, 75, 509–525. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, J.; Yamada, S.; Kawasaki, H. Distribution of vitronectin in plasma and liver tissue: Relationship to chronic liver disease. Hepatology 1994, 20, 1412–1417. [Google Scholar] [CrossRef] [PubMed]
- Stoks, M.; Vieco-Martí, I.; Noguera, I.; Sánchez-Sánchez, M.; Burgos-Panadero, R.; Navarro, S.; Noguera, R. Digital image analysis workflows for evaluation of cell behavior and tumor microenvironment to aid therapeutic assessment in high-risk neuroblastoma. Comput. Biol. Med. 2023, 164, 107364. [Google Scholar] [CrossRef] [PubMed]
- Burgos-Panadero, R.; El Moukhtari, S.H.; Noguera, I.; Rodríguez-Nogales, C.; Martín-Vañó, S.; Vicente-Munuera, P.; Cañete, A.; Navarro, S.; Blanco-Prieto, M.J.; Noguera, R. Unraveling the extracellular matrix-tumor cell interactions to aid better targeted therapies for neuroblastoma. Int. J. Pharm. 2021, 608, 121058. [Google Scholar] [CrossRef] [PubMed]
- Kurozumi, K.; Ichikawa, T.; Onishi, M.; Fujii, K.; Date, I. Cilengitide treatment for malignant glioma: current status and future direction. Neurol. Med. Chir. (Tokyo). 2012, 52, 539–547. [Google Scholar] [CrossRef] [PubMed]
- Reardon, D.A.; Nabors, L.B.; Stupp, R.; Mikkelsen, T. Cilengitide: an integrin-targeting arginine-glycine-aspartic acid peptide with promising activity for glioblastoma multiforme. Expert Opin. Investig. Drugs 2008, 17, 1225–1235. [Google Scholar] [CrossRef]
- Hasturk, O.; Jordan, K.E.; Choi, J.; Kaplan, D.L. Enzymatically crosslinked silk and silk-gelatin hydrogels with tunable gelation kinetics, mechanical properties and bioactivity for cell culture and encapsulation. 2019. [CrossRef]
- Villamón, E.; Piqueras, M.; Meseguer, J.; Blanquer, I.; Berbegall, A.P.; Tadeo, I.; Hernández, V.; Navarro, S.; Noguera, R. NeuPAT: An intranet database supporting translational research in neuroblastic tumors. Comput. Biol. Med. 2013, 43, 219–228. [Google Scholar] [CrossRef]
Figure 1.
VN secretion by NB cells to culture media. A. Origin and main clinical-molecular characteristics of the four NB cell lines. All presented aggressive features as derived from stage 4 patients, being MYCN amplified or ALK mutated and having SCAs. *For more details of PDX origin see [33]. B. Comparison of VN levels secreted to culture media of 2D (monolayer) and 3D HGs cultures by NB cell lines in which VN detection was positive by ELISA (p – value= 0.0052). C. Concentration of VN secreted by the four NB cell lines to culture media of the 3D HGs, and millions of cells calculated with digital analysis in two HGs from which we collected the culture media; blue bars refer to HGs with two weeks of culture, green bars to three weeks of culture, patterned bars to HGs without cross-linked VN, solid bars to HGs with cross-linked VN, and millions of cells estimated in the two HGs are represented with a solid black line.
Figure 1.
VN secretion by NB cells to culture media. A. Origin and main clinical-molecular characteristics of the four NB cell lines. All presented aggressive features as derived from stage 4 patients, being MYCN amplified or ALK mutated and having SCAs. *For more details of PDX origin see [33]. B. Comparison of VN levels secreted to culture media of 2D (monolayer) and 3D HGs cultures by NB cell lines in which VN detection was positive by ELISA (p – value= 0.0052). C. Concentration of VN secreted by the four NB cell lines to culture media of the 3D HGs, and millions of cells calculated with digital analysis in two HGs from which we collected the culture media; blue bars refer to HGs with two weeks of culture, green bars to three weeks of culture, patterned bars to HGs without cross-linked VN, solid bars to HGs with cross-linked VN, and millions of cells estimated in the two HGs are represented with a solid black line.
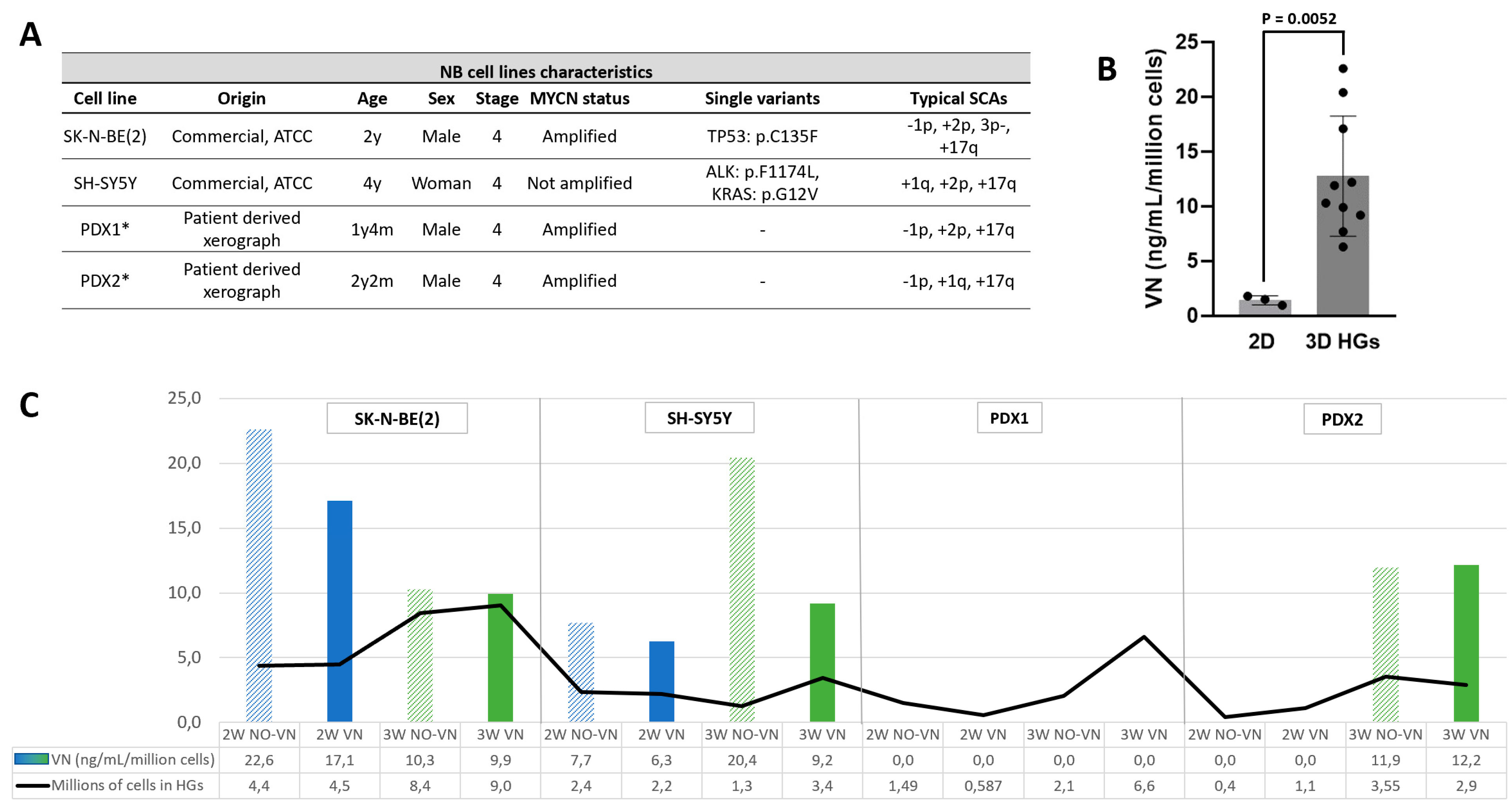
Figure 2.
Plasma VN level of NB cohort and clinic-molecular landscape overview. The top panels show VN levels in plasma stratified by the cutoff of 361 µg/mL in patients older than 18 months, and by quartiles across all dead patients. The central panels show clinical characteristics including age, overall survival (OS), event free survival (EFS) and stage. MYCN status, genomic profile and typSCAs are shown in the bottom panels. See figure legend for color codes. MNNA, MYCN non-amplified; hetMNA, MYCN heterogeneously amplified; MNA, MYCN amplified; NCA, numerical chromosomal alteration; TypSCA, typical segmental chromosomal alterations; AtypSCA, atypical segmental chromosomal alterations.
Figure 2.
Plasma VN level of NB cohort and clinic-molecular landscape overview. The top panels show VN levels in plasma stratified by the cutoff of 361 µg/mL in patients older than 18 months, and by quartiles across all dead patients. The central panels show clinical characteristics including age, overall survival (OS), event free survival (EFS) and stage. MYCN status, genomic profile and typSCAs are shown in the bottom panels. See figure legend for color codes. MNNA, MYCN non-amplified; hetMNA, MYCN heterogeneously amplified; MNA, MYCN amplified; NCA, numerical chromosomal alteration; TypSCA, typical segmental chromosomal alterations; AtypSCA, atypical segmental chromosomal alterations.
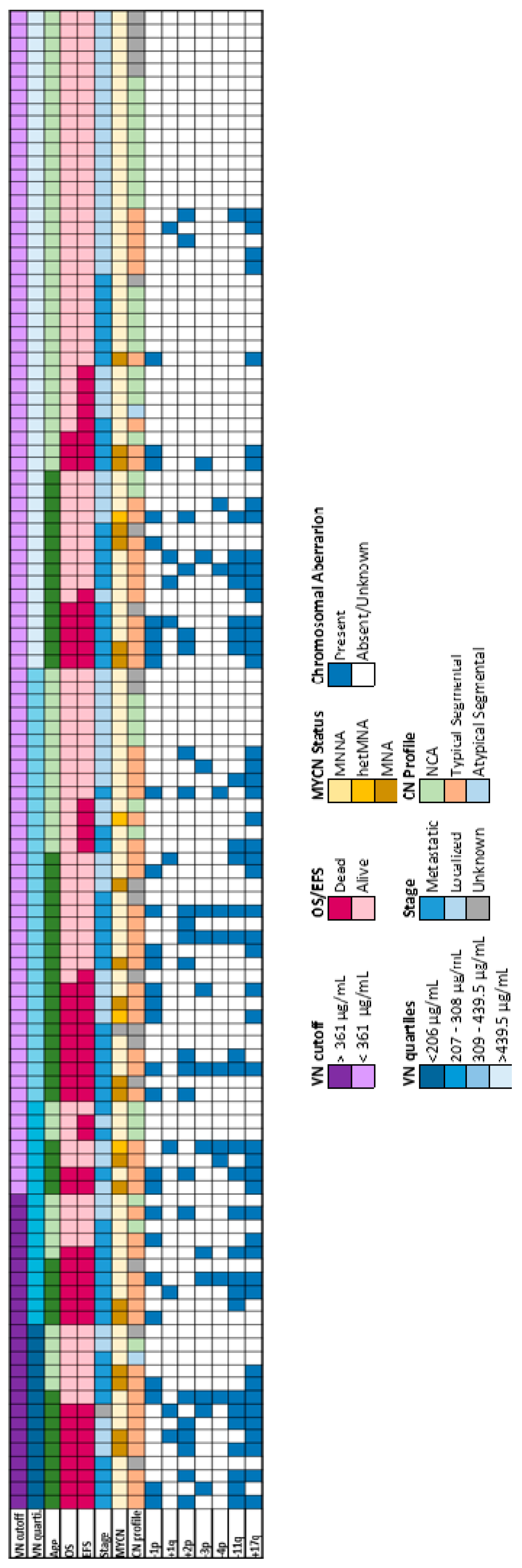
Figure 3.
VN levels in plasma of NB patients. A. Descriptive statistics of patient numbers and VN levels detected by ELISA in plasma of all patients younger and older than 18 months, of the patients who remained alive and of those who died. B. Graphic representation of VN levels in the mentioned groups. VN concentration was significantly different between alive and dead patients older than 18 months (p-value = 0.0074). C. The ROC curve obtained for patients older than 18 months was significant (p-value = 0.0026) and allowed us to establish a cutoff of 361 µL/mL (red arrow), over which patients showed a poor prognosis.
Figure 3.
VN levels in plasma of NB patients. A. Descriptive statistics of patient numbers and VN levels detected by ELISA in plasma of all patients younger and older than 18 months, of the patients who remained alive and of those who died. B. Graphic representation of VN levels in the mentioned groups. VN concentration was significantly different between alive and dead patients older than 18 months (p-value = 0.0074). C. The ROC curve obtained for patients older than 18 months was significant (p-value = 0.0026) and allowed us to establish a cutoff of 361 µL/mL (red arrow), over which patients showed a poor prognosis.
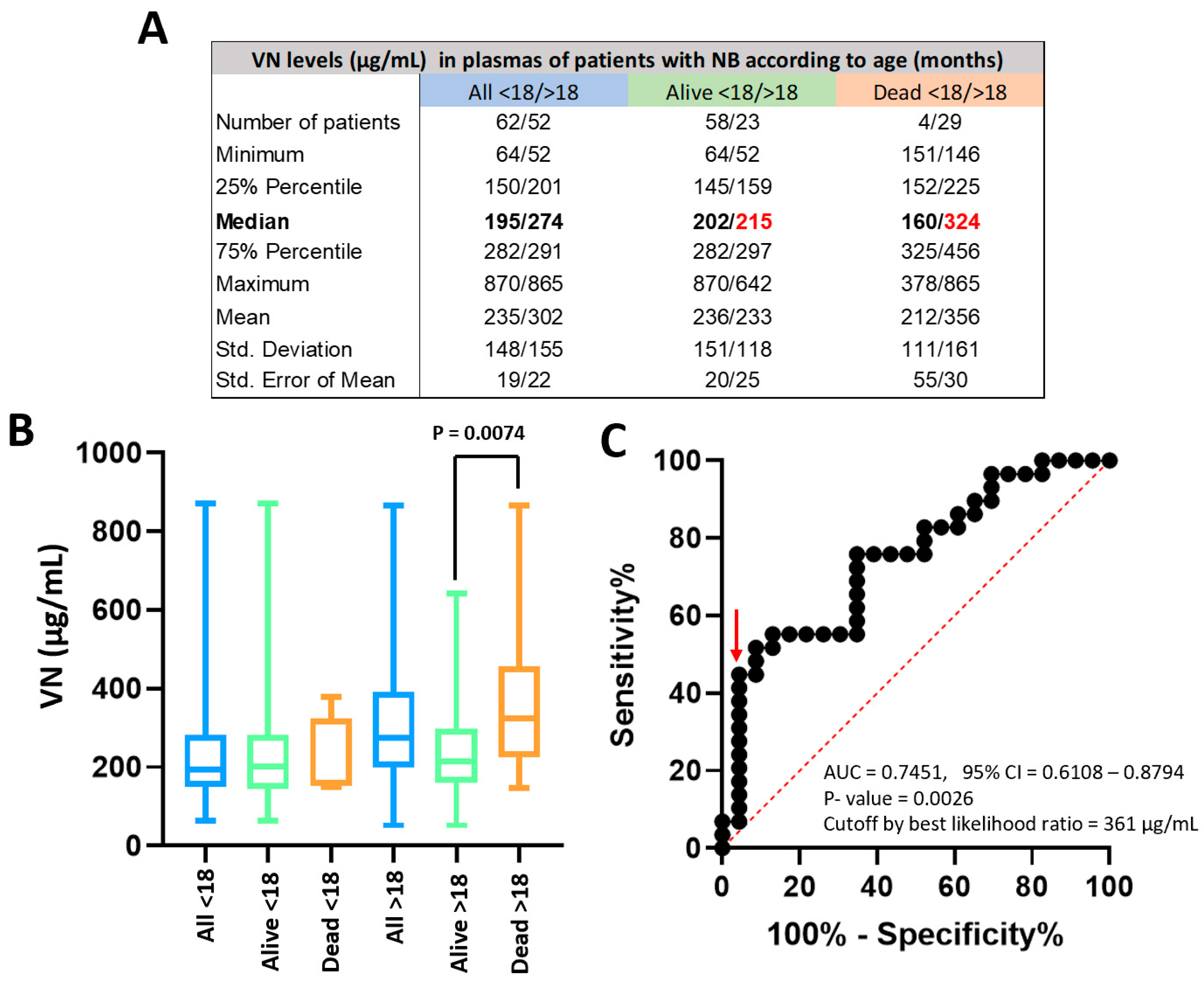
Figure 4.
Survival probabilities of NB patients older than 18 months according to VN levels. Kaplan-Meier curves for A. Overall survival (OS) and B. Event free survival (EFS) reflected the significantly low survival of NB patients with VN plasma levels over the cutoff (p = 0.0018 and p = 0.0027, respectively). Five-year OS for patients with VN levels under and over the cutoff was 53% and 7%, respectively, and for EFS was 45% and 7%, respectively.
Figure 4.
Survival probabilities of NB patients older than 18 months according to VN levels. Kaplan-Meier curves for A. Overall survival (OS) and B. Event free survival (EFS) reflected the significantly low survival of NB patients with VN plasma levels over the cutoff (p = 0.0018 and p = 0.0027, respectively). Five-year OS for patients with VN levels under and over the cutoff was 53% and 7%, respectively, and for EFS was 45% and 7%, respectively.
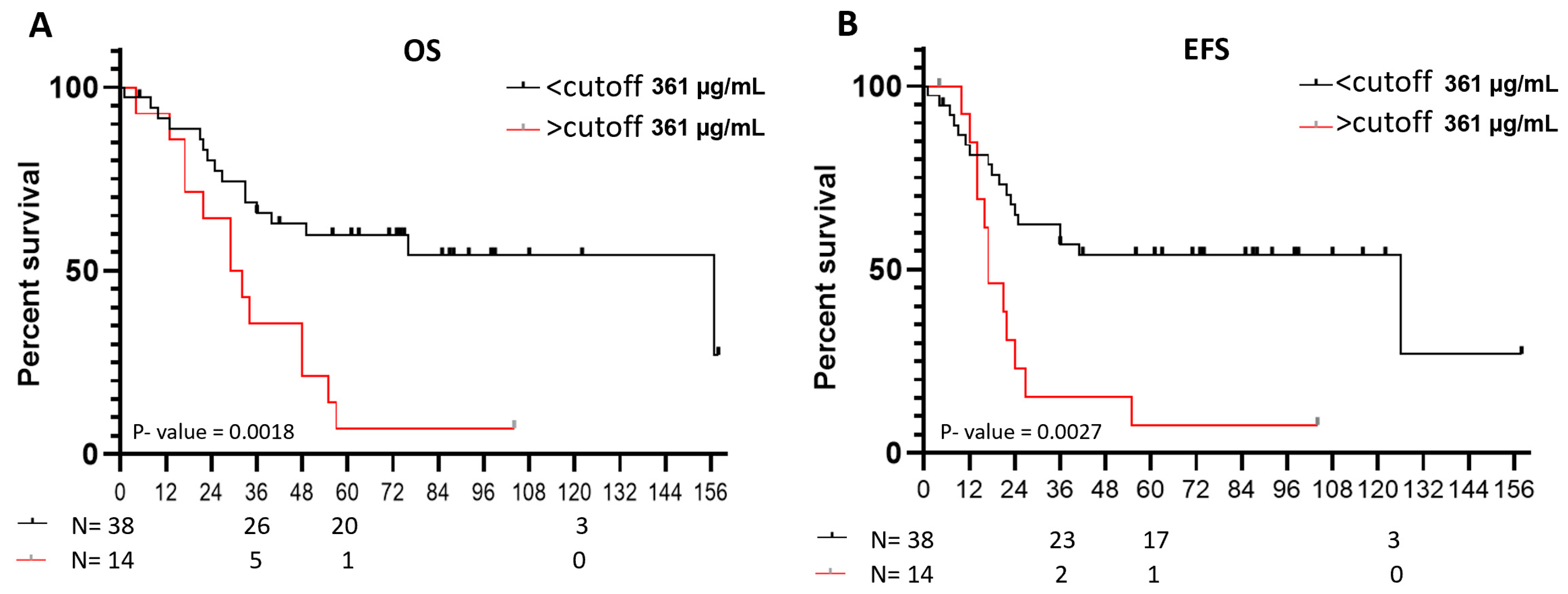

Table 1.
VN level by cutoff is associated with death, stage, genomic profile and 11q loss in NB patients older than 18 months. MYCN status: MNNA, MYCN non-amplified; hetMNA, MYCN heterogeneously amplified; MNA, MYCN amplified; NCA, numerical chromosomal alteration; SCA, segmental chromosomal alterations.
Table 1.
VN level by cutoff is associated with death, stage, genomic profile and 11q loss in NB patients older than 18 months. MYCN status: MNNA, MYCN non-amplified; hetMNA, MYCN heterogeneously amplified; MNA, MYCN amplified; NCA, numerical chromosomal alteration; SCA, segmental chromosomal alterations.
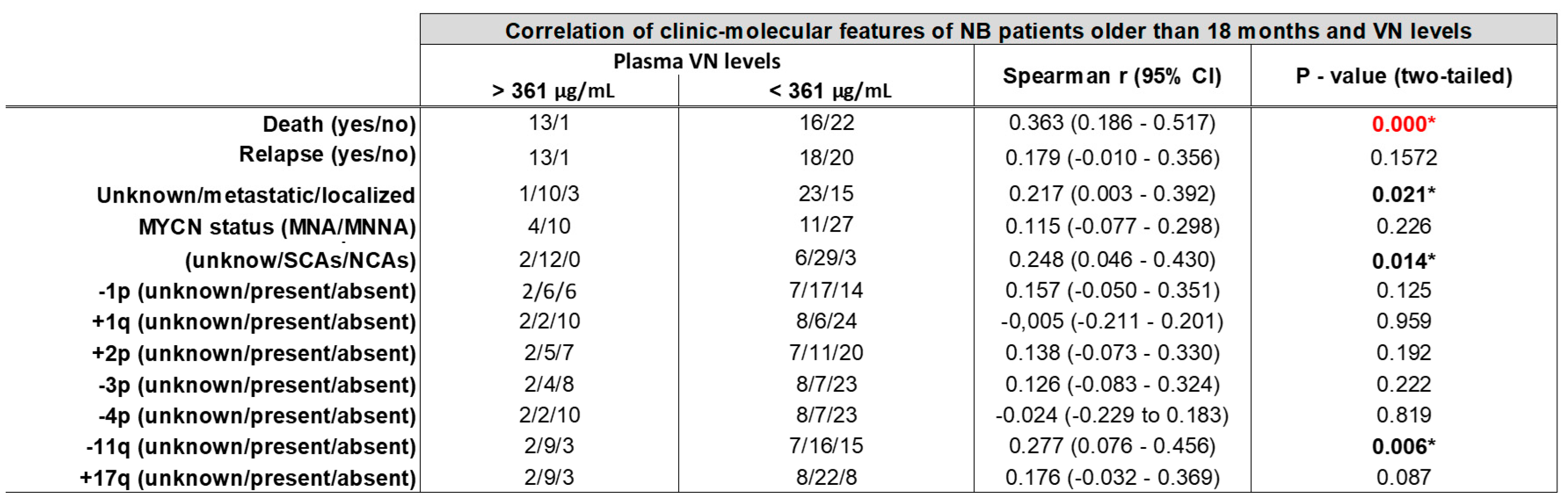 |
Table 2.
Cox regression tests results after applying stepwise forward and backward Wald methods: last step. VN levels dichotomized by cutoff in NB patients older than 18 months, MYCN status (MYCN status: MNNA, MYCN non-amplified; hetMNA, MYCN heterogeneously amplified; MNA, MYCN amplified;) and loss of 4p chromosomic region were the variables maintained in the last step of the Cox regression tests according to EFS and OS; however, only VN levels showed significant p-values. Other variables without significant results included in the Cox regression analysis were age, stage (localized/metastatic), genomic profile (NCAs, chromosomal numeric aberrations/SCAs, chromosomal segmental aberrations) and typical SCAs (-1p, +1q, +2p, 3p-, -4p, -11q, +17q). B: Beta coefficient; SE: Standard Error. Coefficients Exp (B) > 1 indicate that high values of this parameter increase its probability of being an independent poor prognostic factor.
Table 2.
Cox regression tests results after applying stepwise forward and backward Wald methods: last step. VN levels dichotomized by cutoff in NB patients older than 18 months, MYCN status (MYCN status: MNNA, MYCN non-amplified; hetMNA, MYCN heterogeneously amplified; MNA, MYCN amplified;) and loss of 4p chromosomic region were the variables maintained in the last step of the Cox regression tests according to EFS and OS; however, only VN levels showed significant p-values. Other variables without significant results included in the Cox regression analysis were age, stage (localized/metastatic), genomic profile (NCAs, chromosomal numeric aberrations/SCAs, chromosomal segmental aberrations) and typical SCAs (-1p, +1q, +2p, 3p-, -4p, -11q, +17q). B: Beta coefficient; SE: Standard Error. Coefficients Exp (B) > 1 indicate that high values of this parameter increase its probability of being an independent poor prognostic factor.
 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



