Submitted:
12 July 2024
Posted:
12 July 2024
You are already at the latest version
Abstract
The evolution of the H5 highly pathogenic avian influenza (HPAI) viruses has led to the emergence of distinct groups with genetically similar clusters of hemagglutinin (HA) sequences. In this study, a consensus H5 HA sequence was cloned into the baculovirus expression system. The HA protein was expressed in baculovirus-infected insect cells and utilized as the antigen for the production of an oil emulsion-based H5 avian influenza vaccine (rBacH5Con5Mut). 21-day old SPF chickens were immunized with this vaccine and then challenged at 21 days post-vaccination (dpv) with clade 2.3.2.1, clade 2.3.4.4 and clade 7.2 of H5 HPAI virus, respectively. The chicken sera of vaccinated chickens exhibited high hemagglutination inhibition (HI) titers against the rBacH5 vaccine antigen, while lower HI titers were observed against the different challenge virus H5 hemagglutinins. Furthermore, the rBacH5Con5Mut vaccine provided 100% protection from mortality and clinical signs. Virus isolation results showed that oropharyngeal and cloacal shedding was prevented in 100% of the vaccinated chickens when challenged with clade 2.3.2.1 and clade 2.3.4.4 H5 viruses. When the rBacH5Con5Mut vaccine candidate was administrated at one day of age, 100% protection was demonstrated against challenge of clade 2.3.4.4 virus at 3 weeks of age, indicating the potential of this vaccine for hatchery vaccination. Overall, the rBacH5Con5Mut vaccine candidate with a consensus HA antigen and a single dose can protect chickens against different clades of H5 HPAI viruses throughout the rearing period of broiler chickens without a boost, thus fulfilling the criteria for an efficacious broad-spectrum H5 avian influenza vaccine.
Keywords:
avian influenza virus
; H5 subtype
; subunit vaccine
; broad protection
Introduction
Highly pathogenic avian influenza (HPAI) virus infections cause significant economic losses to the poultry industry and pose a severe threat to public health. The H5 HPAI virus has spread among poultry and wild bird populations globally over the past two decades. These viruses have been categorized into 10 different clades based on the evolution of their hemagglutinin (HA) genes [1,2,3,4,5,6], and clade 2 has been further divided into different sub-clades. In China, clades 2.3.2, 2.3.4, and 7 viruses are responsible for most of disease outbreaks in domestic poultry [9]. Most clade 2.3.2 viruses clustered in Hong Kong, Henan, Inner Mongolia and Qinghai [7]. Clade 2.3.4 viruses first emerged in the south and subsequently spread to the north [8]. In contrast, clade 7 viruses were first isolated in northern China and spread from north to south into Hunan and Yunnan provinces [8].
Several countries have implemented a combination of culling and vaccination strategy to prevent and control the spread of H5 HPAI [5,9,10]. Since 2004, a series of recombinant inactivated vaccines have been developed and updated in China to match the antigenicity of the circulating strains of H5 viruses [9]. However, the rapid mutation rate of the H5 subtype HPAI viruses outpaces the time-consuming procedures of screening new candidate strains for the development of traditional inactivated whole-virus vaccines. Therefore, the existing vaccines might not provide adequate protection against the new emerging variants, posing a significant threat to the poultry industry in China [10]. In addition to the inactivated vaccines, a novel Newcastle virus (NDV)-vectored H5 avian influenza bivalent live vaccine has been used in chickens in China since 2006 [11] . In 2018, the first H5 DNA vaccine was approved [12], however, it has not been used yet due to the rapid evolution of the virus.
Due to the high variations in the HA sequences, especially within the neutralizing epitope regions, conventional HA based H5 avian influenza vaccines appear not to be protective against heterologous strains or phylogenetically variant clades of H5. Therefore, there is an urgent need to develop an effective, broadly cross-protective and safe H5 avian influenza vaccine. A strategy to exploit a cocktail of antigenically different H5 virus strains has been tried to provide broad protection [13]. Other strategies are based on choosing immunogens from conserved regions to provide broad protection against different influenza viruses such as HA stalk domain, highly conserved M2 ectodomain (M2e) [12], HA fusion domain (HFD) [13], T-cell epitope of nucleoprotein (TNP) [14], and HA α-helix domain (HαD) [15,16]. A novel “mosaic” approach which used an algorithm to select and recombine potential 9mer to 12mer “chunks” into a full-length protein in an attempt to preserve naturally occurring T cell epitope was introduced previously [17]. This strategy has been applied to generate a modified vaccinia Ankara-vectored vaccine expressing a mosaic H5 HA [17].
Recent studies have shown that highly conserved regions in the HA protein can induce antibodies that provide subtype-independent protection [18,19,20]. Consensus sequences, which are obtained by aligning a population of sequences and selecting the most common residue at each position, is another promising strategy for influenza vaccine development that can elicit cross-reactive immune responses [6,21,22,23]. There are reports of experimental vaccines with consensus sequence that can effectively protect mice [24,25,26], ferrets [26] and chickens [6] against H5N1 infection.
In this study, an insect-cell-derived single consensus H5 HA antigen-based subunit vaccine (rBacH5Con5Mut) has been developed and two mutations are introduced to further increase its antigenicity. This vaccine candidate can not only broadly protect against lethal challenge of clade 2.3.4.4, clade 2.3.2.1 and clade 7.2 H5 HPAI viruses in chickens, but also protect against virus shedding of clade 2.3.2.1 and clade 2.3.4.4 H5 viruses at 5 days post challenge (dpc) which indicated that our vaccine candidate can prevent virus spreading as well. This rBacH5Con5Mut vaccine also showed a satisfactory safety profile and provided 100% protection to one-day-old chickens against mortality, morbidity and virus shedding following challenge with clade 2.3.4.4 H5 virus, indicating that it has the potential to be used in the hatcheries. Taken together, we developed a single consensus H5 HA antigen-based subunit vaccine that provided satisfactory protection against lethal challenge of different clades of the H5 HPAI viruses.
Materials and Methods
Ethics Statement
All animal experiments were carried out in animal biosafety level 3 (ABSL-3) facilities in compliance with the protocols of the Biosafety Committee of the ABSL-3 Laboratory of the South China Agriculture University (SCAU, CNAS BL0011). All animal experiments were reviewed and approved by the Institutional Animal Care and Use Committee of the South China Agriculture University and were carried out in accordance with the approved guidelines (2017A002).
Cells and Viruses
Three H5 HPAI viruses originating from clade 2.3.2.1 (A/Duck/Guangdong/383/2008), clade 2.3.4.4 (A/Goose/Guangdong/079/2013) and clade 7.2 (A/Duck/Shandong/147/2013) were isolated from the mainland of China between 2008 and 2013 and characterized as the challenge strains by SCAU.
Antigen Design and Virus Production
The complete open reading frame (ORF) of the consensus HA sequence was obtained by aligning the avian-origin complete H5 subtype HA sequences (isolated from 2005 to 2013 in China) in GenBank using the Vector NTI software. A total of 150 HA sequences (NCBI 2005-2013) were analyzed and the highest frequency of amino acid in each position was selected to generate the consensus HA. The additional mutations including S120N and S223N were introduced into the selected HA gene as previously described [27]. This HA (rBacH5Con5Mut) ORF was then codon-optimized according to the codon usage frequency of Spodoptera frugiperda (Sf) cells by GeneScript Co., Ltd. (Nanjing, Jiangsu, China). The optimized rBacH5Con5Mut ORF was cloned into the pVL1393 vector plasmid. To generate the recombinant baculovirus (rBVs), the plasmid and recombinant bacmid DNA was co-transfected into Sf9 insect cells according to the manufacturer’s instructions (rAcNPV, Invitrogen, Carlsbad, CA, USA). Single rBV clones were obtained by the limiting dilution method.
The rBacH5Con5Mut virus stocks were produced in SF+ cells. Briefly, the cells were maintained as suspension cultures in EX-CELL 420 insect-serum-free medium (Sigma) at 27 ± 2°C and infected with the virus at a low multiplicity of infection (MOI) of ≤ 0.01. The supernatant was harvested at four days post-infection (dpi).
Phylogenetic Analysis
Full-length HA sequences of H5 HPAI viruses in China were obtained from GenBank (http://www.ncbi.nlm.nih.gov/genomes/FLU/). HA sequences were aligned, and neighbor-joining (NJ) phylogenies were estimated using MEGA 7 software (https://www.megasoftware.net/). NJ methods were used to perform system reconstruction and to calculate confidence values to estimate the number of repetitions (1000).
Western Blot
Samples were mixed with loading buffer (with or without reducing reagent) and denatured by heating for 5 min at 95°C. The proteins were separated by denaturing polyacrylamide gel electrophoresis (SDS-PAGE) and then transferred to polyvinylidene fluoride (PVDF) membranes. After blocking with 5% skim milk in phosphate buffered saline with 0.05% tween (PBST) for 30 mins, the membranes were probed with mouse anti-H5N1 HA monoclonal antibody (SINO BIOLOGICALS), and then with goat anti-mouse IgG-HRP (BIO-RAD). The positive bands were visualized using the Supersignal West Femto Maximum Sensitivity Substrate (Thermo Scientific).
Hemagglutination (HA) Assay
Samples for HA assay were serially diluted 2-fold in 96 well V-bottom plates, and 1% chicken red blood cells (RBCs) were added to each well. The plates were gently mixed and incubated at room temperature for 45 min. The highest dilution that showed complete hemagglutination was recorded as the HA titer for the sample.
Animal Studies for Vaccine Efficacy Evaluation
SF+ cells were infected with HA-encoding recombinant Baculovirus (rBVs) at an MOI of 0.1 and harvested six days later. The clarified whole cell suspension was incubated with BEI (final concentration of 10 mM) at 37°C for 24 h to inactivate the virus. Then the BEI was neutralized by mixing with NaS2O3•5H2O at 37°C for 1h. The inactivated samples were mixed with water-in-oil adjuvant (30% vaccine antigen/70% oil adjuvant, v/v) and emulsified to formulate the oil adjuvanted inactivated vaccine. The content of HA protein in the final vaccine was measured by hemagglutination assay before emulsion.
The commercial trivalent inactivated whole virus-based mineral oil emulsion Re-6-7-8 vaccine was obtained from Weike Biotechnology Co., Ltd., Harbin, China. The six internal genes of these recombinant vaccine strains were derived from A/Puerto Rico/8/1934 virus (PR8, H1N1). The HA and NA genes of Re-6-7-8 were derived from the strains of different subclades of H5 wild-type viruses: Re6 from A/duck/Guangdong/S1322/2006 (clade 2.3.2) [28], Re7 from A/Chicken/Liaoning/S4092/2011 (clade 7.2) [29], and Re8 from A/chicken/Guizhou/4/13 (clade 2.3.4.4) [30].
Animal Study
Immunization and Challenge
One-day-old or 21-day-old White Leghorn SPF chickens were divided into control and immunized groups (10 chickens per group) and injected subcutaneously with 0.3 ml PBS or the oil adjuvanted vaccine respectively. Three weeks after immunization, serum samples were collected for antibody detection, and the birds were challenged intranasally with 106 egg infectious dose 50 (EID50) of the challenge virus strain. Oropharyngeal and cloacal swabs were collected on 5 days post-challenge for virus isolation. All birds were observed for signs of disease (such as loss of appetite, lack of energy, purple comb, hemorrhage of foot scale, palpebral edema, and neurological disorders (twisting the neck)) or death over a period of 14 days after challenge and then humanely euthanized.
Virus Isolation
To assess virus shedding after challenge, virus was isolated from the swabs (see above) and grown in chicken embryos. Briefly, the cotton swabs were taken into labeled tubes pre-loaded with 1 ml PBS+2000 units of pen-strep. The samples were centrifuged at 3000 g for 10 min and the supernatants were transferred to fresh tubes and stored at -80ºC. Each sample was inoculated into the allantoic cavity of three 9 to 11-day-old SPF chicken embryos (0.2 ml per embryo), which were then incubated at 37°C for 72 h. The embryos that were no longer viable within 24 h post-inoculation were discarded. Allantoic fluid was collected from each embryo, and HA activity was measured.
Hemagglutination Inhibition (HI) Assay
Serum samples were collected and inactivated at 56oC for 30min. The antigen used for HI assay is either BEI inactivated rBacH5Con5Mut or formaldehyde inactivated challenge virus antigen. The serum samples were serially diluted 2-fold in 96 well V-bottom plate, mixed with an equal volume of antigen (4 HA units), and incubated at room temperature for 30 min. After adding 1% chicken RBCs, the plates were gently mixed and incubated for 45 min. The HI titer was calculated as the highest dilution of serum causing complete inhibition of hemagglutination.
Generation of Pseudovirus Bearing HA and NA of H5N1 Avian Influenza Virus
H5HA and N1NA derived from A/Goose/Guangdong/079/2013 were codon-optimized for the mammalian expression system. The 293T cells (purchased from ATCC, Cat: PTA-5077) were seeded into 6-well plates and co-transfected with 6 µg plasmid encoding Env-defective, luciferase-expressing HIV-1 (pNL4-3.luc.RE), 1.6 µg H5HA-pVAX1 and 1.6ug N1NA-pVAX1. The medium was removed and replaced with fresh DMEM 4 h later. Supernatants containing the pseudovirus particles were harvested 48 h post-transfection and used for single-cycle infection. VSV-G pseudovirus was generated as a control by co-transfecting plasmids encoding vesicular stomatitis virus G protein (VSV-G-pcDNA3.1) with pNL4-3.luc.RE plasmid. For virus titration, MDCK cells (imported from Boehringer Ingelheim Vetmedica Inc, USA) were infected with 200 µl of each pseudovirus in 24-well culture plates, and the medium was replaced with fresh medium after 24 h. The infected cells were incubated for 48 h and the luciferase activity was detected. The pseudovirus titer was calculated in terms of relative luciferase units (RLUs) tested by Promega GloMax Navigator.
HA Pseudovirus Neutralization Assay
MDCK cells were seeded in 96-well plates at the density of 5000 cells/well in 100 µl DMEM one day before the test. To prepare the virus samples, 104 RLUs of H5N1 pseudo virus were incubated with two-fold serially diluted sera (starting dilution 1:5) for 1 h at 37°C (in a CO2 incubator) in 160 µl antibiotic-supplemented serum-free MEM. The MDCK cells were washed with PBS and incubated with 150 µl of the neutralized sample per well for 24 h. The medium was discarded and 150 µl fresh DMEM with 10% FBS was added to each well, and the cells were incubated for 48 h. The luciferase assay was performed by adding the bright-Glow Luciferase substrate (Promega). The neutralization of HA pseudovirus was calculated and expressed in terms of the half maximal inhibitory concentration (IC50) calculated by using GraphPad Prism 10.
Results
Characterization of H5Nx HA Expressed by Recombinant Baculovirus
The rBacH5Con5Mut virus encoding a consensus H5 HA sequence (Figure 1a) was constructed by co-transfection using a pVL1393 vector with the baculovirus polyhedrin promoter and linearized baculovirus genomic DNA (SapphireTM, baculovirus DNA and transfection kit, Orbigen, USA). Phylogenetic analysis result showed that this consensus H5 HA gene belonged to clade 2.3.4.1 (Figure 1b). S120N and S223N mutations were introduced into the H5 HA consensus sequence to increase the H5 HA antigenicity. Structural modeling showed that these two mutations were both located in the globular head region (Figure 1c). HA expression in rBacH5Con5Mut-infected SF+ cells was confirmed by dual IFA, western blot and HA assays (Figure 1d, 1e and 1f). Bands corresponding to HA0 and HA1 were detected at around 65 kDa and 50kDa respectively in the immunoblot. The HA2 was not detected because the monoclonal antibody cannot bind with HA2. The presence of the HA1 band can be attributed to the partial cleavage of HA0 in the insect cells [31] . Furthermore, dimer (~ 130 kDa) and trimer (~ 195 kDa) forms of HA0, as well as the oligomer form with high molecular weight were detected. The HA titer of H5 HA was 9 log2, and dual IFA assay also confirmed the expression of H5 HA. Moreover, the expression of Gp64 and HA overlapped in every plaque completely, which indicated that the rBacH5Con5Mut virus was a pure virus after plaque purification. Taken together, these results confirmed the successful expression of H5 consensus HA in rBacH5Con5Mut-infected SF+ cells. The expressed consensus H5 HA can be recognized by a H5 HA monoclonal antibody and elicits hemagglutination activity.
Genetic Stability of rBacH5Con5Mut Virus
The rBacH5Con5Mut virus was subjected to plaque purification and its monoclonality was verified as methods described above. To check the genetic stability of rBacH5Con5Mut, the virus was continuously passaged on SF+ cells with 10 passages to P17, and the H5 HA gene was analyzed by PCR and sequencing. The dual IFA assay confirmed the expression of H5 HA protein and the purity of the recombinant virus (Figure 2a). As shown in Figure 2b, there were no deletions in the H5 HA segment and the gene sequencing of the PCR production result showed that there were no mutations in the H5 HA gene when passaged to P17. Furthermore, the rBacH5Con5Mut virus from P7, P12 and P17 showed no significant differences in terms of H5 HA expression level or HA activity (Figure 2c and 2d). These findings indicated that the designed consensus H5 HA gene was stable in recombinant baculovirus for up to 17 passages.
A single Immunization with rBacH5Con5Mut Vaccine Prevented Infection from 2.3.4.4 Clade H5N1 Virus
To test the efficacy of the rBacH5Con5Mut vaccine against widely circulating poultry HPAI strains in China, 21-day old chickens were vaccinated with the rBacH5Con5Mut vaccine and then challenged with the A/Goose/Guangdong/079/2013 (H5N1, 14079) strain, belonging to clade 2.3.4.4, after 21 days post immunization. In addition, a positive control group immunized with the Re-6-7-8 commercial vaccine and a PBS-treated mock control group were included in the study for comparison. Serum samples were collected at 21 days post-vaccination (dpv) from the different groups for serological tests.
As shown in Table 1, rBacH5Con5Mut induced HI geometric mean titer (GMT) of 9.1 log2 against the recombinant H5 (rBacH5) antigen at 21 dpv. The highest HI titer of single sera was 10 log2, while the lowest HI titer was 8 log2. The average HI titer against the standard antigen of the challenge virus strain was 3.6 log2. In contrast, the HI titer of the sera of PBS-treated controls was less than 4 log2 for rBacH5 antigen and 0 log2 for the challenge virus standard antigen. Furthermore, the HI titers of the sera from Re-6-7-8 vaccination group were 6.5 log2 and 8.0 log2 against rBacH5 antigen and challenge strain respectively.
The chickens were challenged at 21 dpv with 106 EID50 of the 14079 virus. Clinical signs and mortality were monitored for 14 days. All PBS-treated chickens died within two days following the challenge. However, none of the chickens vaccinated with rBacH5con5Mut or Re-6-7-8 vaccine died or showed any clinical signs after infection, indicating that both vaccines provided 100% protection from mortality and morbidity (Figure 3a). Virus shedding was tested at 5 dpc by isolating the virus from both oropharyngeal and cloacal swab samples (Table 1). No shedding was observed from the cloacal or oropharyngeal samples of the rBacH5con5Mut and Re-6-7-8 vaccination groups. Taken together, these results clearly indicated this rBacH5Con5Mut subunit vaccine can protect against morbidity, mortality, and virus shedding in response to a lethal challenge of the highly pathogenic clade 2.3.4.4 H5N1 virus.
Pseudovirus-Based Neutralization Assay
In the current experiments, we found that even low HI antibody titers against challenge viruses in the rBacH5Con5Mut-vaccinated chickens could provide full protection. The neuralization antibody titer is applied to further evaluate the immunogenicity of our vaccine candidate. The H5N1 HPAI virus must be handled in biosafety level 3 containment laboratories. To obviate this issue, one possible alternative is to use pseudovirus to replace wild type viruses. The pseudovirus lacks certain gene sequences of the parent virulent virus, and therefore can be handled in biosafety level 2 laboratories. To this end, we performed the H5N1 (14079) pseudovirus-based neutralization test to detect serum neutralization antibody levels against influenza H5N1 (14079). The serum antibody titers were measured at 21 dpv. The average pVN antibody titers were 151 and 3241 respectively in rBacH5Con5Mut vaccination and Re-6-7-8 vaccination group (Figure 3b). Although the pVN titer is lower in rBacH5Con5Mut vaccination group, it still can provide 100% protection against 14079 virus challenging.
The rBacH5Con5Mut Vaccine Was Cross-Protective against Clade 2.3.2.1 H5N1 and Clade 7.2 H5N2 Viruses
We also evaluated the efficacy of rBacH5Con5Mut against H5N1 A/Duck/Guangdong/383/2008 (clade 2.3.2.1) and H5N2 A/Duck/Shandong/147/2013 (clade 7.2) H5 viruses, given that these strains are widely circulating in China. The Re-6-7-8 vaccine-immunized and PBS-treated groups were included as controls, and the experiment was performed as described in the previous section.
In the efficacy study challenged with A/Duck/Guangdong/383/2008 (H5N1), HI test result showed that the immune response to the rBacH5 antigen and the challenge virus on day 21 post-immunization was consistent with the response recorded for the vaccine batch in the previous experiment (Table 1). Sera from the unvaccinated controls demonstrated HI titers of 4 log2 (three chickens), 3 log2 (six chickens) and 2 log2 (seven chickens), which is considered negative, against the antigens. The average HI titers induced by rBacH5Con5Mut against the rBacH5 antigen and challenge virus were 8.0 log2 and 2.1 log2 respectively. In the Re-6-7-8 vaccination group, HI titers of 5.8 log2 and 5.7 log2 were recorded against the challenge virus and rBacH5 antigen respectively. Infection with the clade 2.3.2.1 virus killed all chickens in PBS control group by day 4 post-infection. In contrast, all vaccinated chickens survived after the virus challenge without showing any clinical signs (Figure 3c). Furthermore, the virus isolation test showed that vaccination with rBacH5Con5Mut and Re-6-7-8 prevented both cloacal shedding and oropharyngeal shedding of the clade 2.3.2.1 virus at 5 dpc (Table 1).
Next, we evaluated the efficacy of rBacH5Con5mut against the A/Duck/Shandong/147/2013 (H5N2) virus, which belongs to clade 7.2. Serological testing results showed that the HI titers of the unvaccinated controls against the challenge virus and rBacH5 antigen were 0 log2 and 3.3 log2 respectively. On the other hand, the HI titers of the sera from the rBacH5Con5Mut vaccination group were 8.7 log2 and 1.6 log2 against the vaccine strain and challenge strain respectively, while those of the Re-6-7-8 vaccination group were 6.2 log2 and 6.3 log2. In the unvaccinated group, nine chickens died within 6 dpc, and one bird was sick starting at 4 dpc and died at 14 dpc (Figure 3c). All rBacH5Con5Mut-vaccinated chickens survived the challenge infection without showing any clinical signs during the observation period (Figure 3d). Virus isolation test showed oropharyngeal shedding for one chicken and cloacal shedding for six chickens at 5 dpc. Likewise, virus shedding was observed in the oropharyngeal swab of one chicken from the Re-6-7-8 vaccination group as well (Table 1).
In summary, these results suggested that the rBacH5Con5Mut vaccine can provide cross-protection against lethal challenge of clade 2.3.2.1 and clade 7.2 H5 viruses and prevent shedding of clade 2.3.2.1 prevalent strain. Overall, this novel vaccine candidate exhibits a broad protection profile against HPAI viruses of different clades.
Protective Efficacy of rBacH5Con5Mut Vaccine Delivered to One-Day-Old SPF Chickens against 2.3.4.4 Clade H5N1 Virus
Hatchery immunization is crucial to prevent virus outbreaks in poultry farms. To determine whether rBacH5Con5Mut can be used in hatchery, we immunized a group of one-day-old SPF chickens with 0.3 ml of the rBacH5Con5Mut formulated vaccine product and challenged them three weeks later with 106 EID50 of 14079 H5N1 strain. Chickens injected with PBS were included as the non-immunized control group. Serum samples were collected three weeks post-immunization, and the titers of serum antibodies were measured against rBacH5 and 14079 challenge virus antigens by the HI assay. At week 3 post-immunization, all the chickens immunized with the rBacH5Con5Mut vaccine mounted a positive HI antibody response against rBacH5 and 14079 standard antigens with mean titers of 9.1 log2 and 2.5 log2 respectively (Table 2). In contrast, no HI titers were detected in the non-immunized chickens during the entire observation period. Thus, the rBacH5Con5Mut vaccine candidate induced a positive HI antibody response in one-day-old chickens.
After lethal challenge, all chickens were monitored for 14 days for clinical signs and mortality. The non-immunized chickens showed the typical clinical signs, such as purple comb, hemorrhage of foot scale and palpebral edema, and all birds died within two days, suggesting that 14079 virus infection is highly lethal to these chickens. However, none of the chickens immunized with the rBacH5Con5Mut vaccine showed any clinical symptoms during the observation period, and the survival rate was 100% (Figure 4).
To monitor virus shedding, the oropharyngeal and cloacal swabs were collected at 5 dpc for virus isolation. No shedding could be detected in the immunized group (Table 2). Taken together, these results indicated that the rBacH5Con5Mut vaccine provided complete protection against a lethal challenge of H5N1 virus and virus shedding when administrated to one-day-old SPF chickens.
Discussion
In this study, we developed a consensus H5 HA sequence-based subunit H5 avian influenza vaccine (rBacH5Con5Mut) and comprehensively analyzed its immunogenicity and efficacy in chickens on the basis of serum HI antibody titers, pseudovirus VN antibodies, and cross-protection against lethal challenge of three heterologous subclades (2.3.4.4, 2.3.2.1 and 7.2) of H5 HPAI viruses. The vaccine based on a rationally designed consensus H5 HA antigen can induce broad-spectrum immunity against different clades of H5 avian influenza viruses. By encoding the most conserved residues found in a selected population, the consensus sequences can effectively target multiple clade viruses with different mutation patterns. In contrast, specific virus strain sequence-based subunit vaccines provide only strain-restricted immunity. Consensus sequence-based vaccines have been extensively investigated for eliciting broadly reactive immune responses against different pathogens, including influenza virus [21,32,33], chikungunya virus [34], hepatitis B virus [35], hepatitis C virus [36], HIV-1 [37] and Middle East respiratory virus [38].
The entire ORF of the consensus HA sequence was obtained after aligning from 150 H5 strains isolated between 2005 and 2013. Phylogenetic analysis results showed that this consensus H5 HA belonged to clade 2.3.4.1. The Ser120Asn mutation was added to match the HA sequences of H5N1 HPAI viruses isolated from both humans and poultry in 2003 in order to develop a vaccine against potential pandemics of H5 HPAI viruses. Furthermore, serine at amino acid residue 223 was substituted with asparagine in order to enhance the antigenicity of the vaccine candidate in chickens [39]. Replacement of Ser with Asn will not introduce an additional N-glycosylation site based on the online glycosylation prediction result. It needs to be noted that, the Re-6-7-8 commercial product is a trivalent product and produced by using three antigens while our rBacH5Con5Mut vaccine is generated by using a single antigen. Despite using only a single HA, the efficacy of our subunit vaccine was comparable to that conferred by the inactivated trivalent whole virus vaccine (Re-6-7-8).
A broad-spectrum protective immune response, which is defined as protection against different H5 subtype virus, has been previously reported in ferret and murine models used for the development of vaccines for humans [40,41,42]. In addition, the M2 ectodomain (M2e) has also been evaluated as a broad-spectrum influenza vaccine candidate in chickens [43]. Nevertheless, its clinical application was restricted due to the lack of an appropriate method to evaluate the immune response elicited by the M2e antigens. The HA stalk region has also been used as a target antigen for the development of a universal vaccine for broad-spectrum protection [44].
HI or VN activity is generally considered as an immune correlate for efficacy of influenza vaccines. Interestingly, the HI titers against the challenge viruses were rather low in the rBacH5Con5Mut vaccination group, irrespective of the challenge virus. However, higher HI titers were observed in response to the challenge viruses after Re-6-7-8 vaccination, which can be attributed to the exact antigenic match between these viruses and the Re-6-7-8 vaccine. Spackman et al. found that the HI antibody titers are not always predictive of the protection from challenge infection or viral shedding [45]. In some cases, adequate protection could be observed in the presence of low HI titers [46]. On the other hand, some studies have also reported a clear correlation between HI titer and the level of protection [47]. The exact mechanism underlying the broad-spectrum protective effects of rBacH5Con5Mut is not fully understood. One plausible explanation could be that the exposed epitopes of the recombinant HA are readily recognized by the immune system, resulting in an appropriate broad-spectrum immune response.
Besides HI titers, the neutralizing antibody titers more closely reflect the ability of the antibodies to inhibit viral replication and propagation [48]. In our study, the H5N1 pseudovirus was used to evaluate the presence of neutralizing antibodies. The correlation of measurements performed using this assay was very similar with microneutralization assays using live virus [49,50]. The pVN test results showed that the pVN titer is also very low in rBacH5Con5Mut vaccination group although it can provide 100% protection. This result reinforces the complexity of the immune response required to provide full protection after challenge with antigenic and genetically diverse, rapid, and systemic replicating HPAIVs. Similar results with a lack of correlation of the neutralizing antibody data with protection have been reported in humans against H5N1 infection [51,52]. Recently, Hu et al. also found that the HA-binding non-neutralizing antibodies, but not HI or VN antibodies contribute to the protection against H7N9 infection in chickens and total binding antibody detection can be used as a key analytical method to correlate with protection [53]. This may warrant the necessity for a different assay to evaluate the potency of this vaccine, such as the measurement of H5-specific IgG antibody. The presence of heterosubtypic protection in the absence of strong neutralizing antibodies also indicates the possibility of cell mediated immune (CMI) responses in providing cross-protection.
We also explored the feasibility of vaccination of one-day-old chickens because hatchery vaccination is important in the field, and it has several advantages. First, commercially used vaccination equipment designed for one-day-old chickens can be used directly in the hatchery. Second, given that there are fewer hatcheries compared to poultry farms, vaccination programs in the hatcheries can be more easily conducted, controlled and centralized. Finally, hatchery vaccination allows early onset of immunity in the chickens. The HVT vectored vaccine [54] and fowlpox vectored vaccine [55] have been successfully used in hatcheries. In our study, the rBacH5Con5Mut vaccine was able to confer full protection against H5N1 virus challenge when administered to one-day-old SPF chickens, indicating that it has the potential to be employed in hatcheries as well. Nevertheless, it remains to be ascertained whether rBacH5Con5Mut vaccine can be effective in one-day-old commercial chickens with maternally derived antibodies. The impact of the maternally derived antibodies on efficacy needs to be further evaluated.
In conclusion, by a single dose vaccination, the rBacH5Con5Mut vaccine provided broad-spectrum protection against lethal challenge of three different clades of H5 HPAIV, efficiently prevented or reduced virus shedding, and was also efficacious when applied in one-day-old SPF chickens, indicating its potential for mass vaccination in hatchery. In this study, we only evaluated the efficacy of rBacH5Con5Mut on SPF chickens, the protection and vaccination program in commercial maternally derived antibody positive chickens should be furthered investigated. Furthermore, as a subunit vaccine produced in the baculovirus expression system [27], the ability of rBacH5Con5Mut vaccine to differentiate infected from vaccinated animals shall be investigated. Overall, this rationally designed vaccine candidate appears to fulfill the criteria for a broad protective H5 avian influenza vaccine and could be a useful tool in combating avian influenza virus outbreaks in the future.
References
- Smith, G. J. D., Donis, R. O. & Group, W. H. O. O. for A. H. and A. O. (WHO/OIE/FAO) H. E. W. Nomenclature updates resulting from the evolution of avian influenza A(H5) virus clades 2.1.3.2a, 2.2.1, and 2.3.4 during 2013–2014. Influ. Other Respir. Viruses 9, 271–276 (2015). [CrossRef]
- Claes, F., Morzaria, S. P. & Donis, R. O. Emergence and dissemination of clade 2.3.4.4 H5Nx influenza viruses—how is the Asian HPAI H5 lineage maintained. Curr. Opin. Virol. 16, 158–163 (2016).
- Sun, H. et al. Characterization of clade 2.3.4.4 highly pathogenic H5 avian influenza viruses in ducks and chickens. Vet. Microbiol. 182, 116–122 (2016). [CrossRef]
- SWAYNE, D. E., PAVADE, G., HAMILTON, K., VALLAT, B. & MIYAGISHIMA, K. Assessment of national strategies for control of high-pathogenicity avian influenza and lowpathogenicity notifiable avian influenza in poultry, with emphasis on vaccines and vaccination: -EN- -FR- Évaluation des stratégies nationales de lutte contre l’influenza aviaire hautement pathogène et l’influenza aviaire faiblement pathogène à déclaration obligatoire chez les volailles, et plus particulièrement du recours aux vaccins et à la vaccination -ES- Evaluación de estrategias nacionales de lucha contra la influenza aviar altamente patógena y la influenza aviar levemente patógena de declaración obligatoria en aves de corral, haciendo hincapié en las vacunas y la vacunación. Rev. Sci. Tech. l’OIE 30, 839–870 (2011).
- Swayne, D. E. Impact of Vaccines and Vaccination on Global Control of Avian Influenza. Avian Diseases 56, 818–828 (2012). [CrossRef]
- Chen, M.-W. et al. A consensus–hemagglutinin-based DNA vaccine that protects mice against divergent H5N1 influenza viruses. Proc. Natl. Acad. Sci. 105, 13538–13543 (2008). [CrossRef]
- Bi, Y. et al. Highly pathogenic avian influenza H5N1 Clade 2.3.2.1c virus in migratory birds, 2014–2015. Virol. Sin. 31, 300–305 (2016). [CrossRef]
- Tian, H. et al. Spatial, temporal and genetic dynamics of highly pathogenic avian influenza A (H5N1) virus in China. BMC Infect. Dis. 15, 54 (2015). [CrossRef]
- Li, C., Bu, Z. & Chen, H. Avian influenza vaccines against H5N1 ‘bird flu’ Trends in Biotechnology 32, 147–156 (2014).
- Shi, J., Zeng, X., Cui, P., Yan, C. & Chen, H. Alarming situation of emerging H5 and H7 avian influenza and effective control strategies. Emerg. Microbes Infect. 12, 2155072 (2023).
- Ge, J. et al. Newcastle Disease Virus-Based Live Attenuated Vaccine Completely Protects Chickens and Mice from Lethal Challenge of Homologous and Heterologous H5N1 Avian Influenza Viruses. J. Virol. 81, 150–158 (2007). [CrossRef]
- Jiang, Y. et al. Enhanced protective efficacy of H5 subtype avian influenza DNA vaccine with codon optimized HA gene in a pCAGGS plasmid vector. Antivir. Res. 75, 234–241 (2007). [CrossRef]
- Prabakaran, M. et al. Neutralizing Epitopes of Influenza Virus Hemagglutinin: Target for the Development of a Universal Vaccine against H5N1 Lineages. J. Virol. 84, 11822–11830 (2010). [CrossRef]
- Yewdell, J. W., Bennink, J. R., Smith, G. L. & Moss, B. Influenza A virus nucleoprotein is a major target antigen for cross-reactive anti-influenza A virus cytotoxic T lymphocytes. Proc. Natl. Acad. Sci. 82, 1785–1789 (1985).
- Ekiert, D. C. et al. Antibody Recognition of a Highly Conserved Influenza Virus Epitope. Science 324, 246–251 (2009). [CrossRef]
- Tan, G. S. et al. A Pan-H1 Anti-Hemagglutinin Monoclonal Antibody with Potent Broad-Spectrum Efficacy In Vivo. J. Virol. 86, 6179–6188 (2012). [CrossRef]
- Kamlangdee, A., Kingstad-Bakke, B., Anderson, T. K., Goldberg, T. L. & Osorio, J. E. Broad Protection against Avian Influenza Virus by Using a Modified Vaccinia Ankara Virus Expressing a Mosaic Hemagglutinin Gene. J. Virol. 88, 13300–13309 (2014).
- Wang, T. T. et al. Vaccination with a synthetic peptide from the influenza virus hemagglutinin provides protection against distinct viral subtypes. Proc. Natl. Acad. Sci. 107, 18979–18984 (2010). [CrossRef]
- Sautto, G. A. & Ross, T. M. Hemagglutinin consensus-based prophylactic approaches to overcome influenza virus diversity. Vet. Ital. 55, 195–201 (2019).
- Ma, N. et al. Development of an mRNA vaccine against a panel of heterologous H1N1 seasonal influenza viruses using a consensus hemagglutinin sequence. Emerg. Microbes Infect. 12, 2202278 (2023). [CrossRef]
- Giles, B. M. & Ross, T. M. A computationally optimized broadly reactive antigen (COBRA) based H5N1 VLP vaccine elicits broadly reactive antibodies in mice and ferrets. Vaccine 29, 3043–3054 (2011).
- Wang, G., Yin, R., Zhou, P. & Ding, Z. Combination of the immunization with the sequence close to the consensus sequence and two DNA prime plus one VLP boost generate H5 hemagglutinin specific broad neutralizing antibodies. PLoS ONE 12, e0176854 (2017).
- Zhou, H. et al. Sequential immunization with consensus influenza hemagglutinins raises cross-reactive neutralizing antibodies against various heterologous HA strains. Vaccine 35, 305–312 (2017). [CrossRef]
- Song, J.-M., Rooijen, N. V., Bozja, J., Compans, R. W. & Kang, S.-M. Vaccination inducing broad and improved cross protection against multiple subtypes of influenza A virus. Proc. Natl. Acad. Sci. 108, 757–761 (2011).
- Harada, Y. et al. Inactivated and adjuvanted whole-virion clade 2.3.4 H5N1 pre-pandemic influenza vaccine possesses broad protective efficacy against infection by heterologous clades of highly pathogenic H5N1 avian influenza virus in mice. Vaccine 29, 8330–8337 (2011). [CrossRef]
- Wu, C.-Y. et al. A VLP Vaccine Induces Broad-Spectrum Cross-Protective Antibody Immunity against H5N1 and H1N1 Subtypes of Influenza A Virus. PLoS ONE 7, e42363 (2012). [CrossRef]
- Cavalcanti, M. O. et al. A genetically engineered H5 protein expressed in insect cells confers protection against different clades of H5N1 highly pathogenic avian influenza viruses in chickens. Avian Pathol. 46, 224–233 (2017). [CrossRef]
- Zeng, X. et al. Protective Efficacy of the Inactivated H5N1 Influenza Vaccine Re-6 Against Different Clades of H5N1 Viruses Isolated in China and the Democratic People’s Republic of Korea. Avian Diseases 60, 238–240 (2015). [CrossRef]
- Tian, G., Zeng, X., Li, Y., Shi, J. & Chen, H. Protective Efficacy of the H5 Inactivated Vaccine Against Different Highly Pathogenic H5N1 Avian Influenza Viruses Isolated in China and Vietnam. Avian Diseases 54, 287–289 (2010).
- Zeng, X. et al. Protective Efficacy of an H5N1 Inactivated Vaccine Against Challenge with Lethal H5N1, H5N2, H5N6, and H5N8 Influenza Viruses in Chickens. Avian Diseases 60, 253–255 (2016). [CrossRef]
- Yang, D.-G. et al. Avian Influenza Virus Hemagglutinin Display on Baculovirus Envelope: Cytoplasmic Domain Affects Virus Properties and Vaccine Potential. Mol. Ther. 15, 989–996 (2007). [CrossRef]
- Laddy, D. J. et al. Immunogenicity of novel consensus-based DNA vaccines against avian influenza. Vaccine 25, 2984–2989 (2007). [CrossRef]
- Carter, D. M. et al. Design and Characterization of a Computationally Optimized Broadly Reactive Hemagglutinin Vaccine for H1N1 Influenza Viruses. J. Virol. 90, 4720–4734 (2016). [CrossRef]
- Muthumani, K. et al. Immunogenicity of novel consensus-based DNA vaccines against Chikungunya virus. Vaccine 26, 5128–5134 (2008). [CrossRef]
- Obeng-Adjei, N. et al. Synthetic DNA immunogen encoding hepatitis B core antigen drives immune response in liver. Cancer Gene Ther. 19, 779–787 (2012). [CrossRef]
- Latimer, B. et al. A polyantigenic genotype 1a/1b consensus hepatitis C virus DNA vaccine induces broadly reactive HCV-specific cellular immune responses in both mice and non-human primates (VAC7P.985). The Journal of Immunology (2014).
- Yan, J. et al. Immunogenicity of a novel engineered HIV-1 clade C synthetic consensus-based envelope DNA vaccine. Vaccine 29, 7173–7181 (2011). [CrossRef]
- Muthumani, K. et al. A synthetic consensus anti–spike protein DNA vaccine induces protective immunity against Middle East respiratory syndrome coronavirus in nonhuman primates. Sci. Transl. Med. 7, 301ra132 (2015). [CrossRef]
- Hoffmann, E., Lipatov, A. S., Webby, R. J., Govorkova, E. A. & Webster, R. G. Role of specific hemagglutinin amino acids in the immunogenicity and protection of H5N1 influenza virus vaccines. Proc. Natl. Acad. Sci. 102, 12915–12920 (2005).
- Forrest, H. L. et al. Single- and multiple-clade influenza A H5N1 vaccines induce cross protection in ferrets. Vaccine 27, 4187–4195 (2009). [CrossRef]
- Wang, T. T. et al. Vaccination with a synthetic peptide from the influenza virus hemagglutinin provides protection against distinct viral subtypes. Proc. Natl. Acad. Sci. 107, 18979–18984 (2010). [CrossRef]
- He, F., Prabakaran, M., kumar, S. R., Tan, Y. & Kwang, J. Monovalent H5 vaccine based on epitope-chimeric HA provides broad cross-clade protection against variant H5N1 viruses in mice. Antivir. Res. 105, 143–151 (2014).
- Tang, Y. et al. Chimaeric VP2 proteins from infectious bursal disease virus containing the N-terminal M2e of H9 subtype avian influenza virus induce neutralizing antibody responses to both viruses. Avian Pathol. 42, 260–267 (2013). [CrossRef]
- Bernstein, D. I. et al. Immunogenicity of chimeric haemagglutinin-based, universal influenza virus vaccine candidates: interim results of a randomised, placebo-controlled, phase 1 clinical trial. Lancet Infect. Dis. 20, 80–91 (2020). [CrossRef]
- Spackman, E. et al. Development of a Real-Time Reverse Transcriptase PCR Assay for Type A Influenza Virus and the Avian H5 and H7 Hemagglutinin Subtypes. J. Clin. Microbiol. 40, 3256–3260 (2002). [CrossRef]
- Kumar, M., Chu, H.-J., Rodenberg, J., Krauss, S. & Webster, R. G. Association of Serologic and Protective Responses of Avian Influenza Vaccines in Chickens. Avian Diseases 51, 481–483 (2007).
- Liu, M. et al. Preparation of a standardized, efficacious agricultural H5N3 vaccine by reverse genetics. Virology 314, 580–590 (2003). [CrossRef]
- Temperton, N. J. et al. A sensitive retroviral pseudotype assay for influenza H5N1-neutralizing antibodies. Influ. Other Respir. Viruses 1, 105–112 (2007). [CrossRef]
- Hyseni, I. et al. Characterisation of SARS-CoV-2 Lentiviral Pseudotypes and Correlation between Pseudotype-Based Neutralisation Assays and Live Virus-Based Micro Neutralisation Assays. Viruses 12, 1011 (2020). [CrossRef]
- Tolah, A. M. K. et al. Evaluation of a Pseudovirus Neutralization Assay for SARS-CoV-2 and Correlation with Live Virus-Based Micro Neutralization Assay. Diagnostics 11, 994 (2021). [CrossRef]
- Rowe, T. et al. Detection of Antibody to Avian Influenza A (H5N1) Virus in Human Serum by Using a Combination of Serologic Assays. J. Clin. Microbiol. 37, 937–943 (1999). [CrossRef]
- Schultsz, C. et al. Prevalence of Antibodies against Avian Influenza A (H5N1) Virus among Cullers and Poultry Workers in Ho Chi Minh City, 2005. PLoS ONE 4, e7948 (2009). [CrossRef]
- Hu, Z. et al. Hemagglutinin-Specific Non-neutralizing Antibody Is Essential for Protection Provided by Inactivated and Viral-Vectored H7N9 Avian Influenza Vaccines in Chickens. Front. Vet. Sci. 6, 482 (2020). [CrossRef]
- Reemers, S., Verstegen, I., Basten, S., Hubers, W. & Zande, S. van de. A broad spectrum HVT-H5 avian influenza vector vaccine which induces a rapid onset of immunity. Vaccine 39, 1072–1079 (2021).
- BUBLOT, M. et al. Development and Use of Fowlpox Vectored Vaccines for Avian Influenza. Ann. N. York Acad. Sci. 1081, 193–201 (2006). [CrossRef]
Figure 1.
Design and characterization of rBacH5Con5Munt recombinant virus. (a) A total of 150 H5NX HA sequences were obtained from NCBI database and aligned by Vector NTI software. A consensus H5NX HA sequence was generated and named as H5Con5Mut. (b) Phylogenetic analysis of HA sequences of H5Nx isolated from poultry, location of consensus H5Con5Mut HA and challenge virus used in this study in the tree. (c) The 3D structure model based on consensus HA protein is shown and location of the S120N and S223N mutations are highlighted in red on the structure. (d) H5 HA expression from rBacH5Con5Mut with (R) or without (NR) 10X reducing agent were confirmed by WB. (e) Dual IFA staining of H5 HA and Gp64 by corresponding antibodies. (f) Expression of H5 HA was confirmed by HA assay.
Figure 1.
Design and characterization of rBacH5Con5Munt recombinant virus. (a) A total of 150 H5NX HA sequences were obtained from NCBI database and aligned by Vector NTI software. A consensus H5NX HA sequence was generated and named as H5Con5Mut. (b) Phylogenetic analysis of HA sequences of H5Nx isolated from poultry, location of consensus H5Con5Mut HA and challenge virus used in this study in the tree. (c) The 3D structure model based on consensus HA protein is shown and location of the S120N and S223N mutations are highlighted in red on the structure. (d) H5 HA expression from rBacH5Con5Mut with (R) or without (NR) 10X reducing agent were confirmed by WB. (e) Dual IFA staining of H5 HA and Gp64 by corresponding antibodies. (f) Expression of H5 HA was confirmed by HA assay.
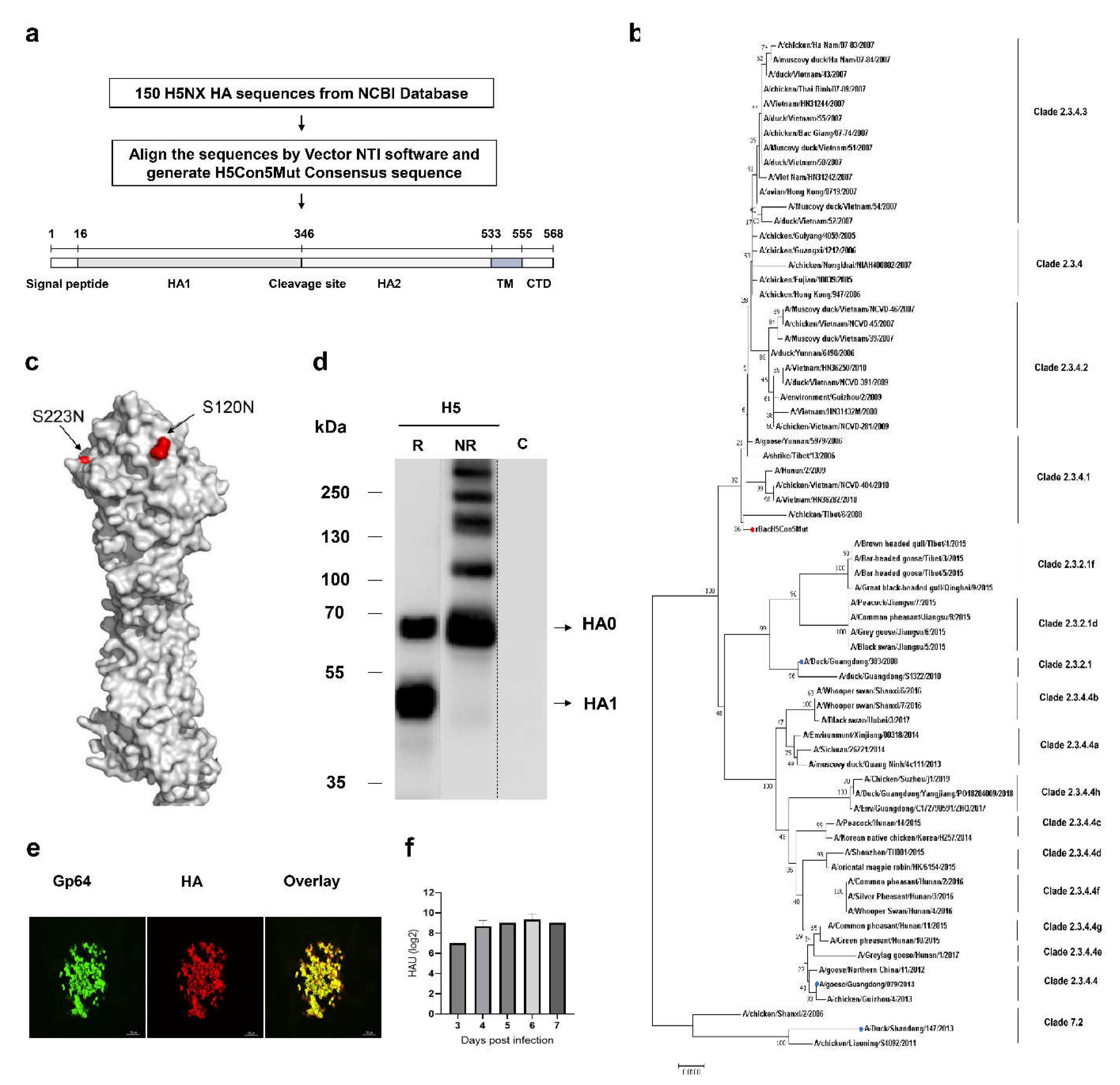
Figure 2.
Genetic stability of HA in rBacH5Con5Mut. The rBacH5Con5Mut virus was continuously passaged in SF+ cells from P7 to P17. (a) Dual IFA staining of H5 HA and gp64 protein of rBacH5Con5Mut P7, P12 and P17 viruses infected in Sf9 cells. (b) The presence of the H5 HA gene in the recombinant baculovirus from different passages was confirmed by PCR (~1800bp). (c and d) H5 HA expression from rBacH5Con5Mut P7, P12 and P17 viruses was confirmed by WB and HA assay.
Figure 2.
Genetic stability of HA in rBacH5Con5Mut. The rBacH5Con5Mut virus was continuously passaged in SF+ cells from P7 to P17. (a) Dual IFA staining of H5 HA and gp64 protein of rBacH5Con5Mut P7, P12 and P17 viruses infected in Sf9 cells. (b) The presence of the H5 HA gene in the recombinant baculovirus from different passages was confirmed by PCR (~1800bp). (c and d) H5 HA expression from rBacH5Con5Mut P7, P12 and P17 viruses was confirmed by WB and HA assay.
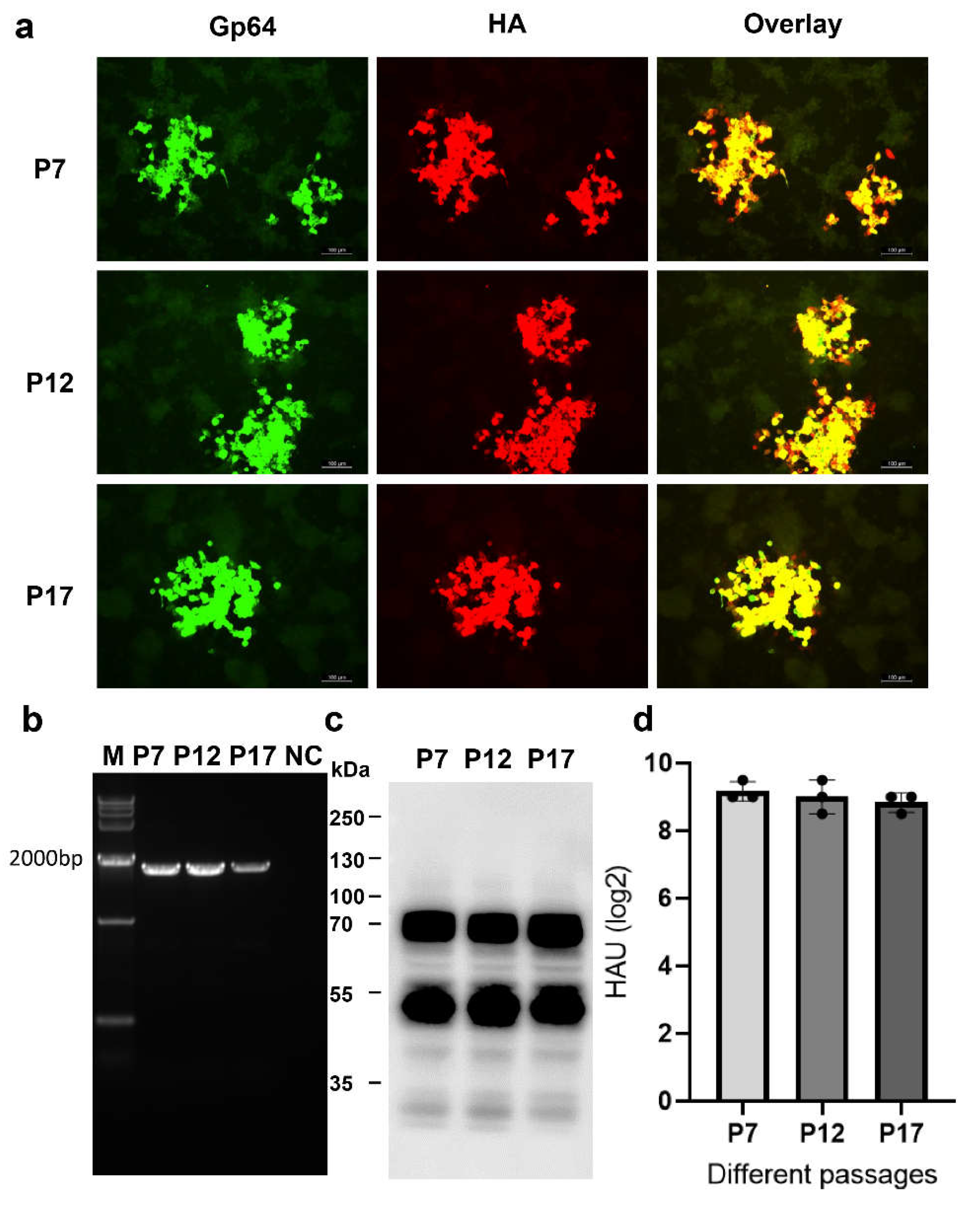
Figure 3.
Survival rate of the chickens intranasally challenged with different strains of H5 highly pathogenic avian influenza virus. Three-week-old SPF chickens were randomly divided into three groups: 10 chickens were immunized with rBacH5Con5Mut, 10 chickens were immunized with the inactivated whole virus Re-6-7-8 vaccine, and 10 chickens were injected with PBS (challenge control group). At 21 dpv, the chickens were infected with (a) clade 2.3.4.4 H5N1 HPAI, (c) clade 2.3.2.1 H5N2 HPAI and (d) clade 7.2 H5N2 HPAI viruses. Clinical signs were observed for 14 days. (b) pVN titers of the chicken serum were tested in 2.3.4.4 challenging study by using 14079-pseudovirus.
Figure 3.
Survival rate of the chickens intranasally challenged with different strains of H5 highly pathogenic avian influenza virus. Three-week-old SPF chickens were randomly divided into three groups: 10 chickens were immunized with rBacH5Con5Mut, 10 chickens were immunized with the inactivated whole virus Re-6-7-8 vaccine, and 10 chickens were injected with PBS (challenge control group). At 21 dpv, the chickens were infected with (a) clade 2.3.4.4 H5N1 HPAI, (c) clade 2.3.2.1 H5N2 HPAI and (d) clade 7.2 H5N2 HPAI viruses. Clinical signs were observed for 14 days. (b) pVN titers of the chicken serum were tested in 2.3.4.4 challenging study by using 14079-pseudovirus.
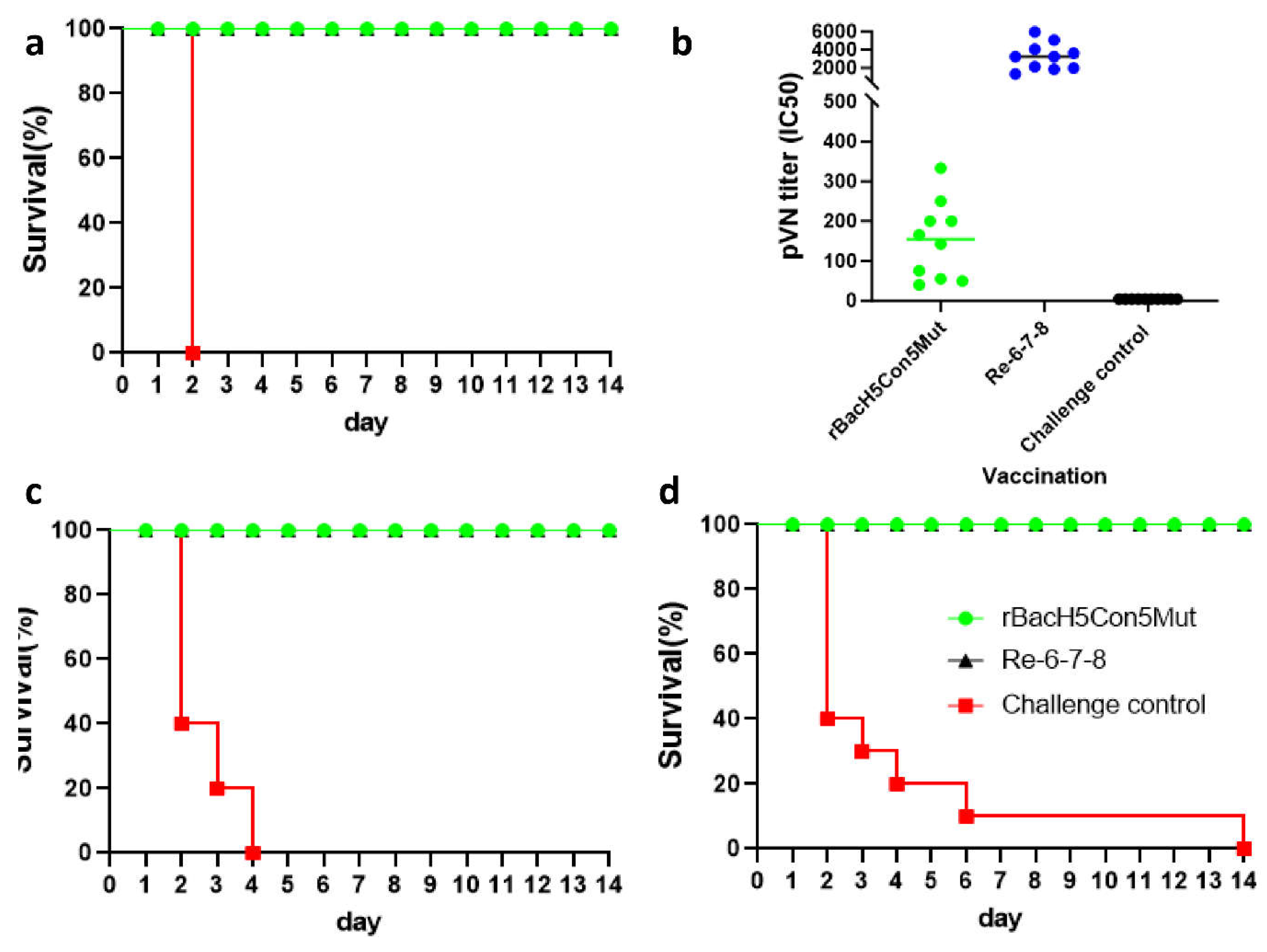
Figure 4.
Survival rate of one-day-old SPF chickens challenged with the highly pathogenic H5 avian influenza virus. Two groups of one-day-old SPF chickens were immunized with the rBacH5Con5Mut vaccine or PBS (control group) respectively. At 21 dpv, the chickens were infected with clade 2.3.4.4 H5N1 HPAIV. Clinical signs were observed for 14 days.
Figure 4.
Survival rate of one-day-old SPF chickens challenged with the highly pathogenic H5 avian influenza virus. Two groups of one-day-old SPF chickens were immunized with the rBacH5Con5Mut vaccine or PBS (control group) respectively. At 21 dpv, the chickens were infected with clade 2.3.4.4 H5N1 HPAIV. Clinical signs were observed for 14 days.
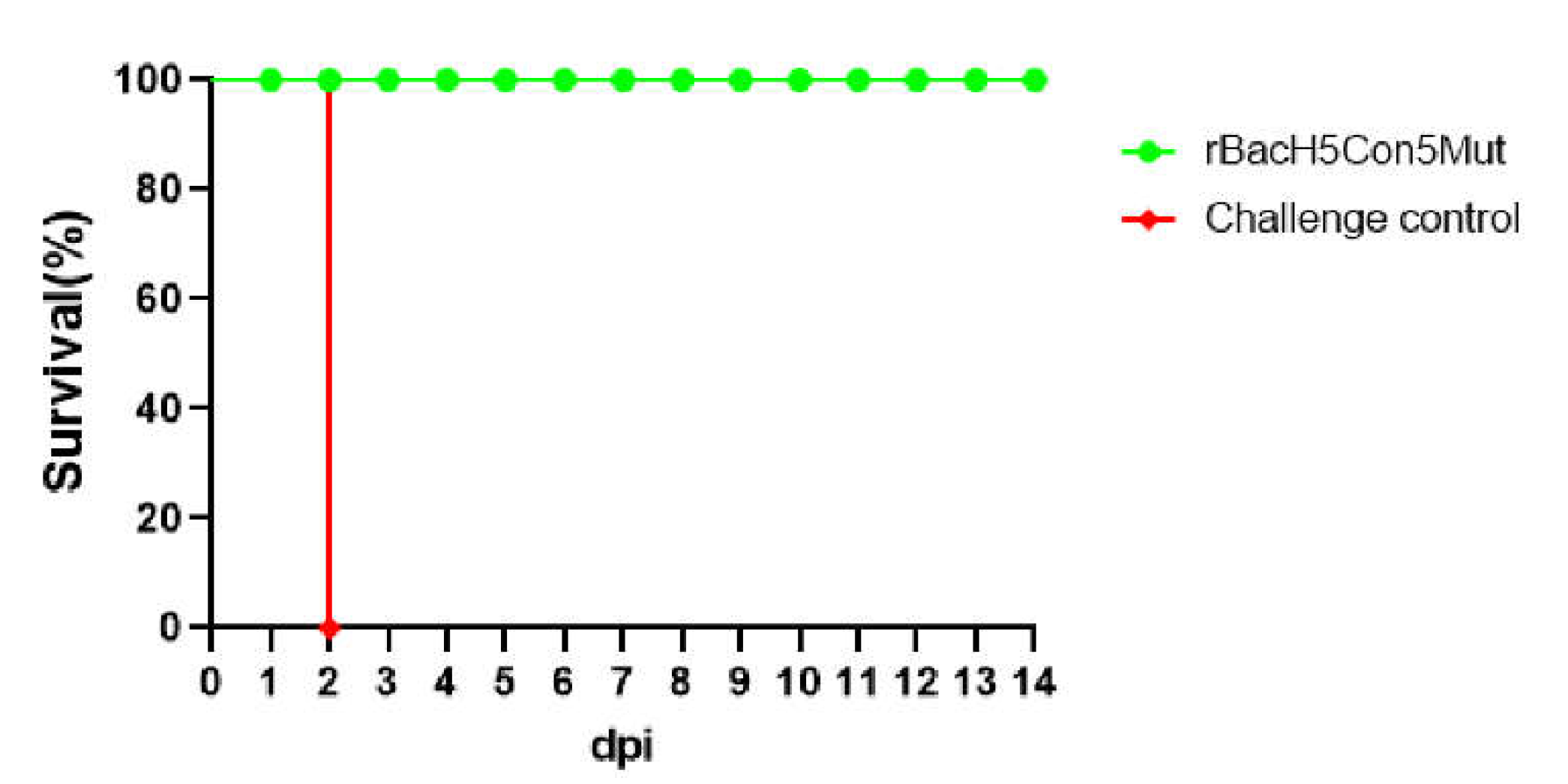

Table 1.
Protective efficacy of the rBacH5Con5Mut vaccine against different highly pathogenic H5 AIV strains in 21-day-old chickens.
Table 1.
Protective efficacy of the rBacH5Con5Mut vaccine against different highly pathogenic H5 AIV strains in 21-day-old chickens.
| Experiment | Vaccination | Challenge virus | Clade | HI titer with different antigens (log2)a | Survival/total | Virus shedding from chickens at five days post challenge | |||
| rBacH5 | CVb | Oropharyngeal | Cloacal | ||||||
| 1 | rBacH5Con5Mut | A/Goose/Guangdong/079/2013 (H5N1, 14079) | 2.3.4.4 | 9.1 | 3.6 | 10/10 | 0/10 | 0/10 | |
| Re6-7-8 | 6.5 | 8 | 10/10 | 0/10 | 0/10 | ||||
| Control | 3.3 | 0 | 0/10 | N/ac | N/a | ||||
| 2 | rBacH5Con5Mut | A/Duck/Guangdong/383/2008 (H5N1, 383) | 2.3.2.1 | 8.0 | 2.1 | 10/10 | 0/10 | 0/10 | |
| Re6-7-8 | 5.7 | 5.8 | 10/10 | 0/10 | 0/10 | ||||
| Control | 3.2 | 0 | 0/10 | N/a | N/a | ||||
| 3 | rBacH5Con5Mut | A/Duck/Shandong/147/2013 (H5N2, 147) | 7.2 | 8.7 | 1.6 | 10/10 | 1/10 | 6/10 | |
| Re6-7-8 | 6.2 | 6.3 | 10/10 | 1/10 | 0/10 | ||||
| Control | 3.3 | 0 | 0/10 | 2/2 | 2/2 | ||||
a The HI titers were measured using the homologous rBacH5Con5Mut virus or the challenge viruses with the antisera collected three weeks after vaccination. b Challenge virus. c All of the chickens in the group died before day 5.
Table 2.
Protective efficacy of the rBacH5Con5Mut vaccine against highly pathogenic H5 AIV strains in one-day-old chickens.
Table 2.
Protective efficacy of the rBacH5Con5Mut vaccine against highly pathogenic H5 AIV strains in one-day-old chickens.
| Vaccination | Challenge virus | Clade | Days of age | HI titer with different antigens (log2)a | Survival/total | Virus shedding from chickens at five days post challenge | |||
| rBacH5 | CVb | Oropharyngeal | Cloacal | ||||||
| rBacH5Con5Mut | A/Goose/Guangdong/079/2013 (H5N1, 14079) | 2.3.4.4 | 1 | 9.1 | 2.5 | 10/10 | 0/10 | 0/10 | |
| Challenge Control | 1 | 2 | 0 | 0/10 | N/ac | N/a | |||
a The HI titers were measured using the homologous rBacH5Con5Mut virus or the challenge viruses with the antisera collected three weeks after vaccination. b Challenge virus. c All of the chickens in the group died before day 5.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



