
Submitted:
17 July 2024
Posted:
17 July 2024
You are already at the latest version
Abstract
Plant activators have emerged as promising alternatives to conventional pesticides for crop disease manage-ment due to their unique mode of action. By priming the plant's innate immune system, these compounds can induce systemic acquired resistance against a broad spectrum of pathogens without directly inhibiting their proliferation. Key advantages of plant activators include prolonged defense activity, lower effective dosages, and negligible risk of developing pathogen resistance. Among the various defensive pathways tar-geted, the salicylic acid (SA) signaling cascade has been extensively explored, leading to the successful devel-opment of commercial activators like benzothiadiazole for widespread application in crop protection. While the action sites of many SA-targeting activators have been preliminarily mapped to different steps along the pathway, a comprehensive understanding of their precise mechanisms remains elusive. This review provides a historical perspective on plant activator development and outlines diverse screening strategies employed across multiple levels, from whole-plant bioassays to molecular and transgenic approaches. We expound on the intricate components, biological relevance, and regulatory circuits governing the SA pathway, while criti-cally examining the structural features, bioactivities, and proposed modes of action of classical activators such as benzothiadiazole derivatives, salicylic acid analogs, and other small molecules. Insights from field trials as-sessing their practical applicability are also discussed. Furthermore, we highlight the current status, challenges, and future prospects in the realm of SA-targeting activator development globally, with a focus on recent en-deavors in China. Collectively, this comprehensive review aims to synthesize existing knowledge and provide a roadmap for future research toward developing more potent and mechanistically understood plant activa-tors that fortify crop immunity against disease.
Keywords:
plant activator
; salicylic acid signal pathway
; systemic acquired resistance
; plant resistance
; action mechanism
1. Introduction
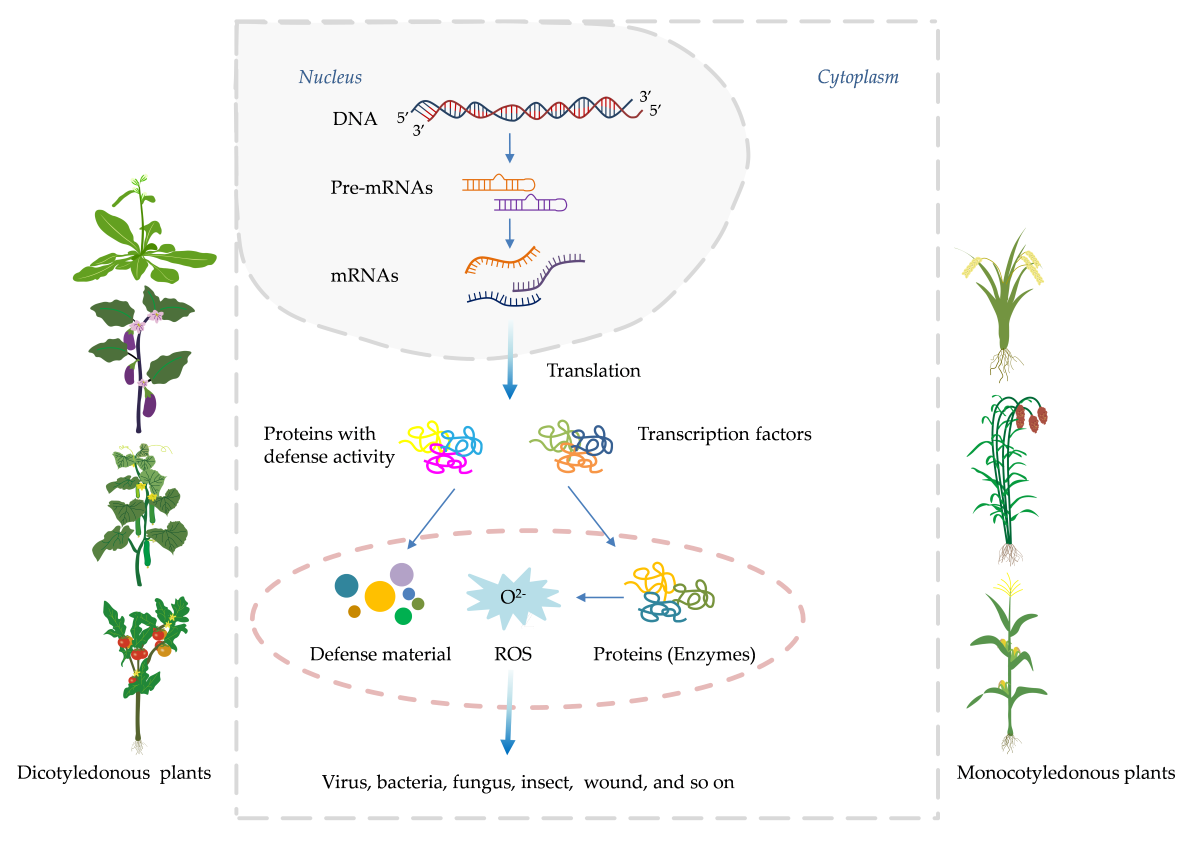
Systemic acquired resistance (SAR) represents an inducible, whole-plant immune response in which localized exposure to a pathogen or an elicitor confers long-lasting, broad-spectrum resistance against subsequent infections [1,2]. This phenomenon underscores the remarkable ability of plants to adapt and fortify their defenses in the face of potential threats [3]. At the heart of SAR lies the salicylic acid (SA) signaling cascade, a pivotal molecular pathway that orchestrates a multifaceted defense program upon activation [4,5,6]. Central to this pathway is the accumulation of pathogenesis-related proteins (PRs) and antimicrobial phytoalexins, accompanied by the potentiation of enzymes involved in oxidative burst and other frontline defense responses (Figure 1) [7,8]. Over the past decades, intensive research has gradually unraveled the intricate network architecture, key components, and regulatory mechanisms governing the SA pathway [8]. Critical players such as the harpin-binding protein (HrBP1), enhanced disease susceptibility proteins (EDS1 and EDS5), the master regulator nonexpressor of pathogenesis-related genes 1 (NPR1), WRKY transcription factors, and the PRs themselves have been identified as essential signaling hubs, amplifiers, and executors of the SA-mediated resistance response [9,10,11]. This expanding molecular blueprint has paved the way for targeted interventions aimed at priming plant immunity (Figure 2) [12]. Capitalizing on this knowledge, researchers and agrochemical companies have actively pursued the development of synthetic "plant activators" compounds capable of stimulating the SA pathway and conferring robust, broad-spectrum disease resistance without directly targeting pathogens [13].
SAR inducible, whole-plant immune response in which localized exposure to a pathogen or an elicitor confers long-lasting, broad-spectrum resistance against subsequent infections [14]. This remarkable ability of plants to "immunize" themselves against potential threats was first reported by Ross in 1961, who observed that localized infection of tobacco leaves with tobacco mosaic virus (TMV) prevented the systemic spread of the same virus upon subsequent inoculation [15]. Subsequent pioneering work by Malamy, et al. [16] and Métraux, et al. [17] demonstrated that SAR is mediated by a long-distance signal produced at the initial infection site, which travels through the vascular system and activates defense responses in distal, uninfected parts of the plant. This seminal finding paved the way for intensive research aimed at elucidating the molecular underpinnings of SAR.
A major breakthrough came when Gaffney, et al. [18] and Malamy, Carr, Klessig and Raskin [16] identified the SA as a crucial signaling molecule required for the establishment of SAR. Exogenous application of the SA or its synthetic analogs was found to be sufficient to induce SAR responses in various plant species. Concurrently, Ward, et al. [19] reported the pivotal role of PR proteins, a suite of antimicrobial proteins whose expression is tightly regulated by the SA pathway, in manifesting SAR. Further dissection of the SA signaling cascade identified key regulatory components such as the transcriptional co-activator NPR1 [20,21], which acts as a master regulator of SAR by orchestrating the expression of PR genes and other defense responses upon perception of the SA signal. Subsequent studies unveiled additional players, including the EDS1 and PAD4 lipase-like proteins [22,23], which function upstream of the SA biosynthesis, and the transcription factor family of WRKY, which acts downstream of NPR1 to regulate PR gene expression [24,25]. The elucidation of the SA-mediated SAR pathway has profound implications for crop protection strategies [26]. By harnessing our understanding of this endogenous defense mechanism, researchers have developed synthetic "plant activators" that can prime the SA pathway and confer broad-spectrum disease resistance without directly targeting pathogens [27]. This approach offers several advantages over conventional pesticides and has led to the successful commercialization of activators like benzothiadiazole for widespread use in agricultural systems [28]. These activators offer several advantages over conventional pesticides, including prolonged defense activation, lower effective dosages, reduced environmental impact, and a negligible risk of developing pathogen resistance [29]. Consequently, numerous the SA-targeting activators, such as benzothiadiazole derivatives, have been successfully commercialized and widely adopted for crop protection globally [30]. Importantly, plant activators have also served as invaluable chemical probes, facilitating the elucidation of SA pathway’s intricate mechanisms and associated physiological phenomena [31]. In this comprehensive review, we trace the historical evolution of plant activator development and elucidate the current understanding of the SA signaling pathway’s architecture and regulation. Furthermore, we critically examine the structural features, bioactivities, proposed modes of action, and field performance of classical and emerging SA-targeting activators (Figure 3). Finally, we highlight the current challenges and future prospects in this burgeoning field, with a particular emphasis on recent endeavors undertaken in China to advance the development of more potent and mechanistically understood plant activators that can fortify crop immunity against diseases.
2. Plants Receptors Trigger an Immune Response
Plants possess cell-surface pattern recognition receptors that can detect danger signals from pathogens or the host itself, triggering an immune response [32]. One subgroup of these receptors is the leucine-rich repeat receptor-like proteins (LRR-RLPs), which play a crucial role in plant immunity [33]. However, the mechanisms underlying ligand recognition and activation of LRR-RLPs have remained elusive. In a recent study, researchers reported in Nicotiana benthamiana, the crystal structure of the LRR-RLP RXEG1 which recognizes the XEG1 xyloglucanase enzyme from the pathogen Phytophthora sojae [33]. The structure revealed that RXEG1 recognizes XEG1 primarily through its amino-terminal and carboxy-terminal loop-out regions. RXEG1, a receptor-like protein in plants, recognizes the pathogen XEG1 (a glycoside hydrolase secreted by Phytophthora) through interactions primarily involving its amino-terminal and carboxy-terminal loop-out regions. This structural insight highlights the specific regions of RXEG1 that are crucial for pathogen recognition, helping to understand the molecular mechanisms behind plant immunity and pathogen evasion strategies [34]. These loops bind to the active site groove of XEG1, inhibiting its enzymatic activity and suppressing Phytophthora infection in N. benthamiana. Binding of XEG1 promotes the association of RXEG1’s LRR domain RXEG1 (LRR) with the co-receptor BAK1, also an LRR-type protein, through RXEG1 (ID) and the last four conserved LRRs [35]. This association triggers RXEG1-mediated immune responses (Figure 1). Structural comparisons of apo-RXEG1 (LRR), XEG1–RXEG1 (LRR), and XEG1–BAK1–RXEG1 (LRR) complexes showed that XEG1 binding induces conformational changes in the N-terminal region of RXEG1 and increases structural flexibility in BAK1-associating regions of RXEG1 (LRR), the RXEG1, characterized by its leucine-rich repeat domain known as RXEG1 (LRR) [36]. These changes allow RXEG1 to undergo a fold-switching mechanism, facilitating the recruitment of BAK1 (LRR). It brassinosteroid-insensitive 1-associated receptor kinase 1, is characterized by its leucine-rich repeat domain (LRR). This domain is crucial for its interactions in plant signaling pathways, particularly in immune responses and development processes. This association triggers RXEG1-mediated immune responses. These changes allow a fold-switching of RXEG1, facilitating the recruitment of brassinosteroid-insensitive 1-associated receptor kinase 1 (BAK1 (LRR), which is essential for initiating downstream signaling cascades in plant defense and development pathways. Chen et al [36] reveal a conserved mechanism of ligand-induced heterodimerization of an LRR-RLP with BAK1 and suggest a dual function for LRR-RLPs in plant immunity direct ligand recognition and co-receptor recruitment for signaling.
The study investigates the role of BIR2, also known as BAK1-interacting receptor-like kinase 2, plays a significant role in plant immune responses by interacting with BAK1 (LRR) and other proteins to regulate signaling pathways involved in defense against pathogens. Meanwhile BIR2 is a receptor-like kinase that recognizes the fungal cell wall protein AVRY567 to initiate PTI responses in N. benthamiana [37]. The E3 ubiquitin ligases SNIPER2a and SNIPER2b target BIR2 for degradation via the 26S proteasome pathway, negatively regulating PTI [38,39]. BAK1 associates with BIR2 upon AVRY567 recognition and protects BIR2 from SNIPER2a/b-mediated degradation [40]. Knock-down of BAK1 leads to enhanced degradation of BIR2 by SNIPER2a/b, compromising AVRY567-triggered PTI responses [41]. The kinase activity of BAK1 is required for its protective role against BIR2 degradation [38]. BAK1 appears to protect BIR2 by phosphorylating it, preventing SNIPER2a/b from ubiquitinating and targeting it for degradation [42]. This study uncovers a novel function for the co-receptor BAK1 in stabilizing receptor-like kinase BIR2 by protecting it from E3 ligase-mediated degradation [43]. This stabilization allows BIR2 to properly initiate and sustain pattern-triggered immune responses upon recognition of the fungal AVRY567 protein in N. benthamiana [38].
A summary of the key points regarding the past, present, and future of plant activators targeting the SA signal pathway:
Early research identified chemicals like SA, 2,6-dichloroisonicotinic acid (INA), and benzo (1, 2, 3) thiadiazole-7-carbothioic acid S-methyl ester (BTH) as activators of the SA signal pathway [44]. These compounds were found to induce SAR, a broad-spectrum resistance against pathogens, by mimicking the effects of pathogen infection [45]. Studies showed that these activators stimulate SA biosynthesis and signaling, leading to the expression of PR genes and other defense responses [46]. Current research focuses on understanding the molecular mechanisms by which these activators regulate the SA pathway and identifying new activators with improved efficacy and specificity [47]. Natural compounds like plant extracts, essential oils, and metabolites are being explored as potential the SA pathway activators with reduced environmental impact [48]. Advanced techniques like transcriptomics, proteomics, and metabolomics are being employed to elucidate the complex signaling networks and identify key components targeted by activators [49]. Activators are being tested for their ability to prime plant defenses against various biotic and abiotic stresses, leading to enhanced crop productivity and sustainability [50].
Future research aims to develop more potent and selective the SA pathway activators with minimal side effects on plant growth and development [51]. Targeted approaches like structure-based drug design and chemical genomics may facilitate the discovery of novel activators with specific modes of action [52]. Integration of activators with other defense-priming strategies, such as induced systemic resistance (ISR) and RNA interference (RNAi), could provide more robust and durable resistance against pathogens (Figure 2) [53]. Exploring the potential of activators in modulating plant-microbe interactions and facilitating beneficial symbioses is an emerging area of interest [54]. Field trials and commercial development of activators as biostimulants or plant protection products are expected to increase, promoting sustainable agriculture practices [55]. In summary, plant activators targeting the SA signal pathway have played a crucial role in enhancing plant immunity, and ongoing research aims to develop more effective and targeted activators for improved crop protection and productivity [56].
2.1. The Past of Plant Activators
2.1. The Usage of Plant Activators
Plant activators play a crucial role in enhancing plant defense mechanisms against a wide array of pathogens [59]. Unlike traditional fungicides, which directly target and kill pathogens, plant activators stimulate the plant’s own immune system, enabling it to resist infections more effectively [60]. Plant activators were initially developed as pesticides to prevent rice blast [61]. For instance, Probenazole (PBZ) was widely applied to control rice blast in Japan in the last century [60,62]. In addition to their use in pest control, plant activators have also served as chemical probes in studying the mechanisms of plant resistance reactions [63]. The recent study discusses the creation of new 5-(thioether)-N-phenyl/benzyl-1, 3, 4-oxadiazole derivatives. These compounds were synthesized and evaluated for their antibacterial properties, showing significant activity against various bacterial strains. The study also explored the antibacterial mechanism, revealing that these derivatives disrupt bacterial cell membranes and inhibit essential enzymes, highlighting their potential as effective antibacterial agents [64]. The research by Ouyang, et al. [65] focuses on the design and synthesis of novel pyrazole derivatives containing oxime esters group. These compounds were evaluated for their antiviral activities, showing significant protective ability. The study also explores the structure-activity relationships to understand how modifications inducing plant host resistance as antiviral agents.
2.1.1. Screening Methods of Plant Activators
The screening of plant activators involves several methodologies to identify compounds that effectively enhance plant resistance [66]. One common approach is using living plants combined with symptom or disease severity observation, where plants are treated with potential activators and then exposed to pathogens; the reduction in disease symptoms indicates the efficacy of the activators (Table 1) [67]. Additionally, the detection of effector markers from the SA signaling pathway is employed, using techniques such as quantitative PCR or Western blotting to measure the expression levels of key markers like PRs, which indicate the activation of the pathway (Table 2) [68,69]. Determining action sites using transgenic or mutant plants is another vital method; genetically modified plants with specific mutations or transgenic expressions help pinpoint where and how activators interact with the plant’s defense mechanisms (Table 3) [70]. Molecular biology screening models further enhance these methods by employing high-throughput techniques such as gene expression profiling, proteomics, and metabolomics to comprehensively analyze the effects of potential activators at the molecular level [71]. Together, these screening methods provide a robust framework for identifying and understanding the mechanisms of effective plant activators [72,73].
2.1.2. Interaction Study of Target-Plant Activator
The study of interactions between plant activators and their targets involves understanding the SA signal pathway, which plays a crucial role in plant immunity [74]. This pathway is significant for its role in SAR, a defense mechanism that provides long-lasting protection against a broad spectrum of pathogens [4]. When a plant activator targets the SA pathway, it typically enhances the production and accumulation of SA, which in turn activates downstream defense genes, such as PRs [75]. These proteins bolster the plant’s immune response, making it more resistant to subsequent pathogen attacks [74]. The interaction studies often focus on how activators influence key components of the SA pathway, including NPR1, a regulatory protein that controls the expression of defense genes, and EDS1, which is involved in signaling processes leading to SAR [76]. By using techniques such as gene expression analysis, protein-protein interaction assays, and mutant plant studies, researchers can elucidate how specific activators modulate the SA pathway, thereby providing insights into their bioactivity and potential effectiveness as plant defense enhancers (Table 4) [77].
Table 4 The proteins physically interacted with SA.
2.2. Plant Activators against the SA Signal Pathway
Plant activators against the SA signal pathway play a crucial role in enhancing plant immunity by inducing various components upstream or downstream of the SA signal pathway [4,78].
2.2.1. HrBP1
So far, rarely research reported HrBP1 and its biology function [79]. HrBP1 was firstly isolated from a cDNA library of Arabidopsis thaliana [9]. This protein was found to locate in the plant cell wall [80]. Activated HrBP1 can prime many signal pathways containing with SA, ethylene (ET), and so on [80]. HrBP1 was participated in forming of the ion channel locating at the plant cell wall with the role of the signal transduction [81]. So far, the sequence of HrBP1 from various species indicated that HrBP1 is very evolutionarily conserved [82]. Harpins are secreted by gram-negative phytopathogenic bacteria harboring the hypersensitive response and pathogenicity (hrp) gene cluster, which were controlled by the secretion channel of three types [83,84]. These proteins possess the characteristics of glycine- or cysteine- rich, thermal stability and proteinases stable. Harpin was firstly isolated from Erwinia amylovora, and identified as an hypersensitivity (HR)-elicitor, then it was also identified from E. chrysanthemi, Pseudomonas syringae, Ralstonia solanacearum, and Xanthomonas, and named to hrpNEch, hrpZ, popA, and harpin-like Hpa1 [85,86,87]. Spraying harpins on the fruit or leaf of apple tree can enhance plant resistance, and then reduce the disease severity of the fruit infected by Penicillium expansum [88]. For citrus black spot caused by Guignardia citricarpa, spraying harpins on the fruit of citrus can reduce the number of the lesion and pycnidia of the pathogen [89]. Spraying harpins on tomato leaf can enhance the activity of POD, and reduce the disease severity caused by phytophthora infestans and Ps. syringae pv. tomato [90]. Pretreated harpin can reduce the numbers of lesions in citrus leaf cauased by X. axonopodis pv. citrumelo and X. axonopodis pv. Citri [91]. Postharvest application of harpin can reduce the size of lesion in hami melons inoculated with Trichothecium roseum [92]. In addition, spraying harpin can promote the growth of plant and increase production during the early growth stage of plants [93]. For the blue mold of apple fruit caused by Pe. expansum, pretreated harpin can intensify cell wall density. The morphological observation found that tannin vacuoles and appositions were accumulated in the epidermal and hypodermal cells, and the growth of mycelial was delayed [94]. Harpins can elicit the alkalinization of plant cells and generate a reactive oxygen species (ROS) burst, and induce plant cell apoptosis when the plant host was infected by the pathogens [95]. Harpin Psph can induce the expression of HIN1 gene, which belongs to tobacco PR gene, as well as the gene of 8-kD salicylic acid-responsive mitogen-activated protein kinase (MAPK) [84]. Meanwhile, harpin can activate plant the SA signal pathway by inducing PR-1 mRNA and increasing the SA amount [96]. Spraying Ps. syringae pv. Syringae 1.3 S on apple peel can induce the accumulation of PR-1a, PR-2, PR-5, and PR-8 [97]. Meanwhile, harpin cannot activate plant the SA signal pathway for NahG transgenic A. thaliana and nim1 mutants plant [98]. The latter is defective in responding to methyl jasmonate or ethylene dependent signal pathway. These results indicated that Harpin act in the upstream of SA in the SA signal pathway [99]. Harpin can bind the HrBP1 to form a protein-protein complex. A study speculated that the complex was changed in the spatial structure, then activate the SA signal pathway [100]. HrpZpss can bind the cell wall of tobacco suspension cell or living cell of tobacco leaf, but was failure to bind the protoplasm without cell wall. These results indicated the action site located on the plant cell wall [100].
2.2.2. Validamycin A and Validoxylamine A (VMA)
These antibiotics, derived from the bacterium Streptomyces hygroscopicus var. limoneus, are known to induce SAR in plants [101]. They function by triggering the SA pathway, leading to increased resistance against a variety of pathogens [46]. VMA, whose chemical name is ([1S-(1α, 4α, 5β, 6α]-1, 5, 6-trideoxy-4-O-β-D-glucopyranosyl-5-(hydroxymethyl)-1-[[4, 5, 6-trihydroxy-3-(hydroxymethyl)-2-cyclohexen-1-yl]amino]-D-chiro-inositol-1L-(1, 3, 4/2, 6)-2, 3-dihydroxy-6-hydroxymethyl-4-[(1S, 4R, 5S, 6S)-4, 5, 6-trihydroxy-3-hydroxymethylcyclohex-2-enylamino] cyclohexyl-3-D-glucopyranoside), is an aminoglucoside compound. The first aminoglycoside, streptomycin was isolated from Streptomyces griseus in 1943; neomycin was isolated from Streptomyces fradiae. This antibiotic was very effective against (Figure 2b) [46,102]. Validoxylamine A (VAA), whose chemical name is ([1S-(1α, 4α, 5β, 6α)]-1, 5, 6-trideoxy-5-(hydroxymethyl)-1-[[4, 5, 6-trihydroxy-3-(hydroxymethyl)-2-cyclohexen-1-yl] amino]-D-chiro-inositol), is an aglycon of VMA [103]. The mechanism of inhibiting pathogens for VMA and VAA involves two types of action: direct inhibition and induction of plant resistance. For instance, VMA can inhibit the growth of fungi in vitro by decreasing trehalase activity [104].
In addition, pretreated by VMA and VAA can reduce disease severity in tomato and cabbage infected by Ps. solanacearum, Xanthomonas campestris pv. campestris and Fusarium oxysporum f. sp. Lycopersici [105]. These results indicated that VMA and VAA possess the inductive activity [106]. VMA and VAA can accumulate the effector marker in the SA signal pathway in tomato leaf, such as PR-1, PR-2, and PR-5. The inductive effect of two compounds is similar to that of BTH [107]. VMA and VAA can also enhance the amount of free SA from the sprayed leaves and the upper leaves in tomato. For the treatment of above two compounds, the amount of free SA in the upper leaves is significantly higher than that of sprayed leaves [108]. Comparing with the content of SA in the upper leaves, the induction ability of VMA and VAA is significantly higher than that of PBZ treatment [109]. Above results indicated that VMA and VAA act at the upstream of SA in the SA signal pathway [106]. The study by Yang, et al. [110] which exhibited significant antiviral activity against the pepper mild mottle virus (PMMoV). Among these, compound A32 showed superior efficacy compared to the standard antiviral agents. The study also included a 3D-QSAR model to understand the structure-activity relationships, indicating the potential of these compounds for further antiviral development. The study of bioassays exposed that many of the title compounds exhibited moral inhibitory activities against TMV in vivo condition [111]. The study explores the creation and antibacterial properties of new benzofuran derivatives that include disulfide groups [112]. These compounds demonstrated significant antibacterial activity against various strains, and mechanistic studies revealed their ability to disrupt bacterial cell walls and interfere with protein synthesis. This research highlights the potential of these novel derivatives as effective antibacterial agents. Bai, et al. [113] study describes the creation of new antiviral compounds by combining indole and 4, 5-dihydro-1H-pyrazoline structures. These novel agents were tested against the potato virus Y (PVY), showing significant antiviral activity. Notably, one compound, D40, exhibited superior curative and protective effects compared to the commercial antiviral agent Ningnanmycin, enhancing key defense enzyme activities and regulating the carbon fixation pathway in plants. Zhang, et al. [114] indicated that some novel mesoionic pyrido [1, 2-a] pyrimidinone compounds featuring a dithioacetal group showed significant insecticidal activity against bean aphids and white-backed planthoppers. However, their effect on disease resistance is not addressed. In particular, compound 34 demonstrated superior insecticidal properties with an LC₅₀ value of 2.80 μg/mL, outperforming the commercial insecticide trifluoropyrimidine. The study suggests that these compounds act on nicotinic acetylcholine receptors, highlighting their potential as new insecticidal agents. Nonetheless, their role in inducing resistance in plants remains unexplored.
2.2.3. Probenazole (PBZ)
PBZ is a synthetic plant activator widely used in agriculture, especially in Japan, to control rice blast disease [115]. It works by stimulating the SA pathway, which enhances the plant’s natural defense mechanisms and increases resistance to the rice blast pathogen [116]. PBZ, whose chemical name is 3-allyloxy-1, 2-benzisothiazole-1, 1-dioxide, was developed as fungicides by Meiji Seika (Janpan) Co., Ltd (Figure 2c) [117]. During the 70-90s of the last century, PBZ was developed as plant protective agent (trade name: Oryzaemate) by Meijiseika Co., Tokyo, Japan, and widely used to prevent and control rice blast and rice bacterial blight in Asia [118]. It can induce the plant resistance against TMV, Pst, and Oidium SP. [119].
PBZ can up-regulate some defense-related genes of rice host, such as peroxidase (POD) and chitinase and PR genes [120,121]. It can also induce the mRNA accumulation of PR-1, PR-2, and PR-5 in A. thaliana leaves, and the amount of PR-1 is increased in the manner of dose dependent [26,122]. PBZ also enhance the activity of defense-related enzymes, such as phenylalanine ammonia-lyase (PAL), POD, and PBZ cannot induce the SA accumulation and up-regulate PR-1 mRNA for Salicylate hydroxylase (NahG) transgenic A. thaliana and the signaling mutants npr1-1 [60,123,124]. Meanwhile, PBZ did not exhibit an inductive effect on NahG transgenic tobacco [125]. These results indicated that PBZ can act upstream of SA in the SA signaling pathway (Figure 3). 1, 2-Benzisothiazol-3(2H)-1, 1-dioxide (BIT), also known as Saccharin, was identified as the active metabolite of PBZ [60]. BIT is found using a cultured parsley cells combined with spectrophotometric measurement of coumarin [126]. Then, spraying BIT can inhibit the growth of Pst tomato DC3000 in A. thaliana leaves (Figure 2d) [122]. Soil drench using BIT can also enhance the plant resistance against tobacco, cucumber and bean infected with TMV, Colletotrichum lagenarium, Uromyces appendiculatu [44]. BIT can accumulate total SA in plant host, and up-regulate the mRNA of PR-1, PR-2, and PR-5 in plant leaves [122,127]. Interestingly, PBZ can induce PBZ1 mRNA, namely PR-10 mRNA, but BIT be failure at the gene for rice suspension cells [128]. In addition, PBZ1 mRNA can be induced by rice blast fungus, but not correlated with the resistance of rice blast [129].
2.2.4. (D, L)-3-Aminobutyric Acid (BABA)
BABA is a non-protein amino acid that induces resistance in plants by activating the SA pathway [130]. It primes the plant’s immune system, leading to a faster and stronger defense response upon pathogen attack. BABA was firstly found to increase resistance of pea plants for the oomycete Aphanomyces euteiches [131]. Then, BABA was found to induce plant resistance against many plant diseases, such as many fungal, bacterial, and viral pathogens [132]. BABA can accumulate the intercellular fluid extract from N. tabacum cv. Xanthi-nc leaves, which was identified as PRs [133]. It can increase the amount of P14a. P14a is a type of peptidylglycine α-amidating monooxygenase (PAM), an enzyme that catalyzes the conversion of glycine-extended peptides to their amidated forms, which are often crucial for their biological activity. P14a likely refers to a specific PAM isoform or variant involved in a particular biological process or the pathway realated with β-1, 3-glucanase and chitinase in tomato plant leaves, and it enhance the activity of these enzymes [134]. For un-sprayed leaf, the amount of PR-1 mRNA was enhanced by BABA treatment; the inductive effect indicated that BABA is a systematic inducer [135]. Nevertheless, BABA cannot enhance plant resistance for NahG transgenic tobacco infected by TMV [60]. The result indicated that BABA act at the up-stream of SA in the SA signal pathway. In addition, many studies represent that BABA is also involved with HR or direct inhibitory effect against pathogens [60,136].
2.2.5. Imprimatins
These synthetic compounds are known to enhance plant immunity by activating the SA pathway [46]. Imprimatins increase the levels of SA and promote the expression of defense-related genes, thereby boosting the plant’s resistance to pathogens [46]. After plant being infected by the pathogens, the level of SA always increases and reaches at 10-20 fold for generating plant defence response [137]. Salicylic acid glucosyltransferase (SAGT) is responsible for transforming SA to its inactive metabolites, 2-O-beta-D-glucose and SA glucose ester [138,139]. So, SAGT possess the role of decreasing the SA level and maintaining a homeostasis of the SA amount in the plant host. A model system of the suspension cultured cells of A. thaliana infected with the bacterial pathogen Pst DC3000 avrRpm1 was used to screen the molecules with the characteristic of only strengthening plant resistance but not triggering plant resistance [140]. Imprimatins A and B were finally screened from above model system (Figure 3a to f), and these compounds can increase the SA level in A. thaliana plant after inoculated by the bacterial avirulent strain or the virulent strain, then generate a strong plant resistance [141]. At an effective concentration of inducing plant disease resistance, imprimatins compounds can block the activity of SAGT mutants in vitro (Figure 4). The results indicated that SAGT can competitively bind with imprimatins to SA [142].
2.2.6. Diuretics
Certain diuretic compounds have been found to act as plant activators by inducing the SA pathway [143]. These compounds enhance the plant’s defense responses, making them more resistant to various diseases. Three diuretics of 3-(butylamino)-4-phenoxy-5-sulfamoylbenzoicacid (Bumnide, MBT) (Figure 3f). 3-benzyl-1, 1-dioxo-6-(trifluoromethyl)-3, 4-dihydro-2H-1, 2, 4-benzothiadiine-7-sulfonamide (Bendroflumethiazide, BFM) (Figure 3g), and 4-chloro-N-(2, 6-dimethyl-1-piperidyl)-3-sulfamoyl-benzamide (Clopamide, CLP) (Figure 3h), were screened via the model system of suspension cultured cells above described [144]. These compounds can promote cell death caused by Pst avrRpm1 in a concentration-dependent manner, but their effects were weaker than that of SA. Three compounds can enhance the resistance of A. thaliana seedlings against Pst avrRpm1 and Pst. After A. thaliana seedlings infected by Pst, three diuretics can accumulate PR-1 mRNA [145]. Differing from the action mechanism of imprimatins A and imprimatins B, MBT and BFM can inhibit the activity of SAGT, and then SA is allowed to be degraded by SAGT. Nevertheless, CLP cannot nearly inhibit SAGT [146]. These upstream activators are crucial for the activation of the SA pathway, which is essential for the plant’s immune response. By inducing the production and accumulation of the SA, these compounds help to strengthen the plant’s defense mechanisms, providing long-lasting protection against a wide range of pathogens.
3. Plant Activators Acted in the Downstream of SA in the SA Signal Pathway
3.1. Downstream Activators
Downstream activators play critical roles in enhancing the SA signaling pathway, which is essential for plant immunity [74].
3.1.1. Benzothiadiazole (BTH)
BTH, also known as acibenzolar-S-methyl, is a synthetic analog of SA that acts downstream in the SA signaling pathway [147]. It mimics SA’s effects, inducing the expression of defense genes and enhancing disease resistance.
BTH, chemically named S-methyl benz [1, 2, 3] thiadiazole-7-carbothioate, was first reported by Schurter, et al. [148,149] (Figure 5a). Subsequent studies by schurter and colleagues systematically explored various bioassay models of plant activators, finding that the cucumber-colletotrichum lagenarium model was superior in resistance response and experimental stability compared to other plant-pathogen models such as cucumber/Ps. lachrymans, wheat/E. graminis, and tomato/Ph. infestans [150]. Field and greenhouse trials confirmed BTH’s high protective activity, leading to its development by Novartis/Syngenta under the trade name “Bion” [151,152]. BTH is metabolized by salicylic acid binding protein 2 (SABP2) into acibenzolar, its active form, which induces SAR in various plants including A. thaliana, tobacco, tomato, cucumber, wheat, sunflower, rice, maize, cotton, and soybean against multiple pathogens like TMV, turnip crinkle virus, wheat powdery mildew, Peronospora sparsa, Ps. syringae, F. oxysporum, Sclerotinia sclerotiorum, and cucumber mosaic virus [44,153,154,155,156,157,158,159,160,161,162,163,164]. BTH also inhibits nematode reproduction, specifically Meloidogyne incognita, when root-dipped [165]. For wheat infected by E. graminis, BTH spray can reduce pathogen proliferation at multiple stages of infection [166]. In tomatoes, BTH increases root cell wall density, reducing F. oxysporum entry [167]. In strawberries, BTH pretreatment increases phenolic compounds in leaves infected by powdery mildew [168]. For sugar beet roots infected by Beet necrotic yellow vein virus (BNYVV), BTH accumulates β-1, 3-glucanase and class IV chitinase in cortical and endodermis cells [169]. BTH induces accumulation of PR-1, PR-2, and PR-5 mRNA in A. thaliana leaves [170]. It also stimulates the expression of resistance-related genes in wheat [171]. Proteomic analyses, such as two-dimensional gel electrophoresis coupled with mass spectrometry, have shown that BTH up-regulates the SA pathway-related proteins in Arctic bramble and induces PAL and PRs expression in rice [171]. BTH induces PR genes in ethylene-response and methyl jasmonate-insensitive mutant plants, indicating that BTH operates independently of JA and ET pathways [141]. BTH also accumulates PR-1 gene expression in NahG transgenic tobacco, suggesting its action is downstream of SA (Figure 6,7) [172]. For BTH’s action mode, acibenzolar can induce PR-1a gene in SABP2-silenced tobacco, confirming SABP2 as a metabolic target [173]. In rice, BTH pretreatment up-regulates OsWRKY45 and OsWRKY13, transcription factors responsible for PRs gene expression [173,174]. This indicates that acibenzolar, the active form of BTH, induces WRKY transcription factors to regulate PRs genes, although the precise regulatory mechanism remains unclear [174].
3.1.2. N-Cyanomethyl-2-Chloroisonicotinamide (NCI)
NCI was first synthesized and identified for its protective activity against the rice/Pyricularia grisea model (Figure 5b) [175]. Subsequent studies have demonstrated NCI’s broad-spectrum protective efficacy against various pathogens in multiple plant species, including Ps. syringae pv. tomato (Pst) and Xanthomonas oryzae [176]. Remarkably, NCI’s protective activity against Py. grisea is significantly higher than that of PBZ [44]. Pretreatment with NCI significantly reduces lesion sizes in NahG transgenic tobacco plants infected by TMV and Oidiopsis lycopersici [177,178].
Additionally, the derivatives of NCI, such as N-methyl-2-chloroisonicotinamide and N-propargyl-2-chloroisonicotinamide, exhibit superior protective activity against rice blast [179]. NCI is known to induce the accumulation of mRNAs for the defense-related genes of PR-1, PR-2, and PR-5 in A. thaliana leaves without altering the levels of SA [177]. Similarly, NCI pretreatment induces these PRs genes in both wild-type and NahG transgenic tobacco leaves [180]. In rice leaves, NCI can also induce the expression of PR-10 (PBZ1 gene), and enhances PAL activity [126,181]. Importantly, pretreatment with NCI does not alter the levels of free and total SA in tobacco leaves. These findings indicate that NCI acts downstream of SA in the SA signaling pathway (Figure 7) [182]. Recent studies supported NCI’s role as an effective plant activator. For instance, research has shown that NCI can enhance resistance in a range of crops, contributing to sustainable agricultural practices by reducing the reliance on traditional chemical pesticides [183]. The recent study about the precise molecular mechanisms of NCI’s action and its integration into plant immune networks continue to be areas of active investigation [184].
3.1.3. 2,6-Dichloroisonicotinic Acid (INA)
INA, a derivative of isonicotinic acid, exhibits notable resistance-inducing properties in various plant species (Figure 5c) [184]. Originally observed in cucumber infected with C. lagenarium, subsequent studies have extended its protective effects to a wide array of plant-pathogen interactions [185]. These include defense responses in tobacco against TMV, A. thaliana against Pst, sugar beet against Cercospora beticola, rose against Sphaerotheca pannosa, barley against E. graminis f. sp. Hordei, and soybean against Sc. sclerotiorum, among others [186]. Field trials with INA have demonstrated its efficacy in enhancing resistance against pathogens such as Uromyces appendiculatus in beans and Alternaria macrospora in cotton seedlings [187]. Furthermore, INA treatments have shown positive effects on seed germination and plant growth, as evidenced by increased shoot and root lengths in INA-treated tomato seeds [188]. Mechanistically, INA elicits the accumulation of PRs gene, including PR-1, PR-2, and PR-5 in A. thaliana and a broader spectrum of PRs gene in tobacco plants [189]. It also induces PRs gene expression in rice, soybean, and tomato, enhancing the activity of defense enzymes such as chitinase and β-1, 3-glucanase [190]. Additionally, INA treatments have been associated with the upregulation of antioxidative enzymes like ascorbate peroxidase (APX) and catalase (CAT), contributing to enhanced defense responses in plants [191]. Furthermore, INA has been shown to accumulate intercellular washing fluid (IWF) in sugar beet leaves, leading to increased defense against pathogens such as C. beticola [192]. At a molecular level, INA primes SAR in tobacco plants by inducing PRs genes in NahG transgenic plant, thereby inhibiting TMV replication without altering the SA content [60]. However, this priming effect is impaired in A. thaliana mutants lacking functional NPR1 or NIM1 protein. Ramadass et al., [61] highlighted the essential role of these proteins in SAR signaling, suggesting the involvement of INA in the SA signaling pathway between SA and NPR1. Indeed, INA has been demonstrated to bind to SABP in vitro, potentially activating the SA signaling cascade [193,194]. Furthermore, affinity chromatography studies have identified INA’s interaction with BTH-binding protein kinase (BBPK), implicating it as a potential target protein for INA-mediated signaling [194]. These findings underscore the need for further investigation into the precise mechanisms by which INA activates the SA signaling pathway and its interactions with key regulatory proteins like SABP and BBPK (Figure 7) [195].
3.1.4. Tiadinil (TDL)
N-(3-chloro-4-methylphenyl)-4-methyl-1, 2, 3-thiadiazole-5-carboxamide, is the another name of TDL was introduced by Nihon Nohyaku Co., Ltd., and registered as a commercial pesticide for combating rice blast disease in Japan in 2003 (Figure 5d) [196]. Upon application, TDL undergoes conversion into its active metabolite, 4-methyl-1, 2, 3-thiadiazole-5-carboxylic acid (SV03), which serves to prime plant SAR (Figure 5e) [197]. Although TDL does not directly inhibit pathogen proliferation in vitro, the bioassays have demonstrated its efficacy in inducing resistance against rice blast and inhibiting the proliferation of pathogens of viral disease, bacterial disease, gray blight and anthracnose of tea tree [196,198]. Moreover, TDL has been shown to enhance the production of certain plant volatiles, indirectly suppressing herbivorous mites [199]. At the molecular level, TDL induces the expression of genes involved in the SA signaling pathway in tobacco leaves without altering the SA levels, suggesting its action downstream of the SA pathway [200]. Similarly, SV03 induces the accumulation of PR-1a, PR-2, and PR-5 mRNA in NahG transgenic tobacco leaves, and exhibits efficacy in inhibiting Pst proliferation and reducing lesion size in tobacco leaves infected with TMV [176, 201]. These findings underscore the potential of TDL and its active metabolite SV03 as effective tools for enhancing plant resistance against various pathogens, providing insights into their mode of action within the SA signaling pathway (Figure 7) [202].
3.1.5. 3-Chloro-1H-Pyrazole-5-Carboxylic Acid (CMPA)
A pyrazole derivative, CMPA, can reduce the disease symptoms of rice bacterial leaf blight in a dose-dependent manner, but it did not inhibit the proliferation of X. oryzae. CMPA can induce PBZ1 mRNA in rice and tobacco, then activate SAR. Nevertheless, CMPA cannt induce SA accumulation in tobacco (Figure 5f,7) [203,204,205].
3.1.6. Benzoylsalicylic Acid (BzSA) and Acetylsalicylic Acid (ASA)
Endogenous SA serves as a key regulator of the SA signaling pathway, influencing SAR against various plant pathogens [206]. External application of SA, particularly during early disease stages or at the seedling stage, has been shown to confer protective effects against a wide range of plant diseases caused by viruses, bacteria, and fungi [207]. This exogenous SA treatment induces the upregulation of PRs genes in tobacco, enhancing resistance against pathogens such as TMV [208,209]. Similarly, in tomato leaves, SA treatment followed by inoculation with Cladosporium fulvum leads to the accumulation of mRNA related to the SA signaling pathway, including extracellular PR-1, intracellular β-1, 3-glucanase, extracellular β-1, 3-glucanase, extracellular chitinase, and extracellular PR-4 [210]. Furthermore, SA plays a pivotal role in regulating R-gene mediated local disease resistance mechanisms [74]. As a derivative of SA, benzoylsalicylic acid (BzSA), also known as 2-(benzoyloxy) benzoic acid, has been isolated from the seed coats of Givotia rottleriformis (Figure 5g) [211]. Spraying BzSA has been demonstrated to reduce lesion number and size in tobacco leaves infected with TMV [212]. Moreover, BzSA treatment induces the accumulation of PR-1 and PR-5 mRNA and upregulates the expression of MYB and WRKY genes in both wild-type tobacco and transgenic NahG tobacco (Figure 7) [213]. Comparative analyses have revealed that BzSA exhibits higher efficacy in regulating marker genes of the SA signaling pathway and inhibiting TMV proliferation compared to SA and ASA (Figure 5h) [214]. Furthermore, injection of tobacco plants with ASA and benzoic acid derivatives has been shown to induce PRs mRNA expression and enhance tobacco resistance to TMV [215]. These findings underscore the potential of SA derivatives such as BzSA and ASA as effective tools for enhancing plant resistance against pathogens, offering valuable insights into their mechanisms of action and their applications in agricultural disease management strategies (Figure 7) [216].
3.1.7 3,5-Dichlorosalicylic Acid (3, 5-DCSA), 4-Chlorosalicylic Acid (4-CSA), and 5-Chlorosalicylic Acid (5-CSA)
3, 5-DCSA, 4-CSA, and 5-CSA have demonstrated the ability to enhance disease resistance in tobacco against TMV infection, with reduction rates in lesion size of 70.0±16.0%, 76.0±11.0%, and 66.0±9.0%, respectively (Figure 5i-k) [217]. However, their potency in inducing disease resistance is slightly weaker compared to that of SA, which achieves a reduction rate in lesion size of 80.3±7.2% [218]. These SA analogs also exhibit the capability to accumulate PR-1 in tobacco leaves, albeit with slightly lower induction effects than SA [219]. These chlorinated derivatives of SA offer potential advantages over SA itself, such as increased stability, prolonged activity, and reduced phytotoxicity (Figure 7) [220]. However, their effectiveness and mechanisms of action may differ, and further research is needed to fully understand their potential as plant activators targeting the SA signal pathway [47].
3.1.8. Dichloroanthranilic Acid (DCA)
DCA, identified through screening using the late/sustained up-regulation in response to Hyaloperonospora parasitica (LURP) cluster as a defense gene marker in A. thaliana, has been found to enhance disease resistance against virulent strains of the oomycete Hpa and Ps. syringae DC3000 (Figure 5l) [221]. DCA induces the accumulation of PR1 and WRKY70 mRNA in plants, but does not lead to the SA accumulation [222]. Notably, DCA’s inductive effect is independent of the NPR1-mediated pathway in npr1 mutants of A. thaliana but is blocked by the WRKY70 mutant, indicating its action downstream of SA in the SA signaling pathway, distinct from other compounds such as BTH, NCI, and INA [221].
3.1.9. Imprimatins C1 and Imprimatins C2
Analogous to SA, imprimatins C1 and imprimatins C2 induce cell death in Arabidopsis cells infected with avirulent Pst-avrrpm1, demonstrating concentration-dependent effects similar to SA (Figure 5m,n) [117]. These compounds, collectively referred to as "imprimatins C" molecules, induce the accumulation of PR-1 mRNA in both wild-type and salicylic acid-induction deficient 2 (sid 2) mutant A. thaliana seedlings, independent of pathogen infection [142]. However, pretreatment with imprimatins C1 fails to induce the SA accumulation in A. thaliana cells, suggesting their action on the downstream of SA in the SA signaling pathway, without activating SA biosynthesis through positive-feedback cycles (Figure 7) [142].
3.1.10. Oxycom™
Oxycom™, a form of peracetic acid, has been observed to elicit a robust resistance response against numerous plant pathogens, particularly affecting leaf, berry, and root diseases [223,224]. For example, pretreatment of tobacco leaves with Oxycom™ has been shown to reduce symptoms caused by Pst infection, with its protective efficacy surpassing that of SA [225]. Furthermore, Oxycom™ demonstrates a distinct ability compared to exogenous SA, significantly enhancing the expression of PR-1a, PR-1g, and PR-3a mRNA in tobacco leaves [226]. Pretreatment with Oxycom™ induces resistance against various pathogens in NahG transgenic tobacco, indicating its action downstream of SA in the SA signaling pathway [178]. Moreover, Oxycom™ has been found to rapidly induce the phosphorylation of Salicylic Acid-Induced Protein Kinase (SIPK) within 1 hour of treatment [227]. There appears to be a correlated dynamic trend between the expression of PR-1a mRNA and SIPK phosphorylation, suggesting that certain defense genes associated with SA signaling pathway may be mediated by SIPK [228]. These findings underscore the potential of Oxycom™ as an effective inducer of plant defense mechanisms, offering insights into its mode of action and its significance in plant disease management strategies (Figure 7) [229].
4. Activators with Unclear Action Sites
Activators with unclear action sites represent a group of compounds that have demonstrated the ability to enhance plant defense mechanisms against pathogens [230,231]. However, the exact pathways through which they exert their effects remain ambiguous [232]. There are some examples of plant activators with unclear or unknown action sites within the SA signaling pathway from previous research. Laminarin is a β-1, 3-glucan derived from brown algae that has been shown to induce defense responses in various plants (Table 1). While laminarin is known to activate the SA pathway, its exact point of action remains unclear. Some studies suggest it may act upstream of the SA biosynthesis, while others propose it acts downstream or in parallel with the SA signaling pathway [233]. Chitosan, a deacetylated derivative of chitin, can induce the SA accumulation and activate defense-related genes in plants [234]. However, the molecular mechanisms by which chitosan triggers the SA pathway are not fully understood [235]. Some researchers suggest it may interact with plant receptors to initiate signaling cascades leading to the SA biosynthesis, while others propose it acts through modulating ROS levels [235]. Hexanoic acid is a natural compound found in plants and has been reported to activate the SA pathway and induce resistance against pathogens [236]. However, the specific targets of hexanoic acid within the SA signaling network remain unclear (Table 2). Some studies suggest it may act upstream of the SA biosynthesis, while others propose it affects downstream components like NPR1 [74]. Azelaic acid is a dicarboxylic acid produced in plants during pathogen infection and has been shown to induce the SA accumulation and the defense responses [237]. However, the precise mechanisms by which azelaic acid activates the SA pathway are not well understood [2]. Some researchers suggest it may act through modulating lipid-derived signaling molecules, while others propose the direct effects on SA biosynthesis or downstream signaling components [238]. Riboflavin (vitamin B2) has been reported to induce resistance against pathogens in various plants, potentially through activating the SA pathway [239]. However, the specific targets and mechanisms by which riboflavin modulates the SA signaling cascade remain unclear, with some studies suggesting it may act through modulating cellular redox status or interacting with unknown receptors [240]. These examples elucidate that while many compounds have been identified as potential activators of the SA signaling pathway, their precise action sites and molecular mechanisms are often not well characterized [241]. Further research is needed to elucidate the specific targets and modes of action of these activators within the complex the SA signaling network [242].
4.1. 3,4-dichloro-2’-Cyano-1, 2-Thiazole-5-Carboxanilide (Isotianil)
Isotianil, initially considered a derivative of PBZ, was discovered through the screening of isothiazole compounds by Bayer CropScience AG, and further developed in collaboration with the Japanese company Sumitomo Chemical Co., Ltd., as a plant activator (Figure 8a) [119, 142]. Demonstrating notable efficacy, Isotianil exhibits protective activity against various rice diseases, including rice blast and bacterial leaf blight [243]. Remarkably, even at very low dosages, isotianil can sustain elevated resistance levels for extended durations [29]. Its inductive capacity surpasses that of PBZ and TDL [244]. Isotianil enhances the abundance and activity of defense-related enzymes such as lipoxygenase and PAL. Additionally, it induces the expression of OsWRKY45, a transcription factor responsible for PR gene expression [245].
4.2. Polypeptide-Product
Polypeptide-product, namely “Pen”, as a water soluble substances, was isolated from Pe. chrysogenum in 1990s [246]. Foliar spraying or soil drench with “Pen” can induce the plant resistance against many pathogens [247]. The pretreated “Pen” can increase the resistance level of many fruits and vegetables for P. viticola, U. necator, V. inaequalis, P. infestans, C. lagenarium, P. cubensis, and P. destructor, and reduce disease severity [248]. When the acidic dry mycelium extract or the neutralized dry mycelium extract were drenched the roots of melon plants, the treatment can also enhance the resistance level against F. oxysporum f. sp. melonis [249]. These two dry mycelium extracts can increase POD activity of plant host (Figure 5). Polypeptide-product can accumulate the mRNA of many defense genes in the cotton leaves, such as PR-1a, PR-1b, PR-2, PR-3, PR-4, PR-5, and so on. Nevertheless, the acurate action site of Polypeptide-product remains unclear [249].
4.3. β,γ-Methyleneadenosine 5’-Triphosphate (AMP-PCP)
AMP-PCP, which is a non-hydrolysable analogue of ATP, can enhance tobacco resistance against Pst and TMV [250]. AMP-PCP can accumulate PR-1, PR-2, and PR-5 in tobacco Xanthinc leaves [251]. Massively used AMP-PCP can inhibit the concentration of extracellular ATP (eATP) [252]. Nevertheless, increasing concentration of ATP can block the expression of PR-1 mRNA [253]. So, a lower concentration or deletion status of eATP can prime plant SAR. The action mechanism may be involved with two aspects: 1) ATP block SA synthesis; 2) ATP promote SA degradation. These two aspects lead to SA reduction. So, as an antagonist of ATP, AMP-PCP can be used for plant activator to activate the SA signal pathway [250].
4.4 2, 2-dichloro-3, 3-Dimethylcyclopropane Carboxylic Acid (DDCC)
DDCC has shown promise in preventing rice diseases caused by Piricularia oryzae and Pyriailaria oryzae (Figure 8b) [254]. Studies have demonstrated that root drenching with DDCC enhances the activity of POD in rice leaves inoculated with Pi. oryzae [255]. Moreover, pretreatment with DDCC can induce the production of melanin compounds at infection sites by Pi. oryzae, effectively inhibiting the growth of pathogens [256]. Furthermore, DDCC has been found to significantly accumulate phytoalexins, specifically momilactones A and B, in inoculation sites and surrounding tissue in rice leaves infected by Py. oryzae. Interestingly, these momilactones were scarcely detected in healthy rice leaves [257]. Momilactones A and B exhibit direct inhibitory effects on fungal growth in vitro, with ED50 values of 4.8 and 0.9 ng ml−1, respectively (Figure 6). Additionally, these compounds promote hyphal disruption in liquid media, suggesting their role as specialized defense materials against pathogens [258]. However, the precise action mechanism of DDCC remains elusive and requires further investigation [259].
4.5 Hyaluronic Acid (HA)
HA, a linear, unbranching polyanionic disaccharide produced by bacteria, consists of alternately joined glucuronic acid (GlcUA) and N-acetyl glucosamine (Glc-NAc) units through β-1-3 and β-1-4 glycosidic bonds [260]. Recent studies have highlighted its potential as a plant activator, showcasing its ability to enhance protective activity and mitigate disease severity in various plant species [261]. For instance, cucumber infected by C. orbiculare, Ps. syringae pv. lachrymans, tomato infected by Pst, Xanthomonas axonopodis pv. vesicatoria, and pepper infected by CMV have all shown improved resistance upon HA treatment [262]. Interestingly, while HA does not directly inhibit these pathogens in vitro, its application can induce SAR in plants. Furthermore, investigations using a tobacco model with transgenic PR-1a promoter-GUS and PDF1.2 promoter-GUS genes have revealed that HA strongly up-regulates these genes [263]. This observation suggests that HA activates both the SA and JA-mediated signaling pathways, distinguishing it from other known plant activators [262]. This dual activation of the SA and JA pathways by HA underscores its potential as a versatile tool for enhancing plant defense mechanisms against a wide range of pathogens [264].
5. The Development of Plant Activator against the SA Signal Pathway in the World at Present
5.1. Plant Activators Developed in China in Recent Years
In recent years, significant strides have been made in China in researching pesticides and plant protection. For example, researchers have synthesized a series of derivatives based on benzo-1, 2, 3-thiadiazole-7-carboxylic acid, with notable findings (Figure 9a) [265]. One such derivative, benzo-1, 2, 3-thiadiazole-7-carboxylic acid 2-(2-hydroxybenzoxyl) ethyl ester, demonstrated the ability to substantially elevate hydrogen peroxide (H2O2) levels and enhance the activity of PAL in taxus chinensis suspension cell cultures [266,267]. Moreover, novel derivatives of benzo-1, 2, 3-thiadiazole carboxylic esters were synthesized, showing promising control efficacy against a spectrum of bacteria and fungi in crops like cucumber, rice, and maize (Figure 9b) [268]. Subsequent research focused on synthesizing benzo-1, 2, 3-thiadiazole-7-carboxylate derivatives containing fluorine, revealing that derivatives with specific functional groups displayed substantial protective activity against pathogens like E. cichoracearum and C. lagenarium (Figure 9c)[268]. Additionally, novel thiadiazole derivatives incorporating thiazole or oxadiazole moieties were synthesized, demonstrating inducible protective activity against pathogens, as reported by Gan, et al. (Figure 9d,e) [269]. Notably, our research group developed an amino phosphonate compound containing benzo-1, 2, 3-thiadiazole, named dufulin, which exhibited superior protective activity across various crops under greenhouse and field conditions (Figure 9f) [269]. Dufulin showed inhibitory effects on the replication of TMV and southern rice black-streaked dwarf virus, while also enhancing the expression of PR-1a mRNA in tobacco leaves and the activity of PAL, POD, and polyphenol oxidase (PPO) in rice leaves or rice suspension cells [9]. Furthermore, the novel pyrimidine-like compound 5-(cyclopropylmethyl)-6-methyl-2-(2-pyridyl) pyrimidin-4-ol (PPA) emerged as a promising plant activator, exhibiting similar inhibitory effects to BTH against A. thaliana disease caused by Ps. syringae pv. maculicola strain (Figure 9g). PPA was found to accumulate mRNA of various genes related to the SA signal pathway in A. thaliana leaves, while also promoting H2O2 accumulation in intracellular sites inoculated with the pathogen, suggesting a novel mechanism of action [270,271]. Finally, 3-acetonyl-3-hydroxyoxindole (AHO), a derivative of isatin isolated from Strobilanthes cusia, demonstrated efficacy in reducing lesion number and size caused by TMV and E. cichoracearum (Figure 9h). AHO was found to induce the accumulation of PR-1a mRNA in N. benthamiana cv. Xanthinc leaves and increase the SA content and PAL activity (Figure 7). However, AHO pretreatment did not lead to the accumulation of SA and PR-1a in NahG transgenic tobacco, suggesting a complex interaction with the SA signal pathway [272].
5.2. Limited Understanding of Molecular Targets
Recent research has made significant strides in finding and understanding the molecular targets of plant activators. However, the advancements are crucial for improving plant resistance and the rational design of new activators [113].
1) Discovery of plant activators with novel structures: Some key bioactive compounds like withaferin-A and withanolide-D were identified from Withania somnifera, which can target specific pathways to inhibit nuclear factor kappa-B, enhancing dopaminergic D2 receptor activity and providing insights into their molecular interactions and potential as stress modulators and immune boosters in plants [273].
2) Discovery of new targets for plant activators: Some studies have demonstrated that silencing certain susceptibility genes (S genes) like REM1.3 using virus-induced gene silencing (VIGS) in plants can enhance resistance to pathogens such as P. infestans. This technique has been effective in crops like potato and soybean, revealing crucial targets for genetic modifications to improve disease resistance [274]. By detecting pathogen-associated molecular patterns (PAMPs), it has been found that Pattern Recognition Receptors (PRRs) and Nucleotide-binding Leucine-rich Repeat Receptors (NLRs) play a significant role in plant immune systems [275]. Advances in understanding these receptors have led to the identification of new molecular targets that can be manipulated to bolster plant defenses against a broad spectrum of pathogens [276].
3) Discovery of new mechanisms in plant resistance responses: Recent work highlights the role of Long non-coding RNAs (lncRNAs) in regulating plant immune responses [277,278,279]. For instance, silencing specific lncRNAs has led to increased resistance to fungal pathogens of tea leaf spot, providing new molecular targets for enhancing crop resistance [280,281,282].
4) New research methods for studying the targets of plant activators: CRISPR/Cas9 was used to edit miR482b and miR482c in tomatoes, which showing the promising results in increasing resistance to P. infestans [283]. At present, this method can identify and modify some critical genetic components involved in plant immunity, paving the way for targeted genetic enhancements [284,285].
5) The crosstalk between the SA signaling pathway and jasmonic acid (JA) signaling pathways and their synergistic role in disease resistance. At present, the interplay between these two pathways has been further elucidated, revealing how their modulation can influence plant responses to biotic stress [286]. Targeting these pathways and crucial nodes would help in designing activators that fine-tune plant immune responses for better protection against pests and diseases [287].
6) New approaches for the development of plant activators: At present, the integration of synthetic biology with traditional breeding techniques has led to the development of new plant activators [288]. These technologies enable precise modifications at the molecular level, enhancing the efficacy and specificity of plant defense mechanisms [289]. These studies collectively underscore the importance of identifying precise molecular targets for the development of effective plant activators, which can significantly enhance crop resilience and yield [290].
6. Future of Plant Activators
6.1. Action Mechanism of Plant Activators
In the past and present, many researchers have developed numerous plant activators targeting the SA signal pathway. However, several unresolved issues require further investigation:
Firstly, while the action sites of some plant activators have been identified upstream or downstream of SA in the SA signal pathway, the precise action mechanisms and targets of these activators remain unclear. For example, while SABP2 converts BTH into acibenzolar, the mechanism by which acibenzolar activates the SA signal pathway and its target are not fully understood [173,174].
Secondly, certain plant activators with diverse molecular structures can target the same protein to induce plant resistance. Therefore, it is speculated that the precise action mechanisms of these activators may differ. For instance, Imprimatins A and B target SAGT, but the spatial sites of interaction in SAGT may vary between two compounds [142].
Lastly, some plant activators can trigger the SA signal pathway while also activating other signal pathways or directly inhibiting microorganisms. Thus, these compounds possess diverse functions in the plant host, and their precise mechanisms require further study. Examples include BABA, HA, and so on [131,263,264].
6.2. Metabolism Mechanism of Plant Activators
The active components and metabolism mechanisms of certain plant activators remain unclear. Some activators are converted into specific metabolites within the plant host, which then induce plant resistance. However, other activators retain disease resistance activity without clear elucidation of their action and metabolism mechanisms. Examples include PBZ and its metabolite BIT, TDL, and its metabolite SV03 [291].
6.3. Screening Models of Plant Activators
The use of models such as the PR-1a or PDF1.2 gene promoters linked with GUS gene has proven effective in screening plant activators, as well as CaBP22 promoter linked with the GUS gene. These models offer advantages in stability and accurate intracellular localization [263]. Moving forward, constructing the models with the promoters of crucial genes upstream of the GUS gene could aid in screening the action sites of plant activators.
6.4. Discovery of Lead Compound Based on Novel Action Target
At present, we had not an accurate information of the action target of many plant activators, except for SABP and SAGT [138,139,140,173,193,194,195]. So, we are unable to design a rational structure template against molecular target. Next step, we should study the action target for these plant activator, and screen novel lead compound from nature.
7. Conclusion
The future of plant activators holds exciting prospects for advancing our understanding of plant defense mechanisms and revolutionizing crop protection strategies. By elucidating the action mechanisms, metabolism, and screening models, and discovering lead compounds based on novel targets, researchers can harness the full potential of plant activators to mitigate crop losses and ensure global food security in the face of evolving biotic stresses.
Author Contributions
CZ, MN designed the review content. MN and CZ wrote the drift. MN and CZ revised the Manuscript. CZ, D.Z, MN, KL and NA edit the final version of Manuscript. CZ, JZ supervision. CZ Project administration. CZ funding acquisition. All authors read and approved the manuscript.
Funding
This work was supported by the National Key Research Development Program of China (2022YFD1700504), the National Natural Science Foundation of China (No.21977023), the China Agriculture Research System of MOF and MARA (CARS-23-C07), and the Central Government Guides Local Science and Technology Development Fund Projects (Qiankehezhongyindi (2023) 001 and Qiankehezhongyindi (2024) 007).
Acknowledgments
This work was financially supported by the National Key Research Development Program of China (2022YFD1700504), the National Natural Science Foundation of China (No.21977023), the China Agriculture Research System of MOF and MARA (CARS-23-C07), and the Central Government Guides Local Science and Technology Development Fund Projects (Qiankehezhongyindi (2023) 001 and Qiankehezhongyindi (2024) 007).
Conflict of Interest
The authors declare that there is no conflict of interests regarding the publication of this article.
Submission Declaration
The current work has not been published previously and is not under consideration for publication elsewhere, that its publication is approved by all authors.
References
- Hayes, J.D.; Strange, R.C. Glutathione S-transferase polymorphisms and their biological consequences. Pharmaco. 2000, 61, 154–166. [Google Scholar] [CrossRef]
- Shine, M.; Xiao, X.; Kachroo, P.; Kachroo, A. Signaling mechanisms underlying systemic acquired resistance to microbial pathogens. Plant Sci. 2019, 279, 81–86. [Google Scholar] [CrossRef]
- Kumar, S.; Thilagam, P.; Shikha, D.; Saikanth, D.; Rahmani, U.; Huded, S.; Panigrahi, C.K. Adapting plant protection strategies to meet the challenges posed by climate change on plant diseases: a review. Int. J. Environ. Clim. Chang. 2023, 13, 25–36. [Google Scholar] [CrossRef]
- Peng, Y.; Yang, J.; Li, X.; Zhang, Y. Salicylic acid: biosynthesis and signaling. Annu. Rev. Plant Biol. 2021, 72, 761–791. [Google Scholar] [CrossRef]
- Saleem, M.; Fariduddin, Q.; Castroverde, C.D.M. Salicylic acid: A key regulator of redox signalling and plant immunity. Plant Physiol. Biochem. 2021, 168, 381–397. [Google Scholar] [CrossRef]
- Tanaka, S.; Han, X.; Kahmann, R. Microbial effectors target multiple steps in the salicylic acid production and signaling pathway. Front. Plant Sci. 2015, 6, 140488. [Google Scholar] [CrossRef]
- Hossain, M.M. Upscaling plant defense system through the application of plant growth-promoting fungi (PGPF). In: Microbial Technology for Agro-Ecosystems. Elsevier. 2024, 61–95. [CrossRef]
- Yang, C.; Dolatabadian, A.; Fernando, W.D. The wonderful world of intrinsic and intricate immunity responses in plants against pathogens. Cana. J. Plant Pathol. 2022, 44, 1–20. [Google Scholar] [CrossRef]
- Chen, Z.; Zeng, M.; Song, B.; Hou, C.; Hu, D.; Li, X.; Wang, Z.; Fan, H.; Bi, L.; Liu, J. Dufulin activates HrBP1 to produce antiviral responses in tobacco. PLoS ONE 2012, 7, e37944. [Google Scholar] [CrossRef]
- Durrant, W.E.; Dong, X. Systemic acquired resistance. Annu. Rev. Phytopathol. 2004, 42, 185–209. [Google Scholar] [CrossRef]
- Yuan, H.; Guo, W.; Zhao, L.; Yu, Y.; Chen, S.; Tao, L.; Cheng, L.; Kang, Q.; Song, X.; Wu, J. Genome-wide identification and expression analysis of the WRKY transcription factor family in flax (Linum usitatissimum L.). BMC Genomics 2021, 22, 375. [Google Scholar] [CrossRef]
- Bentham, A.R.; De la Concepcion, J.C.; Mukhi, N.; Zdrzałek, R.; Draeger, M.; Gorenkin, D.; Hughes, R.K.; Banfield, M.J. A molecular roadmap to the plant immune system. J. Biol. Chem. 2020, 295, 14916–14935. [Google Scholar] [CrossRef]
- Chen, L.; Wang, W.S.; Wang, T.; Meng, X.F.; Chen, T.T.; Huang, X.X.; Li, Y.J.; Hou, B.K. Methyl salicylate glucosylation regulates plant defense signaling and systemic acquired resistance. Plant Physiol. 2019, 180, 2167–2181. [Google Scholar] [CrossRef]
- Gao, Q.M.; Kachroo, A.; Kachroo, P. Chemical inducers of systemic immunity in plants. J. Exp. Bot. 2014, 65, 1849–1855. [Google Scholar] [CrossRef]
- Arya, A.; Sharma, G. Management of host plant resistance through immunization: an overview. J. Hill Agric. 2016, 7, 12–27. [Google Scholar] [CrossRef]
- Malamy, J.; Carr, J.P.; Klessig, D.F.; Raskin, I. Salicylic acid: a likely endogenous signal in the resistance response of tobacco to viral infection. Science 1990, 250, 1002–1004. [Google Scholar] [CrossRef]
- Métraux, J.; Signer, H.; Ryals, J.; Ward, E.; Wyss-Benz, M.; Gaudin, J.; Raschdorf, K.; Schmid, E.; Blum, W.; Inverardi, B. Increase in salicylic acid at the onset of systemic acquired resistance in cucumber. Science 1990, 250, 1004–1006. [Google Scholar] [CrossRef]
- Gaffney, T.; Friedrich, L.; Vernooij, B.; Negrotto, D.; Nye, G.; Uknes, S.; Ward, E.; Kessmann, H.; Ryals, J. Requirement of salicylic acid for the induction of systemic acquired resistance. Science 1993, 261, 754–756. [Google Scholar] [CrossRef]
- Ward, E.R.; Uknes, S.J.; Williams, S.C.; Dincher, S.S.; Wiederhold, D.L.; Alexander, D.C.; Ahl-Goy, P.; Metraux, J.P.; Ryals, J.A. Coordinate gene activity in response to agents that induce systemic acquired resistance. Plant Cell 1991, 3, 1085–1094. [Google Scholar] [CrossRef]
- Cao, H.; Bowling, S.A.; Gordon, A.S.; Dong, X. Characterization of an Arabidopsis thaliana mutant that is nonresponsive to inducers of systemic acquired resistance. Plant Cell. 1994, 6, 1583–1592. [Google Scholar] [CrossRef]
- Shah, J.; Kachroo, P.; Klessig, D.F. The Arabidopsis thaliana ssi1 mutation restores pathogenesis-related gene expression in npr1 plants and renders defensin gene expression salicylic acid dependent. Plant Cell. 1999, 11, 191–206. [Google Scholar] [CrossRef]
- Lapin, D.; Bhandari, D.D.; Parker, J.E. Origins and immunity networking functions of EDS1 family proteins. Annu. Rev. Phytopathol. 2020, 58, 253–276. [Google Scholar] [CrossRef] [PubMed]
- Jirage, D.; Tootle, T.L.; Reuber, T.L.; Frost, L.N.; Feys, B.J.; Parker, J.E.; Ausubel, F.M.; Glazebrook, J. Arabidopsis thaliana PAD4 encodes a lipase-like gene that is important for salicylic acid signaling. Proc. Natl. Acad. Sci. U.S.A. 1999, 96, 13583–13588. [Google Scholar] [CrossRef] [PubMed]
- Eulgem, T.; Somssich, I.E. Networks of WRKY transcription factors in defense signaling. Curr. Opin. Plant Biol. 2007, 10, 366–371. [Google Scholar] [CrossRef] [PubMed]
- Lai, Y.; Eulgem, T. Transcript-level expression control of plant NLR genes. Mol. Plant Pathol. 2018, 19, 1267–1281. [Google Scholar] [CrossRef] [PubMed]
- Thibaud, M.C.; Gineste, S.; Nussaume, L.; Robaglia, C. Sucrose increases pathogenesis-related PR-2 gene expression in Arabidopsis thaliana through an SA-dependent but NPR1-independent signaling pathway. Plant Physiol. Biochem. 2004, 42, 81–88. [Google Scholar] [CrossRef]
- Van Bockhaven, J.; De Vleesschauwer, D.; Höfte, M. Towards establishing broad-spectrum disease resistance in plants: silicon leads the way. J. Exp. Bot. 2013, 64, 1281–1293. [Google Scholar] [CrossRef] [PubMed]
- Qi, G.; Chen, J.; Chang, M.; Chen, H.; Hall, K.; Korin, J.; Liu, F.; Wang, D.; Fu, Z.Q. Pandemonium breaks out: disruption of salicylic acid-mediated defense by plant pathogens. Mol. Plant 2018, 11, 1427–1439. [Google Scholar] [CrossRef] [PubMed]
- Reglinski T, Dann E, Deverall B. Integration of induced resistance in crop production. Induced resistance for plant defence: A sustainable approach to crop protection. 2007, 201–228. [CrossRef]
- Chang, K.; Chen, J.Q.; Shi, Y.X.; Sun, M.J.; Li, P.F.; Zhao, Z.J.; Zhu, W.P.; Li, H.L.; Xu, Y.F.; Li, B.J. The discovery of new scaffold of plant activators: from salicylic acid to benzotriazole Chin. Chem. Lett. 2017, 28, 919–926. [Google Scholar] [CrossRef]
- Chen, Y.; He, B.; Hu, M.; Bao, J.; Yan, W.; Han, X.; Ye, Y. Fluorescent probes for imaging and detection of plant hormones and their receptors. Adv. Agrochem. 2023. [Google Scholar] [CrossRef]
- Zipfel, C. Plant pattern-recognition receptors. Trends. Immunol. 2014, 35, 345–351. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Xu, Y.; Sun, Y.; Wang, H.; Qi, J.; Wan, B.; Ye, W.; Lin, Y.; Shao, Y.; Dong, S. Leucine-rich repeat receptor-like gene screen reveals that Nicotiana RXEG1 regulates glycoside hydrolase 12 MAMP detection. Nat. Commun. 2018, 9, 594. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Wang, Y.; Zhang, X.; Chen, Z.; Xia, Y.; Wang, L.; Sun, Y.; Zhang, M.; Xiao, Y.; Han, Z. Plant receptor-like protein activation by a microbial glycoside hydrolase. Nature 2022, 610, 335–342. [Google Scholar] [CrossRef]
- Snoeck, S.; Garcia, A.G.; Steinbrenner, A.D. Plant receptor-like proteins (RLPs): structural features enabling versatile immune recognition. Physiol. Mol. Plant Pathol. 2023, 125, 102004. [Google Scholar] [CrossRef]
- Chen, Z.; Liu, F.; Zeng, M.; Wang, L.; Liu, H.; Sun, Y.; Wang, L.; Zhang, Z.; Chen, Z.; Xu, Y. Convergent evolution of immune receptors underpins distinct elicitin recognition in closely related Solanaceous plants. Plant Cell 2023, 35, 1186–1201. [Google Scholar] [CrossRef] [PubMed]
- Ma, C.; Liu, Y.; Bai, B.; Han, Z.; Tang, J.; Zhang, H.; Yaghmaiean, H.; Zhang, Y.; Chai, J. Structural basis for BIR1-mediated negative regulation of plant immunity. Cell Res. 2017, 27, 1521–1524. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Zeng, M.; Sun, Y.; Chen, Z.; Chen, Z.; Wang, L.; Cui, J.R.; Zhang, F.; Lv, D.; Chen, X. BAK1 protects the receptor-like kinase BIR2 from SNIPER2a/b-mediated degradation to promote pattern-triggered immunity in Nicotiana benthamiana. Plant Cell 2023, 35, 3566–3584. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Jackson, E.; Liu, X.; Huang, X.; van der Hoorn, R.A.; Zhang, Y.; Li, X. Proteolysis in plant immunity. Plant Cell 2024, koae142. [Google Scholar] [CrossRef] [PubMed]
- Schlöffel, M.A.; Salzer, A.; Wan, W.L.; van Wijk, R.; Del Corvo, R.; Šemanjski, M.; Symeonidi, E.; Slaby, P.; Kilian, J.; Maček, B.; Munnik, T.; Gust, A.A. The BIR2/BIR3-associated phospholipase Dγ1 negatively regulates plant immunity. Plant Physiol. 2020, 183, 371–384. [Google Scholar] [CrossRef] [PubMed]
- van Wersch, R.; Li, X.; Zhang, Y. Mighty dwarfs: Arabidopsis thaliana autoimmune mutants and their usages in genetic dissection of plant immunity. Front. Plant Sci. 2016, 7, 1717. [Google Scholar] [CrossRef]
- Gao, X.; Ruan, X.; Sun, Y.; Wang, X.; Feng, B. BAKing up to survive a battle: functional dynamics of BAK1 in plant programmed cell death. Front. Plant Sci. 2019, 9, 393664. [Google Scholar] [CrossRef]
- Yu, X.; Feng, B.; He, P.; Shan, L. From chaos to harmony: responses and signaling upon microbial pattern recognition. Annu. Rev. Phytopathol. 2017, 55, 109–137. [Google Scholar] [CrossRef] [PubMed]
- Faize, L.; Faize, M. Functional analogues of salicylic acid and their use in crop protection. Agronomy 2018, 8, 5. [Google Scholar] [CrossRef]
- Lü, Z.; Dai, X.; Xu, J.; Liu, Z.; Guo, Y.; Gao, Z.; Meng, F. Medicinal chemistry strategies toward broad-spectrum antiviral agents to prevent next pandemics. Eur. J. Med. Chem. 2024, 271, 116442. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, X. Salicylic acid: biosynthesis, perception, and contributions to plant immunity. Curr. Opin. Plant Biol. 2019, 50, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Klessig, D.F.; Choi, H.W.; Dempsey, D.M.A. Systemic acquired resistance and salicylic acid: past, present, and future. Mol. Plant Microbe Interact. 2018, 31, 871–888. [Google Scholar] [CrossRef]
- Ali, B. Salicylic acid: An efficient elicitor of secondary metabolite production in plants. Biocatal. Agr. Biotech. 2020, 31, 101884. [Google Scholar] [CrossRef]
- Singhal, R.K.; Saha, D.; Skalicky, M.; Mishra, U.N.; Chauhan, J.; Behera, L.P.; Lenka, D.; Chand, S.; Kumar, V.; Dey, P. Crucial cell signaling compounds crosstalk and integrative multi-omics techniques for salinity stress tolerance in plants. Front. Plant Sci. 2021, 12, 670369. [Google Scholar] [CrossRef] [PubMed]
- He, M.; He, C.Q.; Ding, N.Z. Abiotic stresses: general defenses of land plants and chances for engineering multistress tolerance. Front. Plant Sci. 2018, 9, 1771. [Google Scholar] [CrossRef]
- Deng, S.; Zhang, X.; Zhu, Y.; Zhuo, R. Recent advances in phyto-combined remediation of heavy metal pollution in soil. Biotechnol. Adv. 2024, 108337. [Google Scholar] [CrossRef] [PubMed]
- Cao, D.; Zhang, P.; Wang, S. Advances in structure-based drug design: The potential for precision therapeutics in psychiatric disorders. Neuron. 2024, 112, 526–538. [Google Scholar] [CrossRef]
- Tripathi, D.K.; Bhat, J.A.; Antoniou, C.; Kandhol, N.; Singh, V.P.; Fernie, A.R.; Fotopoulos, V. Redox regulation by priming agents towards a sustainable agriculture. Plant Cell Physiol. 2024, pcae031. [Google Scholar] [CrossRef] [PubMed]
- Jain, A.; Sarsaiya, S.; Singh, R.; Gong, Q.; Wu, Q.; Shi, J. Omics approaches in understanding the benefits of plant-microbe interactions. Front. Microbiol. 2024, 15, 1391059. [Google Scholar] [CrossRef] [PubMed]
- Silva, M.D.G.C.; Medeiros, A.O.; Converti, A.; Almeida, F.C.G.; Sarubbo, L.A. Biosurfactants: promising biomolecules for agricultural applications. Sustainability 2024, 16, 449. [Google Scholar] [CrossRef]
- Wang, L.; Bian, L.; Shi, Q.; Li, X.; Sun, Y.; Li, M.; Zhao, A.; Peng, X.; Yu, Y. The Vitis yeshanensis U-box E3 ubiquitin ligase VyPUB21 enhances resistance to powdery mildew by targeting degradation of NIM1-interacting (NIMIN) protein. Plant Cell Rep. 2024, 43, 93. [Google Scholar] [CrossRef] [PubMed]
- Conrath, U. Systemic acquired resistance. Plant Signal. Behave. 2006, 1, 179–184. [Google Scholar] [CrossRef]
- Zhou, M.; Wang, W. Recent advances in synthetic chemical inducers of plant immunity. Front. Plant Sci. 2018, 9, 425103. [Google Scholar] [CrossRef] [PubMed]
- Khoshru, B.; Mitra, D.; Joshi, K.; Adhikari, P.; Rion, M.S.I.; Fadiji, A.E.; Alizadeh, M.; Priyadarshini, A.; Senapati, A.; Sarikhani, M.R. Decrypting the multi-functional biological activators and inducers of defense responses against biotic stresses in plants. Heliyon 2023, 9, e13825. [Google Scholar] [CrossRef]
- Yu, L.; Wang, W.; Zeng, S.; Chen, Z.; Yang, A.; Shi, J.; Zhao, X.; Song, B. Label-free quantitative proteomics analysis of Cytosinpeptidemycin responses in southern rice black-streaked dwarf virus-infected rice. Pestic. Biochem. Physiol. 2018, 147, 20–26. [Google Scholar] [CrossRef]
- Ramadass, M.; Thiagarajan, P. Current trends and emerging technologies for pest control management of rice (Oryza sativa) plants. Environ. Biotech. 2021, 4, 125–179. [Google Scholar]
- Iwai, T.; Seo, S.; Mitsuhara, I.; Ohashi, Y. Probenazole-induced accumulation of salicylic acid confers resistance to Magnaporthe grisea in adult rice plants. Plant Cell Physiol. 2007, 48, 915–924. [Google Scholar] [CrossRef] [PubMed]
- Yokotani, N.; Hasegawa, Y.; Kouzai, Y.; Hirakawa, H.; Isobe, S. Transcriptome analysis of tomato plants following salicylic acid-induced immunity against Clavibacter michiganensis ssp. michiganensis. Plant Biotechnol. 2023, 40, 273–282. [Google Scholar] [CrossRef]
- Zhang, A.; He, H.; Wang, R.; Shen, Z.; Wu, Z.; Song, R.; Song, B. Synthesis, bioactivities, and antibacterial mechanism of 5-(Thioether)-N-phenyl/benzyl-1, 3, 4-oxadiazole-2-carboxamide/amine derivatives. J. Agric. Food Chem. 2024, 72, 1444–1453. [Google Scholar] [CrossRef]
- Ouyang, G.; Chen, Z.; Cai, X. J.; Song, B. A.; Bhadury, P. S.; Yang, S.; Jin, L. H.; Xue, W.; Hu, D. Y.; Zeng, S. Synthesis and antiviral activity of novel pyrazole derivatives containing oxime esters group. Bioorg. Med. Chem. 2008, 16, 9699–9707. [Google Scholar] [CrossRef]
- Wang, Y.; Zhao, Z.; Guo, R.; Tang, Y.; Guo, S.; Xu, Y.; Sun, W.; Tu, H.; Wu, J. Plant antiviral compounds containing pyrazolo [3, 4-d] pyrimidine based on the systemin receptor model. Arab. J. Chem. 2024, 105849. [Google Scholar] [CrossRef]
- El-Shetehy, M.; Moradi, A.; Maceroni, M.; Reinhardt, D.; Petri-Fink, A.; Rothen-Rutishauser, B.; Mauch, F.; Schwab, F. Silica nanoparticles enhance disease resistance in Arabidopsis thaliana plants. Nat. Nanotechnol. 2021, 16, 344–353. [Google Scholar] [CrossRef] [PubMed]
- Qi, P.; Huang, M.; Hu, X.; Zhang, Y.; Wang, Y.; Li, P.; Chen, S.; Zhang, D.; Cao, S.; Zhu, W.A. Ralstonia solanacearum effector targets TGA transcription factors to subvert salicylic acid signaling. Plant Cell 2022, 34, 1666–1683. [Google Scholar] [CrossRef]
- Chen, B.; Zhang, Y.; Yang, J.; Zhang, M.; Ma, Q.; Wang, X.; Ma, Z. The G-protein α subunit GhGPA positively regulates Gossypium hirsutum resistance to Verticillium dahliae via induction of SA and JA signaling pathways and ROS accumulation. Crop J. 2021, 9, 823–833. [Google Scholar] [CrossRef]
- Wani, S.H.; Anand, S.; Singh, B.; Bohra, A.; Joshi, R. WRKY transcription factors and plant defense responses: latest discoveries and future prospects. Plant Cell Rep. 2021, 40, 1071–1085. [Google Scholar] [CrossRef] [PubMed]
- Cui, M.; Cheng, C. High-throughput proteomics: a methodological mini-review. Lab. Invest. 2022, 102, 1170–1181. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Wang, J.; Lv, H.; Peng, Q.; Schreiner, M.; Baldermann, S.; Lin, Z. Integrated proteomic and metabolomic analyses reveal the importance of aroma precursor accumulation and storage in methyl jasmonate-primed tea leaves. Hortic. Res. 2021, 8, 95. [Google Scholar] [CrossRef] [PubMed]
- Lovelace, A.H.; Dorhmi, S.; Hulin, M.T.; Li, Y.; Mansfield, J.W.; Ma, W. Effector identification in plant pathogens. Phytopathology 2023, 113, 637–650. [Google Scholar] [CrossRef] [PubMed]
- Ding, L.N.; Li, Y.T.; Wu, Y.Z.; Li, T.; Geng, R.; Cao, J.; Zhang, W.; Tan, X.L. Plant disease resistance-related signaling pathways: recent progress and future prospects. Int. J. Mol. Sci. 2022, 23, 16200. [Google Scholar] [CrossRef]
- Javed, T.; Wang, W.; Yang, B.; Shen, L.; Sun, T.; Gao, S.J.; Zhang, S. Pathogenesis related-1 proteins in plant defense: regulation and functional diversity. Crit. Rev. Biotechnol. 2024, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Clinton, M.; Qi, G.; Wang, D.; Liu, F.; Fu, Z.Q. Reprogramming and remodeling: transcriptional and epigenetic regulation of salicylic acid-mediated plant defense. J. Exp. Bot. 2020, 71, 5256–5268. [Google Scholar] [CrossRef] [PubMed]
- Altmann, M.; Altmann, S.; Rodriguez, P.A.; Weller, B.; Elorduy Vergara, L.; Palme, J.; Marín-De la Rosa, N.; Sauer, M.; Wenig, M.; Villaécija-Aguilar, J.A. Extensive signal integration by the phytohormone protein network. Nature 2020, 583, 271–276. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.M.; Zhang, Y. Plant immunity: danger perception and signaling. Cell 2020, 181, 978–989. [Google Scholar] [CrossRef] [PubMed]
- Huang, M.; Yan, Y.; Wang, L.; Chen, J.; Liu, T.; Xie, X.; Li, X. Research on the interaction mechanism between α mino-phosphonate derivative QR and harpin-binding protein 1 in tobacco (Nicotiana tabacum) plants. Front. Microbiol. 2021, 12, 621875. [Google Scholar] [CrossRef]
- Showalter, A.M. Structure and function of plant cell wall proteins. Plant Cell 1993, 5, 9. [Google Scholar] [CrossRef] [PubMed]
- Ghouili, E.; Sassi, K.; Hidri, Y.; M’Hamed, H.C.; Somenahally, A.; Xue, Q.; Jebara, M.; Nefissi Ouertani, R.; Riahi, J.; de Oliveira, A.C. Effects of date palm waste compost application on root proteome changes of barley (Hordeum vulgare L.). Plants 2023, 12, 526. [Google Scholar] [CrossRef] [PubMed]
- Huang, M.; Wu, Z.; Li, J.; Ding, Y.; Chen, S.; Li, X. Plant protection against viruses: an integrated review of plant immunity agents. Int. J. Mol. Sci. 2023, 24, 4453. [Google Scholar] [CrossRef] [PubMed]
- Kvitko, B.H.; Collmer, A. Discovery of the Hrp type III secretion system in phytopathogenic bacteria: how investigation of hypersensitive cell death in plants led to a novel protein injector system and a world of inter-organismal molecular interactions within plant cells. Phytopathology 2023, 113, 626–636. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Liu, J.; Ma, W.; Li, X. Characteristics, roles and applications of proteinaceous elicitors from pathogens in plant immunity. Life 2023, 13, 268. [Google Scholar] [CrossRef] [PubMed]
- Sakata, N.; Fujikawa, T.; Uke, A.; Ishiga, T.; Ichinose, Y.; Ishiga, Y. HexR transcription factor contributes to Pseudomonas cannabina pv. alisalensis virulence by coordinating type three secretion system genes. Microorganisms 2023, 11, 1025. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Cao, J.; Cai, X.; Wang, J.; Kong, F.; Wang, D.; Wang, J. Antagonistic activity of volatile organic compounds produced by acid-tolerant Pseudomonas protegens CLP-6 as biological fumigants to control tobacco bacterial wilt caused by Ralstonia solanacearum. Appl. Environ. Microb.. 2023, 89, e01892–01822. [Google Scholar] [CrossRef]
- Zhao, R.; Kang, I.J.; Lee, S. Current status and future prospects in genomic research and breeding for resistance to Xanthomonas citri pv. Glycines in soybean. Agronomy 2023, 13, 490. [Google Scholar] [CrossRef]
- Dai, P.; Li, N.; Li, B.; Wang, S.; Wang, Y.; Meng, X.; Li, B.; Cao, K.; Hu, T. An endophytic strain Bacillus velezensis JZ51 controlled pink mold rot of postharvest apple fruit via antagonistic action and disease resistance induction. Postharvest Biol. Technol. 2024, 210, 112793. [Google Scholar] [CrossRef]
- Ali, S.; Hameed, A.; Muhae-Ud-Din, G.; Ikhlaq, M.; Ashfaq, M.; Atiq, M.; Ali, F.; Zia, Z.U.; Naqvi, S.A.H.; Wang, Y. Citrus canker: a persistent threat to the worldwide citrus industry an analysis. Agronomy 2023, 13, 1112. [Google Scholar] [CrossRef]
- Karačić, V.; Miljaković, D.; Marinković, J.; Ignjatov, M.; Milošević, D.; Tamindžić, G.; Ivanović, M. Bacillus species: excellent biocontrol agents against tomato diseases. Microorganisms 2024, 12, 457. [Google Scholar] [CrossRef]
- Shah, M.H.; Ajaharuddin, S.M.; Rahman, S.S.; Saha, S.; Goswami, S.; Islam, S.; Barik, S.; Hossain, A. Latef, A.A.H.A. Citrus lemon and stressful conditions. In Medicinal plant responses to stressful conditions, 1st Edition; CRCPress: Boca Raton, 2023. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhu, X.; Ma, H.; Ren, X.; Shi, H.; Liu, Z.; Liang, J. Exogenous glucose reduces the incidence of black rot disease in apricot (Prunus armeniaca L.) by regulating energy metabolism and ROS. Sci. Hortic-Amsterdam. [CrossRef]
- Tian, S.; Chen, Y.; Cai, Z.; Wang, Q.; Zhang, J.; Liu, Z.; Li, Y.; Zhao, X. Effects of harpin and carbendazim on antioxidant accumulation in young jujube leaves. Open Chem. 2023, 21, 20220284. [Google Scholar] [CrossRef]
- Delmas, C.E.; Bancal, M.O.; Leyronas, C.; Robin, M.H.; Vidal, T.; Launay, M. Monitoring the phenology of plant pathogenic fungi: why and how? Biol. Rev. 2024. [Google Scholar] [CrossRef]
- Jiang, Y.; Yue, Y.; Wang, Z.; Lu, C.; Yin, Z.; Li, Y.; Ding, X. Plant biostimulant as an environmentally friendly alternative to modern agriculture. J. Agric. Food Chem. 2024, 72, 5107–5121. [Google Scholar] [CrossRef] [PubMed]
- Yuan, X.; Gdanetz, K.; Outwater, C.A.; Slack, S.M.; Sundin, G.W. Evaluation of plant defense inducers and plant growth regulators for fire blight management using transcriptome studies and field assessments. Phytopathology 2023, 113, 2152–2164. [Google Scholar] [CrossRef] [PubMed]
- Khadiri, M.; Boubaker, H.; Lahmamsi, H.; Taoussi, M.; Ezzouggari, R.; Askarne, L.; Farhaoui, A.; Barka, E.A.; Lahlali, R. Challenges in apple preservation: Fungicide resistance and emerging biocontrols. Physiol. Mol. Plant P. 2023, 102205. [Google Scholar] [CrossRef]
- Fu, Y.; Liu, X.; Wang, Q.; Liu, H.; Cheng, Y.; Li, H.; Zhang, Y.; Chen, J. Two salivary proteins Sm10 and SmC002 from grain aphid Sitobion miscanthi modulate wheat defense and enhance aphid performance. Front. Plant Sci. 2023, 14, 1104275. [Google Scholar] [CrossRef] [PubMed]
- Ang, M.C.Y.; Saju, J.M.; Porter, T.K.; Mohaideen, S.; Sarangapani, S.; Khong, D.T.; Wang, S.; Cui, J.; Loh, S.I.; Singh, G.P. Decoding early stress signaling waves in living plants using nanosensor multiplexing. Nat. Commun. 2024, 15, 2943. [Google Scholar] [CrossRef] [PubMed]
- Kongala, S.I.; Mamidala, P. Harpin-loaded chitosan nanoparticles induced defense responses in tobacco. Carbohydr. Polym. Tech. 2023, 5, 100293. [Google Scholar] [CrossRef]
- Jin, L.; Feng, S.; Tang, S.; Dong, P.; Li, Z. Biological control of potato late blight with a combination of Streptomyces strains and biochar. Biol. Control. 2023, 183, 105248. [Google Scholar] [CrossRef]
- Asano, N.; Yamaguchi, T.; Kameda, Y.; Matsui, K. Effect of validamycins on glycohydrolases of Rhizoctonia solani. J. Antibiot. 1987, 40, 526–532. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Ma, X.; Sun, T.; Zhu, Q.-H.; Feng, H.; Li, Y.; Liu, F.; Zhang, X.; Sun, J.; Li, Y. VdPT1 encoding a neutral trehalase of Verticillium dahliae is required for growth and virulence of the pathogen. Int. J. Mol. Sci. 2023, 25, 294. [Google Scholar] [CrossRef]
- Asim, M.; Zhang, Y.; Sun, Y.; Guo, M.; Khan, R.; Wang, X.L.; Hussain, Q.; Shi, Y. Leaf senescence attributes: the novel and emerging role of sugars as signaling molecules and the overlap of sugars and hormones signaling nodes. Crit. Rev. Biotechnol. 2023, 43, 1092–1110. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Wang, J.; Yue, Z.; Wang, J.; Dou, T.; Chen, T.; Li, J.; Dai, H.; Yu, J. Study of cabbage antioxidant system response on early infection stage of Xanthomonas campestris pv. campestris. BMC Plant Bio. 2024, 24, 324. [Google Scholar] [CrossRef] [PubMed]
- Yan, F.; Ma, J.; Peng, M.; Xi, C.; Chang, S.; Yang, Y.; Tian, S.; Zhou, B.; Liu, T. Lactic acid induced defense responses in tobacco against Phytophthora nicotianae. Sci. Rep. 2024, 14, 9338. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.M.; El-Sappah, A.H.; Ali, H.M.; Zandi, P.; Huang, Q.; Soaud, S.A.; Alazizi, E.M.; Wafa, H.A.; Hossain, M.A.; Liang, Y. Pathogenesis-related proteins (PRs) countering environmental stress in plants: a review. S. Afr. J. Bot. 2023, 160, 414–427. [Google Scholar] [CrossRef]
- Kim, T.J.; Lim, G.H. Salicylic acid and mobile regulators of systemic immunity in plants: Transport and metabolism. Plants 2023, 12, 1013. [Google Scholar] [CrossRef]
- Ishikawa, M.; Sharkhuu, N.; Zhang, Y.; Kadota, T.; Ohata, T. Ground thermal and moisture conditions at the southern boundary of discontinuous permafrost, Mongolia. Permafrost Periglac. 2005, 16, 209–216. [Google Scholar] [CrossRef]
- Yang, Y.; Hu, D.; Wang, S.; Wang, Z.; Zu, G.; Song, B. First discovery of novel cytosine derivatives containing a sulfonamide moiety as potential antiviral agents. J. Agr. Food Chem. 2022, 70, 6026–6036. [Google Scholar] [CrossRef] [PubMed]
- Gan, X.; Hu, D.; Li, P.; Wu, J.; Chen, X.; Xue, W.; Song, B. Design, synthesis, antiviral activity and three-dimensional quantitative structure activity relationship study of novel 1, 4-pentadien-3-one derivatives containing the 1, 3, 4-oxadiazole moiety. Pest Manag. Sci. 2016, 72, 534–543. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Liu, Q.; Li, S.; Zeng, L.; Yao, J.; Li, H.; Shen, Z.; Lu, F.; Wu, Z.; Song, B. Design, synthesis, antibacterial activity, and mechanisms of novel benzofuran derivatives containing disulfide moieties. J. Agr. Food Chem. 2024, 72, 10195–10205. [Google Scholar] [CrossRef] [PubMed]
- Bai, L.; Wei, C.; Shen, Z.; He, H.; Yang, X.; Shi, S.; Hu, D.; Song, B. Splicing indoles and 4, 5-dihydro-1 H-pyrazoline structure gave birth to novel antiviral agents: Design, synthesis, and mechanism study. J. Agr. Food Chem. 2023, 71, 7239–7249. [Google Scholar] [CrossRef]
- Zhang, J.; Song, R.; Wu, S.; Cai, D.; Wu, Z.; Liu, Z.; Hu, D.; Song, B. Discovery of pyrido [1, 2-a] pyrimidinone mesoionic compounds incorporating a dithioacetal moiety as novel potential insecticidal agents. J. Agr. Food Chem. 2021, 69, 15136–15144. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, Q.T.; Ueda, K.; Kihara, J.; Ueno, M. Induction of resistance in rice against Magnaporthe oryzae by culture filtrates of Biscogniauxia sp. O821. J. Phytopathol. 2016, 164, 990–995. [Google Scholar] [CrossRef]
- Salman, E.K.; Ghoniem, K.E.; Badr, E.S.; Emeran, A.A. The potential of dimetindene maleate inducing resistance to blast fungus Magnaporthe oryzae through activating the salicylic acid signaling pathway in rice plants. Pest Manag. Sci. 2022, 78, 633–642. [Google Scholar] [CrossRef]
- Jiang, K.; Asami, T. Chemical regulators of plant hormones and their applications in basic research and agriculture. Biosci. Biotechnol. Biochem. 2018, 82, 1265–1300. [Google Scholar] [CrossRef]
- Korbecka-Glinka, G.; Piekarska, K.; Wiśniewska-Wrona, M. The use of carbohydrate biopolymers in plant protection against pathogenic fungi. Polymers 2022, 14, 2854. [Google Scholar] [CrossRef] [PubMed]
- Nakashita, H.; Yasuda, M.; Nitta, T.; Asami, T.; Fujioka, S.; Arai, Y.; Sekimata, K.; Takatsuto, S.; Yamaguchi, I.; Yoshida, S.; Yoshida, S. Brassinosteroid functions in a broad range of disease resistance in tobacco and rice. Plant J. 2003, 33, 887–898. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Chen, B.; Guo, Q.; Shi, L.; He, M.; Qin, Z.; Li, L.; He, P.; Wang, Z.; Hu, D. A time-course proteomic analysis of rice triggered by plant activator BTH. J. Plant Growth. Regul. /: 392–409. https, 1007. [Google Scholar]
- Midoh, N.; Iwata, M. Cloning and characterization of a probenazole-inducible gene for an intracellular pathogenesis-related protein in rice. Plant Cell Physiol. 1996, 37, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Yoshioka, K.; Nakashita, H.; Klessig, D.F.; Yamaguchi, I. Probenazole induces systemic acquired resistance in Arabidopsis thaliana with a novel type of action. Plant J. 2001, 25, 149–157. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Long, S.; Liu, K.; Zhu, T.; Gong, J.; Gao, S.; Wang, R.; Zhang, L.; Liu, T.; Xu, Y. Paclobutrazol ameliorates low-light-induced damage by improving photosynthesis, antioxidant defense system, and regulating hormone levels in tall fescue. Int. J. Mol. Sci. 2022, 23, 9966. [Google Scholar] [CrossRef]
- Osés-Ruiz, M.; Cruz-Mireles, N.; Martin-Urdiroz, M.; Soanes, D.M.; Eseola, A.B.; Tang, B.; Derbyshire, P.; Nielsen, M.; Cheema, J.; Were, V. Appressorium-mediated plant infection by Magnaporthe oryzae is regulated by a Pmk1-dependent hierarchical transcriptional network. Nat microbiol. 2021, 6, 1383–1397. [Google Scholar] [CrossRef] [PubMed]
- Dyakov, Y.T.; Dzhavakhiya, V.; Korpela, T. Molecular basis of plant immunization. Compr. Mol. Phytopathol Elsevier 2007, 423–438. [Google Scholar] [CrossRef]
- Bektas, Y.; Eulgem, T. Synthetic plant defense elicitors. Front. Plant Sci. 2015, 5, 122149. [Google Scholar] [CrossRef]
- Tuteja, N.; Gill, S.S.; Trivedi, P.K.; Asif, M.H.; Nath, P. Plant growth regulators and their role in stress tolerance. Plant Nutr. Abiotic Stress Toler. I Plant Stress 2010, 4, 1–18. [Google Scholar]
- Kim, S.T.; Kim, S.G.; Kang, Y.H.; Wang, Y.; Kim, J.Y.; Yi, N.; Kim, J.K.; Rakwal, R.; Koh, H.J.; Kang, K.Y. Proteomics analysis of rice lesion mimic mutant (spl1) reveals tightly localized probenazole-induced protein (PBZ1) in cells undergoing programmed cell death. J. Proteome Res. 2008, 2008. 7, 1750–1760. [Google Scholar] [CrossRef]
- Vergne, E.; Ballini, E.; Marques, S.; Sidi Mammar, B.; Droc, G.; Gaillard, S.; Bourot, S.; DeRose, R.; Tharreau, D.; Nottéghem, J.L. Early and specific gene expression triggered by rice resistance gene Pi33 in response to infection by ACE1 avirulent blast fungus. New Phytol. 2007, 174, 159–171. [Google Scholar] [CrossRef] [PubMed]
- Jakab, G.; Cottier, V.; Toquin, V.; Rigoli, G.; Zimmerli, L.; Métraux, J.P.; Mauch-Mani, B. β-Aminobutyric acid-induced resistance in plants. Eur. J. Plant Pathol. 2001, 107, 29–37. [Google Scholar] [CrossRef]
- Cohen, Y.; Vaknin, M.; Mauch-Mani, B. BABA-induced resistance: milestones along a 55-year journey. Phytoparasitica 2016, 44, 513–538. [Google Scholar] [CrossRef]
- Enebe, M.C.; Babalola, O.O. The impact of microbes in the orchestration of plants’ resistance to biotic stress: a disease management approach. Appl. Microbiol. Biotechnol. 2019, 103, 9–25. [Google Scholar] [CrossRef] [PubMed]
- Cutt, J.R.; Harpster, M.H.; Dixon, D.C.; Carr, J.P.; Dunsmuir, P.; Klessig, D.F. Disease response to tobacco mosaic virus in transgenic tobacco plants that constitutively express the pathogenesis-related PR1b gene. Virology 1989, 173, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Singh, D.P.; Maurya, S.; Yerasu, S.R.; Chaurasia, A.; Prabha, R.; Shukla, R.; Satnami, L.; Rai, N.; Behera, T.K. Proteomics analysis of wild and cultivated tomato species challenged with Alternaria solani unveiled differential tolerance mechanisms. Research Square 2024. [Google Scholar] [CrossRef]
- Mehra, A.; Mazumdar-Leighton, S. The sublime art of war: Herbivore-induced plant volatiles. Resonance 2024, 29, 29–49. [Google Scholar] [CrossRef]
- Odenbreit, S.; Kavermann, H.; Püls, J.; Haas, R. CagA tyrosine phosphorylation and interleukin-8 induction by Helicobacter pylori are independent from alpAB, HopZ and bab group outer membrane proteins. Int. J. Med. Microbiol. 2002, 292, 257–266. [Google Scholar] [CrossRef]
- Kouzai, Y.; Kimura, M.; Yamanaka, Y.; Watanabe, M.; Matsui, H.; Yamamoto, M.; Ichinose, Y.; Toyoda, K.; Onda, Y.; Mochida, K. Expression profiling of marker genes responsive to the defence-associated phytohormones salicylic acid, jasmonic acid and ethylene in Brachypodium distachyon. BMC Plant Biol. 2016, 16, 1–11. [Google Scholar] [CrossRef]
- Hu, Y.; Zhang, M.; Lu, M.; Wu, Y.; Jing, T.; Zhao, M.; Zhao, Y.; Feng, Y.; Wang, J.; Gao, T. Salicylic acid carboxyl glucosyltransferase UGT87E7 regulates disease resistance in Camellia sinensis. Plant Physiol. 2022, 188, 1507–1520. [Google Scholar] [CrossRef] [PubMed]
- Dean, J.V.; Delaney, S.P. Metabolism of salicylic acid in wild-type, ugt74f1 and ugt74f2 glucosyltransferase mutants of Arabidopsis thaliana. Physiol. Plantarum 2008, 132, 417–425. [Google Scholar] [CrossRef]
- Park, H.J.; Kwon, C.S.; Woo, J.Y.; Lee, G.J.; Kim, Y.J.; Paek, K.H. Suppression of UDP-glycosyltransferase-coding Arabidopsis thaliana UGT74E2 gene expression leads to increased resistance to Psuedomonas syringae pv. tomato DC3000 infection. Plant Pathol. J. 2011, 27, 170–182. [Google Scholar] [CrossRef]
- Zhang, W.; Zhao, F.; Jiang, L.; Chen, C.; Wu, L.; Liu, Z. Different pathogen defense strategies in Arabidopsis: more than pathogen recognition. Cells 2018, 7, 252. [Google Scholar] [CrossRef]
- Noutoshi, Y.; Okazaki, M.; Kida, T.; Nishina, Y.; Morishita, Y.; Ogawa, T.; Suzuki, H.; Shibata, D.; Jikumaru, Y.; Hanada, A. Novel plant immune-priming compounds identified via high-throughput chemical screening target salicylic acid glucosyltransferases in Arabidopsis thaliana. Plant Cell 2012, 24, 3795–3804. [Google Scholar] [CrossRef]
- Palmer, I.A.; Chen, H.; Chen, J.; Chang, M.; Li, M.; Liu, F.; Fu, Z.Q. Novel salicylic acid analogs induce a potent defense response in Arabidopsis thaliana. Int. J. Mol. Sci. 2019, 20, 3356. [Google Scholar] [CrossRef] [PubMed]
- Fürstenberg-Hägg, J.; Zagrobelny, M.; Bak, S. Plant defense against insect herbivores. Int. J. Mol. Sci. 2013, 14, 10242–10297. [Google Scholar] [CrossRef]
- Künstler, A.; Gullner, G.; Ádám, A.L.; Kolozsváriné Nagy, J.; Király, L. The versatile roles of sulfur-containing biomolecules in plant defense-a road to disease resistance. Plants 2020, 9, 1705. [Google Scholar] [CrossRef]
- Wani, M.Y.; Mehraj, S.; Rather, R.; Rani, S.; Hajam, O.; Ganie, N.; Mir, M.; Baqual, M.; Kamili, A.S. Systemic acquired resistance (SAR): A novel strategy for plant protection with reference to mulberry. Int. J. Chem. Stud. 2018, 6, 1184–1192. [Google Scholar]
- Hernández, J.A.; Diaz-Vivancos, P.; Barba-Espín, G.; Clemente-Moreno, M.J. On the role of salicylic acid in plant responses to environmental stresses. Salicylic acid:a multifaceted hormone 2017, 17–34. [CrossRef]
- Reitsma, J.M.; Liu, X.; Reichermeier, K.M.; Moradian, A.; Sweredoski, M.J.; Hess, S.; Deshaies, R.J. Composition and regulation of the cellular repertoire of SCF ubiquitin ligases. Cell 2017, 171, 1326–1339. [Google Scholar] [CrossRef] [PubMed]
- Osés-Ruiz, M.; Cruz-Mireles, N.; Martin-Urdiroz, M.; Soanes, D.M.; Eseola, A.B.; Tang, B.; Derbyshire, P.; Nielsen, M.; Cheema, J.; Were, V. Appressorium-mediated plant infection by Magnaporthe oryzae is regulated by a Pmk1-dependent hierarchical transcriptional network. Nat. Microbiol. 2021, 6, 1383–1397. [Google Scholar] [CrossRef] [PubMed]
- Salotti, I.; Liang, Y.J.; Ji, T.; Rossi, V. Development of a model for Colletotrichum diseases with calibration for phylogenetic clades on different host plants. Front. Plant Sci. 2023, 14, 1069092. [Google Scholar] [CrossRef]
- Von Rad, U.; Mueller, M.J.; Durner, J. Evaluation of natural and synthetic stimulants of plant immunity by microarray technology. New Phytol. 2005, 65, 191–202. [Google Scholar] [CrossRef] [PubMed]
- Mandal, B.; Mandal, S.; Csinos, A.; Martinez, N.; Culbreath, A.; Pappu, H. Biological and molecular analyses of the acibenzolar-S-methyl-induced systemic acquired resistance in flue-cured tobacco against Tomato spotted wilt virus. Phytopathology 2008, 98, 196–204. [Google Scholar] [CrossRef] [PubMed]
- Gozzo, F.; Faoro, F. Systemic acquired resistance (50 years after discovery): moving from the lab to the field. J. Agr. Food Chem. 2013, 61, 12473–12491. [Google Scholar] [CrossRef]
- Cole, D.L. The efficacy of acibenzolar-S-methyl, an inducer of systemic acquired resistance, against bacterial and fungal diseases of tobacco. Crop Prot. 1999, 18, 267–273. [Google Scholar] [CrossRef]
- Frąckowiak, P.; Pospieszny, H.; Smiglak, M.; Obrępalska-Stęplowska, A. Assessment of the efficacy and mode of action of benzo (1, 2, 3)-thiadiazole-7-carbothioic acid s-methyl ester (bth) and its derivatives in plant protection against viral disease. Int. J. Mol. Sci. 2019, 20, 1598. [Google Scholar] [CrossRef] [PubMed]
- Gozzo, F. Systemic acquired resistance in crop protection: from nature to a chemical approach. J. Agr. Food Chem. 2003, 51, 4487–4503. [Google Scholar] [CrossRef] [PubMed]
- Vidhyasekaran, P. ; Bioengineering and molecular manipulation of salicylic acid signaling system to activate plant immune responses for crop disease management. Plant innate immunity signals and signaling systems: bioengineering and molecular manipulation for crop disease management.; Springer: Dordrecht, 2020. [Google Scholar] [CrossRef]
- Schreiber, K.; Desveaux, D. Message in a bottle: chemical biology of induced disease resistance in plants. Plant Pathology J. 2008, 24, 245–268. [Google Scholar] [CrossRef]
- Yang, H.; Wang, S.; Xi, D.; Yuan, S.; Wang, J.; Xu, M.; Lin, H. Interaction between Cucumber mosaic virus and Turnip crinkle virus in Arabidopsis thaliana. J. Phytopathol. 2010, 158, 833–836. [Google Scholar] [CrossRef]
- Zhang, C.; Tang, C.; Wang, Q.; Su, Y.; Zhang, Q. Synergistic effects of oligochitosan and pyraclostrobin in controlling leaf spot disease in Pseudostellaria heterophylla. Antibiotics 2024, 13, 128. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Zhang, Y.; Lin, L.; Ying, S.; Yu, C. Cucumber PGIP2 is involved in resistance to gray mold disease. Gene 2024, 923, 148588. [Google Scholar] [CrossRef]
- Yang, D.; Zhang, X.; Li, Z.; Chu, R.; Shah, S.; Wang, X.; Zhang, X. Antagonistic effect of Bacillus and Pseudomonas combinations against Fusarium oxysporum and their effect on disease resistance and growth promotion in watermelon. J. Appl. Microbiol. 2024, 135, lxae074. [Google Scholar] [CrossRef] [PubMed]
- Tian, B.; Qu, Z.; Mehmood, M.A.; Xie, J.; Cheng, J.; Fu, Y.; Jiang, D. Schizotrophic Sclerotinia sclerotiorum-mediated root and rhizosphere microbiome alterations activate growth and disease resistance in wheat. Microbiol. Spectr. 2023, 11, e00981–00923. [Google Scholar] [CrossRef] [PubMed]
- Zhai, Q.; Pan, Z.; Zhang, C.; Yu, H.; Zhang, M.; Gu, X.; Zhang, X.; Pan, H.; Zhang, H. Colonization by Klebsiella variicola FH-1 stimulates soybean growth and alleviates the stress of Sclerotinia sclerotiorum. J. Integr. Agr. 2023, 22, 2729–2745. [Google Scholar] [CrossRef]
- Molinari, S. Systemic acquired resistance activation in solanaceous crops as a management strategy against root-knot nematodes. Pest Manag. Sci. 2016, 72, 888–896. [Google Scholar] [CrossRef] [PubMed]
- Chicowski, A.S.; Qi, M.; Variz, H.; Bredow, M.; Montes-Serey, C.; Caiazza, F.; Dong, H.; Margets, A.C.; Mejias, J.; Walley, J. A soybean rust effector protease suppresses host immunity and cleaves a 3-deoxy-7-phosphoheptulonate synthase. Plant biol. bioRxiv. 2023, 556260. [Google Scholar] [CrossRef]
- Abdel-Monem, M.O.; Azeiz, A.Z.A.; Khalil, R.; Elsayed, N.A.; Hassan, M.G. Effect of rhizosphere bacteria to reduce Fusarium infection in tomato plant. J. Basic Environ. Sci. 2024, 11, 166–182. [Google Scholar] [CrossRef]
- Abbey, J.; Jose, S.; Percival, D.; Jaakola, L.; Asiedu, S.K. Modulation of defense genes and phenolic compounds in wild blueberry in response to Botrytis cinerea under field conditions. BMC Plant Biol. 2023, 23, 117. [Google Scholar] [CrossRef] [PubMed]
- Burketová, L.; Štillerová, K.; Feltlová, M. Immunohistological localization of chitinase and β-1, 3-glucanase in rhizomania-diseased and benzothiadiazole treated sugar beet roots. Physiol. Mol. Plant Pathol. 2003, 63, 47–54. [Google Scholar] [CrossRef]
- Lawton, K.A.; Friedrich, L.; Hunt, M.; Weymann, K.; Delaney, T.; Kessmann, H.; Staub, T.; Ryals, J. Benzothiadiazole induces disease resistance in Arabidopsis thaliana by activation of the systemic acquired resistance signal transduction pathway. Plant J. 1996, 10, 71–82. [Google Scholar] [CrossRef]
- Zhao, S.; Li, M.; Ren, X.; Wang, C.; Sun, X.; Sun, M.; Yu, X.; Wang, X. Enhancement of broad-spectrum disease resistance in wheat through key genes involved in systemic acquired resistance. Front. Plant Sci. 2024, 15, 1355178. [Google Scholar] [CrossRef] [PubMed]
- Kohler, A.; Schwindling, S.; Conrath, U. Benzothiadiazole-induced priming for potentiated responses to pathogen infection, wounding, and infiltration of water into leaves requires the NPR1/NIM1 gene in Arabidopsis thaliana. Plant Physiol. 2002, 128, 1046–1056. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, Y.; Fukuzawa, N.; Hyodo, A.; Kim, H.; Mashiyama, S.; Ogihara, T.; Yoshioka, H.; Matsuura, H.; Masuta, C.; Matsumura, T. Role of salicylic acid glucosyltransferase in balancing growth and defence for optimum plant fitness. Mol. Plant Pathol. 2020, 21, 429–442. [Google Scholar] [CrossRef] [PubMed]
- Shimono, M.; Koga, H.; Akagi, A.; Hayashi, N.; Goto, S.; Sawada, M.; Kurihara, T.; Matsushita, A.; Sugano, S.; Jiang, C.J. Rice WRKY45 plays important roles in fungal and bacterial disease resistance. Mol. Plant Pathol. 2012, 13, 83–94. [Google Scholar] [CrossRef] [PubMed]
- Toquin, V.; Braun, C.A.; Sirven, C.; Assmann, L.; Sawada, H. Host defense inducers. Research & Development Crop Science, Small Molecules, Pest ControlChemistry, Crop Prot. Compounds., third Edition Insecticides.; Wiley-VCH: Germany, 2019, 2, 959–978. [Google Scholar] [CrossRef]
- Yasuda, R.; Sabatini, B.L.; Svoboda, K. Plasticity of calcium channels in dendritic spines. Nat. Neurosci. 2003, 6, 948–955. [Google Scholar] [CrossRef] [PubMed]
- Van Daele, J.; Blancquaert, D.; Kiekens, F.; Van Der Straeten, D.; Lambert, W.E.; Stove, C.P. Degradation and interconversion of plant pteridines during sample preparation and ultra-high performance liquid chromatography tandem mass spectrometry. Food Chem. 2016, 194, 1189–1198. [Google Scholar] [CrossRef] [PubMed]
- Nakashita, H.; Yoshioka, K.; Yasuda, M.; Nitta, T.; Arai, Y.; Yoshida, S.; Yamaguchi, I. Probenazole induces systemic acquired resistance in tobacco through salicylic acid accumulation. Physiol. Mol. Plant Pathol. 2002, 61, 197–203. [Google Scholar] [CrossRef]
- Saint-Macary, M.E.; Barbisan, C.; Gagey, M.J.; Frelin, O.; Beffa, R.; Lebrun, M.H.; Droux, M. Methionine biosynthesis is essential for infection in the rice blast fungus Magnaporthe oryzae. PLoS ONE 2015, 10, e0111108. [Google Scholar] [CrossRef]
- Nakashita, H.; Yasuda, M.; Nishioka, M.; Hasegawa, S.; Arai, Y.; Uramoto, M.; Yoshida, S.; Yamaguchi, I. Chloroisonicotinamide derivative induces a broad range of disease resistance in rice and tobacco. Plant Cell physiol. 2002, 43, 823–831. [Google Scholar] [CrossRef] [PubMed]
- Nawrath, C.; Métraux, J.P. Salicylic acid induction–deficient mutants of Arabidopsis thaliana express PR-2 and PR-5 and accumulate high levels of camalexin after pathogen inoculation. Plant Cell. 1999, 11, 1393–1404. [Google Scholar] [CrossRef] [PubMed]
- Pokotylo, I.; Kravets, V.; Ruelland, E. Salicylic acid binding proteins (SABPs): the hidden forefront of salicylic acid signalling. Int. J. Mol. Sci. 2019, 20, 4377. [Google Scholar] [CrossRef] [PubMed]
- Chandía, N.P.; Matsuhiro, B. Characterization of a fucoidan from Lessonia vadosa (Phaeophyta) and its anticoagulant and elicitor properties. Int. J. Biol. Macromol. 2008, 42, 235–240. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Wei, H.; Yao, K.; Wu, H.; Huang, T.; Han, M.; Su, T.; Cao, F. Integrative omics studies revealed synergistic link between sucrose metabolic isogenes and carbohydrates in poplar roots infected by Fusarium wilt. Plant Mol. Biol. 2024, 114, 29. [Google Scholar] [CrossRef] [PubMed]
- El-Sharkawy, E.E.; ElSharawy, A.A. The Influence of resistance chemical inducers against anthracnose on cucumber and pepper plants. Catrina 2024, 29, 57–67. [Google Scholar] [CrossRef]
- Thepbandit, W.; Athinuwat, D. Rhizosphere microorganisms supply availability of soil nutrients and induce plant defense. Microorganisms 2024, 12, 558. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Zhao, J.; Liu, Y.; Huang, S.; Zhao, C.; Jiang, Z.; Gu, Y.; Xiao, J.; Wu, Y.; Ying, R. Decipher soil resistance and virulence gene risks in conventional and organic farming systems. J. Hazard. Mater. 2024, 133788. [Google Scholar] [CrossRef] [PubMed]
- Dass, A.; Venkatesh, P.; Choudhary, A.K.; Upadhyay, P.K.; Chandrashekar, A.B.; Francaviglia, R.; Sannagoudar, M.S.; Rangappa, U.M.; Singh, V.K.; Harish, M.N. Soil moisture dynamics, rooting traits, crop and water productivity of wheat under different tillage, irrigation and nutrition conditions. Farming System 2024, 2, 100087. [Google Scholar] [CrossRef]
- Domínguez-Figueroa, J.; Gómez-Rojas, A.; Escobar, C. Functional studies of plant transcription factors and their relevance in the plant root-knot nematode interaction. Front. Plant Sci. 2024, 15, 1370532. [Google Scholar] [CrossRef] [PubMed]
- Riseh, R.S.; Vazvani, M.G.; Vatankhah, M.; Kennedy, J.F. Chitin-induced disease resistance in plants: A review. Int. J. Biol. Macromol. 2024, 131105. [Google Scholar] [CrossRef]
- Mahmoud, R.M.; Fouad, M.S. Approaches to antioxidant defence system: An overview of coping mechanism against lithium/nickel exposure in plants. Lithium and Nickel Contamination in Plants Environ, 2024, 95–138. [CrossRef]
- Yang, T.; Zhao, X.; Bai, J.; Lv, W.; Chen, Q.; Hu, J.; Liu, G.; Zhao, Y.; Zhou, H.; Zhao, M. Transcriptome analysis of genes involved in the pathogenesis mechanism of potato virus Y in potato cultivar YouJin. Front. Microbiol. 2024, 15, 1353814. [Google Scholar] [CrossRef] [PubMed]
- Berlanga, D.J.; Molina, A.; Torres, M.Á. Mitogen-activated protein kinase phosphatase 1 controls broad spectrum disease resistance in Arabidopsis thaliana through diverse mechanisms of immune activation. Front. Plant Sci. 2024, 15, 1374194. [Google Scholar] [CrossRef] [PubMed]
- Yun, S.H.; Khan, I.U.; Noh, B.; Noh, Y.S. Genomic overview of INA-induced NPR1 targeting and transcriptional cascades in Arabidopsis. Nucleic Aci. Res. 2024, gkae019. [Google Scholar] [CrossRef]
- Amokrane, L.; Pokotylo, I.; Acket, S.; Ducloy, A.; Troncoso-Ponce, A.; Cacas, J.L.; Ruelland, E. Phospholipid signaling in crop plants: a field to explore. Plants 2024, 13, 1532. [Google Scholar] [CrossRef]
- Tsubata, K.; Kuroda, K.; Yamamoto, Y.; Yasokawa, N. Development of a novel plant activator for rice diseases, tiadinil. J. Pestic. Sci. 2006, 31, 174–181. [Google Scholar] [CrossRef]
- Hu, Y.; Li, C.Y.; Wang, X.M.; Yang, Y.H.; Zhu, H.L. 1, 3, 4-Thiadiazole: synthesis, reactions, and applications in medicinal, agricultural, and materials chemistry. Chem. Rev. 2014, 114, 5572–5610. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, K.; Ogino, A.; Yamada, K.; Sonoda, R. Induction of disease resistance in tea (Camellia sinensis L.) by plant activators. JARQ-Jpn. Agr. Res. Q. 2010, 44, 391–398. [Google Scholar] [CrossRef]
- Maeda, T.; Ishiwari, H. Tiadinil, a plant activator of systemic acquired resistance, boosts the production of herbivore-induced plant volatiles that attract the predatory mite Neoseiulus womersleyi in the tea plant Camellia sinensis. Exp. Appl. Acarol. 2012, 58, 247–258. [Google Scholar] [CrossRef] [PubMed]
- Qi, X.; Li, K.; Chen, L.; Zhang, Y.; Zhang, N.; Gao, W.; Li, Y.; Liu, X.; Fan, Z. Plant defense responses to a novel plant elicitor candidate LY5-24-2. Int. J. Mol. Sci. 2022, 23, 5348. [Google Scholar] [CrossRef] [PubMed]
- Yasuda, R.; Harvey, C.D.; Zhong, H.; Sobczyk, A.; Van Aelst, L.; Svoboda, K. Supersensitive ras activation in dendrites and spines revealed by two-photon fluorescence lifetime imaging. Nat. Neurosci. 2006, 9, 283–291. [Google Scholar] [CrossRef] [PubMed]
- Nakashita, H. Studies on regulation of plant physiology by pesticides. J. Pestic. Sci. 2021, 46, 393–398. [Google Scholar] [CrossRef] [PubMed]
- Yasuda, M.; Nishioka, M.; Nakashita, H.; Yamaguchi, I.; Yoshida, S. Pyrazolecarboxylic acid derivative induces systemic acquired resistance in tobacco. Biosci. Biotechnol. Biochem. 2003, 67, 2614–2620. [Google Scholar] [CrossRef] [PubMed]
- Nishioka, M.; Nakashita, H.; Yasuda, M.; Yoshida, S.; Yamaguchi, I. Induction of resistance against rice bacterial leaf blight by 3-Chloro-1-methyl-1H-pyrazole-5-carboxylic acid. J. Pestic. Sci. 2005, 30, 47–49. [Google Scholar] [CrossRef]
- Alexandersson, E.; Mulugeta, T.; Lankinen, Å.; Liljeroth, E.; Andreasson, E. Plant resistance inducers against pathogens in Solanaceae species from molecular mechanisms to field application. Int. J. Mol. Sci. 2016, 17, 1673. [Google Scholar] [CrossRef] [PubMed]
- Mishra, S.; Roychowdhury, R.; Ray, S.; Hada, A.; Kumar, A.; Sarker, U.; Aftab, T.; Das, R. Salicylic acid (SA)-mediated plant immunity against biotic stresses: an insight on molecular components and signaling mechanism. Plant Stress 2024, 100427. [Google Scholar] [CrossRef]
- Shalaby, T.A.; Taha, N.A.; Taher, D.I.; Metwaly, M.M.; El-Beltagi, H.S.; Rezk, A.A.; El-Ganainy, S.M.; Shehata, W.F.; El-Ramady, H.R.; Bayoumi, Y.A. Paclobutrazol improves the quality of tomato seedlings to be resistant to Alternaria solani blight disease: biochemical and histological perspectives. Plants 2022, 11, 425. [Google Scholar] [CrossRef] [PubMed]
- Zhu, F.; Xi, D.H.; Yuan, S.; Xu, F.; Zhang, D.W.; Lin, H.H. Salicylic acid and jasmonic acid are essential for systemic resistance against tobacco mosaic virus in Nicotiana benthamiana. Mol. Plant Microbe Interact 2014, 27, 567–577. [Google Scholar] [CrossRef] [PubMed]
- Mur, L.A.; Naylor, G.; Warner, S.A.; Sugars, J.M.; White, R.F.; Draper, J. Salicylic acid potentiates defence gene expression in tissue exhibiting acquired resistance to pathogen attack. Plant J. 1996, 9, 559–571. [Google Scholar] [CrossRef]
- Xu, X.; Chen, Y.; Li, B.; Zhang, Z.; Qin, G.; Chen, T.; Tian, S. Molecular mechanisms underlying multi-level defense responses of horticultural crops to fungal pathogens. Hortic. Res. 2022, 9, uhac066. [Google Scholar] [CrossRef] [PubMed]
- Kamatham, S.; Neela, K.B.; Pasupulati, A.K.; Pallu, R.; Singh, S.S.; Gudipalli, P. Benzoylsalicylic acid isolated from seed coats of Givotia rottleriformis induces systemic acquired resistance in tobacco and Arabidopsis thaliana. Phytochemistry 2016, 126, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Kamatham, S.; Pallu, R.; Pasupulati, A.K.; Singh, S.S.; Gudipalli, P. Benzoylsalicylic acid derivatives as defense activators in tobacco and Arabidopsis thaliana. Phytochemistry 2017, 143, 160–169. [Google Scholar] [CrossRef] [PubMed]
- Cai, Z.; Guo, H.; Shen, S.; Yu, Q.; Wang, J.; Zhu, E.; Zhang, P.; Song, L.; Zhang, Y.; Zhang, K. Generation of the salicylic acid deficient Arabidopsis thaliana via a synthetic salicylic acid hydroxylase expression cassette. Plant Methods 2022, 18, 89. [Google Scholar] [CrossRef] [PubMed]
- Xie, Z.; Chen, Z. Salicylic acid induces rapid inhibition of mitochondrial electron transport and oxidative phosphorylation in tobacco cells. Plant Physiol. 1999, 120, 217–226. [Google Scholar] [CrossRef]
- White, R.F. Acetylsalicylic acid (aspirin) induces resistance to tobacco mosaic virus in tobacco. Virology 1979, 99, 410–412. [Google Scholar] [CrossRef] [PubMed]
- Saberi Riseh, R.; Gholizadeh Vazvani, M. Unveiling methods to stimulate plant resistance against pathogens. Front. Biosci. (Landmark Ed.) IMR Press: 2024, 29, 188. [Google Scholar] [CrossRef] [PubMed]
- Bektas, Y.; Rodriguez-Salus, M.; Schroeder, M.; Gomez, A.; Kaloshian, I.; Eulgem, T. The synthetic elicitor DPMP (2, 4-dichloro-6-{(E)-[(3-methoxyphenyl)imino]methyl}phenol) triggers strong immunity in Arabidopsis thaliana and tomato. Sci. Rep. 2016, 6, 29554. [Google Scholar] [CrossRef] [PubMed]
- Yoo, S.-J.; Choi, H.J.; Noh, S.W.; Cecchini, N.M.; Greenberg, J.T.; Jung, H.W. Genetic requirements for infection-specific responses in conferring disease resistance in Arabidopsis. Front. Plant Sci. 2022, 13, 1068438. [Google Scholar] [CrossRef] [PubMed]
- Palmer, I.A.; Chen, H.; Chen, J.; Chang, M.; Li, M.; Liu, F.; Fu, Z.Q. Novel salicylic acid analogs induce a potent defense response in Arabidopsis. Int. J. Mol. Sci. 2019, 20, 3356. [Google Scholar] [CrossRef] [PubMed]
- Urban, L.; Lauri, F.; Ben Hdech, D.; Aarrouf, J. Prospects for increasing the efficacy of plant resistance inducers stimulating salicylic acid. Agronomy 2022, 12, 3151. [Google Scholar] [CrossRef]
- Knoth, C.; Salus, M.S.; Girke, T.; Eulgem, T. The synthetic elicitor 3, 5-dichloroanthranilic acid induces NPR1-dependent and NPR1-independent mechanisms of disease resistance in Arabidopsis thaliana. Plant Physiol. 2009, 150, 333–347. [Google Scholar] [CrossRef]
- Zhang, N.; Zhou, S.; Yang, D.; Fan, Z. Revealing shared and distinct genes responding to JA and SA signaling in Arabidopsis thaliana by meta-analysis. Front. Plant Sci. 2020, 11, 512053. [Google Scholar] [CrossRef]
- Stenberg, J.A. A conceptual framework for integrated pest management. Trends Plant Sci. 2017, 22, 759–769. [Google Scholar] [CrossRef] [PubMed]
- Jackson, M.P.; Vendeville, B.C.; Schultz-Ela, D.D. Structural dynamics of salt systems. Annu. Rev. Earth Pl. Sc. 1994, 22, 93–117. [Google Scholar] [CrossRef]
- Yang, Z. Small GTPases: versatile signaling switches in plants. Plant Cell 2002, 14, S375–S388. [Google Scholar] [CrossRef] [PubMed]
- Vidhyasekaran, P. Manipulation of reactive oxygen species, redox and nitric oxide signaling systems to activate plant innate immunity for crop disease management. In: plant innate immunity signals and signaling Systems: Bioengineering and molecular manipulation for crop disease management. Springer, 2020, 51–135. [CrossRef]
- Yang, H.; Chen, X.; Yang, R.; Cheng, J.; Chen, Y.; Joosten, M.H.; Du, Y. The potato StMKK5-StSIPK module enhances resistance to Phytophthora pathogens through activating the salicylic acid and ethylene signalling pathways. Mol. Plant Pathol. 2023, 24, 399–412. [Google Scholar] [CrossRef]
- van Loon, L.C.; Rep, M.; Pieterse, C.M. Significance of inducible defense-related proteins in infected plants. Annu. Rev. Phytopathol. 2006, 44, 135–162. [Google Scholar] [CrossRef]
- Carr, J.P.; Lewsey, M.G.; Palukaitis, P. Signaling in induced resistance. Adv. Virus Res. 2010, 76, 57–121. [Google Scholar] [CrossRef]
- de Souza Cândido, E.; Pinto, M.F.S.; Pelegrini, P.B.; Lima, T.B.; Silva, O.N.; Pogue, R.; Grossi-de-Sá, M.F.; Franco, O.L. Plant storage proteins with antimicrobial activity: novel insights into plant defense mechanisms. FASEB J. 2011, 25, 3290–3305. [Google Scholar] [CrossRef] [PubMed]
- AbuQamar, S.; Moustafa, K.; Tran, L.S. Mechanisms and strategies of plant defense against Botrytis cinerea. Crit. Rev. Biotechnol. 2017, 37, 262–274. [Google Scholar] [CrossRef]
- Zeier, J. New insights into the regulation of plant immunity by amino acid metabolic pathways. Plant Cell Environ. 2013, 36, 2085–2103. [Google Scholar] [CrossRef] [PubMed]
- de Borba, M.C.; Velho, A.C.; de Freitas, M.B.; Holvoet, M.; Maia-Grondard, A.; Baltenweck, R.; Magnin-Robert, M.; Randoux, B.; Hilbert, J.L.; Reignault, P. A laminarin-based formulation protects wheat against Zymoseptoria tritici via direct antifungal activity and elicitation of host defense-related genes. Plant Dis. 2022, 106, 1408–1418. [Google Scholar] [CrossRef]
- Coqueiro, D.S.O.; de Souza, A.A.; Takita, M.A.; Rodrigues, C.M.; Kishi, L.T.; Machado, M.A. Transcriptional profile of sweet orange in response to chitosan and salicylic acid. BMC Genomics 2015, 16, 1–14. [Google Scholar] [CrossRef]
- Xing, K.; Zhu, X.; Peng, X.; Qin, S. Chitosan antimicrobial and eliciting properties for pest control in agriculture: a review. Agrono. Sustain. Dev. 2015, 35, 569–588. [Google Scholar] [CrossRef]
- Rabiei, Z.; Hosseini, S.; Dehestani, A.; Pirdashti, H.; Beiki, F. Exogenous hexanoic acid induced primary defense responses in tomato (Solanum lycopersicum L.) plants infected with Alternaria Solani. Sci. Hortic. 2022, 295, 110841. [Google Scholar] [CrossRef]
- Djami-Tchatchou, A.T.; Ncube, E.N.; Steenkamp, P.A.; Dubery, I.A. Similar, but different: structurally related azelaic acid and hexanoic acid trigger differential metabolomic and transcriptomic responses in tobacco cells. BMC Plant Biol. 2017, 17, 1–15. [Google Scholar] [CrossRef]
- Adigun, O.A.; Nadeem, M.; Pham, T.H.; Jewell, L.E.; Cheema, M.; Thomas, R. Recent advances in bio-chemical, molecular and physiological aspects of membrane lipid derivatives in plant pathology. Plant Cell Environ. 2021, 44, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Jiadkong, K.; Fauzia, A.N.; Yamaguchi, N.; Ueda, A. Exogenous riboflavin (vitamin B2) application enhances salinity tolerance through the activation of its biosynthesis in rice seedlings under salinity stress. Plant Sci. 2024, 339, 111929. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Xu, X.; Xie, J.; Yu, W.; Zhang, S.; Wu, H.; Peng, Y.; Ji, H. P-aminobenzoic acid and vitamin B9 enhance rice defence against sheath blight disease caused by Rhizoctonia solani. Plant Pathol. 2024, 73, 90–103. [Google Scholar] [CrossRef]
- Ghallab, D.S.; Shawky, E.; Khalifa, A.A.; Ibrahim, R.S. Insights into the molecular mechanisms of spirulina platensis against rheumatoid arthritis through integrative serum pharmacochemistry and network pharmacology analysis. Food Biosci. 2024, 59, 103902. [Google Scholar] [CrossRef]
- Saleh, R.O.; Al-Ouqaili, M.T.; Ali, E.; Alhajlah, S.; Kareem, A.H.; Shakir, M.N.; Alasheqi, M.Q.; Mustafa, Y.F.; Alawadi, A.; Alsaalamy, A. lncRNA-microRNA axis in cancer drug resistance: particular focus on signaling pathways. Med. Oncol. 2024, 41, 52. [Google Scholar] [CrossRef] [PubMed]
- 243. Reglinski, T.; Havis, N.; Rees, H.J.; de Jong, H. The practical role of induced resistance for crop protection. Phytopathology 2023, 113, 719–731. [Google Scholar] [CrossRef]
- Khokhar, M.; Sharma, S.; Gupta, R. Influence of sowing dates on incidence and severity of post flowering stalk rot of maize caused by Fusarium verticillioides. J. Mycol. Plant Pathol. 2014, 44, 205. [Google Scholar]
- Verma, G.; Khosa, J.; Sharma, M.; Meena, O.; Dhatt, A. Phenotypic and molecular characterization of vernalization sensitivity and pre-mature bolting in onion (Allium cepa L.). Genet. Resour. Crop Evol. [CrossRef]
- Giusti, P.; Vendramin, A.; Zusso, M. The penicillin saga: a different tale. Top Italian Scientists Journal 2024, 1. [Google Scholar] [CrossRef]
- Virk, V.; Deepak, H.; Taneja, K.; Srivastava, R.; Giri, S. Amelioration in nanobiosensors for the control of plant diseases: current status and future challenges. Front. Nanotechnol. 2024; 6, 1310165. [CrossRef]
- Gouveia, C.; Santos, R.B.; Paiva-Silva, C.; Buchholz, G.; Malhó, R.; Figueiredo, A. The pathogenicity of Plasmopara viticola: a review of evolutionary dynamics, infection strategies and effector molecules. BMC Plant Biol. 2024, 24, 327. [Google Scholar] [CrossRef] [PubMed]
- Jemil, N.; Besbes, I.; Gharbi, Y.; Triki, M.A.; Cheffi, M.; Manresa, A.; Nasri, M.; Hmidet, N. Bacillus methylotrophicus DCS1: production of different lipopeptide families, In vitro antifungal activity and suppression of Fusarium wilt in tomato plants. Curr. Microbiol. 2024, 81, 142. [Google Scholar] [CrossRef] [PubMed]
- Harris, F.M.; Mou, Z. Damage-associated molecular patterns and systemic signaling. Phytopathology 2024, 114, 308–327. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.M.; El-Sappah, A.H.; Ali, H.M.; Zandi, P.; Huang, Q.; Soaud, S.A.; Alazizi, E.M.Y.; Wafa, H.A.; Hossain, M.A.; Liang, Y. Pathogenesis-related proteins (PRs) countering environmental stress in plants: a review. S. Afr. J. Bot. 2023, 160, 414–427. [Google Scholar] [CrossRef]
- Wei, H.; Tang, M.; Xu, X. Mechanism of uptake, accumulation, transport, metabolism and phytotoxic effects of pharmaceuticals and personal care products within plants: a review. Sci. Total Environ. 2023, 892, 164413. [Google Scholar] [CrossRef] [PubMed]
- Sajid, Q.ul. ain.; Elçi, E. Knockdown of orthotospovirus-derived silencing suppressor gene by plant-mediated RNAi approach induces viral resistance in tomato. Physiol. Mol. Plant Pathol. 2024, 131, 102264. [Google Scholar] [CrossRef]
- Cartwright, D.; Langcake, P.; Pryce, R.J.; Leworthy, D.P.; Ride, J.P. Chemical activation of host defence mechanisms as a basis for crop protection. Nature 1977, 267, 511–513. [Google Scholar] [CrossRef]
- Zheng, C.; Wang, X.; Xu, Y.; Wang, S.; Jiang, X.; Liu, X.; Cui, W.; Wu, Y.; Yan, C.; Liu, H.; et al. The peroxidase gene OsPrx114 activated by OsWRKY50 enhances drought tolerance through ROS scavenging in rice. Plant Physiol. Bioch. 2023, 204, 108138. [Google Scholar] [CrossRef]
- Zhang, H.; Yang, J.; Liu, M.; Xu, X.; Yang, L.; Liu, X.; Peng, Y.; Zhang, Z. Early molecular events in the interaction between Magnaporthe oryzae and rice. Phytopathol. Res. 2024, 6, 9. [Google Scholar] [CrossRef]
- Sharma, A.; Sharma, A.; Sharma, A.; Kumar, Y.; Sharma, P.; Bhardwaj, R.; Sharma, I. Polyphenol phytoalexins as the determinants of plant disease resistance. In Plant phenolics in biotic stress management; Springer, 2024; 243–274. [CrossRef]
- Rayee, R.; Anh, L.H.; Khanh, T.D.; Xuan, T.D. Potential momilactones in rice stress tolerance and health advantages. Agronomy 2024, 14, 405. [Google Scholar] [CrossRef]
- Arai, H.; Legeai, F.; Kageyama, D.; Sugio, A.; Simon, J.C. Genomic insights into spiroplasma endosymbionts that induce male-killing and protective phenotypes in the pea aphid. FEMS Microbiol. Lett. 2024, 371, fnae027. [Google Scholar] [CrossRef] [PubMed]
- Shikina, E.V.; Kovalevsky, R.A.; Shirkovskaya, A.I.; Toukach, P.V. Prospective bacterial and fungal sources of hyaluronic acid: a review. Comput. Struct. Biotec. 2022, 20, 6214–6236. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Lorente, S.E.; Martí-Guillén, J.M.; Pedreño, M.Á.; Almagro, L.; Sabater-Jara, A.B. Higher plant-derived biostimulants: mechanisms of action and their role in mitigating plant abiotic stress. Antioxidants 2024, 13, 318. [Google Scholar] [CrossRef] [PubMed]
- Park, K.; Paul, D.; Kim, E.; Kloepper, J.W. Hyaluronic acid of Streptococcus sp. as a potent elicitor for induction of systemic resistance against plant diseases. World J. Microbio. Biot. 2008, 24, 1153–1158. [Google Scholar] [CrossRef]
- Sultana, F.; Hossain, M.M.; Kubota, M.; Hyakumachi, M. Elicitation of systemic resistance against the bacterial speck pathogen in Arabidopsis thaliana by culture filtrates of plant growth-promoting fungi. Can. J. Plant Pathol. 2008, 30, 196–205. [Google Scholar] [CrossRef]
- Cui, H.; Qiu, J.; Zhou, Y.; Bhandari, D.D.; Zhao, C.; Bautor, J.; Parker, J.E. Antagonism of transcription factor MYC2 by EDS1/PAD4 complexes bolsters salicylic acid defense in Arabidopsis thaliana effector-triggered immunity. Mol. Plant 2018, 11, 1053–1066. [Google Scholar] [CrossRef] [PubMed]
- Jarecka-Boncela, A.; Spychalski, M.; Ptaszek, M.; Włodarek, A.; Smiglak, M.; Kukawka, R. The effect of a new derivative of benzothiadiazole on the reduction of fusariosis and increase in growth and development of tulips. Agriculture 2023, 13, 853. [Google Scholar] [CrossRef]
- Xu, Y.; Zhao, Z.; Qian, X.; Qian, Z.; Tian, W.; Zhong, J. Novel, unnatural benzo-1, 2, 3-thiadiazole-7-carboxylate elicitors of taxoid biosynthesis. J. Agr. Food Chem. 2006, 54, 8793–8798. [Google Scholar] [CrossRef]
- Xu, R.; Song, F.; Zheng, Z. OsBISAMT1, a gene encoding S-adenosyl-L-methionine: salicylic acid carboxyl methyltransferase, is differentially expressed in rice defense responses. Mol. Biol. Rep. 2006, 33, 223–231. [Google Scholar] [CrossRef] [PubMed]
- Du, Q.S.; Zhao, Z.J.; Zhu, W.P.; Xu, Y.F.; Qian, X.H. Synthesis and biological activity of novel benzo-1, 2, 3-thiadiazole carboxylic ester derivatives. Chin. J. Pestic. Sci.. 2010, 12, 499–502. [Google Scholar]
- Gan, X.; Hu, D.; Wang, Y.; Yu, L.; Song, B. Novel trans-ferulic acid derivatives containing a chalcone moiety as potential activator for plant resistance induction. J. Agr. Food Chem. 2017, 65, 4367–4377. [Google Scholar] [CrossRef] [PubMed]
- Ryu, C.M.; Hu, C.H.; Reddy, M.; Kloepper, J.W. Different signaling pathways of induced resistance by rhizobacteria in Arabidopsis thaliana against two pathovars of Pseudomonas syringae. New Phytol. 2003, 160, 413–420. [Google Scholar] [CrossRef] [PubMed]
- Sun, T.J.; Lu, Y.; Narusaka, M.; Shi, C.; Yang, Y.B.; Wu, J.X.; Zeng, H.Y.; Narusaka, Y.; Yao, N. A novel pyrimidin-like plant activator stimulates plant disease resistance and promotes growth. PLoS ONE 2015, 10, e0123227. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhang, Z.; Jia, Y.; Shen, Y.; He, H.; Fang, R.; Chen, X.; Hao, X. 3-Acetonyl-3-hydroxyoxindole: a new inducer of systemic acquired resistance in plants. Plant Biotechnol. J. 2008, 6, 301–308. [Google Scholar] [CrossRef] [PubMed]
- Bashir, A.; Nabi, M.; Tabassum, N.; Afzal, S.; Ayoub, M. An updated review on phytochemistry and molecular targets of Withania somnifera (L.) Dunal (Ashwagandha). Front. Pharmacol. 2023, 14, 1049334. [Google Scholar] [CrossRef]
- Li, Q.; Zhu, H.; Ai, G.; Yu, J.; Dou, D. Plant genes related to phytophthora pathogens resistance. Phytopathol. Res. 2024, 6, 15. [Google Scholar] [CrossRef]
- Wei, X.; Liu, X.; Zhang, X.; Guo, S.; Shi, J. Structural insights into ligand recognition and receptor activation of plant leucine-rich repeat (LRR) transmembrane receptors. New Crops 2024, 100022. [Google Scholar] [CrossRef]
- Hu, R.; Teng, X.; Li, Y. Unleashing plant synthetic capacity: navigating regulatory mechanisms for enhanced bioproduction and secondary metabolite discovery. Curr. Opin. Biotech. 2024, 88, 103148. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Xu, Y.; Li, Q.; Cao, Y.; Yang, D.; Liu, S.; Wang, X.; Mi, Y.; Liu, Y.; Ding, C. A lncRNA fine-tunes salicylic acid biosynthesis to balance plant immunity and growth. Cell Host Microbe 2022, 30, 1124–1138. [Google Scholar] [CrossRef] [PubMed]
- Huang, J. Zhou, W.; Zhang, X.; Li, Y. Roles of long non-coding RNAs in plant immunity. PLoS Pathog. 2023, 19, e1011340. [Google Scholar] [CrossRef]
- Wu, L.; Liu, S.; Qi, H.; Cai, H.; Xu, M. Research progress on plant long non-coding RNA. Plants 2020, 9, 408. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Huang, H. K.; Xia, Z. Q.; Yang, Y. Q.; Jiang, X. Y.; Yang, Y. Y.; Wang, D. L.; Li, X. L.; Chen, Z. Sequence data, functional annotation, and relationship analysis between mRNAs and long non-coding RNAs from tea leaves during infection by the fungal pathogen Epicoccum sorghinum. Mol. Plant Microbe Interact. 2022, 35, 875–879. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Yang, Y. Y.; Jiang, X. Y.; Yang, Y. Q.; Jiang, S. L.; Guo, D.; Li, Z.; Wang. D. L. ; Chen, Z. The sequence and integrated analysis of competing endogenous RNAs originating from tea leaves infected by the pathogen of tea leaf spot, Didymella segeticola. Plant Dis. 2022, 106, 1286–1290. [Google Scholar] [CrossRef] [PubMed]
- Huang, H. L.; Huang, H. K.; Xia, Z. Q.; Yang, Y. Y.; Jiang, X. Y.; Huang, C.; Yang, Y. Y.; Wang. D. L.; Chen, Z. Sequencing, functional annotation, and interaction prediction of mRNAs and candidate long non-coding RNAs originating from tea leaves during infection by the fungal pathogen causing tea leaf spot, Didymella bellidis. Didymella bellidis. Plant Dis. 2023, 107, 2830–2834.
- Yang, X.; Zhang, L.; Yang, Y.; Schmid, M.; Wang, Y. miRNA mediated regulation and interaction between plants and pathogens. Int. J. Mol. Sci. 2021, 22, 2913. [Google Scholar] [CrossRef] [PubMed]
- Naz, M.; Benavides-Mendoza, A.; Tariq, M.; Zhou, J.; Wang, J.; Qi, S.; Dai, Z.; Du, D. CRISPR/Cas9 technology as an innovative approach to enhancing the phytoremediation: concepts and implications. J. Environ. Manage. 2022, 323, 116296. [Google Scholar] [CrossRef] [PubMed]
- Janik, E.; Niemcewicz, M.; Ceremuga, M.; Krzowski, L.; Saluk-Bijak, J.; Bijak, M. Various aspects of a gene editing system—crispr–cas9. Int. J. Mol. Sci. 2020, 21, 9604. [Google Scholar] [CrossRef] [PubMed]
- Dixit, S.; Sivalingam, P.N.; Baskaran, R.M.; Senthil-Kumar, M.; Ghosh, P.K. Plant responses to concurrent abiotic and biotic stress: unravelling physiological and morphological mechanisms. Plant Physiol. Rep. 2024, 29, 6–17. [Google Scholar] [CrossRef]
- Li, L.; Chen, J.; Sun, Z. Exploring the shared pathogenic strategies of independently evolved effectors across distinct plant viruses. Trends Microbiol. 2024. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Chen, X.; Gong, P.; Long, H.; Wang, J.; Yang, W.; Yao, W. Siraitia grosvenorii as a homologue of food and medicine: a review of biological activity, mechanisms of action, synthetic biology, and applications in future food. J. Agr. Food Chem. 2024, 72, 6850–6870. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Sundin, G.W.; Fuente, L.D.L.; Cubero, J.; Tatineni, S.; Brewer, M.T.; Zeng, Q.; Bock, C.H.; Cunniffe, N.J.; Wang, C. Key challenges in plant pathology in the next decade. Phytopathology 2024, 114, 837–842. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Wang, K.; Zhang, C. Carbon nanomaterials for plant priming through mechanostimulation: emphasizing the role of shape. ACS Nano 2024, 18, 10829–10839. [Google Scholar] [CrossRef] [PubMed]
- Yasuda, M.; Kusajima, M.; Nakajima, M.; Akutsu, K.; Kudo, T.; Yoshida, S.; Nakashita, H. Thiadiazole carboxylic acid moiety of Tiadinil, SV-03, induces systemic acquired resistance in tobacco without salicylic acid accumulation. J. Pestic. Sci. 2006, 31, 329–334. [Google Scholar]
- Patel, T.; Quesada-Ocampo, L.M.; Wehner, T.C.; Bhatta, B.P.; Correa, E.; Malla, S. Recent advances and challenges in management of Colletotrichum orbiculare, the causal agent of watermelon anthracnose. Horticulturae 2023, 9, 1132. [Google Scholar] [CrossRef]
- Kunz, W.; Schurter, R.; Maetzke, T. The chemistry of benzothiadiazole plant activators. Pestic. Sci. 1997, 50, 275–282. [Google Scholar] [CrossRef]
- Friedrich, L.; Lawton, K.; Ruess, W.; Masner, P.; Specker, N.; Rella, M.G.; Meier, B.; Dincher, S.; Staub, T.; Uknes, S. A benzothiadiazole derivative induces systemic acquired resistance in tobacco. Plant J. 1996, 10, 61–70. [Google Scholar] [CrossRef]
- Király, L.; Zechmann, B.; Albert, R.; Bacsó, R.; Schwarczinger, I.; Kolozsváriné Nagy, J.; Gullner, G.; Hafez, Y.M.; Künstler, A. Enhanced resistance to viruses in Nicotiana edwardsonii ‘Columbia’ is dependent on salicylic acid, correlates with high glutathione levels, and extends to plant-pathogenic bacteria and abiotic stress. Mol. Plant Microb. 2024, 37, 36–50. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.; Yoon, H.M.; Jung, J.; Yu, S.; Choi, S.Y.; Bae, H.W.; Cho, Y.H.; Chung, E.H.; Lee, Y. Pleiotropic effects of N-acylhomoserine lactone synthase ExpI on virulence, competition, and transmission in Pectobacterium carotovorum subsp. carotovorum Pcc21. Pest Manag. Sci. 2024, 80, 687–697. [Google Scholar] [CrossRef] [PubMed]
- Testi, S.; Kuhn, M.L.; Allasia, V.; Auroy, P.; Kong, F.; Peltier, G.; Pagnotta, S.; Cazareth, J.; Keller, H.; Panabières, F. The phytophthora parasitica effector AVH195 interacts with ATG8, attenuates host autophagy, and promotes biotrophic infection. BMC Biol. 2024, 22, 100. [Google Scholar] [CrossRef] [PubMed]
- Seth, T.; Asija, S.; Umar, S.; Gupta, R. The intricate role of lipids in orchestrating plant defense responses. Plant Sci. 2024, 338, 111904. [Google Scholar] [CrossRef]
- 299. .Delaney, T.P.; Uknes, S.; Vernooij, B.; Friedrich, L.; Weymann, K.; Negrotto, D.; Gaffney, T.; Gut-Rella, M.; Kessmann, H.; Ward, E.; Ryals, J. A central role of salicylic acid in plant disease resistance. Science 1994, 266, 1247–1250 https://doi101126/science26651881247. [Google Scholar]
- Peghaire, E.; Hamdache, S.; Galien, A.; Sleiman, M.; ter Halle, A.; El Alaoui, H.; Kocer, A.; Richard, C.; Goupil, P. Inducing plant defense reactions in tobacco plants with phenolic-rich extracts from red maple leaves: a characterization of main active ingredients. Forests 2020, 11, 705. [Google Scholar] [CrossRef]
- Yasuda, M.; Kusajima, M.; Nakajima, M.; Akutsu, K.; Kudo, T.; Yoshida, S.; Nakashita, H. Thiadiazole carboxylic acid moiety of tiadinil, SV-03, induces systemic acquired resistance in tobacco without salicylic acid accumulation. J. Pestic. Sci. 2006, 31, 329–334. [Google Scholar] [CrossRef]
- Niki, T.; Mitsuhara, I.; Seo, S.; Ohtsubo, N.; Ohashi, Y. Antagonistic effect of salicylic acid and jasmonic acid on the expression of pathogenesis-related (PR) protein genes in wounded mature tobacco leaves. Plant Cell Physiol. 1998, 39, 500–507. [Google Scholar] [CrossRef]
- Agrawal, G.; Jwa, N.S.; Han, K.-S.; Agrawal, V.; Rakwal, R. Isolation of a novel rice PR4 type gene whose mRNA expression is modulated by blast pathogen attack and signaling components. Plant Physiol. Bioch. 2003, 41, 81–90. [Google Scholar] [CrossRef]
- Uquillas, C.; Letelier, I.; Blanco, F.; Jordana, X.; Holuigue, L. NPR1-independent activation of immediate early salicylic acid-responsive genes in Arabidopsis thaliana. Mol. Plant Microb. 2004, 17, 34–42. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.T.; Bradford, K.J. Class I chitinase and β-1, 3-glucanase are differentially regulated by wounding, methyl jasmonate, ethylene, and gibberellin in tomato seeds and leaves. Plant Physio. 2003, 133, 263–273. [Google Scholar] [CrossRef]
- Mitter, N.; Kazan, K.; Way, H.M.; Broekaert, W.F.; Manners, J.M. Systemic induction of an Arabidopsis thaliana plant defensin gene promoter by tobacco mosaic virus and jasmonic acid in transgenic tobacco. Plant Sci. 1998, 136, 169–180. [Google Scholar] [CrossRef]
- Rai, A.K.; Sahoo, L.P.; Sahoo, M.; Seth, T.; Patra, C. Induced resistance mechanism in plant and its importance in agriculture. Int. J. Plant Soil Sci. 2024, 36, 1–22. [Google Scholar] [CrossRef]
- Li, H.; Liu, Y.; Wu, J.; Chang, K.; Zhang, G.; Zhao, H.; Qiu, N.; Bao, Y. Overexpression of GhGSTF9 enhances salt stress tolerance in transgenic Arabidopsis thaliana. Genes 2024, 15, 695. [Google Scholar] [CrossRef] [PubMed]
- Gaffney, T.; Friedrich, L.; Vernooij, B.; Negrotto, D.; Nye, G.; Uknes, S.; Ward, E.; Kessmann, H.; Ryals, J. Requirement of salicylic Acid for the induction of systemic acquired resistance. Science 1993, 261, 754–756. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, S.A.; Olson, D.G.; Argyros, D.A.; Miller, B.B.; Barrett, T.F.; Murphy, D.M.; McCool, J.D.; Warner, A.K.; Rajgarhia, V.B.; Lynd, L.R. Development of pyrF-based genetic system for targeted gene deletion in Clostridium thermocellum and creation of a pta mutant. Appl. Environ. Microbio. 2010, 76, 6591–6599. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.; Kwok, S.F.; Bleecker, A.B.; Meyerowitz, E.M. Arabidopsis thaliana ethylene-response gene ETR1: similarity of product to two-component regulators. Science 1993, 262, 539–544. [Google Scholar] [CrossRef] [PubMed]
- Angulo, M.; García, M.J.; Alcántara, E.; Pérez-Vicente, R.; Romera, F.J. Comparative study of several Fe deficiency responses in the Arabidopsis thaliana ethylene insensitive mutants ein2-1and ein2-5. Plants 2021, 10, 262. [Google Scholar] [CrossRef] [PubMed]
- Meesters, C.; Mönig, T.; Oeljeklaus, J.; Krahn, D.; Westfall, C.S.; Hause, B.; Jez, J.M.; Kaiser, M.; Kombrink, E. A chemical inhibitor of jasmonate signaling targets JAR1 in Arabidopsis thaliana. Nat. Chem. Bio. 2014, 10, 830–836. [Google Scholar] [CrossRef] [PubMed]
- Bi, Y.M.; Kenton, P.; Mur, L.; Darby, R.; Draper, J. Hydrogen peroxide does not function downstream of salicylic acid in the induction of PR protein expression. Plant J. 1995, 8, 235–245. [Google Scholar] [CrossRef] [PubMed]
- Tian, M.; von Dahl, C.C.; Liu, P.P.; Friso, G.; van Wijk, K.J.; Klessig, D.F. The combined use of photoaffinity labeling and surface plasmon resonance based technology identifies multiple salicylic acid binding proteins. Plant J. 2012, 72, 1027–1038. [Google Scholar] [CrossRef] [PubMed]
- Du, H.; Klessig, D.F. Identification of a soluble, high-affinity salicylic acid-binding protein in tobacco. Plant Physiol. 1997, 113, 1319–1327. [Google Scholar] [CrossRef]
- Me’traux, J.P.; Lamodie’re, E.; Catinot, J.; Lamotte, O.; Garcion, C. Salicylic acid and induced plant defenses. Rec. Adv. Pol-ypheno Res. 2009, 1, 202–210. [Google Scholar]
- Wu, Q.; Wang, X.Z.; Tang, Y.Y.; Yu, H.T.; Ding, Y.F.; De Yang, C.; Cui, F.G.; Zhang, J.C.; Wang, C.T. Molecular cloning and characterization of NPR1 gene from Arachis hypogaea. Mol. Biol. Rep. 2014, 41, 5247–5256. [Google Scholar] [CrossRef]
- Kumar, D. Salicylic acid signaling in disease resistance. Plant Sci. 2014, 228, 127–134. [Google Scholar] [CrossRef] [PubMed]
- Durner, J.; Klessig, D.F. Inhibition of ascorbate peroxidase by salicylic acid and 2, 6-dichloroisonicotinic acid, two inducers of plant defense responses. Proc. Natl. Acad. Sci. U.S.A. 1995, 92, 11312–11316. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.; Tian, M.; Zhang, H.; Li, X.; Wang, Y.; Xia, X.; Zhou, J.; Zhou, Y.; Yu, J.; Shi, K. Salicylic acid binding of mitochon-drial alpha-ketoglutarate dehydrogenase E2 affects mitochondrial oxidative phosphorylation and electron transport chain components and plays a role in basal defense against tobacco mosaic virus in tomato. New Phytol. 2015, 205, 1296–1307. [Google Scholar] [CrossRef] [PubMed]
- Manohar, M.; Tian, M.; Moreau, M.; Park, S.W.; Choi, H.W.; Fei, Z.; Friso, G.; Asif, M.; Manosalva, P.; von Dahl, C.C. Iden-tification of multiple salicylic acid-binding proteins using two high throughput screens. Front. Plant Sci. 2015, 5, 777. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
The genetic basis of plant defense responses.
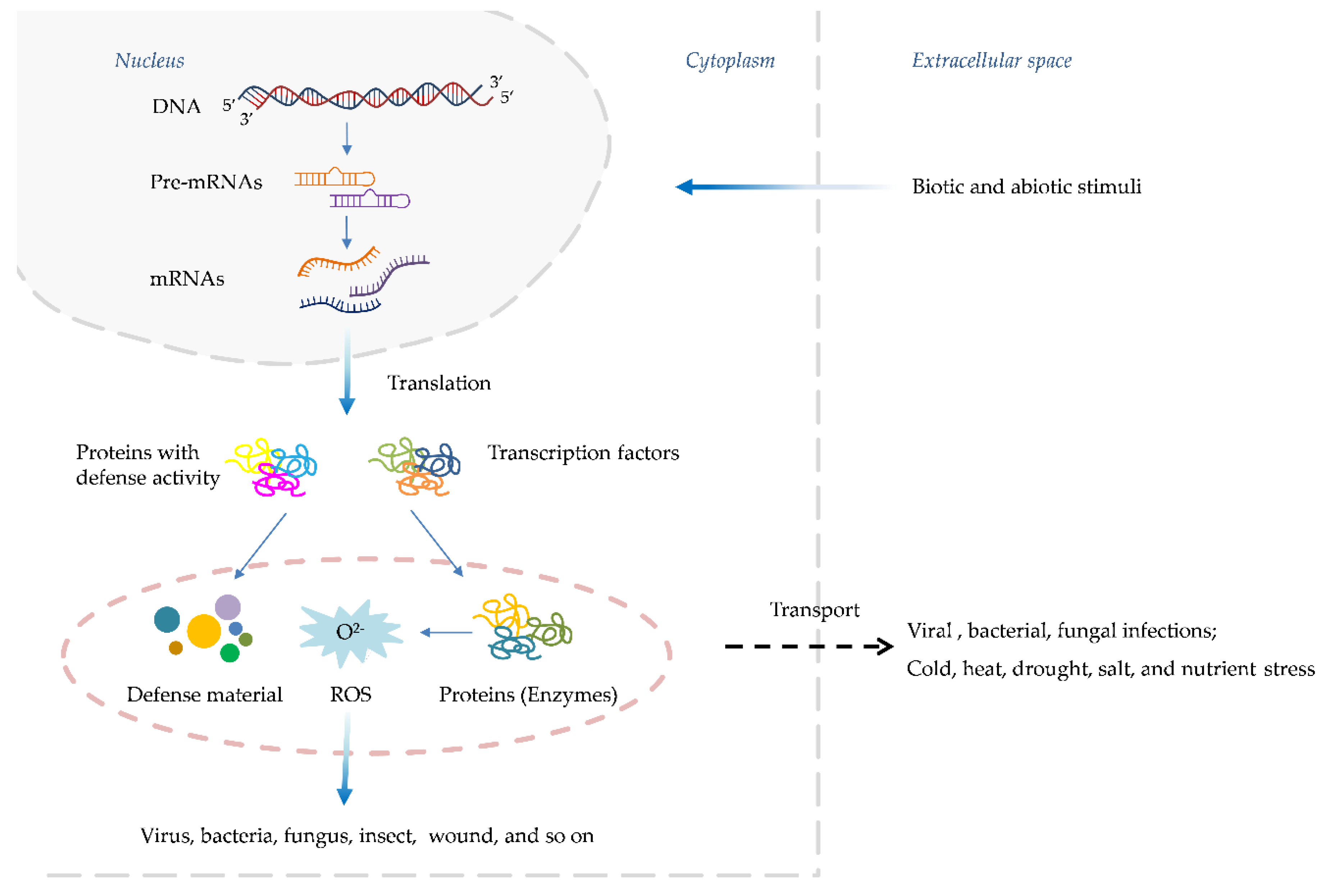
Figure 2.
SA signal pathway and some related regulatory elements in the pathway.
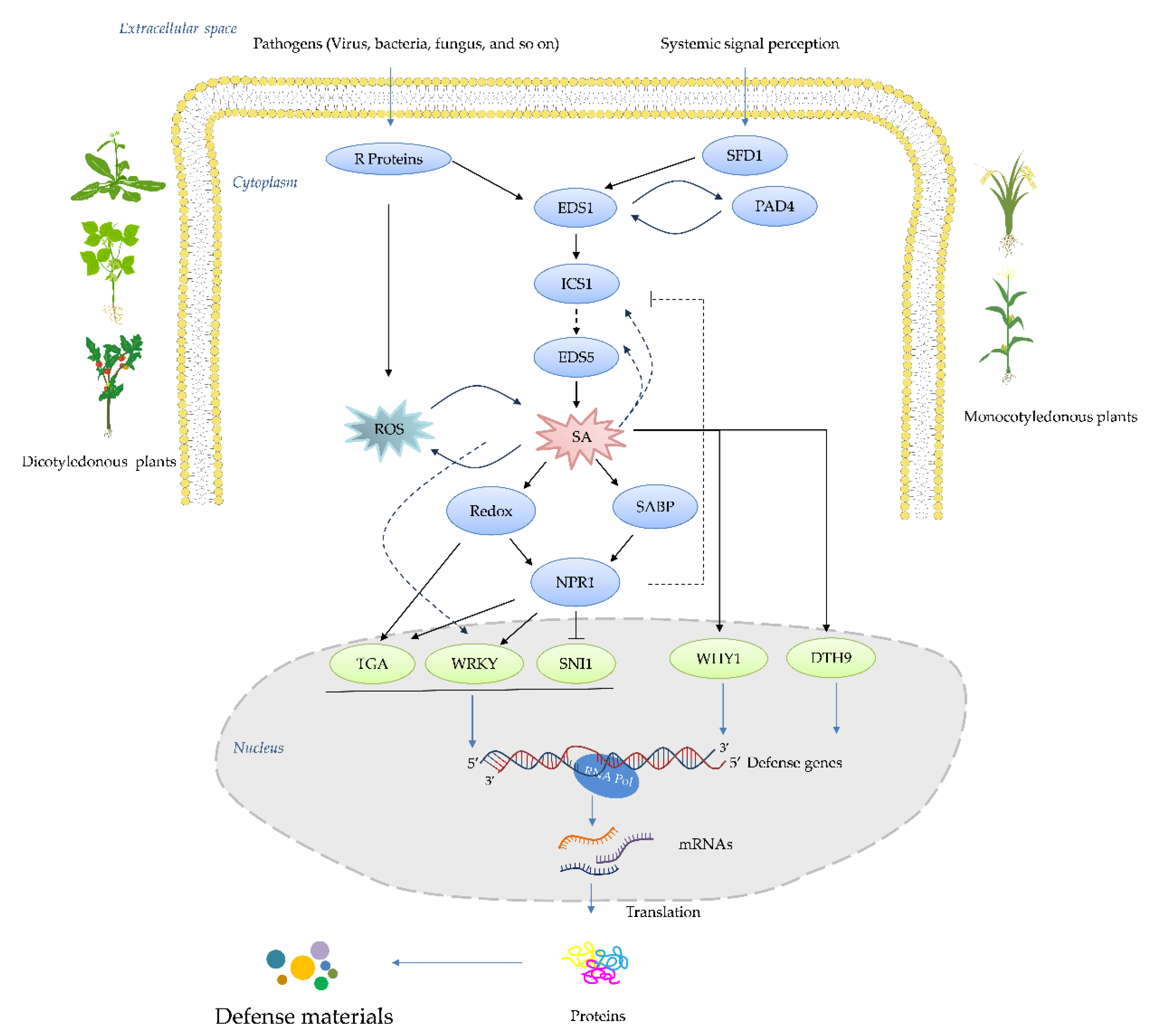
Figure 3.
SA and some molecules act in SA upstream in SA signal pathway. (a) SA. (b) VMA. (c) PBZ. (d) BIT.
Figure 3.
SA and some molecules act in SA upstream in SA signal pathway. (a) SA. (b) VMA. (c) PBZ. (d) BIT.
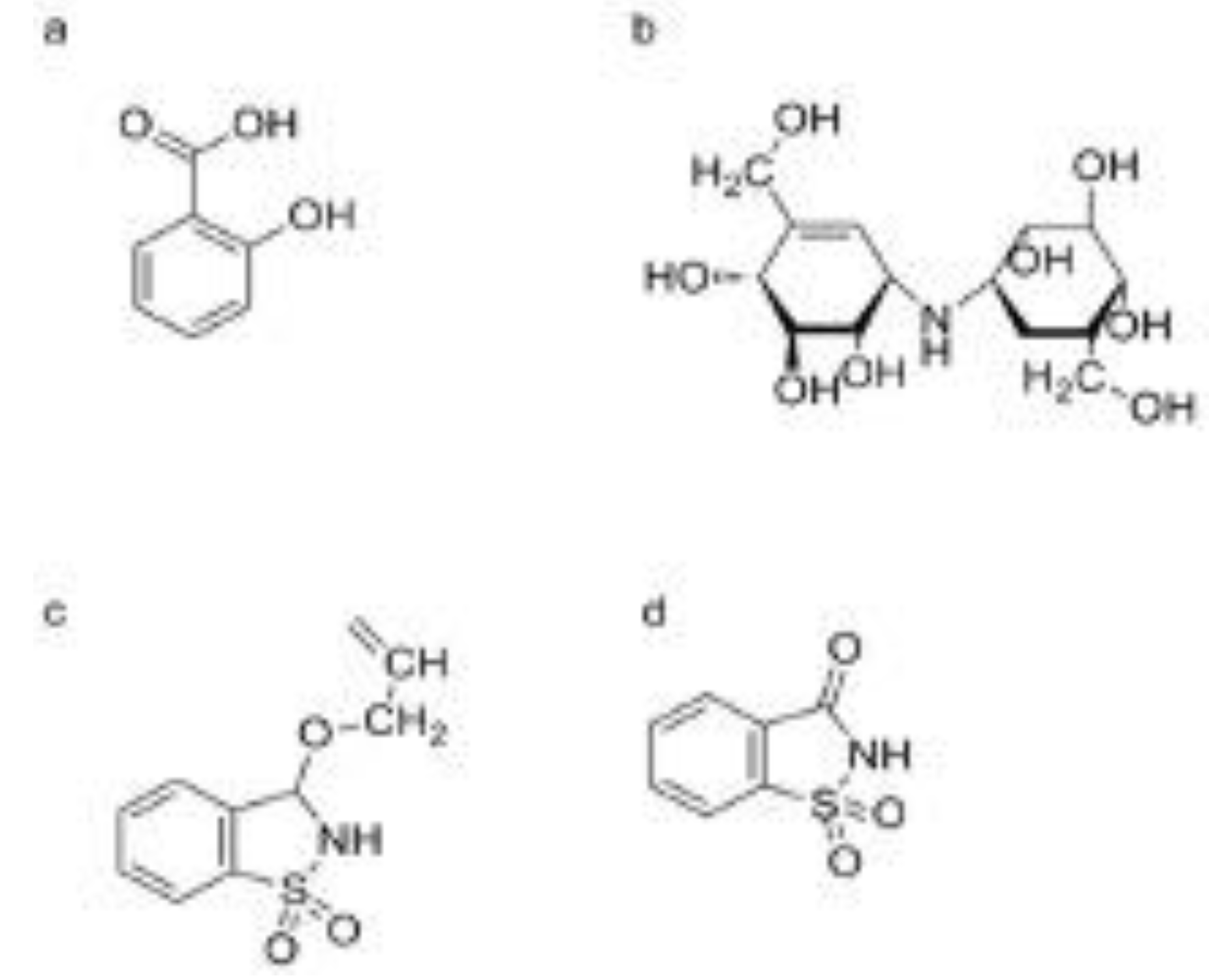
Figure 4.
The molecule structure of Imprimatins and diuretics compounds. (a) Imprimatins A1. (b) Imprimatins A2. (c) Imprimatins A3. (d) Imprimatins B1. (e) Imprimatins B2. (f) Bumetanide. (g) Bendroflumethiazide. (h) Clopamide.
Figure 4.
The molecule structure of Imprimatins and diuretics compounds. (a) Imprimatins A1. (b) Imprimatins A2. (c) Imprimatins A3. (d) Imprimatins B1. (e) Imprimatins B2. (f) Bumetanide. (g) Bendroflumethiazide. (h) Clopamide.
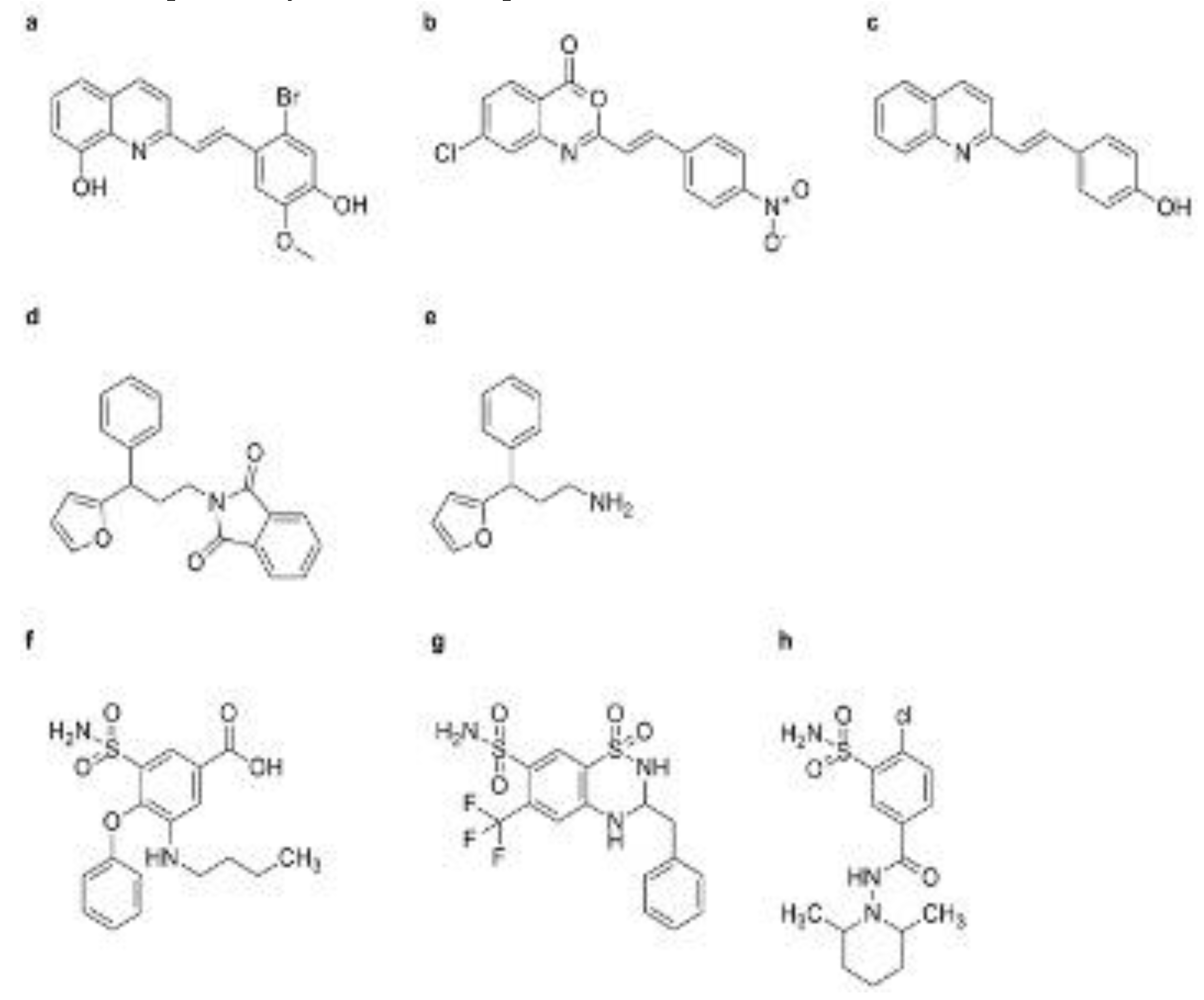
Figure 5.
Some molecules act in SA downstream in SA signal pathway.
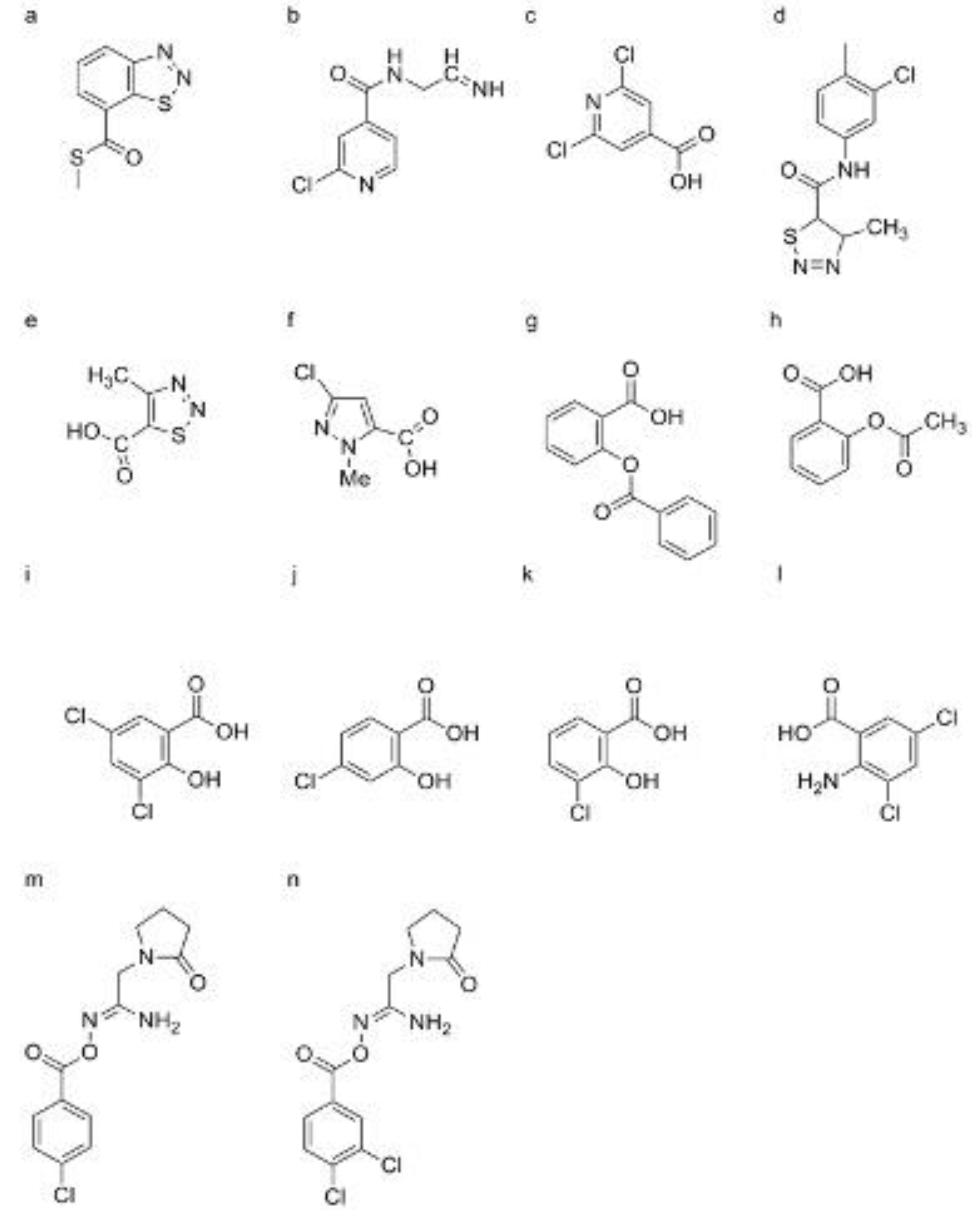
Figure 6.
The action mechanism of BTH.
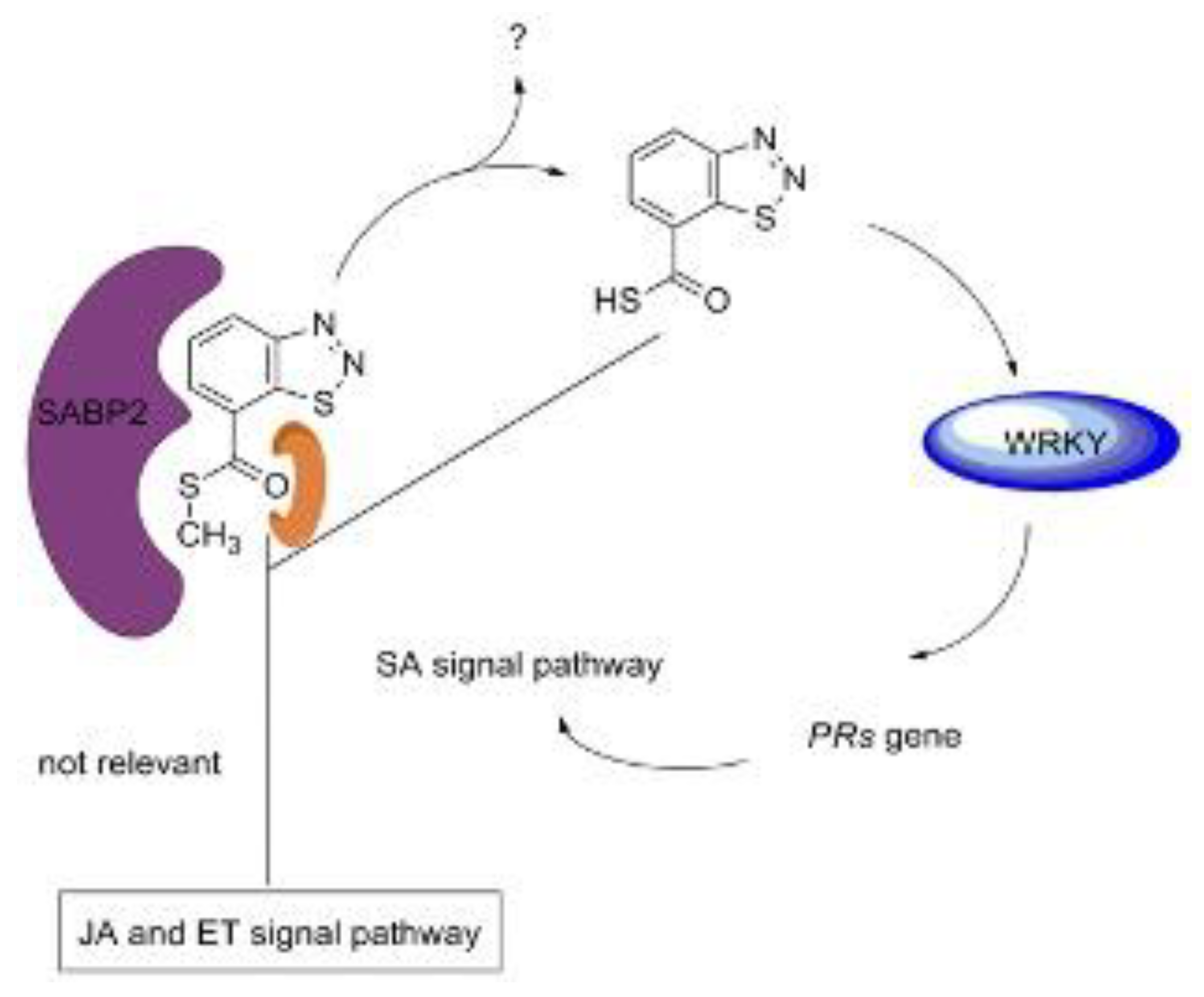
Figure 7.
The action sites of some plant activators against SA signal pathway.
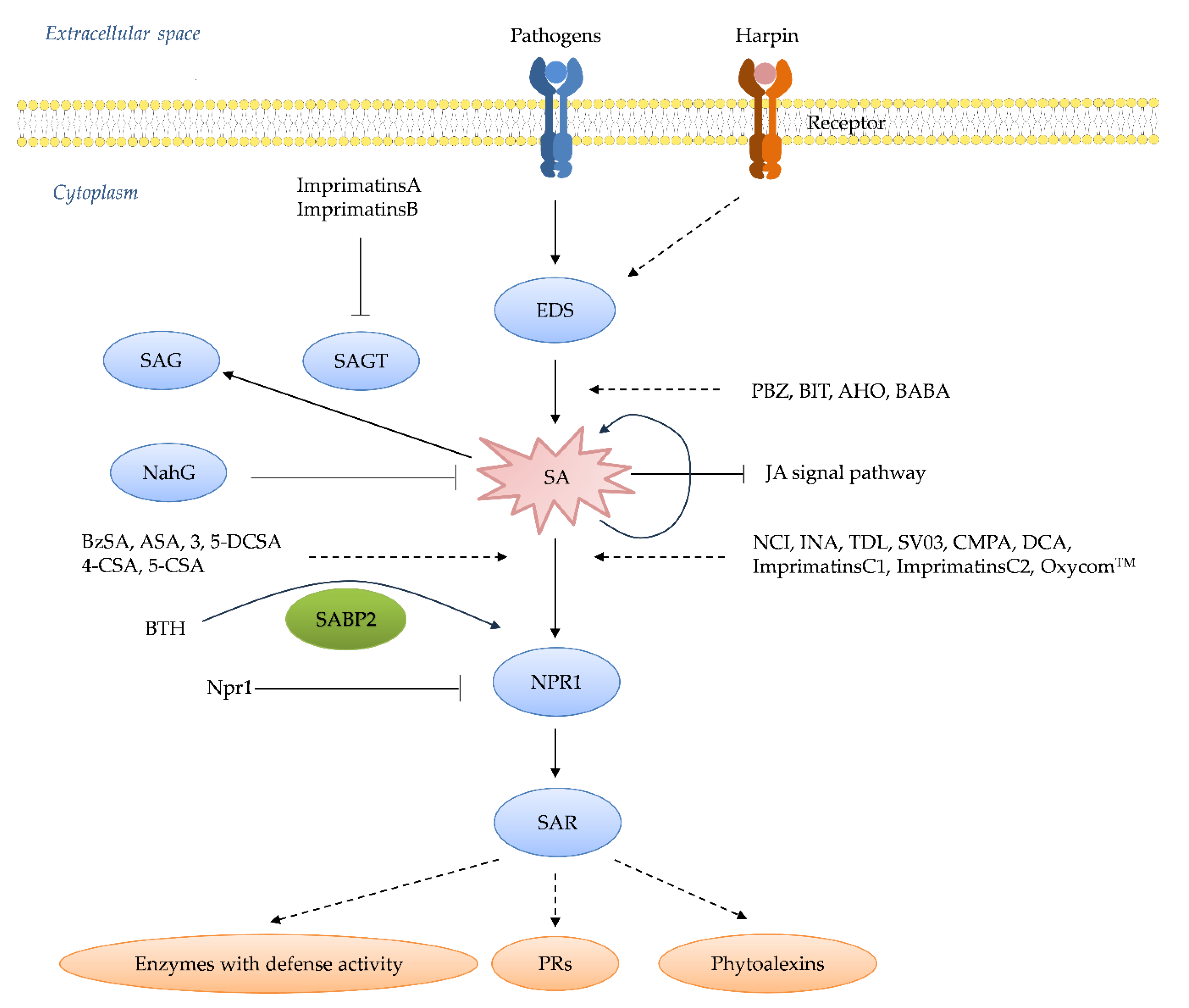
Figure 8.
The molecule structrue of Isotianil (a) and DDCC (b).
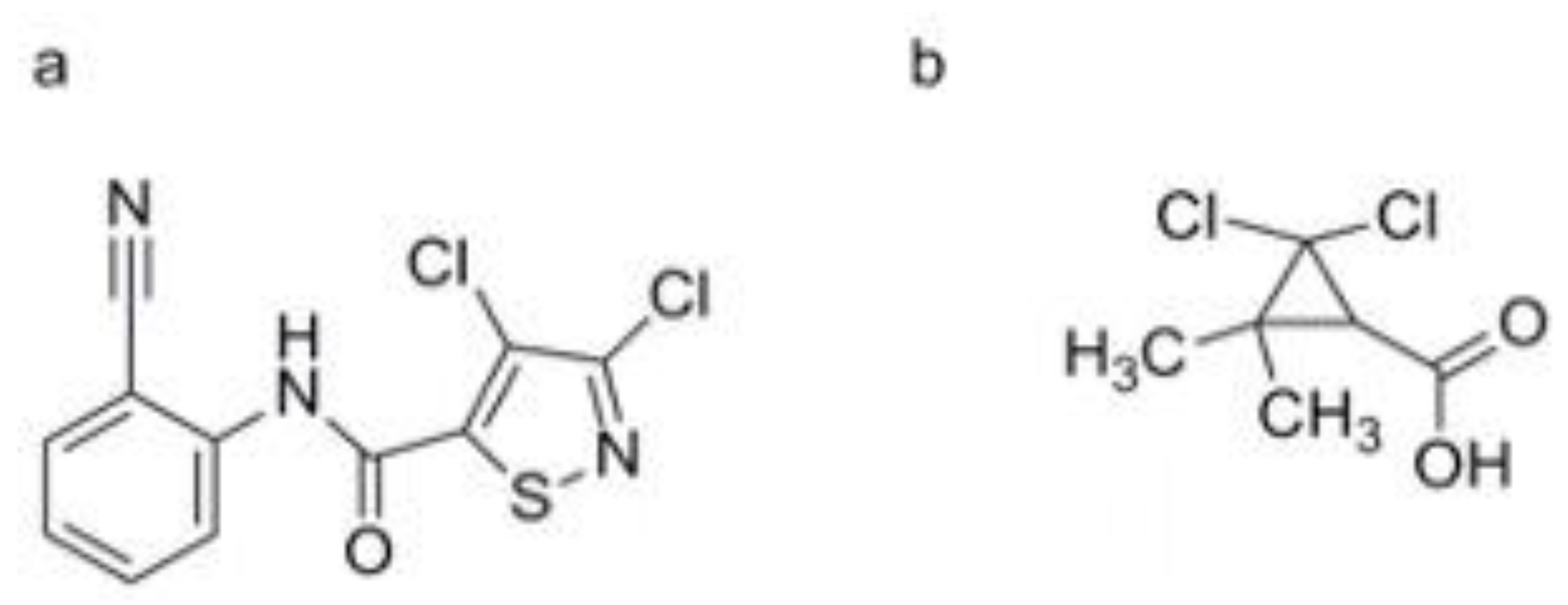
Figure 9.
The plant activators targetting plant SA signal pathway by developed in China.
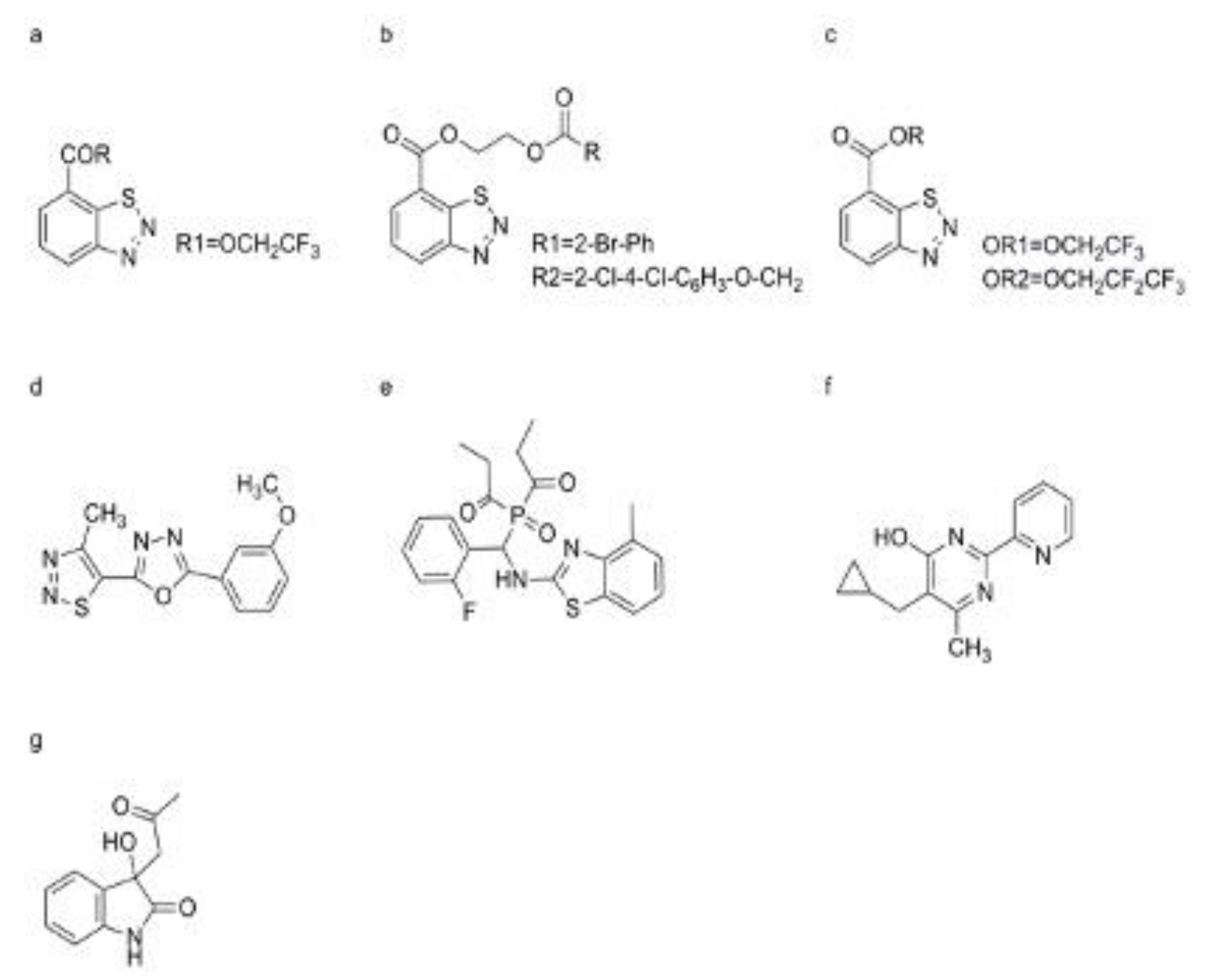

Table 1.
The screening model of the living plant against many phytopathogens.
| Plants | Plant species | Pathogen names | Evaluation index | References |
|---|---|---|---|---|
| Cucumber | No mention | Colletotrichum lagenarium (Paserini) Ell. & Halst | Infected leaf area | [292,293] |
| Tobacco | N. tabacum cv. Xanthi nc | TMV | Lesion size | [294] |
| N. tabacum cv. Xanthi nc | Oidium lycopersici | Lesion size | [119] | |
| N. tabacum cv. Xanthi nc | Pst | CFU or disease severity | [290] | |
| N. tabacum cv. Xanthi nc | Cercospora nicotianae | Infected leaf area | [290,295] | |
| N. tabacum cv. Xanthi nc | Peronospora tabacina | Infected leaf area | [290] | |
| N. tabacum cv. Xanthi nc | Erwinia carotovora | Length of infected stem | [290,296] | |
| N. tabacum cv. Xanthi nc | Phytophthora parasitica | Disease severity | [297] | |
| A. thaliana | Peronospora parasitica pv. Emwa | Disease severity | [171,299] | |
| Rice | Oryza sativa cv. Aichiasahi | M. grisea | Lesion number | [178,298] |
| O. sativa cv. Aichiasahi | X. oryzae pv. oryzae | Lesion length | [202] |
Table 2.
The markers in SA or JA signal pathway and the related methods.
| Plant | Signal pathways | Effector markers | Methods | References | |
|---|---|---|---|---|---|
| Tobacco | SA | SA (free SA) | Organic solvent extract coupled with HPLC | [178,301] | |
| SA (total SA) | Methanol extract coupled with the high-performance liquid chromatographic | ||||
| SA (SAG) | Methanol extract coupled with the high-performance liquid chromatographic | [18] | |||
| Tobacco | SA | PR-1 | Northern blotting | [302] | |
| PR-1 | Western blotting | [172] | |||
| SA | PR-2 | Northern blotting | [302] | ||
| SA | PR-3 | Northern blotting | [303] | ||
| SA | PR-4 | Northern blotting | [304] | ||
| SA | PR-5 | Northern blotting | [155] | ||
| Arabidopsis | SA | PR-1 | Northern blotting, | [170] | |
| SA | PR-2 | Northern blotting, | [170,305] | ||
| SA | PR-5 | Northern blotting, | [170][122] | ||
| Tomato | SA | β-1, 3-glucanase | Western blotting | [306] | |
| Tobacco | JA | PDF1.2 | Northern blotting, | [307] | |
Table 3.
The transgenics plant or mutant gene and their corresponding the detection methods.
| Transgenics or mutants | Plants | Action sites | Signal pathways | References |
|---|---|---|---|---|
| NahG transgenics | Tobacco | SA level | SA | [9,170,294,308,310] |
| SABP2-silenced transgenic | Tobacco | SA level | SA | [111] |
| nim mutant | A. thaliana | Downstream of SA | SA | [299] |
| npr1 mutant | A. thaliana | Downstream of SA | SA | [20] |
| etr1 mutant | A. thaliana | ET | [312] | |
| ein2 mutant | A. thaliana | ET | [313] | |
| jar1 mutant | A. thaliana | JA | [314] |
Table 4.
Proteins interacting with SA in plant systems.
| Proteins interacted with SA | Sources | Ligands | References |
|---|---|---|---|
| Catalase | N. tabacum | 14C-SA | [315] |
| Methyl esterase | N. tabacum | 3H-SA | [316,317] |
| Carbonic anhydrase | N. tabacum | 3H-SA | [318] |
| A protein with ankyrin repeat and BTB/POZ domain | A. thaliana | Genetics/T-DNA mutant | [319] |
| CUL3 adapter protein | A. thaliana | Genetics/NPR1 paralog | [320] |
| CUL3 adapter protein | A. thaliana | Genetics/NPR1 paralog | [320] |
| Ascorbate peroxidase | N. tabacum | 3H-SA | [321] |
| E2 subunit of α-ketoglutarate dehydrogenase 2 | A. thaliana | 4-azido SA, SPR | [322] |
| Glutathione S-transferase 2 | A. thaliana | 4-azido SA, SPR | [1] |
| Glutathione S-transferase 8 | A. thaliana | 4-azido SA, SPR | [321] |
| Glutathione S-transferase 10 | A. thaliana | 4-azido SA, SPR | [322] |
| Glutathione S-transferase 11 | A. thaliana | 4-azido SA, SPR | [93] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



