
Submitted:
16 July 2024
Posted:
18 July 2024
You are already at the latest version
Abstract
Increasing population size and income growth are causing and increasing demand for food and protein of animal origin. Insects are an interesting alternative to conventional animal source proteins as they can convert by-products from the agricultural industry to biomass for commercial feed for livestock and potentially as a food source for human consumption. Microorganisms have been found to affect insects and can be accumulated via horizontal and vertical transmission. This study aimed to identify if the removal of egg- and substrate-associated microorganisms impact larval performance through the following parameters: development of biomass, final biomass, and the survival rate of house fly (Musca domestica) larvae. Four treatments were tested on substrate consisting of 10.81% alfalfa (Medicago sativa), 21.62% wheat bran (Triticum aestivum), and 67.57% water: (A) disinfected eggs and non-autoclaved substrate, (B) non-disinfected eggs and autoclaved substrate, (C) disinfected eggs and autoclaved substrate, and (D) a control without any removal of microbiota. The results showed a significant decrease in final biomass for the treatments with only disinfected eggs, only autoclaved substrate and both when comparing to the control, and a significant decrease in survival rate for B and D. Moreover, the development of biomass showed a significant difference between days within all treatments. Together this suggest that microorganisms of housefly eggs and the growth substrate plays an important role for growth, which is critical in commercial insect production. Further studies must be performed to examine these parameters in more commercially relevant substrates.
Keywords:
Musca domestica
; insect production
; larval performance
; microorganisms
; egg disinfection
; substrate autoclaving
1. Introduction
Rapid global population growth exerts pressure on agricultural production for food and feed [1]. It is estimated that by 2050, food production needs to increase by 70% compared to 2009 and the demand for proteins of animal origin will have increased by 74% [2]. The agricultural industry is currently using 37% of Earth’s land mass for food production. This causes deforestation and as a result it inflicts a loss of biodiversity and increased greenhouse gas emissions [3]. Alone livestock production is responsible for 14% (excluding land-use) of the world’s greenhouse gas emissions. These emissions originate from manure management, ruminant waste on pastures, ruminant emissions, and fertiliser production [3]. In addition, results suggest that the agricultural industry is responsible for 92% of the global water consumption [3].
Moreover, one third of food is lost from primary production to consumption, which has both economic and environmental consequences [4]. In addition, food loss and waste account for 8% of the global greenhouse gas emissions [3]. The loss occurs at several stages of the food supply chain. Already during the production of grain, fish, and fruit, food is discarded. Damaged, spilled, and uneaten food will also be lost during processing, packaging, and consumption [3]. These problems necessitate proper handling of waste and by-products while providing with a sustainable high quality protein source for food and feed.
Insects are an interesting alternative solution for the above-mentioned agricultural related challenges, as it enables the conversion of waste- and by-products from the agricultural industry into biomass that can potentially be used for commercial feed for livestock animals, as a potential food source for humans [5,6], and as fertiliser producers [7].
Currently most of the insects being produced are used as pet feed, fish feed, and aquaculture companies. However, there is a growing interest worldwide for insects as food and feed. The most prominent insects for insect production in Europe are the black soldier fly (Hermetia illucens), yellow mealworm (Tenebrio molitor), lesser mealworm (Alphitobius diaperinus) and the house fly (Musca domestica) [8]. However, current EU legislation imposes challenges for the implementation of insects as food and feed and currently only a few insect species are allowed for human consumption and as feed for livestock [9]. Moreover, only certain feeds are currently allowed as growth substrate in insect production, with current bans on slurry or manure and unprocessed foodstuffs containing meat or fish [10]. These principles illustrate the demand for a greater focus on the commercialisation of insect production.
Insects are an effective commercial food- and feed product because of the high feed conversion rate in comparison to livestock [11,12]. In addition, insects can be valuable in the valorisation of waste- and by-products, such as used grains, household waste, and manure [9]. Specific substrates appear to be more efficient than others. For example, H. illucens larvae fed on grape pomace and spinach showed higher mortality, while a diet of pumpkin and red cabbage showed high bio-conversion rates. The differences are likely due to different nutritional values and digestibility of the substrates [13]. Manure substrates have been found to be effective feed for M. domestica and H. illucens, and other insects have been postulated to be effective in utilising manure as feed [6].
Microorganisms play critical role in the fitness of insects [14]. Some bacterial species exists as either endosymbionts or as ectosymbionts and can be mutualistic, commensalistic or pathogenic [15]. Microorganisms can be acquired in various ways, such as vertically from mother to offspring [16], or horizontally transmitted through substrates [17,18]. Furthermore, free-living bacteria can be acquired throughout the life cycle, and can either become obligate or facultative endosymbionts. These, especially obligate endosymbionts, are entirely dependent on the transmission between generations [15]. Vertically transmitted microbes have shown to be conducive to increased fitness through the influence on factors like reproductive ability or metabolic performance [14,19]. They are also involved in inhibition of pathogens and biofouling, and in oxygen- and nutrient acquisition [14]. Therefore, microorganisms appear to be crucial for many aspects of the insect’s overall fitness during their life cycle.
Bacteria are predominantly present in the digestive system, where they play a role in key aspects of their host’s metabolism—primarily aiding in the acquisition of nutrients such as nitrogen, vitamins, and sterols [15,20]. For example, in honeybees (Apis mellifera), the bacteria Gilliamella apicola helps the breakdown of pollen. Additionally, the bacteria Ishikawaella capsulata aids in growth and reproduction in the Japanese common plataspid stinkbug (Megacopta punctatissima) and is also found to contain pathways for many essential and non-essential amino acids [21]. Larvae of H. illucens have also indicated dependence of microorganisms in the environment, since substrate-associated microorganisms were able to influence the survival, weight, and proportion of pre-pupae in H. illucens [22]. Furthermore, microorganisms in insects have found to improve resistance to environmental disturbance, enhance longevity, and shorten larval development time [23]. Moreover, specific microorganisms play specific roles for hosts. The bacteria Candidatus ishikawealla is important for the bean platasid’s (Megacopta cribraria) development to the adult stage. For aphids, it has been shown that the bacteria Buchnera aphidicola assists in reaching nutritional demands, and that the survival and fecundity of aphids are affected negatively without the presence of this specific bacteria [15]. For the house fly (Musca domestica) it has been determined that different bacteria are present at different life stages. For example, the bacteria strain Weissella dominated colonisation of newly hatched larvae, and at the second larval stage the dominant strains were Stenotrophomonas, Bacillus, and Lactococcus. This suggests that the bacteria have different impacts at different life stages [24]. A specific example of this impact is the bacteria Klebsilla oxytoca’s ability to restrict further oviposition at sites with recently laid eggs for the female flies [24].
Besides in larval intestines, bacteria are also present on the surface of eggs. Mazza et al. (2020) isolated 9 different bacteria present on eggs of H. illucens. Specific combinations of the bacteria present could either negatively or positively impact the weight gain and conversion rates in their larval stage [25]. It is therefore not surprising that several papers have suggested bacteria to play a key role in commercial insect production [9,26].
Eggs are especially sensitive to pathogens and the host can, via microorganisms, inhibit pathogens [14]. Microorganisms within the water flea (Daphnia magna) eggs and in the surrounding environment have shown to result in positive effects on the reproductive ability and development of D. magna. A sterile environment for the eggs has shown to decrease these parameters [14].
M. domestica is an organism which can be used to determine the effects of vertically transmitted and substrate-associated microorganisms, and in addition, the larvae are a promising applicant as an alternative food- and feed-source as it feeds on waste- and by-products, which in turn, creates usable biomass for commercial production [27]. This study aims to quantify the influence of the egg- and substrate-associated microorganisms on M. domestica larvae’s performance. This will be investigated through autoclaved substrate (a combination of wheat bran (Triticum aestivum), alfalfa (Medicago sativa) and water) and surface-disinfected eggs using sodium hypochlorite solution. Tests were performed to examine the effects on the performance of M. domestica larvae through the following parameters: development of biomass, final biomass, and survival rate.
2. Materials and Methods
2.1. Study Organism and Rearing
The flies used in the present study were obtained from a laboratory culture at Aalborg University, established from flies collected from Danish farms in 2019 which were kept in the laboratory. The culture has been kept at 23◦C ± 1 ◦C with 12 hours of light and 12 hours without, for one generation. The flies (F1 generation) were reared in a substrate consisting of 65.75% water, 21.92% wheat bran (Triticum aestivum), 10.96% alfalfa (Medicago sativa), 0.82 % maltose, and 0.55% dry yeast with a density of 0.67 eggs per gram substrate. When pupae were observed, they were placed in a bug dorm. A diet of ad libitum water, and a mix of sugar, powdered sugar, and milk powder were available, when the adult stage was reached.
2.2. Experimental Design
To test how substrate-level microorganisms and the maternally inherited egg surface-level microbes affect development of biomass, final biomass, and survival rate, two experiments were carried out.
Experiment 1: Sodium hypochlorite concentration effects on removal of microbiota from egg-surface
Experiment 1 was conducted to determine which concentration of sodium hypochlorite was the most optimal in ensuring the survival of eggs until the larval stage while also reducing microbiota. Six treatments of various sodium hypochlorite concentrations were used (see Table 1 for concentrations). In all treatments, 50 eggs were collected from the F1 generation, and were disinfected (see Section 2.2.1 for disinfection method). One egg was placed aerobically on a plate of LB agar to examine effectiveness of the disinfection (see Section 2.2.2).
Experiment 2: The effect of disinfection of eggs and substrate on development of biomass, final biomass, and survival rate
Based on experiment 1 it was found that a concentration of 0.006% w/w sodium hypochlorite had the highest rate of survival while also reducing the microbiota on the surface of the eggs. In experiment 2 four different treatments were prepared; (A) one with disinfected eggs and non-autoclaved substrate, (B) one with non-disinfected eggs and autoclaved substrate, (C) one with disinfected eggs and autoclaved substrate, and (D) a control with no disinfection and no autoclaving of eggs and substrate respectively. Thus, 80 containers, each container representing a sample, were prepared; 40 with 50 grams of autoclaved substrate and 40 with 50 grams of non-autoclaved substrate. 8000 eggs were collected; 4000 were disinfected with 0.006% w/w sodium hypochlorite and 4000 were not. Each treatment consisted of 20 replicates of each 100 eggs with 50 grams of substrate. The containers were covered with disinfected and non-disinfected fabric depending on treatment and placed under the same conditions as F1. Every day, the containers were stirred. Four LB agar plates were produced during the disinfection process for the purpose of examining presence of microbiota: one with 25 disinfected eggs, one with 25 non-disinfected eggs, one with autoclaved substrate, and one with non-autoclaved substrate, see Appendix B, Figure A2.
2.2.1. Disinfection and Autoclaving of Eggs and Substrate
Both in experiment 1 and 2, the egg-surface was disinfected with chlorine bleach. The active substance in chlorine bleach is 1-5 % w/w sodium hypochlorite. The disinfection process was carried out by placing the eggs in a sieve, and surface-disinfected by washing the eggs with chlorine bleach, and letting it sit for 4 minutes. Thereafter, the chlorine bleach was rinsed off with ultra-pure water, and the eggs were transferred to a container by pouring ultra-pure water over them, causing them to fall into the container. A disinfected fabric was used to cover the containers of the disinfected eggs and substrate. The process took place under disinfected conditions in a laminar flow cabinet. In the containers there were 50 grams of autoclaved substrate (consisting of 10.81% alfalfa, 21.62% wheat bran, and 67.57% water). The substrate was disinfected in a 1000 millilitre glass container at 120◦C in an autoclave.
To determine the effect of disinfection on hatch rate larvae were counted after six days.
2.2.2. Determination of Bacterial Growth
To determine the bacterial growth, LB agar plates were prepared. In a 250 mL glass container 5 g/L agar (VWR), 2.5 g/L NaCl (VWR), 2.5 g/L tryptone (Flucka), 1 g/L yeast extract (Thermo Scientific), and 250 mL of water were added. The pH was adjusted to 7.0 with NaOH. The LB agar medium was then autoclaved and poured into Petri dishes. Eggs and substrate were placed and left on the agar plates under sterile conditions in a laminar flow cabinet. The plates were incubated at 35 °C and examined after five days. Number of colonies were counted by visual inspection or with the use of a grid to count colonies methodically. These were compared with hatching rates to evaluate the effectiveness of each solution. Hatching rates were calculated with the following formula:
The number of colonies and the corresponding hatching rates are shown in Appendix A, Table A1 for experiment 1. The number of colonies for experiment 2 is shown in Appendix B, Table A2.
2.2.3. Determination of Development of Biomass
In experiment 2, when five larvae or more were observed in a given replicate, larvae were weighed daily. This continued until the day, when a decrease in the mean larvae biomass of a treatment was observed.
2.2.4. Determination of Final Biomass And Survival Rate
After a decrease in mean larval biomass, the surviving larvae and pupae were frozen and afterwards counted to determine the rate of survival. The larvae were weighed individually to determine final biomass. For samples where less than five larvae were present, the larvae were only used for determination of the final biomass and rate of survival. Survival rate was calculated with the following formula:
2.3. Statistical Analysis
All figures and analyses were conducted in R version 4.4.0 using RStudio version 2024.04.1+748 (http://www.rstudio.com) using the following packages: ggplot2 [28], tidyverse [29], dplyr [30], dunn.test [31], and rstatix [32]. Boxplots were made to visualise the development of biomass, final biomass, and survival rate. A Shapiro-Wilk test confirmed that the data followed a non-normal distribution. Therefore, non-parametric tests were used. A Kruskal-Wallis rank sum test was carried out to determine if there was a significant difference of treatments followed by a Dunn’s test to determine the significant differences between the treatments with a Bonferroni correction. Furthermore, a repeated measures ANOVA were used to assess the effect of time on the development of biomass.
3. Results
3.1. Development of Biomass
Disinfected eggs and/or autoclaved substrate resulted in a slower development of biomass (Figure 1). Furthermore, for A (disinfected eggs) and D (control) the median biomass increased from day to day for the first five days. For B (autoclaved substrate) the median biomass increased the first four days, and from day five to day seven. For C (disinfected eggs and autoclaved substrate) the median biomass increased the first two days, and from day four to day six.
There was a significant effect of time on development of biomass for all treatments (Table 2). The p-values of the repeated measures ANOVA show significant difference between means for the days in all treatments. Furthermore, for A and D a substantial F-value indicates a large variance of means between groups in comparison to the variance of means within groups. The smaller F-value for B and C indicates less variance between groups when comparing to variance of means within groups. Moreover, the generalised eta squared value (GES) indicates 76% and 74% of the variance for A and D, respectively, is explained by time. Whereas 36% and 7% of the variance for B and C, respectively, is explained by time.
3.2. Final Biomass
Disinfected eggs and/or autoclaved substrate resulted in a lower final biomass for larvae compared to the control (Figure 2). There was a significant effect of treatment on the final biomass of the larvae (Kruskal-Wallis: p-value of < 2.2 · ).
A significant difference in means for the final biomass of the larvae was found between all treatments (Table 3).
3.3. Survival Rate
Treatment C had a lower median survival rate than the other treatments, B had a lower median survival rate than both A and D, and treatment A had the highest median survival rate (Figure 3). At least one treatment had a significant impact on the survival rate of the larvae (Kruskal-Wallis: p-value of < 2.2 · ).
A significant difference in means for the survival rate of the larvae was found between A and B, A and C, B and D, and C and D. There was no significant difference between B-C and A-D (Table 4).
4. Discussion
Microorganisms can affect insect fitness in various ways [20]. The presence of microorganisms on eggs and in the environment can impact different factors like development, biomass, and survival rate [15,22,23,33]. However, few studies exist on the impact of microbes on the production of M. domestica for food- and feed production. It was expected that disinfection of eggs, autoclaving of substrate, or both would lead to a decreased development of biomass, reduced final biomass, and impaired survival rate for larvae of M. domestica compared to non-disinfected and -autoclaved conditions (control).
For all treatments a significant difference in development of biomass between days were found within each treatment. For disinfected eggs and/or autoclaved substrate a decreased final biomass for the larvae was found. Furthermore, the two treatments with only autoclaved substrate, and both disinfected eggs and autoclaved substrate led to a decreased survival rate.
4.1. The Effect of Disinfected Eggs
It was expected that disinfected eggs would lead to decreased development of biomass, reduced biomass, and impaired survival. This current study found a significant difference between disinfected- and non-disinfected eggs between days in the development of biomass and a significant difference in final biomass, but no significant difference in survival rate.
Schreven et al. (2021) examined the implications a sterile egg surface had on the performance, which includes final biomass and survival rate, of H. illucens. Contrary to the current study, they found no significant effect of sterile eggs on larval survival and biomass. They argue that the microbiota present on eggs were so few compared to the substrate-associated microorganisms, that they had no effect on the overall microbiota composition nor the larval performance [22]. However, Mazza et al. (2019) examined the specific egg-associated bacteria present on the eggs of H. illucens. They found that the bacteria were able to enhance biomass and influence accumulation of crude fat and crude protein depending on the specific bacteria present on the eggs [25]. Similarly, Lam et al. (2009) found bacteria present on eggs of M. domestica contribute as a nutrient reserve [33], which may explain the reduced final biomass observed in this study.
Development of biomass for disinfected eggs did not seem to be influenced in this current study. While significance between days were observed within treatments, the effect size for disinfected eggs and control were similar (GES: 0.768 and 0.739, respectively), indicating the major factor for variance is time for both treatments.
This is not in accordance with a different study by Lam et al. (2009) which uncovered that the presence of bacterial colonies on eggs were associated with suppression of fungal growth in their substrate and showed the removal of said bacteria increased fungal growth which inadvertently affected M. domestica adult fly emergence negatively [34]. While this current study did not investigate emergence, the presence of fungicidal bacteria could be affecting the development of larvae, however, these findings are not sustained in this study, but instead might still be influencing the observed difference in final biomass. As mentioned, this study found no significant difference for the survival rate between the disinfected- and the non-disinfected eggs. However, this is not in accordance with the study by Lam et al. (2009), who observed decreased survival for the sterilised egg treatment [34]. This could be explained by the fact that insects contain egg-associated microbiota which assist with the suppression of fungi in the substrate and protecting against pathogens [14,34]. Therefore, it could be speculated that the removal of microbiota from the egg-surface would reduce survival rate. This study does not find a correlation between survival and egg-associated microbiota.
4.2. The Effect of Autoclaved Substrate
It was expected that autoclaved substrate would lead to a decreased development of biomass, final biomass, and survival rate. This current study found a significant difference between autoclaved- and non-autoclaved substrate in between days in the development of biomass, and a significant decreased final biomass and survival rate.
Similarly, Greenwood et al. (2021) observed that the microorganisms in the substrate significantly affect final biomass, and fat- and protein content of H. illucens larvae [35]. This is in accordance with previous studies, which showed microorganisms from the substrate are connected to gut microbial composition, as 66% of all microorganisms in the H. illucens larvae’s gut originate from the substrate [36]. These concepts give insight into why larvae are affected negatively when microorganisms are removed from the substrate in this current study. Therefore, the difference in development of biomass, final biomass, and survival rate observed, is not unexpected, considering the removal of essential microorganisms.
The decrease in biomass and survival could also be due to decreased availability of nutrients as a result of the Maillard reaction, which might be occurring during autoclaving. This leads to a decrease in the concentration of amino acids, but also a reduced digestibility of amino acids [37]. Another study by Hefnawy (2011) showed that autoclaving lentils (Lens culinaris) led to a decrease in different nutrients like sucrose and raffinose [38]. Therefore, it is possible that the autoclaved substrate in the current study lacks important nutrients which may affect both microorganisms and the larvae. These principles help explain the observed reduction in final biomass and decreased survival rate. Moreover, it might explain the impaired development of biomass, which is supported by the lower effect size when comparing to the control treatment (GES: 0.36 and 0.739, respectively). This suggests other factors than time are influencing the variance observed throughout the development. Somroo et al. (2019) also determined the positive effects of microorganisms on H. illucens larvae. With soybean curd residues as substrate for the larvae and the presence of Lactobacillus buchneri, parameters such as final biomass, fat- and protein content were increased compared to the absence of L. buchneri [39]. Overall, other studies and this current study show how important the substrate’s microorganisms are for larval performance.
4.3. The Effect of Disinfected Eggs and Autoclaved Substrate
It was expected that disinfection of eggs and autoclaving substrate would lead to a decreased development of biomass, final biomass, and survival rate. This current study found a significant difference between disinfected eggs and autoclaved substrate in development of biomass with a low effect size when comparing to control (GES: 0.07 and 0.739, respectively), indicating other factors than time are influencing the development. A significant difference for final biomass and survival rate were found when comparing with the control. The reasons for these findings might be the accumulation of all the previously listed effects of reduced nutrient digestibility and loss of beneficial microbiota.
4.4. Perspectives
It is not certain that the disinfected eggs and the autoclaved substrate were uncontaminated. Agar plates (see Appendix B) revealed that the autoclaved substrate had less colonies compared to the non-autoclaved. However, there were no visible colonies on the disinfected eggs. A beneficial addition to this study would be a 16S ribosomal RNA analysis to identify the quantity of different species of bacteria present in substrate and on eggs. Moreover, while the measure of final biomass was the day after peak biomass, it might have been more accurate to investigate by instar-stages instead, as this would assure all larvae have reached same stage in their life cycle.
Furthermore, some of the samples with autoclaved substrate were infected by fungus. This might be the result of an absence of fungicidal bacteria present on eggs [34]. The survival in samples with fungus were zero which might show the importance of fungicidal bacteria. Doing 16S ribosomal RNA analysis could uncover if these species of bacteria were absent. This study illustrates the importance of microorganism’s influence on M. domestica larvae’s biomass and survival. In addition, it is important to evaluate both the number of bacteria and species present when rearing M. domestica for optimisation of biomass and survival when establishing commercial production of M. domestica larvae for the purpose of maximising outcome.
Future studies are necessary to understand the full scope of the specific bacteria which are relevant for aforementioned parameters. Moreover, it could be interesting to investigate which physical conditions (pH, temperature, humidity etc.) are optimal for enhancing biomass and survival rate for production. Also, to investigate which substrates are most optimal in increasing development of biomass, final biomass, and the survival rate to maximize the outcome. Finally, it is crucial to uncover specifically which microorganisms are present on the eggs and in the substrate, and if possible, to discover the implications of each bacteria on the host.
5. Conclusions
In conclusion, disinfection of eggs, autoclaving of substrate, and both had a significant effect on the final biomass of the larvae of Musca domestica for all treatments in reference to the control. The survival rate decreased significantly for the treatments with both disinfected eggs and autoclaved substrate, and with only autoclaved substrate, but not for only disinfected eggs.
The development of larval biomass was also influenced by treatment with 76.8% and 73.9% of the variance explained by the time factor for only disinfected eggs and the control, respectively. For autoclaved substrate and both disinfected and autoclaved eggs and substrate, 36% and 7% of the variance, respectively, were explained by time indicating that other factors are affecting growth. Further research on more commercially relevant substrates would be crucial to uncover the full effect of microorganisms in insect production.
Author Contributions
Conceptualization, R.M.D, P.A.S., S.B, A.T.M, C.P., and T.M.S.; methodology, R. M. D. and P.A.S; software, R.M.D. and P.A.S.; validation, R.M.D, P.A.S., S.B, A.T.M, C.P; formal analysis, R.M.D. and P.A.S.; investigation, R.M.D. and P.A.S.; resources, S.B, C.P; data curation, R.M.D. and P.A.S.; writing—original draft preparation, R.M.D. and P.A.S.; writing—review and editing, R.M.D, P.A.S., S.B, A.T.M, C.P., and T.M.S.; visualization, R.M.D. and P.A.S.; supervision, S.B., A.T.M, C.P., and T. M. S.; project administration, S.B; funding acquisition, S.B, C.P. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by Aalborg Zoo Conservation Foundation, AZCF: grant number 07-2023. This work was supported by a grant from the Novo Nordic Foundation (Grant no. NNF21OC0070910).
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Conflicts of Interest
The authors declare no conflicts of interest.
Appendix A
Figure A1.
Agar plates with each 1 egg disinfected with different concentrations of sodium hypochlorite for experiment 1. 1 refers to a concentration of 0 % w/w, 2 refers to 0.006% w/w, 3 refers to 0.0075% w/w, 4 refers to 0.01% w/w, 5 refers to 0.015% w/w, and 6 refers to 0.03% w/w sodium hypochlorite. 7 refers to autoclaved substrate.
Figure A1.
Agar plates with each 1 egg disinfected with different concentrations of sodium hypochlorite for experiment 1. 1 refers to a concentration of 0 % w/w, 2 refers to 0.006% w/w, 3 refers to 0.0075% w/w, 4 refers to 0.01% w/w, 5 refers to 0.015% w/w, and 6 refers to 0.03% w/w sodium hypochlorite. 7 refers to autoclaved substrate.

Table A1.
Colonies and hatching rates (%) for each treatment with varying sodium hypochlorite, and colonies in the autoclaved substrate for experiment 1. Asterisk (*) indicates that it is an estimate.
Table A1.
Colonies and hatching rates (%) for each treatment with varying sodium hypochlorite, and colonies in the autoclaved substrate for experiment 1. Asterisk (*) indicates that it is an estimate.
| Disinfection | Colonies | Hatching Rates (%) |
|---|---|---|
| 0% w/w | 7 | 46.94 |
| 0.006% w/w | 0 | 38.78 |
| 0.0075% w/w | 0 | 18.37 |
| 0.001% w/w | 0 | 18.37 |
| 0.015% w/w | 25 * | 0 |
| 0.03% w/w | 20 | 0 |
| Autoclaved | 1 | N/A |
Appendix B
Figure A2.
Agar plates with each 25 eggs for experiment 2. Disinfection occurred with 0.006% w/w sodium hypochlorite. 1 refers to disinfected eggs, 2 refers to non-disinfected eggs, 3 refer to non-disinfected substrate and 4 refers to disinfected substrate.
Figure A2.
Agar plates with each 25 eggs for experiment 2. Disinfection occurred with 0.006% w/w sodium hypochlorite. 1 refers to disinfected eggs, 2 refers to non-disinfected eggs, 3 refer to non-disinfected substrate and 4 refers to disinfected substrate.
Table A2.
Number of colonies for disinfected eggs, non-disinfected eggs, non-autoclaved substrate, and autoclaved substrate for experiment 2.
Table A2.
Number of colonies for disinfected eggs, non-disinfected eggs, non-autoclaved substrate, and autoclaved substrate for experiment 2.
| Treatment | Colonies |
|---|---|
| Disinfected eggs | 0 |
| Non-disinfected eggs | 231 |
| Non-autoclaved | 6 |
| Autoclaved | 14 |
References
- Kraham, S. J. (2017). Environmental impacts of industrial livestock production. International Farm Animal, Wildlife and Food Safety Law, 3–40. [CrossRef]
- FAO. (2009). How to feed the world in 2050. https://www.fao.org/fileadmin/templates/wsfs/ docs/expert paper/How to Feed the World in 2050.pdf.
- Searchinger, T., Waite, R., Hanson, C., & Ranganathan, J. (2019). World resources report - creating sustainable food future. https://research.wri.org/sites/default/files/2019-07/WRRFoodFullReport0.pdf.
- Corrado, S., Caldeira, C., Eriksson, M., Hanssen, O. J., Hauser, H.-E., van Holsteijn, F., Liu, G., Ostergren, K., Parry, A., Secondi, L., Stenmarck, ¨ ˚A., & Sala, S. (2019). Food waste accounting methodologies: Challenges, opportunities, and further advancements. Global Food Security, 20, 93–100. [CrossRef]
- de Carvalho, N. M., Madureira, A. R., & Pintado, M. E. (2019). The potential of insects as food sources – a review. Critical Reviews in Food Science and Nutrition, 60(21), 3642–3652. [CrossRef]
- Cammack, J., Miranda, C., Jordan, H., & Tomberlin, J. (2021). Upcycling of manure with insects: Current and future prospects. Journal of Insects as Food and Feed, 7(5), 605– 619. [CrossRef]
- Varelas, V. (2019). Food wastes as a potential new source for edible insect mass production for food and feed: A review. Fermentation, 5(3), 81. [CrossRef]
- Caparros Megido, R., Francis, F., Haubruge, E., Le Gall, P., Tomberlin, J., Miranda, C., Jordan, H., Picard, C., Pino, M., Ramos-Elordy, J., Katz, E., Barragan Fonseca, K., Neto, E., Ponce-Reyes, R., Wijffels, G., Ghosh, S., Jung, C., Han, Y., Conti, B., & Huis, A. (2024). A worldwide overview of the status and prospects of edible insect production. Entomologia Generalis, 44. [CrossRef]
- Muurmann, A. T., Banovic, M., Gilbert, T. P., Sogari, G., Limborg, M. T., Sicheritz-Pont’en, T., & Bahrndorff, S. (2024). Framework for valorizing waste- and by-products through insects and their microbiomes for food and feed. Food Research International, 187, 114358. [CrossRef]
- Derrien, C., & Boccuni, A. (2018). Current status of the insect producing industry in europe. Springer International Publishing. [CrossRef]
- von Huis, A. (2013). Potential of insects as food and feed in assuring food security. Annual Reviews, 58, 563–583. [CrossRef]
- van Huis, A., & Gasco, L. (2023). Insects as feed for livestock production. Science, 379(6628), 138–139. [CrossRef]
- Ribeiro, N., Costa, R., & Ameixa, O. M. C. C. (2022). The influence of non-optimal rearing conditions and substrates on the performance of the black soldier fly (hermetia illucens). Insects, 13(7), 639. [CrossRef]
- Nyholm, S. V. (2020). In the beginning: Egg–microbe interactions and consequences for animal hosts. Philosophical Transactions of the Royal Society B, 375(1808). [CrossRef]
- Gupta, A., & Nair, S. (2020). Dynamics of insect–microbiome interaction influence host and microbial symbiont. Frontiers in Microbiology, 11, 17–34. [CrossRef]
- Freitak, D., Schmidtberg, H., Dickel, F., Lochnit, G., Vogel, H., & Vilcinskas, A. (2014). The maternal transfer of bacteria can mediate trans-generational immune priming in insects. Virulence, 5(4), 547–554. [CrossRef]
- Li, S.-J., Ahmed, M. Z., Lv, N., Shi, P.-Q., Wan, X.-M., Huang, J.-L., & Qiu, B.-L. (2017). Plant–mediated horizontal transmission of wolbachia between whiteflies. The ISME Journal, 11(4), 1019–1028. [CrossRef]
- Zhao, Y., Wang, W., Zhu, F., Wang, X., Wang, X., & Lei, C. (2017). The gut microbiota in larvae of the housefly musca domestica and their horizontal transfer through feeding. AMB Express, 7(147). [CrossRef]
- Lam, K., Babor, D., Duthie, B., Babor, E.-M., Moore, M., & Gries, G. (2007). Proliferating bacterial symbionts on house fly eggs affect oviposition behaviour of adult flies. Animal Behaviour, 74, 81–92. [CrossRef]
- Dillon, R. J., & Dillon, V. M. (2004). The gut bacteria of insects: Nonpathogenic interactions. Annual Review of Entomology, 49, 71–92. [CrossRef]
- Angel, P., & Moran, N. A. (2013). The gut microbiota of insects – diversity in structure and function. FEMS Microbiological Reviews, 37(5), 699–735. [CrossRef]
- Schreven, S. J. J., de Vries, H., Hermes, G. D. A., Smidt, H., Dicke, M., & van Loon, J. J. A. (2021). Relative contributions of egg-associated and substrate-associated microorganisms to black soldier fly larval performance and microbiota. FEMS Microbiology Ecology, 97(5). [CrossRef]
- Schmidt, K., & Engel, P. (2021). Mechanisms underlying gut microbiota–host interactions in insects. Journal of Experimental Biology, 224(2). [CrossRef]
- de Jonge, N., Michaelsen, T. Y., Ejbye-Ernst, R., Jensen, A., Nielsen, M. E., Bahrndorff, S., & Nielsen, J. L. (2020). Housefly (musca domestica l.) associated microbiota across different life stages. Scientific reports, 10. [CrossRef]
- Mazza, L., Xiao, X., ur Rehman, K., Cai, M., Zhang, D., Fasulo, S., Tomberlin, J. K., Zheng, L., Soomro, A. A., a, Z. Y., & Zhang, J. (2020). Management of chicken manure using black soldier fly (diptera: Stratiomyidae) larvae assisted by companion bacteria. Insects, 102, 312–318. [CrossRef]
- Mannaa, M., Mansour, A., Park, I., Lee, D.-W., & Seo, Y.-S. (2024). Insect-based agri-food waste valorization: Agricultural applications and roles of insect gut microbiotan. Environmental Science and Ecotechnology, 17. [CrossRef]
- Hussein, M., Pillai, V. V., Goddard, J. M., Park, H. G., Kothapalli, K. S., Ross, D. A., Ketterings, Q. M., Brenna, J. T., Milstein, M. B., Marquis, H., Johnson, P. A., Nyrop, J. P., & Selvaraj, V. (2017). Sustainable production of housefly (musca domestica) larvae as a protein-rich feed ingredient by utilizing cattle manure. PLOS one, 12(2). [CrossRef]
- Wickham, H. (2016). Ggplot2: Elegant graphics for data analysis. Springer-Verlag New York. https://ggplot2.tidyverse.org.
- Wickham, H., Averick, M., Bryan, J., Chang, W., McGowan, L. D., Fran¸cois, R., Grolemund, G., Hayes, A., Henry, L., Hester, J., Kuhn, M., Pedersen, T. L., Miller, E., Bache, S. M., M¨uller, K., Ooms, J., Robinson, D., Seidel, D. P., Spinu, V., . . . Yutani, H. (2019). Welcome to the tidyverse. Journal of Open Source Software, 4 (43), 1686. [CrossRef]
- Wickham, H., Fran¸cois, R., Henry, L., M¨uller, K., & Vaughan, D. (2023). Dplyr: A grammar of data manipulation. https://dplyr.tidyverse.org. Rpackage version 1.1.4, https://github.com/tidyverse/dplyr.
- Dinno, A. (2024). Dunn’s test of multiple comparisons using rank sums. https://cran.r-project.org/web/packages/dunn.test/dunn.test.pdf.
- Kassambara, A. (2023). Rstatix: Pipe-friendly framework for basic statistical tests. https://rpkgs.datanovia.com/rstatix/ R package version 0.7.2.
- Lam, K., Geisreiter, C., & Gries, G. (2009). Ovipositing female house flies provision offspring larvae with bacterial food. Entomologia Experimentalis et Applicata, 133(3), 292–295. [CrossRef]
- Lam, K., Thu, K., Tsang, M., Moore, M., & Gries, G. (2009). Bacteria on housefly eggs, musca domestica, suppress fungal growth in chicken manure through nutrient depletion or antifungal metabolites. The Science of Nature, 96, 1127–1132. [CrossRef]
- Greenwood, M. P., Hull, K. L., Brink-Hull, M., & Rhode, C. (2021). Feed and host genetics drive microbiome diversity with resultant consequences for production traits in mass-reared black soldier fly (Hermetia illucens) larvae. Insects, 12(12), 1082. [CrossRef]
- Jiang, C.-L., Jin, W.-Z., Tao, X.-H., Qian Zhang, J. Z., Feng, S.-Y., Xu, X.-H., Li, H.-Y., Wang, Z.-H., & Zhang, Z.-J. (2019). Black soldier fly larvae (hermetia illucens) strengthen the metabolic function of food waste biodegradation by gut microbiome. Microbial Biotechnology, 12(3), 528–543. [CrossRef]
- de Almeida, F. N. (2013). Effects of the maillard reactions on chemical composition and amino acid digestibility of feed ingredients and on pig growth performance. ProQuest Dissertations and Theses Global. https://www.proquest.com/dissertations-theses/effects-maillard-reactions-on-chemical/docview/1467482177/se-2.
- Hefnawy, T. H. (2011). Effect of processing methods on nutritional composition and antinutritional factors in lentils (lens culinaris). Annals of Agricultural Sciences, 56(2), 57– 61. [CrossRef]
- Somroo, A. A., ur Rehman, K., Zheng, L., a, M. C., a, X. X., Hu, S., Mathys, A., Gold, M., Yu, Z., & Zhang, J. (2019). Influence of lactobacillus buchneri on soybean curd residue co-conversion by black soldier fly larvae (hermetia illucens) for food and feedstock production. Waste Management, 86, 114–122. [CrossRef]
Figure 1.
Boxplots with the medians (25%-75% quantiles) of the biomass of larvae after the first five larvae were present in one sample for treatment A (disinfected eggs), B (autoclaved substrate), C (disinfected eggs and autoclaved substrate) and D (control). The x-axis shows time (days) with sample size in parentheses, and the y-axis shows the biomass (mg).
Figure 1.
Boxplots with the medians (25%-75% quantiles) of the biomass of larvae after the first five larvae were present in one sample for treatment A (disinfected eggs), B (autoclaved substrate), C (disinfected eggs and autoclaved substrate) and D (control). The x-axis shows time (days) with sample size in parentheses, and the y-axis shows the biomass (mg).
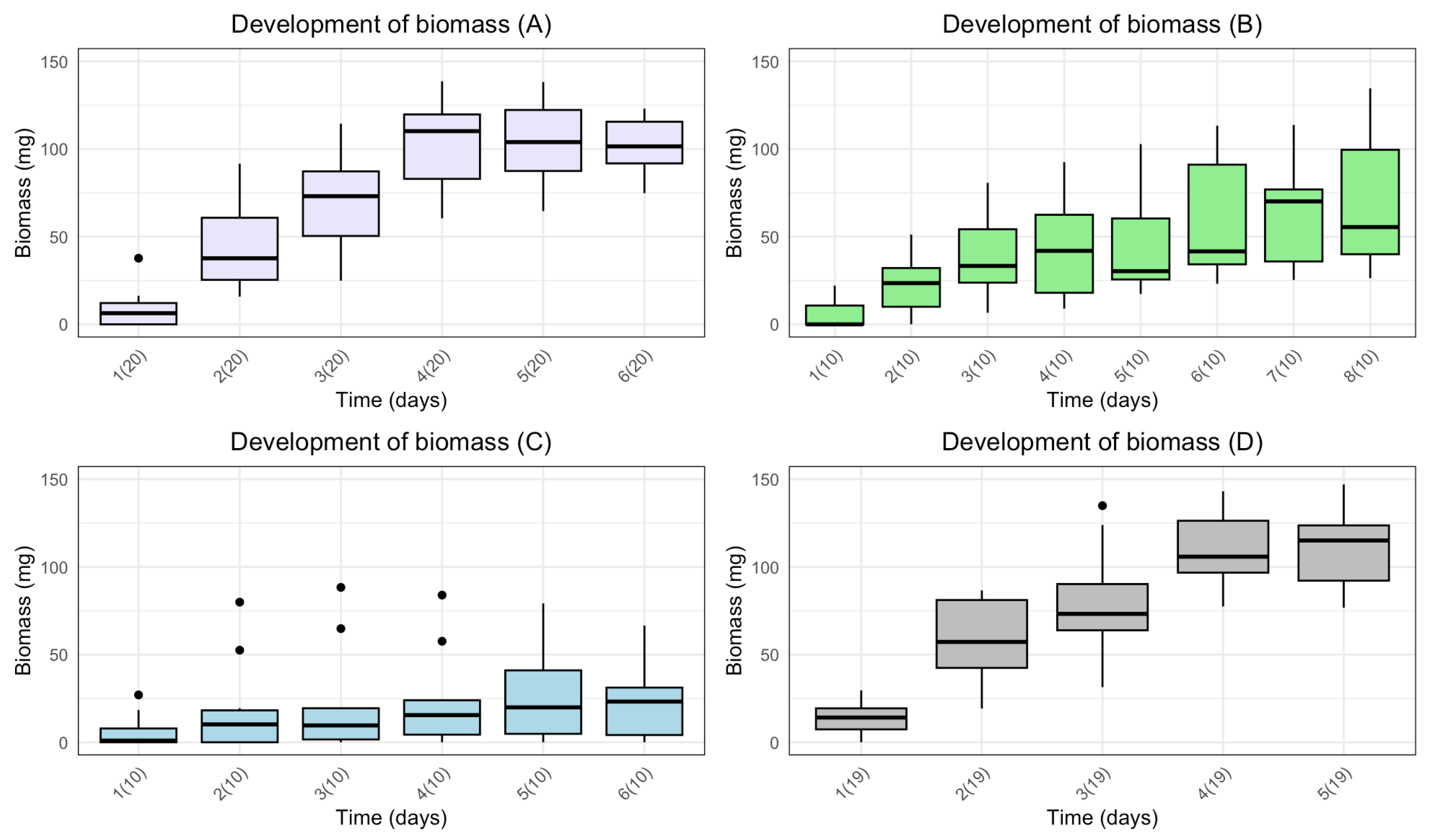
Figure 2.
Boxplots with the medians (25%-75% quantiles) of the final biomass of larvae for treatment A (disinfected eggs), B (autoclaved substrate), C (disinfected eggs and autoclaved substrate) and D (control). The x-axis shows treatments and sample size (N), and the y-axis shows the biomass (mg).
Figure 2.
Boxplots with the medians (25%-75% quantiles) of the final biomass of larvae for treatment A (disinfected eggs), B (autoclaved substrate), C (disinfected eggs and autoclaved substrate) and D (control). The x-axis shows treatments and sample size (N), and the y-axis shows the biomass (mg).
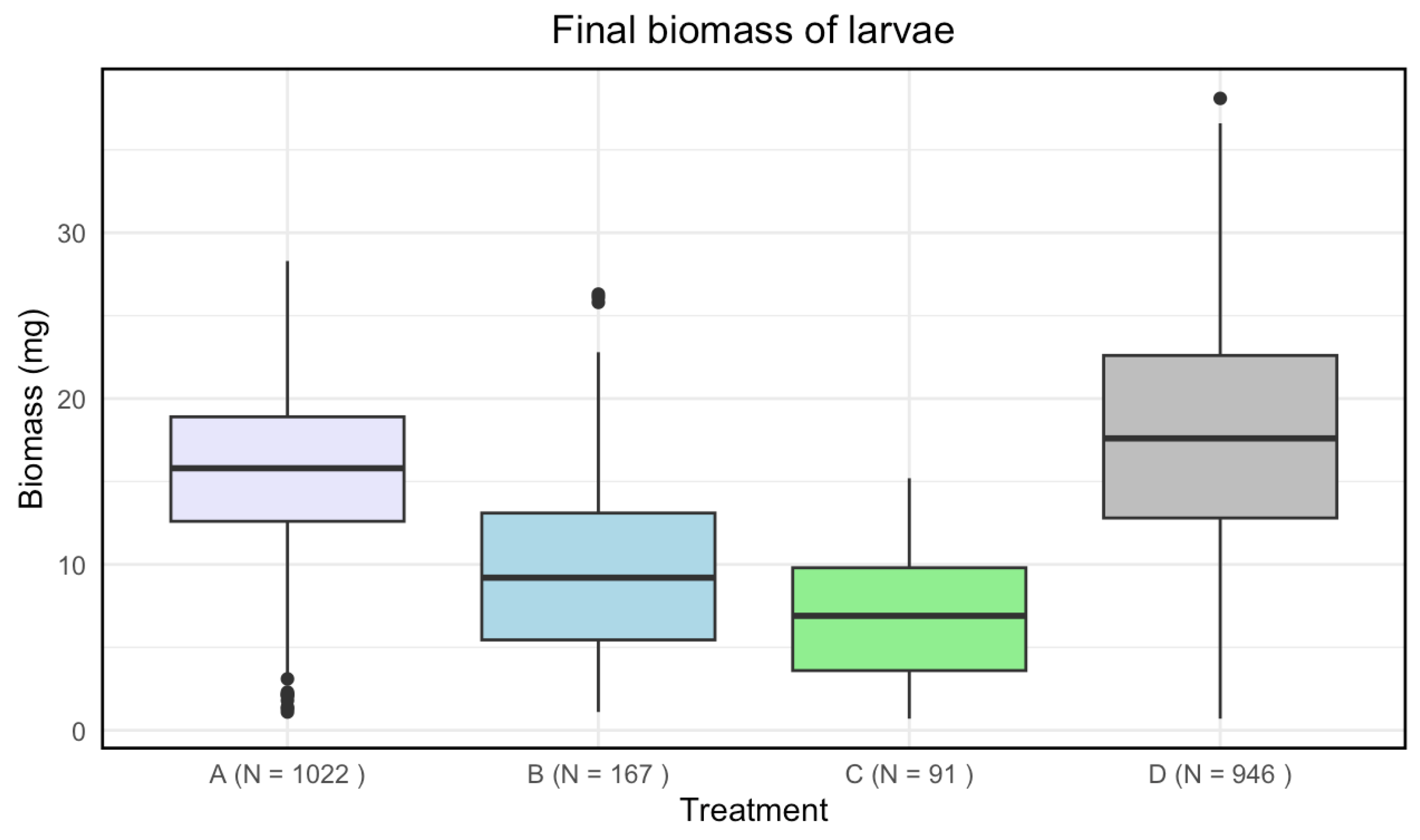
Figure 3.
Boxplots with the medians (25%-75% quantiles) of the survival rate of larvae for treatment A (disinfected eggs), B (autoclaved substrate), C (disinfected eggs and autoclaved substrate) and D (control). The x-axis shows treatments and sample size (N), and the y-axis shows the survival rate (%).
Figure 3.
Boxplots with the medians (25%-75% quantiles) of the survival rate of larvae for treatment A (disinfected eggs), B (autoclaved substrate), C (disinfected eggs and autoclaved substrate) and D (control). The x-axis shows treatments and sample size (N), and the y-axis shows the survival rate (%).
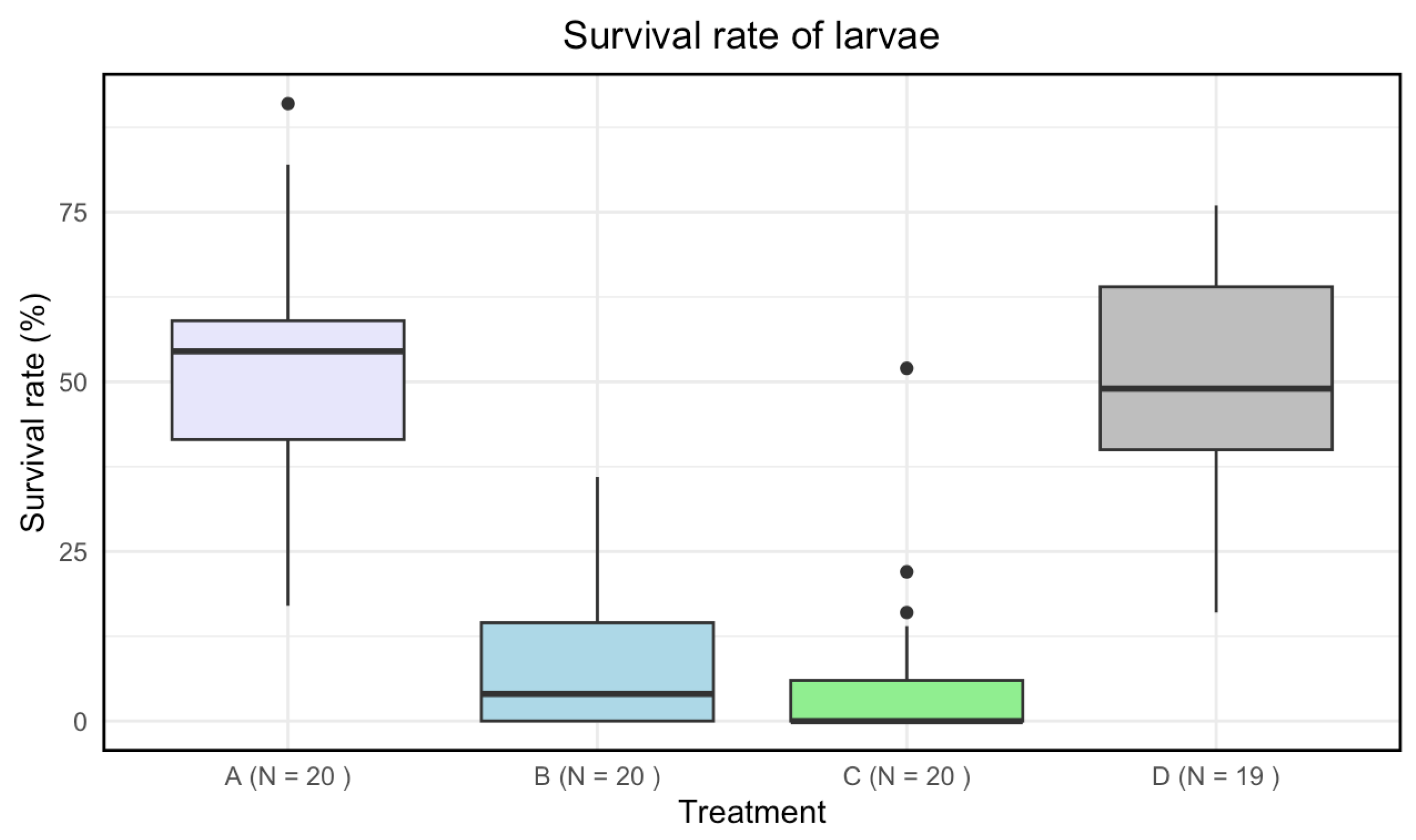
Table 1.
Concentrations of sodium hypochlorite ranged from lowest to highest concentration.
| Sodium Hypochlorite |
|---|
| 0% w/w |
| 0.006% w/w |
| 0.0075% w/w |
| 0.01% w/w |
| 0.015% w/w |
| 0.03% w/w |
Table 2.
Repeated measures ANOVA including p-value, F-value and generalised eta squared (GES) for treatment A (disinfected eggs), B (autoclaved substrate), C (disinfected eggs and autoclaved substrate) and D (control). Significant p-values are indicated with asterisk (*) when p<0.05.
Table 2.
Repeated measures ANOVA including p-value, F-value and generalised eta squared (GES) for treatment A (disinfected eggs), B (autoclaved substrate), C (disinfected eggs and autoclaved substrate) and D (control). Significant p-values are indicated with asterisk (*) when p<0.05.
| Treatment | p-Value | F-Value | GES |
|---|---|---|---|
| A | * | 159.253 | 0.768 |
| B | * | 14.481 | 0.36 |
| C | 0.028 * | 4.375 | 0.07 |
| D | * | 186.309 | 0.739 |
Table 3.
A Kruskal-Wallis indicated a significant effect of all treatments. A Dunn’s test was then performed between treatments, A (disinfected eggs), B (autoclaved substrate), C (disinfected eggs and autoclaved substrate) and D (control). Significant adjusted p-values are indicated with asterisk (*) when p < 0.05.
Table 3.
A Kruskal-Wallis indicated a significant effect of all treatments. A Dunn’s test was then performed between treatments, A (disinfected eggs), B (autoclaved substrate), C (disinfected eggs and autoclaved substrate) and D (control). Significant adjusted p-values are indicated with asterisk (*) when p < 0.05.
| Comparison | Adjusted p-Value |
|---|---|
| A-B | * |
| A-C | * |
| B-C | * |
| A-D | * |
| B-D | * |
| C-D | * |
Table 4.
A Kruskal-Wallis indicated a significant effect of all treatments. A Dunn’s test was then performed between treatments, A (disinfected eggs), B (autoclaved substrate), C (disinfected eggs and autoclaved substrate) and D (control). Significant adjusted p-values are indicated with asterisk (*) when p < 0.05 for both tests.
Table 4.
A Kruskal-Wallis indicated a significant effect of all treatments. A Dunn’s test was then performed between treatments, A (disinfected eggs), B (autoclaved substrate), C (disinfected eggs and autoclaved substrate) and D (control). Significant adjusted p-values are indicated with asterisk (*) when p < 0.05 for both tests.
| Comparison | Adjusted p-Value |
|---|---|
| A-B | * |
| A-C | * |
| B-C | 1 |
| A-D | 1 |
| B-D | * |
| C-D | * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



