
Submitted:
17 July 2024
Posted:
18 July 2024
You are already at the latest version
Abstract
Memory decline is considered a normal part of aging, while the relationship between melatonin deficiency and cognitive function is complex and not fully understood. The present study investigated the role of melatonin deficiency functioning as a hormone at different ages on working and short-term recognition and spatial memory in rats. An age-related decline in memory function was tested in the Y-maze, the object recognition test, and the radial arm maze. Aging was associated with reduced expression of brain-derived neurotrophic factor (BDNF) throughout the hippocampus, whereas pinealectomy exacerbated this process specifically in the CA3 region of 3- and 14-month-old rats. The region-specific reduced expression of the extracellular signal-regulated kinase (ERK)1/2 and pERK1/2 was observed in young adult rats with pinealectomy. However, in middle-aged rats, the expression of these signaling molecules was either downregulated or upregulated in different hippocampal regions. The reduced ratio of pCREB/CREB in the frontal cortex and hippocampus was associated with memory impairment in young adult and middle-aged melatonin-deficient rats, whereas old rats were unaffected. Our study provides insights into the molecular pathways involved in age-related memory changes associated with melatonin deficiency, highlighting the importance of the BDNF / ERK1/2 / CREB pathway in the hippocampus and suggesting a critical period for intervention.
Keywords:
Aging
; Melatonin deficiency
; Memory
; pCREB/CREB
; BDNF
; ERK1/2
; hippocampus
1. Introduction
Memory decline is often regarded as a normal part of the aging process. It is common for people to experience some changes in their memory as they get older. While the tendency for memory to decline with age is considered normal, severe or rapid memory decline may indicate more serious conditions such as Alzheimer’s disease (AD) or other forms of dementia. Weaver et al. [1] reported that up to 28% of relatively healthy older adults in one community had mild cognitive impairment in their responses to memory tests. Although such differences were not associated with age-related changes, education level, mood or health status, the early mild cognitive decline found may indicate early symptoms of AD [2]. Age-related neuroinflammation and vascular damage can disrupt the delicate balance of white matter integrity, affecting the transmission of neural signals across long distances in the brain. This can affect various cognitive functions, including learning, memory, and executive function, which are often impaired in vascular dementia [3].
Melatonin is a hormone produced by the pineal gland in the brain and plays a crucial role in regulating sleep, which indirectly benefits cognitive function. Some studies have investigated the possible relationship between melatonin and cognitive decline [4], particularly in the context of aging and neurodegenerative diseases [5]. Oxidative brain damage and neuroinflammation may be involved in several neurodegenerative diseases, including AD [6]. The potential benefits of melatonin are related to its neuroprotective, antioxidant and anti-inflammatory effects [7,8]. By reducing oxidative stress and inflammation in the brain, melatonin may help to maintain normal cognitive function. Accumulating clinical evidence suggests a strong relationship between cerebrospinal fluid (CSF) melatonin levels and memory impairment. For example, lower CSF melatonin has been shown to precede adverse changes in cognitive function in people at risk of developing AD and in patients with AD [5,9]. However, research on this topic is at an early stage and more studies are needed to provide conclusive evidence. Clinical studies have shown that plasma melatonin levels decrease significantly with age, dropping more than twice in older individuals compared to younger adults [10]. In addition, experimental studies, such as pinealectomy (surgical removal of the pineal gland), have shown that disrupting melatonin production accelerates the aging process [11].
There is a complex relationship between neurotrophic factors, in particular brain-derived neurotrophic factor (BDNF), and the extracellular signal-regulated kinase1/2 (ERK1/2) signalling molecules in the maintenance of cognitive function [12]. Disruption of their function may contribute to cognitive deficits and may be involved in conditions associated with neurodegeneration. ERK1/2 supports the formation of new dendritic spines and regulates several synaptic proteins, contributing to long-term potentiation (LTP), a process associated with learning and memory [13,14]. The phosphorylated ERK1/2 can, through phosphorylation, activate transcription factors critical for cognitive function, such as CREB (cAMP response element-binding protein) [14]. In addition, pCREB could induce the expression of factors essential for the regulation of various aspects of brain function, such as BDNF [15,16]. This neurotrophic factor influences processes closely related to cognition, such as neurogenesis and synaptic plasticity [13,17]. Loss of BDNF can lead to several detrimental effects, including synaptic dysfunction and cognitive impairment. Therefore, the interaction between BDNF, ERK1/2 signalling and downstream transcription factors such as CREB is essential for the maintenance of synaptic plasticity, and learning and memory processes. Disruption of this cascade can lead to cognitive deficits and may be implicated in neurodegenerative diseases. The MT1-ERK-CREB-BDNF signaling pathway represents one of the mechanisms through which melatonin affects memory processing [18].
Recently, we found that pinealectomy in young adult and middle-aged rats resulted in deleterious changes in specific physiological and metabolic functions. However, these effects were not observed in aged rats [19,20]. In the present study, we suggest that that melatonin deficiency, induced by pinealectomy, in rats can lead to cognitive impairments, especially in young adult and middle-aged rats, but not in aged rats. This suggests that melatonin may have a neuroprotective or cognitive-enhancing effect, particularly during certain developmental stages or periods of life. The proposed mechanism involves changes in the BDNF/ERK1/2/CREB pathway within the hippocampus. The hypothesis suggests that melatonin deficiency induced by pinealectomy could disrupt this pathway, leading to alterations in synaptic plasticity, neuronal survival, and gene expression within the hippocampus. These changes may contribute to the observed memory decline in young adult and middle-aged rats.
2. Results
2.1. Experimental Design
The experimental design is shown in Figure 1. At least ten days after surgery, the sham- and pin-operated rats from each group were tested in a battery of memory tests, including the Y-maze Ist trial, the Object Recognition Test (ORT), the Y-maze IInd trial (Experiment #1), and the Radial Arm Maze (RAM). Five days after the last test, rats from each group were divided into two subgroups for either biochemical analysis by ELISA or immunochystochemistry.
2.2. Melatonin Deficiеncy Associated with Pinealectomy Caused an Age-Dependent Impairment of Working and Short-Term Spatial and Recognition Memory, as Measured by the Y-Maze, ORT and RAM Test
The Y-maze test is widely used to assess working and short-term spatial memory in rodents. It requires little or no training and has been used to study memory changes in aged animals [21]. In the first Y-maze test, the middle-aged and old sham rats showed reduced spontaneous alternation behaviour (SAB) and number of arm entries compared to the matched young adult rats (p < 0.001, 3-month-old rats vs. 14- and 18-month-old rats) (Figure 2A,B). Pinealectomy did not affect SAB or motor activity in rats tested at three age stages: young adult, middle-aged and old rats (p > 0.05). In the second Y-maze test, rats were allowed to explore the two unrestrained arms for 10 min during the sample phase. The test phase was performed 5 min later with all arms of the Y-maze open. An age-related decrease in exploration time of the novel arm was observed (p = 0.004 and p < 0.001, 3-month-old sham vs. 14- and 18-month-old sham rats, respectively; p = 0.0007, 14-month-old sham vs. 18-month-old sham rats) (Figure 1C). Pinealectomy impaired short-term spatial memory in young adult and middle-aged rats, which showed a shorter exploration time of the novel arm compared to their matched controls (p < 0.001 and p = 0.002, respectively) (Figure 2C).
Short-term recognition memory was assessed 60 min after the acquisition phase of the object recognition test. The 18-month-old sham rats had a lower DI compared to the 3-month-old rats (p = 0.012), while pinealectomy reduced the DI in the youngest rats compared to their matched controls (p < 0.001) (Figure 3).
Both young adult and middle-aged rats that underwent a sham procedure showed the ability to learn the task (Figure 4A). In the youngest group, working memory errors (WMEs) and double working memory errors (DWMEs) in the RAM test decreased over time (p < 0.05, session five vs. session one within group) (Figure 4AB). Aging was associated with decreased spatial memory as evidenced by increased total WMEs and DWMEs (Figure 4CD). Aging was associated with increased total WMEs (p < 0.001, 3-month-old sham vs. 14- and 18-month-old sham rats) and DWMEs (p < 0.001, 3-month-old sham vs. 18-month-old sham rats).
Pinealectomy exacerbated the age-related decline in spatial memory performance in middle-aged rats. Thus, 14-month-old rats subjected to pinealectomy showed significantly higher total WMEs and DWMEs compared to matched controls (p < 0.001) (Figure 4C,D). Taken together, the results of the RAM test suggest that while aging is associated with a decline in spatial memory performance in rats, pinealectomy exacerbates this decline in middle-aged rats.
2.3. Melatonin Deficiency Induced by Pinealectomy Reduced BDNF in the CA3 Area of the Hippocampus in Young Adult and Middle-Aged Rats
The expression of BDNF in the dorsal hippocampus differed significantly within the sham groups tested at different ages, and a trend towards decreased expression was observed as follows: GrDG (p < 0.001, 3-month-old sham rats vs. 18-month-old sham rats; p = 0.01, 14-month-old sham rats vs. 18-month-old sham rats) (Figure 5; Figure 6A); the CA3 region (p < 0.001, 3-month-old sham rats vs. 18-month-old sham rats; p = 0.002, 14-month-old sham rats vs. 18-month-old sham rats) (Figure 5; Figure 6B), the CA2 region (p = 0.034, p < 0.001, 3-month-old sham rats vs. 14- and 18-month-old sham rats, respectively) (Figure 5; Figure 6C); the CA1 region (p < 0.001, 3-month-old sham rats vs. 18-month-old sham rats) (Figure 5; Figure 6D). However, pinealectomy additionally reduced BDNF expression, specifically in the CA3 region, in the vulnerable age stages, young adult and middle-aged rats (p < 0.001, 3-month-old pin rats vs. sham rats; p = 0.002, 14-month-old pin rats vs. sham rats) (Figure 5; Figure 6B).
2.4. Melatonin Deficiency Induced by Pinealectomy Reduced TrkB Receptor Expression in Middle-Aged Rats in the GrDG and in Young Adult Rats in the CA3 Region of the Hippocampus
Similar to BDNF, a trend towards age-related downregulation of the TrkB receptor was observed in all regions of the dorsal hippocampus examined, as follows: GrDG (p < 0.001, 3- and 14-month-old sham rats vs. 18-month-old sham rats); CA3 (p < 0.001, 3-month-old sham rats vs. 14- and 18-month-old sham rats); CA2 (p < 0.001, 3-month-old sham rats vs. 18-month-old sham rats); CA1 (p < 0.001, 3- vs. 14- and 18-month-old sham rats; p = 0.01, 14-month-old sham rats vs. 18-month-old sham rats) (Figure 7; Figure 8A-D). Removal of the pineal gland further reduced TrkB receptor expression in 14-month-old rats, specifically in the GrDG (p < 0.001 vs. control) (Figure 7; Figure 8A). Similar to BDNF expression, the CA3 region was vulnerable in the young adult rats, where a downregulation was detected after pinealectomy (p = 0.002 vs. 3-month-old sham rats) (Figure 7; Figure 8B).
2.5. Melatonin Deficiency Induced by Pinealectomy Caused Age- and Region-Specific Changes in the Expression of ERK1/2 and Its Phosphorylated form (pERK1/2) in the DG and Dorsal Hippocampus
The expression of extracellular signal-regulated kinase (ERK1/2) and its phosphorylated form (pERK1/2) followed a different pattern. A representative double immunofluorescence in the CA1 region of the dorsal hippocampus for pERK1/2 and NeuN and for pERK1/2 and GFAP in middle-aged sham rats is shown in Figure 9 (A1-A4) and (B1-B4), respectively. We found that the majority of mature NeuN was strongly co-labelled for pERK1/2 (Figure 9A2-A4) but not for GFAP (Figure 9B1-B4) in the CA1 region, so cells expressing these pERK1/2 were considered to be neurons. However, a low intensity of ERK1/2 signal was detected in the DG (Figure 9C1-C4) and as with pERK1/2, GFAP did not co-label for ERK1/2 (Figure 9D1-D4)
While ERK1/2 was scattered throughout the hippocampus, its activated form was detected in selected hippocampal subregions in different age groups and conditions. Similar to the expression of BDNF and its receptor, an age-related decrease was observed for ERK1/2 in GrDG (p < 0.001, 3-month-old sham rats vs. 14- and 18-month-old sham rats); CA3c stratum pyramidale (Py) (p < 0.001, 3-month-old sham rats vs. 14- and 18-month-old sham rats); stratum lucidum (SLu) (p = 0.002, 3-month-old sham rats vs. 18-month-old sham rats, p < 0.001, 14-month-old sham rats vs. 18-month-old sham rats); stratum radiatum (Rad) (p = 0.002, 3-month-old sham rats vs. 18-month-old sham rats, p = 0.003, 14-month-old sham rats vs. 18-month-old sham rats); stratum oriens (Or) (p = 0.006, 3-month-old sham rats vs. 18-month-old sham rats, p < 0.001, 3-month-old pin rats vs. 3-month-old sham rats); Py (p < 0.001, 3-month-old sham rats vs. 18-month-old sham rats; p < 0.001, 14-month-old sham rats vs. 18-month-old sham rats) (Figure 10, Figure 11A,B,D). As for BDNF and the TrkB receptor, pinealectomy in 3-month-old rats reduced ERK1/2 expression in GrDG, CA3c Py and CA3a Or and Py (p < 0.001 vs. sham rats) (Figure 10; Figure 11A,B,D). In contrast, melatonin deficiency in 14-month-old rats caused an increase in ERK1/2 expression in the GrDG, CA3c Py and CA3b Py (p < 0.001 vs. sham rats) (Figure 10, 11; Figure 12A-C). Aging decreased the ERK1/2 expression in both the CA2 Py and CA1 Py (3-month-old sham rats vs. 18-month-old sham rats (Figure 10; Figure 11E,F). PInealectomy reduced the ERK1/2 expression in selected layers of the CA2 field only in young adult rats (Or: *p = 0.044 vs. sham rats, Py: p < 0.001 vs sham rats) and CA1 field (Py: p < 0.001 vs sham rats, Rad: p=0.007, vs. sham rat).
The age-related tendency for decreased pERK1/2 expression was also detected in GrGD (p = 0.017, 3-month-old sham rats vs. 14-month-old sham rats; p < 0.001, 3-month-old sham rats vs. 18-month-old sham rats), PoDG (p < 0.001, 3-month-old sham rats vs. 14- and 18-month-old sham rats) (Figure 12; Figure 13A,B), CA3c region (p < 0. 001, 3-month-old sham rats vs. 14- and 18-month-old sham rats; 14-month-old sham rats vs. 18-month-old sham rats), CA3b region (p < 0.001, 3-month-old sham rats vs. 14- and 18-month-old sham rats), CA3a region (p < 0.001, 3-month-old sham rats vs. 14- and 18-month-old sham rats; 14-month-old sham rats vs. 18-month-old sham rats) (Figure 12; Figure 13C-E).
Pinealectomy resulted in a decreased expression of this signalling molecule in young adult rats in the GrDG, PoDG, CA3c, CA3b, CA3a, CA2, CA1 (p < 0.001, 3-month-old pin rats vs. their matched rats) (Figure 12; Figure 13F,G). As with ERK expression, changes in pERK expression in middle-aged pinealectomized rats were region specific. Thus, melatonin deficiency in this age group resulted in reduced pERK1/2 expression in the CA3a and CA2 regions, respectively (p < 0.001 vs. matched control) (Figure 13E,F). On the contrary, as for ERK1/2 expression, pinealectomy induced an increased expression of the phosphorylated form of this signalling molecule in GrDG (p < 0.001 vs. matched control), CA3c (p = 0.045 vs. matched group) and CA3b (p < 0.001 vs. matched control) (Figure 13A-D).
2.6. Melatonin Deficiency Associated with Pinealectomy Reduced the pCREB/CREB Ratio in the Frontal Cortex and Hippocampus in an Age-Dependent Manner
The ratio of pCREB/CREB in the frontal cortex (FC) and hippocampus was assessed in sham- and pin-operated young adult, middle-aged and old rats. An age-related decrease in the pCREB/CREB ratio was observed both in the FC (3- and 14-month-old sham rats vs. 18 month old matched rats, p = 0.004 and p = 0.008, respectively) and in the hippocampus (3- and 14-month-old sham rats vs. 18-month-old sham rats, p = 0.018 and p < 0.001, respectively) (Figure 14A,B). Furthermore, pinealectomy performed in young adult and middle-aged groups reduced the pCREB/CREB ratio in the FC (3- and 14-month-old pin groups vs. their matched group, p = 0.002 and p = 0.007, respectively) and in the hippocampus (3- and 14-month-old pin groups vs. their matched group, p = 0.036 and p < 0.001, respectively). In contrast, this ratio was not affected in old rats (p > 0.05).
3. Discussion
3.1. The role of ERK/CREB/BDNF Pathway in the Hippocampus in Aging-Related Cognitive Decline
Aging is characterized by a decline in cognitive function and structural degeneration of neural tissue. This process affects regions such as the hippocampus, which is critical for memory formation and retrieval [22]. The hippocampus plays a central role in memory consolidation and spatial navigation. These functions are severely impaired during the progression of diseases such as AD. The age-related decline in hippocampus-related memory appears to be a common phenomenon across species, reinforcing its biological and evolutionary importance [23]. Understanding the mechanisms and factors that contribute to age-related cognitive decline, particularly in the context of memory function, is crucial for developing strategies to promote healthy ageing in older individuals. Research on age-related memory decline in rats suggests that, similar to humans, rodents experience cognitive changes with age [24]. A battery of learning and memory tests was used to investigate the effect of pinealectomy at different ages on working and short-term spatial and recognition memory. We confirmed previous reports in aged rats and mice [25,26,27] of reduced learning capacity as demonstrated by the RAM test during the 5 days of training and impaired responses in memory tasks in aged rats compared to young adult rats. We used the Y-maze (1st trial) and the RAM test to assess working memory and hippocampus-dependent short-term spatial memory (Y-maze 2nd trial) and the ORT test to assess non-hippocampus-dependent short-term recognition memory. Factors such as oxidative stress, inflammation and changes in neurotransmitter levels have been reported to be positively correlated and to contribute to the underlying mechanisms of age-related progressive decline in cognitive function [27,28,29]. In the present study, we found evidence of memory deficits in middle-aged rats compared to young adults, contrary to previous notions that memory impairments only occur in old age (over 18 months in rats). Curiously, Stone et al. [30] reported poor responses in the Y-maze (1st trial) when SAB scores were assessed in 14-month-old Wistar rats. This correlated positively with impaired memory performance in the same rats in old age. We found that middle-aged rats had reduced responses to tasks involving working and short-term spatial memory, but not recognition memory. Oxidative stress has been implicated in age-related cognitive decline, caused by an imbalance between the production of reactive oxygen species (ROS) and the cell’s ability to scavenge them. Increased age-related oxidative stress in the hippocampus, previously reported by our team [20], is consistent with this suggestion, which may contribute to disrupted cellular processes, including BDNF transport, and thus potentially exacerbate hippocampal degeneration [31]. There is a close relationship between BDNF, ERK1/2 and CREB signalling molecules. These molecules are essential for the survival and maintenance of neurons in the hippocampus. They are critical for promoting neuronal growth, synaptic plasticity and overall neural health. Age-related hippocampal degeneration has been linked to a reduction in their expression and function. The proper function of neurons in the hippocampus depends on the transport of BDNF from target neurons [31]. Reduced BDNF levels and subsequent deficits in BDNF transport may lead to decreased ERK1/2 and CREB activation. This may further impair hippocampal function. In 18-month-old rats, behavioral outcomes correlated positively with decreased expression of BDNF, TrkB receptor, ERK1/2 and pERK1/2 in specific regions of the hippocampus, suggesting their involvement in age-related memory decline. Memory decline in middle-aged rats was associated with reduced expression of BDNF in the CA2 region, TrkB receptor in the CA3 and CA1 regions, and pERK1/2 in the GrDG, PoDG and CA3 regions of the dorsal hippocampus. These findings suggest that an early onset of molecular changes in different hippocampal regions may contribute to memory decline even in middle-aged rats.
Consistent with our findings, the age-related decrease in CREB in the hippocampus has been previously observed by other authors [32,33]. Notably, it has been reported that the age-related change in the pCREB/CREB ratio is not directly attributable to changes in neuronal cell number or to gross morphological changes in memory-related brain regions [34,35]. Instead, the observed differences in CREB-related signaling with age may be due to other factors such as changes in signaling pathways, epigenetic modifications or changes in the cellular microenvironment.
Taken together, the demonstrated positive association between reduced expression of BDNF, TrkB receptor, ERK1/2, pERK1/2 in specific hippocampal regions and the hippocampal pCREB/CREB ratio and memory decline underscores the importance of neurotrophic factor signaling and intracellular signaling pathways in maintaining hippocampal function and memory processes across different stages of aging.
3.2. Effect of Pinealetomy on Aging-Related Changes in BDNF, TrkB, ERK, pERK, CREB and pCREB Expression in the Hippocampus
Our results indicate that pinealectomy impairs working memory, short-term recognition and spatial memory in young adult and middle-aged rats, but has no effect on aged rats deficient in hormonal function of melatonin. Pinealectomy-induced susceptibility to cognitive changes in 3- and 14-month-old rats was associated with decreased expression of molecules involved in plastic changes in the hippocampus closely associated with memory processing, including the BDNF/ERK1/2/CREB pathway. The results of the present study are consistent with our previous reports [19,20,36] and those conducted by Pierpaoli [37,38,39]. The evidence for the importance of the pineal gland in physiological processes and its potential involvement in aging-related mechanisms is strengthened by this consistency across studies. In contrast to Pierpaoli [37,38,39], our previous studies [18,19,35] and the present findings suggest an essential role for the intact pineal gland in normal physiological processes not only in young adult but also in middle-aged animals. This is likely to include regulatory functions related to different aspects of physiology, such as hormone secretion, circadian regulation, and possibly other functions that are not yet fully understood. Aged rats with pinealectomy did not differ from matched sham rats in cognitive performance, which can be explained by the already reduced role of melatonin as a hormone in aging [41].
Cecon et al. [42] have shown that melatonin can influence the signaling pathway involved in memory formation through its interaction with several molecular targets. Some studies have suggested that the pineal gland may have an influence on aspects of memory and learning, possibly through the regulation of sleep patterns and circadian rhythms [43]. This includes its potential to influence the consolidation of memories during sleep and its involvement in the regulation of circadian rhythms, which in turn may affect cognitive processes. In the present study, we did not investigate whether melatonin supplementation is able to reverse a memory impairment in vulnerable age periods such as young adult and middle-aged rats, which is a limitation of the work. We can speculate that hormonal treatment may also have a beneficial effect in this paradigm, as has been previously reported for this model in our and other studies. [44]. Our findings are consistent with previous reports that pinealectomy impairs cognitive function in young adult rats [45,46]. However, there is a need to clarify the existing research on the effects of pinealectomy on aging, as the specific effects of pinealectomy on memory function are not well understood. We found that the pineal gland is essential for memory function, even in middle-aged rats in a region-specific mode. In another model of melatonin deficiency in young adult rats exposed to constant light, our team recently reported reduced BDNF expression in the hippocampus [46]. Given the complex relationship between BDNF and cognitive processes, it is plausible that BDNF may play a role in mediating the beneficial effects of melatonin on cognitive functions. BDNF has been implicated in the link between wakefulness and sleep structure. This suggests that it regulates the transitions between different sleep stages and wakefulness. A mechanism by which melatonin may exert its beneficial effects on cognitive function is suggested by its influence on sleep architecture and its potential to modulate BDNF levels. By promoting healthy sleep patterns and potentially increasing BDNF expression or activity, melatonin may improve synaptic plasticity, neurogenesis and overall cognitive performance. Acting through its receptors, particularly the MT1 receptor, melatonin can initiate intracellular signaling cascades, including the ERK pathway [18,47,48]. Once activated, ERK can phosphorylate various proteins within the cell, including CREB, ultimately leading to changes in gene expression and protein synthesis. In addition, the modification of the ERK signaling pathway by melatonin receptors leads to indirect phosphorylation and activation of CREB, specifically at the Ser133 residue. Phosphorylation of CREB at this site is associated with its activation and subsequent transcriptional activity [18]. Activated CREB promotes the expression of BDNF. Therefore, in vitro and in vivo models suggest that MT1-ERK-CREB-BDNF signaling pathway is a universal mechanism by which melatonin mediates memory processing.
Curiously, we found that middle-aged rats with pinealectomy had increased expression of both Erk 1/2 and its phosphorylated form in some regions of the hippocampus, including the GrDG and the CA3c and CA3b subfields, compared to the matched sham group. These changes were not consistent with the memory deficits found in this age group, suggesting that these regions are not critical for cognitive changes associated with melatonin deficiency in this age stage. However, future studies are needed to determine the significance of these alterations in middle-aged rats. We can speculate that they result from an adaptive mechanism that is not observed in young adult rats with pinealectomy. Recently, Pierpaoli and Bulian, 2005 [39] reported that pinealectomy in 14-month-old mice prolonged their lifespan and attenuated the expression of some age-associated blood markers, suggesting that pinealectomy at this age period may delay the aging process. However, our previous studies [19,20,36] and the present work showed that the role of melatonin as a hormone is more complicated and still has a significant impact on most of the functions closely associated with aging.
3.3. Limitation of the Study
Melatonin deficiency induced by pinealectomy may affect a variety of processes, not only BDNF reduction via either the MT receptor [42,49] or non-receptor related mechanisms [50]. In addition, the BDNF/ERK/CREB pathway is involved in the memory processing of several other endogenous molecules or conditions that may trigger this signaling [51,52,53,54]. Although this is a common signaling pathway that promotes cognitive function, melatonin can trigger it directly through a receptor mechanism, i.e., mediated by MT1 receptors. Our results suggest that this molecular mechanism in the action of the melatonin system is an age-dependent and critical in young adult and middle-aged rats. However, direct evidence for the receptor-associated mechanism needs to be confirmed using an MT1 receptor antagonist, which is supposed to mimic the effect of pinealectomy particularly in young adult and middle-aged rats. Additional experiments are needed to address this limitation of the present study.
Materials and Methods
4.1. Animals
Male Wistar rats, obtained from the vivarium of the Institute of Neurobiology, BAS, were employed in this study at three distinct age groups: 3-, 14-, and 18-month-old, respectively. The animals, with body weights ranging from 250–600 g were housed under standard environmental conditions, which included a consistent 12-hour light and 12-hour dark cycle, an average temperature maintained at 21˚, and a humidity level of 45%. They were accommodated in standard plexiglas cages of varying sizes depending on their age in groups of 3-4 rats per cage. Throughout the study, the animals had unrestricted access to both food and water, except during the testing periods. The experiments in the present study were performed in complete agreement with the European Communities Council Directive 2010/63/E.U. Animal experiments were approved by the research project (# 300/N◦5888–0183) of the Bulgarian Food Safety Agency.
4.2. Surgical Procedure
Rats from each age group were divided into two subgroups based on the surgical procedure: sham-operated and the group undergoing pinealectomy (pin)-operated. The surgical process involved anesthesia (2.5% isoflurane), careful mounting onto a stereotaxic apparatus (Stoelting, US) and pinealectomy. The removal of the pineal gland followed the method originally outlined by Hoffman and Reiter [55] and as previously described in our own studies [56].
4.3. Memory Tests
4.3.1. Y-Maze Test
The Y-maze apparatus was configured with three steel arms positioned at 120° angles from one another. Working memory was assessed by the Ist trial. Each rat was positioned in the center of the apparatus and given unrestricted access to explore the three arms for 8 minutes. The sequences of entries into these arms were manually recorded by two individuals who were unaware of the experimental schedule. The percent of spontaneous alternation behavior (SAB), based on visits to three different arms (referred to as a triad), was calculated as follows:
SAB % = (Number of correct entries x 100) / (Total entries (N)—2).
The IInd trial, which occurred at least 5 days after the first one, consisted of two sessions (a pretest and test). During this pretest, which was carried out for 10 minutes, one of the arms was closed off, allowing the tested rat to explore only two arms. After a 30-minute interval, the rat was placed in the arm opposite to one of the arms initially explored. The time spent and the number of entries into the familiar arm and the novel arm was detected.
4.3.2. Object Recognition Test (ORT)
The ORT was conducted following the methodology outlined in our prior study [57]. In brief, 24 hours after habituation to an empty open field (OF) apparatus (50 × 50 × 50 cm), the rat was placed within it alongside two identical plastic objects (referred to as “F”) for 5 minutes (Training session). Sixty minutes later, the rat was reintroduced into the same box for 5 minutes where one of the objects was substituted with a novel object (referred to as “N”) (Test session). The time spent (sec) and the counts (number) each object was explored through sniffing was detected during the Training and Testing session. The discrimination index was calculated as follows: (N) / (N + F).
4.3.3. Radial Arm Maze (RAM)
Spatial learning and memory were evaluated by an 8-arm radial maze (RAM) (Harvard Biosci. Comp., USA). The stainless-steel RAM apparatus is consisting of eight identical arms (42×12×12) radiating from a central octagonal platform (30 cm in diameter), elevated 50 cm off the floor. For the purpose to facilitate spatial navigation, picture signals (circle, square, triangle, and star) were available around the apparatus. A week before the training and the test, rats were put on a diet for at least 15 % of their body weight. Before the test performance, rats were habituated for 3 days (shaping) by exploring the maze for 15 min per day with food pellets in each of the eight arms (shaping). After the habituation, the animals performed the RAM test for 5 days (one session per day) for up to 10 minutes. At the end of each arm, one pellet was placed. The rat was placed in the center of the platform and allowed to freely search for available food. The test was interrupted when the rat ate all the pellets. The following parameters were recorded: number of working memory errors (WMEs)—the number of entries into an empty arm (with a consumed pellet) for the first time in each of 5 sessions; number of double WME (DWMEs): the number of consecutive entries into an empty arm from day 1 to day 5 of testing; time needed to consume all pellets per session; total WMEs, DWMEs and time: a sum of all WMEs, DWMEs and time in 5 sessions.
4.4. Е. nzyme-Linked Immunosorbent Assay (ELISA)
At the conclusion of the testing procedures, the rats were decapitated. Subsequently, their hippocampi were swiftly isolated under ice, rapidly frozen by immersion in liquid nitrogen, and then stored at -20 °C until the ELISA assay. The process of tissue homogenization followed the methodology detailed in a prior study [49].
The ELISA assay was conducted in accordance with the manufacturer’s guidelines provided from Sunlong Biotech Co., Ltd., China (SL1344Ra for Rat Phospho cAMP response element binding protein(P-CREB)) and Elabscience Biotechnology Inc., USA (E-EL-R0289 for Rat CREB1(Cyclic AMP Response Element Binding Protein). The ratio: pCREB/CREB was calculated.
4.5. Immunohistochemistry
After the experiments, Wistar rats from the three different age groups were deeply anaesthetized with urethane (1500 mg.kg-1). They were perfused transcardially first with 150 ml ice-cold 0.05 M phosphate-buffered saline (PBS), followed by 500 ml cold 4% paraformaldehyde (PFA) in 0.1 M phosphate buffer (PB), pH 7.4. After perfusion, the brains were removed and postfixed in the same fixative overnight at 4°C. After that, the tissue blocks were embedded in paraffin and cut into 6 μm thick coronal sections.
4.5.1. BDNF Expression in the Hippocampus
Sham-operated (sham) and experimental rats with pinealectomy aged 3, 14, and 18 months are examined four weeks after the surgical procedure for changes in BDNF/TrkB signaling pathways. We use fragments of the hippocampus of the experimental animals, processed by protocol and included in paraffin. The cuts are mounted on silane-coated slides. The slides are dewaxed alternately with xylene and a descending alcohol battery. An amplifying technique from the combination of ABC and PAP methods is used to identify BDNF/TrkB in the hippocampal layers. Then, we suppressed peroxidase with a solution of methanol and hydrogen peroxide for 30 minutes. We continue dewaxing with 96º, 80º, and 70º alcohol and finish with distilled water. Then, stay in PBS (phosphate-buffered saline) for 15 minutes and in Normal swine serum dissolved in PBS for 30 minutes. This procedure was followed by incubation with a specific anti-BDNF/TrkB antibody (Affinity Biosciences, dissolved 1:100). The duration of incubation was 24 h at 4º in a humid chamber. Then, it was washed with PBS and stayed for 60 minutes in the anti-rabbit HRP, flushed with PBS, and dried. The immune reaction was demonstrated by a highly sensitive PolyQ Stain 2-step detection kit (Quartett, Germany) with DAB as the chromogen. After completion of the reaction, we rinse with PBS, dehydrate in alcohols of upward concentration, and cover with Vecta mount. The specificity of immunostaining was confirmed on control slices in which the primary or secondary antibody was substituted by PBS, or only the peroxidase activity was visualized.
The immunohistochemical manifestations of the investigated signal system are subjected to quantitative analysis of the intensity of reactions by the use of the specialized software Olympus DP-Soft image system.
Quantitative analysis of the intensity of immunohistochemical reactions
Quantitative and statistical analysis of the intensity of the immunohistochemical reaction was performed using specialized software Olympus DP-Soft image system (version 4.1 for Windows) of a microscope Microphot-SA (Nikon, Japan), equipped with a digital camera Camedia-5050Z (Olympus, Japan). The intensity values of the reactions were in the range of 0÷256, with 0 presenting -white and 256—black. The analysis was carried out on hippocampal slices of experimental animals of different ages from the two study groups. The intensity curve of immune reactions in positive cells was calculated on different microscopic fields of slices (10 slices per animal, at magnification x400) of the hippocampus. The mean value of the intensity of antigenic expression for each animal in the groups was calculated, and BDNF/TrkB expression was recorded in relative units (RU).
4.6.2. ERK1/2 and pERK1/2 Expression in the Hippocampus
Immunohistochemistry for visualization of ERK1/2 and pERK1/2 was performed on dewaxed sections using UltraTek HRP Anti-Polyvalent Staining System (AFN600, ScyTek Laboratories, Inc., Logan, UT, USA). After antigen retrieval in Water bath thermostat WB-4MS (at a temperature of 95 degrees for 20 min in 0.01 M citrate buffer, pH 6.0) sections were washed in TBST (Tris Buffered Saline with 0.05% Tween-20, pH 7.6) and endogenous peroxidase was blocked in 3% (v/v) H2O2 in distilled water for 10 min at room temperature (RT). To block nonspecific binding sites, sections were incubated for 10 min with Super Block (ScyTek Laboratories, Inc., Logan, UT, USA) and after two 5-min washes in TBST, biotin block (ScyTek Laboratories, Inc., Logan, UT, USA), pretreatments were performed. All antibodies were diluted in Tris primary antibody diluent (ATG125, ScyTek Laboratories, Inc., USA). The following primary antibodies were incubated overnight at 4°C: rabbit polyclonal anti-ERK1/2 (1:400, Elabscience, E-AB-31374), and mouse monoclonal anti-ERK1/2 (phospho-T202/Y204) (1:600, Antibodies.com Bioworld Technology, A24956). After incubation with the primary antibodies, the sections were incubated sequentially with UltraTek Anti-Polyvalent Biotinylated and UltraTek HRP (ScyTek Laboratories, Inc., USA). DAB Chromogen/Substrate Kit (ScyTek Laboratories, Inc., USA) was used for color development for light microscopy. The sections were counterstained with hematoxylin, dehydrated before mounting and observed under a light research microscope Leica DM1000 equipped with a digital camera Leica DFC 290.
Fluorescent detection used a similar protocol starting from the Superblock pretreatment. The primary antibodies for immunofluorescence were mouse monoclonal [1B7] antibody to NeuN (1:3000, Antibodies.com A85405), mouse monoclonal anti-GFAP antibody (1:500, Elabscience®, E-AB-70205), mouse monoclonal anti-AIF1 antibody (1:1000, Elabscience®, E-AB-70353), rabbit polyclonal anti-ERK1/2 (1:400, Elabscience® E-AB-31374) and rabbit polyclonal anti-ERK1/2 (phospho-T202/Y204) (1:600, Bioworld Technology, A24956). A lipophilic fluorescent stain Hoechst 33342 Ultra Pure grade (CAS 23491-52-3) (1:1000, Santa Cruz Biotechnology, Inc., USA) for DNA labeling was used. Secondary antibodies goat anti-mouse IgG(H+L) (AF594 conjugated) (1:100, Elabscience®, E-AB-1059) and goat anti-rabbit IgG (H+L)(AF488 conjugated) (1:100, Elabscience®, E-AB-1055) were used for 1 hour at room temperature in the dark. The slides were mounted with FluoreGuard Mounting Medium (Hard Set) (FMH-060, ScyTek Laboratories, Inc., USA) and observed under a fluorescent microscope Leica TCS SPE equipped with a Leica Application Suite X (LAS X) Microscope Software, Version 3.5.7.23225 (Leica Microsystems GmbH).
Statistical Analysis
Data were analyzed by a two-way ANOVA with Age and Surgery used as factors with Bonferroni post hoc test. Data from the RAM test were analyzed by repeated ANOVA with additional factor Session. In case data were not normally distributed a non-parametric analysis was conducted (Kruskal-Wallis on ranks followed by the Mann-Whitney U test). The data were presented as mean±S.E.M. SigmaStat® 11.0 and GraphPad Prism 6 software were used for statistics and preparation of figures. The criterion for significant difference was accepted at p ≤ 0.05.
5. Conclusions
Overall, the results of the present study suggest that the effects of pinealectomy on cognitive function are influenced by age and melatonin status, with younger rats being more susceptible to cognitive impairment following pinealectomy, possibly due to disruptions in the BDNF/ERK1/2/CREB pathway in the hippocampus. Pinealectomy impaired several aspects of memory, including working memory, short-term recognition and spatial memory, in both young adult and middle-aged rats. However, the performance of aged rats with pinealectomy on memory tasks did not differ from that of matched sham rats. This lack of difference could be due to the already reduced role of melatonin as a hormone in aging. Thus, its absence due to pinealectomy may have less impact on cognitive function in aged rats compared to younger counterparts. Our study also suggests that pinealectomy-induced susceptibility to cognitive changes in young adult and middle-aged rats may be associated with reduced expression of molecules involved in plastic changes in the hippocampus, a brain region closely associated with memory processing. In particular, the study highlights changes in the BDNF/ERK1/2/CREB pathway, which is crucial for synaptic plasticity and memory formation.
Author Contributions
Conceptualization, J.T.; methodology, J.T.,D.A.,D.B., M.A., ,Z.N., M.M.; software, D.K.,D.A., D.B.; validation, J.T., D.A., N.L., M.A.; formal analysis, D.B.,I.K. D.K., Z.N.; resources, J.T.; data curation, J.T.,D.K..; writing—original draft preparation, J.T.; writing—review and editing, J.T.; visualization, M.M.; D.A.,P.R.; supervision, J.T.; project administration, J.T.; funding acquisition, J.T. All authors have read and agreed to the published version of the manuscript.”.
Funding
This work was supported by the National Science Fund of Bulgaria (research grant # KΠ -06-H41/1, 30 November 2020).
Institutional Review Board Statement
The study was conducted in accordance with the European Communities Council Directive 2010/63/E.U. Animal experiments were approved by the research project (# 300/N◦5888–0183, 2021) of the Bulgarian Food Safety Agency.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data is available in request.
Acknowledgments
We are grateful for the technical support in surgery procedures to Lidia Kortenska.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Weaver Cargin, J.; Maruff, P.; Collie, A.; Masters, C. Mild Memory Impairment in Healthy Older Adults Is Distinct from Normal Aging. Brain and Cognition 2006, 60, 146–155. [CrossRef]
- Grundman, M. Mild Cognitive Impairment Can Be Distinguished From Alzheimer Disease and Normal Aging for Clinical Trials. Arch Neurol 2004, 61, 59. [CrossRef]
- Finger, C.E.; Moreno-Gonzalez, I.; Gutierrez, A.; Moruno-Manchon, J.F.; McCullough, L.D. Age-related immune alterations and cerebrovascular inflammation. Mol Psychiatry 2022, 27, 803-818. [CrossRef]
- Xu, J.; Wang, L.-L.; Dammer, E.B.; Li, C.-B.; Xu, G.; Chen, S.-D.; Wang, G. Melatonin for Sleep Disorders and Cognition in Dementia: A Meta-Analysis of Randomized Controlled Trials. Am J Alzheimers Dis Other Demen 2015, 30, 439–447. [CrossRef]
- Wu, Y.-H.; Feenstra, M.G.P.; Zhou, J.-N.; Liu, R.-Y.; Toranõ, J.S.; Van Kan, H.J.M.; Fischer, D.F.; Ravid, R.; Swaab, D.F. Molecular Changes Underlying Reduced Pineal Melatonin Levels in Alzheimer Disease: Alterations in Preclinical and Clinical Stages. The Journal of Clinical Endocrinology & Metabolism 2003, 88, 5898–5906. [CrossRef]
- Song, T.; Song, X.; Zhu, C.; Patrick, R.; Skurla, M.; Santangelo, I.; Green, M.; Harper, D.; Ren, B.; Forester, B.P.; et al. Mitochondrial Dysfunction, Oxidative Stress, Neuroinflammation, and Metabolic Alterations in the Progression of Alzheimer’s Disease: A Meta-Analysis of in Vivo Magnetic Resonance Spectroscopy Studies. Ageing Research Reviews 2021, 72, 101503. [CrossRef]
- Pandi-Perumal, S.R.; BaHammam, A.S.; Brown, G.M.; Spence, D.W.; Bharti, V.K.; Kaur, C.; Hardeland, R.; Cardinali, D.P. Melatonin Antioxidative Defense: Therapeutical Implications for Aging and Neurodegenerative Processes. Neurotox Res 2013, 23, 267–300. [CrossRef]
- Cho, J.H.; Bhutani, S.; Kim, C.H.; Irwin, M.R. Anti-Inflammatory Effects of Melatonin: A Systematic Review and Meta-Analysis of Clinical Trials. Brain, Behavior, and Immunity 2021, 93, 245–253. [CrossRef]
- Zhou, J.; Liu, R.; Kamphorst, W.; Hofman, M.A.; Swaab, D.F. Early Neuropathological Alzheimer’s Changes in Aged Individuals Are Accompanied by Decreased Cerebrospinal Fluid Melatonin Levels. Journal of Pineal Research 2003, 35, 125–130. [CrossRef]
- Priyadarshini, R.; Raj, G.M.; Shewade, D.G. Pathophysiological and pharmacological modulation of melatonergic system. Int J Basic Clin Pharmacol. 2015, 4, 32–639. [CrossRef]
- Pierpaoli, W.; Bulian, D. The Pineal Aging and Death Program: Life Prolongation in Pre-aging Pinealectomized Mice. Annals of the New York Academy of Sciences 2005, 1057, 133–144. [CrossRef]
- Alonso, M.; Medina, J.H.; Pozzo-Miller, L. ERK1/2 Activation Is Necessary for BDNF to Increase Dendritic Spine Density in Hippocampal CA1 Pyramidal Neurons. Learn. Mem. 2004, 11, 172–178. [CrossRef]
- Tartaglia, N.; Du, J.; Tyler, W.J.; Neale, E.; Pozzo-Miller, L.; Lu, B. Protein Synthesis-Dependent and -Independent Regulation of Hippocampal Synapses by Brain-Derived Neurotrophic Factor. Journal of Biological Chemistry 2001, 276, 37585–37593. [CrossRef]
- Peng, S.; Zhang, Y.; Zhang, J.; Wang, H.; Ren, B. ERK in Learning and Memory: A Review of Recent Research. IJMS 2010, 11, 222–232. [CrossRef]
- Medina, J.H.; Viola, H. ERK1/2: A Key Cellular Component for the Formation, Retrieval, Reconsolidation and Persistence of Memory. Front. Mol. Neurosci. 2018, 11, 361. [CrossRef]
- Josiane; Tatiani; Francielle; Michelle; Alexandra The Involvement of BDNF, NGF and GDNF in Aging and Alzheimer’s Disease. Aging and disease 2015, 6, 331. [CrossRef]
- Albert-Gascó, H.; Ros-Bernal, F.; Castillo-Gómez, E.; Olucha-Bordonau, F.E. MAP/ERK Signaling in Developing Cognitive and Emotional Function and Its Effect on Pathological and Neurodegenerative Processes. IJMS 2020, 21, 4471. [CrossRef]
- Zakaria, R.; Ahmad, A.H.; Othman, Z. The Potential Role of Melatonin on Memory Function: Lessons from Rodent Studies. Folia Biol (Praha) 2016, 62, 181–187.
- Tchekalarova, J.; Hrischev, P.; Ivanova, P.; Boyadjiev, N.; Georgieva, K. Metabolic Footprint in Young, Middle-Aged and Elderly Rats with Melatonin Deficit. Physiology & Behavior 2022, 250, 113786. [CrossRef]
- Tchekalarova, J.; Nenchovska, Z.; Kortenska, L.; Uzunova, V.; Georgieva, I.; Tzoneva, R. Impact of Melatonin Deficit on Emotional Status and Oxidative Stress-Induced Changes in Sphingomyelin and Cholesterol Level in Young Adult, Mature, and Aged Rats. IJMS 2022, 23, 2809. [CrossRef]
- Prieur, E.; Jadavji, N. Assessing Spatial Working Memory Using the Spontaneous Alternation Y-Maze Test in Aged Male Mice. BIO-PROTOCOL 2019, 9. [CrossRef]
- Bettio, L.E.B.; Rajendran, L.; Gil-Mohapel, J. The Effects of Aging in the Hippocampus and Cognitive Decline. Neuroscience & Biobehavioral Reviews 2017, 79, 66–86. [CrossRef]
- Leal, S.L.; Yassa, M.A. Neurocognitive Aging and the Hippocampus across Species. Trends in Neurosciences 2015, 38, 800–812. [CrossRef]
- Foster, T.C. Dissecting the Age-Related Decline on Spatial Learning and Memory Tasks in Rodent Models: N-Methyl-D-Aspartate Receptors and Voltage-Dependent Ca2+ Channels in Senescent Synaptic Plasticity. Progress in Neurobiology 2012, 96, 283–303. [CrossRef]
- Gerrard, J.L.; Burke, S.N.; McNaughton, B.L.; Barnes, C.A. Sequence Reactivation in the Hippocampus Is Impaired in Aged Rats. J. Neurosci. 2008, 28, 7883–7890. [CrossRef]
- Smith, B.M.; Yao, X.; Chen, K.S.; Kirby, E.D. A Larger Social Network Enhances Novel Object Location Memory and Reduces Hippocampal Microgliosis in Aged Mice. Front. Aging Neurosci. 2018, 10, 142. [CrossRef]
- Scali, C.; Giovannini, M.G.; Prosperi, C.; Bartolini, L.; Pepeu, G. TACRINE ADMINISTRATION ENHANCES EXTRACELLULAR ACETYLCHOLINEIN VIVOAND RESTORES THE COGNITIVE IMPAIRMENT IN AGED RATS. Pharmacological Research 1997, 36, 463–469. [CrossRef]
- The American Association for Research into Nervous and Mental Diseases; Albert, M.S. The Ageing Brain: Normal and Abnormal Memory. Phil. Trans. R. Soc. Lond. B 1997, 352, 1703–1709. [CrossRef]
- Haider, S.; Saleem, S.; Perveen, T.; Tabassum, S.; Batool, Z.; Sadir, S.; Liaquat, L.; Madiha, S. Age-Related Learning and Memory Deficits in Rats: Role of Altered Brain Neurotransmitters, Acetylcholinesterase Activity and Changes in Antioxidant Defense System. AGE 2014, 36, 9653. [CrossRef]
- Stone, W.S.; Rudd, R.J.; Parsons, M.W.; Gold, P.E. Memory Scores in Middle-Aged Rats Predict Later Deficits in Memory, Paradoxical Sleep, and Blood Glucose Regulation in Old Age. Experimental Aging Research 1997, 23, 287–300. [CrossRef]
- Shekari, A.; Fahnestock, M. Retrograde Axonal Transport of BDNF and ProNGF Diminishes with Age in Basal Forebrain Cholinergic Neurons. Neurobiology of Aging 2019, 84, 131–140. [CrossRef]
- Countryman, R.A.; Gold, P.E. Rapid Forgetting of Social Transmission of Food Preferences in Aged Rats: Relationship to Hippocampal CREB Activation. Learn. Mem. 2007, 14, 350–358. [CrossRef]
- Morris, K.A.; Gold, P.E. Age-Related Impairments in Memory and in CREB and PCREB Expression in Hippocampus and Amygdala Following Inhibitory Avoidance Training. Mechanisms of Ageing and Development 2012, 133, 291–299. [CrossRef]
- Gallagher, M. Effects of Aging on the Hippocampal Formation in a Naturally Occurring Animal Model of Mild Cognitive Impairment. Experimental Gerontology 2003, 38, 71–77. [CrossRef]
- Rasmussen, T.; Schliemann, T.; Sørensen, J.C.; Zimmer, J.; West, M.J. Memory Impaired Aged Rats: No Loss of Principal Hippocampal and Subicular Neurons. Neurobiology of Aging 1996, 17, 143–147. [CrossRef]
- Tchekalarova, J.; Krushovlieva, D.; Ivanova, P.; Nenchovska, Z.; Toteva, G.; Atanasova, M. The Role of Melatonin Deficiency Induced by Pinealectomy on Motor Activity and Anxiety Responses in Young Adult, Middle-Aged and Old Rats. Behav Brain Funct 2024, 20, 3. [CrossRef]
- Pierpaoli, W. The Pineal Gland: A Circadian or Seasonal Aging Clock? Aging Clin Exp Res 1991, 3, 99–101. [CrossRef]
- Pierpaoli, W. The Pineal Gland as Ontogenetic Scanner of Reproduction, Immunity, and Aging The Aging Clock. Ann NY Acad Sci 1994, 741, 46–49. [CrossRef]
- Pierpaoli, W.; Bulian, D. The Pineal Aging and Death Program. I. Grafting of Old Pineals in Young Mice Accelerates Their Aging. Journal of Anti-Aging Medicine 2001, 4, 31–37. [CrossRef]
- Pierpaoli, W.; Bulian, D. The Pineal Aging and Death Program: Life Prolongation in Pre-aging Pinealectomized Mice. Annals of the New York Academy of Sciences 2005, 1057, 133–144. [CrossRef]
- Wurtman, R.J. Age-Related Decreases in Melatonin Secretion—Clinical Consequences. The Journal of Clinical Endocrinology & Metabolism 2000, 85, 2135–2136. [CrossRef]
- Cecon, E.; Oishi, A.; Jockers, R. Melatonin Receptors: Molecular Pharmacology and Signalling in the Context of System Bias. British J Pharmacology 2018, 175, 3263–3280. [CrossRef]
- De Butte, M.; Fortin, T.; Pappas, B.A. Pinealectomy: Behavioral and Neuropathological Consequences in a Chronic Cerebral Hypoperfusion Model. Neurobiology of Aging 2002, 23, 309–317. [CrossRef]
- Tzoneva, R.; Georgieva, I.; Ivanova, N.; Uzunova, V.; Nenchovska, Z.; Apostolova, S.;Stoyanova, T.; Tchekalarova, J. The Role of Melatonin on Behavioral Changes and Concomitant Oxidative Stress in icvA_1-42 Rat Model with Pinealectomy. Int. J. Mol. Sci. 2021, 22, 12763. [CrossRef]
- Karakaş, A.; Coşkun, H.; Kaya, A. The Effects of Pinealectomy, Melatonin Injections and Implants on the Spatial Memory Performance of Male Wistar Rats. Biological Rhythm Research 2011, 42, 457–472. [CrossRef]
- Tchekalarova, J.; Kortenska, L.; Ivanova, N.; Atanasova, M.; Marinov, P. Agomelatine Treatment Corrects Impaired Sleep-Wake Cycle and Sleep Architecture and Increases MT1 Receptor as Well as BDNF Expression in the Hippocampus during the Subjective Light Phase of Rats Exposed to Chronic Constant Light. Psychopharmacology 2020, 237, 503–518. [CrossRef]
- Shin, E.-J.; Chung, Y.H.; Le, H.-L.T.; Jeong, J.H.; Dang, D.-K.; Nam, Y.; Wie, M.B.; Nah, S.-Y.; Nabeshima, Y.-I.; Nabeshima, T.; et al. Melatonin Attenuates Memory Impairment Induced by Klotho Gene Deficiency Via Interactive Signaling Between MT2 Receptor, ERK, and Nrf2-Related Antioxidant Potential. International Journal of Neuropsychopharmacology 2015, 18. [CrossRef]
- Dash, P.K.; Karl, K.A.; Colicos, M.A.; Prywes, R.; Kandel, E.R. CAMP Response Element-Binding Protein Is Activated by Ca2+/Calmodulin- as Well as CAMP-Dependent Protein Kinase. Proc. Natl. Acad. Sci. U.S.A. 1991, 88, 5061–5065. [CrossRef]
- Mihanfar, A.; Yousefi, B.; Azizzadeh, B. et al. Interactions of melatonin with various signaling pathways: implications for cancer therapy. Cancer Cell Int 2022, 22, 420. [CrossRef]
- Amini, H.; Rezabakhsh, A.; Heidarzadeh, M.; Hassanpour, M.; Hashemzadeh, S.; Ghaderi, S.; Sokullu, E.; Rahbarghazi, R.; Reiter, R.J. An Examination of the Putative Role of Melatonin in Exosome Biogenesis. Front. Cell Dev. Biol. 2021, 9, 686551. [CrossRef]
- Smith. N.K.; Hackett, T.A.; Galli, A.; Flynn, C.R. GLP-1: Molecular mechanisms and outcomes of a complex signaling system. Neurochem Int. 2019 128, 94-105. [CrossRef]
- Kim, M.-S.; Kim, B.-Y.; Kim, J.I.; Lee, J.; Jeon, W.K. Mumefural Improves Recognition Memory and Alters ERK-CREB-BDNF Signaling in a Mouse Model of Chronic Cerebral Hypoperfusion. Nutrients 2023, 15, 3271. [CrossRef]
- Yan, L.; Xu, X.; He, Z.; Wang, S.; Zhao, L.; Qiu, J.; Wang, D.; Gong, Z.; Qiu, X.; Huang, H. Antidepressant-Like Effects and Cognitive Enhancement of Coadministration of Chaihu Shugan San and Fluoxetine: Dependent on the BDNF-ERK-CREB Signaling Pathway in the Hippocampus and Frontal Cortex. Biomed Res Int. 2020, 2020, 2794263. [CrossRef]
- Pak, M.E.; Yang, H.J.; Li, W.; Kim, J.K.; Go, Y. Yuk-Gunja-Tang attenuates neuronal death and memory impairment via ERK/CREB/BDNF signaling in the hippocampi of experimental Alzheimer’s disease model. Front. Pharmacol. 2022, 13, 1014840. [CrossRef]
- Hoffman, R.A., Reiter, R.J. Rapid pinealectomy in hamsters and other small rodents. Anat Rec. 1965, 153, 19-21. [CrossRef]
- Tchekalarova, J.; Nenchovska, Z.; Atanasova, D.; Atanasova, M.; Kortenska, L.; Stefanova, M.; Alova, L.; Lazarov, N. Consequences of Long-Term Treatment with Agomelatine on Depressive-like Behavior and Neurobiological Abnormalities in Pinealectomized Rats. Behavioural Brain Research 2016, 302, 11–28. [CrossRef]
- Shishmanova-Doseva, M.; Tchekalarova, J.; Nenchovska, Z.; Ivanova, N.; Georgieva, K.; Peychev, L. The Effect of Chronic Treatment with Lacosamide and Topiramate on Cognitive Functions and Impaired Emotional Responses in a Pilocarpine-induced Post-status Epilepticus Rat Model. Folia Medica 2020, 62, 723-9. [CrossRef]
Figure 1.
Timeline of experimental steps.
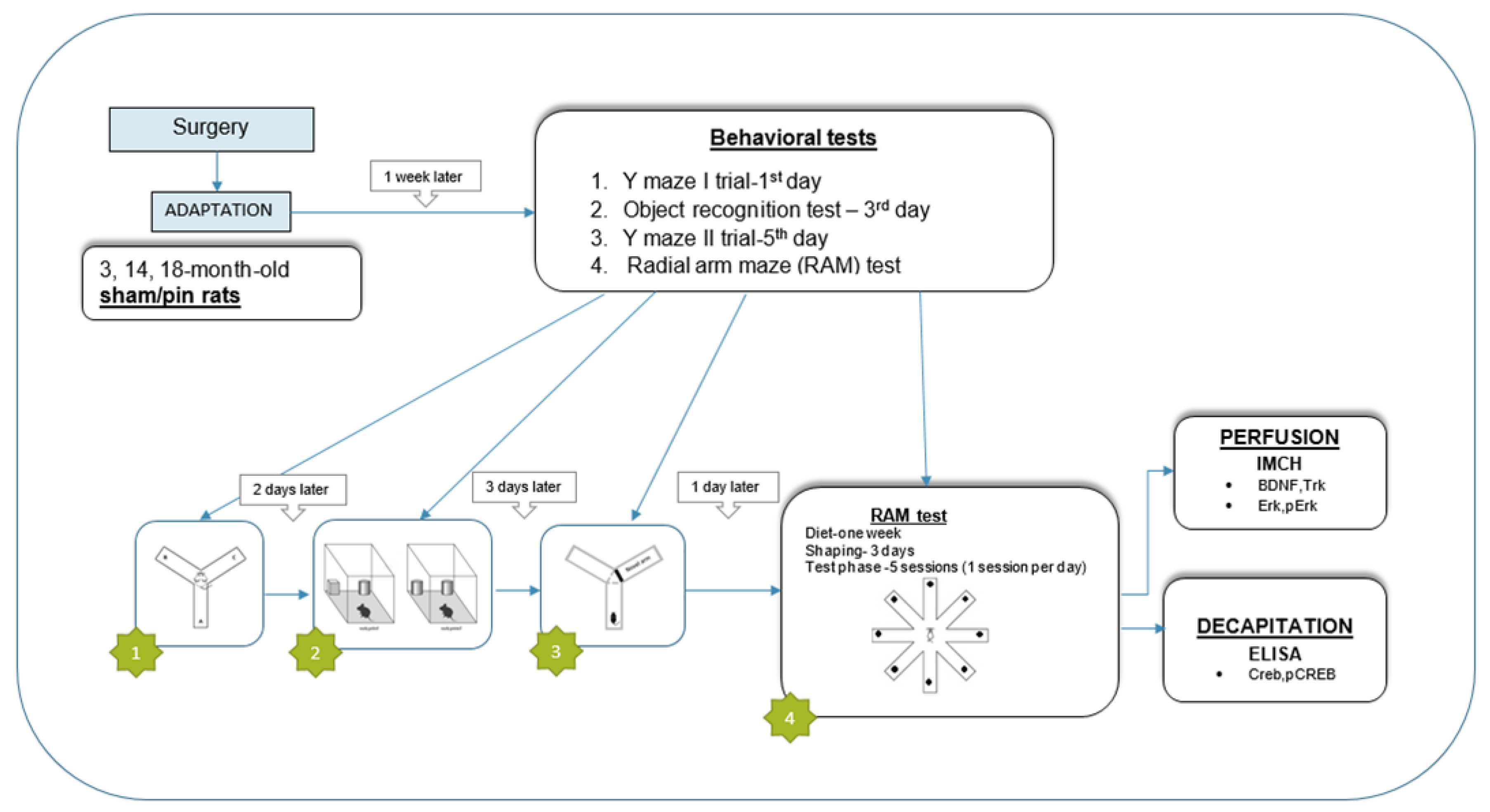
Figure 2.
Effect of aging and pinealectomy on working memory and short-term spatial memory in 3-, 14-, and 18-month-old rats evaluated in the Y-maze test: (A) the Ist trial—Spontaneous alternation behavior (SAB) in %; (B) the Ist trial—number of arm entries; (C) the IInd trial—Discrimination Index (DI). Data are presented as mean ± SEM, n = 10. Two-way ANOVA, Age effect: [F2,59 = 24.87, p < 0.001] (A) and [F2,59 = 26.773, p < 0.001] (B); Age effect: [F2,59 = 24.868, p < 0.001] and Surgery effect: [F1,59 = 57.112, p < 0.001] (C). ***p < 0.001, 3-month-old rats vs. 14-month-old rats and 18-month-old rats, respectively (A,B); **p = 0.004, 3-month-old sham vs. 14-month-old sham rats; ***p < 0.001, 3-month-old sham vs. 18-month-old sham rats; ***p = 0.0007, 14-month-old rats vs. 18-month-old rats; ***p < 0.001, 3-month-old pinealectomy (pin) rats vs. matched control; **p = 0.002, 14-month-old pin rats vs. matched control (C).
Figure 2.
Effect of aging and pinealectomy on working memory and short-term spatial memory in 3-, 14-, and 18-month-old rats evaluated in the Y-maze test: (A) the Ist trial—Spontaneous alternation behavior (SAB) in %; (B) the Ist trial—number of arm entries; (C) the IInd trial—Discrimination Index (DI). Data are presented as mean ± SEM, n = 10. Two-way ANOVA, Age effect: [F2,59 = 24.87, p < 0.001] (A) and [F2,59 = 26.773, p < 0.001] (B); Age effect: [F2,59 = 24.868, p < 0.001] and Surgery effect: [F1,59 = 57.112, p < 0.001] (C). ***p < 0.001, 3-month-old rats vs. 14-month-old rats and 18-month-old rats, respectively (A,B); **p = 0.004, 3-month-old sham vs. 14-month-old sham rats; ***p < 0.001, 3-month-old sham vs. 18-month-old sham rats; ***p = 0.0007, 14-month-old rats vs. 18-month-old rats; ***p < 0.001, 3-month-old pinealectomy (pin) rats vs. matched control; **p = 0.002, 14-month-old pin rats vs. matched control (C).
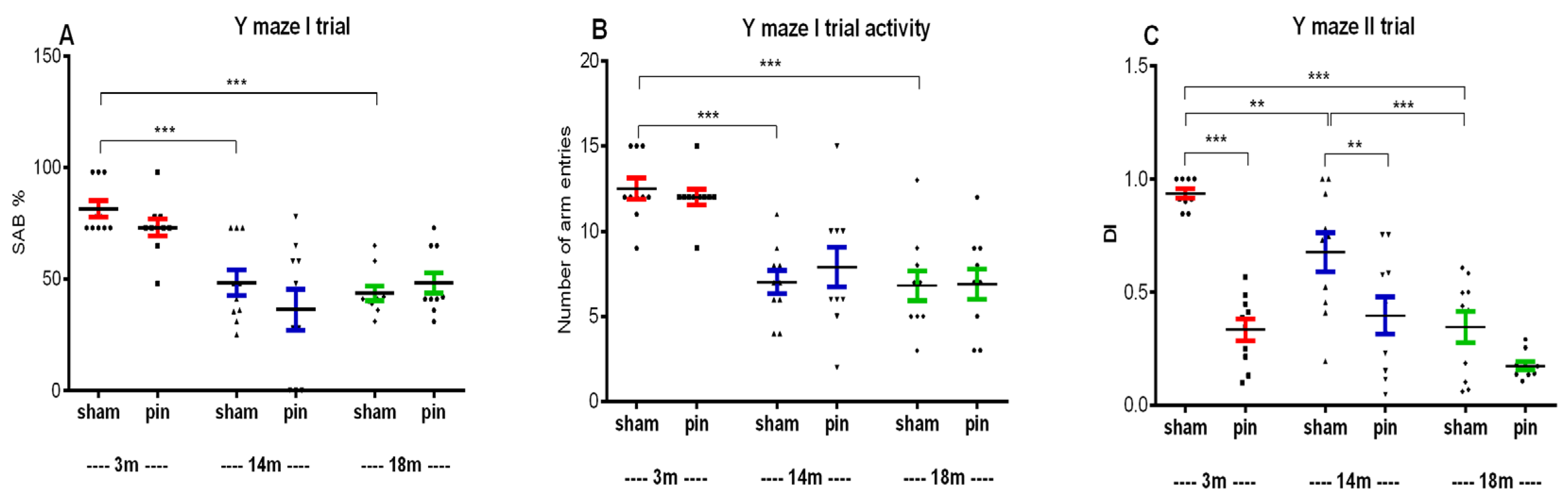
Figure 3.
Effect of pinealectomy on short-term recognition memory in 3-, 14-, and 18-month-old rats measured in the object recognition test (ORT). The DI was assessed in the test phase. Data are presented as mean ± SEM, n = 10. Two-way ANOVA: Age effect: [F2,59 = 3.318, p = 0.044] and Surgery effect: [F1,59 = 8.879, p = 0.004]. *p = 0.012, 3-month-old sham vs. 18-month-old sham rats; ***p < 0.001, 3-month-old pin rats vs. matched controls.
Figure 3.
Effect of pinealectomy on short-term recognition memory in 3-, 14-, and 18-month-old rats measured in the object recognition test (ORT). The DI was assessed in the test phase. Data are presented as mean ± SEM, n = 10. Two-way ANOVA: Age effect: [F2,59 = 3.318, p = 0.044] and Surgery effect: [F1,59 = 8.879, p = 0.004]. *p = 0.012, 3-month-old sham vs. 18-month-old sham rats; ***p < 0.001, 3-month-old pin rats vs. matched controls.
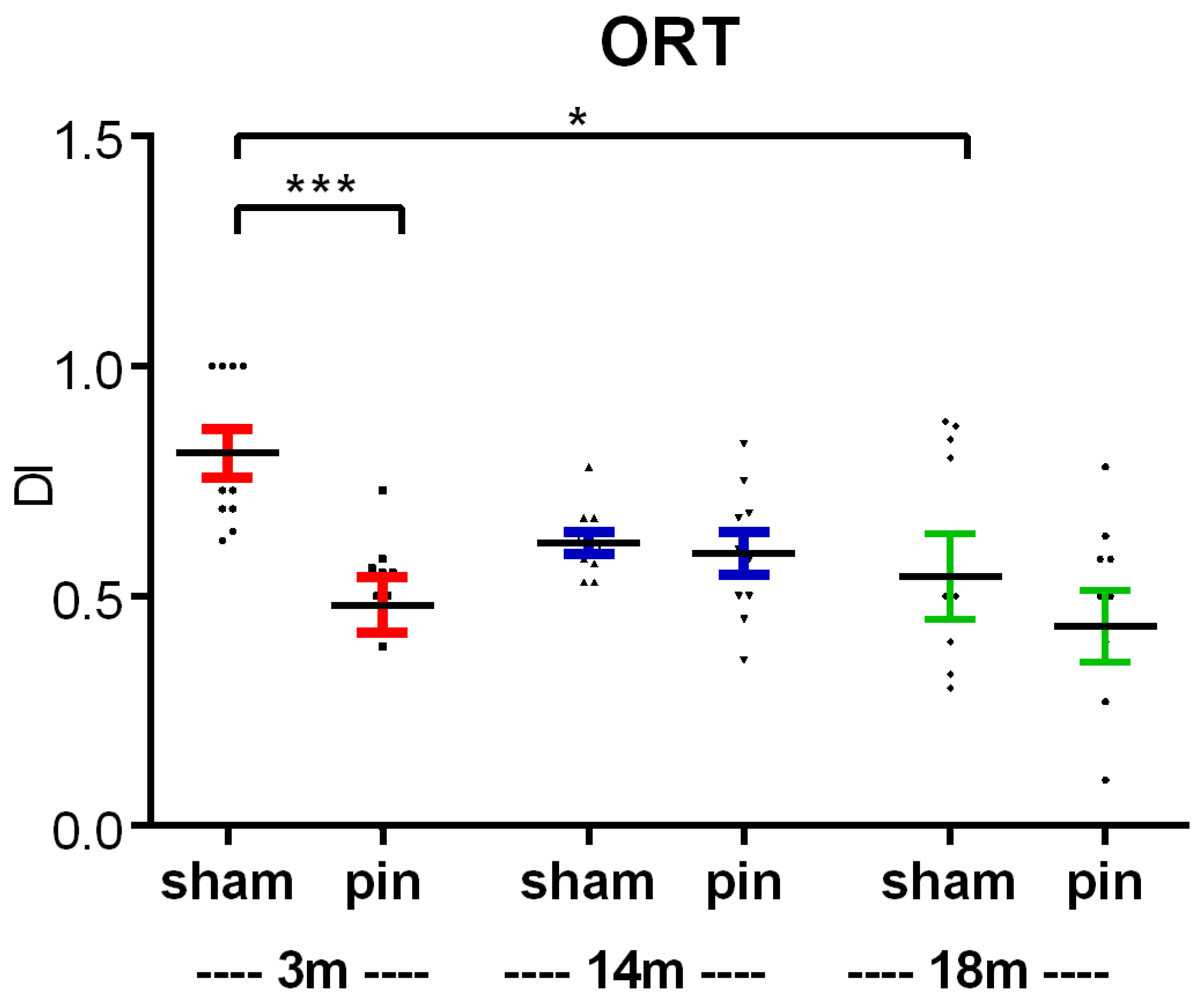
Figure 4.
Effect of pinealectomy on hippocampus-dependent spatial memory in 3-, 14-, and 18-month-old rats measured in the radial arm maze (RAM) test. The repeated two-way ANOVA revealed a significant Group effect: [F5,239 = 7.328, p < 0.001], Time effect [F4,239 = 3.492, p = 0.020] and Group x Time interaction [F20,239 = 1.649, p = 0.049] (A). A main Group effect [F5,239 = 4.534, p = 0.003] was demonstrated for the double working memory errors (DWMEs) (B). Two-way ANOVA demonstrated a main Age effect: [F2,59 = 63,684, p < 0.001] and Surgery effect: [F1,59 = 33,179, p < 0.001] (C); Age effect: [F2,59 = 35,568, p < 0.001], Surgery effect: [F1,59 = 54,402, p < 0.001] and Age x Surgery interaction: [F2,59 = 9,441, p < 0.001] (D); ***p < 0.001, 3-month-old sham vs. 14- and 18-month-old sham rats, ***p < 0.001, 14-month-old pin rats vs. matched control (C); ***p < 0.001, 3-month-old sham vs. 18-month-old sham rats, ***p < 0.001, 14-month-old pin rats vs. matched controls (D).
Figure 4.
Effect of pinealectomy on hippocampus-dependent spatial memory in 3-, 14-, and 18-month-old rats measured in the radial arm maze (RAM) test. The repeated two-way ANOVA revealed a significant Group effect: [F5,239 = 7.328, p < 0.001], Time effect [F4,239 = 3.492, p = 0.020] and Group x Time interaction [F20,239 = 1.649, p = 0.049] (A). A main Group effect [F5,239 = 4.534, p = 0.003] was demonstrated for the double working memory errors (DWMEs) (B). Two-way ANOVA demonstrated a main Age effect: [F2,59 = 63,684, p < 0.001] and Surgery effect: [F1,59 = 33,179, p < 0.001] (C); Age effect: [F2,59 = 35,568, p < 0.001], Surgery effect: [F1,59 = 54,402, p < 0.001] and Age x Surgery interaction: [F2,59 = 9,441, p < 0.001] (D); ***p < 0.001, 3-month-old sham vs. 14- and 18-month-old sham rats, ***p < 0.001, 14-month-old pin rats vs. matched control (C); ***p < 0.001, 3-month-old sham vs. 18-month-old sham rats, ***p < 0.001, 14-month-old pin rats vs. matched controls (D).
Figure 5.
Immunohistochemical localization of BDNF in the dorsal hippocampus of 3-, 14- and 18 -month- old rats. BDNF expression were observed in the hippocampal formation, including in the Dentate Gyrus, granular cell layer (GrDG) and hippocampal cornu ammonis (CA3, CA2 and CA1) fields. Representative images showing BDNF immunostaining in 3-month-old sham rats (A1-A5), 3-month-old pin rats (B1-B5), 14-month-old rats sham rats (C1-C5), 14-month-old pin rats (D1-D5), 18-month-old sham rats (E1-E5) and 18-month-old pin rats (F1-F5). Higher magnifications of the rectangles in all three age groups are shown. Note that the immune response to BDNF in CA3 hippocampal fields is strongly reduced in young adult and middle-aged rats with removed pineal glands (B3,D3). Scale bars: 500 µm (A1, B1, C1, D1, E1, F1), 50 µm (A2-A5; B2-B5; C2-C5; D2-D5; E2-E5; F2-F5).
Figure 5.
Immunohistochemical localization of BDNF in the dorsal hippocampus of 3-, 14- and 18 -month- old rats. BDNF expression were observed in the hippocampal formation, including in the Dentate Gyrus, granular cell layer (GrDG) and hippocampal cornu ammonis (CA3, CA2 and CA1) fields. Representative images showing BDNF immunostaining in 3-month-old sham rats (A1-A5), 3-month-old pin rats (B1-B5), 14-month-old rats sham rats (C1-C5), 14-month-old pin rats (D1-D5), 18-month-old sham rats (E1-E5) and 18-month-old pin rats (F1-F5). Higher magnifications of the rectangles in all three age groups are shown. Note that the immune response to BDNF in CA3 hippocampal fields is strongly reduced in young adult and middle-aged rats with removed pineal glands (B3,D3). Scale bars: 500 µm (A1, B1, C1, D1, E1, F1), 50 µm (A2-A5; B2-B5; C2-C5; D2-D5; E2-E5; F2-F5).
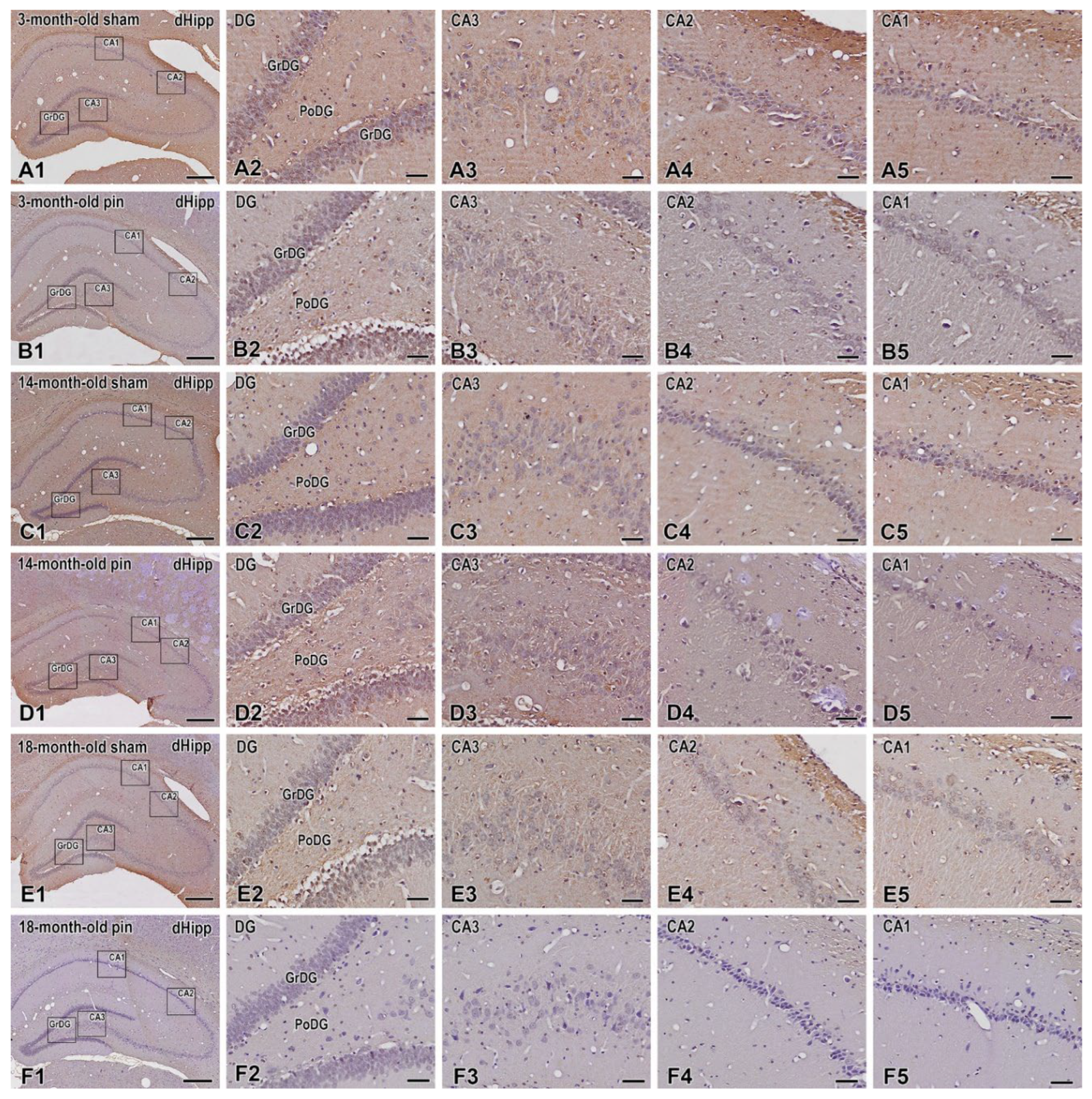
Figure 6.
Effect of pinealectomy on BDNF immunoexpression in the dorsal hippocampus including in the Dentate Gyrus, granular cell layer (GrDG) and hippocampal cornu ammonis (CA3, CA2 and CA1) fields of 3-, 14- and 18 -month- old rats. The data are presented as mean ± SEM, with n = 5 rats per group. ***p < 0.001, 3-month-old rats vs. 18-month-old rats; **p = 0.01, 14-month-old rats vs. 18-month-old rats (A); ***p < 0.001, 3-month-old sham rats vs. 18-month-old sham rats; **p = 0.002, 14-month-old sham rats vs. 18-month-old sham rats; ***p < 0.001, 3-month-old pin rats vs. sham rats; **p = 0.002, 14-month-old pin rats vs. sham rats (B); *p= 0.034, ***p < 0.001, 3-month-old sham rats vs. 14- and 18-month-old sham rats, respectively, (C); ***p < 0.001, 3-month-old sham rats vs. 18-month-old sham rats (D).
Figure 6.
Effect of pinealectomy on BDNF immunoexpression in the dorsal hippocampus including in the Dentate Gyrus, granular cell layer (GrDG) and hippocampal cornu ammonis (CA3, CA2 and CA1) fields of 3-, 14- and 18 -month- old rats. The data are presented as mean ± SEM, with n = 5 rats per group. ***p < 0.001, 3-month-old rats vs. 18-month-old rats; **p = 0.01, 14-month-old rats vs. 18-month-old rats (A); ***p < 0.001, 3-month-old sham rats vs. 18-month-old sham rats; **p = 0.002, 14-month-old sham rats vs. 18-month-old sham rats; ***p < 0.001, 3-month-old pin rats vs. sham rats; **p = 0.002, 14-month-old pin rats vs. sham rats (B); *p= 0.034, ***p < 0.001, 3-month-old sham rats vs. 14- and 18-month-old sham rats, respectively, (C); ***p < 0.001, 3-month-old sham rats vs. 18-month-old sham rats (D).
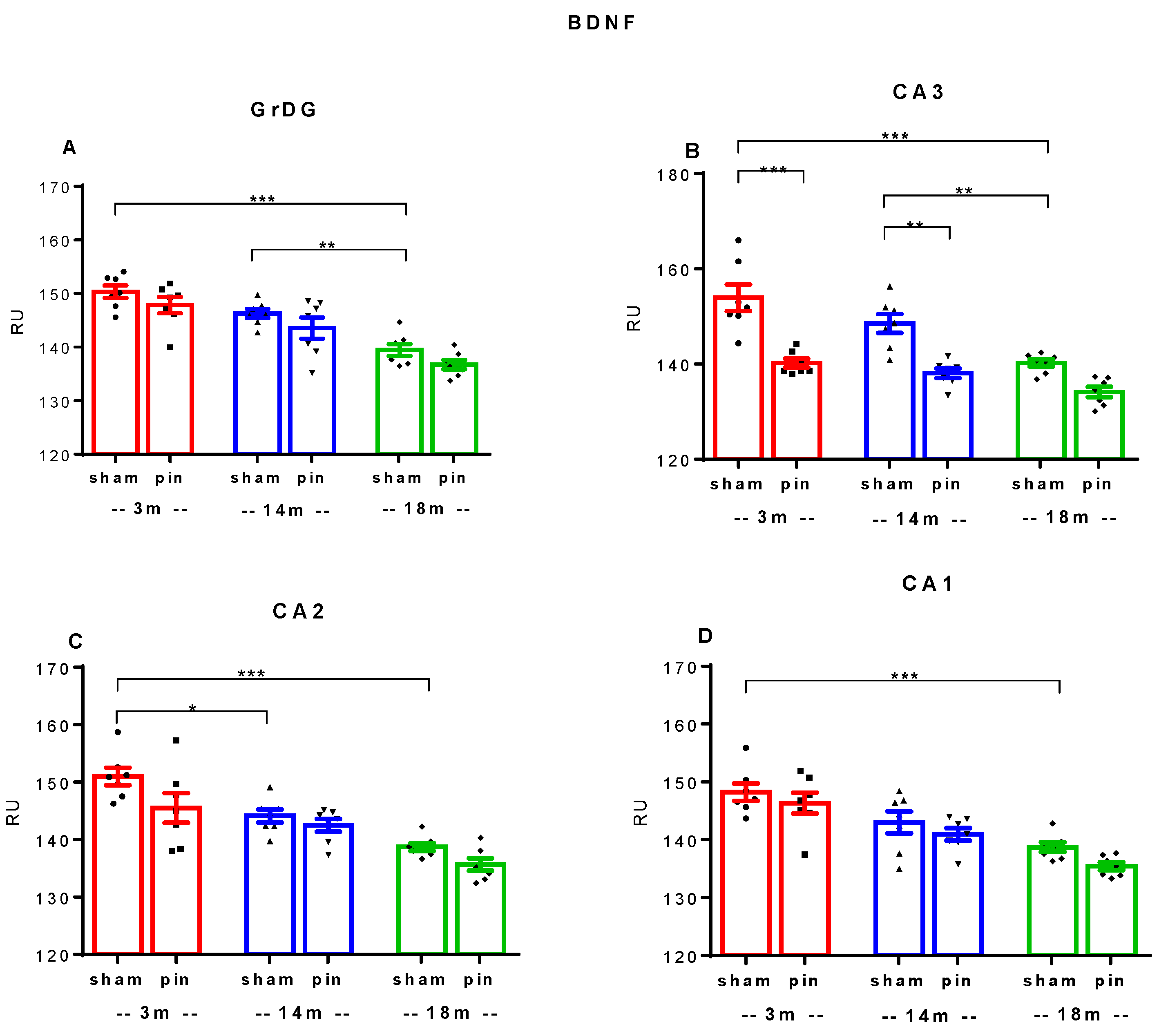
Figure 7.
TrkB immunoexpression in the dorsal hippocampus including in the dentate gyrus, granular cell layer (GrDG) and hippocampal cornu ammonis (CA3, CA2 and CA1) fields of 3-, 14- and 18 -month- old rats. Immunohistochemical localization of TrkB in 3-month-old sham rats (A1-A5), 3-month-old pin rats (B1-B5), 14-month-old rats sham rats (C1-C5), 14-month-old pin rats (D1-D5), 18-month-old sham rats (E1-E5) and 18-month-old pin rats (F1-F5). Higher magnifications of the rectangles in all three age groups are shown. Note the reduced immune reactivity for TrkB in the CA3 region in the young adult rats after pinealectomy (B3). Scale bars: 500 µm (A1, B1, C1, D1, E1, F1), 50 µm (A2-A5; B2-B5; C2-C5; D2-D5; E2-E5; F2-F5).
Figure 7.
TrkB immunoexpression in the dorsal hippocampus including in the dentate gyrus, granular cell layer (GrDG) and hippocampal cornu ammonis (CA3, CA2 and CA1) fields of 3-, 14- and 18 -month- old rats. Immunohistochemical localization of TrkB in 3-month-old sham rats (A1-A5), 3-month-old pin rats (B1-B5), 14-month-old rats sham rats (C1-C5), 14-month-old pin rats (D1-D5), 18-month-old sham rats (E1-E5) and 18-month-old pin rats (F1-F5). Higher magnifications of the rectangles in all three age groups are shown. Note the reduced immune reactivity for TrkB in the CA3 region in the young adult rats after pinealectomy (B3). Scale bars: 500 µm (A1, B1, C1, D1, E1, F1), 50 µm (A2-A5; B2-B5; C2-C5; D2-D5; E2-E5; F2-F5).
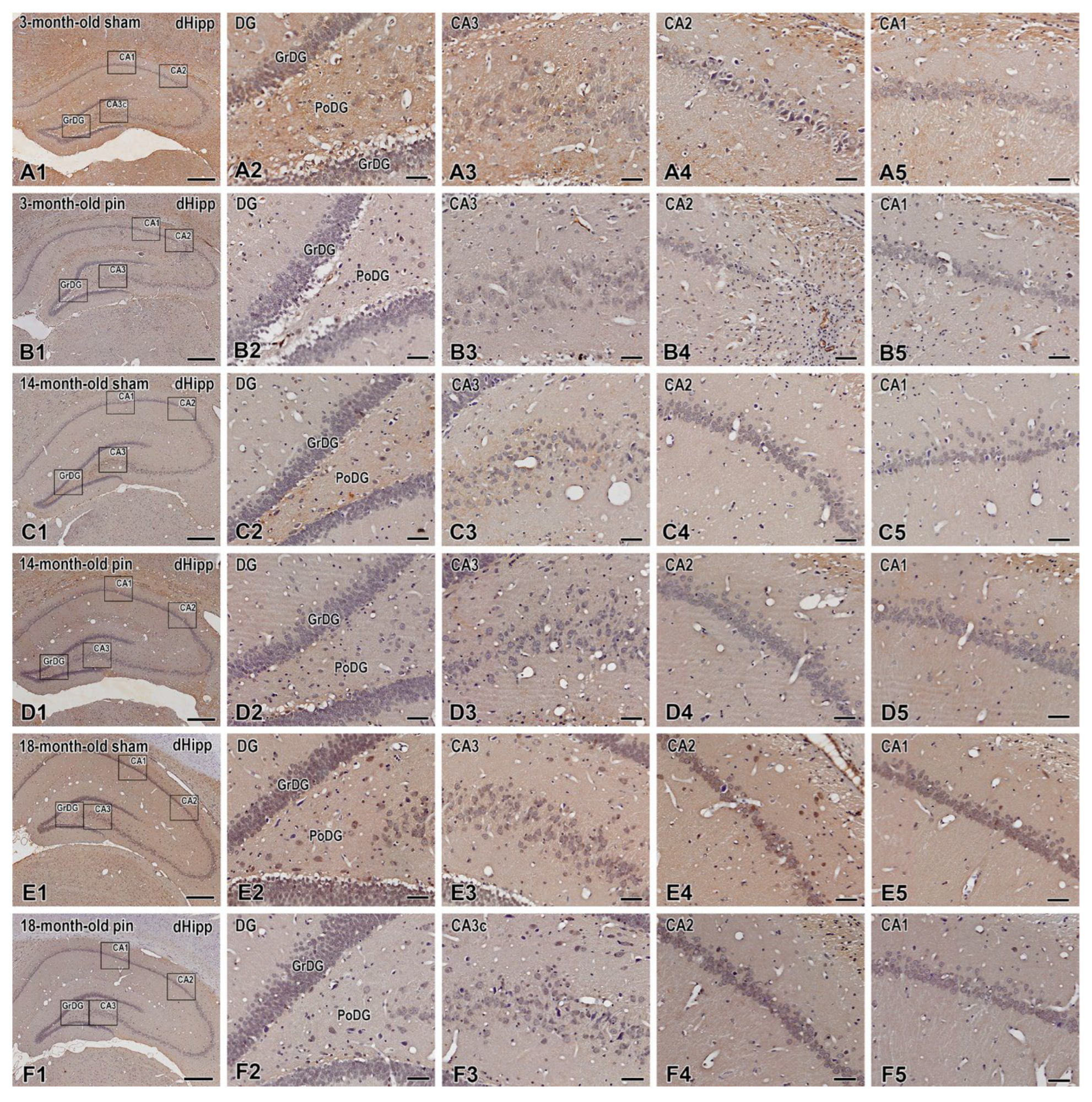
Figure 8.
Effect of pinealectomy on TrkB immunoexpression in the dorsal hippocampus including in the Dentate Gyrus, granular cell layer (GrDG) and hippocampal cornu ammonis (CA3, CA2 and CA1) fields of 3-, 14- and 18 -month- old rats. The data are presented as mean ± SEM, with n = 5 rats per group. In DGgr: A two-way ANOVA indicated a main Age effect: [F1,41 = 39.297, p < 0.001] and Surgery effect: [F2,41 = 132.169, p < 0.001] (A); CA3 region: Age effect: [F1,41 = 16.982, p < 0.001] and Surgery effect: [F2,41 = 32.873, p < 0.001] (B); CA2 region: Age effect: [F1,41 = 4.604, p = 0.039] (C); CA1 region: Age effect: [F1,41 = 9.966, p = 0.003] (D). ***p < 0.001, 3- and 14-month-old sham rats vs. 18-month-old sham rats; ***p < 0.001, 14-month-old pin rats vs. matched rats (A); ***p < 0.001, 3-month-old sham rats vs. 14- and 18-month-old sham rats; **p = 0.002, 3-month-old pin rats vs. matched controls (B); ***p < 0.001, 3-month-old sham rats vs. 18-month-old sham rats (C); ***p < 0.001, 3-month-old sham rats vs. 14- and 18-month-old sham rats; **p = 0.01, 14-month-old sham rats vs. 18-month-old sham rats (C).
Figure 8.
Effect of pinealectomy on TrkB immunoexpression in the dorsal hippocampus including in the Dentate Gyrus, granular cell layer (GrDG) and hippocampal cornu ammonis (CA3, CA2 and CA1) fields of 3-, 14- and 18 -month- old rats. The data are presented as mean ± SEM, with n = 5 rats per group. In DGgr: A two-way ANOVA indicated a main Age effect: [F1,41 = 39.297, p < 0.001] and Surgery effect: [F2,41 = 132.169, p < 0.001] (A); CA3 region: Age effect: [F1,41 = 16.982, p < 0.001] and Surgery effect: [F2,41 = 32.873, p < 0.001] (B); CA2 region: Age effect: [F1,41 = 4.604, p = 0.039] (C); CA1 region: Age effect: [F1,41 = 9.966, p = 0.003] (D). ***p < 0.001, 3- and 14-month-old sham rats vs. 18-month-old sham rats; ***p < 0.001, 14-month-old pin rats vs. matched rats (A); ***p < 0.001, 3-month-old sham rats vs. 14- and 18-month-old sham rats; **p = 0.002, 3-month-old pin rats vs. matched controls (B); ***p < 0.001, 3-month-old sham rats vs. 18-month-old sham rats (C); ***p < 0.001, 3-month-old sham rats vs. 14- and 18-month-old sham rats; **p = 0.01, 14-month-old sham rats vs. 18-month-old sham rats (C).
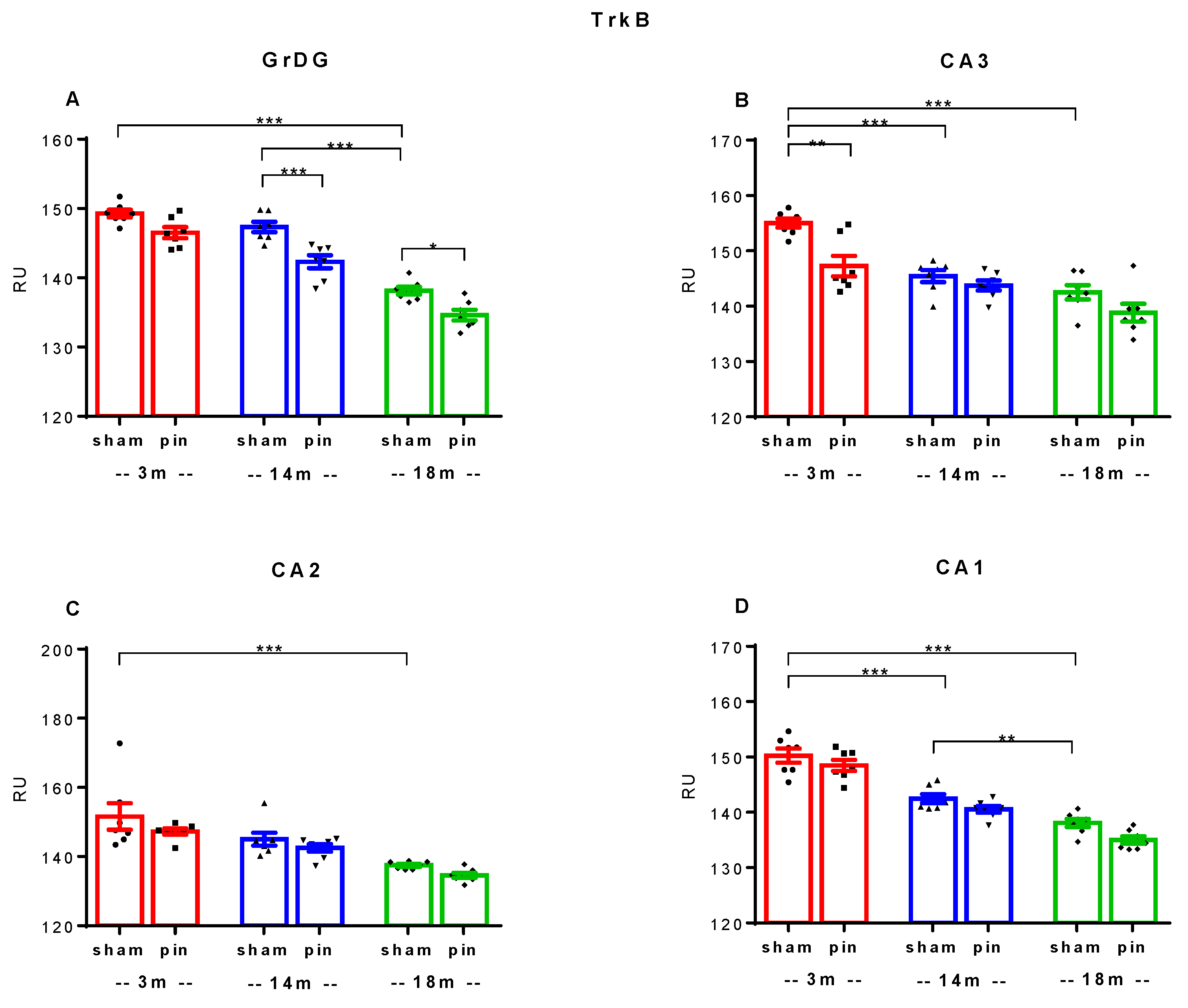
Figure 9.
Double immunofluorescence in CA1 region of dorsal hippocampus for pERK1/2 and NeuN (A1-A4), pERK1/2 and GFAP (B1-B4) in 14-month-old sham rats. The third and fourth columns reflect double immunofluorescence in the same age and experimental group of animals in the dentate gyrus between ERK1/2 and NeuN (C1-C4) and between ERK1/2 and GFAP (D1-D4). Scale bars = 50 µm.
Figure 9.
Double immunofluorescence in CA1 region of dorsal hippocampus for pERK1/2 and NeuN (A1-A4), pERK1/2 and GFAP (B1-B4) in 14-month-old sham rats. The third and fourth columns reflect double immunofluorescence in the same age and experimental group of animals in the dentate gyrus between ERK1/2 and NeuN (C1-C4) and between ERK1/2 and GFAP (D1-D4). Scale bars = 50 µm.
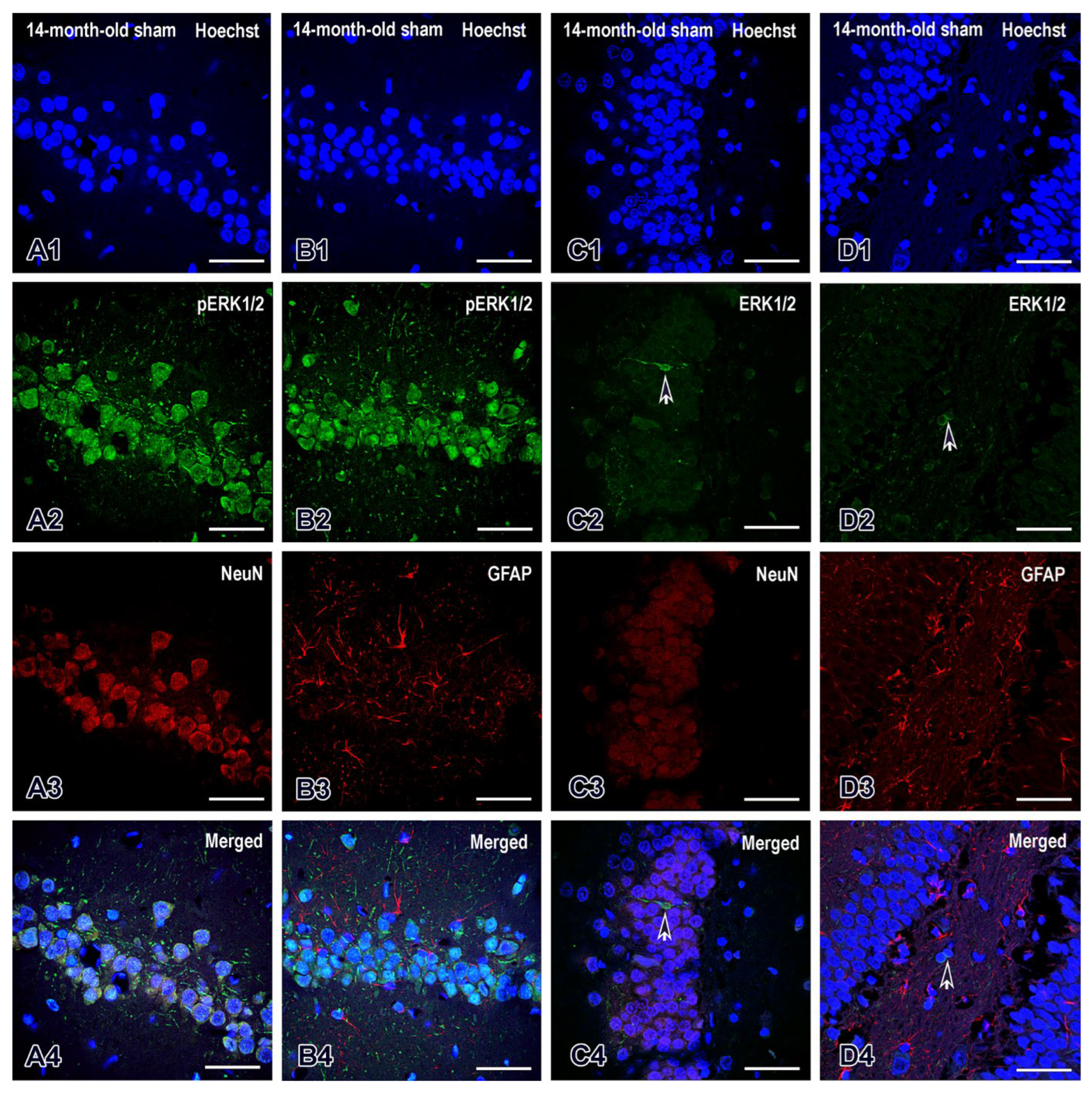
Figure 10.
Immunohistochemical localization of ERK1/2 in the dorsal hippocampus (dHipp) of 3, 14- and 18-month-old rats. Low-power photomicrographs showing the ERK1/2 protein expression in the dentate gyrus (DG) and the CA1-CA3 fields of the hippocampus proper in 3-month-old sham (A1) and 3-month-old pin (B1), 14-month-old sham (C1) and 14-month-old pin (D1) rats, and 18-month-old sham (E1) and 18-month-old pin (F1) rats. Higher magnification of the rectangles in (A1-F1) shows an apparent decrease in ERK1/2 immunoreactivity in the granular layer of the dentate gyrus (GrDG) in 3-month-old pin rats (B2) and in certain layers of the CA1 (B5) and CA3c (B3) of 3-month-old pin rats. Note the increased ERK1/2 expression in GrDG (D2), CA3c stratum pyramidale (Py) of 14-month-old pin rats (D3). Increased ERK1/2 expression in middle-aged pin rats is clearly seen in the GrDG (D2) and CA3c Py (D3) at a higher magnification. Note also the decreased ERK1/2 expression in both the CA2 Py (E4) and CA1 Py (E5) of 18-month-old sham rats. Scale bars: 500 µm (A1, B1, C1, D1, E1, F1), 50 µm (A2-A5; B2-B5; C2-C5; D2-D5; E2-E5; F2-F5).
Figure 10.
Immunohistochemical localization of ERK1/2 in the dorsal hippocampus (dHipp) of 3, 14- and 18-month-old rats. Low-power photomicrographs showing the ERK1/2 protein expression in the dentate gyrus (DG) and the CA1-CA3 fields of the hippocampus proper in 3-month-old sham (A1) and 3-month-old pin (B1), 14-month-old sham (C1) and 14-month-old pin (D1) rats, and 18-month-old sham (E1) and 18-month-old pin (F1) rats. Higher magnification of the rectangles in (A1-F1) shows an apparent decrease in ERK1/2 immunoreactivity in the granular layer of the dentate gyrus (GrDG) in 3-month-old pin rats (B2) and in certain layers of the CA1 (B5) and CA3c (B3) of 3-month-old pin rats. Note the increased ERK1/2 expression in GrDG (D2), CA3c stratum pyramidale (Py) of 14-month-old pin rats (D3). Increased ERK1/2 expression in middle-aged pin rats is clearly seen in the GrDG (D2) and CA3c Py (D3) at a higher magnification. Note also the decreased ERK1/2 expression in both the CA2 Py (E4) and CA1 Py (E5) of 18-month-old sham rats. Scale bars: 500 µm (A1, B1, C1, D1, E1, F1), 50 µm (A2-A5; B2-B5; C2-C5; D2-D5; E2-E5; F2-F5).
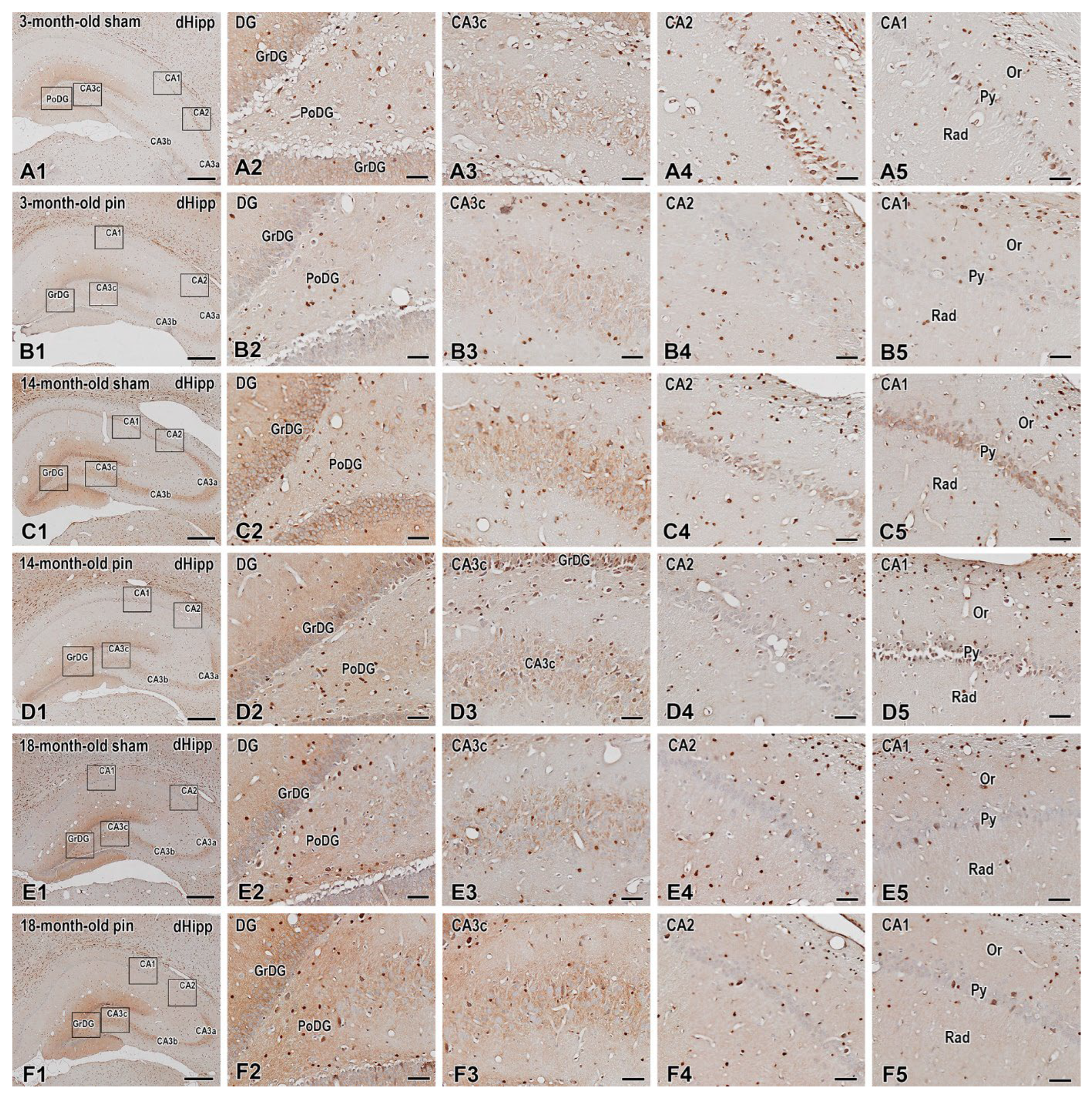
Figure 11.
Effect of pinealectomy on Erk 1/2 immunoexpression in the dorsal hippocampus, including in the DG, CA3c, CA3b, CA3a, CA2 and CA1 region. Data are presented as mean ± SEM, n = 5 rats per group. GrDG: ***p<0.001, 3-month-old sham rats vs. 14- and 18-month-old sham rats, ***p<0.001, 3-month-old pin rats vs. 3-month-old sham rats (A); CA3c: ***p<0.001, 3-month-old pin rats vs. 3-month-old sham rats, ***p<0.001, 14-month-old pin rats vs. 14-month-old sham rats in stratum oriens (Or); ***p<0.001, 3-month-old sham rats vs. 14- and 18-month-old sham rats, ***p<0.001, 3-month-old pin rats vs. 3-month-old sham rats, ***p<0.001, 14-month-old pin rats vs. 14-month-old sham rats in stratum pyramidale (Py); **p=0.002, 3-month-old sham rats vs. 18-month-old sham rats, ***p<0.001, 14-month-old sham rats vs. 18-month-old sham rats in in stratum lucidum (SLu); **p = 0.002, 3-month-old sham rats vs. 18-month-old sham rats, **p=0.003, 14-month-old sham vs. 18-month-old sham in stratum radiatum (Rad) (B). CA3b: ***p<0.001, 14-month-old pin rats vs. 14-month-old sham rats in Or; ***p<0.001, 14-month-old pin rats vs. 14-month-old sham rats in Py (C); **p= 0.006, 3-month-old sham rats vs. 18-month-old sham rats, ***p<0.001, 3-month-old pin rats vs. 3-month-old sham rats in Or; ***p<0.001, 3-month-old sham rats vs. 18-month-old sham rats; ***p<0.001, 14-month-old sham rats vs. 18-month-old sham rats, ***p<0.001, 3-month-old pin rats vs. 3-month-old sham erats in Py (D); CA2: *p= 0.044, 3-month-old pin rats vs. 3-month-old sham rats in Or; ***p<0.001, 3-month-old sham rats vs. 18-month-old sham rats, ***p<0.001, 3-month-old pin rats vs. 3-month-old sham rats in Pyr (E); ***p<0.001, 3-month-old sham rats vs. 14- and 18-month-old sham rats, ***p<0.001, 3-month-old pin rats vs. 3-month-old sham rats in Py; **p=0.007, 3-month-old pin rats vs. 3-month-old sham rats in Rad (F).
Figure 11.
Effect of pinealectomy on Erk 1/2 immunoexpression in the dorsal hippocampus, including in the DG, CA3c, CA3b, CA3a, CA2 and CA1 region. Data are presented as mean ± SEM, n = 5 rats per group. GrDG: ***p<0.001, 3-month-old sham rats vs. 14- and 18-month-old sham rats, ***p<0.001, 3-month-old pin rats vs. 3-month-old sham rats (A); CA3c: ***p<0.001, 3-month-old pin rats vs. 3-month-old sham rats, ***p<0.001, 14-month-old pin rats vs. 14-month-old sham rats in stratum oriens (Or); ***p<0.001, 3-month-old sham rats vs. 14- and 18-month-old sham rats, ***p<0.001, 3-month-old pin rats vs. 3-month-old sham rats, ***p<0.001, 14-month-old pin rats vs. 14-month-old sham rats in stratum pyramidale (Py); **p=0.002, 3-month-old sham rats vs. 18-month-old sham rats, ***p<0.001, 14-month-old sham rats vs. 18-month-old sham rats in in stratum lucidum (SLu); **p = 0.002, 3-month-old sham rats vs. 18-month-old sham rats, **p=0.003, 14-month-old sham vs. 18-month-old sham in stratum radiatum (Rad) (B). CA3b: ***p<0.001, 14-month-old pin rats vs. 14-month-old sham rats in Or; ***p<0.001, 14-month-old pin rats vs. 14-month-old sham rats in Py (C); **p= 0.006, 3-month-old sham rats vs. 18-month-old sham rats, ***p<0.001, 3-month-old pin rats vs. 3-month-old sham rats in Or; ***p<0.001, 3-month-old sham rats vs. 18-month-old sham rats; ***p<0.001, 14-month-old sham rats vs. 18-month-old sham rats, ***p<0.001, 3-month-old pin rats vs. 3-month-old sham erats in Py (D); CA2: *p= 0.044, 3-month-old pin rats vs. 3-month-old sham rats in Or; ***p<0.001, 3-month-old sham rats vs. 18-month-old sham rats, ***p<0.001, 3-month-old pin rats vs. 3-month-old sham rats in Pyr (E); ***p<0.001, 3-month-old sham rats vs. 14- and 18-month-old sham rats, ***p<0.001, 3-month-old pin rats vs. 3-month-old sham rats in Py; **p=0.007, 3-month-old pin rats vs. 3-month-old sham rats in Rad (F).
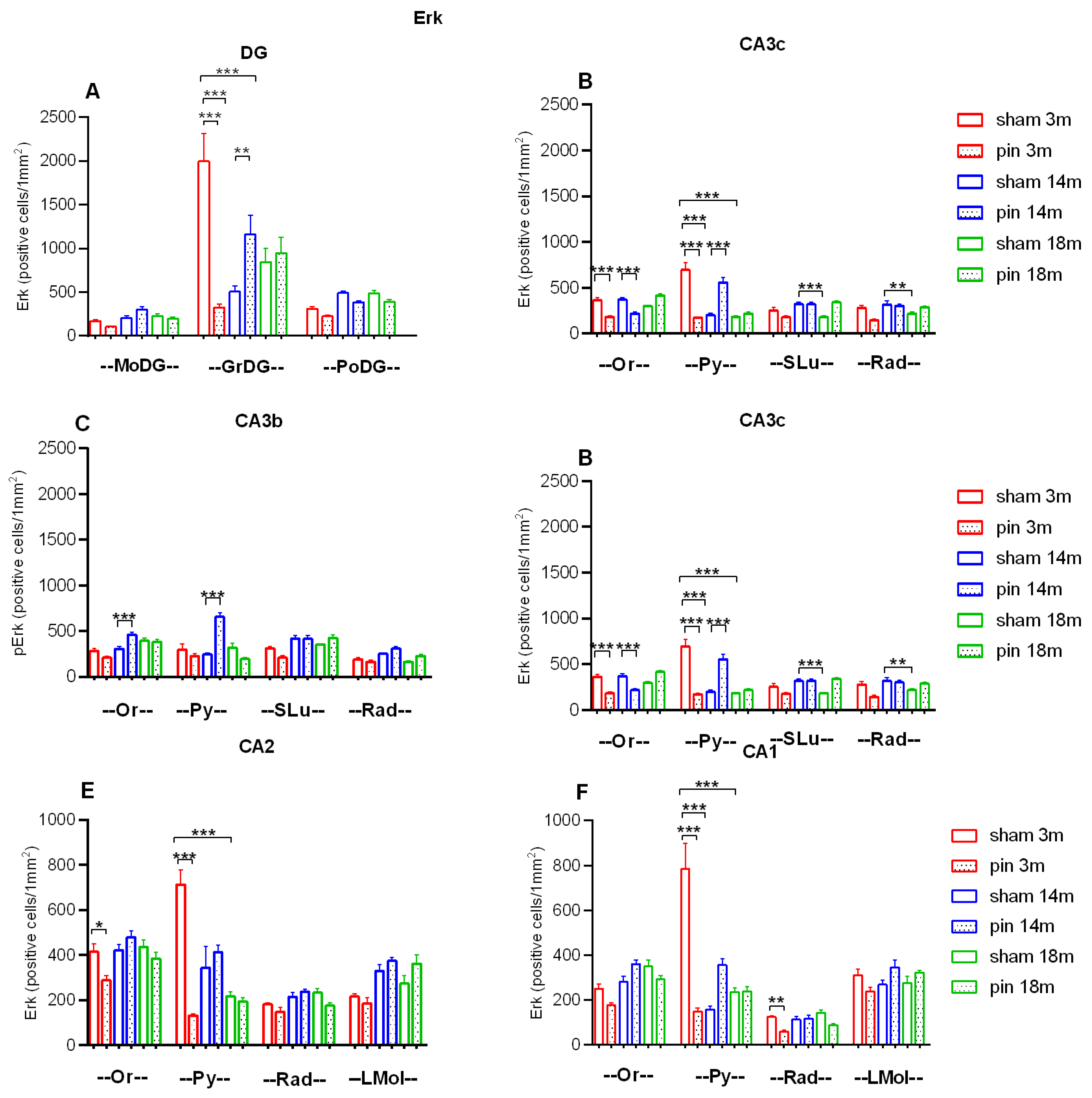
Figure 12.
Immunohistochemical demonstration of pERK1/2 in the dorsal hippocampus (dHipp) of 3-, 14- and 18-month-old rats. Low-power view of the hippocampus in 3-month-old sham (A1) and 3-month-old pin (B1) rats, 14-month-old sham (C1) and 14-month-old pin (D1) rats, and 18-month-old sham (E1) and 18-month-old pin (F1) rats. Insets show at a higher magnification the reduced pERK1/2 expression in the granular layer of the dentate gyrus (GrDG) and the three parts of the CA hippocampal region (E2-E5) in 18-month-old sham rats. Higher magnifications of the boxed areas in (A1-F1) also show the decreased pERK1/2 immunoreactivity in all areas of the hippocampal formation in 3-month-old pin rats (B2-B5). Scale bars = 500 µm (A1, B1, C1, D1, E1, F1); 50 µm (A2-A5, B2-B5, C2-C5, D2-D5, E2-E5, F2-F5).
Figure 12.
Immunohistochemical demonstration of pERK1/2 in the dorsal hippocampus (dHipp) of 3-, 14- and 18-month-old rats. Low-power view of the hippocampus in 3-month-old sham (A1) and 3-month-old pin (B1) rats, 14-month-old sham (C1) and 14-month-old pin (D1) rats, and 18-month-old sham (E1) and 18-month-old pin (F1) rats. Insets show at a higher magnification the reduced pERK1/2 expression in the granular layer of the dentate gyrus (GrDG) and the three parts of the CA hippocampal region (E2-E5) in 18-month-old sham rats. Higher magnifications of the boxed areas in (A1-F1) also show the decreased pERK1/2 immunoreactivity in all areas of the hippocampal formation in 3-month-old pin rats (B2-B5). Scale bars = 500 µm (A1, B1, C1, D1, E1, F1); 50 µm (A2-A5, B2-B5, C2-C5, D2-D5, E2-E5, F2-F5).
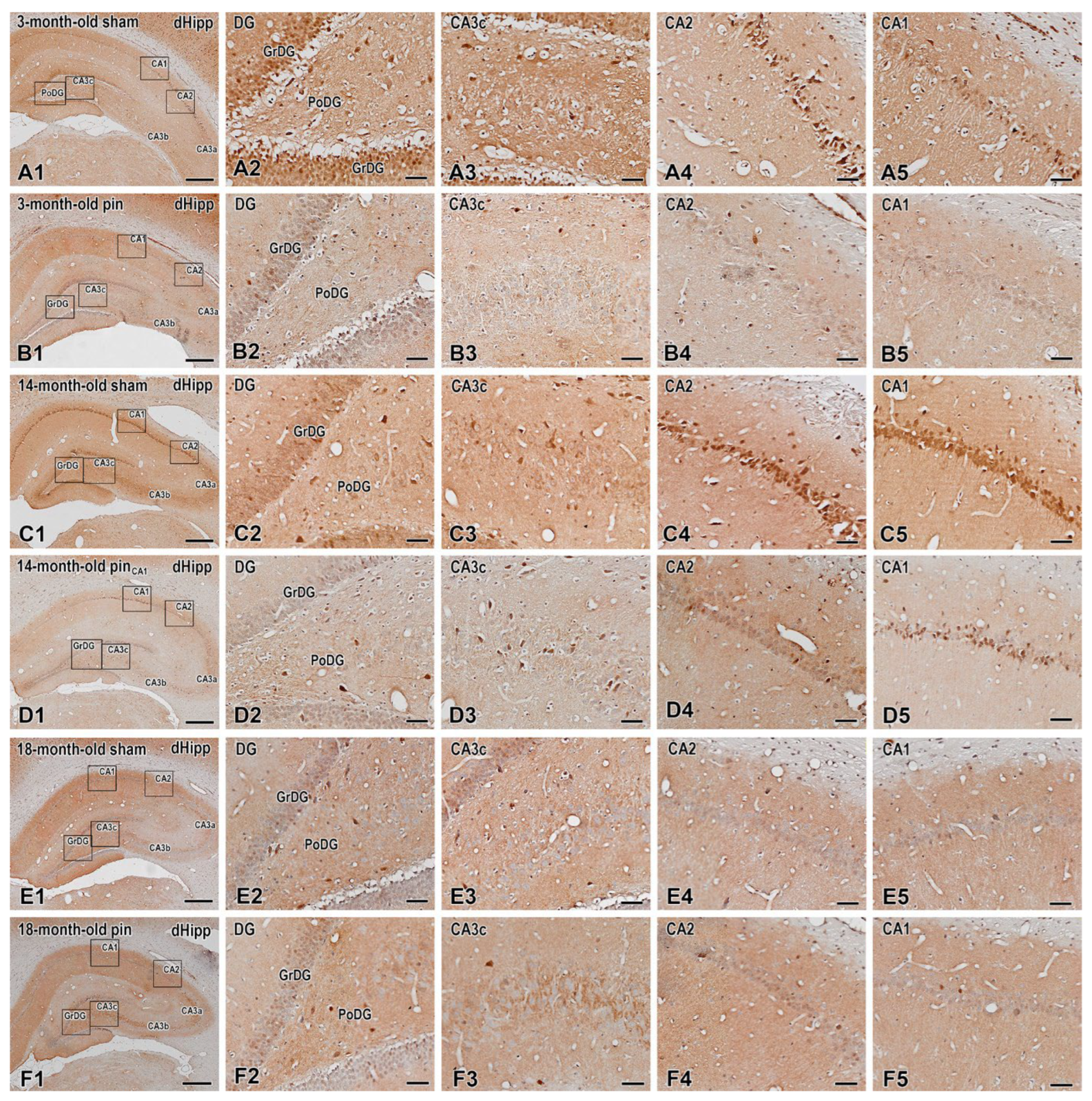
Figure 13.
Effect of pinealectomy on pErk 1/2 immunoexpression in the dorsal hippocampus, including in the GrDG, PoDG, CA3c, CA3b, CA3a, CA2 and CA1 region. Data are presented as mean ± SEM, n = 5 rats per group. **p = 0.017, 3-month-old sham rats vs. 14-month-old sham rats; ***p < 0.001, 3-month-old sham rats vs. 18-month-old sham rats; ***p < 0.001, 3-month-old pin rats vs. their matched rats; ***p<0.001, 14-month-old pin rats vs. their matched rats (A). ***p < 0.001, 3-month-old sham rats vs. 14- and 18-month-old sham rats; ***p < 0.001, 3-month-old pin groups vs. their matched group (B); ***p < 0.001, 3-month-old sham rats vs. 14- and 18-month-old sham rats; ***p < 0.001, 14-month-old rats vs. 18-month-old rats; ***p < 0.001, 3-month-old pin rats vs. their matched rats; *p = 0.045, 14-month-old pin rats vs. their matched rats (C); ***p < 0.001, 3-month-old sham rats vs. 14- and 18-month-old sham rats; ***p < 0.001, 3- and 14-month-old pin rats vs. their matched group (D); ***p < 0.001, 3-month-old sham rats vs. 14- and 18-month-old sham rats; ***p < 0.001, 14-month-old sham rats vs. 18-month-old sham rats; ***p < 0.001, 3- and 14-month-old pin rats vs. their matched group (E); ***p < 0.001, 3-month-old sham rats vs. 18-month-old sham rats; ***p < 0.001, 14-month-old sham rats vs. 18-month-old sham rats; ***p<0.001, 3-month-old pin rats vs. their matched rats; ***p<0.001, 14-month-old pin rats vs. their matched rats (F); ***p < 0.001, 3-month-old sham rats vs. 18-month-old sham rats; ***p < 0.001, 14-month-old sham rats vs. 18-month-old sham rats; ***p < 0.001, 3-month-old pin rats vs. their matched rats (G).
Figure 13.
Effect of pinealectomy on pErk 1/2 immunoexpression in the dorsal hippocampus, including in the GrDG, PoDG, CA3c, CA3b, CA3a, CA2 and CA1 region. Data are presented as mean ± SEM, n = 5 rats per group. **p = 0.017, 3-month-old sham rats vs. 14-month-old sham rats; ***p < 0.001, 3-month-old sham rats vs. 18-month-old sham rats; ***p < 0.001, 3-month-old pin rats vs. their matched rats; ***p<0.001, 14-month-old pin rats vs. their matched rats (A). ***p < 0.001, 3-month-old sham rats vs. 14- and 18-month-old sham rats; ***p < 0.001, 3-month-old pin groups vs. their matched group (B); ***p < 0.001, 3-month-old sham rats vs. 14- and 18-month-old sham rats; ***p < 0.001, 14-month-old rats vs. 18-month-old rats; ***p < 0.001, 3-month-old pin rats vs. their matched rats; *p = 0.045, 14-month-old pin rats vs. their matched rats (C); ***p < 0.001, 3-month-old sham rats vs. 14- and 18-month-old sham rats; ***p < 0.001, 3- and 14-month-old pin rats vs. their matched group (D); ***p < 0.001, 3-month-old sham rats vs. 14- and 18-month-old sham rats; ***p < 0.001, 14-month-old sham rats vs. 18-month-old sham rats; ***p < 0.001, 3- and 14-month-old pin rats vs. their matched group (E); ***p < 0.001, 3-month-old sham rats vs. 18-month-old sham rats; ***p < 0.001, 14-month-old sham rats vs. 18-month-old sham rats; ***p<0.001, 3-month-old pin rats vs. their matched rats; ***p<0.001, 14-month-old pin rats vs. their matched rats (F); ***p < 0.001, 3-month-old sham rats vs. 18-month-old sham rats; ***p < 0.001, 14-month-old sham rats vs. 18-month-old sham rats; ***p < 0.001, 3-month-old pin rats vs. their matched rats (G).
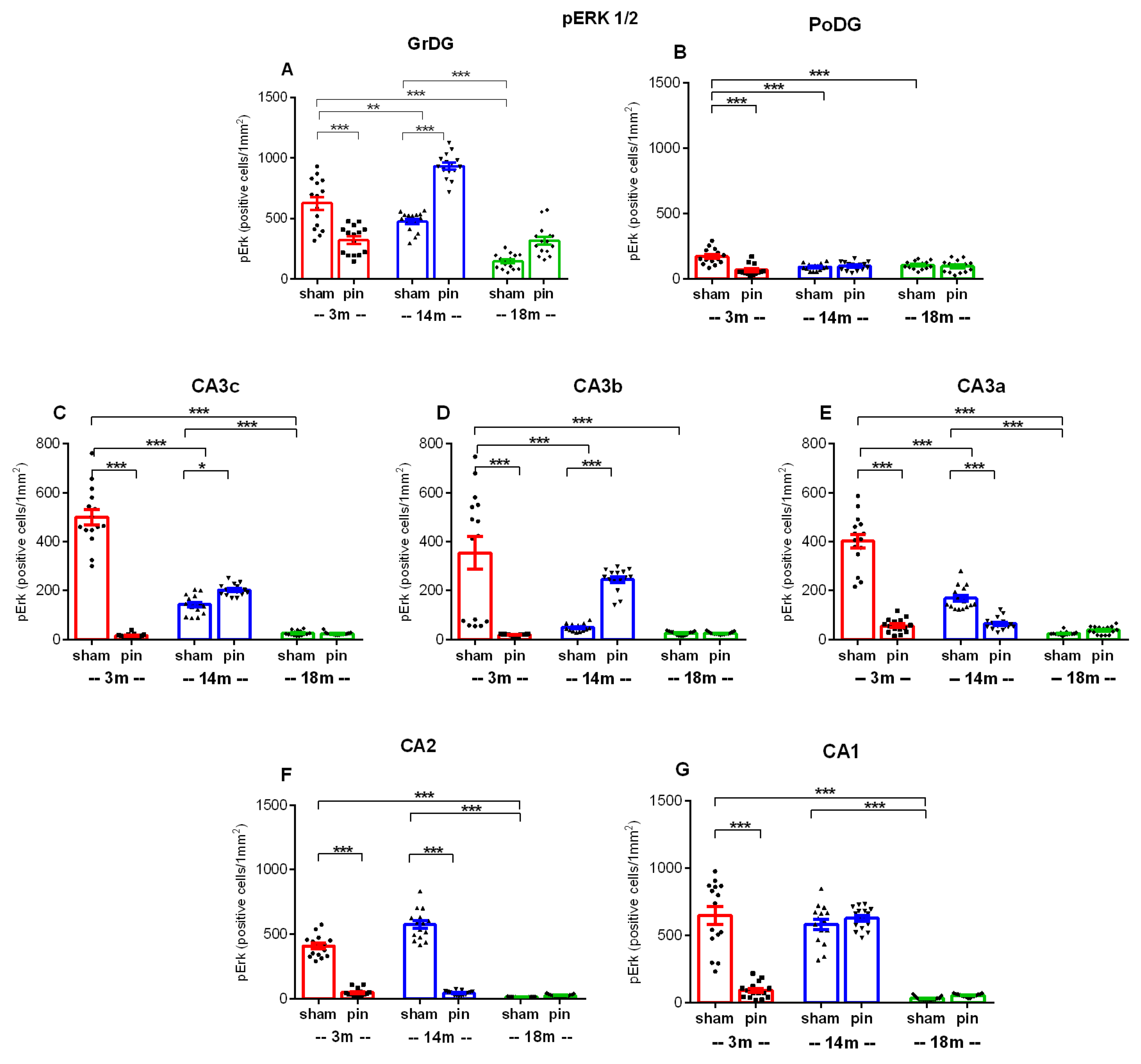
Figure 14.
Effect of pinealectomy on pCREB/CREB ratio in 3-, 14-, and 18-month-old rats measured in the frontal cortex (FC) (A) and the hippocampus (B). Data are presented as mean ± SEM, n = 8. Two-way ANOVA: Age effect: [F1,47 = 16.045, p < 0.001] (A) and [F1,47 = 8.884, p = 0.005](B), respectively; Surgery effect: [F1,47 = 6153, p = 0.005] (A); Age x Surgery interaction: [F2,47 = 6.006, p = 0.005] (B). **p = 0.004 and **p = 0.008, 3- and 14-month-old sham rats vs. 18-month-old sham rats, respectively; **p = 0.002 and **p = 0.007, 3- and 14-month-old pin groups vs. their matched group, respectively (A); *p = 0.018 and ***p < 0.001, 3- and 14-month-old sham rats vs. 18-month-old sham rats, respectively; *p = 0.036 and ***p < 0.001, 3- and 14-month-old pin groups vs. their matched group, respectively (B).
Figure 14.
Effect of pinealectomy on pCREB/CREB ratio in 3-, 14-, and 18-month-old rats measured in the frontal cortex (FC) (A) and the hippocampus (B). Data are presented as mean ± SEM, n = 8. Two-way ANOVA: Age effect: [F1,47 = 16.045, p < 0.001] (A) and [F1,47 = 8.884, p = 0.005](B), respectively; Surgery effect: [F1,47 = 6153, p = 0.005] (A); Age x Surgery interaction: [F2,47 = 6.006, p = 0.005] (B). **p = 0.004 and **p = 0.008, 3- and 14-month-old sham rats vs. 18-month-old sham rats, respectively; **p = 0.002 and **p = 0.007, 3- and 14-month-old pin groups vs. their matched group, respectively (A); *p = 0.018 and ***p < 0.001, 3- and 14-month-old sham rats vs. 18-month-old sham rats, respectively; *p = 0.036 and ***p < 0.001, 3- and 14-month-old pin groups vs. their matched group, respectively (B).
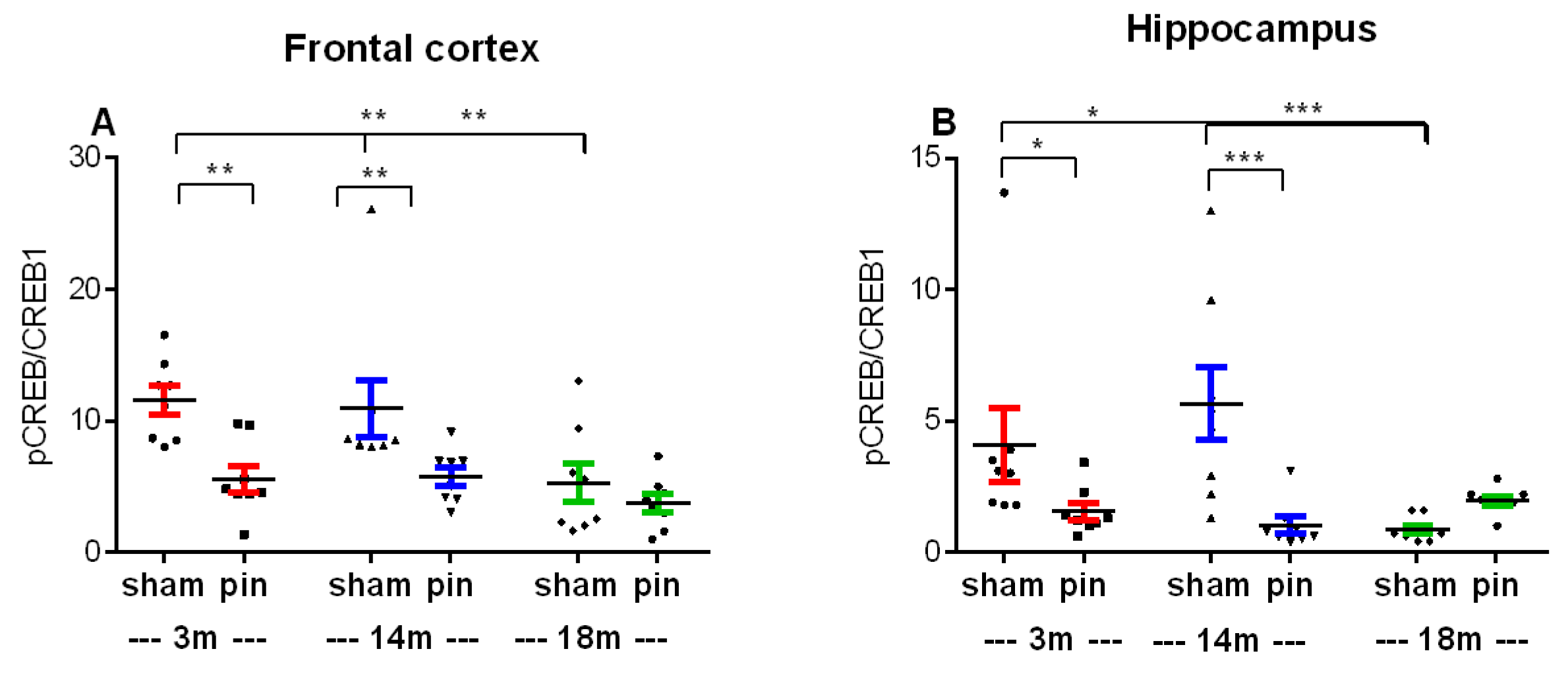
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



