
Submitted:
18 July 2024
Posted:
19 July 2024
You are already at the latest version
Abstract
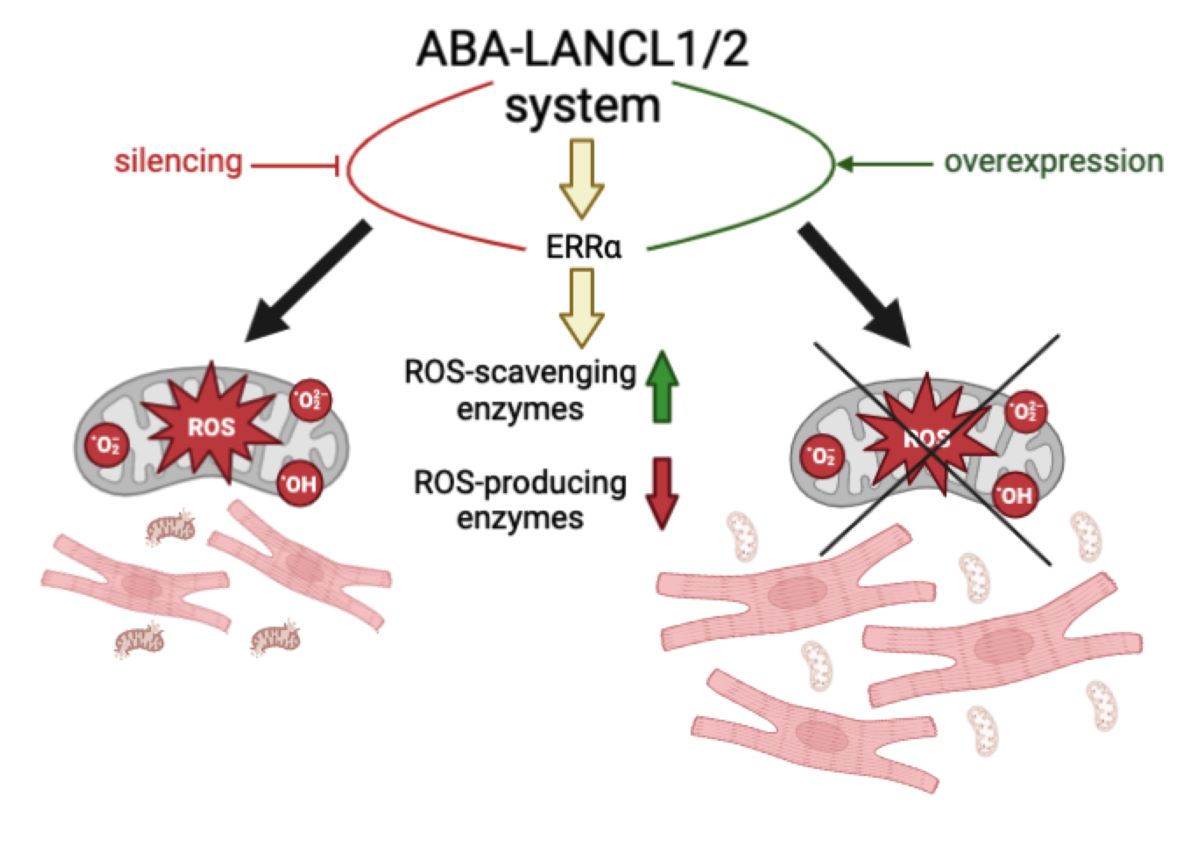
Rat H9c2 cardiomyocytes overexpressing the abscisic acid (ABA) hormone receptors LANCL1 and LANCL2 have an increased mitochondrial proton gradient, respiration and vitality after hypoxia/reoxygenation. Our aim was to investigate the role of the ABA/LANCL1-2 system in ROS turnover in H9c2. H9c2 cells were retrovirally infected to induce overexpression or silencing of LANCL1 and LANCL2, without or with the concomitant silencing of the transcription factor ERRα. Enzymes involved in radical production or scavenging were studied by qPCR and Western blot. Mitochondrial proton gradient and ROS were measured with specific fluorescent probes. ROS generating enzymes decreased, ROS-scavenging enzymes increased, and mitochondrial ROS were reduced in LANCL1/2-overexpressing vs. control cells infected with the empty vector, while the opposite occurred in LANCL1/2-silenced cells. Knock-down of ERRα abrogated all beneficial effects on ROS turnover in LANCL1/2 overexpressing cells. Taken together, these results indicate that the ABA/LANCL1-2 system controls ROS turnover in H9c2 via ERRα. The ABA/LANCL system emerges as a promising target to improve cardiomyocyte mitochondrial function and resilience to oxidative stress.
Keywords:
ROS-producing enzymes
; ROS-scavenging enzymes
; mitochondrial proton gradient
; COX2
; XO
; NOX4
; SOD2
; GPX4
; ERRα
1. Introduction
Abscisic acid (ABA) is an isoprenoid hormone present in unicellular organisms and conserved across kingdoms in modern plants and animals [1]. In mammals, ABA is produced by different cell types and is involved in tissue-specific physiological functions, such as control of blood glucose levels, inflammation, cardiomyocyte energy metabolism, neuroprotection, control of adipocyte browning and energy expenditure, and hemopoietic stem cell regeneration [2,3,4,5,6,7,8].
Two ABA receptors are involved in the metabolic functions of ABA: LANCL1 and LANCL2, belonging to the mammalian LANCL protein family, which comprises three members conserved from bacteria to humans. The ABA-binding capacity of LANCL1 and LANCL2, and their role in mediating the above described functional ABA responses, are well described and documented [9,10]. LANCL1/2 are relatively abundant in all tissues, particularly in the brain and LANCL1 expression in the heart is among the highest in non-neurological tissues [10].
Recently, ABA and its mammalian receptors LANCL1 and LANCL2 have been shown to play an important role in the response of cardiomyocytes to hypoxia [6]. Moreover, in LANCL1/2-overexpressing H9c2 compared with LANCL1/2-silenced cells we observed an improved cell vitality and mitochondrial function after hypoxia/reoxygenation via NO production, with an increased mitochondrial proton gradient (ΔΨ), mitochondrial DNA content and respiration via a signaling pathway involving AMPK and PGC-1α. Overexpression or silencing of LANCL1/2 significantly increased or decreased, respectively, transcription, expression and phosphorylation of AMPK, Akt and eNOS; transcription of NAMPT and Sirt1 [6]. In addition, the NADPH content and the NADPH/NADP ratio were also significantly higher in the overexpressing vs. the silenced H9c2 cells; NADPH is required for NO synthesis and also in the maintenance of the cellular redox balance and antioxidant defense.
In light of this evidence, in this study we studied the role of ABA/LANCL1-2 system in reactive oxygen species (ROS) metabolism in LANCL1/2-overexpressing H9c2 compared with LANCL1/2-silenced rat cardiomyocytes H9c2.
Reactive oxygen species (ROS) such as superoxide anion (O2•-), hydroxyl radical (•OH) and hydrogen peroxide (H2O2) are generated endogenously by processes such as mitochondrial oxidative phosphorylation or by various enzyme systems as Cyclooxygenase-2 (COX2), Xanthine Oxidase (XO) and NADPH oxidase (NOX) or they may derived from exogenous sources like ionizing radiation and bacterial infection [11,12]. ROS can be beneficial as subcellular signaling molecules in gene regulatory and signal transduction pathways but also harmful to cells when ROS levels become unregulated in response to physiological and pathological conditions; in fact free radicals can adversely affect various important classes of biological molecules such as nucleic acids, lipids and proteins, thereby altering the normal redox status leading to increased oxidative stress. The free radicals induced oxidative stress has been reported to be involved in several diseased conditions such as obesity, diabetes mellitus and metabolic disorders, cardiovascular and respiratory diseases, heart failure, neurodegenerative disorders, rheumatoid arthritis and in various cancers [13,14].
To protect themselves from oxidative stress, cells have developed several systems to detoxify ROS. In general, there are two major strategies, which convey partial resistance against oxidative stress to most cell types: non-enzymatic and enzymatic. The first comprises small antioxidant molecules like vitamin E, vitamin C, β-carotene, glutathione (GSH), coenzyme Q and bilirubin, which function to quench ROS. The second consists of ROS-detoxifying enzymes, including superoxide dismutase family (SOD), the seleno-enzyme GSH peroxidase (GPX) and catalase [15].
Transcriptional control of metabolic genes is a crucial mechanism to coordinate ROS homeostasis and, therefore, a better understanding of how ROS metabolism is regulated by specific transcription factors can contribute to uncovering new therapeutic strategies. In particular the estrogen-related receptors (ERRs), transcription factors belonging to the nuclear receptor superfamily are master regulators of cellular energy metabolism and, most recently, are related to ROS metabolism [16,17,18,19]. The ERR subfamily is part of the superfamily of nuclear receptors and comprises three members referred to as ERRα, ERRβ and ERRγ [16]. Most nuclear receptors are ligand-regulated transcription factors that recognize small lipophilic hormones, vitamins and metabolites, and thus provide a direct link between extracellular signals and regulation of gene expression. ERRα, defined as an orphan receptor as its natural activating ligand is not known, is ubiquitously expressed and the most abundantly expressed isoform of its family and it is involved in the regulation of mitochondrial function and thermogenesis [20]. Moreover ERRα regulates the transcription of all enzymes that constitute the TCA cycle and a significant number of target genes that code for proteins involved in oxidative phosphorylation (complex I, II, III, IV, V, coenzyme Q and cytochrome c) [18] and the PGC-1α/ERRα complex regulates transcription of ROS metabolism targeting genes, encoding several enzymes involved in ROS metabolism, such as NOX4, NOX5, XDH, CAT, GPX and SOD2 [16,21,22,23].
We observed an impressive (20-fold) increase of ERRα mRNA levels in LANCL1/2-overexpressing, ABA-treated human differentiated adipocytes, both white and brown, in parallel with an increased mitochondrial function; conversely ERRα expression was significantly decreased in LANCL1-2 silenced white and brown adipocytes, suggesting a correlation between the ABA-LANCL1/2 system and ERRα [7]. Moreover, the ABA/LANCL1-2 system activates the AMPK/PGC-1α axis in skeletal and heart muscle, as well as in adipocytes, increasing expression and also phosphorylation of both proteins [6,7,10,24] and indeed PGC-1α interacts and is a potent coactivator of ERRα [22].
Recently, we investigated the effects of ABA/LANCL system on the mitochondrial oxidative metabolism and structural proteins. The overexpression of LANCL1/2 significantly increased, while silencing conversely reduced the mitochondrial number, OXPHOS complex I, proton gradient, glucose and palmitate-dependent respiration, transcription of uncoupling proteins, expression of proteins involved in cytoskeletal, contractile and electrical functions [25]. These effects are mediated by transcription factor ERRα, upstream of the AMPK/PGC1-α axis and transcriptionally controlled by the LANCL1/2–ABA system [25]. The ABA/LANCL1-2 hormone/receptor system emerges as a new controller of cardiomyocyte “fitness” improving cardiac function and resilience to hypoxic and dysmetabolic conditions via a reciprocal transcriptional stimulation with ERRα [25,26].
The aims of this study were two-fold: i) to investigate the role of the ABA/LANCL1-2 system in ROS production and detoxification by comparing the expression levels of the principal enzymes involved in these pathways and ROS content in LANCL1/2-overexpressing vs. LANCL1/2-silenced rat H9c2 cardiomyocytes; ii) to investigate a possible role for ERRα transcription factors in the ABA/LANCL1-2 signaling pathway in LANCL1/2-overexpressing H9c2 cells.
2. Materials and Methods
2.1. Cell Culture
H9c2 rat cardiomyocytes were purchased from ATCC (LGC Standards s.r.l. Milan, Italy). Cells were incubated in DMEM high glucose (Sigma-Aldrich, Milan, Italy) containing 10% heat-inactivated fetal bovine serum (Sigma-Aldrich, Milan, Italy) and 1% penicillin -streptomycin (Sigma-Aldrich, Milan, Italy) at 37°C in a humidified atmosphere with 5% CO2.
2.2. Lentiviral Cell Transduction
The lentiviral plasmids for silencing rat LANCL1 (SHL1), LANCL2 (SHL2) and ERRα (SHERRα) (plasmid ID: VB010000-0005mme, VB181016-1107sen, VB181016-1124zjp, VB221005-1073jxq) and the respective control vectors (SCR for a control scramble shRNA), were purchased from Vector Builder (Chicago, IL, USA). Lentiviral transductions were performed as described in [6]. hLANCL1 (OVL1) and hLANCL2 (OVL2) were overexpressed in rat H9c2 cardiomyocytes with pBABE vectors, constructed as described in [10], with the empty vector pBABE (Addgene, Watertown, MA, USA) as negative control (PLV).
2.3. qPCR Analysis
After 18 hours of starvation, cardiomyocytes were treated or not with 100 nM ABA for 4 hours and subjected to subsequent analyses. RNA extraction, cDNA synthesis and quantitative real-time PCR (qPCR) were performed as described previously [7]. Gene-specific primers were purchased from Sigma-Aldrich (Milan, Italy) and are listed in Table 1. Comparisons in the gene expression were performed using the iQ5 Optical System Software version 1.0 (Bio-Rad Laboratories, Milan, Italy) by 2-△△Ct method [7]. Hprt1 was used as the housekeeping gene for normalization.
2.4. Western Blot Analysis
After transduction, 1 x 106/well H9c2 cells were plated in 6-well plates and allowed to adhere for 24 hours. After serum deprivation for 18 hours, cardiomyocytes were washed once with HBSS and then incubated for 1 hour at 37°C with or without 100 nM ABA. Thereafter, cells were lysed in ice-cold lysis buffer [20 mM Tris-HCl (pH 7.4), 150 mM NaCl, 1 mM EDTA, 1% NP40 and Protease Inhibitor Cocktail] and briefly sonicated, and the protein concentration was determined according to a standard Bradford assay. Proteins (30 μg) were separated by SDS-PAGE and transferred to nitrocellulose membranes (Bio-Rad, Milan, Italy). Membranes were blocked for 1 hour with TBST containing 5% non-fat dry milk and incubated for 1 hour at room temperature with primary antibodies (Table 2). Following incubation with the appropriate secondary antibodies (Table 2), band intensities were quantified by the Quantity One SW software (Bio-Rad, Milan, Italy) using standard ECL (GE Healthcare, Milan, Italy).
2.5. ROS Detection Assays
Two different fluorescent ROS-sensitive probes were used, dichlorodihydrofluorescein diacetate (H2DCFDA) and MitoSOX Red [27,28]. DCFDA diffuses into the cells and is then deacetylated by cellular esterases to a non-fluorescent compound, which becomes fluorescent upon oxidation by ROS, thus behaving as a “whole cell” sensor of ROS production. MitoSOX Red is instead specifically targeted to mitochondria and allows detection of mitochondrial ROS.
For DCFDA-based assays, H9c2 cells were cultured overnight at 1 x 104/well in a 96-well plate in DMEM high glucose with 10% FBS and 1% penicillin-streptomycin, then were washed once with HBSS and loaded for 45 minutes with 10 μM dichlorodihydrofluorescein diacetate (H2DCFDA) at 37°C in HBSS without or with 100 nM ABA. At the end of incubation, the supernatant was removed, cells were washed once with HBSS and 100 μL HBSS was added to each well. The fluorescence (excitation at 488 nm and emission at 530 nm), monitored with a plate reader (Clariostar plus; BMG Lab Technologies), was calculated by the means fluorescence from 5 acquisitions/well; each experimental condition was assayed in at least 8 wells.
For MitoSOX-based assays, MitoSOX Red reagent (5 mM) (Thermo Fisher Scientific, Waltham, USA) was resuspended in DMSO and diluted in HBSS to obtain a final concentration of 5 μM as working solution. Cells were incubated in 500 μL of working solution for 20 minutes at 37°C in a CO2 incubator and positive staining was evaluated by a Leica TCS SP2 confocal microscope equipped with a x60 N.A. 1.4 oil immersion objective.
2.6. Statistical Analysis
All data were analyzed with the GraphPad Prism software version 7 (GraphPad Software, San Diego, USA). All parameters were tested by two-tailed Student’s t test. P values less than 0.05 were considered significant.
3. Results
3.1. Overexpression and Silencing of LANCL1 or LANCL2 in H9c2 Cells
Recently, it has been reported that the ABA-LANCL1/2 hormone-receptor system plays an important role in NO-mediated cardiomyocyte protection after hypoxia/reoxygenation [6]. In order to study the functions of ABA and its receptors on the control of oxidative stress and ROS metabolism in cardiomyocytes, H9c2 cells transfected to obtain double-silenced or overexpressing cells for both receptors LANCL1 and LANCL2. Overexpression (OVL1+2) or silencing (SHL1+2) of LANCL1/2 were obtained using retroviral or lentiviral infection, respectively, and were confirmed by Western blot. The fold increase of protein expression was approximately 6 for LANCL1 and 30 for LANCL2 (Figure 1A) compared with control cells infected with the empty vector (PLV). Moreover, in silenced cells we observed a reduction of the transcription and expression of both receptors, analyzed by qPCR and Western Blot, respectively 76% for LANCL1 and 95% for LANCL2 (Figure 1B).
3.2. Overexpression of LANCL1 and LANCL2 Decreases, While Their Combined Silencing Increases, Transcription and Expression of Radicals-Generating COX2, XO and NOX4
In order to understand the role of the LANCL proteins in inflammation and ROS production in H9c2, the expression levels of radicals-generating COX2, XO and NOX4 were evaluated in cells overexpressing or silenced for both LANCL1 and LANCL2, treated or not with ABA, using qPCR and Western blot analysis. In LANCL1/2-overexpressing H9c2 cells (OVL1+2), mRNA levels for COX2, XO and NOX4 were all significantly reduced (by 70-80%) compared with controls infected with the empty vector PLV; conversely, in double-silenced cells (SHL1+2), mRNA levels for the same enzymes increased approx. 6-fold over control cells infected with the scrambled sequences (SCR) (Figure 2A). Similar results were observed at the protein level, as measured by Western blot (Figure 2B). These results indicate that LANCL1/2 expression levels inversely control transcription and expression of COX2, XO and NOX4, with low levels of LANCL1/2 expression favoring an inflammatory and carbon and oxygen radicals producing cell phenotype.
3.3. LANCL1/2-Overexpression Increases, While Their Double-Silencing Decreases, Transcription, and Expression of the Radical Scavenging Enzymes SOD2 and GPX4
To further investigate a possible role of the ABA/LANCL1-2 system in ROS metabolism, we analyzed the expression of two important enzymatic antioxidants, superoxide dismutase (SOD2) and glutathione peroxidase (GPX4), that can directly or indirectly catabolize ROS to protect cells. Interestingly, overexpression of LANCL1/2 (OVL1+2) significantly increased transcription of both SOD2 and GPX4, 4- and 5-fold, respectively, as compared with control cells, infected with the empty vector (PLV) (Figure 3A). Moreover, treatment of the cells with 100 nM ABA further significantly increased mRNA levels of both SOD2 and GPX4. Transcription of the same target genes explored in LANCL1/2-overexpressing H9c2 was investigated in the double silenced cells. Results obtained were almost specular: mRNA levels of SOD2 and GPX4 were significantly reduced compared with control cells and did not increase in ABA-treated cells (Figure 3A). Western blot analysis confirmed approx. 80-fold higher protein levels of SOD2 and GPX4 in LANCL1/2-overexpressing vs. double-silenced H9c2 (Figure 3B). These results suggested that the ABA/LANCL system is involved not only in the control of ROS production but also in antioxidant defense. Recently, the NADPH/NADP ratio (2.1 vs 1.6) in LANCL1/2-overexpressing vs. double-silenced H9c2 cells was determined [6] and it was significantly higher in overexpressing cells. NADPH is required for the activity of Glutathione Reductase (GR), which keeps GmSH in the reduced form, essential for the antioxidant activity of Glutathione Peroxidase (GPX). These results are in line with the increased expression of GPX4 in overexpressing cells, suggesting that all enzymatic activities involved in the GSH-mediated detoxification system, which include SOD, GPX and GR, are activated.
3.4. LANCL1/2-Overexpression Decreases and Their Combined Silencing Conversely Increases ROS Content in H9c2 Cells
To further confirm the data obtained on gene and protein expression related to ROS metabolism, we analyzed the ROS content with two different methods, using fluorogenic substrates: i) dichlorodihydrofluorescein diacetate (H2DCFDA), that is deacetylated by cellular esterases into a non-fluorescent compound, which is subsequently oxidized by ROS into 2′,7′-dichlorofluorescein (DCF) (Figure 4A), and ii) the Mitochondrial Superoxide Indicator MitoSOX™, a fluorogenic dye specifically targeting mitochondria in live cells, the oxidation of which by mitochondrial superoxide produces a bright red fluorescence (Figure 4B). In both experiments, an increased basal ROS production was observed in LANCL1-2 silenced H9c2 compared with overexpressing cells, in agreement with the higher expression of enzymes involved in ROS production (NOX4 and XO) and the reduction of antioxidant enzymes (SOD2 and GPX) observed in double-silenced cells. ABA treatment significantly reduced mitochondrial ROS production in the overexpressing, but not in the double-silenced, cells (Figure 4B).
3.5. The ABA/LANCL1-2 System Controls ROS Metabolism via the Transcription Factor ERRα
The results described above indicate that LANCL1/2 proteins control ROS production in H9c2 at least in part by upregulating radical-scavenging enzymes and coenzymes and by reducing the activity of ROS-generating enzymes. The estrogen-related receptors (ERRs) are transcription factors belonging to an “orphan receptor” superfamily, which play important roles in the regulation of mitochondrial oxidative metabolism and in the defense against ROS production [14,15,16,17]. In particular, ERRα is a novel redox sensor and effector of a ROS defense program, regulating the transcription of many enzymes involved in ROS metabolism as a significant number of target genes that code for proteins involved in oxidative phosphorylation (complex I-IV, coenzyme Q and cytochrome c), all of the enzymes that constitute the TCA cycle, NOX4 and NOX5, SOD2 and GPX regulating Sirt3 expression by PGC-1α [16,18,23].
To investigate the role of ERRα in the protection against ROS production observed in LANCL1/2-overexpressing H9c2, ERRα was silenced by lentiviral infection using a vector containing sequences encoding specific shRNAs. As shown in Figure 5A, knockdown of ERRα in LANCL1/2-overexpressing cells (OVL1+2-SHERRα) was confirmed by both immunoblot (Figure 5A, upper panels) and qPCR (Figure 5A, lower panel). Expression of ERRα was reduced by approx. 80% relative to control cells infected with the vector containing scrambled silencing sequences (OVL1+2-SCR) (Figure 5A).
To investigate ROS metabolism in LANCL1/2-overexpressing and ERRα-silenced H9c2 cells (OVL1+2-SHERRα), we firstly evaluated transcription of radicals-generating COX2, NOX4 and XO and of radicals-scavenging SOD2 and GPX4 by qPCR. ERRα silencing in LANCL1/2-overexpressing cells (OVL1+2-SHERRα) significantly increased mRNA levels of all radicals-generating enzymes compared with control cells transfected with the scrambled sequences (OVL1+2-SCR) (Figure 5B). Conversely, transcription of the antioxidant enzymes SOD2 and GPX4 decreased in OVL1+2-SHERRα cells compared with controls (Figure 5C). The increase or decrease of mRNA levels were indeed very marked (logarithmic) as a result of ERRα silencing. Western blot analysis confirmed a significant increase of radicals-generating (Figure 5B, lower panel) and decrease of radicals-scavenging enzymes (Figure 5C, lower panel) in ERRα-silenced LANCL1/2-overexpressing H9c2 compared with control cells, transfected with the scrambled sequences for ERRα. These results suggested that ERRα is necessary for the decreased expression of radicals-generating enzymes and the increased expression of protecting enzymes observed in LANCL1/2-overexpressing vs. double-silenced cells.
3.6. ERRα Silencing Increases ROS Production in LANCL1/2-Overexpressing Cells
We next investigated the ROS content in ERRα-silenced-LANCL1/2-overexpressing H9c2 cells (OVL1+2-SHERRα) compared with control LANCL1/2-overexpressing cells, transfected with the scrambled sequences for ERRα (OVL1+2-SCR). We quantified ROS using the same methods employed to compare ROS content in LANCL1/2-overexpressing vs. double-silenced cells (Figure 4). ERRα silencing significantly increased ROS production in LANCL1/2-overexpressing cells (OVL1+2-SHERRα) vs. controls (OVL1+2-SCR), as determined with both fluorescent probes, the highest increase (2.5 times over control values) being observed with the mitochondrial dye MitoSOX™.These results indicate that ERRα plays a critical role in the reduced ROS content observed in LANCL1/2-overexpressing vs. double-silenced H9c2 cells (Figure 4). Indeed, a 2.5-fold increase would abrogate the significant reduction of ROS content observed in LANCL1/2-overexpressing vs. double-silenced cells shown in Figure 4. Also noteworthy is the fact that treatment with ABA significantly reduces (by 50%) ROS content in LANCL1/2-overexpressing cells, in spite of its stimulation of mitochondrial respiration [6]. Silencing of ERRα abrogates reduction by ABA of mitochondrial ROS production and increases “whole cell” DCFDA fluorescence in LANCL1/2-overexpressing cells.
4. Discussion
Collectively, results obtained in this study outline an unknown regulatory role for the ABA/LANCL1-2 hormone/receptors system in cardiomyocyte protection from ROS-dependent oxidative stress, via the transcription factor ERRα.
A reciprocal relationship emerges between LANCL1/2 expression levels and transcription and expression of critical ROS-scavenging and ROS-producing enzymes, which depends on the transcription factor ERRα. The result of this regulatory mechanism is that LANCL1/2-overexpressing H9c2 have a significantly reduced whole cell and mitochondrial ROS content compared with double-silenced cells, in the face of a significantly steeper mitochondrial proton gradient [6]. Thus, LANCL1/2-overexpressing cells appear to be more protected than double-silenced cells against mitochondrial ROS generation, despite having a higher respiratory chain activity.
Several key ROS-producing and -scavenging enzymes appear to be controlled at the transcriptional and translational level by the ABA/LANCL1-2/ERRα system. Radicals-generating enzymes, COX2, NOX4 and XO were all significantly reduced in LANCL1-2-overexpressing vs. double-silenced H9c2 cells. COX2, the rate-limiting enzyme in the synthesis of prostaglandins and a key player in inflammation and oxidative stress [30], was reduced 15-fold in LANCL1/2-overexpressing compared with double-silenced cells (Figure 2B). mRNA levels of NOX4, the most abundant isoform of the H2O2-generating NADPH oxidases family in cardiomyocytes [31] were 23-fold lower in LANCL1/2-overexpressing vs. double-silenced cells (Figure 2A). Finally, protein levels of Xanthine Oxidase (XO), which produces O2- after sulfhydryl oxidation, as occurs during myocardial ischemia/reperfusion (I/R) [32], were about 6-fold lower in LANCL1/2-overexpressing vs. double-silenced cells (Figure 2B).
Radical-scavenging enzymes were instead increased in LANCL1/2-overexpressing compared with double-silenced H9c2. Transcription and expression of SOD2, a Mn-SOD localized in the mitochondrial matrix, arguably the most important site of cellular ROS production, was 40- and 50-fold higher in LANCL1/2-overexpressing vs. double-silenced cells (Figure 3). The SOD family detoxifies O2− through its conversion to H2O2, which is then reduced to water by GPX, through the oxidation of GSH [33]. GPX4 protein levels were indeed 45-fold higher in LANCL1/2-overexpressing vs. double-silenced cells (Figure 3B). Besides reducing H2O2 and small hydroperoxides in general, GPX4 also has the ability to reduce hydroperoxides in complex lipids (phospholipids, cholesterol and cholesteryl esters), even when they are inserted into biomembranes or lipoproteins [34]. Thus, GPX4 is essential to prevent the accumulation of toxic lipid hydroperoxides, which can trigger a mode of cell death termed ferroptosis, associated with many cardiovascular diseases [35].
The profound effect exerted by the LANCL proteins on radicals-generating and -scavenging enzymes results in a significantly reduced ROS content in the overexpressing compared with the double-silenced H9c2, particularly in mitochondria (Figure 4B). Interestingly, treatment with ABA of LANCL1/2-overexpressing cells further reduced mitochondrial ROS content (Figs. 4B and 6B), an effect which was not observed in double-silenced cells or in ERRα-silenced overexpressing cells. Thus, the beneficial effect of ABA in reducing cardiomyocyte mitochondrial ROS content requires a functional LANCL1/2-ERRα axis.
Results obtained here imply that the ABA/LANCL hormone/receptors system exerts a protective role on (mitochondrial) ROS production and that conversely a reduced activity of this signaling pathway necessarily results in an increased susceptibility of cardiomyocytes to oxidative stress. This conclusion may bear significant consequences in the clinical setting; on the one hand, activation of the LANCL1/2-signaling pathway is attainable by means of pharmacological or nutraceutical compositions titrated in ABA [3]; on the other hand, reduced endogenous LANCL1/2 expression levels may be responsible for an increased susceptibility of cardiomyocytes to oxidative stress, a possibility worth exploring in patients with a poor outcome after cardiac reperfusion.
It should be emphasized that LANCL1/2-overexpressing cells have a significantly steeper proton gradient than double-silenced cells [6]; thus, it could be anticipated that they should also have a higher production of ROS as a consequence of a higher respiratory chain activity. Instead, double-silenced H9c2 cells, despite a much reduced ΔΨ compared with overexpressing cells, produce significantly more ROS. These profound differences in mitochondrial function and ROS generation between overexpressing and double-silenced H9c2 demonstrates the important role played by the LANCL1/2 proteins in the regulation of mitochondrial function. Moreover, the “improved” phenotype of LANCL1/2-overexpressing compared with double-silenced cells regarding ROS metabolism (lower transcriptional and protein levels of radicals-generating enzymes, higher levels of radicals-scavenging enzymes and lower levels of cellular and mitochondrial ROS) is dependent on the transcription factor ERRα, as its knock down abrogates all the above features. A general, cell type-independent role of the LANCL1/2 receptor system in protecting cells against radicals emerges also from very recent reports on hepatocellular carcinoma cells and on testicular cells [36,37].
Taken together with previously published results [4,6,25,26], data reported here allow to outline a multifaceted role of the ABA/LANCL1-2 hormone/receptors system in cardiomyocytes. Targeting this system, via LANCL1/2 overexpression or via stimulation with ABA of endogenous LANCL1-2 proteins (expression in the heart is the highest among non-neurological tissues, [6] ) results in the following responses: i) increased NO production, both under normoxia and hypoxia, via eNOS activation by the AMPK/PGC-1α axis; ii) increased mitochondrial respiration with higher basal and maximal respiration rates, a doubling of the spare respiratory capacity and a steeper proton gradient (∆Ψ) under normoxia and after hypoxia/reoxygenation [6,25]; iii) improved cell vitality after H/R; iv) increased glucose uptake and oxidation, with higher cell NADPH content; v) increased fatty acid-fueled respiration rate [25] vi) increased O2 consumption under normoxia; vii) reduced mitochondrial ROS content resulting from an improved enzymatic expression pattern, with less ROS-producing and more ROS-scavenging enzymes, in the face of an increased mitochondrial electron chain transport activity. These pleiotropic functions controlled by the LANCL proteins in cardiomyocytes require a signaling pathway dependent on the activation of AMPK, PGC-1α and ERRα, arguably among the most important and evolutionarily ancient stress signals [26].
Author Contributions
Conceptualization, S.S., E.Z. and L.S.; methodology, S.S., E.Z. and L.S.; validation, S.S., L.G., M.P., M.M. and B.C.; investigation, S.S., L.G., M.P., M.M. and B.C.; data curation, S.S.; writing—original draft preparation, S.S., E.Z. and L.S.; writing—review and editing, S.S., L.G., M.P., M.M. and B.C., E.Z. and L.S.; visualization, S.S.; supervision, E.Z. and L.S.; funding acquisition, E.Z. and L.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the University of Genova, FRA 2020–2021, to L.S. and to E.Z.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Wang, J.Y.; Lin, P.Y.; Al-Babili, S. On the biosynthesis and evolution of apocarotenoid plant growth regulators. Semin. Cell Dev. Biol. 2021, 109, 3–11. [Google Scholar] [CrossRef] [PubMed]
- Bruzzone, S.; Ameri, P.; Briatore, L.; Mannino, E.; Basile, G.; Andraghetti, G.; Grozio, A.; Magnone, M.; Guida, L.; Scarfì, S.; et al. The plant hormone abscisic acid increases in human plasma after hyperglycemia and stimulates glucose consumption by adipocytes and myoblasts. FASEB J. 2012, 26, 1251–1260. [Google Scholar] [CrossRef] [PubMed]
- Magnone, M.; Leoncini, G.; Vigliarolo, T.; Emionite, L.; Sturla, L.; Zocchi, E.; Murialdo, G. Chronic Intake of Micrograms of Abscisic Acid Improves Glycemia and Lipidemia in a Human Study and in High-Glucose Fed Mice. Nutrients 2018, 10, 1495. [Google Scholar] [CrossRef] [PubMed]
- Spinelli, S.; Magnone, M.; Guida, L.; Sturla, L.; Zocchi, E. The ABA-LANCL hormone-receptor system in the control of glycemia, of cardiomyocyte energy metabolism and in neuro-protection: A new ally in the treatment of diabetes mellitus? Int J Mol Sci 2023, 24, 1199. [Google Scholar] [CrossRef] [PubMed]
- Magnone, M.; Spinelli, S.; Begani, G.; Guida, L.; Sturla, L.; Emionite, L.; Zocchi, E. Abscisic acid improves insulin action on glycemia in insulin-deficient mouse models of type 1 diabetes. Metabolites 2022, 12, 523. [Google Scholar] [CrossRef] [PubMed]
- Spinelli, S.; Guida, L.; Vigliarolo, T.; Passalacqua, M.; Begani, G.; Magnone, M.; Sturla, L.; Benzi, A.; Ameri, P.; Lazzarini, E.; et al. The ABA-LANCL1/2 Hormone-Receptors System Protects H9c2 Cardiomyocytes from Hypoxia Induced Mitochondrial Injury via an AMPK- and NO-Mediated Mechanism. Cells 2022, 11, 2888. [Google Scholar] [CrossRef] [PubMed]
- Spinelli, S.; Cossu, V.; Passalacqua, M.; Hansen, J.B.; Guida, L.; Magnone, M.; Sambuceti, G.; Marini, C.; Sturla, L.; Zocchi, E. The ABA/LANCL1/2 Hormone/Receptor System Controls Adipocyte Browning and Energy Expenditure. Int. J. Mol. Sci. 2023, 24, 3489. [Google Scholar] [CrossRef] [PubMed]
- Scarfì, S.; Fresia, C.; Ferraris, C.; Bruzzone, S.; Fruscione, F.; Usai, C.; Benvenuto, F.; Magnone, M.; Podestà, M.; Sturla, L.; et al. The plant hormone abscisic acid stimulates the proliferation of human hemopoietic progenitors through the second messenger cyclic ADP-ribose. Stem. Cells. 2009, 27, 2469–2477. [Google Scholar] [CrossRef] [PubMed]
- Sturla, L.; Fresia, C.; Guida, L.; Bruzzone, S.; Scarfì, S.; Usai, C.; Fruscione, F.; Magnone, M.; Millo, E.; Basile, G.; et al. LANCL2 Is Necessary for Abscisic Acid Binding and Signaling in Human Granulocytes and in Rat Insulinoma Cells. J Biol Chem 2009, 284, 28045–28057. [Google Scholar] [CrossRef] [PubMed]
- Spinelli, S.; Begani, G.; Guida, L.; Magnone, M.; Galante, D.; D’Arrigo, C.; Scotti, C.; Iamele, L.; De Jonge, H.; Zocchi, E.; et al. LANCL1 binds abscisic acid and stimulates glucose transport and mitochondrial respiration in muscle cells via the AMPK/PGC-1α/Sirt1 pathway. Mol. Metab. 2021, 53, 101263. [Google Scholar] [CrossRef] [PubMed]
- Sies, H.; Jones, D.P. Reactive oxygen species (ROS) as pleiotropic physiological signaling agents. Nat Rev Mol Cell Biol 2020, 21, 363–383. [Google Scholar] [CrossRef] [PubMed]
- Cadenasa, S. ROS and redox signaling in myocardial ischemia-reperfusion injury and cardioprotection. Free Radic. Biol. Med. 2018, 117, 76–89. [Google Scholar] [CrossRef] [PubMed]
- Shenshu Yang, S.; Lian, G. ROS and diseases: Role in metabolism and energy supply. Mol. Cell. Biochem. 2020, 467, 1–12. [Google Scholar] [CrossRef]
- Valko, M.; Leibfritz, D.; Moncol, J.; Cronin, M.T.K.; Mazur, M.; Telser, J. Free radicals and antioxidants in normal physiological functions and human disease. Int J Biochem Cell Biol 2007, 39, 44–84. [Google Scholar] [CrossRef] [PubMed]
- He, L.; He, T.; Farrar, S.; Jia, L.; Liu, T.; Ma, X. Antioxidants maintain cellular redox homeostasis by elimination of reactive oxygen species. Cell Physiol Biochem 2017, 44, 532–553. [Google Scholar] [CrossRef] [PubMed]
- Scholtes, C.; Giguère, V. Transcriptional Regulation of ROS Homeostasis by the ERR Subfamily of Nuclear Receptors. Antioxidants 2021, 10, 437. [Google Scholar] [CrossRef]
- Huss, J.M.; Garbacz, W.G.; Xie, W. Constitutive activities of estrogen-related receptors: Transcriptional regulation of metabolism by the ERR pathways in health and disease. Biochim Biophys Acta 2015, 9, 1912–1927. [Google Scholar] [CrossRef] [PubMed]
- Vernier, M.; Dufour, C.R.; McGuirk, S.; Scholtes, C.; Li, X.; Bourmeau, G.; Kuasne, H.; Park, M.; St-Pierre, J.; Audet-Walsh, E.; et al. Estrogen-related Receptors are Targetable ROS sensors. Genes Dev 2020, 34, 544–559. [Google Scholar] [CrossRef] [PubMed]
- Giguère, V. Transcriptional Control of Energy Homeostasis by the Estrogen-related Receptors. Endocr. Rev. 2008, 29, 677–696. [Google Scholar] [CrossRef] [PubMed]
- Villena, J.A.; Hock, M.B.; Chang, V.Y.; Barcas, J.E.; Gigue, V.; Kralli, A. Orphan nuclear receptor estrogen-related receptor is essential for adaptive thermogenesis. Proc Natl Acad Sci U S A 2007, 104, 1418–1423. [Google Scholar] [CrossRef] [PubMed]
- Deblois, G.; Smith, H.W.; Tam, I.S.; Gravel, S.P.; Caron, M.; Savage, P.; Labbé, D.P.; Bégin, L.R.; Tremblay, M.L.; Park, M.; et al. ERRα Mediates Metabolic Adaptations Driving Lapatinib Resistance in Breast Cancer. Nat. Commun. 2016, 7, 12156. [Google Scholar] [CrossRef] [PubMed]
- Scarpulla, R.C. Metabolic control of mitochondrial biogenesis through the PGC-1 family regulatory network. Biochim Biophys Acta 2011, 1813, 1269–1278. [Google Scholar] [CrossRef] [PubMed]
- Kong, X.; Wang, R.; Xue, Y.; Liu, X.; Zhang, H.; Chen, Y.; Fang, F.; Chang, Y. Sirtuin 3, a new target of PGC-1alpha, plays an important role in the suppression of ROS and mitochondrial biogenesis. PLoS ONE 2010, 22, e11707. [Google Scholar]
- Magnone, M.; Emionite, L.; Guida, L.; Vigliarolo, T.; Sturla, L.; Spinelli, S.; Buschiazzo, A.; Marini, C.; Sambuceti, G.; De Flora, A.; et al. Insulin-independent stimulation of skeletal muscle glucose uptake by low-dose abscisic acid via AMPK activation. Sci. Rep. 2020, 10, 1454. [Google Scholar] [CrossRef] [PubMed]
- Spinelli, S.; Guida, L.; Passalacqua, M.; Magnone, M.; Cossu, V.; Sambuceti, G.; Marini, C.; Sturla, L.; Zocchi, E. Abscisic Acid and Its Receptors LANCL1 and LANCL2 Control Cardiomyocyte Mitochondrial Function, Expression of Contractile, Cytoskeletal and Ion Channel Proteins and Cell Proliferation via ERRα. Antioxidants 2023, 12, 1692. [Google Scholar] [CrossRef] [PubMed]
- Spinelli, S.; Bruschi, M.; Passalacqua, M.; Guida, L.; Magnone, M.; Sturla, L.; Zocchi, E. Estrogen-Related Receptor α: A Key Transcription Factor in the Regulation of Energy Metabolism at an Organismic Level and a Target of the ABA/LANCL Hormone Receptor System. Int J Mol Sci 2024, 25, 4796. [Google Scholar] [CrossRef] [PubMed]
- Gomes, A.; Fernandes, E.; Lima, J.L. Fluorescence probes used for detection of reactive oxygen species. J Biochem Biophys Methods 2005, 65, 45–80. [Google Scholar] [CrossRef] [PubMed]
- Robinson, K.M.; Janes, M.S.; Pehar, M.; Monette, J.S.; Ross, M.F.; Hagen, T.M.; Murphy, M.P.; Beckmanet, J.S. Selective fluorescent imaging of superoxide in vivo using ethidium-based probes. Proc Natl Acad Sci U S A 2006, 103, 15038–15043. [Google Scholar] [CrossRef] [PubMed]
- Vigliarolo, T.; Guida, L.; Millo, E.; Fresia, C.; Turco, E.; de Flora, A.; Zocchi, E. Abscisic acid transport in human erythrocytes. J. Biol. Chem. 2015, 290, 13042–13052. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Borisenko, G.G.; Osipov, A.; Martin, I.; Chen, R.; Shvedova, A.A.; Sorokin, A.; Tyurina, Y.Y.; Potapovich, A.; Tyurin, V.A.; et al. Arachidonic acid-induced carbon-centered radicals and phospholipid peroxidation in cyclo-oxygenase-2-transfected PC12 cells. J Neurochem 2004, 90, 1036–1049. [Google Scholar] [CrossRef] [PubMed]
- Sofiullah, S.S.M.; Murugan, D.D.; Muid, S.A.; Seng, W.Y.; Abdul Kadir, S.Z.S.A.; Abas, R.; Ridzuan, N.R.A.; Zamakshshari, N.H.; Woon, C.K. Natural bioactive compounds targeting NADPH Oxidase pathway in cardiovascular diseases. Molecules 2023, 28, 1047. [Google Scholar] [CrossRef] [PubMed]
- Granger, D.N.; Kvietys, P.R. Reperfusion injury and reactive oxygen species: The evolution of a concept. Redox Biol. 2015, 6, 524–551. [Google Scholar] [CrossRef] [PubMed]
- Brigelius-Flohé, R.; Maiorino, M. Glutathione peroxidases. Biochim Biophys Acta 2013, 5, 3289–3303. [Google Scholar] [CrossRef] [PubMed]
- Forcina, G.C.; Dixon, S.J. GPX4 at the Crossroads of Lipid Homeostasis and Ferroptosis. Proteomics 2019, 18, e1800311. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Yan, Y.; Qi, C.; Liu, J.; Li, L.; Wang, J. The Role of Ferroptosis in Cardiovascular Disease and Its Therapeutic Significance. Front Cardiovasc Med 2021, 8, 733229. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Tsui, Y.M.; Ho, D.W.; Chung, C.Y.; Sze, K.M.; Lee, E.; Cheung, G.C.; Zhang, V.X.; Wang, X.; Lyu, X.; et al. LANCL1, a cell surface protein, promotes liver tumor initiation through FAM49B-Rac1 axis to suppress oxidative stress. Hepatology 2024, 79, 323–340. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Wang, J.; Shi, S.; Lan, X.; Cheng, X.; Li, L.; Zou, Y.; Jia, L.; Liu, W.; Luo, Q.; et al. LanCL2 Implicates in Testicular Redox Homeostasis and Acrosomal Maturation. Antioxidants 2024, 13, 534. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Overexpression and silencing of LANCL1/2 in H9c2 rat cardiomyocytes. LANCL1 and LANCL2 were overexpressed (A) or silenced (B) in H9c2 cells by lentiviral infection.(A) Left panel, representative Western blots of LANCL1 and LANCL2 protein expression in cells overexpressing both LANCL proteins (OVL1+2) or infected with the empty vector (PLV); right panel, densitometric quantitation of the LANCL proteins expression in the same cell types normalized on PLV control and relative to vinculin. Values are normalized against vinculin, as housekeeping protein. (B) Left panel, representative Western blots of LANCL1 and LANCL2 in cells silenced for the expression of both proteins (SHL1+2) or infected with the scrambled sequences (SCR); upper right panel, densitometric quantitation of the LANCL proteins in the same cell types, normalized on SCR control and relative to vinculin; lower right panel, LANCL1/2 mRNA levels relative to control in LANCL1/2-silenced cells. Values are normalized against vinculin, as housekeeping protein. The exposure time of Western Blot panel of Figure 1A,B are different and higher in silenced H9c2 silenced cells. Data are the mean±SD from at least three experiments. *p<0.001 relative to control cells (PLV for overexpression or SCR for silencing) by unpaired t test.
Figure 1.
Overexpression and silencing of LANCL1/2 in H9c2 rat cardiomyocytes. LANCL1 and LANCL2 were overexpressed (A) or silenced (B) in H9c2 cells by lentiviral infection.(A) Left panel, representative Western blots of LANCL1 and LANCL2 protein expression in cells overexpressing both LANCL proteins (OVL1+2) or infected with the empty vector (PLV); right panel, densitometric quantitation of the LANCL proteins expression in the same cell types normalized on PLV control and relative to vinculin. Values are normalized against vinculin, as housekeeping protein. (B) Left panel, representative Western blots of LANCL1 and LANCL2 in cells silenced for the expression of both proteins (SHL1+2) or infected with the scrambled sequences (SCR); upper right panel, densitometric quantitation of the LANCL proteins in the same cell types, normalized on SCR control and relative to vinculin; lower right panel, LANCL1/2 mRNA levels relative to control in LANCL1/2-silenced cells. Values are normalized against vinculin, as housekeeping protein. The exposure time of Western Blot panel of Figure 1A,B are different and higher in silenced H9c2 silenced cells. Data are the mean±SD from at least three experiments. *p<0.001 relative to control cells (PLV for overexpression or SCR for silencing) by unpaired t test.
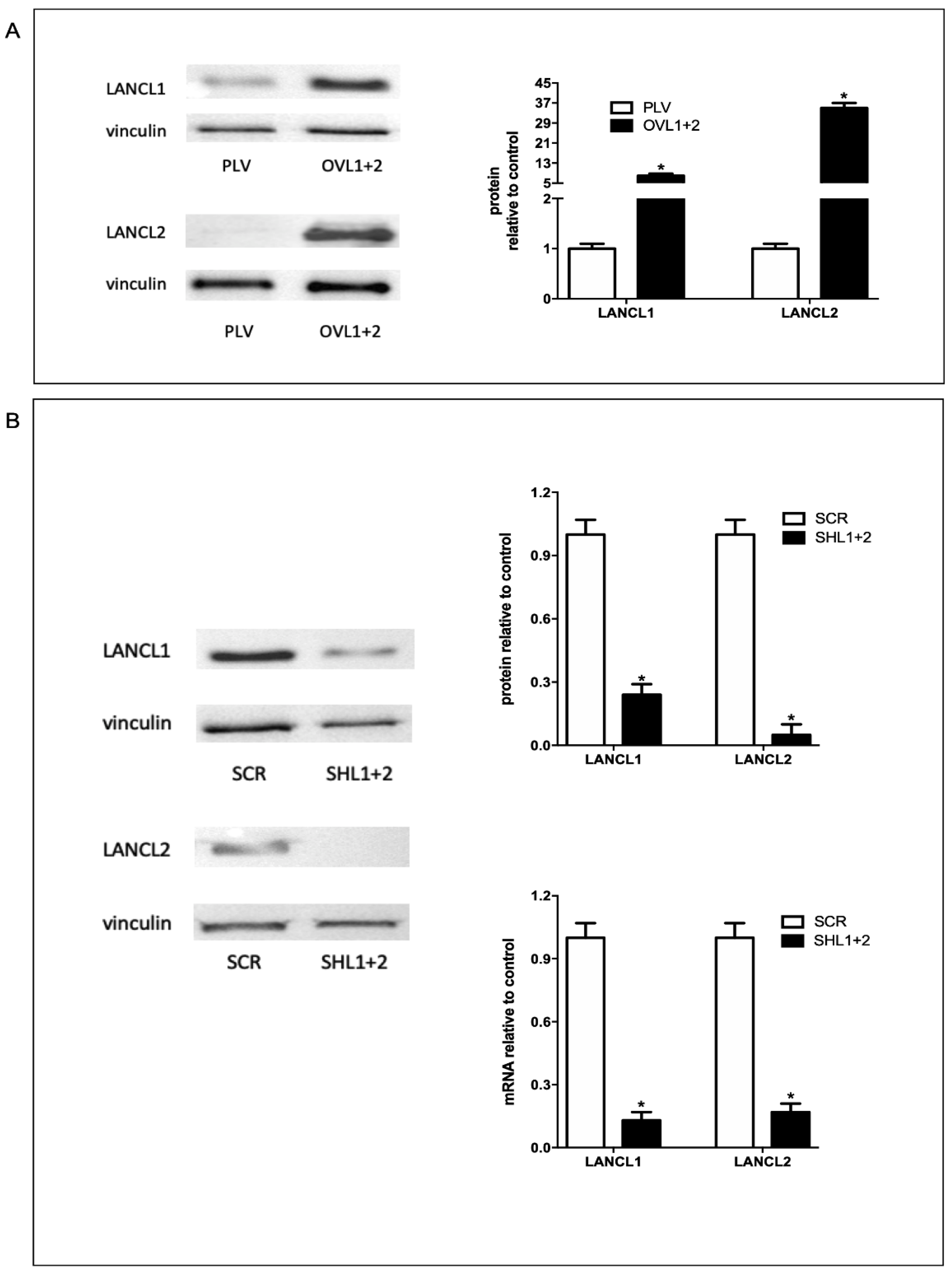
Figure 2.
Radicals-generating enzymes are reduced in LANCL1/2-overexpressing vs. double-silenced H9c2. (A) qPCR analysis of the transcription of radicals-generating enzymes (COX2, XO and NOX4) in cells overexpressing (OVL1+2) or silenced (SHL1+2) for LANCL1 and LANCL2 proteins and incubated in the absence or in the presence of 100 nM ABA for 4 hours. Results are expressed relative to control ABA-untreated, PLV cells. *p<0.001 relative to untreated control cells (PLV or SCR) and $p<0.02 relative to ABA-untreated OVL1+2 or SHL1+2 cells by unpaired t test. (B) Upper panel: representative Western blot image of COX2 and XO proteins in LANCL1/2-overexpressing or silenced cells, treated or not with 100 nM ABA for 4 hours. Lower panel: histograms are the mean±SD from at least three experiments. Results are expressed relative to ABA-untreated OVL1+2 cells. Values are normalized against vinculin, as housekeeping protein. Data are *p<0.001 relative to untreated OVL1+2 cells and $p<0.02 relative to ABA-untreated SHL1+2 cells by unpaired t test.
Figure 2.
Radicals-generating enzymes are reduced in LANCL1/2-overexpressing vs. double-silenced H9c2. (A) qPCR analysis of the transcription of radicals-generating enzymes (COX2, XO and NOX4) in cells overexpressing (OVL1+2) or silenced (SHL1+2) for LANCL1 and LANCL2 proteins and incubated in the absence or in the presence of 100 nM ABA for 4 hours. Results are expressed relative to control ABA-untreated, PLV cells. *p<0.001 relative to untreated control cells (PLV or SCR) and $p<0.02 relative to ABA-untreated OVL1+2 or SHL1+2 cells by unpaired t test. (B) Upper panel: representative Western blot image of COX2 and XO proteins in LANCL1/2-overexpressing or silenced cells, treated or not with 100 nM ABA for 4 hours. Lower panel: histograms are the mean±SD from at least three experiments. Results are expressed relative to ABA-untreated OVL1+2 cells. Values are normalized against vinculin, as housekeeping protein. Data are *p<0.001 relative to untreated OVL1+2 cells and $p<0.02 relative to ABA-untreated SHL1+2 cells by unpaired t test.
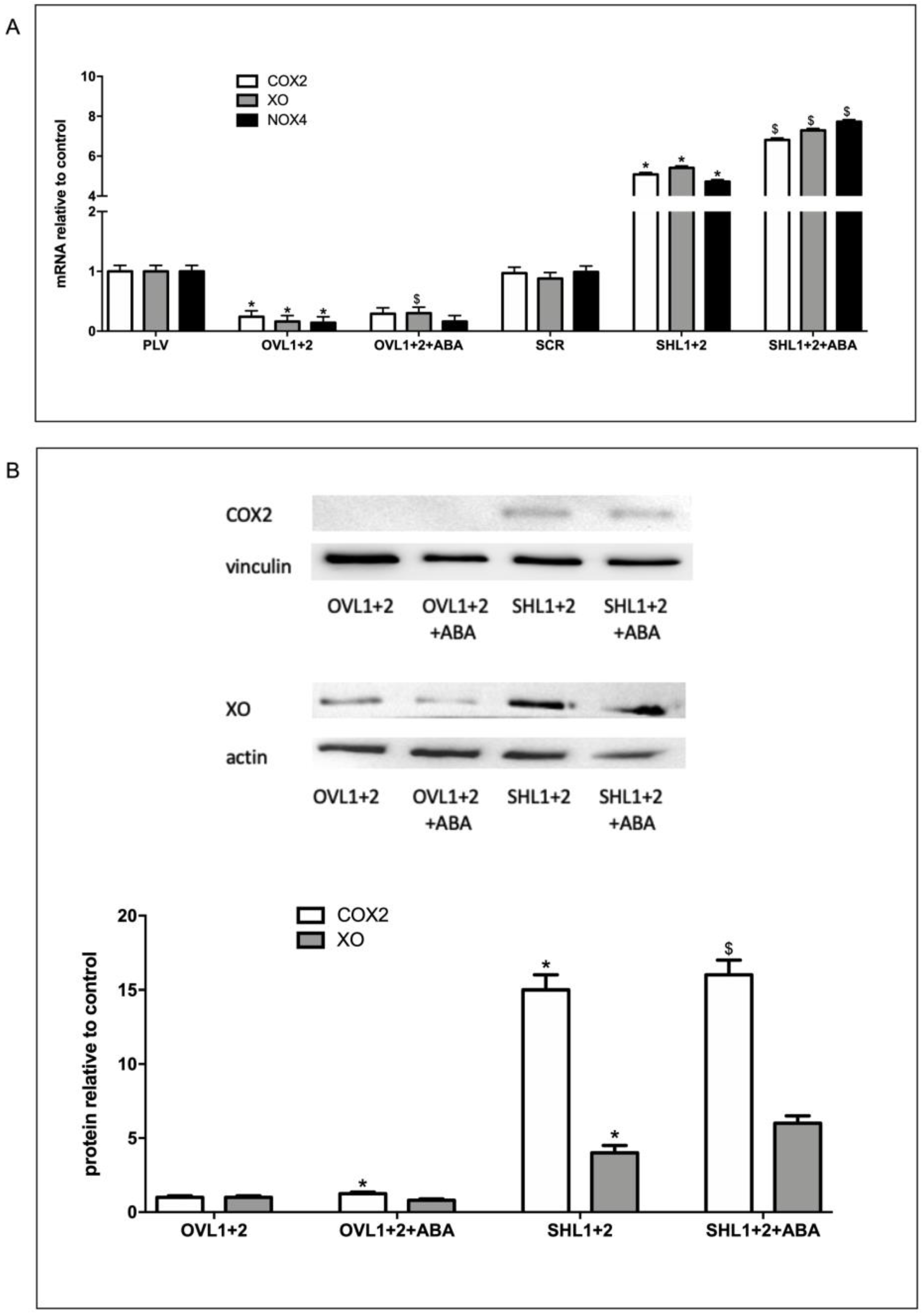
Figure 3.
Radicals-scavenging enzymes are increased in LANCL1/2-overexpressing vs. double-silenced H9c2. (A) qPCR analysis of the transcription of radicals-scavenging enzymes (SOD2 and GPX4) in cells overexpressing (OVL1+2) or silenced (SHL1+2) for LANCL1 and LANCL2 and incubated in the absence or in the presence of 100 nM ABA for 4 hours. Results are expressed relative to control ABA-untreated PLV cells. *p<0.001 relative to untreated control cells (PLV or SCR) and $p<0.02 relative to ABA-untreated OVL1+2 or SHL1+2 cells by unpaired t test. (B) Upper panel: a representative Western blot image of SOD2 and GPX4 in LANCL1/2-overexpressing or silenced cells, treated or not with 100 nM ABA for 4 hours. Lower panel: histograms are the mean±SD from at least three experiments. Results are expressed relative to ABA-untreated OVL1+2 cells. Values are normalized against vinculin, as housekeeping protein. Data are*p<0.001 relative to untreated OVL1+2 cells by unpaired t test.
Figure 3.
Radicals-scavenging enzymes are increased in LANCL1/2-overexpressing vs. double-silenced H9c2. (A) qPCR analysis of the transcription of radicals-scavenging enzymes (SOD2 and GPX4) in cells overexpressing (OVL1+2) or silenced (SHL1+2) for LANCL1 and LANCL2 and incubated in the absence or in the presence of 100 nM ABA for 4 hours. Results are expressed relative to control ABA-untreated PLV cells. *p<0.001 relative to untreated control cells (PLV or SCR) and $p<0.02 relative to ABA-untreated OVL1+2 or SHL1+2 cells by unpaired t test. (B) Upper panel: a representative Western blot image of SOD2 and GPX4 in LANCL1/2-overexpressing or silenced cells, treated or not with 100 nM ABA for 4 hours. Lower panel: histograms are the mean±SD from at least three experiments. Results are expressed relative to ABA-untreated OVL1+2 cells. Values are normalized against vinculin, as housekeeping protein. Data are*p<0.001 relative to untreated OVL1+2 cells by unpaired t test.
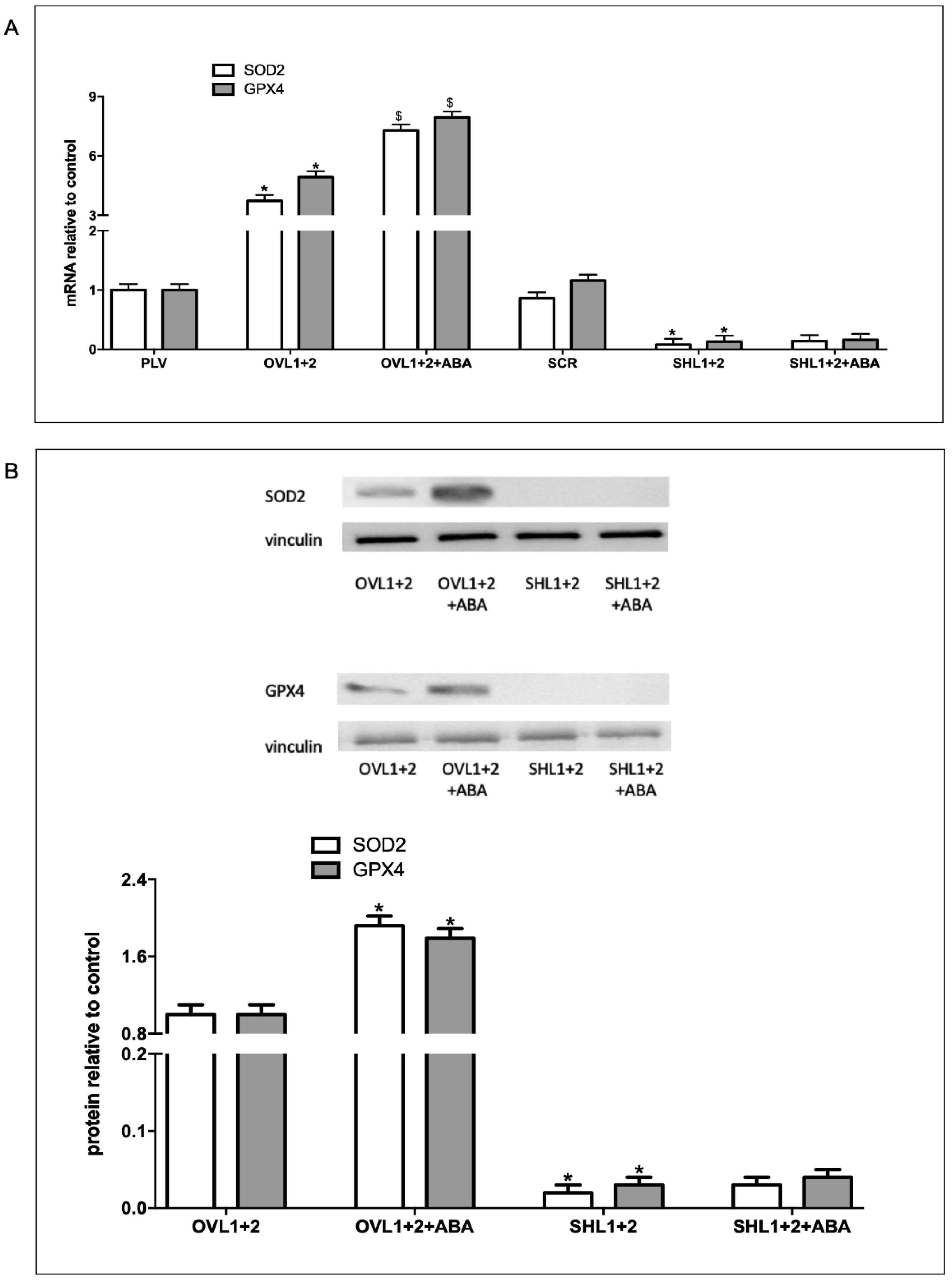
Figure 4.
LANCL1/2-overexpressing cells have a reduced ROS content compared with double-silenced H9c2. (A) Intracellular ROS production measured by DCF fluorimetric analysis in H9c2 cells. Results are expressed as fluorescence intensity in arbitrary units. (B) Mitochondrial superoxide anions were detected by confocal microscopy on MitoSOX-loaded H9c2 overexpressing (OVL1+2) or silenced (SHL1+2) for LANCL1 and LANCL2 proteins and incubated in the absence or in the presence of 100 nM ABA for 4 hours. Upper panel, representative images of the cells; lower panel, histograms represent mean cell fluorescence recorded in three independent experiments, relative to ABA-untreated OVL1+2 cells. *p<0.05 relative to untreated OVL1+2 cells and $p<0.02 relative to ABA-untreated SHL1+2 cells by unpaired t test.
Figure 4.
LANCL1/2-overexpressing cells have a reduced ROS content compared with double-silenced H9c2. (A) Intracellular ROS production measured by DCF fluorimetric analysis in H9c2 cells. Results are expressed as fluorescence intensity in arbitrary units. (B) Mitochondrial superoxide anions were detected by confocal microscopy on MitoSOX-loaded H9c2 overexpressing (OVL1+2) or silenced (SHL1+2) for LANCL1 and LANCL2 proteins and incubated in the absence or in the presence of 100 nM ABA for 4 hours. Upper panel, representative images of the cells; lower panel, histograms represent mean cell fluorescence recorded in three independent experiments, relative to ABA-untreated OVL1+2 cells. *p<0.05 relative to untreated OVL1+2 cells and $p<0.02 relative to ABA-untreated SHL1+2 cells by unpaired t test.
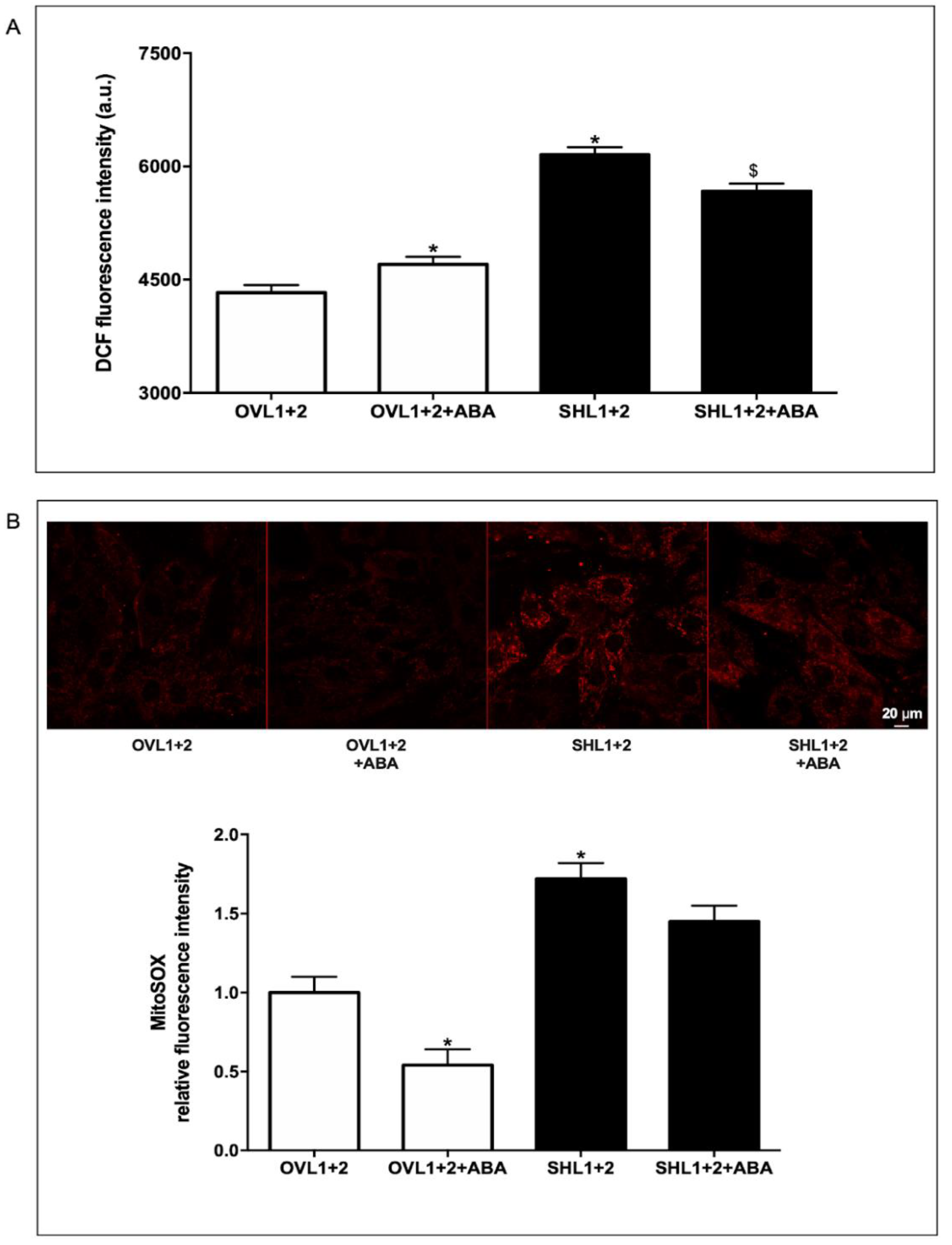
Figure 5.
ERRα silencing in LANCL1/2-overexpressing cells increases radicals-generating and decreases radicals-scavenging enzymes. (A) OVL1+2 cells were silenced for the expression of ERRα by lentiviral infection. Upper left panel, representative Western blot of ERRα in OVL1+2 cells silenced for the expression of ERRα (OVL1+2-SHERRα) or infected with the empty vector (OVL1+2-SCR); upper right panel, densitometric quantitation of ERRα in the same cell types. Values are normalized against vinculin, as housekeeping protein; lower panel, ERRα mRNA levels relative to control OVL1+2-SCR cells in ERRα-silenced cells (OVL1+2-SHERRα). *p<0.001 relative to control OVL1+2-SCR cells by unpaired t test. (B) qPCR analysis (upper panel) and Western blot analysis (lower panels) of radicals-generating enzymes (COX2, XO and NOX4) in OVL1+2 cells silenced for ERRα (OVL1+2-SHERRα) and incubated in the absence or in the presence of 100 nM ABA for 4 hours. Results are expressed relative to ABA-untreated OVL1+2-SCR cells. *p<0.001 relative to ABA-untreated OVL1+2-SCR cells by unpaired t test. (C) qPCR analysis (upper panel) and Western blot analysis (lower panels) of radicals-scavenging enzymes (SOD2 and GPX4) in OVL1+2 cells silenced for ERRα (OVL1+2-SHERRα) and incubated in the absence or in the presence of 100 nM ABA for 4 hours. Results are expressed relative to control ABA-untreated OVL1+2-SCR cells. *p<0.001 relative to untreated OVL1+2-SCR cells by unpaired t test.
Figure 5.
ERRα silencing in LANCL1/2-overexpressing cells increases radicals-generating and decreases radicals-scavenging enzymes. (A) OVL1+2 cells were silenced for the expression of ERRα by lentiviral infection. Upper left panel, representative Western blot of ERRα in OVL1+2 cells silenced for the expression of ERRα (OVL1+2-SHERRα) or infected with the empty vector (OVL1+2-SCR); upper right panel, densitometric quantitation of ERRα in the same cell types. Values are normalized against vinculin, as housekeeping protein; lower panel, ERRα mRNA levels relative to control OVL1+2-SCR cells in ERRα-silenced cells (OVL1+2-SHERRα). *p<0.001 relative to control OVL1+2-SCR cells by unpaired t test. (B) qPCR analysis (upper panel) and Western blot analysis (lower panels) of radicals-generating enzymes (COX2, XO and NOX4) in OVL1+2 cells silenced for ERRα (OVL1+2-SHERRα) and incubated in the absence or in the presence of 100 nM ABA for 4 hours. Results are expressed relative to ABA-untreated OVL1+2-SCR cells. *p<0.001 relative to ABA-untreated OVL1+2-SCR cells by unpaired t test. (C) qPCR analysis (upper panel) and Western blot analysis (lower panels) of radicals-scavenging enzymes (SOD2 and GPX4) in OVL1+2 cells silenced for ERRα (OVL1+2-SHERRα) and incubated in the absence or in the presence of 100 nM ABA for 4 hours. Results are expressed relative to control ABA-untreated OVL1+2-SCR cells. *p<0.001 relative to untreated OVL1+2-SCR cells by unpaired t test.
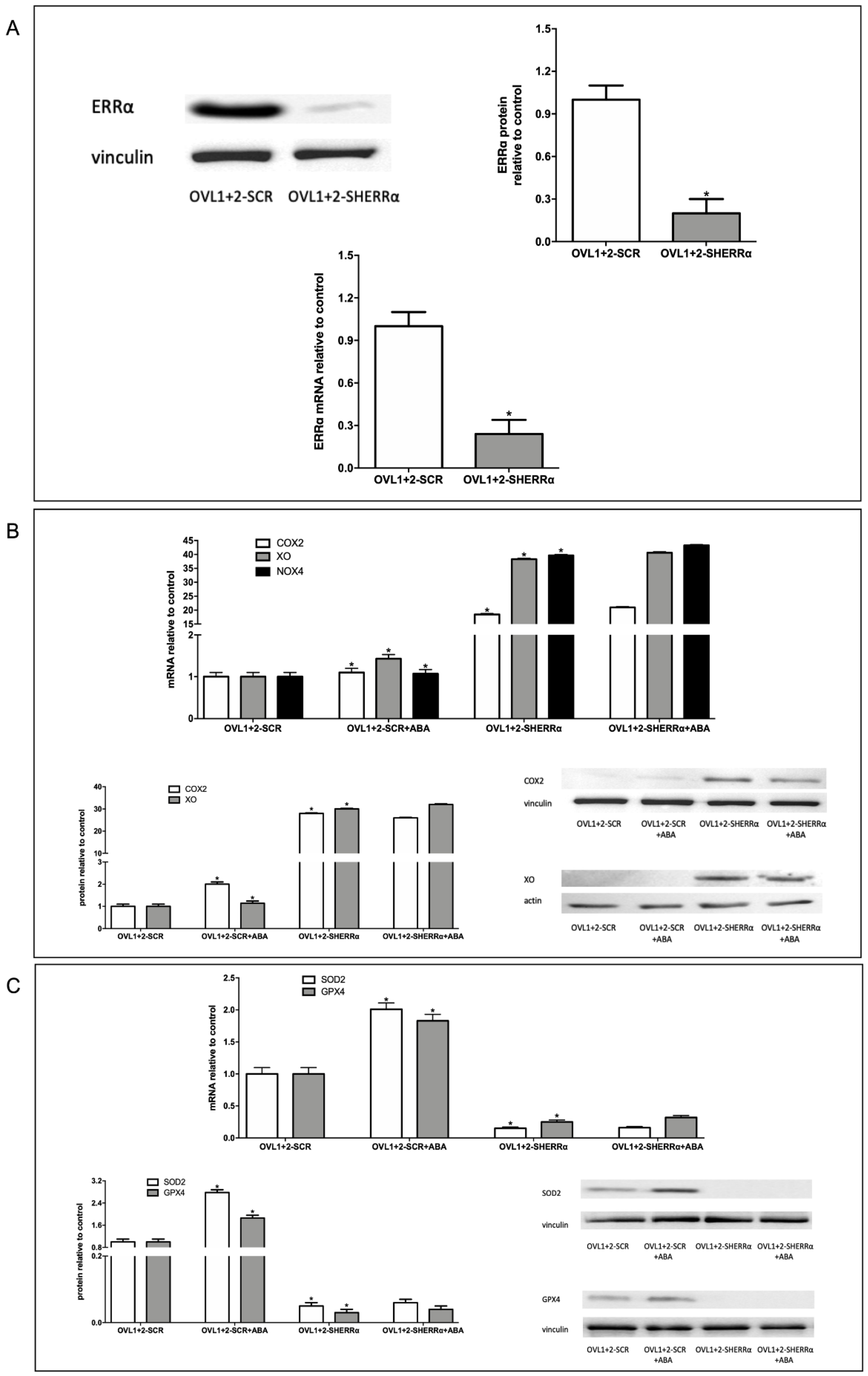
Figure 6.
ERRα silencing increases ROS production in LANCL1/2-overexpressing H9c2. (A) Intracellular ROS production measured by DCF fluorimetric analysis in H9c2 cells. Results are expressed as fluorescence intensity in arbitrary units with respect to OVL1+2-SCR cells. (B) Mitochondrial superoxide anions were detected on MitoSOX-loaded cells by confocal microscopy in H9c2 cells overexpressing LANCL1 and LANCL2 proteins and silenced (OVL1+2-SHERRα) or not (OVL1+2) for ERRα and incubated in the absence or in the presence of 100 nM ABA for 4 hours. Upper panel, representative confocal microscopy of the cells; lower panel, histogram summarized quantitative data of the mean±SD of three independent experiments. *p<0.05 relative to untreated OVL1+2-SCR cells by unpaired t test.
Figure 6.
ERRα silencing increases ROS production in LANCL1/2-overexpressing H9c2. (A) Intracellular ROS production measured by DCF fluorimetric analysis in H9c2 cells. Results are expressed as fluorescence intensity in arbitrary units with respect to OVL1+2-SCR cells. (B) Mitochondrial superoxide anions were detected on MitoSOX-loaded cells by confocal microscopy in H9c2 cells overexpressing LANCL1 and LANCL2 proteins and silenced (OVL1+2-SHERRα) or not (OVL1+2) for ERRα and incubated in the absence or in the presence of 100 nM ABA for 4 hours. Upper panel, representative confocal microscopy of the cells; lower panel, histogram summarized quantitative data of the mean±SD of three independent experiments. *p<0.05 relative to untreated OVL1+2-SCR cells by unpaired t test.
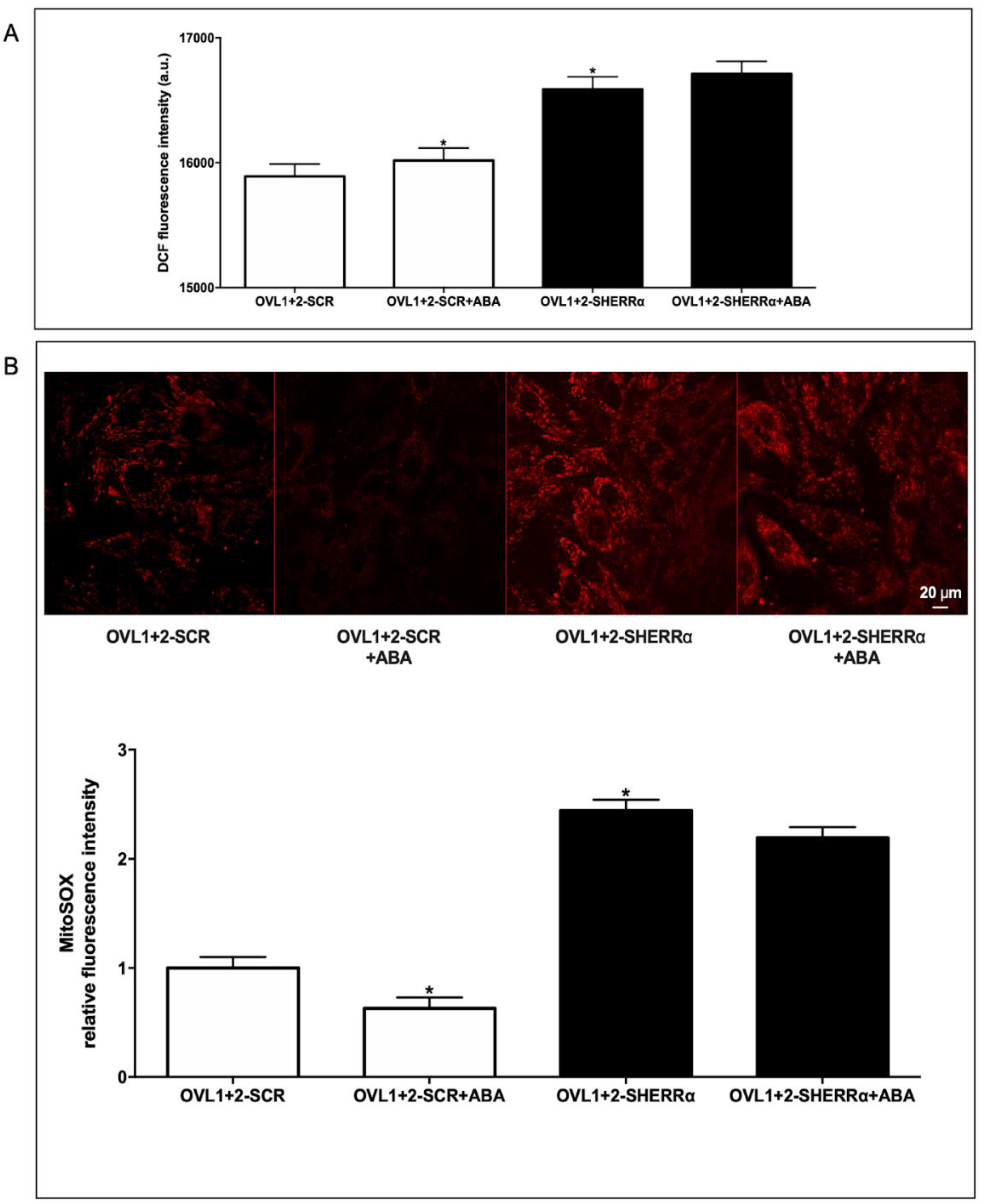

Table 1.
Primer sequences used to amplify rat target genes.
| Rat genes | Accession N. | Forward Primer 5’-3’ | Reverse Primer 5’-3’ |
|---|---|---|---|
| Lancl1 | NM_053723 | TCTTGCTCCTCATCCTGCTCATC | CACTGTACTCGCCGAAGGTCTC |
| Lancl2 | NM_001014187 | GGTGCCACGGTGCTCCAG | CCTCGCTGCCAAATCACATCAC |
| Sod2 | NM_017051 | TAAGGGTGGTGGAGAACCCA | ACCTTGGACTCCCACAGACA |
| Nox4 | NM_053524 | CTGTACAACCAAGGGCCAGA | GCTCTGCTCAAACACAATCCT |
| Gpx4 | NM_017165 | CCGTCTGAGCCGCTTATTGA | AATCATCGCGGGATGCACA |
| Cox2 | S67722 | GTGAAAACTGTACTACGCCGAG | TACTGTGTTTGGGGTGGGCT |
| Xor | NM_017154 | TCCCTGCGTTTGGTAGCATC | CCAGGAAAAGAGGTGGCTCC |
| Hprt1 | NM_012583 | TTGGTCAAGCAGTACAGCCC | TGGCCTGTATCCAACACTTCG |
Table 2.
Primary and secondary antibodies used for Western blot.
| Primary Antibody | Host | Concentrations | Manufacturer |
|---|---|---|---|
| Anti-LANCL1 | Rabbit | 1:250 | Novus Biologicals |
| Anti-LANCL2 | Mouse | 1:1000 | Reference [29] |
| Anti-XO | Mouse | 1:100 | Santa Cruz Biotechnology Inc., California |
| Anti-COX2 | Goat | 1:200 | Santa Cruz Biotechnology Inc., California |
| Anti-SOD2 | Rabbit | 1:5000 | Abcam |
| Anti-GPX4 | Mouse | 1:100 | Santa Cruz Biotechnology Inc., California |
| Anti-ERRα | Mouse | 1:200 | Santa Cruz Biotechnology Inc., California |
| Anti-vinculin | Rabbit | 1:1000 | Cell Signaling Technology, Danvers, MA |
| Anti-Actin | Mouse | 1:1000 | Santa Cruz Biotechnology Inc., California |
| Secondary Antibody | Concentrations | Manufacturer | |
| Anti-Mouse | 1:2000 | Santa Cruz Biotechnology Inc., California | |
| Anti-Rabbit | 1:1000 | Santa Cruz Biotechnology Inc., California | |
| Anti-Goat | 1:1000 | Santa Cruz Biotechnology Inc., California | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



