
Submitted:
21 July 2024
Posted:
23 July 2024
You are already at the latest version
Abstract
BReast CAncer gene 1 (BRCA1) and BReast CAncer gene 2 (BRCA2) encode for tumor suppressor proteins which are critical regulators of the homologous recombination (HR) pathway, the most precise and important DNA damage response mechanism. Dysfunctional HR proteins cannot repair double-stranded DNA breaks in mammalian cells, a situation called HR deficiency. Since their identification, pathogenic variants and other alterations of BRCA1 and BRCA2 genes have been associated with an increased risk of developing mainly breast and ovarian cancer. Interestingly, HR deficiency is also detected in tumors not carrying BRCA1/2 mutations, a condition termed “BRCAness”. One of the main mechanisms causing the BRCAness phenotype is the methylation of the BRCA1/2 promoters and this epigenetic modification is associated with carcinogenesis and poor prognosis mainly among patients with breast and ovarian cancer. BRCA1 promoter methylation has been suggested as an emerging biomarker of great predictive significance, especially concerning Poly (ADP-ribose) Polymerase inhibitors (PARP inhibitor-PARPi) responsiveness, along with or beyond BRCA1/2 mutations. However, as its clinical exploitation is still insufficient, the impact of BRCA1/2 promoter methylation status needs to be further evaluated. The current review aims to gather the latest findings about the mechanisms that underline BRCA1/2 function as well as the molecular characteristics of tumors associated with BRCA1/2 defects, by focusing on DNA methylation. Furthermore, we critically analyze their translational meaning and the validity of BRCA methylation biomarkers in predicting treatment response and we suggest a diagnostic pipeline that could be implemented in liquid biopsy to aid precision pharmacotherapy in BRCA-associated tumors.
Keywords:
BRCA1
; BRCA2
; promoter methylation
; breast cancer
; ovarian
; cancer
; liquid biopsy
; PARP inhibitors
Introduction
BReast CAncer gene 1 (BRCA1) and BReast CAncer gene 2 (BRCA2) encode for proteins that are well-known mediators of DNA damage response and particularly of double-strand breaks (DSBs) through homologous recombination (HR) [1,2]. Since the discovery of BRCA1 and BRCA2 genes in the early 1990s, it was demonstrated that individuals carrying germline BRCA1/2 mutations had a much higher lifetime risk of developing a malignancy such as Breast Cancer (BrCa), Ovarian Cancer (OvCa), Prostate Cancer (PrCa), and Pancreatic Cancer (PaCa) compared to the general population [3,4,5,6,7]. Interestingly, the risk probability of carcinogenesis varies and depends on the type and position of the actual mutation within the BRCA1/2 genes [8,9]. While the correlation between BRCA1/2 mutations and higher risk for tumorigenesis is well-established, the conclusions regarding impact on the survival are still under debate and characterized by conflicting results [10,11,12,13,14]. Later, the major importance of BRCA1 promoter methylation was also highlighted in different types of cancer as it represents an alternative silencing mechanism of the BRCA1 gene [15,16,17]. In general, aberrant epigenetic regulation affecting gene expression independently of DNA sequence is very common in cancer [18]. Specifically, hypermethylation of the 5′ promoter region of genes is a frequent epigenetic event in cancer cells leading generally to gene silencing [19,20]. Interestingly, BRCA1 promoter methylation was identified only in tumor tissue, indicating its potential oncogenic role [15,16]. Different clinical studies have demonstrated that patients with BrCa, OvCa, PrCa, and PaCa harboring BRCA1/2 mutations or other aberrations leading to malfunction could receive clinical benefit with the use of PARPi), such as olaparib and rucaparib, thus succeeding a major advance of precision medicine for these tumor types [21,22,23,24,25,26,27]. Today, additional pieces of clinical research work have shown the BRCA1/2 impact on a person’s lifetime risk of developing specific types of cancer and highlight the potential of the aberrant methylation of these genes as prognostic and predictive biomarkers [28,29,30,31]. Therefore, on the eve of precision medicine, the understanding of the tumors with BRCA1/2 aberrations and their distinct traits remains of utmost significance.
BRCA1/2 molecular mechanism of DNA damage response during Homologous Recombination
BRCA1 and BRCA2 are proteins with a critical role in maintaining genomic stability by responding to Double-Strand Breaks (DSBs) through the HR pathway [1,32]. It is of note, that BRCA1 targets effectively every DSB through HR1. Repair through HR takes place in the late S phase and G2 of the cell cycle [33]. The ATR and ATM kinases recognize this DNA damage and initiate the repair process by phosphorylating downstream DNA repair-related targets such as BRCA1 [34]. BRCA1 is a multifunctional nuclear phosphoprotein composed of diverse domains such as BRCA C-terminal (BRCT) domain, which participates in many biological processes [1,35]. HR is considered an error-free DNA damage response mechanism and is mediated by BRCA1/2 and other effectors, as follows: BRCA1 binds to the DSBs through a protein complex composed of Mre11, Rad50, and NBS1 (MRN complex) as well as CtIP [36]. Then, this BRCA1-containing multi-protein complex promotes DNA resection at the 5’ end of DSBs, creating single-strand DNA (ssDNA) [37]. Then, BRCA1 employs Rad51, an important factor with recombinase function, through its interaction with BRCA2 and PALB2 and drives it to the ssDNA, where it takes place the HR process [38]. A schematic representation of the main events of HR is illustrated in Figure 1.
DSB: Double-Strand Breaks (DSB)
BRCA1/2-mutated cancers
BRCA1-mutated tumors include all tumors which exhibit a pathogenic mutation and not a Variant of Unknown Significance (VUS) in the BRCA1 gene [39]. Mutations in BRCA1 gene are detected in different cancers, such as in BrCa (about 5% to 10% of all cases), OvCa (about 20% of cases), PaCa (about 5% to 10% of all cases), and to a lesser extent in PrCa (about 1% to 5% of all cases) [40,41,42,43,44,45]. In clinicopathological settings, these tumors display some distinct features: BRCA1-mutated BrCa is more often associated with the basal-like triple-negative phenotype (ER-/PR-/HER2-), mutated p53, immune cell infiltration (mainly T-cell lymphocytes), whereas BRCA2-mutated BrCa presents the following features: luminal type, ER+/PR+/HER2- profile, intense immunogenicity and better survival rates [39,46,47,48]. Apart from mutations, other genetic aberrations of BRCA1/2 take an active role in carcinogenesis. For example, researchers analyzed 36 Formalin-Fixed Paraffin-Embedded (FFPE) OvCa samples by next-generation sequencing (NGS) and found 15 BRCA1 and 12 BRCA2 variants as well as important loss-of-function due to CNV of BRCA1/2 genes [49].
Loss of heterozygosity (LOH) is also a key concept in tumorigenesis. It refers to the loss of an allele, usually through a mutation, and then the loss of the second allele due to genetic imbalance/rearrangements, epigenetic regulation, or other mechanisms [50]. LOH is strongly correlated to BRCA1/2 status as it was found to be relatively frequent in BrCa and OvCa bearing BRCA1/2 mutations [51]. Moreover, BRCA1 mutation carriers presented BRCA1 promoter methylation and to a great extent exhibited also LOH. In BRCA1/2 mutation carriers, LOH is associated with better survival rates and therapeutic implications, as the absence of BRCA1/2 function due to LOH renders tumors sensitive to PARPi and mainly to platins [51], as LOH is necessary for tumor sensitivity to platins and LOH absence is connected with a worse prognosis under this type of treatment [51].
BRCA1-like cancers
Following the establishment of the pivotal role of BRCA1/2 mutations in carcinogenesis, another emerging group of tumors was associated with BRCA1 dysfunction, the so-called BRCA1-like (or BRCAness) tumors, which do not exhibit a distinct mutation in the BRCA1 gene but share altogether common traits [52,53]. BRCA1-like tumors are HR deficient and present chromosomal breaks, DNA methylation, copy number variations (CNV), and genomic instability [52,54]. Recent studies focus on describing every aspect of the BRCA1-like tumors, in order to identify cancer subgroups with distinct characteristics rendering them candidates for efficient therapeutic strategies [48,54].
A broad spectrum of DNA damage response genes directly or indirectly linked to HR status has been identified including ATM, STK11, TP53, PTEN, CDH1, CHEK2, BARD1, BRIP1, MRE11, RAD50, NBS1, RAD51C/D, ATR, BAP1, BLM, CDK12, FANCA, FANCC, FANCD2, KRAS, and PALB2 [3,53,55]. However, only a few of the aforementioned genes are found often mutated, such as TP53, which is mutated in 84% of all BRCA1-like tumors and could serve as a valuable biomarker for stratifying BRCA1-like tumors [48]. Takamatsu et al, showed that BRCA1/2 wild-type cancers which present alteration in other HR genes associated with elevated genomic scar scores (model predicting homologous recombination deficiency). This score differed significantly by sex and the presence of somatic TP53 mutations and was associated with HR deficiency and
treatment response to DNA-damaging agents [56]. Alternatively, the evaluation of foci formation (a biomarker of HR repair) of 4 key HR proteins (BRCA1, Rad51, γH2AX and 53BP1) on DNA is recommended in order to detect possible HR deficiency and BRCA1-like tumors [57]. As mentioned above, BRCA1 and Rad51 are key mediators of HR and thus their foci formation is present in HR-proficient cells, whereas γH2AX and 53BP1 as conventional DNA damage markers build foci in HR-deficient cells [58]. Interestingly, researchers proved that the positive BRCA1 and RAD51 foci formation is associated with non-response to olaparib therapy in a study featuring Patient Derived Xenograft (PDX)-derived Triple Negative Breast Cancer (TNBC) samples with BRCA1/2 defect and could be used as a predictive marker in the TNBC [59]. The above studies, pointing out the importance of analyzing a panel of HR genes to identify HR deficiency. Also, identifying a BRCA1-like tumor and distinguishing it from a BRCA1-mutated tumor is not as simple as anticipated on the genetic level, making it clear that a multidimensional approach would be more suitable in studying BRCA1-like tumors.
BRCA1/2 promoter methylation in cancers
It is widely accepted that aberrant gene promoter methylation represents an epigenetic event exhibiting an oncogenic role by repressing gene expression in numerous cancers [60,61,62,63]. Specifically, locus-specific hypermethylation takes place on sites rich in CGs of the promoter region of tumor suppressor genes such as BRCA1, leading to BRCA1 transcripts and BRCA1 protein levels downregulation [64]. In ΒrCa tissues, BRCA1 promoter hypermethylation has been identified in 9 to 24% of all cases [65,66]. In particular, the prevalence of BRCA1 promoter methylation is increased in TNBC [67,68,69]. In general, an individual with BRCA1 promoter methylation, an event being particularly encountered in East Asia than Caucasians, has a 4.6 higher risk of developing BrCa than baseline, according to a meta-analysis [28] including 19,084 individuals, which associated BRCA1 promoter methylation with BrCa occurrence, recurrence, prognosis, and therapy response [28,70]. BRCA1 promoter methylation was present in 44.4% of malignant and 9.7% of normal tissues [28]. Despite the strong evidence that hypermethylation of BRCA1 promoter is detected mainly in cancer tissue, several studies presented contradictory results, suggesting that BRCA1 promoter methylation levels in normal tissue might be equal to or exceed the methylation levels of cancer tissues [71,72,73].
On the other hand, BRCA2 promoter hypermethylation is rarely encountered in BrCa and OvCa and no statistically significant correlation has been observed to clinical end-points [28], whereas in ovarian tumor samples, BRCA2 promoter methylation was confirmed in only 4.6% of the cases [31].
From a technical point of view, the overall approach for the quantification of BRCA1 promoter methylation differs between studies likely due to the determination of different cut-offs, different handling and pre-analytical procedures, lack of a common validation assay and quality of the biomaterial, eventually leading to discrepancies in calling a sample hypermethylated or not [74]. For example, in one study, 5% of the TNBC tissue samples showed methylation levels over 80% and were classified as high-methylated while 25% of them demonstrated methylation levels between 30% and 80% respectively, classified as low-methylated [75]. In another study focusing on OvCa tissues, researchers considered as a cut-off value the 15% methylation for calling a sample methylated [31]. There are also different methodologies to determine methylation (pyrosequencing, Methylation Specific PCR, restriction enzyme-based methods, droplet digital PCR, Genome-Wide Methylation Assays) and therefore the results have to be interpreted according to the used assay to avoid discrepant results between studies [31,64]. It is clear that BRCA1 promoter methylation should be examined quantitatively and in relation to methylation zygosity, as samples, many times are misidentified as hypermethylated without adequate methylation levels [64]. Methylation zygosity describes the methylation status of all epialleles (alleles that are variably expressed due to epigenetic modifications). ‘Homozygous methylation’ refers to the situation when all epialleles in a cell have highly methylated promoters resulting in gene silence. ‘Heterozygous methylation’ describes a mix of highly methylated and unmethylated epialleles coexisting within each cell. In these cells, gene expression is active due to the presence of unmethylated epialleles, despite the presence of highly methylated epialleles [76]. An important factor that affects the methylation rate determination is neoplastic cellularity. Tumor cells exhibit drastically different methylation levels thus, sufficient tumor cellularity will lead to higher mean methylation in cancerous in relation to healthy tissue [29,31,64]. Collectively, for what is concerned with measuring BRCA1 methylation, for valid conclusions to be drawn, the establishment of a widely accepted unified analytical procedure is of utmost importance.
BRCA1/2 promoter methylation in different cancer types
The following section will provide insight into current studies linking the BRCA1/2 promoter methylation status and other BRCA1/2-related genetic modifications to certain cancer types. Data from major studies assessing BRCA1 promoter methylation levels among patients with different types of cancer have been included, to frame the whole spectrum of BRCA1 promoter methylation applications in clinical settings. Table 1 presents the percentages of BRCA1 methylation reported in BrCa, OvCa, PrCa, and PaCa and correlations that have been made with the disease state.
BrCa
BrCa is the most frequently diagnosed malignancy in women globally. In addition to the study mentioned above [65], a meta-analysis by Wu et al. featuring data from 3,205 women suffering from BrCa, reported that BRCA1 methylation in tumor tissues was statistically significantly correlated to poor prognosis in terms of overall survival [87]. Interestingly, the researchers also concluded that the handling and storage of cancerous tissue could affect the tissue quality influencing the methylation results [87]. In another study by Chen et al., 139/536 (26.0%) tumor samples deriving from patients with sporadic BrCa exhibited BRCA1 promoter methylation. Interestingly, the scientists observed a worse 5-year Disease Free Survival (DFS) for patients bearing tumors with BRCA1 methylation in a statistically significant manner [78]. A meta-analysis in patients with BrCa showed that BRCA1 promoter methylation status was similar between tumor tissue and peripheral blood cells, thus encouraging its potential use as a blood-based biomarker [28]. However, a study that analyzed BRCA1 methylation in the blood of early BrCa in younger patients found that only 2 out of 154 blood cell samples presented hypermethylation of BRCA1 promoter [88]. According to these findings, someone can speculate that BRCA1 promoter methylation is a rare event in the early onset of BrCa, but more studies are needed for definite conclusions to be drawn. On the other hand, BRCA2 methylation has a very low incidence, about 4% and no correlation was observed with BrCa, according to a meta-analysis [28]. TNBC is a subtype of BrCa lacking the ER, PR, and HER-2 receptors, and thus not responding to hormonal therapy (like tamoxifen or aromatase inhibitors) or therapies that target HER2 receptors (like Herceptin) [89]. TNBC accounts for about 10% to 20% of all BrCa cases and may be either hereditary or sporadic [90]. TNBC is stimulated by mechanisms, such as point mutations, large rearrangements, and gene promoter methylation, and interestingly shares the same clinicopathological characteristics with the BRCA1-mutated tumors [28,67]. Multiple studies confirmed that BRCA1 promoter methylation and BRCA1 mutation status are almost mutually exclusive, thus tumors featuring BRCA1 promoter methylation are not linked to BRCA1 gene mutations, although there are some rare exceptions observed [28,29,31,43,67,68,69,91,92,93]. Interestingly, according to a study, 62% of BRCA1-mutated and 50% of BRCA1 promoter methylated cancers appear to be TNBC, whereas 40% to 70% of TNBC is estimated to be HR deficient [68]. Another study analyzed 237 TNBC tissues identifying hypermethylation of BRCA1 promoter in the 57/237 (24.1%) of samples [66]. Interestingly, 89.5% of the hypermethylated cases harbored concurrent LOH of BRCA1 and patients with TNBC harboring BRCA1 promoter methylation presented a significantly longer DFS than non-altered patients [66]. An immense potential of BRCA1 methylation as an early biomarker for TNBC (also HGSOC), was highlighted in a study showing that BRCA1 promoter methylation aberrations can be detected in white blood cells almost 5 years earlier than usually expected, paving the way for timely interventions and a better therapeutic outcome [69].
It is clear that BRCA1 promoter methylation is a strong candidate both as a prognostic and a predictive biomarker; nevertheless, intratumor heterogeneity and differences in epialleles render BRCA1 promoter methylation as a marker only partially effective. It is well-known that the dynamic evolution of a tumor leads to different tumor cell subpopulations with distinct genetic, epigenetic, and phenotypic traits. The different epialleles in these subpopulations could determine the response to treatment as in the case of BRCA1 mutations. Scientists now focus their attempts on deep sequencing to catch all sample epialleles. In a relevant study, researchers using bisulfite sequencing found lower methylation in epialleles of core breast tumors than in tumor periphery, [94]. These methylation differences were rendered to the hypoxic microenvironment of the tumor’s, core leading to this heterogeneous phenotype; such tumor biology aspects need to be considered for developing effective treatment schemes [94]. On the other hand, the combination of the BRCA1 promoter methylation status with other markers has been used to assess prognosis and therapy response with more accuracy. In TNBC, researchers revealed that the combination of low pRb expression levels, high p16 expression levels, PTEN absence, and BRCA1 promoter methylation exhibited a similar phenotype to BRCA1-mutated tumors [70].
Collectively, BRCA1 promoter methylation is detected frequently in BrCa, especially in TNBC, and has been associated with survival and other prognostic and therapy response end-points. Further studies analyzing all epialleles at a cellular level and/or combined with additional markers are awaited towards the establishment of BRCA1 promoter methylation as a useful tool in the clinical management of BrCa.
OvCa
Although first identified in breast, BRCA1 mutations and other gene aberrations were soon shown to have a significant role also in OvCa [95]. The presence of germline BRCA1 and BRCA2 mutations in patients with OvCa ranges from 5% to 20%, also somatic mutations are rare (2% and 8%, respectively) [95]. OvCa is the second cancer type that has been extensively studied as regards to BRCA1 promoter methylation status. A recent meta-analysis of 15 studies concluded that BRCA1 promoter methylation was present in 430/2636 tumors (16.3%). However, methylation percentages were not consistent between studies, ranging from 6.2% to 73.7% [43], and this is probably attributed similarly to breast cancer to variations in analytical methods and different methylation cut-offs used in each study. Nevertheless, BRCA1 promoter-methylated tumors share similar clinicopathological characteristics with the BRCA1-mutated as they are associated with younger age and advanced disease but no correlation with survival or platinum sensitivity has been reported [43]. In general, studies are not in agreement regarding a possible correlation between BRCA1 methylation and survival [79,80,81]. OvCa patients with homozygous BRCA1 promoter methylation showed higher PFS than patients bearing BRCA1-mutated tumours [43,49]. Another study showing BRCA1 promoter hypermethylation in 17/88 (19.3%) OvCa and BRCA2 methylation in 4/86 (4.6%) reported no correlation with clinicopathological characteristics (age, stage, histology type) [31]. Interestingly, BRCA1/2 promoter methylation is never observed in non-neoplastic ovarian tissue at any histological type, confirming its cancer-specific role [31].
HGSOC, a most lethal OvCa subtype accounting for 70% to 80% of OvCa cases is linked to rapid intraperitoneal spread [96]. The majority of BRCA1 promoter methylation cases concern younger patients with HGSOC of advanced stage [43]. A study including 172 HGSOC tissues, concluded that the combined examination of BRCA1/2 sequencing, CNVs, and methylation could lead to a more accurate diagnosis of “BRCAness” phenotype, with an estimated Area Under the Curve (AUC) of 0.77 and accuracy of 0.75, thus worthy to be validated in bigger cohorts of patients [49]. Interestingly, another study using HGSOC-derived PDX models harboring BRCA1 mutations showed a response to rucaparib and so did two chemo-naive HGSOC-PDX models with homozygous BRCA1 methylation [97]. Moreover, the donor-patients responded to rucaparib as well [97]. On the other hand, two PDX models with heterozygous BRCA1 methylation presented some BRCA1 mRNA and protein expressions and failed to respond to the rucaparib, suggesting that it is homozygous BRCA1 methylation that predicts PARPi sensitivity [97]. The above results again highlight the significance of assessing BRCA1 methylation zygosity very carefully to predict clinical outcomes. The zygosity status is thus considered an emerging factor of clinical significance to support decisions for different therapeutic strategies [29,49].
Collectively, these results point out a potential predictive and to a lesser extent prognostic role for BRCA gene methylation in OvCa. Survival rates in relation to BRCA1 methylation should be further studied for conclusions to be drawn. For sure, a determining factor is the quantitative analysis in terms of methylation zygocity as it is of utmost importance for guiding treatment options.
Prostate Cancer (PrCa)
PrCa is the most frequent cancer in men. Although the majority of PrCa cases present an indolent clinical course, PrCa remains a leading cause of cancer-related deaths [98]. Germline mutations in BRCA1/2 genes increase significantly the risk of developing PrCa. Although BRCA2 mutations have been found only in 1-3% of cases, BRCA2 mutation carriers are two-fold to four-fold more likely to develop an aggressive tumor at a younger age compared to the general population [99]. Genetic alterations affecting BRCA1 gene and representing part of BRCAness phenotype also seem to play a role in PrCa development and metastasis [100]. In PrCa, BRCA1 promoter methylation status has not been considered of the same clinical importance as in BrCa and OvCa, as there are controversial results between studies. BRCA1 promoter methylation was absent in all of the 31 prostate cancer samples examined, although other BRCA1 aberrations, such as BRCA1 imbalance, could bear some value in evaluating PrCa prognosis [82]. Another study examined BRCA1 promoter methylation both in non-malignant and malignant tissues, reporting contrasting results; Specifically, BRCA1 promoter methylation was observed in all malignant tissues (prostate intraepithelial neoplasia, peri-tumor tissue, and adenocarcinoma) but also in 15/17 normal samples [17]. Clearly, further studies are required to enlighten the topic and reveal any significance.
Pancreatic Cancer (PaCa)
Generally, PaCa is characterized by poor prognosis [101]. Pancreatic Ductal Adenocarcinoma (PDAC), the predominant form of PaCa is a highly aggressive tumor with rising incidence and the lowest survival rate amongst all the major cancers. Germline BRCA1/2 mutations are detected in approximately 5–10% of cases of hereditary PDAC and approximately 3% of cases of sporadic PDAC [45]. Similarly, with PrCa, BRCA2 mutations seem to be associated with an increased risk of PDAC development [45]. Regarding BRCA mutations and survival, the few studies exploring possible associations have presented controversial findings [45,102,103,104]. Moreover, the findings supporting the role of BRCA1 promoter methylation in PaCa are not conclusive yet. Indeed, Peng et al. examined surgical samples of PDAC, reporting BRCA1 promoter methylation in more than half of the cancerous samples (60.3%) [84]. However, Zhou et al. evaluated the promoter methylation status of BRCA1 and BRCA2 in the peripheral blood lymphocytes of 655 patients suffering from PaCa and reported BRCA1 promoter methylation levels ranging from 0.0% to 3.3%, and the BRCA2 from 0.0% to 7.6%. As the mean values were extremely low (0.3% and 0.1% respectively), the researchers considered the occurrence of BRCA1 and BRCA2 promoter methylation in PaCa as highly unlikely [83]. Abdallah et al. assessed the promoter methylation levels of BRCA1 in 121 FFPE PDAC samples by using different analytical methods to exclude possible low sensitivity and observed no methylation in any of the PDAC samples [85]. In 2022, Zhen-Lin et al. examined tissue samples from patients with PDAC and reached similar conclusions. The mean BRCA1 promoter methylation levels were found to be low (3.62%). To ensure the results, an additional detection method was used by which the unmethylated status of BRCA1 promoter was confirmed. Thus, they concluded in concordance with previous studies that BRCA1 promoter methylation was rather unusual [86].
BRCA1/2 methylation in liquid biopsy as a predictive biomarker
The emergence of liquid biopsy has revolutionized clinical oncology, introducing an alternative to traditional tissue sampling for exploring genetic aberrations and dynamic changes in the tumor [105,106,107,108]. Some of its most significant advantages are its non-invasive character and the powerful potential for effective disease monitoring by repeated sampling for controlling therapy efficacy and resistance onset [109,110]. In cancer, circulating tumor DNA (ctDNA) is an important blood component released in the bloodstream by dying tumor cells, reflecting molecular patterns of the cancer cells. It is mainly comprised of around 150 bp nucleic acid fragments and because of its relatively short length, an increased tumor volume is required for accurate assessment [88,110]. The application of liquid biopsy in assessing the BRCA1 promoter methylation status is on the rise, especially in OvCa. A study evaluating BRCA1 promoter methylation status in plasma cfDNA from patients with OvCa before and during treatment observed occurrence at 60% before treatment, and a 24% epigenetic shift to the unmethylated state during treatment, which was correlated to OvCa recurrence. Researchers concluded that BRCA1 promoter methylation in cfDNA can be used as a marker for treatment monitoring (Elazezy et al. 2021). In a relevant study, researchers found cfDNA hypermethylated BRCA1 in about 57% of OvCa patients of all cancer stages, suggesting its use as a diagnostic and prognostic marker [111]. Similarly, the hypermethylation of BRCA1 and RASSF1A was detected in 68% of the tumor tissues but also in the corresponding cfDNA in all stages of OvCa, being present in the majority of early-stage OvCa cfDNAs, suggesting an early event in OvCa [112] and making BRCA1 an ideal marker for OvCa monitoring in liquid biopsy. Melnikov et al, used the methylation of a five-gene panel (BRCA1, HIC1, PAX5, PGR & THBS1) for OvCa detection in cfDNA, reaching a sensitivity of 85% and a specificity of 61% [113]. These results indicate the importance of using multiple methylation biomarkers in cfDNA to achieve maximum effectiveness in cancer detection.
As far as BrCa concerns, studies of BRCA1/2 methylation in liquid biopsy are less. Cristall et al, introduced the mDETECT method for detecting ctDNA to manage TNBC. This assay examined many common hypermethylated genome regions including BRCA1 promoter, reaching an AUC of 0.97 for detecting a tumor with a sensitivity of 93% and a specificity of 100%. Interestingly, BRCA1 promoter methylation was present in cfDNA of about 25% of TNBC cases and 5% of healthy samples [114]. Liu et al found that cfDNA methylation frequency was higher (but still low) in patients with breast ductal cancers than in healthy individuals [115]. Low cfDNA BRCA1 methylation frequency (below 5%) was also reported in BrCa by Sturgeon et al. However, BRCA1 methylation was more often present in lymph-node-positive patients [116]. According to a meta-analysis, the hypermethylation of BRCA1 in cfDNA, among other markers, was associated with poor prognosis in ER+/PR+ BrCa [117]. In a recent work by Yen et al, researchers introduced the Guardant INFINITY, a cfDNA-based test that simultaneously examined BRCA1 methylation and genomic alterations for the management of advanced BrCa. In specific, 3% of patients had germline mutations in BRCA1, BRCA2, or ATM and almost 9% of patients had methylated the BRCA1 gene. Only one patient presented concomitant methylation and mutation at the BRCA1 gene [118]. Interestingly, methylation of BRCA1 was not detected in the 3210 cancer-free samples, implying the great specificity of BRCA1 methylation as a biomarker for cancer detection and monitoring.
In PaCa, only one recent study in cfDNA is available. Unlike PaCa tissue where methylation is low, Koukaki et al identified high methylation levels of BRCA1 and BRCA2 in plasma cfDNA, ranging between 46% and 63% in a group of 105 PaCa patients, associated further with poorer survival [119]. The evaluation of CTCs, although challenging as CTCs are extremely few (1 cancer cell:10 billion healthy cells) [120] presents another liquid biopsy alternative BRCA1 loss is linked to vimentin and cytokeratin-positive CTCs, showing an EMT stimulation through BRCA1 loss [59,65]. Unfortunately, there is no available study examining the BRCA1 methylation status in CTCs. This could be due to technical reasons as CTCs counts are low. Perhaps, analysis of methylation in CTCs could be applicable in metastatic cancer where CTCs are more abundant.
Based on these limited observations presented above, the highest BRCA1/2 methylation percentages in liquid biopsy were reported in PaCa. Then, was more often detectable in OvCa than BrCa, but more studies are needed to confirm results. In OvCa, BRCA1 methylation correlated with diagnosis and treatment monitoring but in BrCa the detectable methylation was correlated to specific cancer subtypes and poor prognosis. We believe that it is of utmost importance the design of new larger liquid biopsy-based studies in those and other cancer types, such as in PrCa, to explore BRCA1/2 methylation as predictive liquid biopsy biomarkers to aid treatment decisions in a minimally invasive manner, which also allows dynamic monitoring.
BRCA1/2 methylation and treatment strategies
Through the evaluation of BRCA1 promoter methylation in tumor tissue or liquid biopsy and as this assessment becomes more concrete in terms of methylation zygosity and methylation levels, specific groups of patients are identified, who are likely to experience clinical benefit from a specific treatment strategy [31]. PARPis (including olaparib, rucaparib, veliparib, talazoparib, niraparib) are considered a primary treatment option for patients with BRCA1 mutations and especially for TNBC and HGSOC [29,30,48,49,59,69,75,121,122,123]. However, not all tumors in these cancer subtypes are sensitive to PARPi due to tumor heterogeneity [124,125]. Consequently, not all TNBC patients carrying BRCA1 Wild Type (WT) will benefit from PARPi, as much as non-TNBC patients, carrying a BRCA1 mutation [46,124]. It has been shown that PARPi is also effective in those patients presenting homozygous BRCA1 methylated tumors [39,49,75,123,126,127]. Thus, all alleles of BRCA1 must be evaluated. Homozygous BRCA1 methylation carriers (and not heterozygous) show similar treatment outcomes as BRCA1 mutation carriers [128]. In general, BRCA1-methylated tumors present similarities to BRCA1-mutated tumors as regards to the HR pathway activity but are substantially less differentiated according to their pathological traits [68,69]. A study recommends that methylation levels for multiple genes engaged in the HR pathway need to be evaluated, to recognize eligible patients for PARPi treatment [49]. Interestingly, secondary BRCA1 mutations occurring within the BRCA1 ring domain can lead to platinum and PARPi resistance [129,130]. Partially predictive for PARPi effectiveness are also the LOH status ofBRCA1/2 mutations implying a defective HR [39,75]. To identify a possible correlation between BRCA1 promoter methylation status and LOH, studies in PDX models were conducted using a suitable scoring system for measuring LOH. They confirmed that LOH is linked to homozygous BRCA1 promoter methylation that could induce sufficient HR deficiency to permit PARPi activity [29]. The truth is that heterozygous BRCA1 promoter methylation carriers cannot have a significantly improved clinical status under PARPi treatment due to remaining BRCA1 activity. It is of note, that low methylation levels may be attributed either to a monoclonal cancer with heterozygous BRCA1 promoter methylation status or a heterogenous cancer with some cells exhibiting homozygous BRCA1 promoter methylation status [51,75]. The complete or almost complete loss of BRCA1/2 system activity is a requirement for HR deficiency and thus PARPi sensitivity [51]. According to a study, BRCA1/2 deficient status and consequently HR deficiency can be determined through the absence of BRCA1 and Rad51 [59,88]. Other studies suggest the simultaneous evaluation of BRCA1 methylation and BRCA1 protein expression or PALB2 promoter methylation alone as predictive for therapy response [59,124,131,132,133].
In OvCa, patients that have BRCA1 hypermethylation are very likely to have high genomic instability, being good candidates for PARPi therapy. On the other hand, low levels of methylation were associated with poor outcomes post-platinum [79]. In a relevant study, TNB patients with BRCA1-methylated tumors were sensitive to adjuvant chemotherapy and had better survival as compared with TNB patients with BRCA1-unmethylated tumors [30]. A patient with TNBC presenting high BRCA1 promoter methylation levels and a BRCA2 VUS experienced a complete response after Olaparib/Eribulin combination treatment [75]. Rucaparib was evaluated in 9 cell lines of BrCa, OvCa and PaCa of various BRCA1/2 statuses such as methylation, LOH, and mutation [134]. Particularly, cytotoxic effect was caused in UACC3199, a BrCa cell line methylated at BRCA1 promoter, being equal to or even exceeding carboplatin efficiency. The importance of BRCA1 promoter methylation for PARPi efficiency is thereby confirmed. Furthermore, a study noted that BRCA1 and BRCA2 methylation frequencies varied between CpG sites across their promoters. Some CpG sites were methylated more frequently in BRCA1/2 mutated cancers, while others were more often methylated in sporadic carcinomas, suggesting the use of BRCA methylation as a screening test to identify patients with BRCA germline mutation or BRCAness who may benefit from therapies such as PARPi [135].
In contrast to mutations, methylation status can change due to tumor microenvironment over the lifespan of a tumor or during treatment [136]. This might lead to the emergence of PARPi treatment resistance either during treatment or at recurrence [39,43,93,137]. Retention of homozygous BRCA1 methylation, a shift to heterozygous BRCA1 methylation, or complete loss of BRCA1 methylation may be observed following chemotherapy, e.g. under cisplatin/rucaparib treatment [29]. Loss of BRCA1 promoter methylation restores BRCA1 function and thus HR activity [29,93,137], driving PARPi treatment resistance. Methylation reversion in recurrent tumors is associated with resistance and shorter PFS, as illustrated in studies of paired primary-recurrent ovarian tumors [137]. To the best of our knowledge, studies analyzing BRCA1 methylation in cfDNA in relation to treatment response are missing. Only in one recent report, researchers used methylation and mutation analysis to assess how clinical resistance to PARPis developed in a cohort of 35 metastatic BrCa bearing BRCA1/2 mutations. Guardant INFINITY (explained above) was employed to analyze tumors' DNA and corresponding cfDNA. Results showed that the most common resistance mechanism was BRCA1/2 reversion mutation and less frequent alterations in the 53BP1-Shieldin pathway [138]. Liquid biopsy seems to be also promising in PrCa, but available data concern only gene sequencing results and no methylation. In a phase II study of abiraterone acetate in chemotherapy-naive metastatic castration-resistant prostate cancer patients, the targeted sequencing of BRCA1, BRCA2, and other 11 genes in cfDNA after one cycle of treatment could be indicative of cancer prognosis and treatment response [139].
Conclusions
BRCA1 promoter methylation status is a promising predictive and prognostic biomarker in BrCa and OvCa but also in PrCa and PaCa is worthy of further attention. Apart from germline/somatic BRCA1/2 mutations, other aberrations can lead to tumors bearing similar features, a phenotype called BRCA1-like or “BRCAness”. Specifically, BRCA1 promoter methylation, a cancer-specific mechanism, accounts for most cases of BRCA1-like tumors. It has become clear from several studies that to predict treatment response in PARPi, BRCA1 promoter methylation needs to be assessed quantitatively, both concerning methylation levels and in terms of methylation zygosity. This is why some researchers point out the term hypermethylation, thus showing that methylation levels must exceed a certain cut-off, to be of clinical, prognostic, or therapeutic significance. A combination of a comprehensive evaluation of BRCA1 methylation, Rad51 foci formation, and BRCA1 protein expression analysis in tumor samples is considered predictive for “BRCAness”, although other genes may be of significance as well, e.g. PTEN. Currently, liquid biopsy as a cancer monitoring tool has attracted particular interest in clinical oncology. Evaluating BRCA1/2 in tumor-derived material in the blood can demonstrate an early diagnosis and predict therapy response thus, leading to personalized solutions for effective treatment. The analysis of BRCA1/2 methylation in liquid biopsy could reveal how methylation patterns are influenced by cancer evolution and treatment and moreover, define patient subgroups at different time-points which may benefit from PARPi. In Figure 2, we suggest a diagnostic pipeline that could be implemented in liquid biopsy to aid precision pharmacotherapy in BRCA-associated tumors. PARPi is a relatively new therapy with a particular effect in tumors with identified BRCA1/2 or HR deficiency. PARPi therapy is often combined with other chemotherapy agents and stands in the epicenter targeting the underlying molecular mechanisms. As genetic testing becomes less expensive and more comprehensive, validation, optimization, and unifying of assays analyzing BRCA1/2 methylation alone or combined with other biomarkers in a clinical setting are expected to change the scenery in prognosis and predicting treatment response in multiple cancer types.
References
- Roy, R.; Chun, J.; Powell, S.N. BRCA1 and BRCA2: different roles in a common pathway of genome protection. Nat. Rev. Cancer 2011, 12, 68–78. [Google Scholar] [CrossRef] [PubMed]
- Prakash, R.; Zhang, Y.; Feng, W.; Jasin, M. Homologous Recombination and Human Health: The Roles of BRCA1, BRCA2, and Associated Proteins. Cold Spring Harb. Perspect. Biol. 2015, 7, a016600–a016600. [Google Scholar] [CrossRef] [PubMed]
- Guo, M.; Wang, S.M. The BRCAness Landscape of Cancer. Cells 2022, 11, 3877. [Google Scholar] [CrossRef] [PubMed]
- Miki, Y.; Swensen, J.; Shattuck-Eidens, D.; Futreal, P.A.; Harshman, K.; Tavtigian, S.; Liu, Q.; Cochran, C.; Bennett, L.M.; Ding, W.; et al. A strong candidate for the breast and ovarian cancer susceptibility gene BRCA1. Science 1994, 266, 66–71. [Google Scholar] [CrossRef] [PubMed]
- Black, D.M.; Solomon, E. The search for the familial breast/ovarian cancer gene. Trends Genet. 1993, 9, 22–26. [Google Scholar] [CrossRef] [PubMed]
- Wooster, R.; Weber, B.L. Breast and Ovarian Cancer. New Engl. J. Med. 2003, 348, 2339–2347. [Google Scholar] [CrossRef] [PubMed]
- Pilarski, R. The Role of BRCA Testing in Hereditary Pancreatic and Prostate Cancer Families. Am. Soc. Clin. Oncol. Educ. Book 2019, 39, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Rebbeck, T.R.; Mitra, N.; Wan, F.; Sinilnikova, O.M.; Healey, S.; McGuffog, L.; Mazoyer, S.; Chenevix-Trench, G.; Easton, D.F.; Antoniou, A.C.; et al. Association of Type and Location ofBRCA1andBRCA2Mutations With Risk of Breast and Ovarian Cancer. JAMA 2015, 313, 1347–1361. [Google Scholar] [CrossRef] [PubMed]
- Kuchenbaecker, K.B.; Hopper, J.L.; Barnes, D.R.; Phillips, K.-A.; Mooij, T.M.; Roos-Blom, M.-J.; Jervis, S.; Van Leeuwen, F.E.; Milne, R.L.; Andrieu, N.; et al. Risks of Breast, Ovarian, and Contralateral Breast Cancer for BRCA1 and BRCA2 Mutation Carriers. JAMA 2017, 317, 2402–2416. [Google Scholar] [CrossRef]
- Copson, E.R.; Maishman, T.C.; Tapper, W.J.; I Cutress, R.; Greville-Heygate, S.; Altman, D.G.; Eccles, B.; Gerty, S.; Durcan, L.T.; Jones, L.; et al. Germline BRCA mutation and outcome in young-onset breast cancer (POSH): a prospective cohort study. Lancet Oncol. 2018, 19, 169–180. [Google Scholar] [CrossRef]
- Goodwin, P.J.; Phillips, K.-A.; West, D.W.; Ennis, M.; Hopper, J.L.; John, E.M.; O'Malley, F.P.; Milne, R.L.; Andrulis, I.L.; Friedlander, M.L.; et al. Breast Cancer Prognosis in BRCA1 and BRCA2 Mutation Carriers: An International Prospective Breast Cancer Family Registry Population-Based Cohort Study. J. Clin. Oncol. 2012, 30, 19–26. [Google Scholar] [CrossRef]
- Schmidt, M.K.; Van Den Broek, A.J.; Tollenaar, R.A.; Smit, V.T.; Westenend, P.; Brinkhuis, M.; Oosterhuis, W.J.W.; Wesseling, J.; Janssen-Heijnen, M.L.; Jobsen, J.J.; et al. Breast Cancer Survival of BRCA1/BRCA2 Mutation Carriers in a Hospital-Based Cohort of Young Women. J. Natl. Cancer Inst. 2017, 109. [Google Scholar] [CrossRef]
- Kurian, A.W.; Abrahamse, P.; Bondarenko, I.; Hamilton, A.S.; Deapen, D.; Gomez, S.L.; Morrow, M.; Berek, J.S.; Hofer, T.P.; Katz, S.J.; et al. Association of Genetic Testing Results With Mortality Among Women With Breast Cancer or Ovarian Cancer. JNCI J. Natl. Cancer Inst. 2021, 114, 245–253. [Google Scholar] [CrossRef]
- Bolton, K.L.; Chenevix-Trench, G.; Goh, C.; Sadetzki, S.; Ramus, S.J.; Karlan, B.Y.; Lambrechts, D.; Despierre, E.; Barrowdale, D.; McGuffog, L.; et al. Association Between BRCA1 and BRCA2 Mutations and Survival in Women With Invasive Epithelial Ovarian Cancer. JAMA 2012, 307, 382–390. [Google Scholar] [CrossRef]
- Dobrovic, A.; Simpfendorfer, D. Methylation of the BRCA1 gene in sporadic breast cancer. . 1997, 57, 3347–50. [Google Scholar]
- Catteau, A.; Harris, W.H.; Xu, C.-F.; Solomon, E. Methylation of the BRCA1 promoter region in sporadic breast and ovarian cancer: correlation with disease characteristics. Oncogene 1999, 18, 1957–1965. [Google Scholar] [CrossRef]
- Rabiau, N.; Thiam, M.O.; Satih, S.; Guy, L.; Kemeny, J.-L.; Boiteux, J.-P.; Fontana, L.; Bignon, Y.-J.; Bernard-Gallon, D. Methylation analysis of BRCA1, RASSF1, GSTP1 and EPHB2 promoters in prostate biopsies according to different degrees of malignancy. . 2009, 23, 387–91. [Google Scholar]
- Sharma, S.; Kelly, T.K.; Jones, P.A. Epigenetics in cancer. Carcinogenesis 2010, 31, 27–36. [Google Scholar] [CrossRef]
- Herman, J.G.; Baylin, S.B. Gene Silencing in Cancer in Association with Promoter Hypermethylation. New Engl. J. Med. 2003, 349, 2042–2054. [Google Scholar] [CrossRef]
- Baylin, S.B. DNA methylation and gene silencing in cancer. Nat. Clin. Pr. Oncol. 2005, 2, S4–S11. [Google Scholar] [CrossRef]
- Tutt, A.N.; Garber, J.E.; Kaufman, B.; Viale, G.; Fumagalli, D.; Rastogi, P.; Gelber, R.D.; de Azambuja, E.; Fielding, A.; Balmaña, J.; et al. Adjuvant Olaparib for Patients with BRCA1- or BRCA2-Mutated Breast Cancer. New Engl. J. Med. 2021, 384, 2394–2405. [Google Scholar] [CrossRef]
- Arora, S.; Balasubramaniam, S.; Zhang, H.; Berman, T.; Narayan, P.; Suzman, D.; Bloomquist, E.; Tang, S.; Gong, Y.; Sridhara, R.; et al. FDA Approval Summary: Olaparib Monotherapy or in Combination with Bevacizumab for the Maintenance Treatment of Patients with Advanced Ovarian Cancer. Oncol. 2020, 26, e164–e172. [Google Scholar] [CrossRef]
- Tung, N.; Garber, J.E. PARP inhibition in breast cancer: progress made and future hopes. npj Breast Cancer 2022, 8, 1–5. [Google Scholar] [CrossRef]
- Wu, Y.; Xu, S.; Cheng, S.; Yang, J.; Wang, Y. Clinical application of PARP inhibitors in ovarian cancer: from molecular mechanisms to the current status. J. Ovarian Res. 2023, 16, 1–15. [Google Scholar] [CrossRef]
- Mateo, J.; de Bono, J.S.; Fizazi, K.; Saad, F.; Shore, N.; Sandhu, S.; Chi, K.N.; Agarwal, N.; Olmos, D.; Thiery-Vuillemin, A.; et al. Olaparib for the Treatment of Patients With Metastatic Castration-Resistant Prostate Cancer and Alterations in BRCA1 and/or BRCA2 in the PROfound Trial. J. Clin. Oncol. 2024, 42, 571–583. [Google Scholar] [CrossRef]
- Fizazi, K.; Piulats, J.M.; Reaume, M.N.; Ostler, P.; McDermott, R.; Gingerich, J.R.; Pintus, E.; Sridhar, S.S.; Bambury, R.M.; Emmenegger, U.; et al. Rucaparib or Physician’s Choice in Metastatic Prostate Cancer. New Engl. J. Med. 2023, 388, 719–732. [Google Scholar] [CrossRef]
- Reiss, K.A.; Mick, R.; O'Hara, M.H.; Teitelbaum, U.; Karasic, T.B.; Schneider, C.; Cowden, S.; Southwell, T.; Romeo, J.; Izgur, N.; et al. Phase II Study of Maintenance Rucaparib in Patients With Platinum-Sensitive Advanced Pancreatic Cancer and a Pathogenic Germline or Somatic Variant in BRCA1, BRCA2, or PALB2. J. Clin. Oncol. 2021, 39, 2497–2505. [Google Scholar] [CrossRef]
- Li, S.; He, Y.; Li, C.; Liu, X.; Shen, Y.B.; Wu, Y.; Bai, N.; Li, Q.B. The association between the methylation frequency of BRCA1/2 gene promoter and occurrence and prognosis of breast carcinoma A meta-analysis. Medicine 2020, 99, e19345. [Google Scholar] [CrossRef]
- Kondrashova, O.; Topp, M.; Nesic, K.; Lieschke, E.; Ho, G.Y.; Harrell, M.I.; Zapparoli, G.V.; Hadley, A.; Holian, R.; Boehm, E.; et al. Methylation of all BRCA1 copies predicts response to the PARP inhibitor rucaparib in ovarian carcinoma. Nat. Commun. 2018, 9, 1–16. [Google Scholar] [CrossRef]
- Xu, Y.; Diao, L.; Chen, Y.; Liu, Y.; Wang, C.; Ouyang, T.; Li, J.; Wang, T.; Fan, Z.; Fan, T.; et al. Promoter methylation of BRCA1 in triple-negative breast cancer predicts sensitivity to adjuvant chemotherapy. Ann. Oncol. 2013, 24, 1498–1505. [Google Scholar] [CrossRef]
- Sahnane, N.; Carnevali, I.; Formenti, G.; Casarin, J.; Facchi, S.; Bombelli, R.; Di Lauro, E.; Memoli, D.; Salvati, A.; Rizzo, F.; et al. BRCA Methylation Testing Identifies a Subset of Ovarian Carcinomas without Germline Variants That Can Benefit from PARP Inhibitor. Int. J. Mol. Sci. 2020, 21, 9708. [Google Scholar] [CrossRef]
- Gorodetska, I.; Kozeretska, I.; Dubrovska, A. BRCA Genes: The Role in Genome Stability, Cancer Stemness and Therapy Resistance. J. Cancer 2019, 10, 2109–2127. [Google Scholar] [CrossRef]
- Moynahan, M.E.; Jasin, M. Mitotic homologous recombination maintains genomic stability and suppresses tumorigenesis. Nat. Rev. Mol. Cell Biol. 2010, 11, 196–207. [Google Scholar] [CrossRef]
- Mandel, P. & Metais, P. [Not Available]. Comptes rendus des seances de la Societe de biologie et de ses filiales 142, 241-243 (1948).
- A Clapperton, J.; A Manke, I.; Lowery, D.M.; Ho, T.; Haire, L.F.; Yaffe, M.B.; Smerdon, S.J. Structure and mechanism of BRCA1 BRCT domain recognition of phosphorylated BACH1 with implications for cancer. Nat. Struct. Mol. Biol. 2004, 11, 512–518. [Google Scholar] [CrossRef]
- Her, J.; Lee, N.S.; Kim, Y.; Kim, H. Factors forming the BRCA1-A complex orchestrate BRCA1 recruitment to the sites of DNA damage. Acta Biochim. et Biophys. Sin. 2016, 48, 658–664. [Google Scholar] [CrossRef]
- Zhao, F.; Kim, W.; Kloeber, J.A.; Lou, Z. DNA end resection and its role in DNA replication and DSB repair choice in mammalian cells. Exp. Mol. Med. 2020, 52, 1705–1714. [Google Scholar] [CrossRef]
- Zhao, W.; Steinfeld, J.B.; Liang, F.; Chen, X.; Maranon, D.G.; Ma, C.J.; Kwon, Y.; Rao, T.; Wang, W.; Sheng, C.; et al. BRCA1–BARD1 promotes RAD51-mediated homologous DNA pairing. Nature 2017, 550, 360–365. [Google Scholar] [CrossRef]
- Shah, J.B.; Pueschl, D.; Wubbenhorst, B.; Fan, M.; Pluta, J.; D’andrea, K.; Hubert, A.P.; Shilan, J.S.; Zhou, W.; Kraya, A.A.; et al. Analysis of matched primary and recurrent BRCA1/2 mutation-associated tumors identifies recurrence-specific drivers. Nat. Commun. 2022, 13, 1–19. [Google Scholar] [CrossRef]
- Fu, X.; Tan, W.; Song, Q.; Pei, H.; Li, J. BRCA1 and Breast Cancer: Molecular Mechanisms and Therapeutic Strategies. Front. Cell Dev. Biol. 2022, 10, 813457. [Google Scholar] [CrossRef]
- Petrucelli, N. , Daly, M. B. & Pal, T. in GeneReviews((R)) (eds M. P. Adam et al.) (1993).
- Casaubon, J. T. , Kashyap, S. P. in StatPearls ( 2024.
- Kalachand, R.D.; Stordal, B.; Madden, S.; Chandler, B.; Cunningham, J.; Goode, E.L.; Ruscito, I.; I Braicu, E.; Sehouli, J.; Ignatov, A.; et al. BRCA1Promoter Methylation and Clinical Outcomes in Ovarian Cancer: An Individual Patient Data Meta-Analysis. JNCI J. Natl. Cancer Inst. 2020, 112, 1190–1203. [Google Scholar] [CrossRef]
- Messina, C.; Cattrini, C.; Soldato, D.; Vallome, G.; Caffo, O.; Castro, E.; Olmos, D.; Boccardo, F.; Zanardi, E. BRCA Mutations in Prostate Cancer: Prognostic and Predictive Implications. J. Oncol. 2020, 2020, 1–7. [Google Scholar] [CrossRef]
- Lai, E.; Ziranu, P.; Spanu, D.; Dubois, M.; Pretta, A.; Tolu, S.; Camera, S.; Liscia, N.; Mariani, S.; Persano, M.; et al. BRCA-mutant pancreatic ductal adenocarcinoma. Br. J. Cancer 2021, 125, 1321–1332. [Google Scholar] [CrossRef]
- Miklikova, S.; Trnkova, L.; Plava, J.; Bohac, M.; Kuniakova, M.; Cihova, M. The Role of BRCA1/2-Mutated Tumor Microenvironment in Breast Cancer. Cancers 2021, 13, 575. [Google Scholar] [CrossRef]
- Hill, S.J.; Clark, A.P.; Silver, D.P.; Livingston, D.M. BRCA1 Pathway Function in Basal-Like Breast Cancer Cells. Mol. Cell. Biol. 2014, 34, 3828–3842. [Google Scholar] [CrossRef]
- Severson, T.M.; Peeters, J.; Majewski, I.; Michaut, M.; Bosma, A.; Schouten, P.C.; Chin, S.-F.; Pereira, B.; Goldgraben, M.A.; Bismeijer, T.; et al. BRCA1-like signature in triple negative breast cancer: Molecular and clinical characterization reveals subgroups with therapeutic potential. Mol. Oncol. 2015, 9, 1528–1538. [Google Scholar] [CrossRef]
- Aref-Eshghi, E.; McGee, J.D.; Pedro, V.P.; Kerkhof, J.; Stuart, A.; Ainsworth, P.J.; Lin, H.; Volodarsky, M.; McLachlin, C.M.; Sadikovic, B. Genetic and epigenetic profiling of BRCA1/2 in ovarian tumors reveals additive diagnostic yield and evidence of a genomic BRCA1/2 DNA methylation signature. J. Hum. Genet. 2020, 65, 865–873. [Google Scholar] [CrossRef]
- Nichols, C.A.; Gibson, W.J.; Brown, M.S.; Kosmicki, J.A.; Busanovich, J.P.; Wei, H.; Urbanski, L.M.; Curimjee, N.; Berger, A.C.; Gao, G.F.; et al. Loss of heterozygosity of essential genes represents a widespread class of potential cancer vulnerabilities. Nat. Commun. 2020, 11, 2517. [Google Scholar] [CrossRef]
- Maxwell, K.N.; Wubbenhorst, B.; Wenz, B.M.; De Sloover, D.; Pluta, J.; Emery, L.; Barrett, A.; Kraya, A.A.; Anastopoulos, I.N.; Yu, S.; et al. BRCA locus-specific loss of heterozygosity in germline BRCA1 and BRCA2 carriers. Nat. Commun. 2017, 8, 1–11. [Google Scholar] [CrossRef]
- Byrum, A.K.; Vindigni, A.; Mosammaparast, N. Defining and Modulating ‘BRCAness’. Trends Cell Biol. 2019, 29, 740–751. [Google Scholar] [CrossRef]
- Murai, J.; Pommier, Y. BRCAness, Homologous Recombination Deficiencies, and Synthetic Lethality. Cancer Res. 2023, 83, 1173–1174. [Google Scholar] [CrossRef] [PubMed]
- Schouten, P.C.; Marmé, F.; Aulmann, S.; Sinn, H.-P.; van Essen, H.F.; Ylstra, B.; Hauptmann, M.; Schneeweiss, A.; Linn, S.C. Breast Cancers with aBRCA1-like DNA Copy Number Profile Recur Less Often Than Expected after High-Dose Alkylating Chemotherapy. Clin. Cancer Res. 2015, 21, 763–770. [Google Scholar] [CrossRef] [PubMed]
- Turner, N.; Tutt, A.; Ashworth, A. Hallmarks of 'BRCAness' in sporadic cancers. Nat. Rev. Cancer 2004, 4, 814–819. [Google Scholar] [CrossRef] [PubMed]
- Takamatsu, S.; Brown, J.; Yamaguchi, K.; Hamanishi, J.; Yamanoi, K.; Takaya, H.; Kaneyasu, T.; Mori, S.; Mandai, M.; Matsumura, N. Utility of Homologous Recombination Deficiency Biomarkers Across Cancer Types. JCO Precis. Oncol. 2022, 6, e2200085. [Google Scholar] [CrossRef] [PubMed]
- Cruz, C.; Castroviejo-Bermejo, M.; Gutiérrez-Enríquez, S.; Llop-Guevara, A.; Ibrahim, Y.; Gris-Oliver, A.; Bonache, S.; Morancho, B.; Bruna, A.; Rueda, O.; et al. RAD51 foci as a functional biomarker of homologous recombination repair and PARP inhibitor resistance in germline BRCA-mutated breast cancer. Ann. Oncol. 2018, 29, 1203–1210. [Google Scholar] [CrossRef] [PubMed]
- Bunting, S.F.; Callén, E.; Wong, N.; Chen, H.-T.; Polato, F.; Gunn, A.; Bothmer, A.; Feldhahn, N.; Fernandez-Capetillo, O.; Cao, L.; et al. 53BP1 Inhibits Homologous Recombination in Brca1-Deficient Cells by Blocking Resection of DNA Breaks. Cell 2010, 141, 243–254. [Google Scholar] [CrossRef] [PubMed]
- Velazquez, C.; Orhan, E.; Tabet, I.; Fenou, L.; Orsetti, B.; Adélaïde, J.; Guille, A.; Thézénas, S.; Crapez, E.; Colombo, P.-E.; et al. BRCA1-methylated triple negative breast cancers previously exposed to neoadjuvant chemotherapy form RAD51 foci and respond poorly to olaparib. Front. Oncol. 2023, 13, 1125021. [Google Scholar] [CrossRef]
- Bouras, E.; Karakioulaki, M.; Bougioukas, K.I.; Aivaliotis, M.; Tzimagiorgis, G.; Chourdakis, M. Gene promoter methylation and cancer: An umbrella review. Gene 2019, 710, 333–340. [Google Scholar] [CrossRef] [PubMed]
- Panagopoulou, M.; Karaglani, M.; Balgkouranidou, I.; Biziota, E.; Koukaki, T.; Karamitrousis, E.; Nena, E.; Tsamardinos, I.; Kolios, G.; Lianidou, E.; et al. Circulating cell-free DNA in breast cancer: size profiling, levels, and methylation patterns lead to prognostic and predictive classifiers. Oncogene 2019, 38, 3387–3401. [Google Scholar] [CrossRef]
- Panagopoulou, M.; Fanidis, D.; Aidinis, V.; Chatzaki, E. ENPP2 Methylation in Health and Cancer. Int. J. Mol. Sci. 2021, 22, 11958. [Google Scholar] [CrossRef]
- Panagopoulou, M.; Esteller, M.; Chatzaki, E. Circulating Cell-Free DNA in Breast Cancer: Searching for Hidden Information towards Precision Medicine. Cancers 2021, 13, 728. [Google Scholar] [CrossRef] [PubMed]
- Ibragimova, I. & Cairns, P. Assays for hypermethylation of the BRCA1 gene promoter in tumor cells to predict sensitivity to PARP-inhibitor therapy. [CrossRef]
- Birgisdottir, V.; Stefansson, O.A.; Bodvarsdottir, S.K.; Hilmarsdottir, H.; Jonasson, J.G.; Eyfjord, J.E. Epigenetic silencing and deletion of the BRCA1 gene in sporadic breast cancer. Breast Cancer Res. 2006, 8, R38–R38. [Google Scholar] [CrossRef] [PubMed]
- Glodzik, D.; Bosch, A.; Hartman, J.; Aine, M.; Vallon-Christersson, J.; Reuterswärd, C.; Karlsson, A.; Mitra, S.; Niméus, E.; Holm, K.; et al. Comprehensive molecular comparison of BRCA1 hypermethylated and BRCA1 mutated triple negative breast cancers. Nat. Commun. 2020, 11, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Brianese, R.C.; Nakamura, K.D.d.M.; Almeida, F.G.d.S.R.d.; Ramalho, R.F.; Barros, B.D.d.F.; e Ferreira, E.N.; Formiga, M.N.d.C.; de Andrade, V.P.; de Lima, V.C.C.; Carraro, D.M. BRCA1 deficiency is a recurrent event in early-onset triple-negative breast cancer: a comprehensive analysis of germline mutations and somatic promoter methylation. Breast Cancer Res. Treat. 2018, 167, 803–814. [Google Scholar] [CrossRef] [PubMed]
- A Stefansson, O.; Hilmarsdottir, H.; Olafsdottir, K.; Tryggvadottir, L.; Sverrisdottir, A.; Johannsson, O.T.; Jonasson, J.G.; E Eyfjord, J.; Sigurdsson, S. BRCA1 Promoter Methylation Status in 1031 Primary Breast Cancers Predicts Favorable Outcomes Following Chemotherapy. JNCI Cancer Spectr. 2020, 4, pkz100. [Google Scholar] [CrossRef] [PubMed]
- Lønning, P.E.; Nikolaienko, O.; Pan, K.; Kurian, A.W.; Eikesdal, H.P.; Pettinger, M.; Anderson, G.L.; Prentice, R.L.; Chlebowski, R.T.; Knappskog, S. Constitutional BRCA1 Methylation and Risk of Incident Triple-Negative Breast Cancer and High-grade Serous Ovarian Cancer. JAMA Oncol. 2022, 8, 1579–1587. [Google Scholar] [CrossRef] [PubMed]
- Stefansson, O.A.; Jonasson, J.G.; Olafsdottir, K.; Hilmarsdottir, H.; Olafsdottir, G.; Esteller, M.; Johannsson, O.T.; Eyfjord, J.E. CpG island hypermethylation ofBRCA1and loss of pRb as co-occurring events in basal/triple-negative breast cancer. Epigenetics 2011, 6, 638–649. [Google Scholar] [CrossRef] [PubMed]
- Vu, T.L.; Nguyen, T.T.; Doan, V.T.H.; Vo, L.T.T. Methylation Profiles of BRCA1, RASSF1A and GSTP1 in Vietnamese Women with Breast Cancer. 19, 1887. [Google Scholar] [CrossRef]
- Parrella, P.; Poeta, M.L.; Gallo, A.P.; Prencipe, M.; Scintu, M.; Apicella, A.; Rossiello, R.; Liguoro, G.; Seripa, D.; Gravina, C.; et al. Nonrandom Distribution of Aberrant Promoter Methylation of Cancer-Related Genes in Sporadic Breast Tumors. Clin. Cancer Res. 2004, 10, 5349–5354. [Google Scholar] [CrossRef] [PubMed]
- Kontorovich, T.; Cohen, Y.; Nir, U.; Friedman, E. Promoter methylation patterns of ATM, ATR, BRCA1, BRCA2 and P53 as putative cancer risk modifiers in Jewish BRCA1/BRCA2 mutation carriers. Breast Cancer Res. Treat. 2008, 116, 195–200. [Google Scholar] [CrossRef]
- Sahnane, N.; Rivera, D.; Libera, L.; Carnevali, I.; Banelli, B.; Facchi, S.; Gismondi, V.; Paudice, M.; Cirmena, G.; Vellone, V.G.; et al. Pyrosequencing Assay for BRCA1 Methylation Analysis. J. Mol. Diagn. 2023, 25, 217–226. [Google Scholar] [CrossRef]
- Kawachi, A.; Yamashita, S.; Okochi-Takada, E.; Hirakawa, A.; Tsuda, H.; Shimomura, A.; Kojima, Y.; Yonemori, K.; Fujiwara, Y.; Kinoshita, T.; et al. BRCA1 promoter methylation in breast cancer patients is associated with response to olaparib/eribulin combination therapy. Breast Cancer Res. Treat. 2020, 181, 323–329. [Google Scholar] [CrossRef] [PubMed]
- Geissler, F.; Nesic, K.; Kondrashova, O.; Dobrovic, A.; Swisher, E.M.; Scott, C.L.; Wakefield, M.J. The role of aberrant DNA methylation in cancer initiation and clinical impacts. Ther. Adv. Med Oncol. 2024, 16. [Google Scholar] [CrossRef] [PubMed]
- Lønning, P.E.; Nikolaienko, O.; Pan, K.; Kurian, A.W.; Eikesdal, H.P.; Pettinger, M.; Anderson, G.L.; Prentice, R.L.; Chlebowski, R.T.; Knappskog, S. Constitutional BRCA1 Methylation and Risk of Incident Triple-Negative Breast Cancer and High-grade Serous Ovarian Cancer. JAMA Oncol. 2022, 8, 1579–1587. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Zhou, J.; Xu, Y.; Li, Z.; Wen, X.; Yao, L.; Xie, Y.; Deng, D. BRCA1promoter methylation associated with poor survival in Chinese patients with sporadic breast cancer. Cancer Sci. 2009, 100, 1663–1667. [Google Scholar] [CrossRef] [PubMed]
- Blanc-Durand, F.; Tang, R.; Pommier, M.; Nashvi, M.; Cotteret, S.; Genestie, C.; Le Formal, A.; Pautier, P.; Michels, J.; Kfoury, M.; et al. Clinical Relevance of BRCA1 Promoter Methylation Testing in Patients with Ovarian Cancer. Clin. Cancer Res. 2023, 29, 3124–3129. [Google Scholar] [CrossRef] [PubMed]
- Pradjatmo, H.; Dasuki, D.; Anwar, M.; Mubarika, S.; Harijadi, H. Methylation Status and Immunohistochemistry of BRCA1 in Epithelial Ovarian Cancer. Asian Pac. J. Cancer Prev. 2014, 15, 9479–9485. [Google Scholar] [CrossRef] [PubMed]
- Ruscito, I.; Dimitrova, D.; Vasconcelos, I.; Gellhaus, K.; Schwachula, T.; Bellati, F.; Zeillinger, R.; Benedetti-Panici, P.; Vergote, I.; Mahner, S.; et al. BRCA1 gene promoter methylation status in high-grade serous ovarian cancer patients – A study of the tumour Bank ovarian cancer (TOC) and ovarian cancer diagnosis consortium (OVCAD). Eur. J. Cancer 2014, 50, 2090–2098. [Google Scholar] [CrossRef] [PubMed]
- Bednarz, N.; Eltze, E.; Semjonow, A.; Rink, M.; Andreas, A.; Mulder, L.; Hannemann, J.; Fisch, M.; Pantel, K.; Weier, H.-U.G.; et al. BRCA1 Loss Preexisting in Small Subpopulations of Prostate Cancer Is Associated with Advanced Disease and Metastatic Spread to Lymph Nodes and Peripheral Blood. Clin. Cancer Res. 2010, 16, 3340–3348. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.; Porter, N.; Borges, M.; Gauthier, C.; Ferguson, L.; Huang, B.; Nanda, N.; He, J.; Laheru, D.; Hruban, R.H.; et al. Examination of ATM, BRCA1, and BRCA2 promoter methylation in patients with pancreatic cancer. Pancreatology 2021, 21, 938–941. [Google Scholar] [CrossRef]
- Peng, D.-F.; Kanai, Y.; Sawada, M.; Ushijima, S.; Hiraoka, N.; Kitazawa, S.; Hirohashi, S. DNA methylation of multiple tumor-related genes in association with overexpression of DNA methyltransferase 1 (DNMT1) during multistage carcinogenesis of the pancreas. Carcinog. 2006, 27, 1160–1168. [Google Scholar] [CrossRef]
- Abdallah, R.; Zhao, S.; Garinet, S.; Hormigos, K.; Le Corre, D.; Cros, J.; Toralla, K.P.; Bats, A.S.; Augustin, J.; Bachet, J.-B.; et al. BRCA1 and RAD51C promotor methylation in human resectable pancreatic adenocarcinoma. Clin. Res. Hepatol. Gastroenterol. 2022, 46, 101880. [Google Scholar] [CrossRef] [PubMed]
- Zheng-Lin, B.; Rainone, M.; Varghese, A.M.; Yu, K.H.; Park, W.; Berger, M.; Mehine, M.; Chou, J.; Capanu, M.; Mandelker, D.; et al. Methylation Analyses Reveal Promoter Hypermethylation as a Rare Cause of “Second Hit” in Germline BRCA1-Associated Pancreatic Ductal Adenocarcinoma. Mol. Diagn. Ther. 2022, 26, 645–653. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Wang, F.; Xu, R.; Zhang, S.; Peng, X.; Feng, Y.; Wang, J.; Lu, C. Promoter methylation of BRCA1 in the prognosis of breast cancer: a meta-analysis. Breast Cancer Res. Treat. 2013, 142, 619–627. [Google Scholar] [CrossRef] [PubMed]
- Azzollini, J.; Pesenti, C.; Pizzamiglio, S.; Fontana, L.; Guarino, C.; Peissel, B.; Plebani, M.; Tabano, S.; Sirchia, S.M.; Colapietro, P.; et al. Constitutive BRCA1 Promoter Hypermethylation Can Be a Predisposing Event in Isolated Early-Onset Breast Cancer. Cancers 2019, 11, 58. [Google Scholar] [CrossRef] [PubMed]
- de Ruijter, T.C.; Veeck, J.; de Hoon, J.P.J.; van Engeland, M.; Tjan-Heijnen, V.C. Characteristics of triple-negative breast cancer. J. Cancer Res. Clin. Oncol. 2010, 137, 183–192. [Google Scholar] [CrossRef] [PubMed]
- Almansour, N.M. Triple-Negative Breast Cancer: A Brief Review About Epidemiology, Risk Factors, Signaling Pathways, Treatment and Role of Artificial Intelligence. Front. Mol. Biosci. 2022, 9, 836417. [Google Scholar] [CrossRef]
- Bednarz-Knoll, N.; Eltze, E.; Semjonow, A.; Brandt, B. BRCAness in prostate cancer. Oncotarget 2019, 10, 2421–2422. [Google Scholar] [CrossRef] [PubMed]
- Swisher, E.M.; Lin, K.K.; Oza, A.M.; Scott, C.L.; Giordano, H.; Sun, J.; Konecny, G.E.; Coleman, R.L.; Tinker, A.V.; O’Malley, D.M.; et al. Rucaparib in relapsed, platinum-sensitive high-grade ovarian carcinoma (ARIEL2 Part 1): an international, multicentre, open-label, phase 2 trial. Lancet Oncol. 2017, 18, 75–87. [Google Scholar] [CrossRef]
- Elazezy, M.; Prieske, K.; Kluwe, L.; Oliveira-Ferrer, L.; Peine, S.; Müller, V.; Woelber, L.; Schmalfeldt, B.; Pantel, K.; Joosse, S.A. BRCA1 promoter hypermethylation on circulating tumor DNA correlates with improved survival of patients with ovarian cancer. Mol. Oncol. 2021, 15, 3615–3625. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, Y.; Huang, Y.; Chen, C.; Zhang, X.; Xing, Y.; Gu, Y.; Zhang, M.; Cai, L.; Xu, S.; et al. Intratumor heterogeneity of breast cancer detected by epialleles shows association with hypoxic microenvironment. Theranostics 2021, 11, 4403–4420. [Google Scholar] [CrossRef]
- Ashour, M.; Shafik, H.E. Frequency of germline mutations in BRCA1 and BRCA2 in ovarian cancer patients and their effect on treatment outcome. Cancer Manag. Res. 2019, ume 11, 6275–6284. [Google Scholar] [CrossRef]
- Bowtell, D.D.L. The genesis and evolution of high-grade serous ovarian cancer. Nat. Rev. Cancer 2010, 10, 803–808. [Google Scholar] [CrossRef] [PubMed]
- Nesic, K.; et al. BRCA1 secondary splice-site mutations drive exon-skipping and PARP inhibitor resistance. medRxiv : the preprint server for health sciences. [CrossRef]
- Litwin, M.S.; Tan, H.-J. The Diagnosis and Treatment of Prostate Cancer. JAMA 2017, 317, 2532–2542. [Google Scholar] [CrossRef] [PubMed]
- Rajwa, P.; Quhal, F.; Pradere, B.; Gandaglia, G.; Ploussard, G.; Leapman, M.S.; Gore, J.L.; Paradysz, A.; Tilki, D.; Merseburger, A.S.; et al. Prostate cancer risk, screening and management in patients with germline BRCA1/2 mutations. Nat. Rev. Urol. 2023, 20, 205–216. [Google Scholar] [CrossRef] [PubMed]
- Omari, A.; Nastały, P.; Stoupiec, S.; Bałabas, A.; Dąbrowska, M.; Bielińska, B.; Huss, S.; Pantel, K.; Semjonow, A.; Eltze, E.; et al. Somatic aberrations of BRCA1 gene are associated with ALDH1, EGFR, and tumor progression in prostate cancer. Int. J. Cancer 2018, 144, 607–614. [Google Scholar] [CrossRef] [PubMed]
- Bilici, A. Prognostic factors related with survival in patients with pancreatic adenocarcinoma. World J. Gastroenterol. 2014, 20, 10802–12. [Google Scholar] [CrossRef] [PubMed]
- Lal, G.; Liu, G.; Schmocker, B.; Kaurah, P.; Ozcelik, H.; A Narod, S.; Redston, M.; Gallinger, S. Inherited predisposition to pancreatic adenocarcinoma: role of family history and germ-line p16, BRCA1, and BRCA2 mutations. . 2000, 60, 409–16. [Google Scholar] [PubMed]
- Iqbal, J.; Ragone, A.; Lubinski, J.; Lynch, H.T.; Moller, P.; Ghadirian, P.; Foulkes, W.D.; Armel, S.; Eisen, A.; Neuhausen, S.L.; et al. The incidence of pancreatic cancer in BRCA1 and BRCA2 mutation carriers. Br. J. Cancer 2012, 107, 2005–2009. [Google Scholar] [CrossRef] [PubMed]
- Golan, T.; Sella, T.; O'Reilly, E.M.; Katz, M.H.G.; Epelbaum, R.; Kelsen, D.P.; Borgida, A.; Maynard, H.; Kindler, H.; Friedmen, E.; et al. Overall survival and clinical characteristics of BRCA mutation carriers with stage I/II pancreatic cancer. Br. J. Cancer 2017, 116, 697–702. [Google Scholar] [CrossRef]
- Tivey, A.; Church, M.; Rothwell, D.; Dive, C.; Cook, N. Circulating tumour DNA — looking beyond the blood. Nat. Rev. Clin. Oncol. 2022, 19, 600–612. [Google Scholar] [CrossRef]
- Panagopoulou, M.; Karaglani, M.; Balgkouranidou, I.; Pantazi, C.; Kolios, G.; Kakolyris, S.; Chatzaki, E. Circulating cell-free DNA release in vitro: kinetics, size profiling, and cancer-related gene methylation. J. Cell. Physiol. 2019, 234, 14079–14089. [Google Scholar] [CrossRef] [PubMed]
- Panagopoulou, M.; Drosouni, A.; Fanidis, D.; Karaglani, M.; Balgkouranidou, I.; Xenidis, N.; Aidinis, V.; Chatzaki, E. ENPP2 Promoter Methylation Correlates with Decreased Gene Expression in Breast Cancer: Implementation as a Liquid Biopsy Biomarker. Int. J. Mol. Sci. 2022, 23, 3717. [Google Scholar] [CrossRef] [PubMed]
- Papadakis, V.M.; Cheimonidi, C.; Panagopoulou, M.; Karaglani, M.; Apalaki, P.; Katsara, K.; Kenanakis, G.; Theodosiou, T.; Constantinidis, T.C.; Stratigi, K.; et al. Label-Free Human Disease Characterization through Circulating Cell-Free DNA Analysis Using Raman Spectroscopy. Int. J. Mol. Sci. 2023, 24, 12384. [Google Scholar] [CrossRef] [PubMed]
- Nikanjam, M.; Kato, S.; Kurzrock, R. Liquid biopsy: current technology and clinical applications. J. Hematol. Oncol. 2022, 15, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Heidrich, I.; Ačkar, L.; Mossahebi Mohammadi, P.; Pantel, K. Liquid biopsies: Potential and challenges. Int. J. Cancer 2021, 148, 528–545. [Google Scholar] [CrossRef] [PubMed]
- S, S.K.; Swamy, S.N.; Premalatha, C.S.; Pallavi, V.R.; Gawari, R. Aberrant Promoter Hypermethylation of RASSF1a and BRCA1 in Circulating Cell-Free Tumor DNA Serves as a Biomarker of Ovarian Carcinoma. Asian Pac. J. Cancer Prev. 2019, 20, 3001–3005. [Google Scholar] [CrossRef] [PubMed]
- de Caceres, I.I.; Battagli, C.; Esteller, M.; Herman, J.G.; Dulaimi, E.; Edelson, M.I.; Bergman, C.; Ehya, H.; Eisenberg, B.L.; Cairns, P. Tumor Cell-Specific BRCA1 and RASSF1A Hypermethylation in Serum, Plasma, and Peritoneal Fluid from Ovarian Cancer Patients. Cancer Res. 2004, 64, 6476–6481. [Google Scholar] [CrossRef] [PubMed]
- Melnikov, A.; Scholtens, D.; Godwin, A.; Levenson, V. Differential Methylation Profile of Ovarian Cancer in Tissues and Plasma. J. Mol. Diagn. 2009, 11, 60–65. [Google Scholar] [CrossRef] [PubMed]
- Cristall, K.; Bidard, F.-C.; Pierga, J.-Y.; Rauh, M.J.; Popova, T.; Sebbag, C.; Lantz, O.; Stern, M.-H.; Mueller, C.R. A DNA methylation-based liquid biopsy for triple-negative breast cancer. npj Precis. Oncol. 2021, 5, 1–13. [Google Scholar] [CrossRef]
- Liu, L.; Sun, L.; Li, C.; Li, X.; Zhang, Y.; Yu, Y.; Xia, W. Quantitative detection of methylation of FHIT and BRCA1 promoters in the serum of ductal breast cancer patients. Bio-Medical Mater. Eng. 2015, 26, S2217–S2222. [Google Scholar] [CrossRef]
- Sturgeon, S.R.; Balasubramanian, R.; Schairer, C.; Muss, H.B.; Ziegler, R.G.; Arcaro, K.F. Detection of promoter methylation of tumor suppressor genes in serum DNA of breast cancer cases and benign breast disease controls. Epigenetics 2012, 7, 1258–1267. [Google Scholar] [CrossRef] [PubMed]
- de Ruijter, T.C.; van der Heide, F.; Smits, K.M.; Aarts, M.J.; van Engeland, M.; Heijnen, V.C.G. Prognostic DNA methylation markers for hormone receptor breast cancer: a systematic review. Breast Cancer Res. 2020, 22, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Yen, J.; Chen, S.; Jenkins, C.; Overstreet, B.; Fu, Y.; Zhao, J.; Jiang, T.; Drusbosky, L.; Pettitt, S.; Dorschner, M.; et al. Abstract 6603: BRCA1 promoter methylation in sporadic breast cancer patients detected by liquid biopsy. Cancer Res. 2023, 83, 6603–6603. [Google Scholar] [CrossRef]
- Koukaki, T.; Balgkouranidou, I.; Biziota, E.; Karayiannakis, A.; Bolanaki, H.; Karamitrousis, E.; Zarogoulidis, P.; Deftereos, S.; Charalampidis, C.; Ioannidis, A.; et al. Prognostic significance of BRCA1 and BRCA2 methylation status in circulating cell-free DNA of Pancreatic Cancer patients. J. Cancer 2024, 15, 2573–2579. [Google Scholar] [CrossRef] [PubMed]
- Vasseur, A.; Kiavue, N.; Bidard, F.-C.; Pierga, J.-Y.; Cabel, L. Clinical utility of circulating tumor cells: an update. Mol. Oncol. 2021, 15, 1647–1666. [Google Scholar] [CrossRef] [PubMed]
- Stordal, B.; Timms, K.; Farrelly, A.; Gallagher, D.; Busschots, S.; Renaud, M.; Thery, J.; Williams, D.; Potter, J.; Tran, T.; et al. BRCA1/2 mutation analysis in 41 ovarian cell lines reveals only one functionally deleterious BRCA1 mutation. Mol. Oncol. 2013, 7, 567–579. [Google Scholar] [CrossRef] [PubMed]
- Cortesi, L.; Rugo, H.S.; Jackisch, C. An Overview of PARP Inhibitors for the Treatment of Breast Cancer. Target. Oncol. 2021, 16, 255–282. [Google Scholar] [CrossRef]
- Moschetta, M.; George, A.; Kaye, S.B.; Banerjee, S. BRCA somatic mutations and epigenetic BRCA modifications in serous ovarian cancer. Ann. Oncol. 2016, 27, 1449–1455. [Google Scholar] [CrossRef] [PubMed]
- Jacot, W.; Thezenas, S.; Senal, R.; Viglianti, C.; Laberenne, A.-C.; Lopez-Crapez, E.; Bibeau, F.; Bleuse, J.-P.; Romieu, G.; Lamy, P.-J. BRCA1 promoter hypermethylation, 53BP1 protein expression and PARP-1 activity as biomarkers of DNA repair deficit in breast cancer. BMC Cancer 2013, 13, 523–523. [Google Scholar] [CrossRef]
- Zhu, X.; Chen, L.; Huang, B.; Li, X.; Yang, L.; Hu, X.; Jiang, Y.; Shao, Z.; Wang, Z. Efficacy and mechanism of the combination of PARP and CDK4/6 inhibitors in the treatment of triple-negative breast cancer. J. Exp. Clin. Cancer Res. 2021, 40, 1–18. [Google Scholar] [CrossRef]
- Gelmon, K.A.; Tischkowitz, M.; Mackay, H.; Swenerton, K.; Robidoux, A.; Tonkin, K.; Hirte, H.; Huntsman, D.; Clemons, M.; Gilks, B.; et al. Olaparib in patients with recurrent high-grade serous or poorly differentiated ovarian carcinoma or triple-negative breast cancer: a phase 2, multicentre, open-label, non-randomised study. Lancet Oncol. 2011, 12, 852–861. [Google Scholar] [CrossRef] [PubMed]
- Veeck, J.; Ropero, S.; Setien, F.; Gonzalez-Suarez, E.; Osorio, A.; Benitez, J.; Herman, J.G.; Esteller, M. BRCA1 CpG Island Hypermethylation Predicts Sensitivity to Poly(Adenosine Diphosphate)- Ribose Polymerase Inhibitors. J. Clin. Oncol. 2010, 28, e563–e564. [Google Scholar] [CrossRef] [PubMed]
- Kondrashova, O.; Topp, M.; Nesic, K.; Lieschke, E.; Ho, G.Y.; Harrell, M.I.; Zapparoli, G.V.; Hadley, A.; Holian, R.; Boehm, E.; et al. Methylation of all BRCA1 copies predicts response to the PARP inhibitor rucaparib in ovarian carcinoma. Nat. Commun. 2018, 9, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Dimitrova, D.; Ruscito, I.; Olek, S.; Richter, R.; Hellwag, A.; Türbachova, I.; Woopen, H.; Baron, U.; Braicu, E.I.; Sehouli, J. Germline mutations of BRCA1 gene exon 11 are not associated with platinum response neither with survival advantage in patients with primary ovarian cancer: understanding the clinical importance of one of the biggest human exons. A study of the Tumor Bank Ovarian Cancer (TOC) Consortium. Tumor Biol. 2016, 37, 12329–12337. [Google Scholar] [CrossRef]
- Drost, R.; Dhillon, K.K.; Van Der Gulden, H.; Van Der Heijden, I.; Brandsma, I.; Cruz, C.; Chondronasiou, D.; Castroviejo-Bermejo, M.; Boon, U.; Schut, E.; et al. BRCA1185delAG tumors may acquire therapy resistance through expression of RING-less BRCAJ. Clin. Investig. 2016, 126, 2903–2918. [Google Scholar] [CrossRef] [PubMed]
- Jacot, W.; Lopez-Crapez, E.; Mollevi, C.; Boissière-Michot, F.; Simony-Lafontaine, J.; Ho-Pun-Cheung, A.; Chartron, E.; Theillet, C.; Lemoine, A.; Saffroy, R.; et al. BRCA1 Promoter Hypermethylation is Associated with Good Prognosis and Chemosensitivity in Triple-Negative Breast Cancer. Cancers 2020, 12, 828. [Google Scholar] [CrossRef] [PubMed]
- Buisson, R.; Dion-Côté, A.-M.; Coulombe, Y.; Launay, H.; Cai, H.; Stasiak, A.Z.; Stasiak, A.; Xia, B.; Masson, J.-Y. Cooperation of breast cancer proteins PALB2 and piccolo BRCA2 in stimulating homologous recombination. Nat. Struct. Mol. Biol. 2010, 17, 1247–1254. [Google Scholar] [CrossRef] [PubMed]
- Annunziata, C.M.; O'Shaughnessy, J. Poly (ADP-Ribose) Polymerase as a Novel Therapeutic Target in Cancer. Clin. Cancer Res. 2010, 16, 4517–4526. [Google Scholar] [CrossRef] [PubMed]
- Drew, Y.; Mulligan, E.A.; Vong, W.-T.; Thomas, H.D.; Kahn, S.; Kyle, S.; Mukhopadhyay, A.; Los, G.; Hostomsky, Z.; Plummer, E.R.; et al. Therapeutic Potential of Poly(ADP-ribose) Polymerase Inhibitor AG014699 in Human Cancers With Mutated or Methylated BRCA1 or BRCA2. JNCI J. Natl. Cancer Inst. 2010, 103, 334–346. [Google Scholar] [CrossRef]
- Vos, S.; Moelans, C.B.; van Diest, P.J. BRCA promoter methylation in sporadic versus BRCA germline mutation-related breast cancers. Breast Cancer Res. 2017, 19, 64. [Google Scholar] [CrossRef]
- Guo, M.; Peng, Y.; Gao, A.; Du, C.; Herman, J.G. Epigenetic heterogeneity in cancer. Biomark. Res. 2019, 7, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Prieske, K.; Prieske, S.; Joosse, S.A.; Trillsch, F.; Grimm, D.; Burandt, E.; Mahner, S.; Schmalfeldt, B.; Milde-Langosch, K.; Oliveira-Ferrer, L.; et al. Loss of BRCA1 promotor hypermethylation in recurrent high-grade ovarian cancer. Oncotarget 2017, 8, 83063–83074. [Google Scholar] [CrossRef] [PubMed]
- Harvey-Jones, E.; Raghunandan, M.; Robbez-Masson, L.; Thanussuyah, A.; Liccardo, R.; Yablonovitch, A.; Cai, M.; Drusbosky, L.; Dorschner, M.; Pardo, L.M.; et al. Abstract 6094: Longitudinal analysis of PARP inhibitor and platinum resistance in BRCA1/2m breast cancer using liquid biopsy. Cancer Res. 2023, 83, 6094–6094. [Google Scholar] [CrossRef]
- Jayaram, A.; Wingate, A.; Wetterskog, D.; Wheeler, G.; Sternberg, C.; Jones, R.; Berruti, A.; Lefresne, F.; Lahaye, M.; Thomas, S.; et al. Plasma tumor gene conversions after one cycle abiraterone acetate for metastatic castration-resistant prostate cancer: a biomarker analysis of a multicenter international trial. Ann. Oncol. 2021, 32, 726–735. [Google Scholar] [CrossRef]
Figure 1.
BRCA1 and BRCA2 molecular mechanisms of DNA damage response during in homologous recombination. i) A double-strand DNA molecule without errors. ii) The DSB of DNA is typically caused by external factors such as ionizing radiation or chemotherapeutic drugs but also naturally due to the accumulation of reactive oxygen species. Most of the time, this DNA damage leads to the uneven loss of numerous nucleotides resulting in two DNA strands that are incompatible near the breakage point. iii) ATM and ATR kinases are activated in the presence of DSB initiating the repair process by phosphorylating a set of DNA repair targets. One of the main phosphorylation targets is the BRCA1, which in turn associates with the Mre11, Rad50, and NBS1 (MRN complex) as well as CtIP, eventually forming a multi-protein BRCA1 complex. This complex approaches the DSBs and initiates DNA end resection, creating single-strand DNA (ssDNA) overhangs. iv) After DNA end resection is completed, BRCA1 recruits PALB2 and BRCA2 which in turn promotes the loading of the recombination enzyme RAD51 to the ssDNA. v) The second BRCA1-based protein complex starts the DNA strand invasion and homologous repair mechanism, restoring the genetic information lost at the breakage. vi) After this multi-step but accurate process is successfully completed, two identical or almost identical DNA double strands are formed.
Figure 1.
BRCA1 and BRCA2 molecular mechanisms of DNA damage response during in homologous recombination. i) A double-strand DNA molecule without errors. ii) The DSB of DNA is typically caused by external factors such as ionizing radiation or chemotherapeutic drugs but also naturally due to the accumulation of reactive oxygen species. Most of the time, this DNA damage leads to the uneven loss of numerous nucleotides resulting in two DNA strands that are incompatible near the breakage point. iii) ATM and ATR kinases are activated in the presence of DSB initiating the repair process by phosphorylating a set of DNA repair targets. One of the main phosphorylation targets is the BRCA1, which in turn associates with the Mre11, Rad50, and NBS1 (MRN complex) as well as CtIP, eventually forming a multi-protein BRCA1 complex. This complex approaches the DSBs and initiates DNA end resection, creating single-strand DNA (ssDNA) overhangs. iv) After DNA end resection is completed, BRCA1 recruits PALB2 and BRCA2 which in turn promotes the loading of the recombination enzyme RAD51 to the ssDNA. v) The second BRCA1-based protein complex starts the DNA strand invasion and homologous repair mechanism, restoring the genetic information lost at the breakage. vi) After this multi-step but accurate process is successfully completed, two identical or almost identical DNA double strands are formed.
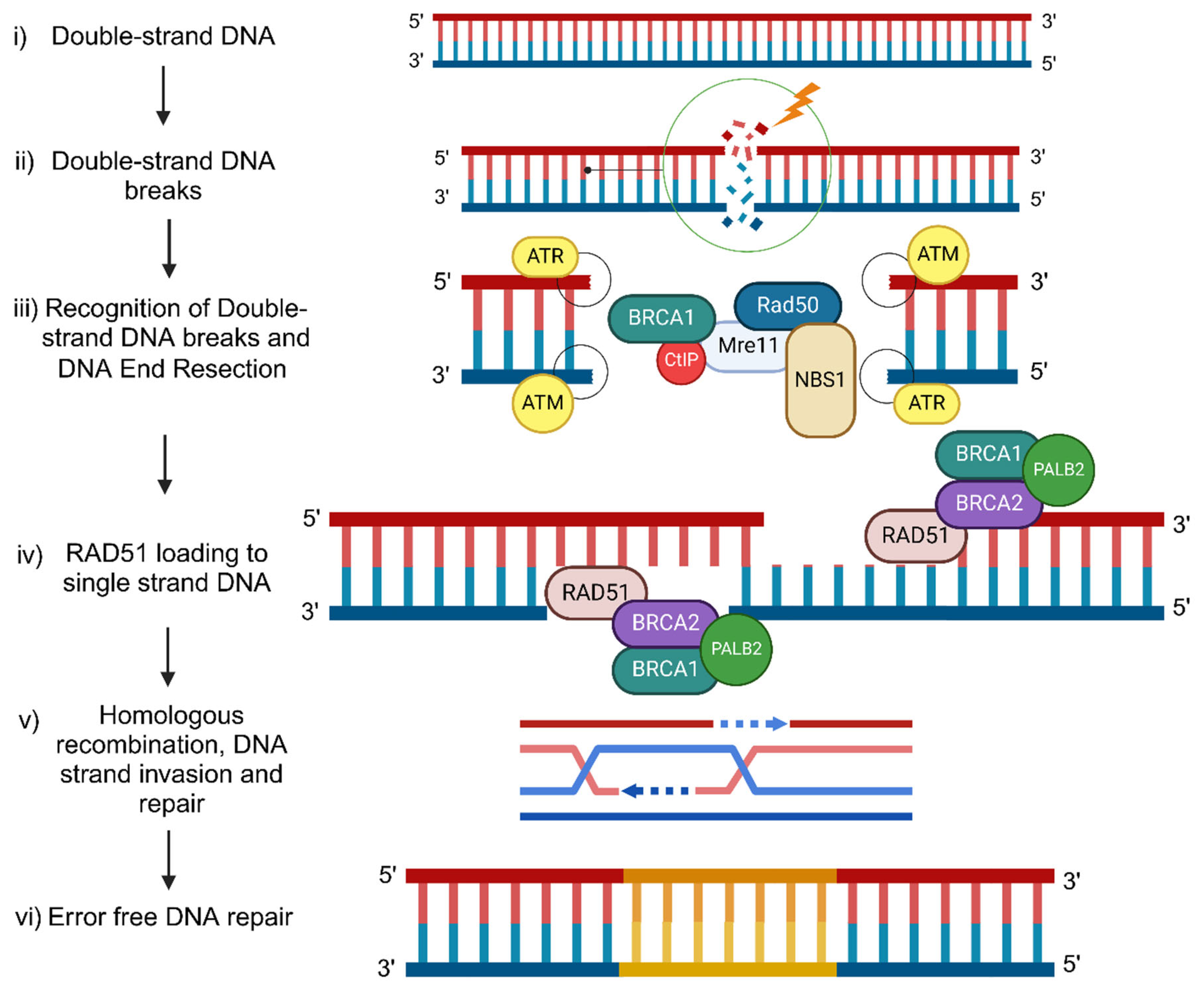
Figure 2.
A suggested diagnostic pipeline to identify cancer patients who will benefit from PARPi treatment. Abbreviation: PARPi: PARP inhibitors
Figure 2.
A suggested diagnostic pipeline to identify cancer patients who will benefit from PARPi treatment. Abbreviation: PARPi: PARP inhibitors
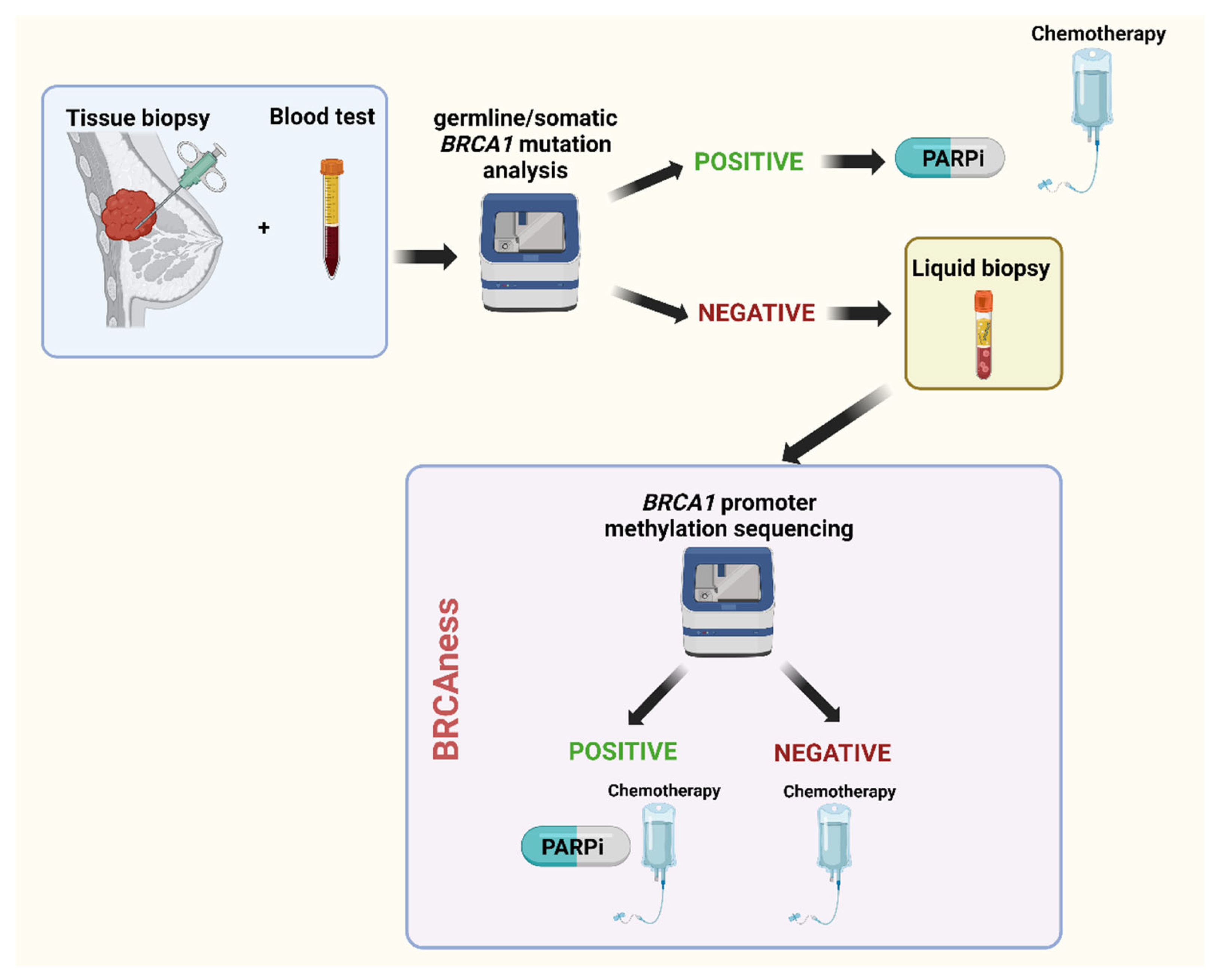

Table 1.
BRCA1 methylation percentages in tissue and blood cells among cancer types and their correlations with the disease's clinically significant end-points.
Table 1.
BRCA1 methylation percentages in tissue and blood cells among cancer types and their correlations with the disease's clinically significant end-points.
| Cancer type | Biomaterial | BRCA1 methylation (%) | Correlation | Reference |
|---|---|---|---|---|
| BrCa | Tissue | 9.1 | Diagnosis at a young age | Birgisdottir et al [65] |
| 3.0 | Improved Survival after chemotherapy | Stefansson et al [68] | ||
| 12.4 | Incidence of TNBC | Lonning et al [77] | ||
| 26.0 | Worse Survival | Chen et al [78] | ||
| 24.1 | Improved Survival after chemotherapy | Glodzik et al [66] | ||
| TNBC | Tissue | 20.6 | Improved Survival | Brianese et al [67] |
| OvCa | Tissue | 16.3 | Young age, Advanced stage, Improved Survival | Kalachand et al [43] |
| 19 (high methylation) 14 (low methylation) |
High methylation with GIS and PARPi treatment option | Durand et al [79] | ||
| 89.9 | None | Pradjatmo et al [80] | ||
| 5.2 | Partially BRCAness prediction | Aref-Eshghi et al [49] | ||
| 19.3 | None | Sahnane et al | ||
| HGSOC | Tissue | 14.8 | Young age | Ruscito et al [81] |
| PrCa | Tissue | 100.0 | None | Rabiau et al [17] |
| 0.0 | None | Bednarz et al [82] | ||
| PaCa | blood lymphocytes | 0.3 | None | Zhou et al [83] |
| Tissue | 60.3 | Poorer tumor differentiation, protein expression levels |
Peng et al. [84] | |
| PaCa | Tissue | 0.0 | None | Abdalah et al. [85] |
| PaCa | Tissue & Blood lymphocytes | 3.6 | None | Zhen-Lin et al. [86] |
Abbreviations: BrCa: Breast Cancer; TNBC: Triple-Negative Breast Cancer; OvCa: Ovarian Cancer; HGSOC: High-Grade Serous Ovarian Cancer, PaCa: Pancreatic Cancer, GIS: Genomic Instability; PARPi: PARP inhibitor.
140. Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



