
Submitted:
19 July 2024
Posted:
24 July 2024
You are already at the latest version
Abstract
The bacterium Yersinia ruckeri causes enteric redmouth disease in salmonids and hence has substantial economic implications for the farmed fish industry. The Norwegian Y. ruckeri outbreak isolate NVH_3758 carries a relatively uncharacterized plasmid, pYR4, which encodes both type 4 pili and a type 4 secretion system. In this study, we demonstrate that pYR4 does not impose a growth burden on the Y. ruckeri host bacterium, nor does the plasmid contribute to twitching motility (an indicator of type 4 pilus function) or virulence in a Galleria mellonella larval model of infection. However, we show that pYR4 is conjugative. We also reveal, through mutagenesis, that pYR4 encodes a functional post-segregational killing system, HigBA, that is responsible for plasmid maintenance within Y. ruckeri. This is the first toxin-antitoxin system to be characterized for this organism. Whilst further work is needed to elucidate the virulence role of pYR4 and whether it contributes to bacterial disease under non-laboratory conditions, our results suggest that the plasmid possesses substantial stability and transfer mechanisms that imply importance within the organism. These results add to our understanding of the mobile genetic elements and evolutionary trajectory of Y. ruckeri as an important commercial pathogen, with consequences for human food production.

Keywords:
conjugation
; toxin-antitoxin system
; virulence plasmid
; Yersinia ruckeri
1. Introduction
Farming of aquatic organisms such as fish and shellfish, or aquaculture, is an important and growing industry to meet the worlds nutritional needs. In 2021, the amount of fish produced through aquaculture equaled the amount of fish caught from wild fisheries [1], and the proportion of farmed fish is now exceeding that of wild-captured fish [2]. However, the crowded and stressful conditions in fish farms predispose the livestock to infections by bacterial, viral, and parasitic pathogens [3]. Infections in aquaculture have been estimated to cost the global economy over $6 billion per year [4].
One such fish pathogen is the bacterium Yersinia ruckeri, the causative agent of enteric redmouth disease (ERM). ERM affects mainly salmonid fish, including commercially important species such as Atlantic salmon and rainbow trout [5]. Though ERM has been controlled to some extent through vaccination, outbreaks caused by bio-serotypes not covered or poorly controlled by existing vaccines have occurred in recent years [6,7,8]. In addition, the pathogenesis of ERM is not well understood. Although several virulence factors of Y. ruckeri have been identified [9], their role in the disease has not been fully characterized. Therefore, investigation into the disease mechanisms of Y. ruckeri is important for understanding its pathogenesis and developing novel control measures.
The human pathogenic Yersinia species rely on plasmid-mediated virulence for causing disease [10]. Similarly, most Y. ruckeri isolates contain plasmids [11,12]. Recently, the plasmid pYR4 was discovered in the Norwegian outbreak strain NVH_3758, encoding a type 4 pilus (TFP) cluster and a type 4 secretion system (T4SS) [13]. Type 4 pili are contractile fimbrial adhesins that mediate twitching motility but are also often involved in biofilm formation, pathogenesis, or DNA uptake [14]. T4SSs (distinct structures from TFP) include conjugation apparatus [15], which is why pYR4 is presumed conjugative [13]. The tra locus of Y. ruckeri 150R, encoding a T4SS, has been implicated in virulence [16]. The region encoding this T4SS locus as well as a pil locus encoding TFP is highly similar to regions on the plasmids pYR3 and pYR4 and so is also likely plasmid-borne [13]. Due to the presence of the TFP and T4SS loci on pYR4, it has been suspected to be virulence plasmid. Efforts to cure pYR4 were unsuccessful [13], suggesting it is stably maintained in the population.
Bioinformatic examination of pYR4 revealed a toxin-antitoxin system locus, higBA. Toxin-antitoxin (TA) systems, also known as addiction factors, encode a stable toxin and an antitoxin that is degraded quickly. If the antitoxin gene is present, the antitoxin pool can be regenerated, and the toxin is not able to exert its toxicity on the cell. However, if the antitoxin gene is lost (e.g. through curing of the plasmid encoding it), the stable toxin persists while any remaining antitoxin is degraded, leading to toxic effects resulting in cessation of growth or death of the cell. Therefore, the host bacteria become “addicted” to the plasmid because its maintenance is essential for a healthy growing population [17,18]. The HigBA module is part of a wider group of TA systems known as type II TA systems, in which both the toxin and antitoxin are proteins, and the antitoxin binds directly to and sequesters the toxin [19]. The higBA operon is unusual in that the toxin gene is transcribed upstream of the antitoxin, contrary to most type II TA operons [20]. The HigB protein itself is a ribosome-dependent mRNA endoribonuclease and is a member of the RelE family of toxins [21].
We hypothesized that the HigBA TA system of pYR4 contributes to its stability. In this study, we demonstrate that the higBA locus is a functional TA system in Y. ruckeri and that its deletion results in a reduction in the stability of pYR4. This is, to our knowledge, the first characterization of a TA system in Y. ruckeri. We further provide evidence to show that pYR4 is a conjugative plasmid. However, using a simple infection model, larvae of the greater wax moth (Galleria mellonella), we did not observe any difference in virulence between the wild-type and the plasmid cured strains. The status of pYR4 as a virulence plasmid thus remains unresolved.
2. Materials and Methods
2.1. Bacterial Strains and Growth Conditions
The bacterial strains used in this study were E. coli DH5α and Y. ruckeri NVH_3758 [22,23]. Bacteria were grown in lysogeny broth (LB) [24] and supplemented with antibiotics where necessary; ampicillin at 100 µg/mL, chloramphenicol at 25 µg/mL and neomycin and streptomycin at 50 µg/mL. 1.5 % (w/v) agar was added to prepare solid media. For experiments in minimal medium, M9 was used [25], either with glycerol at 0.2% rather than glucose, or supplemented with iron(II) sulphate (10 µM), thiamine (5 µg/mL), BME vitamins (Sigma) and A5 trace metals (Sigma). E. coli was propagated at 37 °C and Y. ruckeri at 30 °C unless otherwise indicated.
2.2. Cloning
For toxicity assays, the higB gene was amplified using Y. ruckeri NVH_3758 genomic DNA as the template using Q5 polymerase (New England Biolabs). Whole genome DNA was extracted from Y. ruckeri using the Wizard® Genomic DNA Purification Kit (Promega). Primers (Table A1 in the Appendix) were designed to include overhangs for Gibson assembly [26] of the insert into the arabinose-inducible vector pBAD33 [27]. To produce the antitoxin construct, the entire operon (higBA, including upstream and downstream regions) was amplified as above with overhangs complementary to the pGM101 vector [28]. Alternatively, variants of the higBA operon were cloned using mutagenic primers to produce pGM101::ΔhigB (lacking the higB gene entirely) or pGM101::higBmut (containing a higB start codon ATG → CTG mutation). Vectors were linearised by PCR and digested with DpnI to remove the circular template. The constructs were then assembled using the NEBuilder HiFi DNA Assembly kit (New England Biolabs), and the reaction mixtures were transformed into E. coli DH5α. Transformants were selected for using chloramphenicol (pBAD33) or ampicillin (pGM101), and insert-positive clones were screened for using colony PCR. All constructs were verified by Sanger sequencing.
2.3. Creating a higB Knockout and Introducing a Selection Marker into pYR4
To knock out the higB gene in Y. ruckeri, the upstream (US) and downstream (DS) regions (600 bp in length) of the higBA operon were amplified by PCR. In addition, the higBA promoter-higA sequence was amplied from pGM101::ΔhigB as well as a chloramphenicol resistance cassette (cat gene plus its promoter and flanking FRT sites) from pKD3 [29]. Primers (Table A1 in the Appendix) were designed such that these PCR products could be annealed through Gibson assembly to yield US-higA-cat-DS as a single DNA fragment lacking the higB gene (Figure A1a in the Appendix). This fragment was produced using the NEBuilder HiFi Assembly kit, and the assembled reaction was used as a template for amplifying the US-higA-cat-DS fragment. To generate the knockout, the λ Red helper plasmid pMJH65 [30] was transformed into NVH_3758 by electroporation and transformants were selected for on LB + ampicillin. A transformant colony was grown in LB at 30 °C and the λ Red recombination proteins were induced by the addition of 0.2% arabinose for an hour before harvesting the cells for electroporation. 1 µg of the US-higA-cat-DS fragment was used for electroporation; the recovery medium in which the bacteria were incubated for 2 hours at 30 °C was supplemented with magnesium and arabinose (LB + 10 mM MgCl2 + 0.2% arabinose). The bacteria were then plated on LB + chloramphenicol. Transformants were screened by PCR to identify clones with either insertion of the cat gene after the intact higBA operon (pYR4::cat higB+) or deletion of higB and insertion of the cat gene after higA (pYR4::cat ΔhigB) (Figure A1b in the Appendix). pMJH65 was removed by growing the bacteria overnight without selection and plating for single colonies, followed by screening for ampicillin-sensitive clones. The correctness of the insertions (either cat or ΔhigB cat) was verified by Sanger sequencing of the amplified hig locus. pMJH65 was a gift from Mark Liles (Addgene plasmid #67273 ; http://n2t.net/addgene:67273 ; RRID:Addgene_67273)
2.3. Plasmid Curing and Plasmid Loss Experiments
To cure pYR4, we initially grew bacteria at an elevated temperature (37 °C) for extended periods and tested for the presence of pYR4 by PCR. However, this did not result in any observable plasmid loss, and neither did transient repeated heat shocks at higher temperatures (up to 47 °C). Plasmid curing only occurred after introduction of the pGM101::ΔhigB into NVH_3758 by electroporation and subsequent culture of a confirmed transformant at 37 °C in LB + ampicillin, followed by testing for the presence of the plasmid by PCR. Once a pYR4-negative clone had been verified, the pGM101::ΔhigB plasmid was cured by culturing in the absence of ampicillin overnight, diluting the culture 1:10,000 the following day in fresh medium, and reculturing, over the course of three days. Then the culture was plated for single colonies and ampicillin-sensitive clones were screened for by streaking individual colonies first on LB +ampicillin and then LB; clones that grew on LB but not LB + ampicillin were chosen. That these were Y. ruckeri was confirmed by 16S rDNA PCR.
To determine the effect of the higB deletion, the NVH_3758 strains with pYR4::cat derivatives (either higB+ or ΔhigB) were inoculated in biological quadruplicate in LB and grown overnight at 37 °C to stress the cells and expedite plasmid loss. The following day, the cultures were diluted 1:10,000 in fresh medium and regrown at 37 °C. This was continued for a total of 10 days. To quantify plasmid loss, dilution series of the cultures were plated onto LB with no selection and LB + chloramphenicol; only bacteria still retaining the plasmid would grow on the selection plate. We then calculated the percentage of resistant colonies in the overall viable count of the cultures.
2.4. Conjugation
To prepare recipient cells for conjugation experiments, spontaneous streptomycin-resistant mutants were isolated by plating cultures of Y. ruckeri NVH_3758 pYR4- onto LB medium with 50 µg/mL of streptomycin. After two days of culture at 30 °C, colonies were restreaked onto streptomycin medium to verify the resistance. Alternatively, we introduced a neomycin resistance marker-containing plasmid (pGM101neo (Whelan & McVicker, unpublished)) by electroporation and selection on neomycin (50 µg/mL). This plasmid was determined to be stably kept in the population during overnight culture even in the absence of selection. For conjugation, the streptomycin or neomycin-resistant pYR4- strains were used as the recipient, mixed with the pYR4::cat higB+ donor. The strains were grown in liquid medium separately overnight with selection, and then 1 mL of the cultures were pelleted at 3,000 x g for 5 minutes, washed and then resuspended in 500 µL PBS. The donor and recipient strains were then mixed at different ratios (1:1, 2:1, 1:2) in a total of 100 µL of PBS, and this was spotted onto an LB plate without selection and grown overnight at 30 °C. The bacteria were then scraped off the plate, diluted in PBS and plated for single colonies on counterselection plates containing chloramphenicol to select for the pYR4::cat plasmid and neomycin/streptomycin to select for the recipient, then grown overnight at 30 °C. Putative transconjugants from the chloramphenicol-neomycin plates were screened by PCR for both the pYR4 plasmid and the pGM101neo plasmid.
2.5. Toxicity Assays
Bacterial viability in response to toxin-antitoxin production was assayed according to [28]. Briefly, cells containing pBAD33::higB together with either pGM101, pGM101::higBA, pGM101::ΔhigB or pGM101::higBmut were grown overnight in LB + ampicillin + chloramphenicol + 0.2 % glucose to repress toxin expression. Cultures were then subcultured and grown to OD600nm = 0.1 in fresh media at 37 °C, before pelleting and resuspending the bacteria in prewarmed LB + ampicillin + chloramphenicol + 1 % arabinose to induce toxin gene expression. At 0, 15, 30, 60 and 180 minutes post induction, samples were serially diluted in PBS before plating onto LB agar + ampicillin + chloramphenicol + 0.2 % glucose for quantification of the number of viable CFU at each timepoint.
2.6. Twitching Motility Assays
Twitching motility was assayed for using a macroscopic twitching assay, essentially as described in [31]. Briefly, bacteria were stabbed into the centre of a 1 % (w/v) LB agar plate down to the agar-plastic interface. The plates were then incubated overnight at 25 °C, 30 °C and 37 °C, with three biological triplicates for all temperatures, after which the interstitial halo was measured (no developer was added). As a positive control, we used Pseudomonas aeruginosa PA14.
2.7. Galleria Infection Assays
For in vivo infection assays, we used Galleria mellonella larvae as a model (purchased from LiveFoods Direct, UK). Upon arrival, the larvae were checked and any blackening or pupating larvae were discarded. The larvae were then weighed and ones between 200 and 300 mg were used in the experiments. For infections, Y. ruckeri NVH_3758 pYR4+ and pYR4- strains were cultured overnight at 30 °C; the following day subcultures were prepared and grown till an OD600nm of 0.4-0.5. These cultures were then diluted to specific CFU/mL in PBS, based on a standard curve relating CFU/mL to OD600nm. In addition to live bacteria, we also included heat-killed NVH_3758 pYR4+ (20 minutes at 80 °C) as a control to test whether bacterial cell components posed any toxicity to the larvae. 10 Galleria were injected with each sample using 27G needles and 1 mL syringes with a mechanical syringe pump (Cole-Parmer, UK) at a rate of 13.21 mL/h and a volume of 10 µL per injection. Injections were made on the bottom left proleg into the haemocoel of the larvae. Inoculum doses were checked by diluting and plating from 10 µL drops from the syringe. Sterile PBS-injected and uninjected larvae were used as negative controls. 5 larvae from each group were then incubated in Petri dishes at either 30 °C or 25 °C for 96 hours. Scoring of the larvae’s health was done according to the scale of Serrano et al. [32] , with higher scores indicating healthy larvae.
3. Results
3.1. pYR4 Carries Several Putative TA-Related Genes
The 80.8 kb putative virulence plasmid pYR4 from Y. ruckeri strain NVH_3758 (Figure 1A) contains genes potentially encoding a T4SS and TFP [13]. In addition, a manual analysis of pYR4 identified several sequences of interest for plasmid stabilisation, such as genes potentially encoding two partitioning systems (StbAB and ParAG) and TA systems. TA systems and TA-related/orphaned proteins encoded within the plasmid’s so-called “stability cluster” include a putative type IV system toxin (CbtA) and its associated XRE-family antidote, lone antitoxins from the Phd, MqsA and ParD families and an Abi-family protein. The plasmid also encodes an intact HigBA type II TA system; pYR4 HigB is 39.25% identical (50.46% similar) to the HigB-2 toxin of the Vibrio cholerae N16961 superintegron, whereas pYR4 HigA is 23.46% identical (37.75% similar) to the Vibrio HigA-2 antitoxin [21]. Interestingly, the cbtA gene is truncated by an IS6-family transposase in the pYR4-like plasmid pYR3 found in Y. ruckeri strain CSF007-82 (Figure 1B), removing the first 29 amino acids at the protein’s N-terminus and presumably the gene’s promoter, and creating a region of sequence diversity between the two plasmids upstream of the cbtA pseudogene. We therefore focused our investigation on the conserved higBA locus, which is shared between pYR3 and pYR4.
3.2. pYR4 HigBA Is a Functional Postsegregational Killing System
In order to ascertain whether pYR4 higBA encodes a bona fide toxin-antitoxin system, both toxin and antitoxin activity were assessed. The toxin gene higB and its native ribosome binding site were cloned into the arabinose-inducible promoter on pBAD33, creating pHigB. Due to the inverted nature of the operon, three antitoxin-expressing constructs (Figure 2A) were cloned separately into compatible vector pGM101 to test their ability to abrogate toxicity: one containing the entire wild type operon (pGM101::higBA), one containing the operon with a deletion of the higB gene (pGM101::ΔhigB) and one containing the entire operon with an A→C mutation in the start codon of higB (pGM101::higBmut), preventing its translation. Each antitoxin construct included the operon’s native promoter in order to take advantage of conditional cooperativity [33,34].
Expression of the toxin gene from pBAD33 in E. coli DH5α in the absence of antitoxin resulted in approximately 100-fold loss of cell viability at 180 mins post induction relative to strains carrying any of the three antitoxin variants (Figure 2B; P = 0.0003). Curing pYR4 from NVH_3758 (see below) allowed us to also test the effect of the TA system in its native host. Here, the effect of the toxin was far more dramatic (10,000-fold reduction in viability) but was still abolished by any of the three antitoxin constructs (Figure 2C; P < 0.0001). Hence, HigBA encoded on pYR4 is a functional and cognate toxin-antitoxin pair.
In order to test pYR4 stability under laboratory conditions, we grew Y. ruckeri NVH_3758 for several weeks with daily subculturing into fresh media. At various timepoints during the experiment, cultures were serially diluted and plated to obtain single colonies, then those colonies tested for the presence of pYR4 using a multiplex PCR targeting key plasmid genes repA, pilN and higB (Figure A2 in the Appendix). No evidence of plasmid loss was observed. The same was true when cultures were briefly heat-shocked to induce stress-based plasmid loss at temperatures up to 47 °C, the maximum temperature for which we obtained viable bacteria after recovery and plating. pYR4 seems, therefore, to be highly stable, in agreement with previous observations [13].
Given that we had previously shown that HigA is able to prevent HigB toxicity and hence interfere with TA system function (Figure 2), we transformed NVH_3758 with pGM101::higBmut and tested plasmid loss in the resulting transformant cells using the multiplex PCR as before. Results showed approximately 30 % of colonies lacked repA and pilN (though retained the higB band, as this was present on the antitoxin vector; Figure A3 in the Appendix) within ten days of subculture, sharply contrasting our attempts at plasmid curing without the use of HigA. Subsequent growth in non-selective media resulted in spontaneous curing of the pGM101::higBmut vector (assessed by PCR and ampicillin sensitivity), resulting in NVH_3758_pYR4–.
Lastly, as direct proof of TA-mediated plasmid addiction, we used λ Red recombineering to construct a ΔhigB deletion at the native pYR4 locus tagged with a chloramphenicol resistance marker, and the corresponding chloramphenicol-resistant higB+ control strain. After ten daily subcultures, pYR4ΔhigB retention (measured by chloramphenicol resistance) was only 41 % of that of the higB+ plasmid (Figure 3; P < 0.0001). These results, alongside the relative ease of pYR4 curing via the pGM101::higBmut construct, are consistent with HigBA being a functional postsegregational killing system that enhances stability of pYR4.
3.3. Qualitative Evidence of pYR4 Conjugation
Since pYR4 putatively encodes a T4SS, we surmised that it may be conjugative. Analysis via MOB-Typer [35] failed to identify any conjugation machinery but suggested that the plasmid may be mobilisable; however, since MOB-Typer is by the authors’ own admission focussed on Enterobacteriaceae plasmids, its lack of recognition of Yersinia mobilisation/conjugation elements is not surprising.
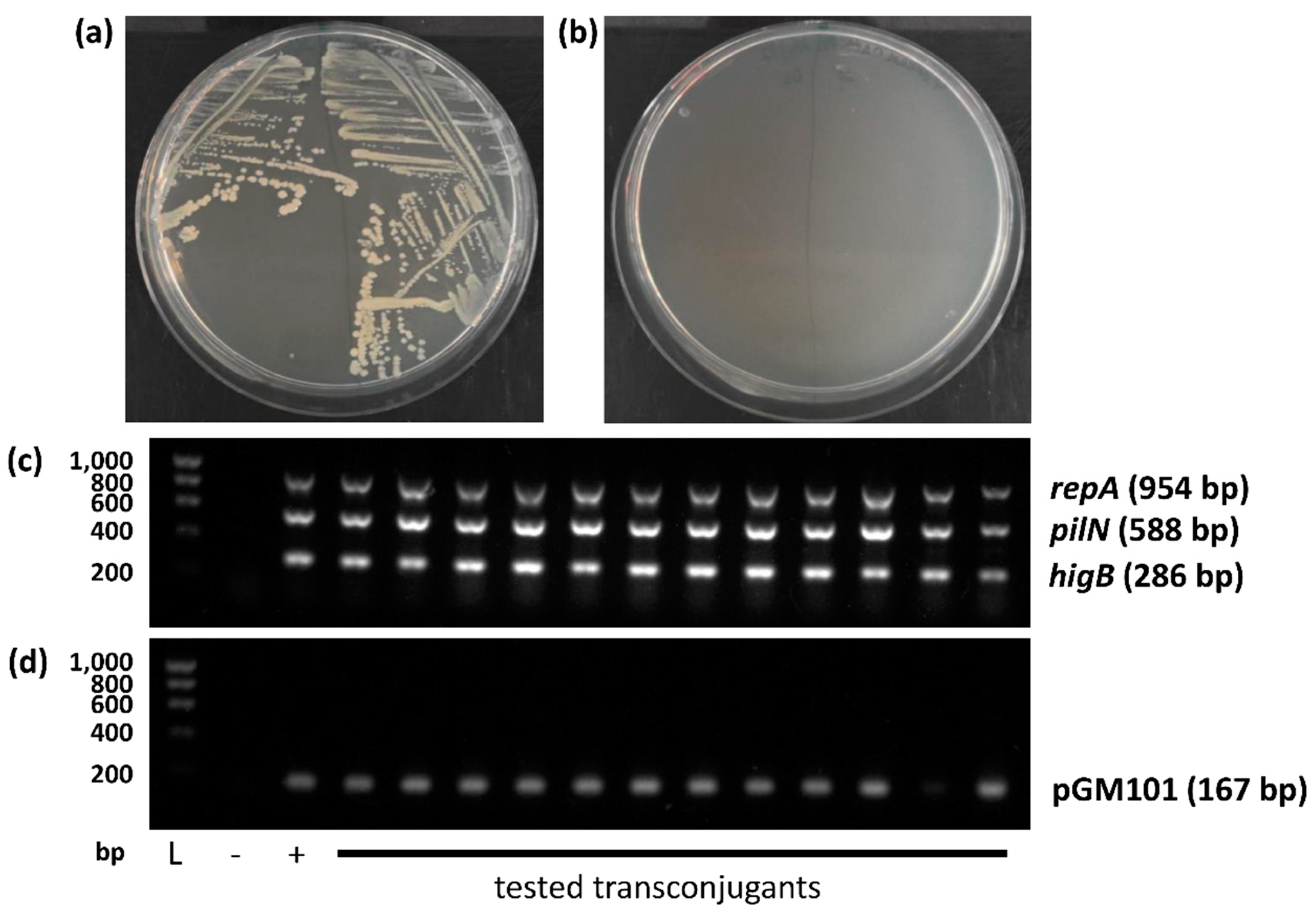
In order to experimentally test the conjugative capacity of pYR4, we employed the higB+ chloramphenicol-resistant variant produced previously in this work and tested its ability to transfer into either a recipient strain harbouring pGM101neo, conferring neomycin resistance, or a recipient strain harbouring a spontaneous streptomycin resistance mutation generated through growth on streptomycin in our laboratory. In both cases, results were impossible to quantify due to an extremely high level of background growth post-conjugation. However, by PCR, we identified transconjugant colonies containing both pYR4 and pGM101neo in the relevant experiment, therefore giving qualitative evidence of successful conjugation (Figure 4). Notably, such colonies continued to grow healthily and provide positive PCR results for both recipient and donor genes when they were restreaked onto fresh medium and re-analysed, whereas colonies taken from background growth did not grow well after restreaking.
Figure 4.
Qualitative evidence of pYR4 conjugation. (a) Restreak of two representative transconjugant colonies. (b) Restreak of two representative background colonies. (c-d) PCR to detect (c) pYR4 and (d) pGM101neo. L: Ladder; size (bp) shown left. Amplicon sizes shown right. + and – in (c) are pre-conjugation donor and recipient colonies, respectively. + and – in (d) are pre-conjugation recipient and donor colonies, respectively.
Figure 4.
Qualitative evidence of pYR4 conjugation. (a) Restreak of two representative transconjugant colonies. (b) Restreak of two representative background colonies. (c-d) PCR to detect (c) pYR4 and (d) pGM101neo. L: Ladder; size (bp) shown left. Amplicon sizes shown right. + and – in (c) are pre-conjugation donor and recipient colonies, respectively. + and – in (d) are pre-conjugation recipient and donor colonies, respectively.
3.4. Virulence-Related Phenotypes Encoded by pYR4
We assessed the burden of carrying pYR4 in various laboratory media but found no difference between NVH_3758 and NVH_3758_pYR4– in either growth rate or yield, regardless of media (Figure A4 in the Appendix).
Due to the presence of the putative TFP encoded by pYR4, we assessed twitching motility of Y. ruckeri NVH_3758 and NVH_3758_pYR4– on solid media at a range of temperatures. Results (Figure 5) showed poor motility of Y. ruckeri at all temperatures under these experimental conditions regardless of the presence or absence of pYR4 (P > 0.95), whilst a Pseudomonas aeruginosa PA14 positive control was highly motile (P < 0.0001 compared to both Y. ruckeri strains).
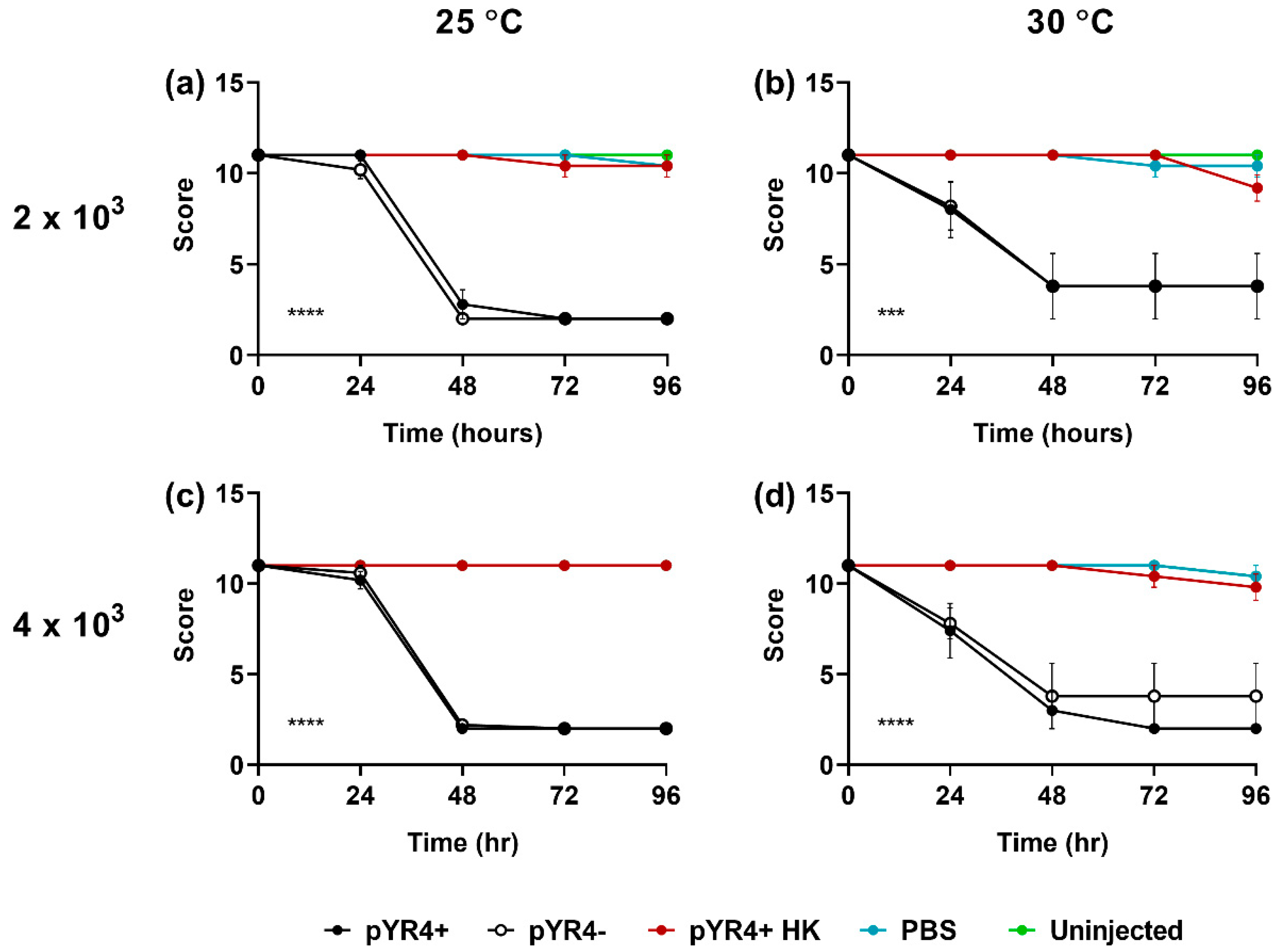
Lastly, we assessed the contribution of pYR4 to Y. ruckeri virulence by comparing NVH_3758 and NVH_3758_pYR4– in a Galleria mellonella infection assay at a range of temperatures and infectious doses. Results (Figure 6) showed that G. mellonella larvae succumbed rapidly to Y. ruckeri infection at all temperatures and doses tested, regardless of the presence or absence of pYR4 (P ≥ 0.7288 between the two score curves in Figure 6; survival curves shown in Figure A5 in the Appendix). On the contrary, larvae left uninjected or those injected with PBS only or heat-killed Y. ruckeri remained healthy (overall effect of injection type: P ≤ 0.0001 under all conditions).
4. Discussion
In this study, we investigated the role of the higBA locus in maintaining the plasmid pYR4 in the Y. ruckeri strain NVH_3758. We demonstrated that HigBA is a functional TA system that contributes directly to the stability of pYR4, as deletion of this system led to significant loss of the plasmid in a relatively short time (6 days, representing approximately 60 generations). Furthermore, providing the higA antitoxin gene in trans in pYR4+ cells allowed efficient curing of the plasmid; something that had failed during several previous efforts, both in this study and a previous one [13]. However, neutralizing the HigB toxin only led to ~50% loss of the plasmid even after a 10-day incubation, suggesting other components also contribute to the stability of pYR4. One of these could be the intact CbtA TA system also identified on pYR4; future work will determine its role in stabilizing pYR4 and any compound effects this may have with the HigBA system.
HigBA was originally characterized as a plasmid maintenance element in Proteus spp. [36] but has since been found in a wide range of pathogenic bacteria species including V. cholerae [37] and both enterohaemorrhagic and uropathogenic E. coli [38]. HigB is part of the RelE superfamily of endoribonuclease toxins and is encoded upstream of its cognate antitoxin, which is unusual for type II TA systems [20]. This gene organization is conserved on pYR4 and the related Y. ruckeri plasmid, pYR3. Type II TA systems, including HigBA, autoregulate operon expression via binding of the TA complex to their own promoters, in a process known as conditional cooperativity [33,34]. We cloned higA under the control of its own promoter and showed that this construct was able to abrogate toxicity of HigB produced from an induced pBAD33 promoter, suggesting that this regulation mechanism remains intact for the pYR4 HigBA module.
The higBA locus is part of a ‘stability cluster’ found in pYR4, and our re-analysis of this region shows that some of this cluster, including higBA, is shared with pYR3. Therefore, it is reasonable to assume that also pYR3 is stabilized by higBA. The cbtA type IV TA locus we identified is also shared between these two plasmids, but it is intact only in pYR4, which bolsters the argument for higBA being the main stabilizing factor of both plasmids. The stability of other Y. ruckeri plasmids has not been investigated; it is therefore not known whether the high stability of pYR4 (and presumably pYR3) mediated by higBA is unusual among Y. ruckeri plasmids, or if the others are similarly stable through some other mechanisms. Of note, pYR3 and pYR4 also encode one and two putative partitioning systems, respectively, within their stability clusters. These may contribute to plasmid maintenance, though the systems differ between the two plasmids.
Based on the presence of a T4SS, pYR4 was previously presumed to be conjugative [13]. However, the related tra operon was also implicated in virulence in a different strain of Y. ruckeri, and deletion of traI led to reduction in virulence in a fish model [16]. The pYR4 T4SS has similarities to the Icm/Dot T4SS of Legionella pneumophila, which translocates effector proteins into eukaryotic host cells [39]. The pYR4 tra thus encodes an ‘expanded’ T4SS, which are generally virulence-related but can also be conjugation systems [15]. We provide evidence here that pYR4 T4SS is conjugative, rather than a virulence-related effector delivery system. We were able to observe transfer of antibiotic resistance gene-tagged pYR4 into a pYR4-negative recipient. Though we could not quantify the rate of transfer due to background growth, regardless of multiple attempts with different recipient strain markers, we could identify genuine transconjugants by PCR. These transconjugants were able to grow well when restreaked on counterselection medium, in contrast to PCR-negative colonies on the original counterselection plates. This demonstrates transfer of the plasmid and strongly suggests that the T4SS mediates conjugation. pYR4 does not encode any obvious effector proteins, further strengthening the argument that the T4SS is purely conjugative [13]. However, we cannot currently conclusively rule out whether the T4SS translocates effectors encoded on the chromosome.
If the T4SS is not virulence-related, this suggests that the TFP encoded by an adjacent locus might be responsible for the reduction in virulence observed in a previous study [16]. In our study, we did not observe twitching motility, but this is not the only function attributed to TFP. The pYR4-encoded pil locus could be involved in biofilm formation, adhesion to particular surfaces, or possibly aid T4SS-mediated conjugation by bringing bacterial cells into close proximity. TFP have been demonstrated to promote conjugation for some plasmids and other mobile genetic elements [40,41,42]. It is also possible that we did not test for twitching motility under the right conditions. We tested three temperatures, which are close to the optimum growth temperature of 28 °C but higher than the temperatures in which Y. ruckeri generally causes disease in fish, in waters below 20 °C [43]. Previously, the T4SS of Y. ruckeri 150R was found to be more highly expressed at 18 °C compared with 28 °C [16], and more recently higher expression was observed for many other genes at 18 °C [44]. Therefore, it is possible that the TFP may be active in generating twitching motility at lower temperatures than those we tested.
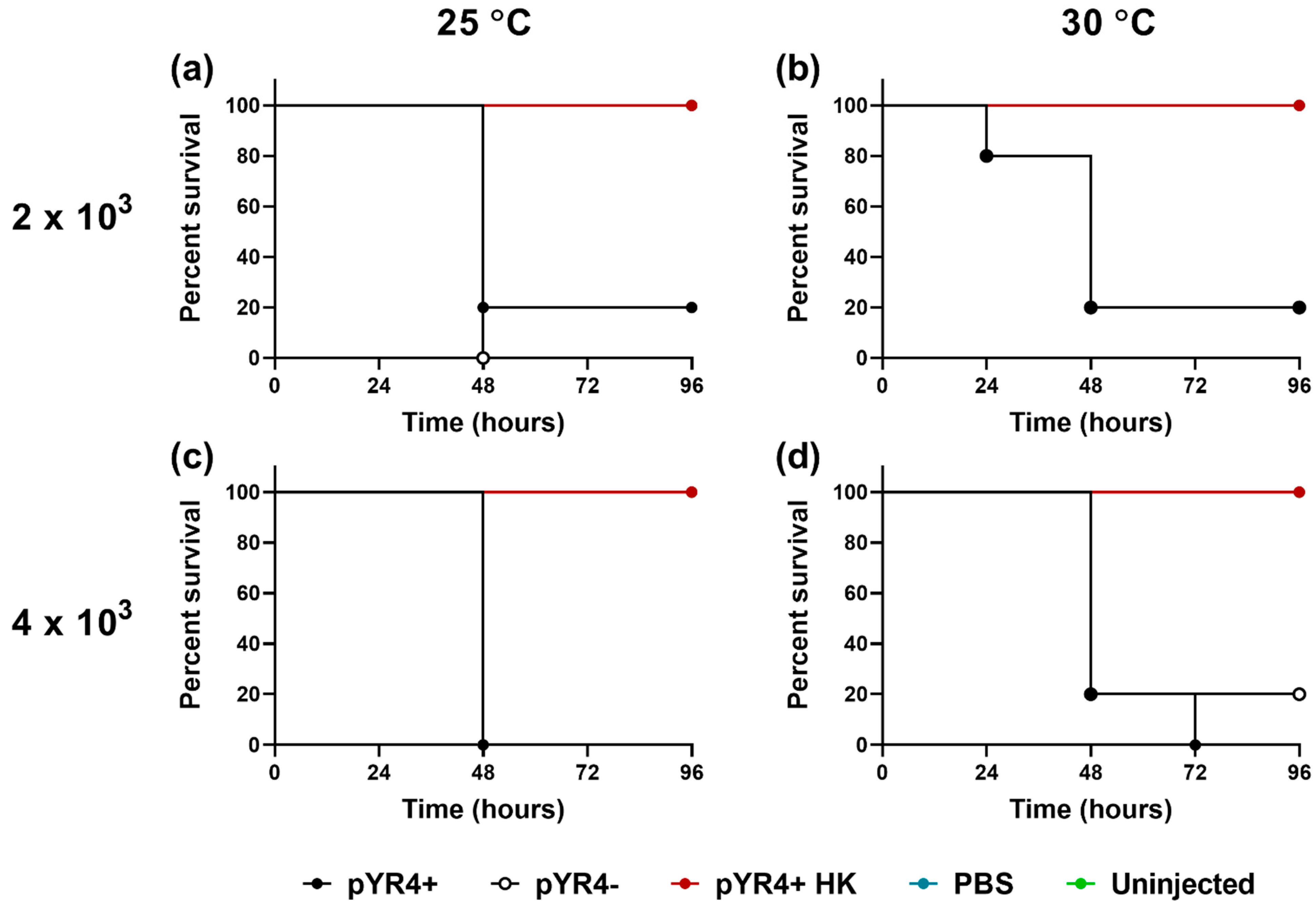
To test the hypothesis of pYR4 being a virulence plasmid, we performed infection experiments using the Galleria mellonella larvae model. This model has been used before for Y. ruckeri [45]. Previous experiments were performed at the mammalian body temperature of 37 °C, higher than the temperature optimum of Y. ruckeri. In our experiments, we tested lower temperatures, 30 °C and 25 °C. At these temperatures, ≥80% of larvae succumbed within 48 hours of infection, even with a dose (2 x 103 CFU) lower than one that resulted in only ~20 % mortality after 96 hours at 37 °C. This suggests that Y. ruckeri is indeed more virulent at lower temperatures, as proposed before [44]. We did not observe major differences in overall health scores (Figure 5) or mortality rates (Figure A4) between the tested temperatures. Nonetheless, our results suggest that if the G. mellonella model is used in the future for assessing Y. ruckeri virulence, temperatures below 30 °C should be preferred.
Contrary to our hypothesis, we did not see any effect of curing pYR4 on virulence. Again, this could be because we tested virulence at a suboptimal temperature, and the role of this plasmid would only become evident at even lower temperatures. A more probable reason is that the G. mellonella model is a very crude one, with bacteria injected directly into the haemocoel of the larvae. This is a poor mimic for the infection route in the natural fish host, where the bacteria probably enter through the gills [5], and the G. mellonella model may bypass early steps in the infection process where pYR4 plays a role. Therefore, to conclusively determine whether pYR4 (and by extension pYR3 and others) are virulence plasmids, experiments should be conducted in salmonid fish.
5. Conclusions
We have shown that the higBA locus of pYR4 encodes a functional TA system, the first to be characterized for Y. ruckeri. This system contributes to the high stability of this plasmid and deleting the toxin component higB or adding the higA antitoxin in trans allowed curing the plasmid efficiently. We further provided qualitative evidence to show that pYR4 is conjugative, suggesting the T4SS is involved in horizontal gene transfer rather than virulence. Surprisingly, deletion of pYR4 did not show a significant effect in Y. ruckeri virulence in the Galleria mellonella model. This may be because the model is not a good mimic for the natural fish host, or other conditions such as temperature were not optimal. Further research is needed to identify the role, if any, of pYR4 and other Y. ruckeri plasmids in pathogenesis.
Author Contributions
Conceptualization, G.M. and J.C.L.; infection assays, F.F. and M.S.; plasmid stability and conjugation, M.S.; cloning and toxicity assays, F.F. and V.W.; plasmid curing, C.M.; bioinformatics, L.H.; writing—original draft preparation, G.M. and J.C.L.; writing—review and editing, G.M. and J.C.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable
Data Availability Statement
All data are presented in the paper.
Acknowledgments
We thank Prof. Dirk Linke (University of Oslo) for providing the Y. ruckeri NVH_3758 strain. We are grateful to other members of the Antimicrobial Resistance, Omics and Microbiota group for support and discussions.
Conflicts of Interest
The authors declare no conflicts of interest.
Appendix A

Table A1.
Primers used in this study.
| Primer | Sequence (5’->3’) | Description |
|---|---|---|
| GMntu043 | GCATTTTTATCCATAAGATTAGCGG | pBAD33 MCS (forward) |
| GMntu044 | GCGTTCTGATTTAATCTGTATCAGG | pBAD33 MCS (reverse) |
| GMntu045 | CACCGTCATCACCGAAACG | pGM101 insert site (forward) |
| GMntu046 | CTGTTTTATCAGACCGCTTCTGC | pGM101 insert site (reverse) |
| GMmbiol057 | GGATCCTCTAGAGTCGACGCAAGGGACGAGCGTTAATGG | higB with pBAD33 overlap (forward) |
| GMmbiol058 | AAGCTTGCATGCCTGCAGCGGGGGCTTGTTATTCGATTTG | higB with pBAD33 overlap (reverse) |
| GMmbiol059 | AATCGAATAACAAGCCCCCGCTGCAGGCATGCAAGCTT | pBAD33 with higB overlap (forward) |
| GMmbiol060 | CATTAACGCTCGTCCCTTGCGTCGACTCTAGAGGATCC | pBAD33 with higB overlap (reverse) |
| GMmbiol061 | GCATGGCGGCTAAAGTTGTG | pYR4 repA (forward) |
| GMmbiol062 | GCTAGATTATGCCTGCTCGC | pYR4 repA (reverse) |
| GMmbiol063 | CCATGAGCAAGGGCGAATTGCT | pYR4 pilN (forward) |
| GMmbiol064 | GCTTCAGTCATCACGCTGACAT | pYR4 pilN (reverse) |
| GMmbiol065 | CCCGTATTTAGCAGGCGAAGAG | pYR4 higB (forward) |
| GMmbiol066 | GCTGACTTATCGATTTCAGGAC | pYR4 higB (reverse) |
| GMmbiol067 | CAGCGGAAAGTAGCTTG | Y. ruckeri 16S rDNA (forward) |
| GMmbiol068 | TGTTCAGTGCTATTAACACTTAA | Y. ruckeri 16S rDNA (reverse) |
| GMmbiol069 | CGCCCACAGGGTGCGCCGCTCGAAGCGGCATGCATTTACG | pGM101 with higBA overlap (forward) |
| GMmbiol070 | TTGCACTTTTTGAGATGATTCCTTCGCGCGCGAATTGATC | pGM101 with higBA overlap (reverse) |
| GMmbiol071 | GATCAATTCGCGCGCGAAGGAATCATCTCAAAAAGTGCAATTATTGC | higBA with pGM101 overlap (forward) |
| GMmbiol072 | CGTAAATGCATGCCGCTTCGAGCGGCGCACCCTGTGG | higBA with pGM101 overlap (reverse) |
| GMmbiol073 | TCAGCAAGGGACGAGCGTTACAAGCCCCCGCACTGCG | ΔhigB variant (forward) |
| GMmbiol074 | CCCCGCAGTGCGGGGGCTTGTAACGCTCGTCCCTTGCTGA | ΔhigB variant (reverse) |
| GMmbiol075 | GCAAGGGACGAGCGTTATACGAGTATCTAGAATTCATTGAG | higB mutation variant (forward) |
| GMmbiol076 | ATGAATTCTAGATACTCGTATAACGCTCGTCCCTTGCTGACCG | higB mutation variant (reverse) |
| MS103 | GGGAGAGCTCAAAAAAGCGACTTTAGCC | Upstream higBA (forward) |
| MS104 | TGAGATGATTATGACAGGTTATGAATTGC | Upstream higBA (reverse) |
| MS105 | AACCTGTCATAATCATCTCAAAAAGTGCAATTATTGCACTATTTTATATTTTTATTTAGCGAGCGTATACC | ΔhigA (forward) |
| MS106 | TAATTCCCATGAGCGGCGCACCCTGTGG | ΔhigA (reverse) |
| MS107 | TGCGCCGCTCATGGGAATTAGCCATGGTCC | Chloramphenicol cassette (forward) |
| MS108 | TGGCCAGTAAGTGTAGGCTGGAGCTGCTTC | Chloramphenicol cassette (reverse) |
| MS109 | CAGCCTACACTTACTGGCCACTTCCGTG | Downstream higBA (forward) |
| MS110 | TACCGCATGCAGCCGAAGCATATGTTTTG | Downstream higBA (reverse) |
Figure A1.
Construct design and PCR to detect successful mutagenesis of pYR4. (a) Map of the US-higA-cat-DS fragment used for lambda Red integration into pYR4. (b) PCR of the higBA region of pYR4. L: Ladder; size (bp) shown left. NT: no template control. WT: wild type pYR4 amplicon (2,280 bp). C: transformant with cat cassette integration downstream of intact higBA locus (3,313 bp). Δ: transformant with cat cassette integration downstream of higB deletion (2,989 bp).
Figure A1.
Construct design and PCR to detect successful mutagenesis of pYR4. (a) Map of the US-higA-cat-DS fragment used for lambda Red integration into pYR4. (b) PCR of the higBA region of pYR4. L: Ladder; size (bp) shown left. NT: no template control. WT: wild type pYR4 amplicon (2,280 bp). C: transformant with cat cassette integration downstream of intact higBA locus (3,313 bp). Δ: transformant with cat cassette integration downstream of higB deletion (2,989 bp).
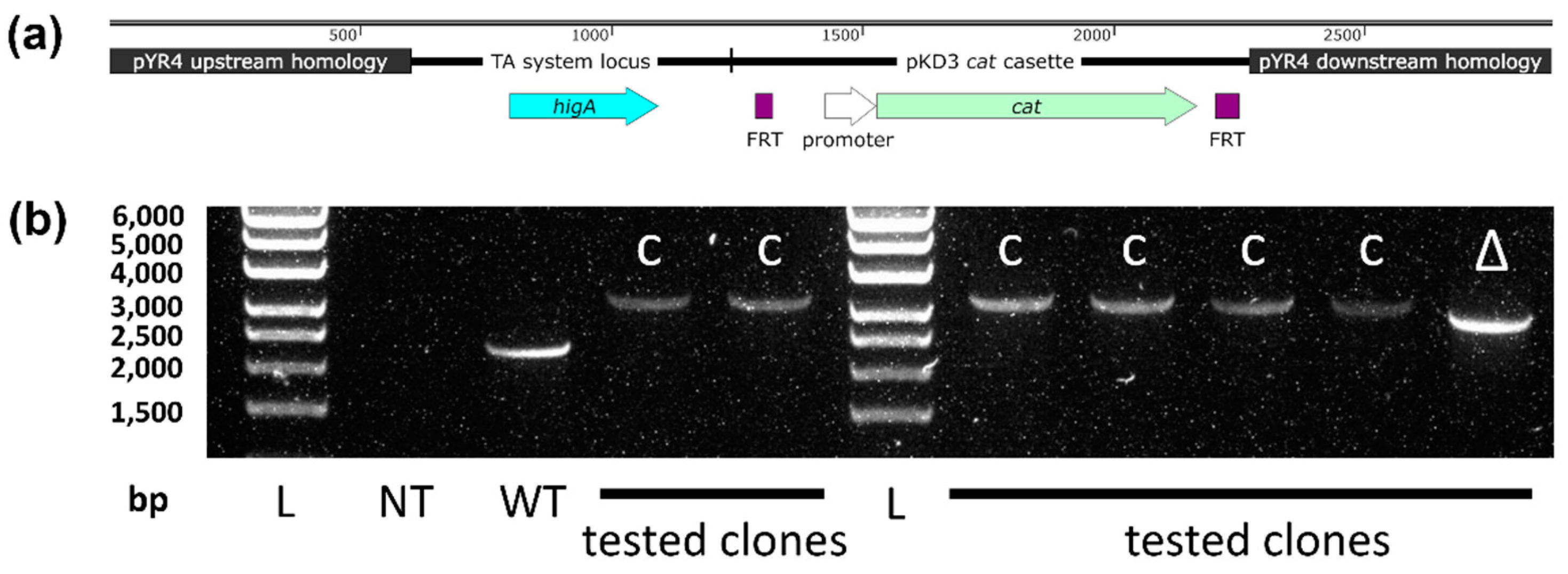
Figure A2.
Multiplex PCR design to detect pYR4. Taq amplification of individual gene targets and combined multiplex reaction are shown for both purified genomic DNA template and bacterial colony template. L: Ladder; size (bp) shown left. Amplicon sizes shown right.
Figure A2.
Multiplex PCR design to detect pYR4. Taq amplification of individual gene targets and combined multiplex reaction are shown for both purified genomic DNA template and bacterial colony template. L: Ladder; size (bp) shown left. Amplicon sizes shown right.
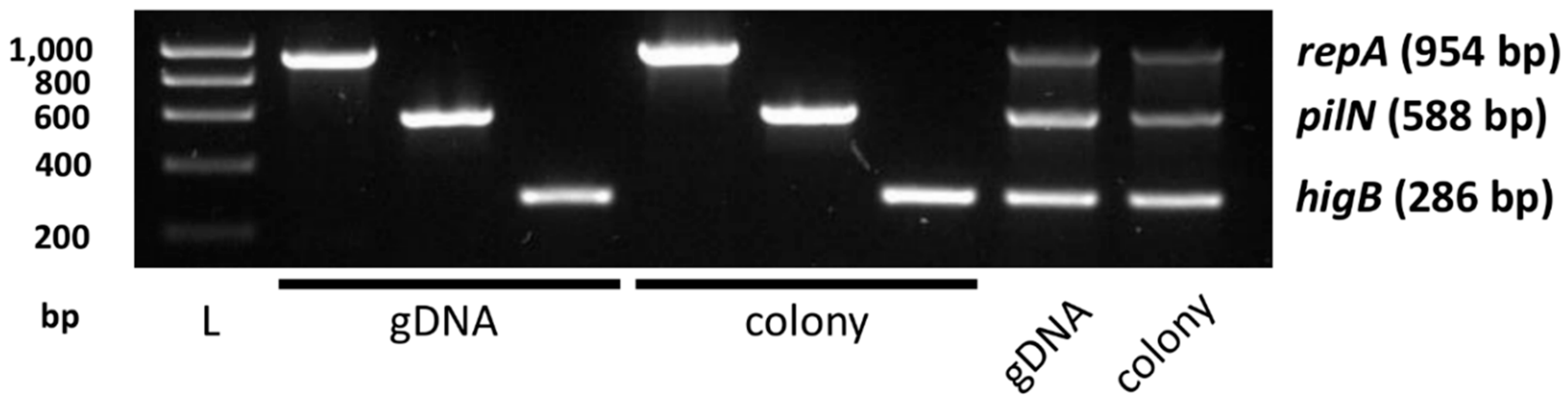
Figure A3.
Curing of pYR4 and helper plasmid from Y. ruckeri NVH_3758. Agarose gel electrophoresis showing (a) multiplex PCR results for colonies containing pGM101::higBmut after ten days of growth (white arrows denote pYR4-cured colonies; note the presence of the higB band due to pGM101::higBmut carriage); and eight fully plasmid-cured colonies tested for (b) pYR4 (via multiplex PCR), (c) pGM101::higBmut and (d) Y. ruckeri 16S rDNA. L: Ladder; size (bp) shown left. M: multiplex PCR positive control (unmodified Y. ruckeri NVH_3758 gDNA template). pG: pGM101::higBmut positive control (miniprep template). Amplicon sizes (bp) shown right.
Figure A3.
Curing of pYR4 and helper plasmid from Y. ruckeri NVH_3758. Agarose gel electrophoresis showing (a) multiplex PCR results for colonies containing pGM101::higBmut after ten days of growth (white arrows denote pYR4-cured colonies; note the presence of the higB band due to pGM101::higBmut carriage); and eight fully plasmid-cured colonies tested for (b) pYR4 (via multiplex PCR), (c) pGM101::higBmut and (d) Y. ruckeri 16S rDNA. L: Ladder; size (bp) shown left. M: multiplex PCR positive control (unmodified Y. ruckeri NVH_3758 gDNA template). pG: pGM101::higBmut positive control (miniprep template). Amplicon sizes (bp) shown right.
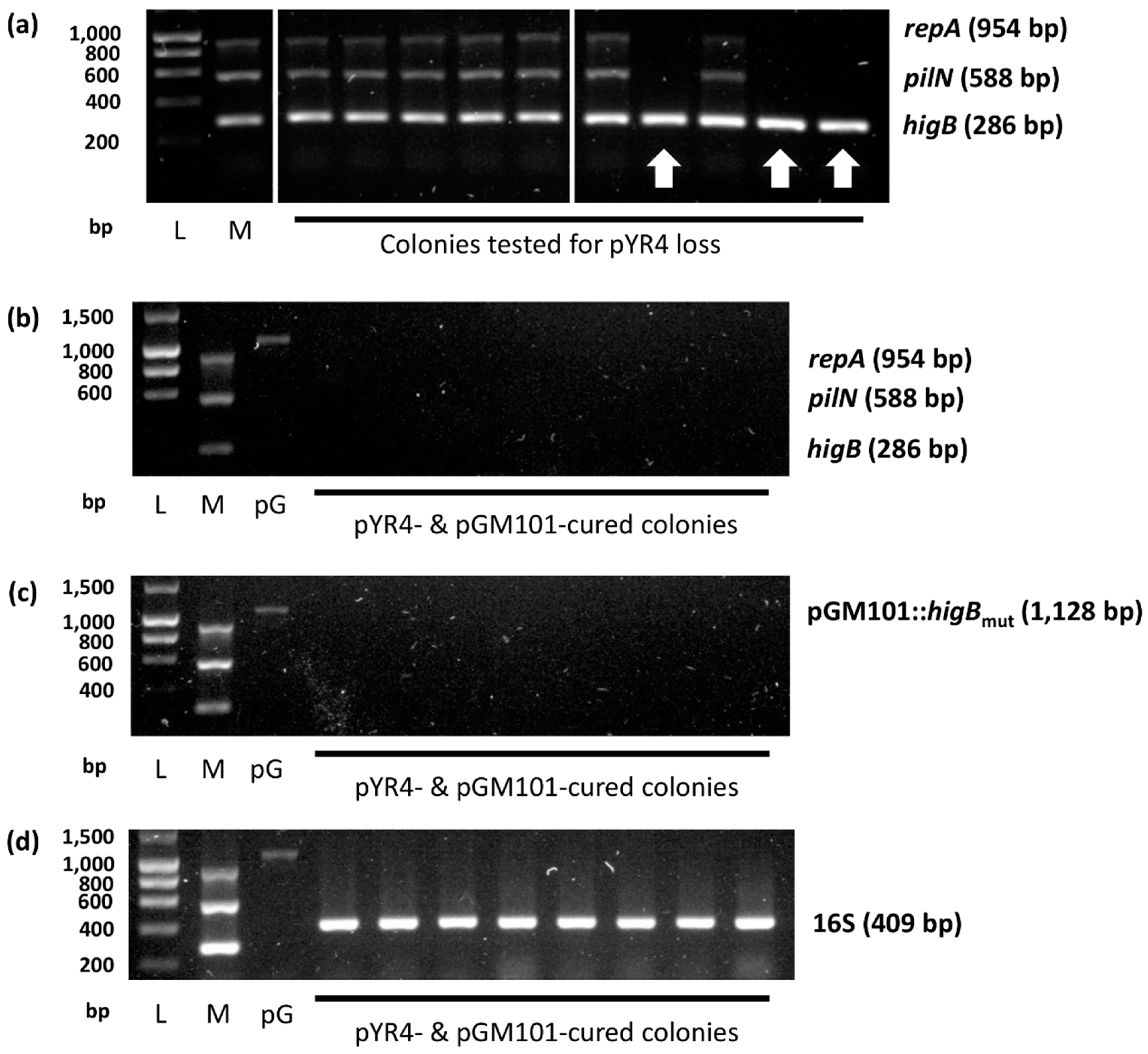
Figure A4.
Effect of pYR4 on growth. Mean OD600nm from three technical replicates of strains NVH_3758 (“pYR4+”) or NVH_3758_pYR4– (“pYR4-“) grown at 30 °C in LB (data repeated in each panel) compared to (a) M9 supplemented with glucose, thiamine, trace metals and vitamins; (b) M9 supplemented with magnesium and glycerol; (c) DMEM supplemented with glutamine; and (d) DMEM supplemented with glutamine and 10 % FBS. Representative data shown from multiple independent experiments.
Figure A4.
Effect of pYR4 on growth. Mean OD600nm from three technical replicates of strains NVH_3758 (“pYR4+”) or NVH_3758_pYR4– (“pYR4-“) grown at 30 °C in LB (data repeated in each panel) compared to (a) M9 supplemented with glucose, thiamine, trace metals and vitamins; (b) M9 supplemented with magnesium and glycerol; (c) DMEM supplemented with glutamine; and (d) DMEM supplemented with glutamine and 10 % FBS. Representative data shown from multiple independent experiments.
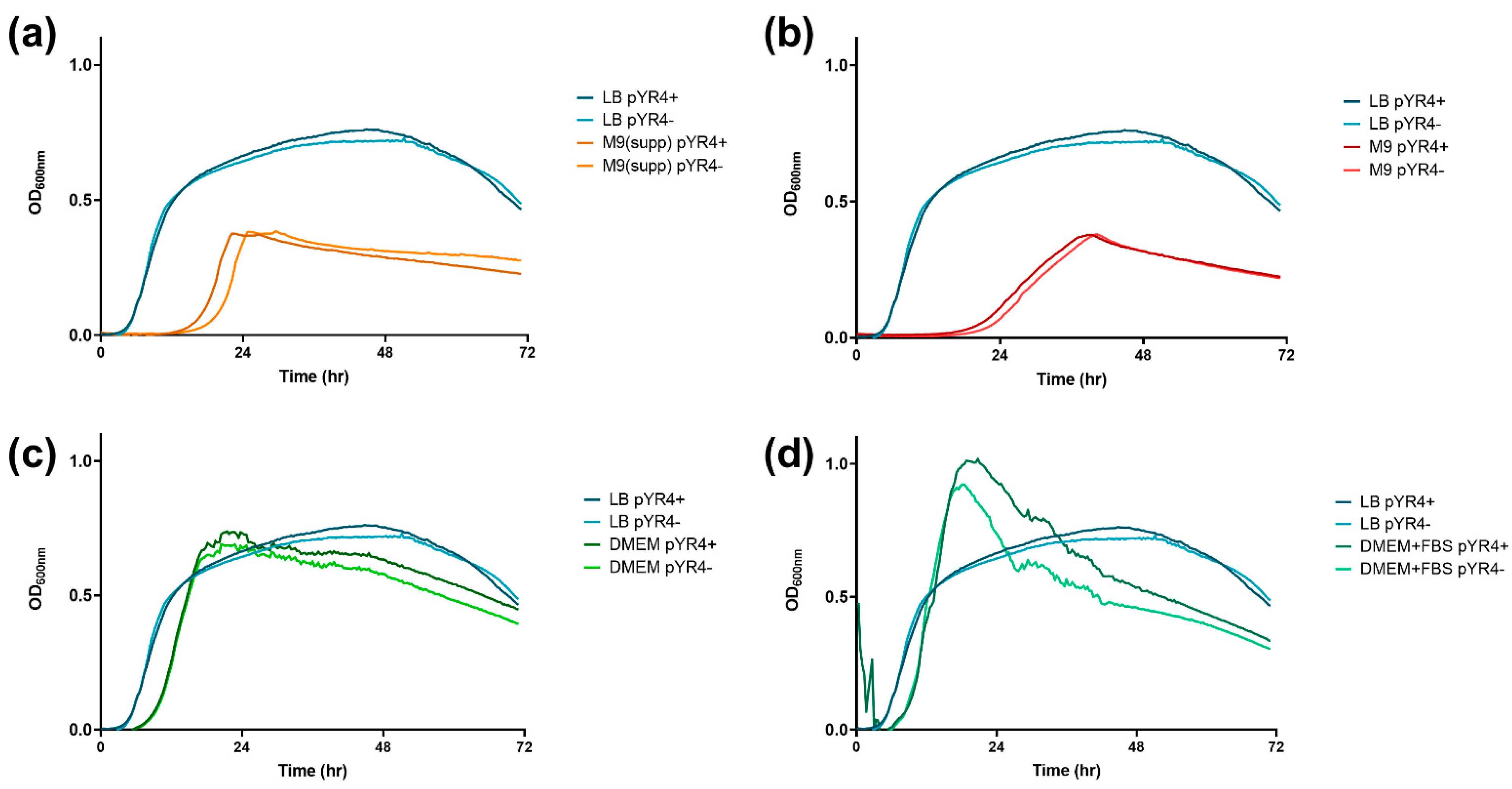
Figure A5.
The impact of pYR4 on Galleria mellonella survival. Larvae were left uninjected or were injected with 2 x 103 (a, b) or 4 x 103 (c, d) CFU Y. ruckeri NVH_3758 (“pYR4+”) or NVH_3758_pYR4– (“pYR4--”), or an equivalent volume of PBS or heat-killed Y. ruckeri (“pYR4+ HK”), as indicated. Larvae were incubated at either 25 °C (a, c) or 30 °C (b, d) for 96 hours and their survival recorded. Assay conducted on n = 5 larvae per experimental condition. Note that these data form part of the Galleria scoring experiment in Figure 5 and are shown here for convenience of comparison only.
Figure A5.
The impact of pYR4 on Galleria mellonella survival. Larvae were left uninjected or were injected with 2 x 103 (a, b) or 4 x 103 (c, d) CFU Y. ruckeri NVH_3758 (“pYR4+”) or NVH_3758_pYR4– (“pYR4--”), or an equivalent volume of PBS or heat-killed Y. ruckeri (“pYR4+ HK”), as indicated. Larvae were incubated at either 25 °C (a, c) or 30 °C (b, d) for 96 hours and their survival recorded. Assay conducted on n = 5 larvae per experimental condition. Note that these data form part of the Galleria scoring experiment in Figure 5 and are shown here for convenience of comparison only.
References
- FAO Fishery and Aquaculture Statistics – Yearbook 2021, FAO Yearbook of Fishery and Aquaculture Statistics: Rome, 2024.
- OECD/FAO. Fish. In OECD-FAO Agricultural Outlook 2023-2032OECD Publishing: Paris, 2023; pp. 214-224.
- Segner, H.; Sundh, H.; Buchmann, K.; Douxfils, J.; Sundell, K.S.; Mathieu, C.; Ruane, N.; Jutfelt, F.; Toften, H.; Vaughan, L. Health of farmed fish: its relation to fish welfare and its utility as welfare indicator. Fish Physiol Biochem 2011, 38, 85-105, DOI 10.1007/s10695-011-9517-9. Available online: https://link.springer.com/article/10.1007/s10695-011-9517-9.
- Stentiford, G.D.; Sritunyalucksana, K.; Flegel, T.W.; Williams, B.A.P.; Withyachumnarnkul, B.; Itsathitphaisarn, O.; Bass, D. New paradigms to help solve the global aquaculture disease crisis. PLoS Pathog 2017, 13, e1-e1006160, DOI 10.1371/journal.ppat.1006160. Available online: https://www.ncbi.nlm.nih.gov/pubmed/28152043.
- Kumar, G.; Menanteau-Ledouble, S.; Saleh, M.; El-Matbouli, M. Yersinia ruckeri, the causative agent of enteric redmouth disease in fish. Vet Res 2015, 46, 103, DOI 10.1186/s13567-015-0238-4. Available online: https://www.ncbi.nlm.nih.gov/pubmed/26404907.
- Fernandez-espinel, C.; Medina-morillo, M.; Irgang, R.; Sotil, G.; Araya-león, H.; Flores-dominick, V.; Romalde, J.L.; Avendaño-herrera, R.; Yunis-aguinaga, J. Co-existence of two Yersinia ruckeri biotypes and serotype O1a retrieved from rainbow trout (Oncorhynchus mykiss) farmed in Puno, Peru. J Fish Dis 2022, 46, 157-163, DOI 10.1111/jfd.13730. Available online: https://onlinelibrary.wiley.com/doi/abs/10.1111/jfd.13730.
- Riborg, A.; Colquhoun, D.J.; Gulla, S. Biotyping reveals loss of motility in two distinct Yersinia ruckeri lineages exclusive to Norwegian aquaculture. J Fish Dis 2022, 45, 641-653, DOI 10.1111/jfd.13590. Available online: https://onlinelibrary.wiley.com/doi/abs/10.1111/jfd.13590.
- Fouz, B.; Zarza, C.; Amaro, C. First description of non-motile Yersinia ruckeri serovar I strains causing disease in rainbow trout, Oncorhynchus mykiss (Walbaum), cultured in Spain. J Fish Dis 2006, 29, 339-346, DOI 10.1111/j.1365-2761.2006.00723.x. Available online: https://api.istex.fr/ark:/67375/WNG-03SWBFVW-X/fulltext.pdf.
- Wrobel, A.; Leo, J.C.; Linke, D. Overcoming fish defences: the virulence factors of Yersinia ruckeri. Genes 2019, 10, 700, DOI 10.3390/genes10090700. Available online: https://www.ncbi.nlm.nih.gov/pubmed/31514317.
- Carniel Arniel, E. Plasmids and Pathogenicity Islands of Yersinia. Curr Top Microbiol Immunol 2002, 264, 89-108, DOI 10.1007/978-3-642-56031-6_6. Available online: http://link.springer.com/10.1007/978-3-642-56031-6_6.
- Andreopoulos, W.B.; Geller, A.M.; Lucke, M.; Balewski, J.; Clum, A.; Ivanova, N.N.; Levy, A. Deeplasmid: deep learning accurately separates plasmids from bacterial chromosomes. Nucleic Acids Res 2021, 50, e17, DOI 10.1093/nar/gkab1115. Available online: https://www.ncbi.nlm.nih.gov/pubmed/34871418.
- Riborg, A.; Gulla, S.; Fiskebeck, E.Z.; Ryder, D.; Verner-Jeffreys, D.W.; Colquhoun, D.J.; Welch, T.J. Pan-genome survey of the fish pathogen Yersinia ruckeri links accessory- and amplified genes to virulence. PLoS ONE 2023, 18, e0285257, DOI 10.1371/journal.pone.0285257. Available online: https://www.ncbi.nlm.nih.gov/pubmed/37167256.
- Wrobel, A.; Ottoni, C.; Leo, J.C.; Linke, D. pYR4 from a Norwegian isolate of Yersinia ruckeri is a putative virulence plasmid encoding both a type IV pilus and a type IV secretion system. Front Cell Infect Microbiol 2018, 8, 373, DOI 10.3389/fcimb.2018.00373. Available online: https://www.frontiersin.org/articles/10.3389/fcimb.2018.00373/full.
- Pelicic, V. Type IV pili: e pluribus unum? Mol Microbiol 2008, 68, 827-837, DOI 10.1111/j.1365-2958.2008.06197.x. Available online: https://onlinelibrary.wiley.com/doi/abs/10.1111/j.1365-2958.2008.06197.x.
- Costa, T.R.D.; Patkowski, J.B.; Macé, K.; Christie, P.J.; Waksman, G. Structural and functional diversity of type IV secretion systems. Nat Rev Microbiol 2023, 22, 170-185, DOI 10.1038/s41579-023-00974-3. Available online: https://www.ncbi.nlm.nih.gov/pubmed/37814112.
- Méndez, J.; Fernández, L.; Menéndez, A.; Reimundo, P.; Pérez-Pascual, D.; Navais, R.; Guijarro, J.A. A chromosomally located traHIJKCLMN operon encoding a putative type IV secretion system is involved in the virulence of Yersinia ruckeri. Appl Environ Microbiol 2009, 75, 937-945, DOI 10.1128/AEM.01377-08. Available online: http://aem.asm.org/content/75/4/937.abstract.
- Qiu, J.; Zhai, Y.; Wei, M.; Zheng, C.; Jiao, X. Toxin–antitoxin systems: Classification, biological roles, and applications. Microbiol Res 2022, 264, 127159, DOI 10.1016/j.micres.2022.127159. Available online: https://dx.doi.org/10.1016/j.micres.2022.127159.
- Harms, A.; Brodersen, D.E.; Mitarai, N.; Gerdes, K. Toxins, targets, and triggers: An overview of toxin-antitoxin biology. Mol Cell 2018, 70, 768-784, DOI 10.1016/j.molcel.2018.01.003. Available online: https://dx.doi.org/10.1016/j.molcel.2018.01.003.
- Chan, W.T.; Espinosa, M.; Yeo, C.C. Keeping the wolves at bay: Antitoxins of prokaryotic type II toxin-antitoxin systems. Front Mol Biosci 2016, 3, 9, DOI 10.3389/fmolb.2016.00009. Available online: https://www.frontiersin.org/journals/molecular-biosciences/articles/10.3389/fmolb.2016.00009/full.
- Tian, Q.B.; Ohnishi, M.; Murata, T.; Nakayama, K.; Terawaki, Y.; Hayashi, T. Specific protein–DNA and protein–protein interaction in the hig gene system, a plasmid-borne proteic killer gene system of plasmid Rts1. Plasmid 2001, 45, 63-74, DOI 10.1006/plas.2000.1506. Available online: https://dx.doi.org/10.1006/plas.2000.1506.
- Christensen-Dalsgaard, M.; Gerdes, K. Two higBA loci in the Vibrio cholerae superintegron encode mRNA cleaving enzymes and can stabilize plasmids. Mol Microbiol 2006, 62, 397-411, DOI 10.1111/j.1365-2958.2006.05385.x. Available online: https://onlinelibrary.wiley.com/doi/abs/10.1111/j.1365-2958.2006.05385.x.
- Wrobel, A.; Ottoni, C.; Leo, J.C.; Gulla, S.; Linke, D. The repeat structure of two paralogous genes, Yersinia ruckeri invasin (yrInv) and a “Y. ruckeri invasin-like molecule”, (yrIlm) sheds light on the evolution of adhesive capacities of a fish pathogen. Journal of structural biology 2018, 201, 171-183, DOI 10.1016/j.jsb.2017.08.008. Available online: https://dx.doi.org/10.1016/j.jsb.2017.08.008.
- Wasteson, U.; Hvaal, A.; Serum, H.; Myhr, E.; Fossum, K. Antibacterial spectrum and some other characteristics of an antimicrobial factor produced by Yersinia ruckeri. Acta Vet Scand 1989, 30, 253-257, DOI 10.1186/BF03548029. Available online: https://www.ncbi.nlm.nih.gov/pubmed/2629502.
- Bertani, G. Studies on lysogenesis. I. The mode of phage liberation by lysogenic Escherichia coli. Journal of bacteriology 1951, 62, 293-300, DOI 10.1128/jb.62.3.293-300.1951. Available online: https://www.ncbi.nlm.nih.gov/pubmed/14888646.
- AnonymousM9 minimal medium (standard). Cold Spring Harb Protoc 2010, 2010, pdb.rec12295, DOI 10.1101/pdb.rec12295. Available online: https://cshprotocols.cshlp.org/content/2010/8/pdb.rec12295.
- Gibson, D.G.; Chuang, R.; Hutchison, C.A.; Venter, J.C.; Smith, H.O.; Young, L. Enzymatic assembly of DNA molecules up to several hundred kilobases. Nat Methods 2009, 6, 343-345, DOI 10.1038/nmeth.1318. Available online: http://dx.doi.org/10.1038/nmeth.1318.
- Guzman, L.M.; Belin, D.; Carson, M.J.; Beckwith, J. Tight regulation, modulation, and high-level expression by vectors containing the arabinose PBAD promoter. J Bacteriol 1995, 177, 4121-4130, DOI 10.1128/jb.177.14.4121-4130.1995. Available online: http://jb.asm.org/content/177/14/4121.abstract.
- Mcvicker, G.; Tang, C.M. Deletion of toxin–antitoxin systems in the evolution of Shigella sonnei as a host-adapted pathogen. Nat Microbiol 2016, 2, 16204, DOI 10.1038/nmicrobiol.2016.204. Available online: https://www.nature.com/articles/nmicrobiol2016204.
- Datsenko, K.A.; Wanner, B.L. One-step inactivation of chromosomal genes in Escherichia coli K-12 using PCR products. Proc Natl Acad Sci U S A 2000, 97, 6640-6645, DOI 10.1073/pnas.120163297. Available online: https://www.jstor.org/stable/122690.
- Hossain, M.J.; Thurlow, C.M.; Sun, D.; Nasrin, S.; Liles, M.R. Genome modifications and cloning using a conjugally transferable recombineering system. Biotechnol Rep 2015, 8, 24-35, DOI 10.1016/j.btre.2015.08.005. Available online: https://www.sciencedirect.com/science/article/pii/S2215017X15000478.
- Turnbull, L.; Whitchurch, C.B. Motility assay: Twitching motility. Methods Mol Biol 2014, 1149, 73-86, DOI 10.1007/978-1-4939-0473-0_9. Available online: http://link.springer.com/10.1007/978-1-4939-0473-0_9.
- Serrano, I.; Verdial, C.; Tavares, L.; Oliveira, M. The virtuous Galleria mellonella model for scientific experimentation. Antibiotics 2023, 12, DOI 10.3390/antibiotics12030505.
- Schureck, M.A.; Meisner, J.; Hoffer, E.D.; Wang, D.; Onuoha, N.; Ei Cho, S.; Lollar, P.; Dunham, C.M. Structural basis of transcriptional regulation by the HigA antitoxin. Mol Microbiol 2019, 111, 1449-1462, DOI 10.1111/mmi.14229. Available online: https://onlinelibrary.wiley.com/doi/abs/10.1111/mmi.14229.
- Overgaard, M.; Borch, J.; Jørgensen, M.G.; Gerdes, K. Messenger RNA interferase RelE controls relBE transcription by conditional cooperativity. Mol Microbiol 2008, 69, 841-857, DOI 10.1111/j.1365-2958.2008.06313.x. Available online: https://onlinelibrary.wiley.com/doi/abs/10.1111/j.1365-2958.2008.06313.x.
- Robertson, J.; Nash, J.H.E. MOB-suite: software tools for clustering, reconstruction and typing of plasmids from draft assemblies. Microb Genom 2018, 4, e000206, DOI 10.1099/mgen.0.000206. Available online: https://search.datacite.org/works/10.6084/m9.figshare.6177188.
- Tian, Q.B.; Ohnishi, M.; Tabuchi, A.; Terawaki, Y. A new plasmid-encoded proteic killer gene system: cloning, sequencing, and analyzing hig locus of plasmid Rts1. Biochem Biophys Res Commun 1996, 220, 280-284, DOI 10.1006/bbrc.1996.0396. Available online: https://www.ncbi.nlm.nih.gov/pubmed/8645296.
- Christensen-Dalsgaard, M.; Gerdes, K. Two higBA loci in the Vibrio cholerae superintegron encode mRNA cleaving enzymes and can stabilize plasmids. Mol Microbiol 2006, 62, 397-411, DOI 10.1111/j.1365-2958.2006.05385.x. Available online: https://onlinelibrary.wiley.com/doi/abs/10.1111/j.1365-2958.2006.05385.x.
- Pandey, D.P.; Gerdes, K. Toxin-antitoxin loci are highly abundant in free-living but lost from host-associated prokaryotes. Nucleic Acids Res 2005, 33, 966-976, DOI 10.1093/nar/gki201. Available online: https://api.istex.fr/ark:/67375/HXZ-H200PTKH-F/fulltext.pdf.
- Yang, Y.; Mei, L.; Chen, J.; Chen, X.; Wang, Z.; Liu, L.; Yang, A. Legionella pneumophila-mediated host posttranslational modifications. J Mol Cell Biol 2023, 15, mjad032, DOI 10.1093/jmcb/mjad032. Available online: https://www.ncbi.nlm.nih.gov/pubmed/37156500.
- Allard, N.; Neil, K.; Grenier, F.; Rodrigue, S. The type IV pilus of plasmid TP114 displays adhesins conferring conjugation specificity and is important for DNA transfer in the mouse gut microbiota. Microbiol Spectr 2022, 10, e0230321, DOI 10.1128/spectrum.02303-21. Available online: https://www.ncbi.nlm.nih.gov/pubmed/35293798.
- Carter, M.Q.; Chen, J.; Lory, S. The Pseudomonas aeruginosa pathogenicity island PAPI-1 is transferred via a novel type IV pilus. J Bacteriol 2010, 192, 3249-3258, DOI 10.1128/jb.00041-10.
- Ishiwa, A.; Komano, T. PilV adhesins of plasmid R64 thin pili specifically bind to the lipopolysaccharides of recipient cells. Journal of Molecular Biology 2004, 343, 615-625, DOI 10.1016/j.jmb.2004.08.059. Available online: https://dx.doi.org/10.1016/j.jmb.2004.08.059. [CrossRef]
- Guijarro, J.A.; Cascales, D.; García-Torrico, A.I.; García-Domínguez, M.; Méndez, J. Temperature-dependent expression of virulence genes in fish-pathogenic bacteria. Front Microbiol 2015, 6, 700, DOI 10.3389/fmicb.2015.00700.
- Mendez, J.; Cascales, D.; Garcia-Torrico, A.I.; Guijarro, J.A. Temperature-dependent gene expression in Yersinia ruckeri: Tracking specific genes by bioluminescence during in vivo colonization. Front Microbiol 2018, 9, 1098, DOI 10.3389/fmicb.2018.01098. Available online: https://www.ncbi.nlm.nih.gov/pubmed/29887855.
- Wrobel, A.; Saragliadis, A.; Pérez-Ortega, J.; Sittman, C.; Göttig, S.; Liskiewicz, K.; Spence, M.H.; Schneider, K.; Leo, J.C.; Arenas, J.; Linke, D. The inverse autotransporters of Yersinia ruckeri, YrInv and YrIlm, contribute to biofilm formation and virulence. Environ Microbiol 2020, 22, 2939-55, DOI 10.1111/1462-2920.15051. Available online: https://www.ncbi.nlm.nih.gov/pubmed/32372498.
Figure 1.
Map of pYR4 and its stability cluster. Predicted gene functions are indicated by colours: dark blue, TFP and T4SS; grey, mobile genetic elements and insertion sequences; yellow, plasmid replication; cyan, stability elements; green, genes of other predicted function; lavender, hypothetical ORFs of unknown function. (a) Linearised map of pYR4. GC content (%) is shown above by colour (right) and graph height (left). Yellow bar shows the region expanded in the lower panel. (b) Expanded map of the ~18 kb stability cluster of pYR4 and its comparison to pYR3, indicating individual putative plasmid maintenance elements. Truncation of the cbtA gene in pYR3 (top) relative to pYR4 (bottom), and hence the start of sequence divergence of the plasmids, is marked by a vertical red arrow. Alignment anchored on the higBA locus. pYR3 ORFs beyond the putative stability cluster are not shown.
Figure 1.
Map of pYR4 and its stability cluster. Predicted gene functions are indicated by colours: dark blue, TFP and T4SS; grey, mobile genetic elements and insertion sequences; yellow, plasmid replication; cyan, stability elements; green, genes of other predicted function; lavender, hypothetical ORFs of unknown function. (a) Linearised map of pYR4. GC content (%) is shown above by colour (right) and graph height (left). Yellow bar shows the region expanded in the lower panel. (b) Expanded map of the ~18 kb stability cluster of pYR4 and its comparison to pYR3, indicating individual putative plasmid maintenance elements. Truncation of the cbtA gene in pYR3 (top) relative to pYR4 (bottom), and hence the start of sequence divergence of the plasmids, is marked by a vertical red arrow. Alignment anchored on the higBA locus. pYR3 ORFs beyond the putative stability cluster are not shown.
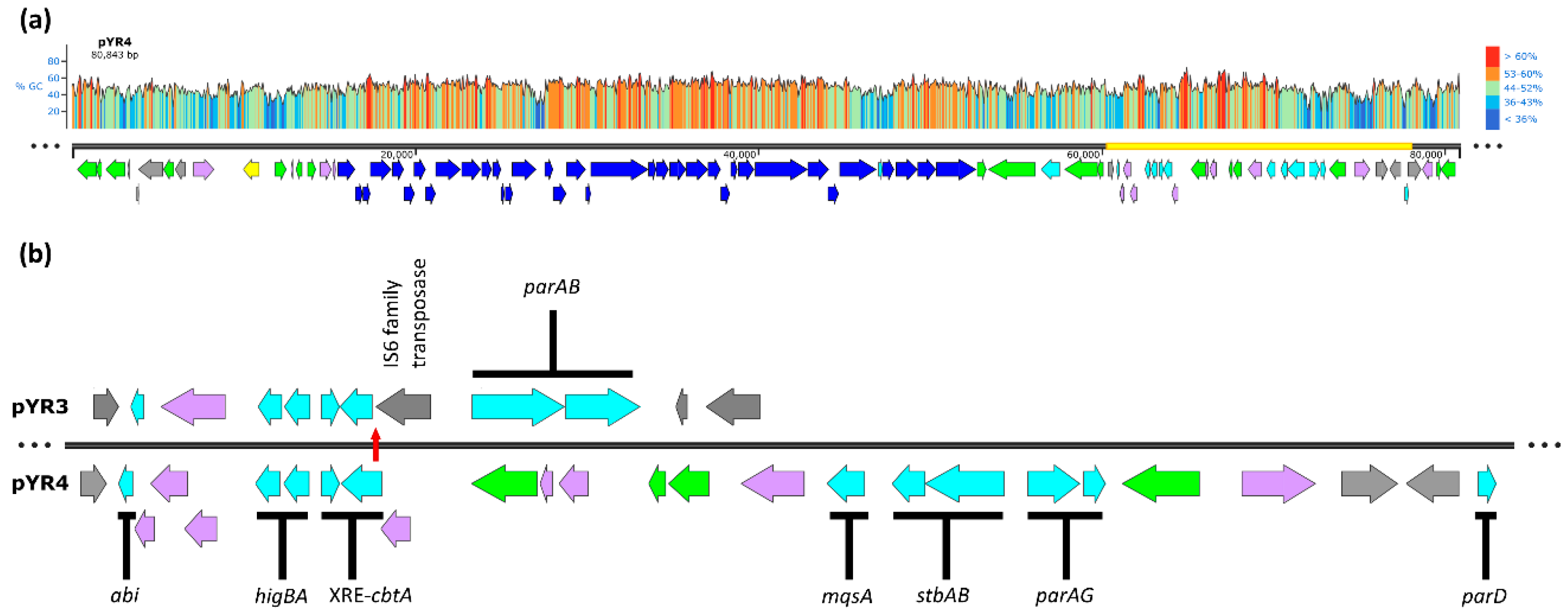
Figure 2.
pYR4 encodes a functional toxin-antitoxin, HigBA. (a) Map of the 812 bp region included in pGM101::higBA and its derivatives, indicating the deletion and mutation in pGM101::ΔhigB and pGM101::higBmut, respectively. Putative σ70 promoter predicted by SoftBerry BPROM (REF). (b-c) Viability of bacteria after induction of higB expression from pBAD33 in the presence of antitoxin-encoding vectors or empty vector control, as indicated, in either (b) E. coli DH5α or (c) Y. ruckeri NVH_3758_pYR4–. *** P < 0.001; **** P < 0.0001; overall effect of strain calculated by two-way ANOVA (n = 3 biological replicates). Bars show standard error of the mean.
Figure 2.
pYR4 encodes a functional toxin-antitoxin, HigBA. (a) Map of the 812 bp region included in pGM101::higBA and its derivatives, indicating the deletion and mutation in pGM101::ΔhigB and pGM101::higBmut, respectively. Putative σ70 promoter predicted by SoftBerry BPROM (REF). (b-c) Viability of bacteria after induction of higB expression from pBAD33 in the presence of antitoxin-encoding vectors or empty vector control, as indicated, in either (b) E. coli DH5α or (c) Y. ruckeri NVH_3758_pYR4–. *** P < 0.001; **** P < 0.0001; overall effect of strain calculated by two-way ANOVA (n = 3 biological replicates). Bars show standard error of the mean.
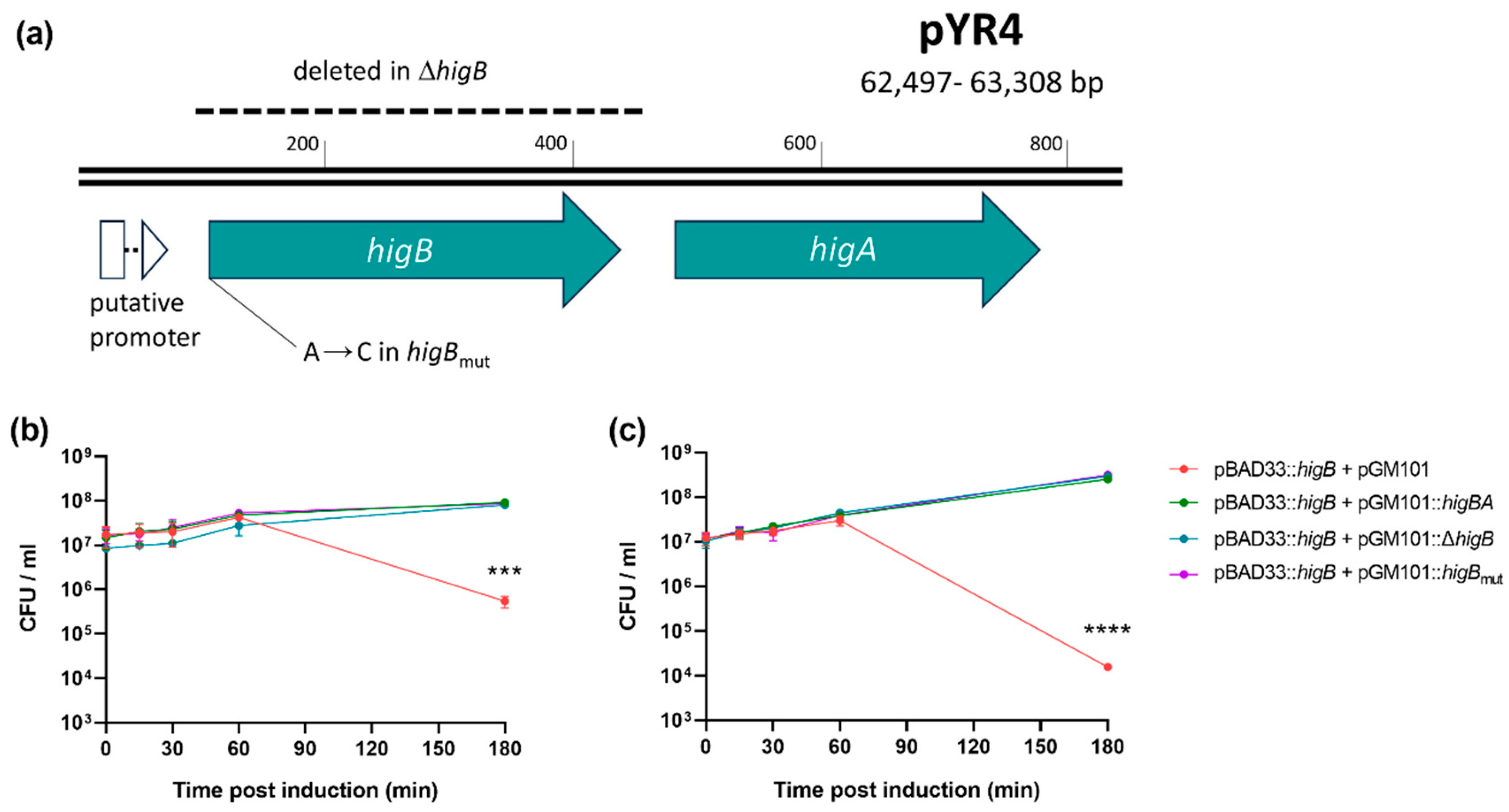
Figure 3.
Plasmid loss of pYR4 encoding (higB+) or lacking (ΔhigB) a functional HigBA TA system. Per cent plasmid-containing cells after the indicated number of daily subcultures in liquid media, calculated as the proportion of chloramphenicol-resistant versus total CFU grown on solid media. ns, not significant; * P < 0.05; **** P < 0.0001 by two-way ANOVA with Sidak’s multiple comparisons tests (n ≥ 3 biological replicates). Error bars show standard error of the mean.
Figure 3.
Plasmid loss of pYR4 encoding (higB+) or lacking (ΔhigB) a functional HigBA TA system. Per cent plasmid-containing cells after the indicated number of daily subcultures in liquid media, calculated as the proportion of chloramphenicol-resistant versus total CFU grown on solid media. ns, not significant; * P < 0.05; **** P < 0.0001 by two-way ANOVA with Sidak’s multiple comparisons tests (n ≥ 3 biological replicates). Error bars show standard error of the mean.
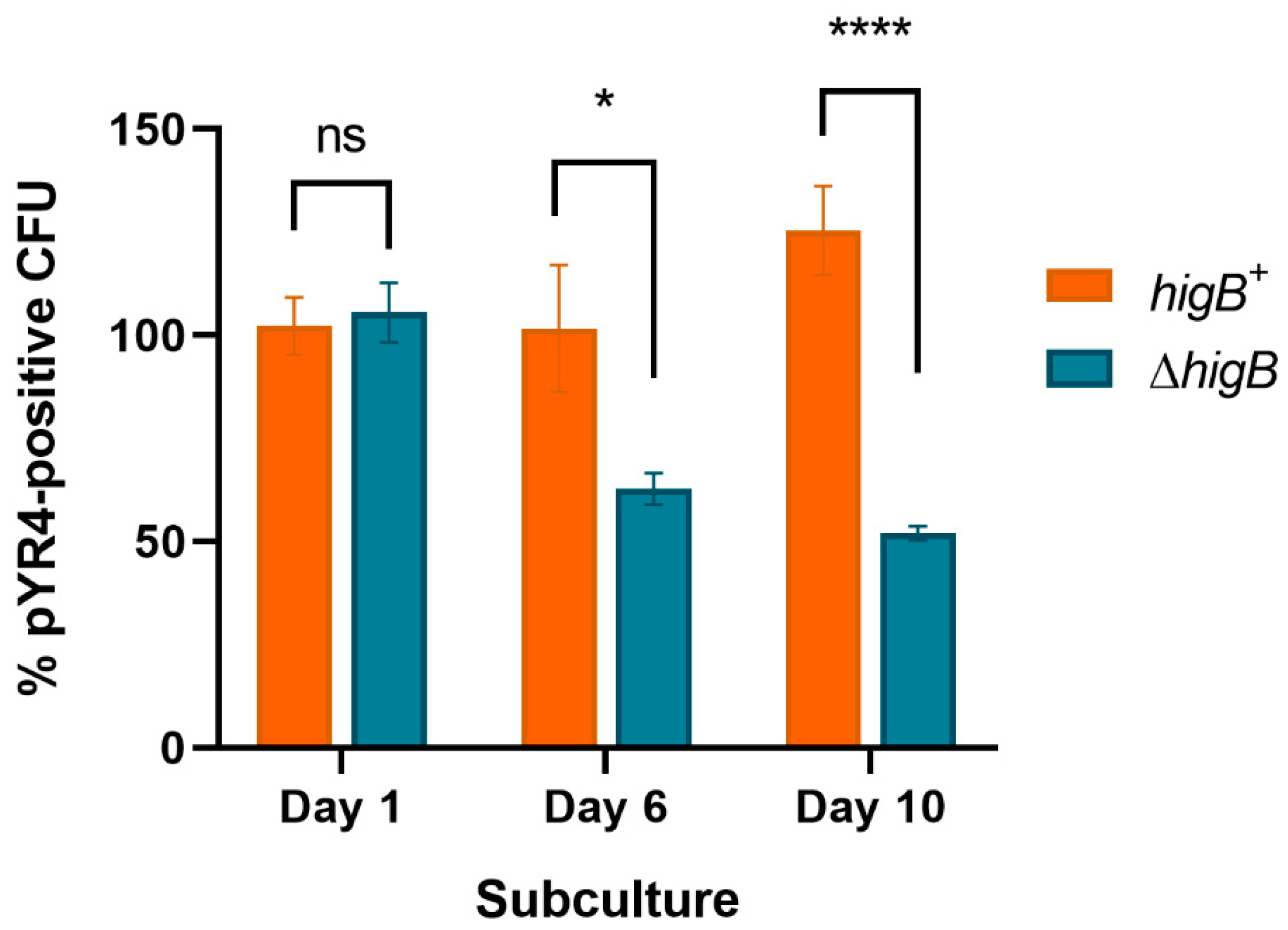
Figure 5.
Twitching motility of Y. ruckeri carrying (+) or not carrying (–) pYR4. Motility “halo” from stab point on solid media measured at the indicated temperatures after growth for 24 hours. P. aeruginosa PA14 included as a highly-motile control. ns, not significant; **** P < 0.0001 by two-way ANOVA with Tukey’s multiple comparisons tests (n = 3 biological replicates). Error bars show standard error of the mean.
Figure 5.
Twitching motility of Y. ruckeri carrying (+) or not carrying (–) pYR4. Motility “halo” from stab point on solid media measured at the indicated temperatures after growth for 24 hours. P. aeruginosa PA14 included as a highly-motile control. ns, not significant; **** P < 0.0001 by two-way ANOVA with Tukey’s multiple comparisons tests (n = 3 biological replicates). Error bars show standard error of the mean.
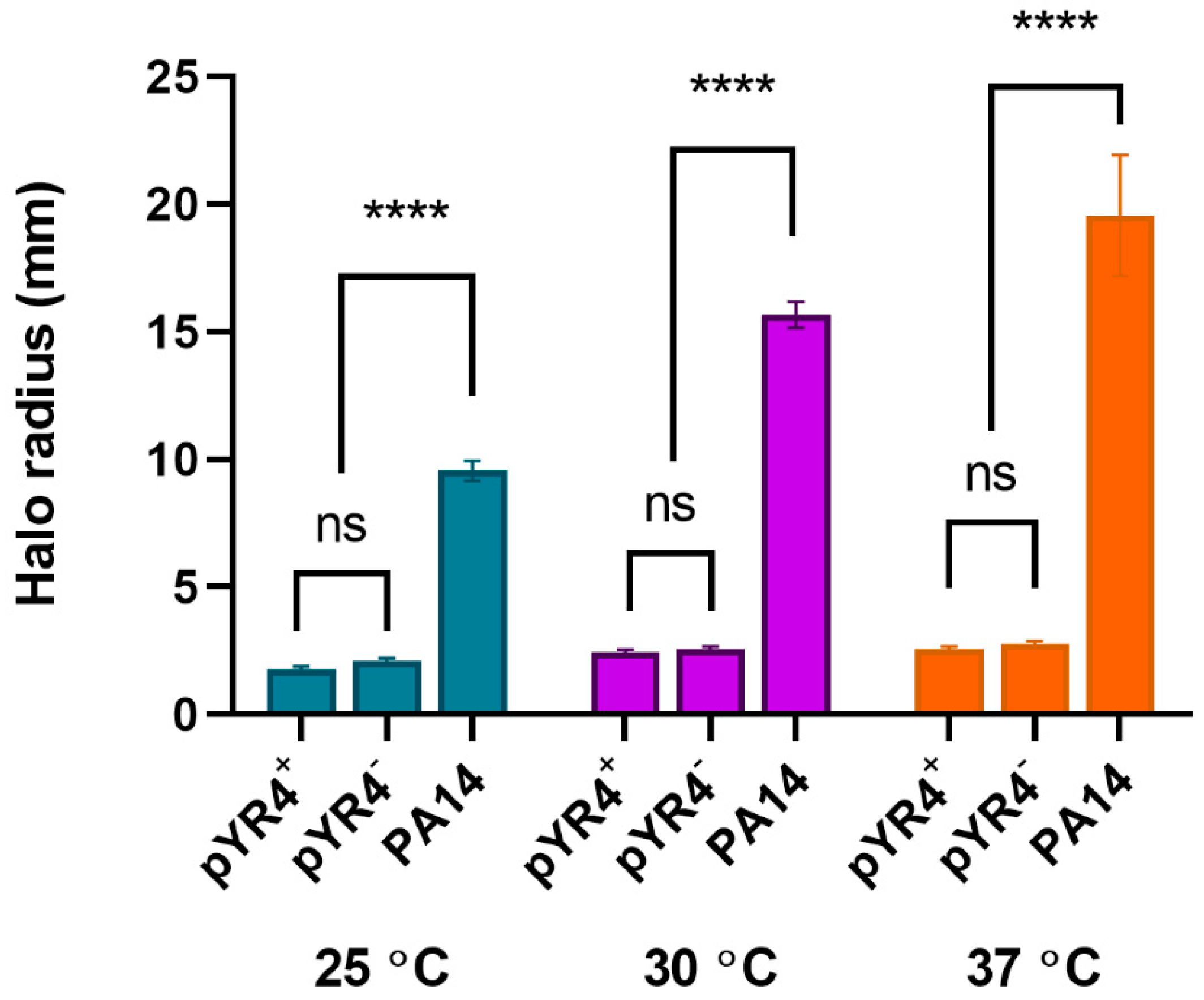
Figure 6.
The impact of pYR4 in a Galleria mellonella model of infection. Larvae were left uninjected or were injected with 2 x 103 (a, b) or 4 x 103 (c, d) CFU Y. ruckeri NVH_3758 (“pYR4+”) or NVH_3758_pYR4– (“pYR4--”), or an equivalent volume of PBS or heat-killed Y. ruckeri (“pYR4+ HK”), as indicated. Larvae were incubated at either 25 °C (a, c) or 30 °C (b, d) for 96 hours and their health scored every 24 hours according to the Galleria scoring system by Serrano et al. [32]. Bars show standard error of the mean. Statistical analysis by two-way ANOVA with Dunnett’s post test; overall effect of injection type shown in bottom left of each graph: *** P < 0.001; **** P < 0.0001. Assay conducted on n = 5 larvae per experimental condition.
Figure 6.
The impact of pYR4 in a Galleria mellonella model of infection. Larvae were left uninjected or were injected with 2 x 103 (a, b) or 4 x 103 (c, d) CFU Y. ruckeri NVH_3758 (“pYR4+”) or NVH_3758_pYR4– (“pYR4--”), or an equivalent volume of PBS or heat-killed Y. ruckeri (“pYR4+ HK”), as indicated. Larvae were incubated at either 25 °C (a, c) or 30 °C (b, d) for 96 hours and their health scored every 24 hours according to the Galleria scoring system by Serrano et al. [32]. Bars show standard error of the mean. Statistical analysis by two-way ANOVA with Dunnett’s post test; overall effect of injection type shown in bottom left of each graph: *** P < 0.001; **** P < 0.0001. Assay conducted on n = 5 larvae per experimental condition.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



