Submitted:
22 July 2024
Posted:
24 July 2024
You are already at the latest version
Abstract
The soybean cyst nematode (SCN), Heterodera glycines Ichinohe, is one of the most destructive plant-parasitic nematodes in soybean production worldwide. The use of resistant soybean is the most effective alternative for its management. However, SCN-resistant soybean cultivars with increased yield and favorable agronomic traits remain limited in the market. Here, we developed a new SCN-resistant soybean cultivar Nongqing 28 from the cross the female parent cultivar ‘An 02-318’ × a male parent line F2 (Hei 99-980 × America Xiaoheidou). Resistance evaluation suggested that Nongqing 28 displayed stable resistance to SCN race 3 in pot assays and the 4-year field experiments, including inhibition of SCN development and reduction in female and cyst numbers. The average yield of Nongqing 28 were 2,593 kg/ha and 2,660 kg/ha in the 2-year regional trails and the 1-year production trials, with yield increase of 6.2% and 8.1% compared with the local cultivar Nengfeng 18, respectively. The average seed-fat contents in Nongqing 28 reached 21.26%. Additionally, RNA-seq analysis revealed that the resistance of Nongqing 28 to SCN infection is involved in pathogen perception and defense activation, such as reactive oxygen species burst, calcium-mediated defense signaling, hormonal signaling, the MAPK signaling cascade, and phenylpropanoid biosynthesis. In summary, this study provides a detailed characterization of a novel SCN-resistant soybean cultivar with high-oil and yield potential.
Keywords:
soybean cyst nematode
; Heterodera glycines
; soybean
; resistance
; defense signaling
1. Introduction
Soybean (Glycine max (L.) Merrill) is an important legume crop that supplies essential oils and vegetable proteins for humans and animals and serves as a source of biofuel. However, soybean yield is substantially impacted by several biotic constraints, including soybean cyst nematode (SCN), Heterodera glycines, Ichinohe. Currently, SCN has spread worldwide and has emerged as one of the most destructive pathogens for soybean production globally [1,2]. In United States, it was previously estimated that SCN caused up to $32 billions of soybean yield losses from 1996 through 2016 [3]. In China, more than $120 million of soybean production is lost annually due to SCN infestation [1]. Notably, this pathogen is particularly prevalent in two main soybean producing regions, the Huang-Huai Valleys in Central-East China and Northeast China [4,5].
To reduce the yield losses caused by SCN, the use of cultural, chemical, and biological methods are the recommended management actions [6]. Generally, rotating soybean with non-host plants such as corn or wheat is partially effective because SCN cysts display resistant to adverse conditions, protecting the developing eggs for many years in soil. Additionally, crop rotation can lead to a decrease in the total soybean production. The use of chemical nematicides is limited and, in some cases, prohibited because of their environmental harm. In this context, the cultivation of SCN-resistant soybean cultivars provides an economically and environmentally viable option to control SCN [6].
Surveys indicated that most of the commercial SCN-resistant soybean varieties were derived from limited resistance sources such as PI 548402 (Peking), PI 88788, and PI 437654 [7,8]. Rhg1 (Peking-type rhg1-a and PI88788-type rhg1-b) and Rhg4 (GmSHMT08) are two major-effect quantitative trait loci (QTL) conferring these varieties resistance to SCN [9,10]. However, overuse of these resistant cultivars led to a genetic shift in SCN populations, eventually resulting in the development of virulent SCN races or HG types that can overcome some SCN-resistant sources [6]. To broaden the genetic basis for breeding SCN-resistant cultivars, big efforts have been made to identify new resistant varieties and QTLs. For example, few resistant soybean sources lacking the Rhg1 or Rhg4 loci such as PI 567516C [11] and Pingliang Xiaoheidou [12] have been identified. To date, more than 200 QTLs, including new SCN-resistant QTLs such as qSCN-PL10 [12] and SCN_7_1 [13], have been reported and mapped to 19 chromosomes (https://www.soybase.org/).
Heilongjiang is the largest province in total soybean areas planted, contributing to over 40% of the total soybean production in China. Since the launch of the ‘Soybean Planting Expansion Plan’, Heilongjiang has continuously increased its total planting area of soybeans. In 2022, it reached 4.93 million hectares from 3.90 million hectares in 2021, representing a 17.1 percent increase. Heilongjiang also has a longer history of SCN infestation, with SCN firstly reported in 1899 in Anda County of Heilongjiang [14]. Survey data demonstrated that the infestation of SCN in Heilongjiang has spread to at least 63 soybean-planting counties [15,16]. Heilongjiang possesses highly genetically diverse SCN populations, including eleven different HG types [5,16]. Moreover, it has been reported that the continuous planting of a single type of resistant cultivar such as ‘Peking’ and ‘Franklin’ has resulted in shifts in dominant SCN populations in this region [17].
However, the publicly available utilized resistance sources are limited for developing soybean resistant varieties in China [16,17,18]. Although more than 30 commercially registered soybean varieties with SCN resistance have been developed in Heilongjiang since 1990 [1,19], few SCN-resistant varieties with increased yield and favorable agronomic traits have been reported [20]. With this objective, we developed a new SCN-resistant soybean cultivar, Nongqing 28, from a cross between the female parent cultivar ‘An 02-318’ and a male parent line F2 (Hei 99-980 × America Xiaoheidou). We determined the characteristic of SCN resistance and agronomic traits in Nongqing 28. Furthermore, we conducted the transcriptome sequencing to gain insights into the molecular mechanisms of SCN resistance in Nongqing 28.
2. Materials and Methods
2.1. Soybean Cultivars and Nematode Source
Four soybean cultivars, including Heihe 99-980, Hefeng 25, Hartwig, and An 02-318, were used for breeding of Nongqing 28 in this study. A resistant soybean PI 437456 and two susceptible soybean cultivars, Lee and Hefeng 50, were used as controls in the resistance assessment to SCN. SCN HG Type 0 (race 3) was originally isolated from an SCN-infested field in Daqing County, Heilongjiang. The virulence of SCN population were determined using the race scheme [21]and the HG type test [22].
2.2. Breeding Nongqing 28 and Selection
Soybean pedigree breeding was employed for develop Nongqing 28 through crosses between An02-318 as the male parent and the female parental line F2 (Heihe 99-980 × America Xiaoheidou) in Anda County, Heilongjiang (Figure 1). The SCN-resistant cultivar An02-318 was developed from the cross the high-yielding parent Hefeng 25 and the SCN-resistant parent Hartwig before 2006 by the Daqing branch of Heilongjiang Academy of Agricultural Sciences. The hybrid seeds of the F2 (F2-1) generation of Heihe 99-980 × America Xiaoheidou was produced in 2006. The cross An02-318 and an individual F2-1 line of Hei 99-980 × America Xiaoheidou was made by artificial hybridization in Anda, and F0 seeds were harvested in the fall of 2007. Seeds of subsequent generations from F1 to F5 were self-bred in Anda. From 2012 to 2014, the seeds of generations from F6 to F8 were planted in SCN-infested fields for disease resistance testing in Daqing. During 2015-2017, the seeds of F8 generations were collected for field tests. During 2018-2022, seed yield, agronomic trials, and SCN resistance were assessed through regional and production trials at different locations.
2.3. Assessment of SCN Resistance
Evaluation of SCN resistance was performed using a pot assay in a greenhouse at Northeast Institute of Geography and Agroecology, Chinese Academy of Sciences, and by the Daqing branch of Heilongjiang Academy of Agricultural Sciences in SCN-infested fields. For the pot assay, two-week-old soybean plants in pots (7.0 cm diameter × 10.3 cm height) containing sterilized sand and soil (2:1) were inoculated with 1,000 second-stage juveniles (J2) of HG Type 0 (each plant) and maintained under controlled greenhouse conditions at 22-28 ℃ and a 16-hour light/8-hour dark photoperiod. At 5 and 10 days post inoculation (dpi), nematode development in the collected roots was surveyed by acid fusion staining [23]. The number of nematodes at the different developmental stages (J2, J3, and/or J4) was counted using a stereomicroscope. At 21 dpi, the white females on roots for each plant were count under a stereomicroscope. Ten replicates were included at each time point, and the experiment was repeated three times.
For the field assay, soybean seeds were planted in SCN-infested fields in May during 2018-2022, with the susceptible soybean cultivar Lee used as the control. Approximately 45 days after germination, soybean plants were harvested, and SCN females were extracted from both roots and soil. The numbers of females were counted under a stereomicroscope. The female index (FI) was calculated as follows: FI= (average number of females on each soybean cultivar/ average number of females on the susceptible check Lee) × 100 [22].
2.4. Analysis of Seed Protein and Fat Contents
Seeds were collected for each trial location during harvest and analyzed for protein and oil content. The crude protein content in soybean seeds was determined according to the national standards of China (GB2905-82) using the Semi-micro Kjeldahl method. The seed crude fats were measured according to the method described in the national standards of China (GB 2906-82).
2.5. RNA-Seq Analysis
Two-week-old Nongqing 28 plants were inoculated with SCN HG type 0. At 10 dpi, three biological roots samples from both SCN-infected and non-infected plants were prepared for RNA extraction and library construction. Total RNA was extracted using a commercial kit according to the manufacturer’s instructions. RNA integrity, purity, and concentrations were assessed using an Agilent 4200 system (Agilent Technologies, Waldbron, Germany) with a Nanodrop One (Thermo Fisher Scientific, MA, USA). Library construction and sequencing were performed by Guangdong Meg Gene Biotechnology Co., LTD (Guangzhou, China) using the illumina platform with 150-bp paired-end reads. High-quality reads were mapped against the Williams 82 soybean reference genome (Glycine max Wm82.a4.v1) with Hisat2 (v2.2.1). Gene expression was assessed using DESeq2 (v1.34.0). Only those genes with a false discovery rate (FDR) ≤0.05 and |log2(fold change)|≥ 1 were considered differentially expressed genes (DEGs).
2.6. Quantitative Real-Time (qRT)-PCR Analysis
Total RNA was purified from soybean roots by using an RNAPrep Pure Micro Kit (TianGen Biotech, Beijing, China) following the manufacturer’s instructions, including a DNase treatment step. First-strand cDNA was synthesized from 1.0 μg of total RNA using FastKing gDNA Dispelling RT SuperMix FastKing Kit (TianGen Biotech, Beijing, China). PCR reactions were performed in the LightCycler® 480 System with AceQ qPCR SYBR Green Master Mix (Vazyme, Nanjing, China) according to the manufacturer’s procedure. The primers used for qRT-PCR are listed in Supplementary Table S3. PCR cycles began with 5 minutes at 95 °C, followed by 40 cycles of 10 seconds at 95 °C and 30 seconds at 60 °C. The 2-∆∆Ct method was applied to calculate the relative expression of specific genes using GmUBQ3 as a reference control. All experiments were conducted with three independent biological replicates, each with three technical replicates.
2.7. Data Analysis
Data were subjected to one-way ANOVA using SPSS version 17.0 software (Chicago, IL, USA). Tukey’s HSD test (P < 0.05) was used for overall pairwise comparisons. The error bars in the figures represent the SE of means, and the significance level was set at p < 0.05.
3. Results
3.1. Main Characters of Nongqing 28
Nongqing 28 is a highly homozygous recombinant inbred line derived from an individual F13 generation single plant from the cross An 02-318 × the female parental line F2 (Heihe 99-980 × America Xiaoheidou) through 5-year screening in the SCN-infested fields, 2-year regional trials (2018-2019), and 1-year production trial (2019). Nongqing 28 (Reg. No. ‘Heishendou 20200006’) was released by Daqin branch of Heilongjiang Academy of Agricultural Sciences in 2020 as a new soybean cultivar due to its high seed yield potential and SCN resistance in Heilongjiang Province.
The main agronomic traits of Nongqing 28 were investigated in two continuous years (2018 and 2019). Nongqing 28 has white flowers, round leaves, gray pubescence, sickle-shaped pods, and a determinate growth habit. The growth period of Nongqing 28 is about 123 days, making it suitable for cultivation in the first accumulated temperature zone of Heilongjiang. The mature plant height is about 97 cm without branches (Figure 2). The seeds have a brown hilum and are yellow with smooth seed coats (Figure 2). The 100-seed weight of Nongqing 28 is approximately 19.0 g. Additionally, the average contents of protein and oil in the seeds of Nongqing 28 are 38.2% and 21.2%, respectively (Table 1). In 2018 and 2019, Nongqing28 was tested in regional trails, where the average yield of Nongqing28 was 2,593 kg/ha, resulting in a yield increase of 6.2% compared to the local cultivar Nengfeng 18 (Table 2). In 2019, Nongqing 28 was tested in the production trials, where it produce an average yield of 2,660 kg/ha, increasing by 8.1% at five locations compared with the local cultivar Nengfeng 18 (Table 3). These data suggest that Nengfeng18 increased the yield, exceeding the test local soybean cultivar.
3.2. SCN Resistance of Nongqing 28
In the field assay during 2018-2021, SCN reproduction were significantly inhibited in Nongqing 28, compared to the susceptible cultivars Lee and Hefeng 50. The average number of cysts in Lee and Hefeng 50 were up to 69 and 67 in the 4-year field experiments, respectively (Table 3). However, the average number of cysts in Nongqing 28 were about 3, with FI values ranging from 3.08 to 4.76 (FI < 10%), suggesting a resistance level close to that of the resistant control PI 437456 (Table 3). These results indicated that Nongqing 28 exhibited resistant to SCN under field conditions.
The development of SCN in Nongqing 28 was further assessed in the greenhouse (Figure 2 and Table 4). During the early developmental stage, similar sizes of SCN in the roots of Nongqing 28, PI 437456 and Hefeng 50 were observed at 5 dpi. However, the number of J3 in Nongqing 28 and PI 437456 was lower than in Hefeng 50 roots (Figure 3 and Table 4). By 10 dpi, fewer J2 developed into J3/J4 in the roots of Nongqing 28 and PI 437456 compared to the number of nematodes grown to J3 or J4 in Hefeng 50 roots (Figure 2, Table 4). By 21 dpi, the average number of white females in Hefeng 50 was 51, whereas the average female numbers in Nongqing 28 was 13; almost no SCN had advanced to females in PI 437456 (Table 4). Notably, the levels of resistance to SCN infection in PI 437456 was higher than in Nongqing 28. These results were consistent with the results from field tests, indicating that the development of SCN in Nongqing 28 was inhibited.
3.3. Transcription changes in Nongqing 28 in response to SCN infection
To investigate the resistance mechanism of Nongqing 28, we performed RNA-seq analysis with the SCN-infected and non-infected root tissues of Nongqing 28 at 10 dpi. A total of 2,808 upregulated genes and 1,523 downregulated genes were identified between the infected and uninfected roots in Nongqing 28 (Figure 4A-B and Supplementary Table S1). Gene ontology (GO) enrichment analysis demonstrated that the significantly upregulated genes were involved in GO terms mainly including defence response, plant responses to biotic stimuli and oxidative stress, plant hormone-related processes, reactive oxygen species (ROS) metabolic process, regulation of protein phosphatase activity and dephosphorylation, and secondary metabolic biosynthesis such as phenylpropanoid, monocarboxylic acid, oxylipin, and flavonoid (Figure 4C). The mostly affected molecular function (MF) terms were represented by heme binding, ADP binding, peroxidase activity, monooxygenase activity, quercetin 3-O-glucosyltransferase activity, hormone binding, and oxidoreductase activity, signaling receptor activity, and protein phosphatase activity (Supplementary Table S1). In the cell components (CC) category, plant cell wall (GO:0009505 and GO:0005618) and external encapsulating structure (GO:0030312) are the top enriched groups (Supplementary Table S1). The KEGG analysis revealed that the most enriched terms among the upregulated DEGs were associated with phenylpropanoid biosynthesis, ABC transporters, hormone biosynthesis (Brassinosteroid and Zeatin), and secondary metabolic biosynthesis pathways such as Isoflavonoid, flavone, tropane, piperidine, pyridine alkaloid, Linoleic acid, glucosinolate, cutin, suberine and wax (Figure 4D and Supplementary Table S1).
Furthermore, we identified key DEGs involved in soybean immune responses (Supplementary Table S2). For instance, a majority of DEGs encoding PRR proteins were upregulated in Nongqing 28 upon SCN infection, including receptor-like protein kinase (RLK), leucine-rich repeat (LRR)-RLK, lectin domain containing receptor kinase (LecRLK), serine/threonine-protein kinase (STK), Receptor-like protein (RLP), and cell wall-associated receptor kinase (WAK). Of the 44 NBS-LRR type resistance genes (NLR), which play critical roles in plant disease resistance, 41 NLR DEGs were found to be significantly induced by SCN (Figure 5A). Plant defence-related hormones ethylene (ET), jasmonic acid (JA), and salicylate acid (SA) play important roles in plant response to nematode stresses. Here, 87 DEGs involved in the biosynthesis and signaling of these three plant hormones were identified in Nongqing 28 during SCN infection. Significantly upregulated genes related to JA synthesis encoded lipoxygenases (12 DEGs), 12-oxophytodienoate reductase (1 DEG), and JA methyltransferase (4 DEGs). In the SA pathway, 7 2-oxoglutarate Fe(II)-dependent oxygenase (2OG oxygenase), 3 glucosyltransferase, 1 regulatory protein NPR1, and 2 MACPF domain-containing proteins exhibited upregulated expression in response to SCN infection. Among 44 DEGs encoding proteins related to ET biosynthesis and signaling in Nongqing 28, 36 DEGs were upregulated, and including 1-aminocyclopropane-1-carboxylate oxidases (ACO), 1-aminocyclopropane-1-carboxylate synthases (ACS), and ethylene-responsive transcription factors (ERF) (Figure 5B and Supplementary Table S1). In addition, other defence-related genes involved in ROS metobolic and signaling, calcium-mediating signaling, and MAPK cascades were also induced in SCN-infected roots of Nongqing 28. These proteins included respiratory burst oxidase homolog proteins (RBOH), glutathione S-transferase (GST), peroxidase, heat shock proteins (HSP), calcium-binding proteins (CaM), mitogen-activated protein kinase kinase kinases (MAPKKK), and pathogenesis-related proteins (PR), chitinase, and endo-1,3(4)-beta-glucanase (Figure 5C-D and Supplementary Table S1).
To validate the results from the RNA-seq data, six upregulated DEGs related to plant defense were selected for qRT-PCR analysis in Nongqing 28 and Hefeng 50 at 10 dpi (Figure 6). These genes included GmLecRK02 (Glyma.02G221900), GmCHI03 (Glyma.03G024600), GmPR6 (Glyma.20G205700), GmWRKY40 (Glyma.14G103100), GmNIMIN1 (Glyma.10G010100), and GmLRR-RLK (Glyma.10G069500). In agreement with the RNA-seq data, all six genes, except GmNIMIN1, displayed upregulated expression in Nongqing 28 roots after SCN infection (Figure 6). In contrast, the less significant induction in trancript levels of GmWRKY40, GmLecRK02, and GmNIMIN1 was observed in the susceptible soybean Hefeng 50 at 10 dpi. Notably, the expression of GmCHI03, GmLRR-RLK, and GmPR6 were downrelated by SCN in Hefeng 50 roots (Figure 6).
4. Discussion
The soybean planting area in China has steadily increased due to launching China’s soybean revitalization plan policy, particularly in Heilongjiang. In 2022, the total soybean planting area reached 4.93 million ha, accounting for 47% of the total soybean harvested area in China. The occurrence of SCN has been consistently frequent and severe in soybean-producing regions of Heilongjiang. Recent survey of SCN population density and virulence phenotypes in Heilongjiang indicated that over 50% of field samples tested positive for SCN. HG type 0 and 7 (race 3) are the most prevalent SCN population phenotypes in Heilongjiang [5,16]. Therefore, planting resistant soybean cultivars is a major method for controlling this pest. Currently, a serials of SCN-resistant, such as ‘KangXian’, ‘Nongqingdou’, and ‘Heinong 531’, have been developed and released in the market in Heilongjiang [15,24]. However, SCN virulence changes due to the overuse of soybean cultivars with the same resistant source has been reported in this area [17,25,26]. Thus, it is crucial to breed new soybean cultivars with different resistance sources in Heilongjiang. In this study, we provided a novel SCN-resistant cultivar, Nongqing 28, which exhibits stable resistance to SCN race 3 in the regional trials. Compared to the local cultivar Nengfeng 18, Nongqing 28 has high yields in both regional trials (2,593 kg/ha) and production trials (average yield of 2,660 kg/ha). The yield of Nongqing 28 is stably higher than the average yield of ‘KangXian’ soybean cultivars (2,082 kg/ha–2,513 kg/ha) in Heilongjiang. Additionally, Nongqing 28 features a yellow seed coat and brown hilum, which are more attractive in the market compared to other SCN-resistant cultivars with less desirable black hilum [24,27]. Furthermore, Nongqing 28 has an average fat content of 21.2% in the seed, meeting the National Standards for the second-level high-oil soybean variety (≥ 21% fat content in seed). Its high yield potential and desirable agronomic characteristics make it a promising candidate for use as a donor in developing SCN-resistant cultivars for breeding programs.
The resistance of Nongqing 28 to SCN was further characterized in a controlled greenhouse conditions. Results of acid fuchsin staining revealed that SCN development was significantly inhibited in Nongqing 28 compared to the susceptible cultivar at 10 dpi and 21 dpi. This suggests that a defense reaction occurred in the roots of Nongqing 28 in response to SCN infection, and further confirmed the reliable resistance of Nongqing 28 to SCN observed in field trials, effectively suppressing the SCN lifecycle. Similarly, the inhibition of SCN development has also been found in previous research on other resistant soybean cultivars including Heinong 531, Kangxian 12, and Pendou 158 [15], and Hatiwag [28]. However, SCN infection was not affected in Nongqing 28 at early nematode development stage (5dpi). In comparison, a rapid and strong resistant reaction was observed in Heinong 531 at 3dpi, which carried Peking -type SCN resistance. These findings suggest the SCN resistance of Nongqing 28 derived from PI 437654 is genetically distinct from SCN-resistant soybean cultivars with Peking background.
Our RNA-seq data provides important information on gene expression changes to further elucidate the resistance mechanism in Nongqing 28. The most enriched GO terms and pathways mainly included plant defence response, responses to biotic stimuli, plant-pathogen interaction, secondary metabolism (phenylpropanoid and favonoid biosynthesis), hormone metabolism, ROS metabolic process, and protein phosphorylation and dephosphorylation. We observed significant upregulation of genes involved in pathogen perception and defense activation in Nongqing 28 upon SCN infection, such as RLK, STKs, NBS-LRR, WAKs and RLPs. Additionally, DEGs related to plant defense signaling, including PRs, calcium-binding protein, MAPK, RBOH, GST, Peroxidase, and HSPs, were also activated in Nongqing 28. These findings suggest that an adequate recognition of the pathogen and activation of defense mechanisms may contribute to the observed resistance of Nongqing 28 to SCN. In accordance with these findings, a set of RNA-seq data showed the activation gene expression of soybean PTI and ETI components in resistant soybean cultivars in response to SCN infection [13,29,30,31,32,33]. Recent experimental evidences further supported the importance of MAPK genes (GmMAPK3 and GmMAPK6) and RLK genes (GmCDL1, GmLecRK02g, and GmLecRK08g) as the important PTI components contributed to SCN resistance through amplifying immune signaling [34,35]. The GmMAPK3/MAPK6-mediated phosphorylation of GmCDL1 activated basal immune responses and increase soybean resistance to SCN [35]. There has been some evidence for the involvement of SA, JA, and ET in soybean defense against SCN. For example, overexpression of specific genes in SA and signaling pathways, such as NPR1, TGA2, salicylic acid methyltransferase gene SAMT, confers soybean resistance to SCN [36,37,38]. Guo et al. [39] reported that inhibition of JA signaling decreased the Rhg1a-mediated resistance to SCN. ET-insensitive soybean mutant etr1-1 exhibited enhanced host resistance to SCN infection compared to the control soybean [40]. The present work showed that the expression levels of some specific DEGs in SA, JA, ET pathways were elevated in SCN-infected roots of Nongqing 28. Similarly, previous transcriptomic profiling revealed that many SA, JA, and ET related- genes were upregulated in other resistant soybean cultivars in the presence of SCN, suggesting the SA, JA and ET signaling might be the conserved defense components required for SCN resistance in soybean [29,30,31,32,33]. Therefore, SA, JA, and ET pathways may play critical role in Nongqing 28 against SCN, although this contribution did not offer a complete protection to this pest.
5. Conclusions
The present work reveals that Nongqing 28 is a new SCN-resistant soybean cultivar with high yield and favorable agronomic traits. This line can be used to enhance productivity and SCN resistance in the main soybean-producing areas of Heilongjiang. Furthermore, we used RNA-seq to demonstrate that the basal defense responses were activated in SCN-infected Nongqing 28 roots. The PTI and ETI signaling components, including several RLK subfamilies, cell wall-related receptors (WAKs), ROS genes (RBOHs), and MAPK family genes, CaM genes, NBS-LRRs (ETI-associated defenses), and JAs/ET and SA signaling, might contribute to the resistance of Nongqing 28 to SCN.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Table S1: List of all DEGs and Gene Ontology (GO) enrichment terms and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway analysis of upregulated differentially expressed genes in Nongqing 28 inoculated with SCN; Table S2: List of DEGs involved in the defence-related pathwayss in Nongqing 28 inoculated with SCN; Table S2: List of gene primers for RT-qPCR used in this study.
Author Contributions
Conceptualization, C.Z.; Y.H and J.W.; methodology, C.Z.; software, C.Z. and Y.H.; validation, Y.H and J.W; formal analysis, C.Z.; Y.H and J.W; investigation, C.Z.; Y.H.; G.C.; B.L.; J.Y.; Y.W.; J.L.; L.M.; writing—original draft preparation,Y.H; writing—review and editing,Y.H and J.W.; supervision, Y.H and J.W. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Scientific Institution Basal Research Fund of Heilongjiang province (CZKYF2023-1-B004), Leap Engineering Major Needs Science and Technology Innovation Key Program in Heilongjiang Province (CX23ZD04), Daqing Comprehensive Test Station of National Soybean Industrial Technology System (CARS-04-CES07), the Youth Innovation Promotion Association of CAS (Grant No. 2020236), and Jilin Science and Technology Department Project (20230405053RC).
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Peng, D.L.; Jiang, R.; Peng, H.; Liu, S.M. Soybean cyst nematodes: a destructive threat to soybean production in China. Phytopathol Res 2021, 3, 19. [Google Scholar] [CrossRef]
- Tylka, G.L.; Marett, C.C. Known Distribution of the Soybean Cyst Nematode, Heterodera glycines, in the United States and Canada in 2020. Plant Health Progress. 2021, 22, 72–74. [Google Scholar] [CrossRef]
- Bandara, A.Y.; Weerasooriya, D.K.; Bradley, C.A.; Allen, T.W.; Esker, P.D. Dissecting the economic impact of soybean diseases in the United States over two decades. PLoS One. 2020, 15, e0231141. [Google Scholar] [CrossRef] [PubMed]
- Lian, Y.; Koch, G.; Bo, D.; Wang, J.; Nguyen, H.T.; Li, C.; Lu, W.G. The Spatial Distribution and Genetic Diversity of the Soybean Cyst Nematode, Heterodera glycines, in China: It Is Time to Take Measures to Control Soybean Cyst Nematode. Front Plant Sci. 2022, 13, 927773. [Google Scholar] [CrossRef]
- You, J.; Wang, J.J.; Tian, R.Z.; Wang, S.R.; Yu, Y.; Xu, L.J.; Zhou, C.J.; Pan, F.J.; Chen, J.S.; Sui, Y.Y.; Hu, Y.F. Survey of Heterodera glycines population abundance and virulence phenotypes during 2021-2022 in Heilongjiang Province. Plant Dis 2024. [Google Scholar] [CrossRef] [PubMed]
- Arjoune, Y.; Sugunaraj, N.; Peri, S.; Nair, S.V.; Skurdal, A.; Ranganathan, P.; Johnson, B. Soybean cyst nematode detection and management: A review. Plant Methods. 2022, 18, 6–12. [Google Scholar] [CrossRef]
- Brucker, E.; Carlson, S.; Wright, E.; Niblack, T.; Diers, B. Rhg1 alleles from soybean PI 437654 and PI 88788 respond diferentially to isolates of Heterodera glycines in the greenhouse. Theor Appl Genet. 2005, 111, 44–49. [Google Scholar] [CrossRef]
- Kofsky, J.; Zhang, H.; Song, B.H. Novel resistance strategies to soybean cyst nematode (SCN) in wild soybean. Sci Rep. 2021, 11, 1–13. [Google Scholar]
- Cook, D.E.; Lee, T.G.; Guo, X.; Melito, S.; Wang, K.; Bayless, A.M.; Wang, J.; Hughes, T.J.; Willis, D.K.; Clemente, T.E.; Diers, B.W.; Jiang, J.; Hudson, M.E.; Bent, A.F. Copy number variation of multiple genes at Rhg1 mediates nematode resistance in soybean. Science. 2012, 338, 1206–1209. [Google Scholar] [CrossRef]
- Liu, S.; Kandoth, P.K.; Warren, S.D.; Yeckel, G.; Heinz, R.; Alden, J.; Yang, C.; Jamai, A.; El-Mellouki, T.; Juvale, P.S.; Hill, J.; Baum, T.J.; Cianzio, S.; Whitham, S.A.; Korkin, D.; Mitchum, M.G.; Meksem, K. A soybean cyst nematode resistance gene points to a new mechanism of plant resistance to pathogens. Nature. 2012, 492, 256–260. [Google Scholar] [CrossRef]
- Vuong, T.D.; Sleper, D.A.; Shannon, J.G.; Nguyen, H.T. Novel quantitative trait loci for broad-based resistance to soybean cyst nematode (Heterodera glycines Ichinohe) in soybean PI 567516C. Theor Appl Genet. 2021, 121, 1253–1266. [Google Scholar] [CrossRef] [PubMed]
- Guo, W.; Chen, J.S.; Zhang, F.; Li, Z.Y.; Chen, H.F.; Zhang, C.J.; Chen, L.M.; Yuan, S.L.; Li, R.; Cao, D.; Hao, Q.N.; Chen, S.L.; Shan, Z.H.; Yang, Z.L.; Zhang, X.J.; Qiu, Z.; You, Q.B.; Dai, W.J.; Zhou, X.A.; Shen, X.J.; Jiao, Y.Q. Characterization of Pingliang xiaoheidou (ZDD 11047), a soybean variety with resistance to soybean cyst nematode Heterodera glycines. Plant Mol Biol. 2020, 103, 253–267. [Google Scholar] [CrossRef] [PubMed]
- Wei, H.; Lian, Y.; Li, J.; Li, H.; Song, Q.; Wu, Y.; Lei, C.; Wang, S.; Zhang, H.; Wang, J.; Lu, W. Identification of Candidate Genes Controlling Soybean Cyst Nematode Resistance in “Handou 10” Based on Genome and Transcriptome Analyzes. Front Plant Sci 2022, 13, 860034. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.Z.; Li, J.Q.; Zhang, D.S. History and status of soybean cyst nematode in China. Int J Nematol. 1997, 7, 18–25. [Google Scholar]
- Wang, J.; Kong, L.; Zhang, L.; Shi, X.; Yu, B.; Li, J.; Zhang, B.; Gao, M.; Liu, X.; Li, X.; Gao, Y.; Peng, D.; Liu, S. Breeding a Soybean Cultivar Heinong 531 with Peking-Type Cyst Nematode Resistance, Enhanced Yield, and High Seed-Oil Contents. Phytopathology. 2022, 112, 1345–1349. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Zhou, Y.; Wang, Y.; Fan, H.; Liu, X.; Wang, D.; Zhao, D.; Duan, Y.; Zhu, X.; Chen, L. Characterization of Virulence Phenotypes of Heterodera glycines in Heilongjiang, Northeast China. Plant Dis. 2021, 105, 2056–2060. [Google Scholar] [CrossRef]
- Tian, Z.Y.; Gao, G.J.; Zhou, C.J.; Du, Z.Q.; Wu, Y.K.; Wang, M.Z.; Yang, L.; Li, J.Y. Study on the variation of soybean cyst nematode. Soybean Sci. 2007, 26, 290–292. [Google Scholar]
- Ma, Y.S.; Wang, W.H.; Wang, L.X.; Ma, F.M.; Wang, P.W.; Chang, R.Z.; Qiu, L.J. Genetic diversity of soybean and the establishment of a core collection focused on resistance to soybean cyst nematode. J Integr Plant Biol. 2006, 48, 722–731. [Google Scholar] [CrossRef]
- Meng, F.L.; Yu, J.Y.; Li, C.J.; Huang, M.H.; Zhao, L.; Wang, X.; Jiang, Y.; Qin, R.F.; Wang, C.L. Research progress on occurrence and management of soybean cyst nematode in Northeast China. Journal of Northeast Agricultural University 2022, 53, 87–94. (in Chinese). [Google Scholar]
- Wang, J.; Kong, L.; Zhang, L.; Shi, X.; Yu, B.; Li, J.; Zhang, B.; Gao, M.; Liu, X.; Li, X.; Gao, Y.; Peng, D.; Liu, S. Breeding a Soybean Cultivar Heinong 531 with Peking-Type Cyst Nematode Resistance, Enhanced Yield, and High Seed-Oil Contents. Phytopathology. 2022, 112, 1345–1349. [Google Scholar] [CrossRef]
- Riggs, R.D.; Schmitt, D.P. Complete Characterization of the Race Scheme for Heterodera glycines. J Nematol. 1988, 20, 392–395. [Google Scholar] [PubMed]
- Niblack, T.L.; Arelli, P.R.; Noel, G.R.; Opperman, C.H.; Orf, J.H. A revised classification scheme for genetically diverse populations of Heterodera glycines. J Nematol. 2002, 34, 279–288. [Google Scholar] [PubMed]
- Byrd, D.W.; Kirkpatrick, T.; Barker, K.R. An improved technique for clearing and staining plant tissues for detection of nematodes. J Nematol. 1983, 15, 142–143. [Google Scholar]
- Sun, M.M.; Wang, P.; Lv, S.X. New soybean varieties released in Heilongjiang Province in 2015. Soybean Sci 2015, 34, 918–920, (article in Chinese). [Google Scholar]
- Li, Y.H.; Li, X.B.; Tian, Z.Y.; Gao, J.G.; Cai, J.L.; Du, Z.Q. Study on the race about continuous plant varieties with resistance to SCN. Soybean Sci. 1998, 17, 370–372, (article in Chinese). [Google Scholar]
- Yu, B.S.; Wang, J.J.; Cui, L.W. Effects of Continuous Planting of Resistant SoybeanCultivars on Races of Soybean Cyst Nematode. Heilongjiang Agricultural Science 1999, 3, 5–7, (article in Chinese). [Google Scholar]
- Song, X.J. New soybean varieties released in Heilongjiang Province in 2012. Soybean Sci. 2012, 31, 504–510, (article in Chinese). [Google Scholar]
- Anand, S.C. Registration of ‘Hartwig’ soybean. Crop Sci. 1992, 32, 1069–1070. [Google Scholar] [CrossRef]
- Klink, V.P.; Overall, C.C.; Alkharouf, N.W.; Macdonald, M.H.; Matthews, B.F. Microarray detection call methodology as a means to identify and compare transcripts expressed within syncytial cells from soybean (Glycine max) roots undergoing resistant and susceptible reactions to the soybean cyst nematode (Heterodera glycines). J Biomed Biotechnol. 2010, 2010, 491217. [Google Scholar] [CrossRef]
- Klink, V.P.; Hosseini, P.; Matsye, P.D.; Alkharouf, N.W.; Matthews, B.F. Differences in gene expression amplitude overlie a conserved transcriptomic program occurring between the rapid and potent localized resistant reaction at the syncytium of the Glycine max genotype Peking (PI 548402) as compared to the prolonged and potent resistant reaction of PI 88788. Plant Mol Biol. 2011, 75, 141–165. [Google Scholar]
- Mazarei, M.; Liu, W.; Al-Ahmad, H.; Arelli, P.R.; Pantalone, V.R.; Stewart, Jr. C.N. Gene expression profiling of resistant and susceptible soybean lines infected with soybean cyst nematode. Theor Appl Genet. 2011, 123, 1193–1206. [Google Scholar] [CrossRef]
- Wan, J.; Vuong, T.; Jiao, Y.; Joshi, T.; Zhang, H.; Xu, D.; Nguyen, H.T. Whole-genome gene expression profiling revealed genes and pathways potentially involved in regulating interactions of soybean with cyst nematode (Heterodera glycines Ichinohe). BMC Genomics. 2015, 16, 148. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Kjemtrup-Lovelace, S.; Li, C.; Luo, Y.; Chen, L.P.; Song, B.H. Comparative RNA-Seq Analysis Uncovers a Complex Regulatory Network for Soybean Cyst Nematode Resistance in Wild Soybean (Glycine soja). Scientific reports. 2017, 7, 9699. [Google Scholar] [CrossRef] [PubMed]
- Khatri, R.; Pant, S.R.; Sharma, K.; Niraula, P.M.; Lawaju, B.R.; Lawrence, K.S.; Alkharouf, N.W.; Klink, V.P. Glycine max Homologs of DOESN’T MAKE INFECTIONS 1, 2, and 3 Function to Impair Heterodera glycines Parasitism While Also Regulating Mitogen Activated Protein Kinase Expression. Front Plant Sci. 2022, 13, 842597. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zhu, Q.; Tan, Y.; Deng, M.; Zhang, L.; Cao, Y.; Guo, X. Mitogen-activated protein kinases MPK3 and MPK6 phosphorylate receptor-like cytoplasmic kinase CDL1 to regulate soybean basal immunity. Plant Cell. 2024, 36, 963–986. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; Mazarei, M.; Zhao, N.; Zhu, J.J.; Zhuang, X.; Liu, W.; Pantalone, V.R.; Arelli, P.R.; Stewart, C.N. Jr.; Chen, F. Overexpression of a soybean salicylic acid methyltransferase gene confers resistance to soybean cyst nematode. Plant Biotechnol J. 2013, 11, 1135–1145. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; Mazarei, M.; Zhao, N.; Hatcher, C.N.; Wuddineh, W.A.; Rudis, M.; Tschaplinski, T.J.; Pantalone, V.R.; Arelli, P.R.; Hewezi, T.; Chen, F.; Stewart, C.N., Jr. Transgenic soybean overexpressing GmSAMT1 exhibits resistance to multiple-HG types of soybean cyst nematode Heterodera glycines. Plant Biotechnol J. 2016, 14, 2100–2109. [Google Scholar] [CrossRef]
- Matthews, B.F.; Beard, H.; Brewer, E.; Kabir, S. MacDonald MH, Youssef RM. Arabidopsis genes, AtNPR1, AtTGA2 and AtPR-5, confer partial resistance to soybean cyst nematode (Heterodera glycines) when overexpressed in transgenic soybean roots. BMC Plant Biol 2014, 14, 96. [Google Scholar] [CrossRef]
- Guo, W.; Zhang, F.; Bao, A.; You, Q.; Li, Z.; Chen, J.; Cheng, Y.; Zhao, W.; Shen, X.; Zhou, X.; Jiao, Y. The soybean Rhg1 amino acid transporter gene alters glutamate homeostasis and jasmonic acid-induced resistance to soybean cyst nematode. Mol Plant Pathol. 2019, 20, 270–286. [Google Scholar] [CrossRef]
- Tucker, M.L.; Xue, P.; Yang, R. 1-Aminocyclopropane-1-carboxylic acid (ACC) concentration and ACC synthase expression in soybean roots, root tips, and soybean cyst nematode (Heterodera glycines)-infected roots. J Exp Bot. 2010, 61, 463–472. [Google Scholar] [CrossRef]
Figure 1.
Pedigree chart of soybean cultivar Nongqing 28. An 02-318 derived from Hartwig is resistant to soybean cyst nematode (SCN), but Hei 99-980 is susceptible to SCN.
Figure 1.
Pedigree chart of soybean cultivar Nongqing 28. An 02-318 derived from Hartwig is resistant to soybean cyst nematode (SCN), but Hei 99-980 is susceptible to SCN.
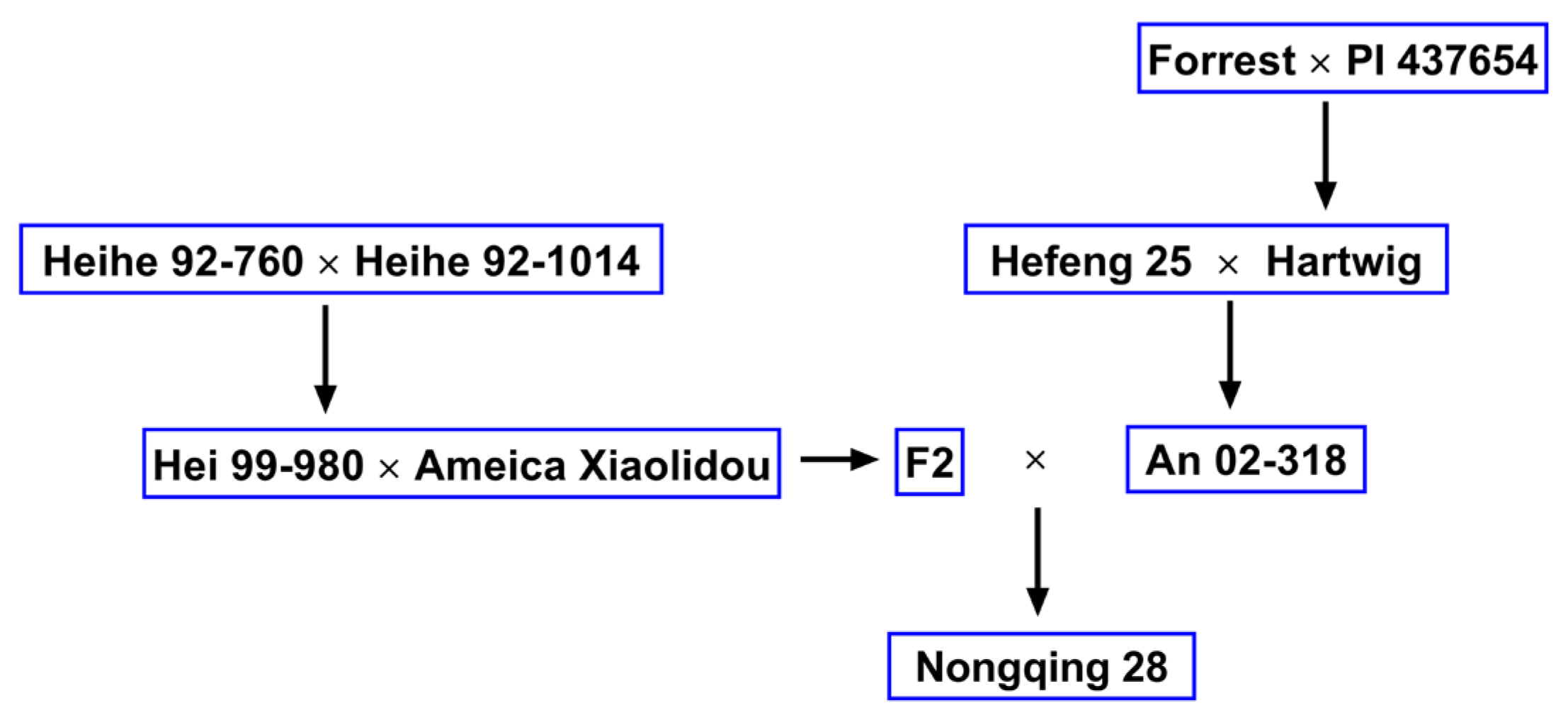
Figure 2.
The phenotypes and seeds of soybean cultivar Nongqing 28 in maturity stage.

Figure 3.
The comparable development of soybean cyst nematode on Nongqing28, PI 437456, and Heifeng 50 roots at 5 and 10 days post inoculation (dpi).
Figure 3.
The comparable development of soybean cyst nematode on Nongqing28, PI 437456, and Heifeng 50 roots at 5 and 10 days post inoculation (dpi).
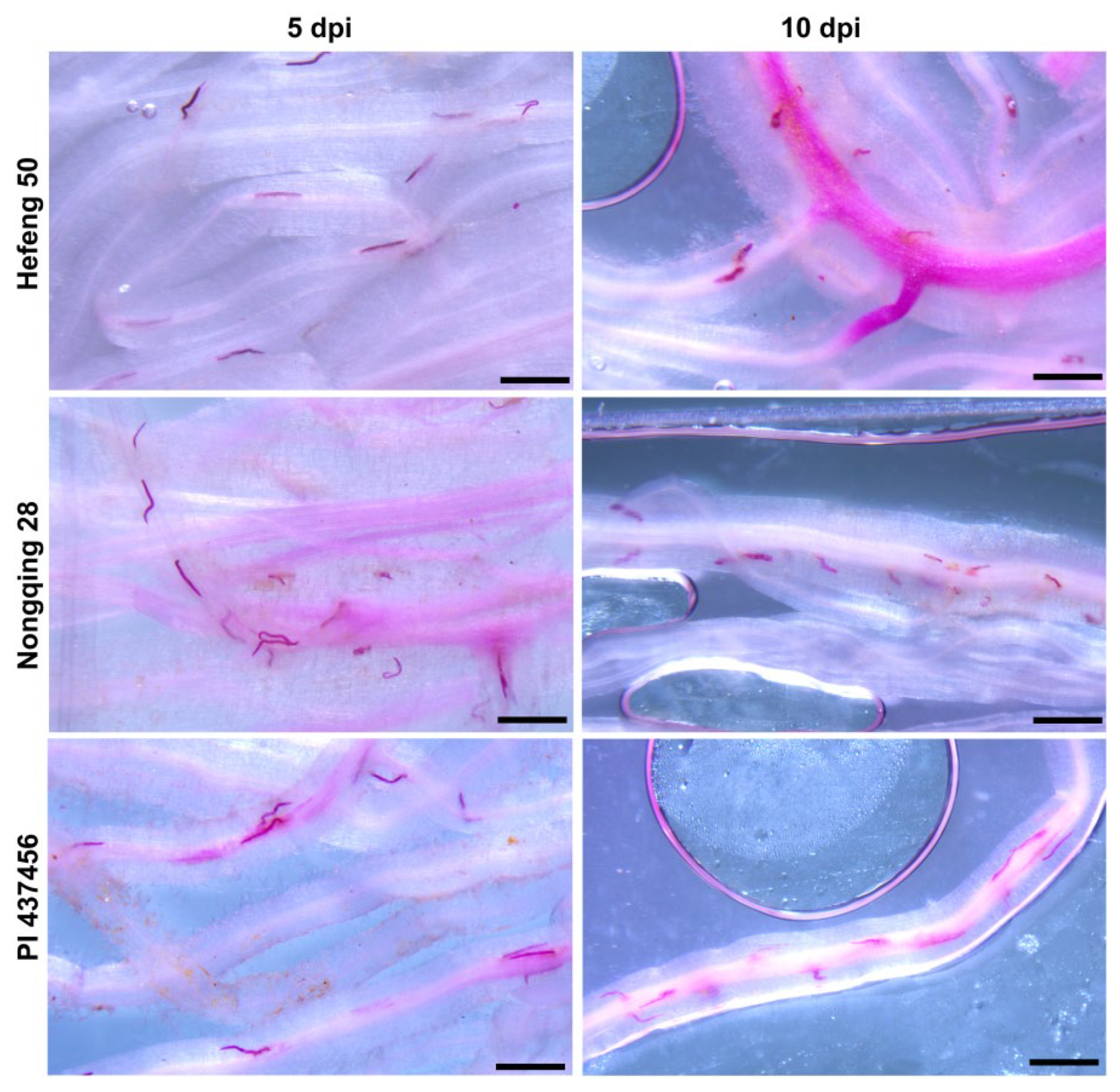
Figure 4.
Transcriptome response of Nongqing28 to soybean cyst nematode at 10 days post inoculation (dpi). (A) The number of differentially expressed genes (DEGs). (B) Volcano plots indicating all soybean genes of Nongqing 28 at 10 dpi. The red and green dots represent up- and downregualted genes. (C) Top 30 enriched Gene Ontology (GO) biological processes (BP) of the upregulated DEGs. (D) Top 20 most enriched KEGG terms of upregulated DEGs.
Figure 4.
Transcriptome response of Nongqing28 to soybean cyst nematode at 10 days post inoculation (dpi). (A) The number of differentially expressed genes (DEGs). (B) Volcano plots indicating all soybean genes of Nongqing 28 at 10 dpi. The red and green dots represent up- and downregualted genes. (C) Top 30 enriched Gene Ontology (GO) biological processes (BP) of the upregulated DEGs. (D) Top 20 most enriched KEGG terms of upregulated DEGs.

Figure 5.
Heatmap of the representative DEGs involved in the defence-related pathways. (A) DEGs encoding for proteins with role in perception and defense signaling including receptor-like protein kinase (RLK), serine/threonine-protein kinase (STK), Receptor-like protein (RLP), TIR-NBS-LRR type disease resistance protein. (B) DEGs encoding for proteins responsible for ROS metabolic and signaling including respiratory burst oxidase homolog protein (RBOH), glutathione S-transferase (GST), Peroxidase, heat shock protein (HSP). (C) DEGs involved in defence-related plant hormone signaling including salicylic acid (SA), jasmonic acid (JA), and ET. (D) DEGs involved in other defense proteins including cell wall-associated receptor kinase (WAK), pathogenesis-related proteins (PRs), calcium-binding protein (CaM), and mitogen-activated protein kinase (MAPK). Individual genes are listed and colors represent the log2 fold change value based on the comparison of the transcript levels between SCN-inoculated and non-inoculated roots in Nongqing 28. Red represents upregulated DEGs and green downregulated DEGs.
Figure 5.
Heatmap of the representative DEGs involved in the defence-related pathways. (A) DEGs encoding for proteins with role in perception and defense signaling including receptor-like protein kinase (RLK), serine/threonine-protein kinase (STK), Receptor-like protein (RLP), TIR-NBS-LRR type disease resistance protein. (B) DEGs encoding for proteins responsible for ROS metabolic and signaling including respiratory burst oxidase homolog protein (RBOH), glutathione S-transferase (GST), Peroxidase, heat shock protein (HSP). (C) DEGs involved in defence-related plant hormone signaling including salicylic acid (SA), jasmonic acid (JA), and ET. (D) DEGs involved in other defense proteins including cell wall-associated receptor kinase (WAK), pathogenesis-related proteins (PRs), calcium-binding protein (CaM), and mitogen-activated protein kinase (MAPK). Individual genes are listed and colors represent the log2 fold change value based on the comparison of the transcript levels between SCN-inoculated and non-inoculated roots in Nongqing 28. Red represents upregulated DEGs and green downregulated DEGs.
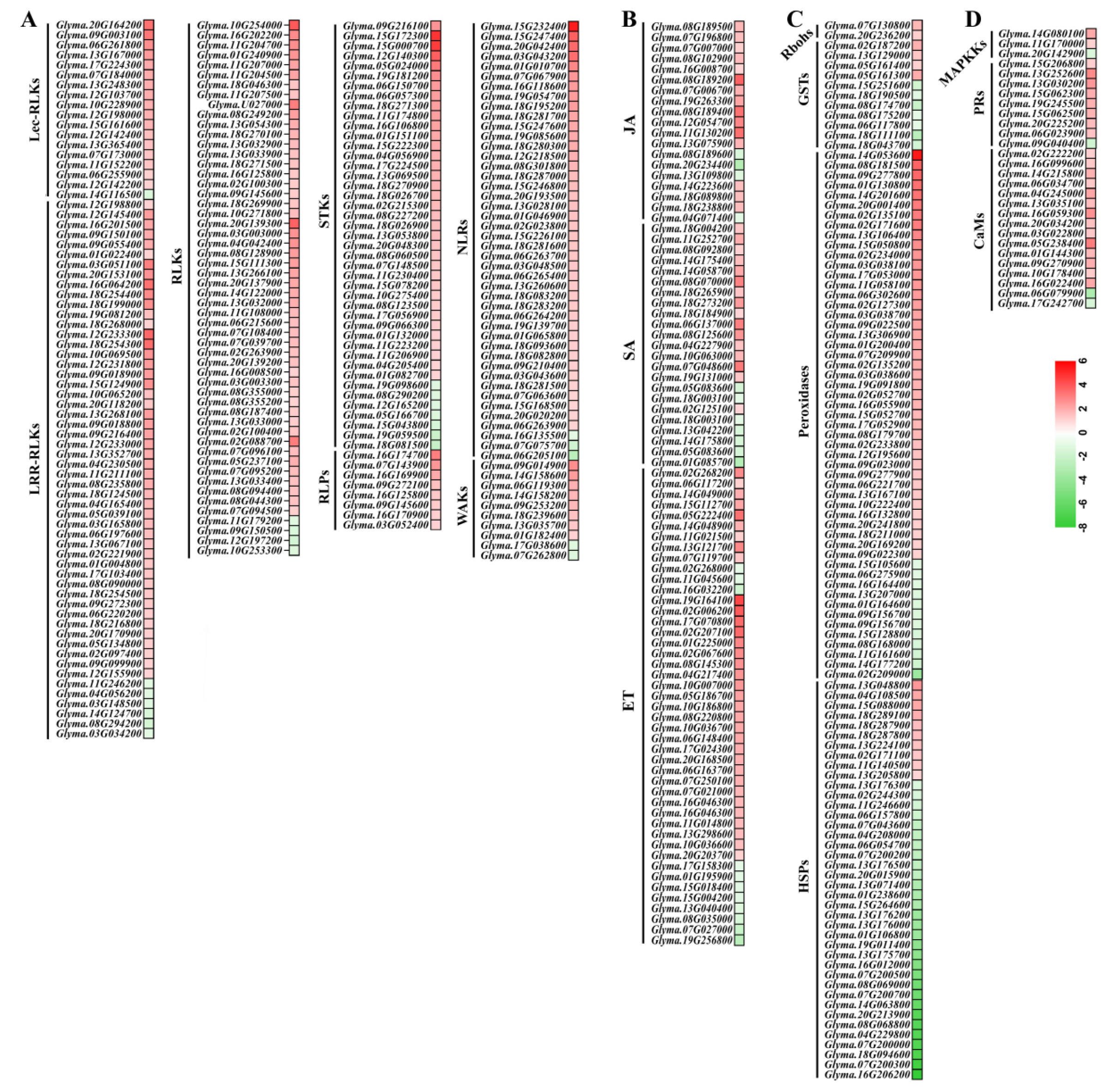
Figure 6.
Validation of DEGs in Nongqing 28 and Hefeng 50 at 10 dpi by quantity reverse transcription RT-qPCR. The expression levels of target genes are relative to the level of expression of soybean genes in the non-inoculated control plants using GmUBQ3 as reference gene. The data are presented as the means of three replicates ± SE. Asterisks indicate a statistically significant difference between soybeans genes in SCN-infected plants and the non-infected control plants (Student’s t-test, * p < 0.5).
Figure 6.
Validation of DEGs in Nongqing 28 and Hefeng 50 at 10 dpi by quantity reverse transcription RT-qPCR. The expression levels of target genes are relative to the level of expression of soybean genes in the non-inoculated control plants using GmUBQ3 as reference gene. The data are presented as the means of three replicates ± SE. Asterisks indicate a statistically significant difference between soybeans genes in SCN-infected plants and the non-infected control plants (Student’s t-test, * p < 0.5).
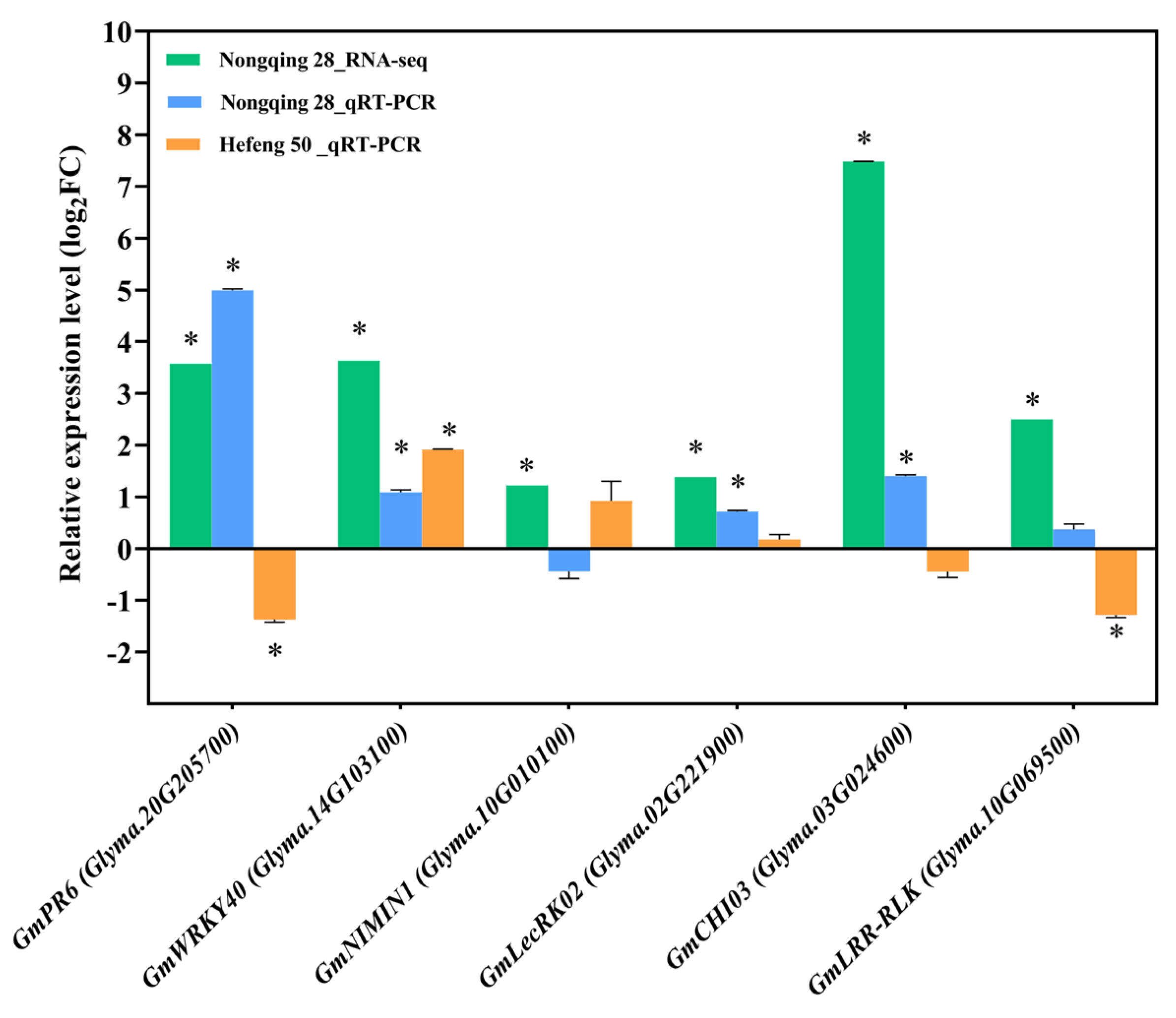

Table 1.
Seed yield and agronomic trials of Nongqing 28 in regional tests in 2018 and 2019.
| Year | Trial locations | Height (cm) |
Seed yield (Kg/ha) |
Increase (%) | Oil contents | Protein contents |
|---|---|---|---|---|---|---|
| 2018 | Anda Dapeng Seeds Co.Ltd. | 92 | 2471.2 | 1.3 | 18.93 | 41.26 |
| Longjiang Branch of Qishan Seeds Co.Ltd. | 105 | 2725.0 | 6.2 | 22.03 | 38.41 | |
| Daqing Seeds Management Bureau | 91 | 2250.0 | 0.7 | 22.97 | 37.53 | |
| Dumeng Seeds Management Bureau | 90 | 2956.7 | 7.9 | 21.24 | 40.13 | |
| Tailai Seeds Station | 92 | 2426.0 | 9.1 | 20.78 | 39.21 | |
| Qiqihar Branch of Heilongjiang Academy of Agricultural Sciences | 91 | 2421.2 | -0.5 | 19.67 | 40.17 | |
| mean | 93.5 ± 2.3 | 2542 ± 103.9 | 4.1 ± 1.7 | 20.9 ± 0.6 | 39.5 ± 0.6 | |
| 2019 | Anda Dapeng Seeds Co.Ltd. | 107 | 2677.9 | 11.8 | 22.41 | 36.69 |
| Qiqihar Branch of Heilongjiang Academy of Agricultural Sciences | 109 | 2623.3 | 1.3 | 20.17 | 37.93 | |
| Longjiang Branch of Qishan Seeds Co.Ltd. | 82 | 2732.4 | 14.4 | 21.72 | 36.43 | |
| Qinggang Experimental Station of Beifeng Seeds Co.Ltd. | 109 | 2550.0 | 1.9 | 22.01 | 35.36 | |
| Tailai Seeds Station | 101 | 2521.3 | 7.0 | 21.61 | 36.73 | |
| Mean | 101.6 ± 5.1 | 2621 ± 39.1 | 7.3 ± 2.6 | 21.6 ± 0.4 | 36.6 ± 0.4 | |
| Total | 97.2 ± 2.8 | 2593 ± 61.9 | 6.2 ± 1.5 | 21.2 ± 0.4 | 38.2 ± 0.6 |
Nengfeng 18 was used as the control. Each trial location has a randomized complete design with three plots (three replicates) in each year. The seed yield was converted based on the production of each plot (20 m2). The results show the yield increase of Nongqing 28, compared with the local soybean cultivar Nengfeng 18.
Table 2.
Seed yield of Nongqing 28 in production trials in 2019.
| Trial locations | Seed yield (kg/ha) | Increase (%) |
|---|---|---|
| Anda Dapeng Seeds Co.Ltd. | 2523.3 | 12.7 |
| Qiqihar Branch of Heilongjiang Academy of Agricultural Sciences | 2663.8 | 1.5 |
| Longjiang Branch of Qishan Seeds Co.Ltd. | 2467.5 | 11.4 |
| Qinggang Experimental Station of Beifeng Seeds Co.Ltd. | 2764.4 | 5.7 |
| Tailai Seeds Station | 2880 | 9.3 |
| Total | 2660 ± 75.9 | 8.1 ± 2.0 |
Nengfeng 18 was set as the control. The area of each location in production trials was about 200 or 300 m2. The results show the yield increase of Nongqing 28, compared with the local soybean cultivar Nengfeng 18.
Table 3.
Reaction of Nongqing 28 to soybean cyst nematode in fields during 2018-2021.
| soybean cultivars |
2018 | 2019 | 2020 | 2021 | ||||
|---|---|---|---|---|---|---|---|---|
| Cystsa | FI (%)b | Cystsa | FI (%)b | Cystsa | FI (%)b | Cystsa | FI (%)b | |
| Lee | 63 ± 4.15A | 100 | 65 ± 5.57A | 100 | 69 ± 4.74A | 100 | 68 ± 6.53A | 100 |
| Hefeng 50 | 47 ± 2.55A | 74.60 | 52 ± 4.16A | 80.00 | 67 ± 4.06A | 97.1 | 58 ± 3.90A | 85.29 |
| Nongqing 28 | 3 ± 0.42B | 4.76 | 2 ± 0.43B | 3.08 | 3 ± 0.50B | 4.35 | 3 ± 0.46B | 4.41 |
| PI 437654 | 1 ± 0.33B | 1.59 | 2 ± 0.60B | 3.08 | 1 ± 0.42B | 1.45 | 2 ± 0.72B | 2.94 |
Hefeng 50 and Lee were used as the susceptible controls, PI 437456 was set as the resistant control. aData are mean ± SE of 20 replicates. bFI= (average number of females on the soybean cultivar/average number of females on the susceptible check of Lee) × 100.
Table 4.
Comparison of soybean cyst nematode development in Nongqing 28 and Heifeng 50.
| Soybean cultivars | 5 dpi | 10 dpi | 21 dpi | |||
|---|---|---|---|---|---|---|
| J2 | J3 | J2 | J3 | J4 | Female | |
| Hefeng 50 | 205 ± 8.14 a | 18 ± 3.21 a | 107 ± 8.77 a | 73 ± 3.31 a | 5 ± 0.53 a | 51 ± 5.93 a |
| Nongqing 28 | 193 ± 6.51 a | 12 ± 1.53 b | 95 ± 9.69 a | 41 ± 3.01 b | 2 ± 0.43 a | 13 ± 1.73 b |
| PI 437654 | 165 ± 4.84 b | 3 ± 0.88 c | 83 ± 4.67 a | 23 ± 2.92 c | 0 ± 0 b | 0 ± 0.10 c |
The experiment was repeated three times, and ten replicates were included at each time. Hefeng 50 were used as the susceptible control. Data are mean ± SE, *denote significant difference (P < 0.05) compared to the susceptible control. dpi, days post-inoculation; J2, second stage juveniles; J3, third stage juveniles; J4, fourth stage juveniles.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



