Submitted:
23 July 2024
Posted:
24 July 2024
You are already at the latest version
Abstract
Jerusalem artichoke juice is valued for its nutritional content and health benefits. Spontaneous fermentation enhances its flavor, quality, and functional components through microbial metabolic activities. This study used high-throughput sequencing to analyze microbial community changes and LC-MS and GC-MS to detect secondary metabolites and flavor compounds during fermentation. During the natural fermentation, beneficial bacteria like Lactobacillus and Pediococcus increased, promoting lactic acid production and inhibiting harmful bacteria, while environmental bacteria decreased. Similarly, fungi shifted from environmental types like Geosmithia and Alternaria to fermentation-associated Pichia and Penicillium. A total of 1, 666 secondary metabolites were identified, with 595 upregulated and 497 downregulated. Key metabolic pathways included phenylpropanoid biosynthesis, with significant increases in phenylalanine, tryptophan, and related metabolites. Lipid and nucleotide metabolism also showed significant changes. Flavor compounds, including 134 identified alcohols, esters, acids, and ketones, mostly increased in content after fermentation. Notable increases were seen in Phenylethyl Alcohol, Ethyl Benzenepropanoate, 3-Methylbutyl Butanoate, Ethyl 4-Methylpentanoate, 5-Ethyl-3-Hydroxy-4-Methyl-2(5H)-Furanone, Ethyl Decanoate, Hexanoic Acid, 1-Octanol. γ-aminobutyric acid (GABA) and other functional components enhanced the health value of the juice. This study provides insights into microbial and metabolic changes during fermentation, aiding in optimizing processes and improving the quality of fermented Jerusalem artichoke juice for functional food development.
Keywords:
Jerusalem artichoke
; Spontaneous fermentation
; Microbial diversity
; Secondary metabolites
; Flavor compounds
1. Introduction
Jerusalem artichoke (Helianthus tuberosus L.) tubers are widely recognized for their high inulin content [1,2]. Inulin, which constitutes 10-25% of the fresh tuber weight, is a functional dietary fiber that acts as a prebiotic, promoting the growth of beneficial gut bacteria such as bifidobacteria and lactobacilli, thereby improving gut health [3,4]. Inulin significantly reduces blood glucose levels in diabetic patients and enhances insulin sensitivity [2,5]. Its hydrolysis product, fructose, does not cause a rapid increase in blood sugar levels, making it suitable for diabetic patients and those requiring glycemic control [6,7,8]. Additionally, inulin aids in the absorption of minerals like calcium and magnesium, promoting bone health and playing a crucial role in preventing and treating osteoporosis [9,10,11,12,13].
Jerusalem artichoke tubers are also rich in antioxidants and anti-inflammatory compounds, which protect body cells by reducing oxidative stress and inhibiting inflammatory responses [14,15,16,17]. The high content of vitamins C, E, and polyphenolic compounds in the tubers helps prevent cardiovascular diseases and cancer [15,18]. Furthermore, Jerusalem artichoke extracts can be used in fermented fruit and vegetable juices to boost probiotic content and functionality [15,18,19]. Jerusalem artichoke tubers improve the moisture retention and texture of fermented bread, increasing dietary fiber content and catering to health-conscious consumers [20,21,22,23]. The components of Jerusalem artichoke juice, rich in inulin and other fermentable sugars, serve as substrates for microbial metabolism, leading to the production of various primary and secondary metabolites [24].
Spontaneous fermentation is an effective food processing method that utilizes natural microbial communities from raw materials, equipment, and the environment [25,26]. This method preserves the original flavor of the ingredients while producing beneficial metabolites through microbial activity, enhancing the nutritional and health value of the food [27,28]. Secondary metabolites play crucial roles in fermentation. Phenolic acids, flavonoids, and alkaloids exhibit antioxidant, antimicrobial, and anticancer properties. For example, flavonoids have strong antioxidant activity, protecting cells from oxidative stress [29,30]. Phenolic acids like chlorogenic acid and caffeic acid have anti-inflammatory and antimicrobial effects, enhancing food preservation and safety [31,32,33]. Additionally, alkaloids produced during fermentation, such as biotin and pyridoxine, improve the nutritional value of the food and enhance its health benefits [34,35]. Studies have shown that adding inulin to spontaneously fermented products like yogurt, kimchi, and fermented fruit and vegetable juices increases the probiotic content and enhances health benefits. Inulin addition significantly improves the survival rate of probiotics and sensory properties in yogurt. During fermentation, inulin is metabolized by lactic acid bacteria, producing organic acids (e.g., lactic and acetic acids) and other metabolites that enhance the taste and nutritional value of yogurt [36,37,38,39,40].
The flavor of spontaneously fermented foods primarily arises from various metabolites produced by microbes during fermentation. Different microorganisms generate a variety of volatile compounds, such as alcohols, esters, aldehydes, ketones, and organic acids, through metabolic pathways, creating complex and unique flavors and aromas. In the early stages of fermenting beer, sauerkraut, cocoa beans, carrot juice, and kimchi, common enterobacteria metabolize carbohydrates to produce various organic acids and alcohols, forming distinctive flavors [25,41,42]. Spontaneously fermented foods, free from artificial additives or industrial starter cultures, are more natural and pure, thus increasingly favored by consumers. Jerusalem artichoke tubers show broad application potential in spontaneously fermented foods, preserving the original flavor of the ingredients while generating various beneficial metabolites through microbial activity. With the growing demand for healthy and natural foods, the application prospects of Jerusalem artichoke tubers in spontaneously fermented foods are promising [43]. However, research on the microbial diversity and metabolic characteristics of spontaneously fermented Jerusalem artichoke juice remains limited.
In-depth studies on these characteristics are essential for understanding the dynamic changes in microbial communities and the mechanisms of metabolite production during spontaneous fermentation of Jerusalem artichoke juice. This study aims to systematically analyze the structural and functional changes in microbial communities during the spontaneous fermentation of Jerusalem artichoke juice, elucidating the metabolic pathways of key microbes and their impact on product flavor and nutritional value. This will help optimize fermentation processes, improving the quality and functionality of fermented Jerusalem artichoke products. Additionally, by investigating the secondary metabolites produced during spontaneous fermentation, such as phenolic compounds, flavonoids, and alkaloids, we can better understand their health benefits and application potential in foods. Given the rich nutritional value and health benefits of Jerusalem artichoke tubers, this study will provide a scientific basis for developing high-value-added fermented products and promote the application and development of Jerusalem artichoke in functional foods.
2. Materials and Methods
2.1. Sample Preparation
Fresh Jerusalem artichoke tubers were sourced from Yuzhong experimental station of Lanzhou University, Lanzhou, China (35◦56′N, 104◦09′E, 1750 m asl). The tubers were thoroughly washed with tap water, juiced with distilled water at a 1:2 weight ratio, and filtered through an 80-mesh sieve to remove solid impurities. Each liter of juice was transferred to a 2 L sterilized Erlenmeyer flask, and 50 g of white sugar were added. The flasks were placed in a 25°C incubator for fermentation, which lasted for 30 days to ensure sufficient fermentation time (JASF). Unfermented juice samples served as control groups for comparative analysis (CK).
2.2. Determination of Microbial Diversity
To assess microbial diversity in the fermented juice, samples were pretreated and total microbial DNA was extracted. Samples (0.2-0.5 g) were quickly thawed, placed in 2 mL centrifuge tubes with extraction lysis solution, and homogenized using a Tissuelyser-48 at 60 Hz. Total DNA was extracted using the OMEGA Soil DNA Kit (M5635-02) according to the manufacturer’s protocol. DNA quality was assessed using a NanoDrop NC2000 spectrophotometer and 0.8% agarose gel electrophoresis.
The bacterial 16S rRNA gene V3-V4 regions were amplified by PCR using primers 338F (5’-ACTCCTACGGGAGGCAGCA-3’) and 806R (5’-GGACTACHVGGGTWTCTAAT-3’). The PCR mixture included 5 μL of 5× buffer, 0.25 μL of Fast pfu DNA polymerase (5 U/μL), 2 μL of dNTPs (2.5 mM), 1 μL of each primer (10 μM), and 1 μL of DNA template, with a final volume of 25 μL. The PCR conditions were 98°C for 5 min, followed by 25 cycles of 98°C for 30 s, 53°C for 30 s, and 72°C for 45 s, with a final extension at 72°C for 5 min. PCR products were purified using Vazyme VAHTS DNA Clean Beads and quantified with the Quant-iT PicoGreen dsDNA Assay Kit on a BioTek FLx800 fluorometer. Libraries were prepared using the Illumina TruSeq Nano DNA LT Library Prep Kit and sequenced on the Illumina NovaSeq platform (2×250 bp) [44]. Data analysis was performed using QIIME2 2019.4, which included sequence demultiplexing, primer trimming, quality filtering, denoising, merging, and chimera removal using DADA2. Amplicon sequence variants (ASVs) were classified using the Greengenes database. Alpha diversity metrics were calculated, and data visualization and statistical analysis were performed using R software, including PCoA, NMDS, and PCA. Taxonomic composition, differential analysis, and correlation network construction were conducted using SparCC, and functional predictions were made with PICRUSt2.
2.3. Analysis of Secondary Metabolites
Samples were thawed, vortexed, and 100 μL of each sample was mixed with 100 μL of 70% methanol containing internal standards. The mixture was vortexed for 15 minutes at 4°C and then centrifuged. The supernatant was filtered through a 0.22 μm membrane and stored for subsequent LC-MS/MS analysis. Samples were analyzed in both positive and negative ion modes using an ACQUITY Premier HSS T3 column with a gradient of solvent A (0.1% formic acid in water) and solvent B (0.1% formic acid in acetonitrile). The gradient conditions and parameters were optimized to ensure comprehensive metabolite detection. Mass spectrometry was conducted on an Agilent 6545A QTOF, with specific ion source parameters set for each mode.
Mass spectrometry data were converted to mzXML format using ProteoWizard. XCMS software was employed for peak extraction, alignment, and retention time correction. Peaks with more than 50% missing values were filtered out. The remaining peaks were normalized and identified through custom and public databases. Metabolomics data were analyzed using PLS-DA with SIMCA-P 14.0 software, and differential metabolites were identified based on VIP scores and manual inspection.
2.4. Analysis of the Volatile Flavor Compounds Using HS-SPME-GC-MS
To explore metabolite differences in fermented Jerusalem artichoke juice, control (CK) and fermented (JASF) samples were collected, frozen in liquid nitrogen, and stored at -80°C. Each sample (500 mg or 1 mL) was mixed with a saturated NaCl solution and 20 μL of internal standard (10 μg/mL hexane) and extracted using HS-SPME for GC-MS analysis.
GC-MS analysis was performed on an Agilent 8890-7000D system equipped with a DB-5MS column. Headspace extraction was conducted at 60°C for 15 minutes using a 120 µm DVB/CWR/PDMS fiber, followed by desorption at 250°C for 5 minutes. High-purity helium was used as the carrier gas at a flow rate of 1.2 mL/min. The temperature program started at 40°C (held for 3.5 minutes), ramped to 100°C at 10°C/min, then to 180°C at 7°C/min, and finally to 280°C at 25°C/min, where it was held for 5 minutes. Mass spectrometry was conducted in electron impact ionization (EI) mode. Metabolites were identified and quantified using a custom database, with accuracy ensured through retention time and selective ion monitoring (SIM).
3. Results and Discussion
3.1. Changes of Microbial Diversity during Fermentation
3.1.1. Bacterial Diversity
A detailed analysis of the microbial communities before (CK group) and after fermentation (JASF group) of Jerusalem artichoke juice revealed significant changes in microbial diversity (Figure 1A). The Chao1 index was approximately 483 in the CK group and decreased to about 314 in the JASF group. Similarly, the Observed_species index dropped from about 443 in the CK group to around 307 in the JASF group. The Faith_pd index decreased from about 35 in the CK group to 19 in the JASF group, suggesting that certain microbial communities became dominant during fermentation, leading to a reduction in overall diversity. The Simpson and Shannon indices showed that the diversity and evenness in the CK group (0.26 and 1.5, respectively) were significantly lower than those in the JASF group (0.57 and 2.3, respectively). Additionally, the Pielou_e index indicated better species evenness in the CK group (0.17) compared to the JASF group (0.28). The Goods-coverage index showed better sequencing depth in the CK and JASF group, nearing 1.000. These data suggest that microbial diversity and evenness significantly decreased during fermentation, likely due to changes in substrate utilization.
Heatmap analysis (Figure 1B) further highlighted significant differences in microbial community structures between the CK and JASF groups. During the natural fermentation of Jerusalem artichoke tuber juice, the microbial community structure underwent substantial changes. Prior to fermentation, environmental bacteria such as Flavobacterium, Sphingomonas, and Luteimonas predominated, participating in the initial degradation of organic matter in the tuber juice. Post-fermentation, beneficial bacteria like Lactobacillus and Pediococcus increased significantly, promoting lactic acid production, lowering pH, and thereby inhibiting the growth of harmful bacteria and stabilizing the fermentation environment [45,46]. Enterobacter and Bacillus also proliferated during fermentation, while the former includes some potential pathogens, the latter are mostly beneficial, although certain species like Bacillus cereus may cause food poisoning. Therefore, understanding the dynamic changes of beneficial and pathogenic bacteria during fermentation is crucial for ensuring the safety of the fermented product and optimizing the process.
The species diversity and abundance of bacteria at the phylum and genus levels during the spontaneous fermentation of Jerusalem artichoke tubers are displayed in Figure 1C. The microbial community is highly diverse, involving multiple phyla and genera, which is essential for a balanced fermentation process, contributing to the development of flavors, textures, and possibly health benefits of the fermented product. Proteobacteria is the most dominant phylum across all conditions, particularly high in CK and CK-JASF, indicating its significant role in fermentation. However, Proteobacteria in the CK group might include potential pathogens that start to decrease as fermentation progresses, suggesting the need to control such pathogens during natural fermentation. Firmicutes are notable in the JASF condition, suggesting their involvement in breaking down complex carbohydrates and producing fermentation end-products like lactic acid. Genera such Gluconobacter, Lysinibacillus and Pediococcus in JASF are important for lactic acid production, crucial for lowering pH and inhibiting spoilage organisms [47,48,49]. Differences in microbial composition across CK, JASF, and CK-JASF highlight the impact of environmental factors and fermentation stages on microbial dynamics.
3.1.2. Fungal Diversity
The α-diversity index analysis of fungi (Figure 2A) shows significant changes in fungal community structure during fermentation. The Chao1 index indicates a significant decrease in fungal richness, from approximately 150 in the CK group to about 32 in the JASF group (p = 0.0039). The Shannon index reveals a decrease in diversity and evenness, dropping from about 4.0 in the CK group to 3.3 in the JASF group (p = 0.0039), while the Pielou’s evenness index increases from 0.56 to 0.68 (p = 0.0039). The Simpson index remains similar between groups at around 0.86 (p = 0.87), suggesting little change in the dominance of fungal communities. The number of observed species decreases significantly from about 148 to 30 (p = 0.0039), consistent with the Chao1 results. The Goods-coverage index, nearly 1.000 for both groups (p = 0.52), indicates sufficient sequencing depth. Overall, while fungal richness decreases after fermentation, evenness increases, leading to a more uniform community structure.
Heatmap analysis reveals distinct fungal community structures before (CK group) and after (JASF group) fermentation (Figure 2B). During the natural fermentation of Jerusalem artichoke tuber juice, significant changes in the fungal community structure were observed. Before fermentation (CK group), environmental fungi such as Geosmithia, Paecilomyces, Brunneochlamydosporium, Alternaria, and Cladosporium dominated, participating in the initial degradation of organic matter. After fermentation (JASF group), there was a notable increase in Pichia and Penicillium. Pichia significantly proliferated during fermentation, contributing to the stability and flavor development of the fermented product [50,51]. Although Penicillium was not abundant before fermentation, it increased post-fermentation and could impact the characteristics of the final product. The fermentation process, characterized by lower pH, organic acid production, and reduced oxygen levels, favored the growth of specific yeasts and molds while inhibiting plant pathogens like Fusarium and Alternaria. This dynamic shift not only ensures the safety of the fermented product but also enhances its flavor and quality.
Figure 2C displays the species diversity and abundance of fungi at the phylum and genus levels across three conditions (CK, JASF, and CK-JASF). At the phylum level, Ascomycota is dominant in all conditions, especially in CK and CK-JASF, indicating its significant role during fermentation. This phylum is known for its capability to produce various enzymes and secondary metabolites that contribute to the breakdown of complex carbohydrates and the development of flavors [52]. Basidiomycota is present in lower amounts, with the highest abundance in the CK group and a significant decrease in the JASF group. This reduction could be due to the competitive exclusion by Ascomycota members that thrive in the fermentation environment. Mortierellomycota is only present in trace amounts, indicating a minimal role in the fermentation process. At the genus level, Saccharomyces is highly abundant in CK and CK-JASF, playing a crucial role in fermentation due to its well-known fermentation capabilities, including ethanol production and flavor enhancement. Tetracladium and Pseudogymnoascus are present in the CK condition but less abundant in JASF. This suggests that these genera might be involved in the initial stages of fermentation but are outcompeted or less active in later stages. Pichia and Penicillium show a significant increase in the JASF group, suggesting their importance in later fermentation stages. Pichia is known for its ability to ferment a wide range of sugars and produce aromatic compounds, while Penicillium can contribute to the breakdown of more complex organic substances and potentially produce antimicrobial compounds that inhibit spoilage organisms. Other genera like Cephalotrichum, Pleospora, Neocallimastix, Alternaria, and Tausonia contribute to the overall fungal diversity. Cephalotrichum and Pleospora might be involved in the degradation of plant material, while Neocallimastix, although traditionally associated with rumen fungi, might play a similar role in the breakdown of fibrous material in the tubers. Alternaria and Tausonia can produce a variety of metabolites that may influence the flavor profile of the fermented juice [53,54].
3.2. Dynamic Changes of Secondary Metabolites during Fermentation
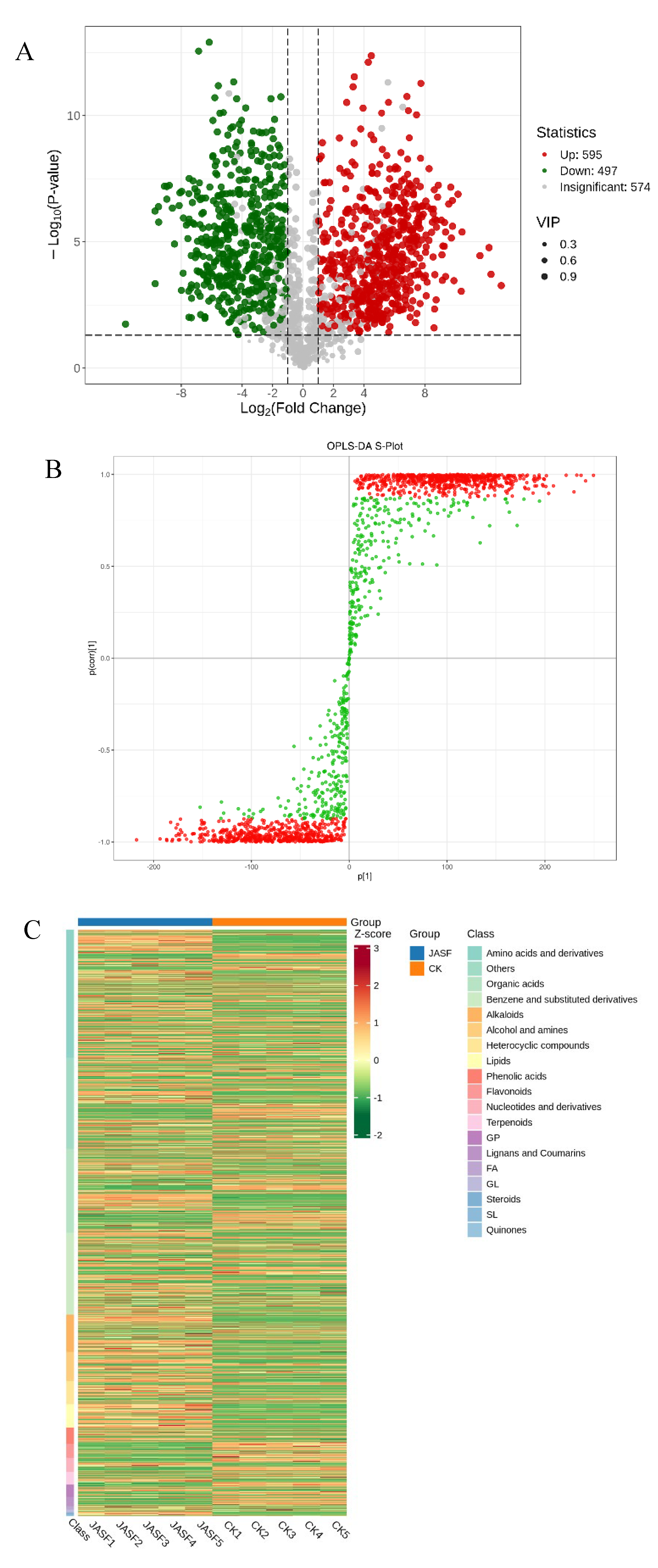
The Volcano Plot (Figure 3A) illustrates the relative abundance differences of metabolites between the CK and JASF groups and the statistical significance of these differences. In this plot, 595 metabolites are significantly upregulated in the experimental group, 497 metabolites are significantly downregulated, and 574 metabolites show no significant difference. The size of the dots indicates the VIP (Variable Importance in Projection) score, with larger dots representing higher VIP scores and suggesting greater importance in distinguishing between the two groups. The OPLS-DA S-plot (Figure 3B) provides a visual representation of the metabolites that contribute most significantly to the differences between the CK and JASF groups. Metabolites located closer to the top right and bottom left corners exhibit more significant differences and have VIP values greater than 1, indicating they are key biomarkers for the fermentation process. These metabolites play a critical role in distinguishing between the CK and JASF groups. Although metabolites with smaller VIP values contribute less to differentiating the two groups, they still affect the overall metabolic profile and may interact with key metabolites.
Further insights can be drawn from the heatmap of differential secondary metabolites between the CK and JASF groups (Figure 3C). Metabolites are categorized into various classes such as amino acids and derivatives, organic acids, alkaloids, benzene and substituted derivatives, alcohols and amines, heterocyclic compounds, phenolic acids, flavonoids, terpenoids, and more, each color-coded for easy identification [55]. The heatmap shows distinct patterns of metabolite abundance between the two groups, with the CK group generally exhibiting higher levels of certain metabolites compared to the JASF group, and vice versa for others. Notably, amino acids and derivatives, organic acids, and phenolic acids are prominently represented, indicating their significant roles in the fermentation process. The CK group tends to have higher levels of certain amino acids, organic acids, and phenolic acids, essential for the initial stages of fermentation, while the JASF group shows increased levels of other metabolites, such as specific alkaloids and terpenoids, which might be more relevant in later stages of fermentation or specific to the JASF conditions. This heatmap highlights the dynamic changes in secondary metabolite profiles between the CK and JASF groups, suggesting different active metabolic pathways and processes in each group.
The KEGG pathway enrichment analysis highlights significant metabolic pathway changes between the CK and JASF groups during the natural fermentation of Jerusalem artichoke tuber juice (Figure 3D). Phenylalanine metabolism, galactose metabolism, and tryptophan metabolism pathways are notably enriched, reflecting substantial gene expression changes in these areas. The biosynthesis of phenylpropanoids pathway is also significantly enriched, indicating an increase in secondary metabolites that contribute to flavor and antioxidant properties. The enrichment of drug metabolism via cytochrome P450 and fructose and mannose metabolism pathways underscores active microbial metabolism throughout fermentation. Enhanced degradation pathways for compounds, including aminobenzoate and caprolactam, suggest increased organic compound breakdown and transformation. Additionally, the biosynthesis of various plant secondary metabolites and changes in taste transduction pathways highlight the complexity of biochemical reactions. The metabolism of 2-oxocarboxylic acids and aromatic compounds further emphasizes the dynamic nature of metabolic activities during fermentation. These enriched pathways illustrate the intricate microbial interactions and metabolic processes that enhance the flavor, nutritional value, and safety of the fermented product.
3.3. Dynamic Changes of Volatile Flavor Compounds during Fermentation
During the spontaneous fermentation of Jerusalem artichoke juice, significant and diverse changes were observed in alcohol compounds, reflecting the complex biochemical reactions and microbial metabolic activities during fermentation. For example, the content of 3-Undecanol increased from 0.68 μg/L before fermentation to 24.29 μg/L after fermentation, and 1-Octanol increased from 3.79 μg/L to 170.37 μg/L. These significant increases indicate the microbial breakdown and utilization of sugars and other organic substances in Jerusalem artichoke during fermentation. Phenylethyl Alcohol showed a particularly significant increase from 113.08 μg/L to 15,675.14 μg/L, with its relative odor activity value (rOAV) also rising markedly. This increase suggests that Phenylethyl Alcohol, associated with floral and fruity aromas, contributes significantly to the post-fermentation product’s fragrance, likely making the juice more appealing to consumers. This change could be attributed to the metabolic activities of yeasts and other microbes, which produce a large amount of secondary metabolites, including alcohols and esters, during sugar breakdown. The total alcohol content increased from 552.14 μg/L to 17,155.1 μg/L post-fermentation, indicating the significant role of fermentation in enhancing the flavor complexity and aroma concentration of Jerusalem artichoke juice. This substantial change reflects vigorous microbial metabolic activity, leading to the generation of numerous new compounds and significant improvements in sensory properties.
During the spontaneous fermentation of Jerusalem artichoke juice, aldehydes showed significant changes, reflecting complex biochemical reactions and microbial metabolic activities. Comparing data before and after fermentation, 23 aldehyde compounds exhibited notable changes. The content of 10-Undecenal increased from 1.15 μg/L before fermentation to 15.20 μg/L after fermentation, (E,E)-2,4-Nonadienal increased from 10.71 μg/L to 59.63 μg/L, and 2,5-Dimethylbenzaldehyde increased from 21.22 μg/L to 118.21 μg/L. These significant increases indicate the microbial transformation and utilization of precursor substances in Jerusalem artichoke during fermentation. In contrast, Benzeneacetaldehyde decreased from 98.40 μg/L to 36.35 μg/L, suggesting a different metabolic pathway or consumption during the fermentation process. The total aldehyde content increased from 865.39 μg/L to 1,172.78 μg/L post-fermentation, highlighting the fermentation process’s role in enhancing the flavor complexity and aroma concentration of Jerusalem artichoke juice.
During the spontaneous fermentation of Jerusalem artichoke juice, acid compounds also showed significant changes, demonstrating the impact of microbial metabolic activity and diversity on flavor substances. Analyzing the changes in five major acid compounds provides deeper insights into their contributions to post-fermentation flavor. Hexanoic Acid increased from 0.23 μg/L before fermentation to 177.36 μg/L after fermentation, with its rOAV increasing from 0 to 0.06. This significant increase could be due to yeast breakdown of fatty acids during fermentation. Hexanoic Acid, a common fatty acid with strong fatty and fruity aromas, is often found in fermented beverages. 9-Decenoic acid increased from 0.64 μg/L to 43.76 μg/L. This acid, known for its unique spicy and fruity aromas, may result from the microbial conversion of unsaturated fatty acids. 4-Aminobutanoic acid (γ-aminobutyric acid) increased from 17.57 μg/L to 78.55 μg/L. γ-Aminobutyric acid, an important neurotransmitter with various bioactivities, is possibly produced through microbial amino acid metabolism, significantly enhancing the product’s health value and flavor. The total acid content increased from 33.27 μg/L to 382.24 μg/L post-fermentation, not only enhancing the sourness and overall flavor but also potentially improving the product’s antioxidant activity and health benefits.
Ketone compounds also exhibited significant changes during the spontaneous fermentation of Jerusalem artichoke juice, revealing the importance of microbial metabolism in flavor substance formation. Analyzing the changes in 24 ketone compounds, we found that Acetophenone increased from 1.28 μg/L before fermentation to 86.86 μg/L after fermentation, with its contribution to flavor becoming more significant. 5-Ethyl-3-Hydroxy-4-Methyl-2(5H)-Furanone increased significantly from 4.13 μg/L to 303.74 μg/L, a highly significant increase, with strong caramel and fruity aromas forming an important aroma substance during fermentation. 2-Methylcyclohexanone increased from 3.56 μg/L to 41.88 μg/L, reflecting its role in the flavor profile post-fermentation. 1-Nonen-3-one increased from 8.76 μg/L to 216.16 μg/L, contributing its unique metallic and sweet aromas commonly found in fruits and vegetables. 4-Undecanone increased from 28.48 μg/L to 170.27 μg/L, further enhancing the flavor profile. In contrast, 3-Methyl-4-Heptanone and 6-Methyl-3,5-Heptadien-2-one decreased from 170.38 μg/L to 72.88 μg/L and from 214.76 μg/L to 167.15 μg/L, respectively. Overall, most ketone compounds significantly increased during fermentation, with the total ketone content rising from 926.95 μg/L to 2,195.82 μg/L. This significant increase mainly results from microbial conversion and metabolic activity of precursor substances in Jerusalem artichoke juice during fermentation. Significance analysis showed that 20 out of 24 ketone compounds had statistically significant changes (p < 0.05), reflecting active microbial metabolism during fermentation and further demonstrating the key role of microbes in flavor substance formation.
Ester compounds also exhibited significant changes during the spontaneous fermentation of Jerusalem artichoke juice, revealing the importance of microbial metabolism in flavor substance formation. Analyzing the changes in 61 ester compounds, various esters showed significant changes before and after fermentation. Ethyl Decanoate increased from 0.06 μg/L before fermentation to 162.31 μg/L after fermentation, contributing significantly to post-fermentation flavor. Ethyl Hexanoate increased from 0.10 μg/L to 160.55 μg/L, enhancing the fruity and sweet aromas of the juice. Octyl Acetate increased from 0.35 μg/L to 61.37 μg/L, adding to the overall fruity aroma. 1-Methylbutyl Butanoate increased from 0.38 μg/L to 69.98 μg/L, and Ethyl Hexadecanoate increased from 0.56 μg/L to 67.52 μg/L, both contributing to the complex aroma profile. Ethyl Benzenepropanoate saw a significant increase from 0.86 μg/L to 2,088.29 μg/L, with its floral and sweet aromas enhancing the overall flavor. Ethyl 4-Methylpentanoate increased from 0.59 μg/L to 492.01 μg/L, adding significantly to the fruity aroma. Geranyl Formate increased from 1.16 μg/L to 138.01 μg/L, known for its strong fruity aromas. Pentyl Butanoate increased from 1.22 μg/L to 279.13 μg/L, adding banana and fruity aromas to the juice. 1-Isothiocyanato-2-Butene increased from 2.56 μg/L to 311.28 μg/L, and Butyl Butanoate increased from 12.14 μg/L to 233.19 μg/L, both significantly enhancing the aroma. 3-Methylbutyl Butanoate increased significantly from 12.74 μg/L to 1,406.55 μg/L, adding to the rich, fruity, and sweet aromas. Ethyl Butanoate increased from 189.73 μg/L to 1,056.87 μg/L, contributing to the rich aroma profile. In contrast, some ester compounds showed a decrease during fermentation. (E)-Methyl 3-Hexenoate, Ethyl Tiglate, and Methyl 2-Octynoate saw reductions in their concentrations, indicating changes in the microbial metabolic pathways during fermentation, affecting the overall flavor profile. Overall, most ester compounds significantly increased during fermentation, with the total ester content rising from 1,190.37 μg/L to 10,850.27 μg/L. This substantial increase mainly results from microbial conversion and metabolic activity of precursor substances during fermentation. Significance analysis showed that most of the 61 ester compounds had statistically significant changes (p < 0.05), reflecting active microbial metabolism during fermentation and further demonstrating the key role of microbes in flavor substance formation. High-content ester compounds formed during fermentation, such as hexyl acetate and ethyl decanoate, contributed significantly to the overall aroma and flavor of the fermented product.
During the spontaneous fermentation of Jerusalem artichoke juice, significant changes in various compounds were observed, reflecting the complex biochemical reactions and microbial activities. Microbial diversity plays a crucial role, with initial stages dominated by Flavobacterium, Sphingomonas, and Luteimonas breaking down complex substances like inulin, leading to flavor development [56]. As fermentation progresses, Lactobacillus, and Pediococcus become dominant, utilizing organic acids and producing flavor compounds [57,58]. Fungal shifts from Geosmithia and Alternaria to Pichia and Penicillium also contribute significantly, particularly in producing alcohols and esters [59]. Alcohols like Phenylethyl Alcohol, which saw a dramatic increase, are produced by yeast through the Ehrlich pathway, while aldehydes like 10-Undecenal and 2,5-Dimethylbenzaldehyde result from microbial oxidation of alcohols and amino acids. Acids such as Hexanoic Acid and 4-Aminobutanoic acid increase through microbial fermentation, enhancing the juice’s health benefits and antioxidant activity [60]. Ketones, including 5-Ethyl-3-hydroxy-4-methylfuran-2(5H)-one, form via microbial metabolism of fatty acids and amino acids, while esters like Ethyl Decanoate and Ethyl Hexanoate result from microbial esterification, contributing fruity and sweet aromas [61]. The significant role of inulin, a prebiotic, selectively promotes specific microbes, influencing community dynamics and metabolic activities, while reducing sugars are efficiently utilized by Saccharomyces and Pichia, enhancing the fruity and floral aromas [62]. These findings highlight the importance of microbial diversity, metabolic pathways, and substrate utilization in forming complex flavor profiles, providing insights for optimizing fermentation processes and improving the quality and flavor of fermented Jerusalem artichoke juice.

Table 1.
Changes of volatile flavor compounds during spontaneous fermentation.
| Volatile Compounds | RI | CAS | rOAV | Content(μg/L) | p-Value | |||
|---|---|---|---|---|---|---|---|---|
| CK | JASF | CK | JASF | |||||
| Alcohols | ||||||||
| 1 | cis-2-Furanmethanol, 5-Ethenyltetrahydro-α,α,5-Trimethyl | 1074 | 5989-33-3 | 0–1 | 0–1 | 0.28±0.02 | 5.30±0.60 | 0.0142 |
| 2 | 3-Undecanol | 1400 | 6929-08-4 | 0–1 | >1 | 0.68±0.02 | 24.29±0.30 | 0.0002 |
| 3 | 2,3-Dimethyl-2-Butanol | 720 | 594-60-5 | 0–1 | 0–1 | 1.36±0.05 | 7.58±0.57 | 0.0097 |
| 4 | 4,4-Dimethyl-2-Pentanol | 812 | 6144-93-0 | 0–1 | 0–1 | 1.36±0.03 | 7.58±0.50 | 0.0058 |
| 5 | α,α,4-Trimethyl-3-Cyclohexene-1-Methanethiol | 1283 | 71159-90-5 | >1 | >1 | 2.83±0.12 | 15.75±1.09 | 0.0056 |
| 6 | 2-Ethyl-1-Hexanol | 1029 | 104-76-7 | 0–1 | 0–1 | 3.59±0.24 | 20.01±1.18 | 0.0058 |
| 7 | 1-Octanol | 1070 | 111-87-5 | 0–1 | >1 | 3.79±0.06 | 170.37±14.04 | 0.0070 |
| 8 | 1-Undecanol | 1371 | 112-42-5 | 0–1 | 0–1 | 4.13±0.11 | 7.73±0.40 | 0.0188 |
| 9 | 2-Mercaptoethanol | 723 | 60-24-2 | 0–1 | 0–1 | 6.70±0.38 | 37.32±1.46 | 0.0035 |
| 10 | 1-Decanol | 1272 | 112-30-1 | 0–1 | >1 | 8.36±0.60 | 46.58±3.68 | 0.0119 |
| 11 | 6-Undecanol | 1281 | 23708-56-7 | 0–1 | >1 | 8.43±0.49 | 46.95±2.60 | 0.0030 |
| 12 | 2-Nonanol | 1099 | 628-99-9 | 0–1 | 0–1 | 10.32±0.60 | 4.20±0.21 | 0.0045 |
| 13 | 3-Methyl-4-Heptanol | 997 | 1838-73-9 | 0–1 | >1 | 14.79±0.59 | 207.21±9.49 | 0.0027 |
| 14 | 5-Hexen-1-ol | 868 | 821-41-0 | 0–1 | 0–1 | 17.70±0.85 | 15.85±0.93 | 0.4025 |
| 15 | 2-Butoxyethanol | 905 | 111-76-2 | 0–1 | 0–1 | 18.04±0.76 | 100.47±5.98 | 0.0044 |
| 16 | 2-Heptanol | 900 | 543-49-7 | 0–1 | >1 | 23.58±1.58 | 131.35±13.45 | 0.0189 |
| 17 | 6-Ethenyltetrahydro-2,2,6-Trimethyl-2H-Pyran-3-ol | 1173 | 14049-11-7 | 0–1 | 0–1 | 25.69±0.26 | 154.34±13.49 | 0.0112 |
| 18 | 2-Undecanol | 1301 | 1653-30-1 | >1 | >1 | 27.97±1.81 | 155.82±12.67 | 0.0081 |
| 19 | trans,cis-2,6-Nonadien-1-ol | 1170 | 28069-72-9 | >1 | >1 | 31.48±1.00 | 126.11±9.14 | 0.0078 |
| 20 | Phenylethyl Alcohol | 1116 | 60-12-8 | 0–1 | >1 | 113.08±10.17 | 15675.14±834.64 | 0.0028 |
| 21 | Hotrienol | 1106 | 20053-88-7 | >1 | >1 | 227.97±7.85 | 195.16±9.53 | 0.1997 |
| Aldehydes | ||||||||
| 22 | 3-Cyclohexene-1-Carboxaldehyde | 958 | 100-50-5 | 0–1 | 0–1 | 0.39±0.02 | 2.16±0.01 | 0.0002 |
| 23 | (Z)-3-Phenylacrylaldehyde | 1219 | 57194-69-1 | 0–1 | 0–1 | 0.50±0.02 | 2.79±0.05 | 0.0002 |
| 24 | 5-Methyl-2-Thiophenecarboxaldehyde | 1118 | 13679-70-4 | 0–1 | >1 | 0.99±0.05 | 5.49±0.17 | 0.0014 |
| 25 | 10-Undecenal | 1297 | 112-45-8 | 0–1 | >1 | 1.15±0.03 | 15.20±0.46 | 0.0011 |
| 26 | (Z,Z)-3,6-Nonadienal | 1100 | 21944-83-2 | >1 | >1 | 1.18±0.13 | 6.60±0.27 | 0.0029 |
| 27 | 2-Ethyl-2-Hexenal | 999 | 645-62-5 | 0–1 | 0–1 | 1.97±0.10 | 2.59±0.09 | 0.0450 |
| 28 | Glutaraldehyde | 895 | 111-30-8 | >1 | >1 | 4.65±0.34 | 25.91±0.48 | 0.0001 |
| 29 | Piperonal | 1334 | 120-57-0 | 0–1 | 0–1 | 5.07±0.22 | 3.17±0.22 | 0.0020 |
| 30 | (E)-4-Decenal | 1198 | 65405-70-1 | 0–1 | >1 | 6.00±0.30 | 39.83±4.79 | 0.0177 |
| 31 | 3-Methylbenzaldehyde | 1070 | 620-23-5 | 0–1 | 0–1 | 9.27±0.73 | 51.65±0.22 | 0.0005 |
| 32 | Heptanal | 901 | 111-71-7 | >1 | >1 | 9.50±0.43 | 52.90±3.14 | 0.0067 |
| 33 | 2-Nonenal | 1161 | 2463-53-8 | >1 | >1 | 10.29±0.71 | 57.32±3.67 | 0.0056 |
| 34 | (E,E)-2,4-Octadienal | 1115 | 30361-28-5 | 0–1 | 0–1 | 10.71±0.80 | 59.63±4.57 | 0.0096 |
| 35 | 4-(1-Methylethenyl)-1-Cyclohexene-1-Carboxaldehyde | 1274 | 2111-75-3 | 0–1 | >1 | 11.52±0.75 | 64.19±3.76 | 0.0063 |
| 36 | (Z)-6-Nonenal | 1104 | 2277-19-2 | >1 | >1 | 14.56±0.33 | 7.88±0.67 | 0.0056 |
| 37 | 2,5-Dimethylbenzaldehyde | 1154 | 5779-94-2 | 0–1 | 0–1 | 17.47±0.29 | 3.89±0.36 | 0.0002 |
| 38 | (E,E)-2,4-Nonadienal | 1216 | 5910-87-2 | >1 | >1 | 21.22±1.28 | 118.21±11.23 | 0.0161 |
| 39 | Tridecanal | 1513 | 10486-19-8 | 0–1 | 0–1 | 45.38±2.90 | 53.60±0.37 | 0.0887 |
| 40 | (S)-4-(1-Methylethenyl)-1-Cyclohexene-1-Carboxaldehyde | 1243 | 18031-40-8 | >1 | >1 | 49.49±2.36 | 34.55±0.13 | 0.0240 |
| 41 | Benzeneacetaldehyde | 1046 | 122-78-1 | >1 | >1 | 98.40±2.14 | 36.35±1.20 | 0.0026 |
| 42 | Nonanal | 1105 | 124-19-6 | >1 | >1 | 117.08±2.46 | 90.79±4.02 | 0.0526 |
| 43 | (Z)-2-Decenal | 1252 | 2497-25-8 | >1 | >1 | 170.39±13.31 | 128.60±2.42 | 0.0627 |
| 44 | 4-(1,1-Dimethylethyl)benzenepropanal | 1521 | 18127-01-0 | >1 | >1 | 258.20±14.06 | 309.48±23.63 | 0.2868 |
| Acids | ||||||||
| 45 | Hexanoic Acid | 987 | 142-62-1 | 0–1 | 0–1 | 0.23±0.01 | 177.36±8.39 | 0.0022 |
| 46 | 9-Decenoic Acid | 1360 | 14436-32-9 | 0–1 | 0–1 | 0.64±0.06 | 43.76±4.64 | 0.0111 |
| 47 | 4-Methyloctanoic Acid | 1232 | 54947-74-9 | 0–1 | 0–1 | 2.35±0.03 | 13.06±0.81 | 0.0054 |
| 48 | (E)-2-Hexenoic Acid | 1045 | 13419-69-7 | 0–1 | 0–1 | 12.48±0.95 | 69.51±1.08 | 0.0001 |
| 49 | 4-Aminobutanoic Acid | 1190 | 56-12-2 | 0–1 | 0–1 | 17.57±1.88 | 78.55±2.82 | 0.0010 |
| Ketones | ||||||||
| 50 | 2-Undecanone | 1295 | 112-12-9 | 0–1 | >1 | 0.58±0.03 | 10.70±0.42 | 0.0020 |
| 51 | Isophorone | 1123 | 78-59-1 | 0–1 | 0–1 | 1.24±0.10 | 6.92±0.39 | 0.0029 |
| 52 | 1-(4-Methylphenyl)ethanone | 1183 | 122-00-9 | 0–1 | 0–1 | 1.25±0.05 | 6.00±0.09 | 0.0001 |
| 53 | Acetophenone | 1068 | 98-86-2 | 0–1 | >1 | 1.28±0.13 | 86.86±10.04 | 0.0131 |
| 54 | 2-Dodecanone | 1395 | 6175-49-1 | 0–1 | 0–1 | 1.47±0.10 | 4.93±0.23 | 0.0022 |
| 55 | (E,E)-3,5-Octadien-2-one | 1073 | 30086-02-3 | >1 | >1 | 2.31±0.04 | 12.87±1.30 | 0.0150 |
| 56 | (E)-5,9-Undecadien-2-one, 6,10-Dimethyl | 1453 | 3796-70-1 | 0–1 | 0–1 | 3.35±0.11 | 2.62±0.06 | 0.0502 |
| 57 | 2-Octanone | 991 | 111-13-7 | 0–1 | 0–1 | 3.50±0.28 | 19.48±1.15 | 0.0030 |
| 58 | 2-Methylcyclohexanone | 953 | 583-60-8 | 0–1 | 0–1 | 3.56±0.09 | 41.88±2.23 | 0.0036 |
| 59 | 5-Ethyl-3-Hydroxy-4-Methyl-2(5H)-Furanone | 1195 | 698-10-2 | >1 | >1 | 4.13±0.33 | 303.74±23.36 | 0.0060 |
| 60 | 1-(4,5-Dihydro-2-Thiazolyl)ethanone | 1106 | 29926-41-8 | >1 | >1 | 4.64±0.06 | 25.86±2.12 | 0.0096 |
| 61 | 3-Butylisobenzofuran-1(3H)-one | 1656 | 6066-49-5 | 0–1 | 0–1 | 5.33±0.39 | 3.17±0.21 | 0.0427 |
| 62 | 1-Nonen-3-one | 1076 | 24415-26-7 | >1 | >1 | 8.76±0.79 | 216.16±8.62 | 0.0015 |
| 63 | 3-Octen-2-one | 1016 | 1669-44-9 | >1 | >1 | 11.08±0.28 | 61.71±4.26 | 0.0079 |
| 64 | 3-Decanone | 1187 | 928-80-3 | 0–1 | >1 | 13.10±0.32 | 72.95±2.82 | 0.0022 |
| 65 | 4-(2,6,6-Trimethylcyclohexa-1,3-Dienyl)but-3-en-2-one | 1485 | 1203-08-3 | >1 | >1 | 14.06±1.51 | 6.96±0.06 | 0.0403 |
| 66 | 1-(2,6,6-Trimethyl-1,3-Cyclohexadien-1-yl)-2-Buten-1-one | 1362 | 23696-85-7 | >1 | >1 | 17.67±0.33 | 17.37±1.11 | 0.8407 |
| 67 | 1-(2-Thienyl)ethanone | 1092 | 88-15-3 | >1 | >1 | 17.95±0.88 | 43.99±0.52 | 0.0010 |
| 68 | 4-Undecanone | 1208 | 14476-37-0 | 0–1 | >1 | 28.48±2.43 | 170.27±9.34 | 0.0024 |
| 69 | 2-Sec-Butylcyclohexanone | 1220 | 14765-30-1 | 0–1 | >1 | 75.95±3.34 | 423.10±13.08 | 0.0022 |
| 70 | 2-Hydroxy-3,4-Dimethyl-2-Cyclopenten-1-one | 1075 | 21835-00-7 | >1 | >1 | 141.02±4.04 | 136.87±10.86 | 0.8055 |
| 71 | 3-Methyl-4-Heptanone | 928 | 15726-15-5 | >1 | >1 | 170.38±17.84 | 72.88±1.21 | 0.0287 |
| 72 | 3,4-Dimethyl-1,2-Cyclopentadione | 1109 | 13494-06-9 | >1 | >1 | 181.10±9.76 | 281.39±4.42 | 0.0192 |
| 73 | 6-Methyl-3,5-Heptadien-2-one | 1107 | 1604-28-0 | >1 | >1 | 214.76±8.41 | 167.15±14.82 | 0.1383 |
| Esters | ||||||||
| 74 | Ethyl Decanoate | 1396 | 110-38-3 | 0–1 | >1 | 0.06±0.00 | 162.31±13.54 | 0.0069 |
| 75 | Ethyl Hexanoate | 999 | 123-66-0 | 0–1 | >1 | 0.10±0.01 | 160.55±3.85 | 0.0006 |
| 76 | Octyl Acetate | 1210 | 112-14-1 | 0–1 | 0–1 | 0.35±0.05 | 61.37±5.29 | 0.0074 |
| 77 | 1-Methylbutyl Butanoate | 970 | 60415-61-4 | 0–1 | >1 | 0.38±0.02 | 69.98±5.37 | 0.0059 |
| 78 | Ethyl Hexadecanoate | 1993 | 628-97-7 | 0–1 | 0–1 | 0.56±0.01 | 67.52±4.04 | 0.0036 |
| 79 | Ethyl 4-Methylpentanoate | 969 | 25415-67-2 | 0–1 | >1 | 0.59±0.05 | 492.01±27.89 | 0.0032 |
| 80 | 1-Methylpropyl 2-Methylbutanoate | 971 | 869-08-9 | 0–1 | 0–1 | 0.78±0.05 | 4.32±0.12 | 0.0021 |
| 81 | Ethyl Benzenepropanoate | 1353 | 2021-28-5 | 0–1 | >1 | 0.86±0.05 | 2088.29±67.40 | 0.0010 |
| 82 | 4-tert-Butylcyclohexyl Acetate | 1368 | 32210-23-4 | 0–1 | 0–1 | 0.90±0.08 | 23.44±1.30 | 0.0037 |
| 83 | Ethyl 9-Decenoate | 1388 | 67233-91-4 | 0–1 | 0–1 | 1.00±0.00 | 34.92±3.26 | 0.0091 |
| 84 | 2-Ethylhexyl Acrylate | 1220 | 103-11-7 | 0–1 | 0–1 | 1.14±0.08 | 14.04±0.53 | 0.0019 |
| 85 | Geranyl Formate | 1301 | 105-86-2 | 0–1 | 0–1 | 1.16±0.09 | 138.01±2.15 | 0.0002 |
| 86 | Pentyl Butanoate | 1077 | 540-18-1 | 0–1 | 0–1 | 1.22±0.07 | 279.13±7.96 | 0.0008 |
| 87 | Hexyl Acetate | 1013 | 142-92-7 | 0–1 | 0–1 | 1.24±0.06 | 6.90±0.09 | 0.0002 |
| 88 | Pentyl 2-Methylbutanoate | 1142 | 68039-26-9 | 0–1 | >1 | 1.25±0.06 | 24.67±1.29 | 0.0028 |
| 89 | Methyl 4-Methoxybenzoate | 1373 | 121-98-2 | 0–1 | 0–1 | 1.58±0.08 | 3.45±0.11 | 0.0096 |
| 90 | trans-3-Methyl-4-Octanolide | 1288 | 39638-67-0 | 0–1 | 0–1 | 1.86±0.07 | 0.41±0.02 | 0.0034 |
| 91 | Methyl Anthranilate | 1349 | 134-20-3 | 0–1 | >1 | 1.92±0.12 | 100.47±0.93 | 0.0001 |
| 92 | Butyl 2-Hydroxybenzoate | 1436 | 2052-14-4 | 0–1 | 0–1 | 1.93±0.11 | 2.13±0.13 | 0.4387 |
| 93 | Hexyl 2-Methylbutanoate | 1236 | 10032-15-2 | 0–1 | 0–1 | 2.00±0.11 | 1.62±0.04 | 0.0323 |
| 94 | 1,2-Ethanediol, Diacetate | 991 | 111-55-7 | 0–1 | 0–1 | 2.03±0.14 | 11.28±1.06 | 0.0145 |
| 95 | 2-Ethylhexyl Methacrylate | 1296 | 688-84-6 | 0–1 | 0–1 | 2.15±0.17 | 28.94±1.30 | 0.0029 |
| 96 | 1-Isothiocyanato-2-Butene | 1070 | 2253-93-2 | 0–1 | 0–1 | 2.56±0.03 | 311.28±18.03 | 0.0034 |
| 97 | Phenyl Acetate | 1062 | 122-79-2 | 0–1 | 0–1 | 2.61±0.12 | 10.34±0.86 | 0.0091 |
| 98 | 2-Ethylhexyl Acetate | 1185 | 103-09-3 | 0–1 | 0–1 | 2.94±0.11 | 16.37±0.50 | 0.0009 |
| 99 | 1-Ethylpropyl Acetate | 793 | 620-11-1 | 0–1 | >1 | 3.10±0.34 | 194.85±4.87 | 0.0006 |
| 100 | δ-Dodecalactone | 1720 | 713-95-1 | 0–1 | 0–1 | 3.45±0.17 | 4.04±0.04 | 0.0464 |
| 101 | Methyl Heptanoate | 1024 | 106-73-0 | 0–1 | >1 | 3.56±0.12 | 19.85±0.98 | 0.0044 |
| 102 | Ethyl Dodecanoate | 1595 | 106-33-2 | 0–1 | 0–1 | 4.27±0.20 | 23.78±1.75 | 0.0095 |
| 103 | 3-Phenylpropyl Acetate | 1373 | 122-72-5 | 0–1 | 0–1 | 4.28±0.07 | 6.10±0.56 | 0.0701 |
| 104 | 3-Methylphenylmethyl Butanoate | 1396 | 103-38-8 | 0–1 | >1 | 5.49±0.50 | 30.60±1.68 | 0.0027 |
| 105 | cis-2-Methyl-5-(1-Methylethenyl)-2-Cyclohexen-1-ol Acetate | 1362 | 1205-42-1 | 0–1 | >1 | 5.63±0.24 | 31.37±5.71 | 0.0463 |
| 106 | Butyl Hexanoate | 1189 | 626-82-4 | 0–1 | 0–1 | 6.39±0.30 | 32.25±0.62 | 0.0004 |
| 107 | Pentyl Acetate | 916 | 628-63-7 | 0–1 | 0–1 | 6.69±0.32 | 37.27±2.74 | 0.0072 |
| 108 | 4-Methylphenyl Acetate | 1171 | 140-39-6 | 0–1 | >1 | 8.00±0.33 | 36.87±1.79 | 0.0051 |
| 109 | 3-Methylbutyl Butanoate | 1046 | 109-19-3 | 0–1 | 0–1 | 8.41±0.45 | 46.84±1.05 | 0.0015 |
| 110 | Ethyl Nonanoate | 1295 | 123-29-5 | 0–1 | >1 | 9.00±0.58 | 50.16±2.23 | 0.0038 |
| 111 | Methyl Thiocyanate | 702 | 556-64-9 | 0–1 | >1 | 9.51±0.92 | 52.95±1.39 | 0.0026 |
| 112 | Ethyl 3-Methylpentanoate | 960 | 5870-68-8 | >1 | >1 | 9.52±0.93 | 295.96±2.07 | 0.0000 |
| 113 | Tetrahydro-6-Pentyl-2H-Pyran-2-one | 1502 | 705-86-2 | 0–1 | 0–1 | 10.89±0.36 | 43.16±2.44 | 0.0069 |
| 114 | Methyl Decanoate | 1326 | 110-42-9 | >1 | >1 | 11.03±0.30 | 17.51±0.57 | 0.0172 |
| 115 | 2-Methylbutyl 2-Methylbutanoate | 1105 | 2445-78-5 | 0–1 | >1 | 11.63±1.03 | 148.48±6.08 | 0.0023 |
| 116 | 2-Phenylethyl 3-Methylbutanoate | 1491 | 140-26-1 | >1 | >1 | 11.65±1.16 | 64.87±3.29 | 0.0043 |
| 117 | Butyl Butanoate | 996 | 109-21-7 | 0–1 | >1 | 12.14±0.26 | 233.19±15.59 | 0.0049 |
| 118 | Ethyl Pentanoate | 902 | 539-82-2 | 0–1 | >1 | 12.16±0.18 | 67.76±3.27 | 0.0033 |
| 119 | Ethyl Benzeneacetate | 1247 | 101-97-3 | 0–1 | 0–1 | 12.52±1.44 | 71.91±3.06 | 0.0016 |
| 120 | Dihydro-5-Propyl-2(3H)-Furanone | 1156 | 105-21-5 | 0–1 | 0–1 | 12.71±0.46 | 70.78±3.50 | 0.0028 |
| 121 | 3-Methylbutyl Butanoate | 1056 | 106-27-4 | 0–1 | >1 | 12.74±0.12 | 1406.55±18.28 | 0.0002 |
| 122 | n-Amyl Isovalerate | 1110 | 25415-62-7 | 0–1 | 0–1 | 18.84±0.91 | 473.98±30.74 | 0.0044 |
| 123 | Isothiocyanatoethane | 796 | 542-85-8 | 0–1 | 0–1 | 20.83±0.96 | 205.69±30.54 | 0.0247 |
| 124 | Propyl Propanoate | 810 | 106-36-5 | 0–1 | >1 | 22.72±0.54 | 126.54±7.70 | 0.0051 |
| 125 | 3-Methyl-1-Butanol Acetate | 878 | 123-92-2 | >1 | >1 | 24.93±0.67 | 471.94±35.83 | 0.0066 |
| 126 | Propyl Butanoate | 899 | 105-66-8 | >1 | >1 | 30.20±0.85 | 168.23±4.58 | 0.0010 |
| 127 | (E)-Methyl 3-Hexenoate | 920 | 13894-61-6 | 0–1 | 0–1 | 34.56±1.43 | 7.70±0.67 | 0.0009 |
| 128 | n-Butyl Tiglate | 1134 | 7785-66-2 | >1 | >1 | 51.65±2.22 | 287.71±2.48 | 0.0001 |
| 129 | Isopentyl Hexanoate | 1250 | 2198-61-0 | 0–1 | 0–1 | 59.91±1.01 | 39.01±0.71 | 0.0023 |
| 130 | Butyl Acetate | 815 | 123-86-4 | >1 | >1 | 99.48±5.70 | 554.18±17.65 | 0.0017 |
| 131 | 2-Butoxyethyl Acetate | 1090 | 112-07-2 | 0–1 | 0–1 | 102.40±4.71 | 199.72±6.11 | 0.0118 |
| 132 | Ethyl Tiglate | 939 | 5837-78-5 | >1 | >1 | 150.65±11.45 | 66.08±2.19 | 0.0223 |
| 133 | Methyl 2-Octynoate | 1202 | 111-12-6 | >1 | >1 | 156.55±12.81 | 91.76±6.52 | 0.0758 |
| 134 | Ethyl Butanoate | 802 | 105-54-4 | >1 | >1 | 189.73±15.80 | 1056.87±79.69 | 0.0109 |
4. Conclusion
This study revealed the changes in microbial communities, metabolic transformations, and flavor characteristics during the spontaneous fermentation of Jerusalem artichoke juice. During fermentation, the dominant microbial groups shifted from Enterobacteriaceae and Bacillaceae to Lactobacillaceae and Acetobacteraceae, with significant increases in Actinobacteria and Bacteroidetes post-fermentation. Secondary metabolites, including those involved in phenylalanine and tryptophan metabolic pathways, were significantly enriched, along with enhanced lipid and nucleotide metabolic activities, indicating increased metabolic activity during fermentation. Notably, the significant increase in phenylpropanoid and aromatic amino acid metabolites highlighted the importance of these pathways in improving the quality of fermented products. Flavor analysis showed significant increases in alcohols, esters, acids, and ketones, imparting unique and rich aromas and health benefits to the fermented Jerusalem artichoke juice. Specifically, the significant increases in Phenylethyl Alcohol, Ethyl Benzenepropanoate, 3-Methylbutyl Butanoate, Ethyl 4-Methylpentanoate, 5-Ethyl-3-Hydroxy-4-Methyl-2(5H)-Furanone, Ethyl Decanoate, Hexanoic Acid, 1-Octanol, GABA enhanced the overall flavor and sensory quality.
The advantage of spontaneous fermentation lies in its ability to retain the natural flavor and nutritional components of the raw materials without adding exogenous strains, producing a wealth of secondary metabolites. The dynamic changes in microbial populations and metabolic activities during fermentation are closely related, with specific groups such as Lactobacillaceae and Acetobacteraceae playing key roles in the later stages of fermentation, significantly affecting the flavor and quality of Jerusalem artichoke juice.
However, the downside of spontaneous fermentation is its sensitivity to environmental factors, making it difficult to control the composition of microbial communities and the changes in metabolic products, potentially leading to product quality instability. These findings provide a scientific basis for optimizing fermentation processes, strain enhancement, and new technology applications, helping to further improve the quality and market competitiveness of fermented Jerusalem artichoke products.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Conceptualization, T.Z.; methodology, T.Z; software, Z.L.; validation, Z.L., and T.Z.; formal analysis, X.L.; investigation, C.C., and X.L.; resources, C.C.; data curation, T.Z.; writing—original draft preparation, T.Z.; writing—review and editing, Y.M.; visualization, C.C.; supervision, Y.M.; project admin-istration, Y.M.; funding acquisition, T.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Gansu Academy of Agricultural Sciences Scientific Research Conditions Construction and Achievement Transformation Project (2022GAAS50) and the Science & Technology Project of Gansu Agricultural and Rural Affairs Department (GNKJ-2020-4).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data is contained within the article or supplementary material: The data presented in this study are available in insert article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Chyc, M.; Ogonowski, J. Jerusalem artichoke as a prospective raw material for industry. Przemysl Chemiczny 2015, 94, 578–582. [Google Scholar]
- Qin, Y.-Q.; Wang, L.-Y.; Yang, X.-Y.; Xu, Y.-J.; Fan, G.; Fan, Y.-G.; Ren, J.-N.; An, Q.; Li, X. Inulin: properties and health benefits. Food & Function 2023, 14, 2948–2968. [Google Scholar] [CrossRef]
- Riva, A.; Rasoulimehrabani, H.; Cruz-Rubio, J.M.; Schnorr, S.L.; von Baeckmann, C.; Inan, D.; Nikolov, G.; Herbold, C.W.; Hausmann, B.; Pjevac, P.; et al. Identification of inulin-responsive bacteria in the gut microbiota via multi-modal activity-based sorting. Nature Communications 2023, 14. [Google Scholar] [CrossRef] [PubMed]
- Tawfick, M.M.; Xie, H.; Zhao, C.; Shao, P.; Farag, M.A. Inulin fructans in diet: Role in gut homeostasis, immunity, health outcomes and potential therapeutics. International Journal of Biological Macromolecules 2022, 208, 948–961. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, C.M.; Davy, B.M.; Ponder, M.A.; McMillan, R.P.; Hughes, M.D.; Hulver, M.W.; Neilson, A.P.; Davy, K.P. Prebiotic Inulin Supplementation and Peripheral Insulin Sensitivity in adults at Elevated Risk for Type 2 Diabetes: A Pilot Randomized Controlled Trial. Nutrients 2021, 13. [Google Scholar] [CrossRef] [PubMed]
- Guimaraes, J.B.; Rodrigues, V.F.; Pereira, I.S.; Manso, G.M.d.C.; Elias-Oliveira, J.; Leite, J.A.; Waldetario, M.C.G.M.; de Oliveira, S.; Gomes, A.B.d.S.P.; Faria, A.M.C.; et al. Inulin prebiotic ameliorates type 1 diabetes dictating regulatory T cell homing via CCR4 to pancreatic islets and butyrogenic gut microbiota in murine model. Journal of Leukocyte Biology 2024, 115, 483–496. [Google Scholar] [CrossRef] [PubMed]
- Jia, S.; Li, J.; Yu, B.; Li, M.; Cui, B. Improvement of myocardial injury and gut microbiota disturbance in type 2 diabetic mice by inulin with various degrees of polymerization. Food Bioscience 2023, 51. [Google Scholar] [CrossRef]
- Tang, Z.; Shao, T.; Gao, L.; Yuan, P.; Ren, Z.; Tian, L.; Liu, W.; Liu, C.; Xu, X.; Zhou, X.; et al. Structural elucidation and hypoglycemic effect of an inulin-type fructan extracted from Stevia rebaudiana roots. Food & Function 2023, 14, 2518–2529. [Google Scholar] [CrossRef]
- Bakirhan, H.; Karabudak, E. Effects of inulin on calcium metabolism and bone health. International Journal for Vitamin and Nutrition Research 2023, 93, 85–96. [Google Scholar] [CrossRef]
- Gomez-Betancur, A.M.; Carmona-Tamayo, R.; Martinez-Alyarez, O.L.; Casanova-Yepes, H.; Torres-Oquendo, J.D. Effect of fat substitution using long-chain inulin and fortification with microencapsulated calcium in the rheological and sensory properties of yogurt mousse. Journal of Food Process Engineering 2022, 45. [Google Scholar] [CrossRef]
- Sulejmani, E.; Boran, O.S.; Huppertz, T.; Hayaloglu, A.A. Rheology, microstructure and sensory properties of low-fat milk jam: Influence of inulin type, sucrose content, sodium bicarbonate and calcium chloride. International Dairy Journal 2021, 123. [Google Scholar] [CrossRef]
- Kowalczyk, M.; Znamirowska-Piotrowska, A.; Buniowska-Olejnik, M.; Zagula, G.; Pawlos, M. Bioavailability of Macroelements from Synbiotic Sheep’s Milk Ice Cream. Nutrients 2023, 15. [Google Scholar] [CrossRef] [PubMed]
- Omori, K.; Miyakawa, H.; Watanabe, A.; Nakayama, Y.; Lyu, Y.; Ichikawa, N.; Sasaki, H.; Shibata, S. The Combined Effects of Magnesium Oxide and Inulin on Intestinal Microbiota and Cecal Short-Chain Fatty Acids. Nutrients 2021, 13. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, J.; Nagashima, T. Preparation of dried chips from Jerusalem artichoke (Helianthus tuberosus) tubers and analysis of their functional properties. Food Chemistry 2011, 126, 922–926. [Google Scholar] [CrossRef]
- Showkat, M.M.; Falck-Ytter, A.B.; Straetkvern, K.O. Phenolic Acids in Jerusalem Artichoke (Helianthus tuberosus L.): Plant Organ Dependent Antioxidant Activity and Optimized Extraction from Leaves. Molecules 2019, 24. [Google Scholar] [CrossRef] [PubMed]
- Dias, N.S.; Ferreira, J.F.S.; Liu, X.; Suarez, D.L. Jerusalem artichoke (Helianthus tuberosus L.) maintains high inulin, tuber yield, and antioxidant capacity under moderately-saline irrigation waters. Industrial Crops and Products 2016, 94, 1009–1024. [Google Scholar] [CrossRef]
- Mu, Y.; Gao, W.; Lv, S.; Li, F.; Lu, Y.; Zhao, C. The antioxidant capacity and antioxidant system of Jerusalem artichoke (Helianthus tuberosus L.) tubers in relation to inulin during storage at different low temperatures. Industrial Crops and Products 2021, 161. [Google Scholar] [CrossRef]
- Kolniak-Ostek, J.; Kita, A.; Peksa, A.; Wawrzyniak, A.; Hamulka, J.; Jeznach, M.; Danilcenko, H.; Jariene, E. Analysis of the content of bioactive compounds in selected flours and enriched extruded corn products. Journal of Food Composition and Analysis 2017, 64, 147–155. [Google Scholar] [CrossRef]
- Zalan, Z.; Hudacek, J.; Toth-Markus, M.; Husova, E.; Solichova, K.; Hegyi, F.; Plockova, M.; Chumchalova, J.; Halasz, A. Sensorically and antimicrobially active metabolite production of Lactobacillus strains on Jerusalem artichoke juice. Journal of the Science of Food and Agriculture 2011, 91, 672–679. [Google Scholar] [CrossRef] [PubMed]
- Celik, I.; Isik, F.; Gursoy, O.; Yilmaz, Y. Use of Jerusalem artichoke (Helianthus Tuberosus) tubers as a natural source of inulin in cakes. Journal of Food Processing and Preservation 2013, 37, 483–488. [Google Scholar] [CrossRef]
- Radovanovic, A.; Stojceska, V.; Plunkett, A.; Jankovic, S.; Milovanovic, D.; Cupara, S. The use of dry Jerusalem artichoke as a functional nutrient in developing extruded food with low glycaemic index. Food Chemistry 2015, 177, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Diaz, A.; Bomben, R.; Dini, C.; Vina, S.Z.; Garcia, M.A.; Ponzi, M.; Comelli, N. Jerusalem artichoke tuber flour as a wheat flour substitute for biscuit elaboration. Lwt-Food Science and Technology 2019, 108, 361–369. [Google Scholar] [CrossRef]
- Diaz, A.; Garcia, M.A.; Dini, C. Jerusalem artichoke flour as food ingredient and as source of fructooligosaccharides and inulin. Journal of Food Composition and Analysis 2022, 114. [Google Scholar] [CrossRef]
- Dimitrovski, D.; Velickova, E.; Dimitrovska, M.; Langerholc, T.; Winkelhausen, E. Synbiotic functional drink from Jerusalem artichoke juice fermented by probiotic Lactobacillus plantarum PCS26. Journal of Food Science and Technology-Mysore 2016, 53, 766–774. [Google Scholar] [CrossRef] [PubMed]
- Sooresh, M.M.; Willing, B.P.; Bourrie, B.C.T. Opportunities and challenges of understanding community assembly in spontaneous food fermentation. Foods 2023, 12. [Google Scholar] [CrossRef] [PubMed]
- Gao, Q.; Song, Y.; Liang, Y.; Li, Y.; Chang, Y.; Ma, R.; Cao, X.; Wang, S. Dynamics of physicochemical properties, functional compounds and antioxidant capacity during spontaneous fermentation of Lycium ruthenicum Murr. (Qinghai-Tibet Plateau) natural vinegar. Foods 2022, 11. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Z.; Zhou, Q.; Chen, Q.; Gao, J.; Wu, Y.; Yang, F.; Zhong, K.; Gao, H. Improvement of physicochemical characteristics, flavor profiles and functional properties in Chinese radishes via spontaneous fermentation after drying. Journal of Food Science 2023, 88, 1292–1307. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Sun, F.; Wang, W.; Liu, Y.; Wang, J.; Sun, J.; Mu, J.; Gao, Z. Effects of spontaneous fermentation on the microorganisms diversity and volatile compounds during ‘Marselan’ from grape to wine. Lwt-Food Science and Technology 2020, 134. [Google Scholar] [CrossRef]
- Kumar, S.; Chhabra, V.; Shenoy, S.; Daksh, R.; Ravichandiran, V.; Swamy, R.S.; Kumar, N. Role of flavonoids in modulation of mitochondria dynamics during oxidative stress. Mini-Reviews in Medicinal Chemistry 2024, 24, 908–919. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Zhang, X.; Wang, S.; Gao, X.; Zhang, X. Research progress on extraction and detection technologies of flavonoid compounds in foods. Foods 2024, 13. [Google Scholar] [CrossRef] [PubMed]
- Jin, L.; Xiumin, L.; Xiang, Z.; Bang-Ce, Y. Recent advances in microbial metabolic engineering for production of natural phenolic acids. Journal of Agricultural and Food Chemistry 2024, 72, 4538–4551. [Google Scholar]
- Yuan, G.; Shuai, M.; Min, D.; Meng, W.; Xiaoyuan, F. Progress in research on the biosynthesis pathway and metabolic regulation of phenolic acids. Food Science, China 2018, 39, 286–293. [Google Scholar]
- Yang, X.; Lan, W.; Sun, X. Antibacterial and antioxidant properties of phenolic acid grafted chitosan and its application in food preservation: A review. Food Chemistry 2023, 428. [Google Scholar] [CrossRef] [PubMed]
- Saini, A.; Seni, K.; Chawla, P.A.; Chawla, V.; Ganti, S.S. An insight into recent updates on analytical techniques for bioactive alkaloids. Phytochemical Analysis 2024, 35, 423–444. [Google Scholar] [CrossRef] [PubMed]
- Zhu, R.; Zhang, J.; Lyu, Y.; Chen, Y.; Han, S.; Wang, H. Efficacy of substances containing 3 types of active ingredientssaponins, flavones, and alkaloids in regulation of cytokines in autoimmune diseases a systematic review and Meta-analysis based on animal studies. Journal of Traditional Chinese Medicine 2024, 44, 417–426. [Google Scholar] [CrossRef]
- Kusuma, G.D.; Paseephol, T.; Sherkat, F. Prebiotic and rheological effects of Jerusalem artichoke inulin in low-fat yogurt. Australian Journal of Dairy Technology 2009, 64, 159–163. [Google Scholar]
- Yi, H.; Zhang, L.; Hua, C.; Sun, K.; Zhang, L. Extraction and enzymatic hydrolysis of inulin from Jerusalem artichoke and their effects on textural and sensorial characteristics of yogurt. Food Bioprocess Tech 2010, 3, 315–319. [Google Scholar] [CrossRef]
- Canbulat, Z.; Ozcan, T. Effects of short-chain and long-chain inulin on the quality of probiotic yogurt containing Lactobacillus Rhamnosus. Journal of Food Processing and Preservation 2015, 39, 1251–1260. [Google Scholar]
- Guven, M.; Yasar, K.; Karaca, O.B.; Hayaloglu, A.A. The effect of inulin as a fat replacer on the quality of set-type low-fat yogurt manufacture. International Journal of Dairy Technology 2005, 58, 180–184. [Google Scholar] [CrossRef]
- Ehsani, J.; Mohsenzadeh, M.; Khomeiri, M.; Ghasemnezhad, A. Chemical characteristics, and effect of inulin extracted from artichoke (Cynara scolymus L.) root on biochemical properties of synbiotic yogurt at the end of fermentation. Iranian Journal of Chemistry & Chemical Engineering-International English Edition 2018, 37, 219–230. [Google Scholar]
- Balcazar-Zumaeta, C.R.; Castro-Alayo, E.M.; Cayo-Colca, I.S.; Idrogo-Vasquez, G.; Munoz-Astecker, L.D. Metabolomics during the spontaneous fermentation in cocoa (Theobroma cacao L.): An exploraty review. Food Research International 2023, 163. [Google Scholar] [CrossRef] [PubMed]
- Luis Navarrete-Bolanos, J. Improving traditional fermented beverages: How to evolve from spontaneous to directed fermentation. Engineering in Life Sciences 2012, 12, 410–418. [Google Scholar] [CrossRef]
- Yao, Z.; Zhu, Y.; Wu, Q.; Xu, Y. Challenges and perspectives of quantitative microbiome profiling in food fermentations. Critical Reviews in Food Science and Nutrition 2024, 64, 4995–5015. [Google Scholar] [CrossRef] [PubMed]
- Callahan, B.J.; Wong, J.; Heiner, C.; Oh, S.; Theriot, C.M.; Gulati, A.S.; McGill, S.K.; Dougherty, M.K. High-throughput amplicon sequencing of the full-length 16S rRNA gene with single-nucleotide resolution. Nucleic Acids Research 2019, 47. [Google Scholar] [CrossRef] [PubMed]
- Qi, Y.; Huang, L.; Zeng, Y.; Li, W.; Zhou, D.; Xie, J.; Xie, J.; Tu, Q.; Deng, D.; Yin, J. Pediococcus pentosaceus: screening and application as probiotics in food processing. Front Microbiol 2021, 12, 762467. [Google Scholar] [CrossRef] [PubMed]
- Peter, S.B.; Qiao, Z.; Godspower, H.N.; Ajeje, S.B.; Xu, M.; Zhang, X.; Yang, T.; Rao, Z. Biotechnological innovations and therapeutic application of Pediococcus and lactic acid bacteria: the next-generation microorganism. Frontiers in Bioengineering and Biotechnology 2022, 9, 802031. [Google Scholar] [CrossRef] [PubMed]
- Medina, E.; Pérez-Díaz, I.M.; Breidt, F.; Hayes, J.; Franco, W.; Butz, N.; Azcarate-Peril, M.A. Bacterial ecology of fermented cucumber rising pH spoilage as determined by nonculture-based methods. Journal of food science 2016, 81, 121–129. [Google Scholar] [CrossRef] [PubMed]
- Sengun, I.Y.; Doyle, M.P. Microbiology of fermented foods. In Acetic Acid Bacteria; CRC Press: 2017, pp. 213-260.
- Paramithiotis, S. Microorganisms associated with food fermentation. In Bioactive Compounds in Fermented Foods; CRC Press: 2021, pp. 3-47.
- Zhao, H.; Li, Y.; Liu, L.; Zheng, M.; Feng, Z.; Hu, K.; Tao, Y. Effects of inoculation timing and mixed fermentation with Pichia fermentans on Oenococcus oeni viability, fermentation duration and aroma production during wine malolactic fermentation. Food Research International 2022, 159, 111604. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Nguyen, T.T.H.; Jin, J.; Lim, J.; Lee, J.; Piao, M.; Mok, I.-K.; Kim, D. Brewing of glucuronic acid-enriched apple cider with enhanced antioxidant activities through the co-fermentation of yeast (Saccharomyces cerevisiae and Pichia kudriavzevii) and bacteria (Lactobacillus plantarum). Food Sci Biotechnol 2021, 30, 555–564. [Google Scholar] [CrossRef] [PubMed]
- Debeljak, P.; Baltar, F. Fungal diversity and community composition across ecosystems. Journal of Fungi 2023, 9, 510. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Tang, J.; Zhang, Z.; Wu, Z.; Zhong, A.; Li, Z.; Wang, Y. Correlation between flavor compounds and microorganisms of Chaling natural fermented red sufu. Lwt 2022, 154, 112873. [Google Scholar] [CrossRef]
- Wang, X.; Schlatter, D.C.; Glawe, D.A.; Edwards, C.G.; Weller, D.M.; Paulitz, T.C.; Abatzoglou, J.T.; Okubara, P.A. Native yeast and non-yeast fungal communities of Cabernet Sauvignon berries from two Washington State vineyards, and persistence in spontaneous fermentation. International Journal of Food Microbiology 2021, 350, 109225. [Google Scholar] [CrossRef] [PubMed]
- Cao, H.; Chen, B.H.; Inbaraj, B.S.; Chen, L.; Alvarez-Rivera, G.; Cifuentes, A.; Zhang, N.; Yang, D.J.; Simal-Gandara, J.; Wang, M. Preventive potential and mechanism of dietary polyphenols on the formation of heterocyclic aromatic amines. Food Frontiers 2020, 1, 134–151. [Google Scholar] [CrossRef]
- Beygmoradi, A.; Homaei, A. Marine microbes as a valuable resource for brand new industrial biocatalysts. Biocatal Agr Biotech 2017, 11, 131–152. [Google Scholar] [CrossRef]
- Hu, Y.; Zhang, L.; Wen, R.; Chen, Q.; Kong, B. Role of lactic acid bacteria in flavor development in traditional Chinese fermented foods: A review. Critical reviews in food science and nutrition 2022, 62, 2741–2755. [Google Scholar] [CrossRef] [PubMed]
- Song, G.; He, Z.; Wang, X.; Zhao, M.; Cao, X.; Lin, X.; Ji, C.; Zhang, S.; Liang, H. Improving the quality of Suancai by inoculating with Lactobacillus plantarum and Pediococcus pentosaceus. Food Research International 2021, 148, 110581. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, H.; Hassane, A.; Atta, O.; Song, Y. Deep learning strategies for active secondary metabolites biosynthesis from fungi: Harnessing artificial manipulation and application. Biocatal Agr Biotech 2021, 38, 102195. [Google Scholar] [CrossRef]
- Grewal, J. Gamma-aminobutyric acid (GABA): A versatile bioactive compound. Eur. J. Mol. Clin. Med 2020, 7, 3068–3075. [Google Scholar]
- Tang, M.; Wang, Z.; Luo, J.; Zhu, T.; Song, F.; Chen, H. Preparation, chemical profiles, antioxidative activities, and angiotensin-converting enzyme 2 inhibitory effect of date fruit vinegar. Journal of Food Science 2024, 89, 684–700. [Google Scholar] [CrossRef] [PubMed]
- Keșa, A. L.; Pop, C.R.; Mudura, E.; Salanță, L.C.; Pasqualone, A.; Dărab, C.; Burja-Udrea, C.; Zhao, H.; Coldea, T.E. Strategies to improve the potential functionality of fruit-based fermented beverages. Plants 2021, 10, 2263. [Google Scholar] [CrossRef]
Figure 1.
Composition and diversity of bacterial communities during spontaneous fermentation. α-diversity index analysis of bacteria (A). Heatmap of genus-level species composition for co-clustering (B). Species diversity and abundance of bacteria at the phylum and genus levels (C).
Figure 1.
Composition and diversity of bacterial communities during spontaneous fermentation. α-diversity index analysis of bacteria (A). Heatmap of genus-level species composition for co-clustering (B). Species diversity and abundance of bacteria at the phylum and genus levels (C).
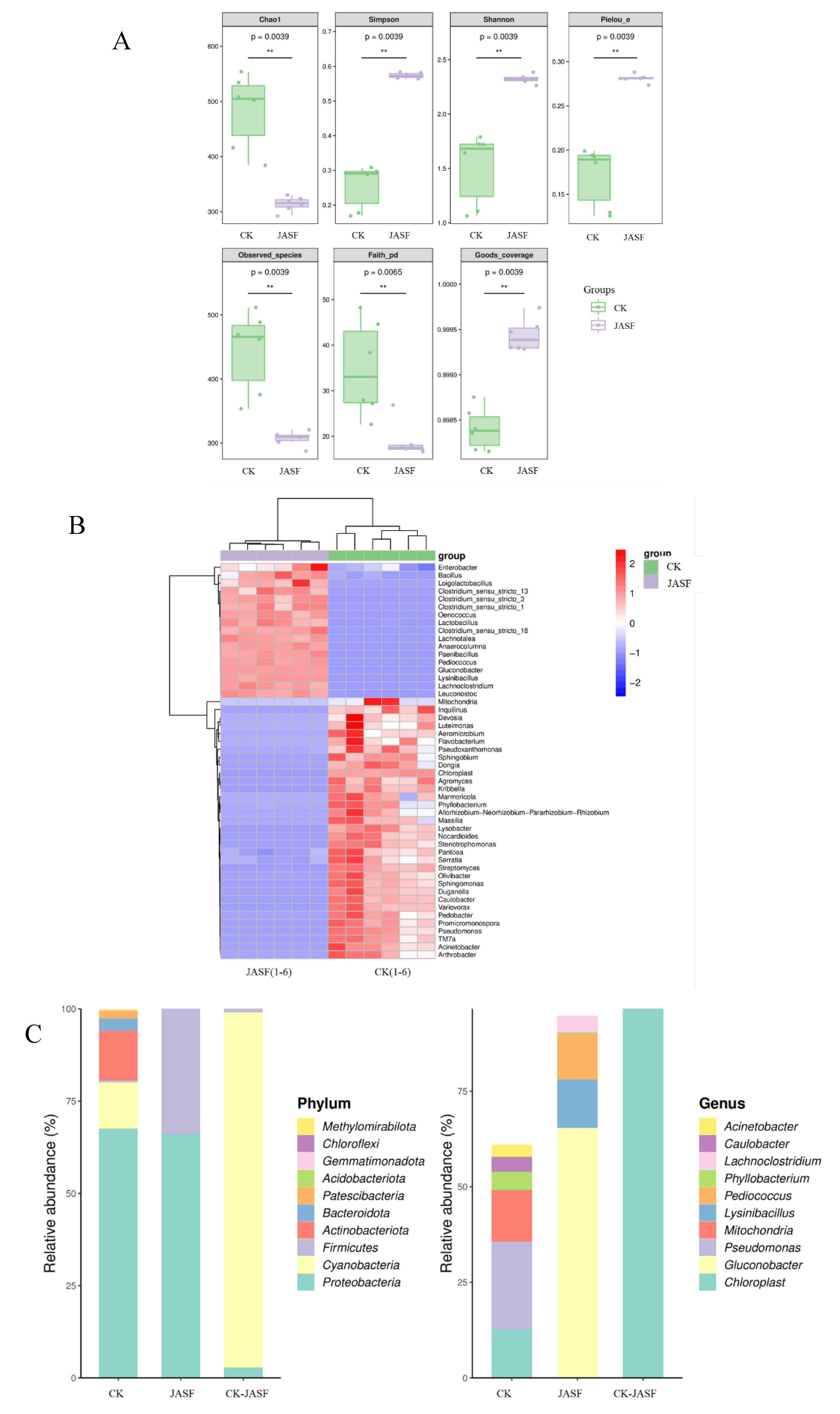
Figure 2.
Composition and diversity of fungal communities during spontaneous fermentation. α-diversity index analysis of fungi (A). Heatmap of genus-level species composition for co-clustering (B). Species diversity and abundance of fungi at the phylum and genus levels (C).
Figure 2.
Composition and diversity of fungal communities during spontaneous fermentation. α-diversity index analysis of fungi (A). Heatmap of genus-level species composition for co-clustering (B). Species diversity and abundance of fungi at the phylum and genus levels (C).
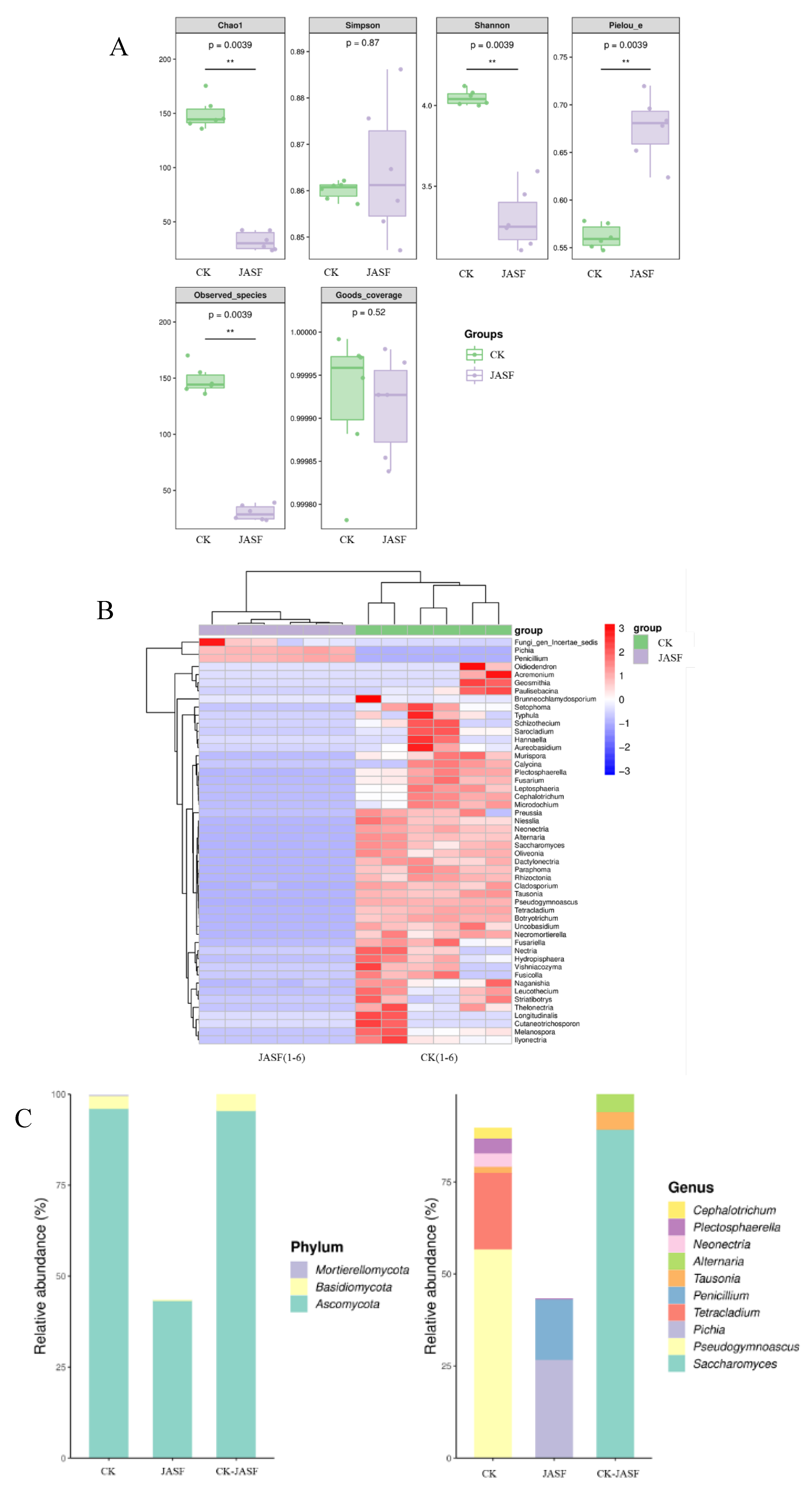
Figure 3.
Dynamic changes of secondary metabolites during fermentation. Volcano plot (A). The OPLS-DA S-plot (B). Heatmap of the differential secondary metabolites (C). Enrichment of the differential secondary metabolites (D).
Figure 3.
Dynamic changes of secondary metabolites during fermentation. Volcano plot (A). The OPLS-DA S-plot (B). Heatmap of the differential secondary metabolites (C). Enrichment of the differential secondary metabolites (D).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



