
Submitted:
24 July 2024
Posted:
25 July 2024
You are already at the latest version
Abstract
Background and aim: Apigenin (API), a dietary flavone subclass of flavonoid found in herbs, fruits, and vegetables, exhibits various pharmacological effects. However, its role in triggering oxidative stress- induced apoptosis in uterine cells remains unclear. This study aimed to evaluate the oxidative stress-mediated apoptosis induced by apigenin in rat uterine leiomyoma ELT3 cells. Material and methods: ELT3 cells were treated with 50 and 100 µM API for 48 hours. We assessed oxidative stress markers (reactive oxygen species, malondialdehyde), antioxidant defense (glutathione reductase activity), cell proliferation, and apoptotic gene expression (p53, Bax, Bcl-2, caspase-3, caspase-9) using relevant assays and real-time PCR. Results: API treatment decreased cell proliferation and glutathione reductase activity in ELT3 cells. This coincided with increased levels of reactive oxygen species, malondialdehyde, and lactate dehydrogenase activity, indicating oxidative stress and cell damage. Furthermore, apigenin upregulated pro-apoptotic genes (p53, Bax, caspase-3, caspase-9) and downregulated the anti-apoptotic gene Bcl-2, suggesting activation of the intrinsic apoptotic pathway. Conclusion: API induces apoptosis in rat uterine leiomyoma cells through a mechanism involving oxidative stress-mediated activation of the intrinsic apoptotic pathway.
Keywords:
apigenin
; apoptosis
; ELT3 cells
; oxidative stress
; reactive oxygen species
1. Introduction
Apigenin (4′, 5, 7, -trihydroxyflavone; API) is a bioactive flavone subclass of flavonoids. It is found in a wide range of fruits, vegetables and herbs including oranges, grapefruits, celery, parsley, onions, green tea, wheat sprouts, chamomile, spearmint, and thyme [1,2,3,4,5,6,7,8,9,10]. This compound exhibits a diverse array of biological activities such as anti-inflammatory [6,11,12,13,14,15,16,17,18], antioxidant [5,6,14,16,18], antimicrobial [11,12,13,15,17], anticancer [6,11,12,13,14,15,17,19], hepatoprotective [20], renoprotective [5], and neuroprotective effects [21]. Additionally, API has been noted for its chemopreventive properties and its ability to inhibit tumor growth [6,19]. Phytosteroids are a class of specialized metabolites, derived from plants that can bind to steroid receptors in animals. API has been identified as a phytosteroids specially phytoprogestin [22,23,24]. Its biological effects have been demonstrated in progesterone-sensitive tissues such as the uterus and breast cancer [25]. Oxidative stress and apoptosis play key roles in various physiological and pathological processes. These processes can be triggered by numerous factors, including physical elements, chemical treatments, and toxic substances [26,27]. Leiomyomas account for 90% of all canine tumors, with the canine uterus being the most common site. These tumors are often difficult to distinguish from other malignant tumors [28]. Traditional antineoplastic therapies often provoke side effects, which require additional pharmacological interventions, leading to a cascade of drug-related side effects [29]. Therefore, alternative herbal remedies are a promising approach to reduce these side effects and provide effective treatments. This study employs rat uterine leiomyoma ELT3 cells (ELT3 cells) as an in vitro model to investigate the oxidative stress and apoptosis induced by apigenin. This research aims to explore the potential of apigenin as a therapeutic agent for treating uterine leiomyomas while minimizing adverse effects associated with conventional therapies.
2. Materials and Methods
2.1. Ethical Approval
No experiments were conducted on live animals, so ethical approval was unnecessary.
2.2. Study Period and Location
The study was conducted from March to December 2022 at the Department of Veterinary Technology, Kasetsart University, Thailand.
2.3. Cells and Chemicals
ELT3 cells were obtained from the American Type Culture Collection (Manassas, VA, USA). The MTS cell proliferation assay kit, Lactate Dehydrogenase (LDH)-cytotoxicity assay kit II, 2′7′-dichlorofluorescin diacetate (DCFDA) cellular Reactive Oxygen Species (ROS) detection assay kit, lipid peroxidation (MDA) assay kit and Glutathione reductase (GR) assay kit were purchased from Abcam. API and dimethyl sulfoxide (DMSO) were purchased from Sigma-Aldrich. A 5 mM stock solution of API was prepared in DMSO and further diluted to final concentrations of 20-160 µM using serum-free culture medium. All treatments ensured the amount of DMSO added to the cell culture was less than 0.1%.
2.4. Cell Culture and Assays
ELT3 cells were cultured in complete Dulbecco’s Modified Eagle Medium/Nutrient Mixture F-12 (DMEM/F-12) supplemented with 5% horse serum, 2.5% fetal bovine serum (FBS), 100 U/mL penicillin, and 100 U/mL streptomycin. The culture was maintained in a humidified incubator with 5% CO2 at 37°C. For the assay, the culture was seeded in 96-well plates at density of 5-10 × 103 cells/ well in a final volume of 200 µL/well. Cell proliferation and cytotoxicity were assessed using MTS assay and LDH leakage assay. Intracellular ROS production, lipid peroxidation (MDA levels), and glutathione reductase (GR) activity were measured following the methods described by Sathaporn et al. [30].
2.5. Determination of Apoptotic-Related Gene Expression
To evaluate the mRNA expression levels of genes associated with apoptosis. The mRNA expression levels were determined using quantitative RT-PCR (qRT-PCR). ELT3 cells were treated with 100 µM API for 48 h. Total RNA was extracted using TRIZOL® reagent (Invitrogen, USA) following the manufacturer’s instructions. Subsequently, the cDNA was synthesized from 2 µg of total RNA in 20 µL reaction mixture using the high-capacity cDNA Reverse Transcription kit (Applied Biosystems, CA) according to the manufacturer’s protocol. qRT-PCR was performed to quantify the mRNA expression of target genes: Tp53, Bax, Bcl2, caspase-3 and caspase-9. GAPDH was used as a housekeeping reference gene for normalization. The specific primer sequences used for qRT-PCR are listed in Table 1. Each reaction contained 50 ng of cDNA, 0.25 µM of each forward and reverse primer, and Luna® Universal qPCR Master mix (New England Biolabs) in a final volume of 10 µL. The thermal cycling conditions included an initial denaturation step at 95°C for 1 minute, followed by 44 cycles of denaturation at 95°C for 15 seconds and annealing/extension at 60°C for 30 seconds. Cycle threshold (Ct) values were normalized to GAPDH expression using the 2-ΔΔCt method for relative quantification of mRNA levels.
2.6. Statistical Analysis
All experiments were performed at least three times to ensure reproducibility. Data were analyzed using GraphPad Prism 5 software (GraphPad, San Diego, CA). For comparisons between two groups, a paired t-test was employed. All data are expressed as the mean ± S.D. Statistical significance was set at p < 0.05.
3. Results
3.1. API Decreased the Viability of ELT3 Cells
The morphological changes are observed in ELT3 cells following API treatment in Figure 1. Cell viability percentage of ELT3 cells was assessed using an MTS assay in response to API are presented in Figure 2. The half-maximal inhibitory concentration (IC50) values were 104.85, 63.74 and 60.25 µM at 24, 48 and 72 h of incubation, respectively. These findings indicate that API exhibits increasing potency against ELT3 cells with longer incubation times. According to the previous study, we used the API concentrations of 50 and 100 µM for 24, 48 and 72 h to test TM4 mouse Sertoli cells which is the cells in reproductive system [30], moreover the researchers reported the API concentration of 0-100 µM for 24-114 h to test human hepatoma cells and human leukemia cell line [31,32]. So, the treatment of 48 h was selected as the optimal time and the optimal concentration of API of 50 and 100 µM was used in further experiments.
3.2. API Increased LDH Activity of ELT3 Cells
LDH leakage of ELT3 cells in response to API is shown in Figure 3. API treatment significantly increased LDH activity at concentration 50 and 100 µM in ELT3 cells compared at when compared to both the control group and the DMSO-treated group. This suggests that API exposure leads to a dose-dependent increase in cell death or membrane damage in ELT3 cells.
3.3. API Increased ROS Production of ELT3 Cells
The ROS production of ELT3 cells in response to API is shown in Figure 4. API treatment significantly increased ROS levels in ELT3 cells at 50 and 100 µM when compared to the control group. This finding suggests that API may induce oxidative stress in these cells.
3.4. API Increased MDA Levels of ELT3 Cells
The MDA levels of ELT3 cells in response to API are shown in Figure 5. API treatment significantly increased MDA levels in ELT3 cells at 50 and 100 µM when compared to the control group. This suggests that API treatment promotes lipid peroxidation and contributes to oxidative stress in ELT3 cells.
3.5. API Decreased GR Activity of ELT3 Cells
The GR activity of ELT3 cells in response to API is shown in Figure 6. API treatment significantly decreased GR activity in ELT3 cells at 50 and 100 µM when compared to the control and DMSO treatment. This suggests that API may impair the antioxidant defense mechanisms of ELT3 cells, potentially contributing to the observed increase in oxidative stress.
3.6. API Induced Intrinsic Apoptotic Pathway
API treatment induces apoptosis (programmed cell death) in ELT3 cells through a mechanism involving oxidative stress. As shown in Figure 7, API treatment with concentration at 50 and 100 µM significantly upregulated the expression of pro-apoptotic genes Tp53, Bax, caspase-9, and caspase-3 compared to the control group. Conversely, API treatment significantly downregulated the expression of the anti-apoptotic gene Bcl-2. These alterations in gene expression are indicative of the activation of the intrinsic apoptotic pathway, a well-established cell death mechanism regulated by proteins like p53 (Tp53), Bax, and Bcl-2 [33]. Caspase-3 acts as a key executioner caspase in apoptosis [34], and its upregulation further supports API-induced cell death in ELT3 cells.
4. Discussion
The present study demonstrates the oxidative stress, especially apoptosis induced by API on ELT3 cells. ELT3 cells are a well-established in vitro model system for investigating uterine leiomyomas. These cells, a uterine epithelial-like cell, are derived from the uterine epithelium of Eker rats and exhibit responsiveness to estrogen-like agents [35]. Importantly, ELT3 cells share characteristics with tumor-derived cells, making them a valuable tool for preclinical studies aimed at identifying novel therapeutic agents for uterine leiomyomas [36]. By employing this model, we can explore the potential effects of API on these tumors in a controlled laboratory setting.
Cytotoxicity assays are essential tools for indicating the ability of a compound to induce cell death [37]. In this study, we employed two complementary assays: the MTS assay and the LDH leakage assay. The MTS assay offers a convenient method to evaluate cell viability through a single step directly in the culture medium, eliminating the need for multiple steps required in the traditional MTT assay. The permeabilization of plasma membrane is a key point for necrotic cells. This event can be quantified in tissue culture by measuring the release of the LDH [38]. We observed that API exhibited cytotoxicity in ELT3 cells in a dose- and time-dependent manner, indicating a decrease in cell viability with increasing API concentration and exposure time. This finding aligns with previous research demonstrating API’s ability to reduce cell proliferation in various human and animal cell lines, including hepatocellular carcinoma cells (Hep3B, HepG2), TM4 Sertoli cells, leukemia cells, cholangiocarcinoma cells and breast carcinoma cells [16,30,31,32,39,40]. Additionally, the LDH leakage assay confirmed API’s cytotoxic effect by showing a significant increase in LDH release from ELT3 cells, a marker of cell membrane damage or death.
Oxidative stress arises from an imbalance between production and accumulation of ROS within cells. Increased ROS generation and lipid peroxidation are indicators of oxidative stress [41]. In this study demonstrate that API treatment significantly elevates ROS levels in ELT3 cells. This finding aligned with many studies that described API-induced ROS levels in different human and animal cells, including TM4 Sertoli cells, human hepatocarcinoma HepG2 cells, human leukemia cells, human breast carcinoma cells and human hepatocarcinoma Hep3B and HepG2 cells [30,31,32,40,42]. Furthermore, API significantly increased the levels of MDA in ELT3 cells. MDA is a byproduct of lipid peroxidation, a process triggered by excessive ROS. This indicates that API treatment promotes lipid peroxidation, further contributing to oxidative stress in these cells [43].
GR enzyme is a key antioxidant enzyme that helps maintain cellular redox balance. Several studies suggest that the interplay between ROS production and antioxidant systems plays a crucial role in cell death [44]. Notably, our data presented a significant decrease in GR activity in API-treated ELT3 cells. This reduced GR activity suggests that API may impair the antioxidant defense mechanisms of ELT3 cells, potentially exacerbating the observed oxidative stress.
Programmed cell death (PCD) encompasses various mechanisms with generally characterized by distinct morphological characteristics and energy-dependent biochemical features [45]. While this study did not explore all PCD pathways, our findings strongly suggest that API triggers apoptosis in ELT3 cells. Apoptosis is a controlled cell death process crucial for maintaining tissue homeostasis. Ferroptosis is a new type of PCD, which occurs with iron dependence [46]. Necroptosis is an alternative mode of regulated cell death mimic of apoptosis and necrosis [47]. Autophagy is the recycle of cellular components to maintain metabolism under stress conditions [48]. Previous studies support the anticarcinogenic properties of API through its ability to modulate cellular responses to oxidative stress and induce both autophagy and apoptosis [49]. Notably, studies have demonstrated API’s capacity to induce apoptosis and autophagy in HCT116 colon cancer cells [50].
Apoptosis, a PCD, plays a vital role to eliminate unwanted cells and maintaining tissue homeostasis in biological systems. It is regulated by two main pathways; the extrinsic pathway (death receptor-mediated pathway) which requires the cell surface receptor and the intrinsic pathway which is mediated by mitochondria (mitochondrial-mediated pathway) [51,52]. Our findings suggest that API likely triggers apoptosis in ELT3 cells via the intrinsic pathway. This is supported by the observed upregulation of the Tp53. p53, a well-known tumor suppressor protein, can promote apoptosis through various mechanisms, including both transcriptional and non-transcriptional activities [53]. Further investigation into the specific downstream targets of p53 activation in API-treated ELT3 cells would be valuable in fully elucidating the intrinsic apoptotic pathway involved. The extrinsic and intrinsic pathway involve some transcriptional activity [54]. Apoptosis assays can be classified into six major groups [45]. There are various methods to assess apoptosis. A common approach involves measuring caspase activity, a family of enzymes crucial for executing the cell death program [45]. Additionally, real-time PCR allows researchers to analyze gene expression changes. This includes evaluating pro-apoptotic genes like Bax and anti-apoptotic genes like Bcl-2, which provide insights into the apoptotic signaling cascade [55]. Our findings demonstrate that API treatment upregulates the expression of the pro-apoptotic gene p53 (Tp53). This, along with the previously observed decrease in the anti-apoptotic gene Bcl-2 expression, suggests that API disrupts the balance between pro-apoptotic and anti-apoptotic factors, potentially leading to apoptosis in ELT3 cells via the intrinsic pathway mediated by p53. In this study, Tp53 promotes API-induced ROS generation, further activates Bax gene expression, and inhibit Bcl2 gene expression. Mitochondria are the major targets of excessive ROS and excess ROS induce cell death by promoting the intrinsic apoptotic pathway [56,57]. The disruption of mitochondria facilitates the activation of caspase9/3 cascade and induction of apoptosis [58,59]. API-treated cells show upregulation of caspase9/3 gene expression confirming the involvement of intrinsic apoptotic pathway. Previous studies reported the API induces apoptosis in various tumor cell lines through activation of mitochondria-mediated pathway via intracellular generation of ROS as an activator [60,61,62,63] whereas Sanjay, 2001 reported that API was safe for normal versus human prostate carcinoma cells [64]. Our findings suggest its potential to induce apoptosis in ELT3 cells. This difference might be due to variations in cell type, treatment concentration, or other experimental conditions. Further investigation is needed to fully understand the multifaceted effects of API across different cell lines and tissues.
5. Conclusions
This study demonstrates that API exposure triggers oxidative stress and induces apoptosis in ELT3 cells, a uterine epithelial cell line, via the intrinsic pathway. These findings are characterized by decreased cell viability, increased LDH release, elevated ROS and MDA levels, and altered gene expression patterns. Specifically, API treatment upregulated pro-apoptotic genes (Tp53, Bax, caspase-9, caspase-3) and downregulated the anti-apoptotic gene Bcl-2. These observations suggest that API may have potential as a therapeutic agent for uterine tumors. However, further in vivo studies are necessary to confirm the efficacy and safety of API in this context. Additionally, exploring the underlying mechanisms by which API modulates p53 and the apoptotic pathway could provide valuable insights for future drug development efforts.
Author Contributions
Conceptualization, S.S.; methodology, S.S., N.M. and Ch.S.; Validation, S.S., N.M., Ch.S., C.S. and K.O.; Writing-original draft, S.S; Writing-review and editing, S.S., N.M., Ch.S., C.S. and K.O. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by Disease-Free Animals and People Safe from Rabies project According to the Professor’s wishes Her Royal Highness Princess Chulabhorn Walailak Archarajkumari Krom Phra Sri Swangwattana Worakattiyarajnari funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The authors confirm that the data supporting the findings of this study are available within the article.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Bak, M.J.; Gupta, S.D.; Wahler, J. Role of dietary bioactive natural products in estrogen receptor-positive breast cancer. Semin. Cancer Biol, 2016; 40–41. [Google Scholar] [CrossRef]
- Duthie, G.; Crozier, A. Plant-derived phenolic antioxidants. Curr. Opin. Clin. Nutr. Metab. Care., 2000, 3, 447–451. [Google Scholar] [CrossRef] [PubMed]
- Manach, C.; Scallbert, A.; Morand, C. Polyphenols: Food sources and bioavaiability. Am. J. Clin. Nutr., 2004, 79, 727–747. [Google Scholar] [CrossRef] [PubMed]
- Peterson, J. , Dwyer, J. Flavonoids: dietary occurrence and biochemical activity. Nutr. Res., 1998, 18, 1995–2018. [Google Scholar] [CrossRef]
- Sharma, H. , Kanwal, R., Bhaskaran, N. Plant flavone apigenin binds to nucleic acid bases and reduces oxidative DNA damage in prostate epithelial cells. PloS One., 2014, 9, e91588. [Google Scholar] [CrossRef]
- Shukla, S. , Gupta, S. Apigenin: a promising molecule for cancer prevention. Pharm. Res., 2010, 27, 962–978. [Google Scholar] [CrossRef] [PubMed]
- Tsanova-Sanova, S. , Ribarova, F. Flavonoids and flavones in some Bulgarian plant foods. Pol. J. Food Nut. Sci, 2013; 63, 173–177. [Google Scholar] [CrossRef]
- Zhang, Q. , Zhou, M.M., Chen, P.L. Optimization of ultrasonic assisted enzymatic hydrolysis for the extraction of luteolin and apigenin from celery. J. Food Sci., 2011, 76, C680–C685. [Google Scholar] [CrossRef]
- Zhou, Y. , Zheng, J., Li, Y. Natural polyphenols for prevention and treatment of cancer. Nutrients., 2016, 8, 515. [Google Scholar] [CrossRef]
- Zhu, Y. , Wu, J., Li, S. Apigenin inhibits migration and invasion via modulation of epithelial mesenchymal transition in prostate cancer. Mol. Med. Rep., 2015, 11, 1004–1008. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, K. , Mandal, M. Oxidative stress triggered by naturally occurring flavone apigenin results in senescence and chemotherapeutic effect in human colirectal cancer cells. Redox Biol., 2015, 5, 153–162. [Google Scholar] [CrossRef]
- Bao, Y.Y. , Zhou, S.H., Fan, J. Anticancer mechanism of apigenin and the implications of GULT-1 expression in head and neck cancers. Future Oncol., 2013, 9, 1353–1364. [Google Scholar] [CrossRef]
- Bian, M. , Zhang, Y., Du, X. Apigenin-7-diglucuronide protects retinas against bright light-induced photoreceptor degeneration through inhibition of retinal oxidative stress and inflammation. Brain Res., 2017, 1663, 141–150. [Google Scholar] [CrossRef]
- Birt, D.F. , Walker, B., Tibbles, M.G. Anti-mutagenesis and anti-promotion by apigenin, robinetin and indole-3-carbinol. Carcinogenesis., 1986, 7, 959–963. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.Y. , Liang, J.Y., Guo, X.J. 5-fluorouracil combined with apigenin enhances anticancer activity through mitochondrial membrane potential (∆Ѱm)-mediated apoptosis in hepatocellular carcinoma. Clin. Exp. Pharmacol. Physiol., 2014, 42, 146–153. [Google Scholar] [CrossRef] [PubMed]
- Kang, O.H. , Lee, J.H., Kwon, D.Y. Apigenin inhibits release of inflammatory mediators by blocking the NF-kappa B activation pathways in the HMC-1 cells. Immunopharmacol. Immunotoxicol., 2011, 33, 473–479. [Google Scholar] [CrossRef] [PubMed]
- Kaur, P. , Shukla, S., Gupta, S. Plant flavonoid apigenin inactivates AKt to trigger apoptosis in human prostate cancer: an in vitro and in vivo study. Carcinogenesis., 2008, 29, 2210–2217. [Google Scholar] [CrossRef] [PubMed]
- Saeed, M. , Kadioglu, O. , Khalid, H. Activity of dietary flavonoid, apigenin, against multidrug-resistant tumor cell as determined by pharmacogenomics and molecular docking. J. Nutr. Biochem., 2015, 26, 44–59. [Google Scholar] [CrossRef] [PubMed]
- Xu, X. , Li, M., Chen, W. Apigenin attenuates oxidative injury in ARPE-19 cells through activation of Nrf2 pathway. Oxid. Med. Cell Longev, 2016, 4378461. [CrossRef]
- Yang, J. , Wang, X. Y., Xue, J. Protective effect of apigenin on mouse acute liver injury induced by acetaminophen is associated with increment of hepatic glutathione reductase activity. Food Funct., 2013, 4, 939–943. [Google Scholar] [CrossRef]
- Wang, C.N. , Chi, C.W., Lin, Y.L. The neuroprotective effects of phytoeatrogen on amyloid beta protein-induced toxicity are mediated by abrogating the activation of caspase cascade in rat cortical neurons. J. Biol. Chem., 2001, 276, 5287–5295. [Google Scholar] [CrossRef]
- Rosenberg, R.S. , Grass, L., Jenkins, D.J. Modulation of androgen and progesterone receptors by phytochemicals in breast cancer cell lines. Biochem. Biophys. Res. Commun., 1998, 248, 935–939. [Google Scholar] [CrossRef]
- Stroheker, T. , Picard, K., Lhuguenot, J.C. Steroid activities comparison of natural and food wrap compounds in human breast cancer cell lines. Food Chem. Toxicol., 2004, 42, 887–897. [Google Scholar] [CrossRef]
- Toh, M.F. , Mendonca, E., Eddie, S.L. Kaempferol exhibits progestogenic effects in ovariectomized rats. J. Steroids Horm. Sci., 2014, 5, 136. [Google Scholar] [CrossRef] [PubMed]
- Dean, M. , Murphy, B.T., Burdette, J.E. Phytosteroids beyond estrogens: regulators of reproductive and endocrine function in natural products. Mol. Cell Endocrinol., 2017, 442, 98–105. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.H. , Chan, W.H. Protective effects of liquiritienin against citrinin-triggered, oxidative-stress-mediated apoptosis and disruption of embryonic development in mouse blastocyst. Int. J. Mol. Sci., 2017, 18. [CrossRef]
- Liu, F. , Du, K.J., Fang, Z. Chemical and biological insights into uranium-induced apoptosis of rat hepatic cell line. Radiat. Environ. Biophys., 2015, 54, 207–216. [Google Scholar] [CrossRef] [PubMed]
- Corey, F.S. , Lawrence, J.A. Tumors of the female reproductive system. Withrow and MacEwen’s Small Animal Clinical Oncology 5th Ed, 2013, 532-537.
- Nicole, L.S. , Susan, S.W. Antineoplastic therapy side effects and polypharmacy in older adults with cancer. Top Geriatr. Rehabil, 2019; 35, 15–30. [Google Scholar] [CrossRef]
- Jittapalapong, S.; Poompoung, T.; Sutjarit, S. Apigenin induces oxidative stress in mouse Sertoli TM4 cells. Vet World 2021, 14, 3132–3137. [Google Scholar] [CrossRef] [PubMed]
- Valdameri, G., Lima, M.T., Worfel, P.R. Involvement of catalase in the apoptotic mechanism induced by apigenin in HepG2 human hepatoma cells. Chem. Biol. Interact., 2011, 193(2), 180-189. [CrossRef]
- Jayasooriya, R.G.P.T. , Kang, S.H., Kang, C.H. Apigenin decreases cell viability and telomerase activity in human leukemia cell lines. Food Chem. Toxicol., 2012, 50, 2605–2611. [Google Scholar] [CrossRef] [PubMed]
- Mohammed, A.R. , Shawkey, A.E., Rabeh, A.O. Expression of P53, BAX, and BCL-2 in human malignant melanoma and squamous cell carcinoma cells after tea tree oil treatment in vitro. Cytotech., 2019, 71, 461–473. [Google Scholar] [CrossRef]
- Huang, K.H. , Fang, W.L., Li, A.F.Y., Liang, P.H., Wu, C.W., Shyr, Y.M., Yang, M.H. Caspase-3, a key apoptotic protein, as a prognostic marker in gastric cancer after curative surgery. In. J. Surg., 2018, 52, 258–263. [Google Scholar] [CrossRef]
- Yu, J.J., Robb, V.A., Morrison, T.A., Ariazi, E.A., Karbowniczek, M., Astrinidis, A., Wang, C., Hernadez-Cuebas, L., Seeholzer, L.F., Nicolas, E., Hensley, H., Jordan, V.C., Walker, C.L., Henske, E.P. Estrogen promotes the survival and pulmonary metastasis of tuberin-null cells. Proc. Natl. Acad. Sci., 2009, 106(8), 2635–2640. [CrossRef]
- Jeffrey, I.L. Uterine Leiomyoma in the Eker Rat: A Unique Model for Important Diseases of Women. Genes Chromosomes. Cancer., 2003, 38(4), 349-56. [CrossRef]
- Weyermann, J. , Lochmann, D., Zimmer, A. A practical note on the use of cytotoxicity assays. Int. J. Pharm., 2005, 288, 369–376. [Google Scholar] [CrossRef]
- Francis, K.M.C. , Kenta, M., Maria, J.D.R. Detection of Necrosis by Release of Lactate Dehydrogenase (LDH) Activity. Methods Mol. Biol., 2013, 979, 65–70. [Google Scholar] [CrossRef]
- Subhasitanont, P. , Chokchaichamnankit, D., Chiablaem, K. Apigenin inhibits growth and induces apoptosis in human cholangiocarcinoma cells. Oncol. Lett., 2017, 14, 4361–4371. [Google Scholar] [CrossRef]
- Harrison, M.E. , Coombs, M.R.P., Delaney, L.M. Exposure of breast cancer cells to a subcytotoxic dose of apigenin causes growth inhibition, oxidative stress, and hypophosphorylation of Akt. Exp. Mol. Pathol., 2014, 97, 211–217. [Google Scholar] [CrossRef] [PubMed]
- Xiaozhen, L. , Shaoping, N., Danfei, H. Mitogen-activated protein kinase and Akt pathways are involved in 4-n-nonyphenol induced apoptosis in mouse Sertoli ELT3 cells. Environ. Toxicol. Pharmacol., 2015, 39, 815–824. [Google Scholar] [CrossRef]
- Kang, C.H. , Molagoda, I.M.N., Choi, Y.H. Apigenin promotes TRAIL-mediated apoptosis regardless of ROS generation. Food Chem. Toxicol., 2018, 11, 623–630. [Google Scholar] [CrossRef]
- Garwel, S.; Maria, W.; Elzbieta, N. Malondialdehyde (MDA) as a lipid peroxidation marker. Wiad Lek 2004, 57, 453–455. [Google Scholar]
- Anastasiya, V.S. Anastasiya, V.S., Anna, V.K., Olga, L.K., Maria, V.S., Nataliya, V.M., George, S.K., Alexy, A.D. ROS generation and antioxidant defense systems in normal and malignant cells. Oxid. Med. Cell Longev., 2019, 1-17. [CrossRef]
- Susan, E. Apoptosis: A Review of Programmed Cell Death. Toxico.l Pathol., 2007, 35(4), 495–516. [CrossRef]
- Li, J.; Cao, F.; Yin, H.L.; Huang, Z.J.; Lin, Z.T.; Mao, N.; Sun, B.; Wang, G. Ferroptosis: past, present and future. Cell Death Dis. 2020, 11, 1–13. [Google Scholar] [CrossRef]
- Dhuriya, Y.K.; Sharma, D. Necroptosis: a regulated inflammatory mode of cell death. J. Neuroinflammation 2018, 15, 1–9. [Google Scholar] [CrossRef]
- Xie, Z. , Klionsly, D.J. Autophagosome formation: core machinery and adaptations. Nat. Cell Biol., 2007, 9, 1102–1109. [Google Scholar] [CrossRef]
- Sung, B., Chung, H.Y., Kim, N.D. Role of apigenin in cancer prevention via the induction of apoptosis and autophagy. J. Cancer Prev., 2016, 21(4), 216-226. [CrossRef]
- Lee, Y. , Sung, B., Kang, Y.J., Kim, D.H., Jang, J.Y., Hwang, S.Y., Kim, M., Lim, H.S., Toon, J.H., Chung, H.Y., Kim, N.D. Apigenin-induced apoptosis is enhanced by inhibition of autophagy formation in HCT116 human colon cancer cells. Int. J. Oncol., 2014, 44, 1599–1606. [Google Scholar] [CrossRef] [PubMed]
- Fulda, S. , Debatin, K.M. Extrinsic versus intrinsic apoptosis pathways in anticancer chemotherapy. Oncogene., 2006, 25, 4798–4811. [Google Scholar] [CrossRef]
- Fulda, S. Molecular pathways: targeting death receptors and smacmimetric. Clin. Cancer Res., 2014, 20, 3915–3920. [Google Scholar] [CrossRef]
- Zhao, X. , Cong, X., Zheng, L. Dioscin, a natural steroid saponin, shows remarkable protective effect against acetaminophen-induced liver damage in vitro and in vivo. Toxicol. Lett., 2012, 214, 69–80. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, S., Pascal, D., Anne, M., Marlene, B., Valerie, D.C., Veronique, B., Evelyne, M. A comprehensive study of p53 transcriptional activity in thymus and spleen of γ irradiated mouse: High sensitivity of genes involved in the two main apoptotic pathways. Int. J. Radiat. Biol., 2006, 82(11), 761-770. [CrossRef]
- Lopez, J. , Tait, S.W.G. Mitochondrial apoptosis: killing cancer using the enemy within. Br. J. Canc., 2015, 112, 957–962. [Google Scholar] [CrossRef] [PubMed]
- Indo, H.P. , Davidson, M. , Yen, H.C. Evidence of ROS generation by mitochondria in cells with impaired electron transport chain and mitochondrial DNA damage. Mitochondrion., 2007, 7, 106–118. [Google Scholar] [CrossRef] [PubMed]
- Marchi, S. Marchi, S., Giorgim, C., Suski, J.M. Mitochondria-ros crosstalk in the control of cell death and aging. J. Signal. Transduct., 2012, 329635. [CrossRef]
- Anuradha, C.D. , Kanno, S., Hirano, S. Oxidative damage to mitochondria is a preliminary step to caspase-3 activation in fluoride-induced apoptosis in HL-60 cells. Free Radic. Biol. Med., 2001, 31, 367–373. [Google Scholar] [CrossRef]
- Chen, M. , Guerrero, A.D., Huang, L. Caspase-9-induced mitochondrial disruption through cleavage of anti-apoptotic BCL-2 family members. J. Biol. Chem., 2007, 282, 33888–33895. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.F. , Chie, Y.J., Yang, M.S. Apigenin induces apoptosis in human lung cancer H460 cells through caspase-and mitochondria-dependent pathways. Hum. Exp. Toxicol., 2011, 30, 1053–1061. [Google Scholar] [CrossRef] [PubMed]
- Patel, D.; Shukla, S.; Gupta, S. Apigenin and cancer chemoprevention: progress, potential and promise (review). Int. J. Oncol., 2007, 30, 233–245. [Google Scholar] [CrossRef] [PubMed]
- Ren, W. , Qiao, Z., Wang, H. Flavonoids: promising anticancer agents. Med. Res. Rev., 2003, 23, 519–534. [Google Scholar] [CrossRef] [PubMed]
- Torkin, R. , Lavoie, J.F., Kaplan, D.R. Induction of caspase-dependent, p53-mediated apoptosis by apigenin in human neuroblastoma. Mol. Cancer Ther., 2005, 4, 1–11. [Google Scholar] [CrossRef]
- Sanjay, G., Furrukh, A., Hasan, M. Selective Growth-Inhibitory, Cell-Cycle Deregulatory and Apoptotic Response of Apigenin in Normal versus Human Prostate Carcinoma Cells. Biochem. Biophys. Res. Commun., 2001, 287(4), 914-920. [CrossRef]
Figure 1.
Morphological changes of ELT3 cells after 48 h API treatment at 0-160 µM. Phase-contrast images were taken at 4x magnification. (A) 0 µM; (B) DMSO; (C) 20 µM; (D) 40 µM; (E) 60 µM; (F) 80 µM; (G) 100 µM; (H) 120 µM; (I) 140 µM; (J) 160 µM.
Figure 1.
Morphological changes of ELT3 cells after 48 h API treatment at 0-160 µM. Phase-contrast images were taken at 4x magnification. (A) 0 µM; (B) DMSO; (C) 20 µM; (D) 40 µM; (E) 60 µM; (F) 80 µM; (G) 100 µM; (H) 120 µM; (I) 140 µM; (J) 160 µM.

Figure 2.
The effect of API on cell viability of ELT3 cells. The results represent mean± S.D. *p < 0.05, ** p < 0.005 and *** p < 0.001 significantly different from no treatment. (A) 24h; (B); 48h; (C) 72h.
Figure 2.
The effect of API on cell viability of ELT3 cells. The results represent mean± S.D. *p < 0.05, ** p < 0.005 and *** p < 0.001 significantly different from no treatment. (A) 24h; (B); 48h; (C) 72h.
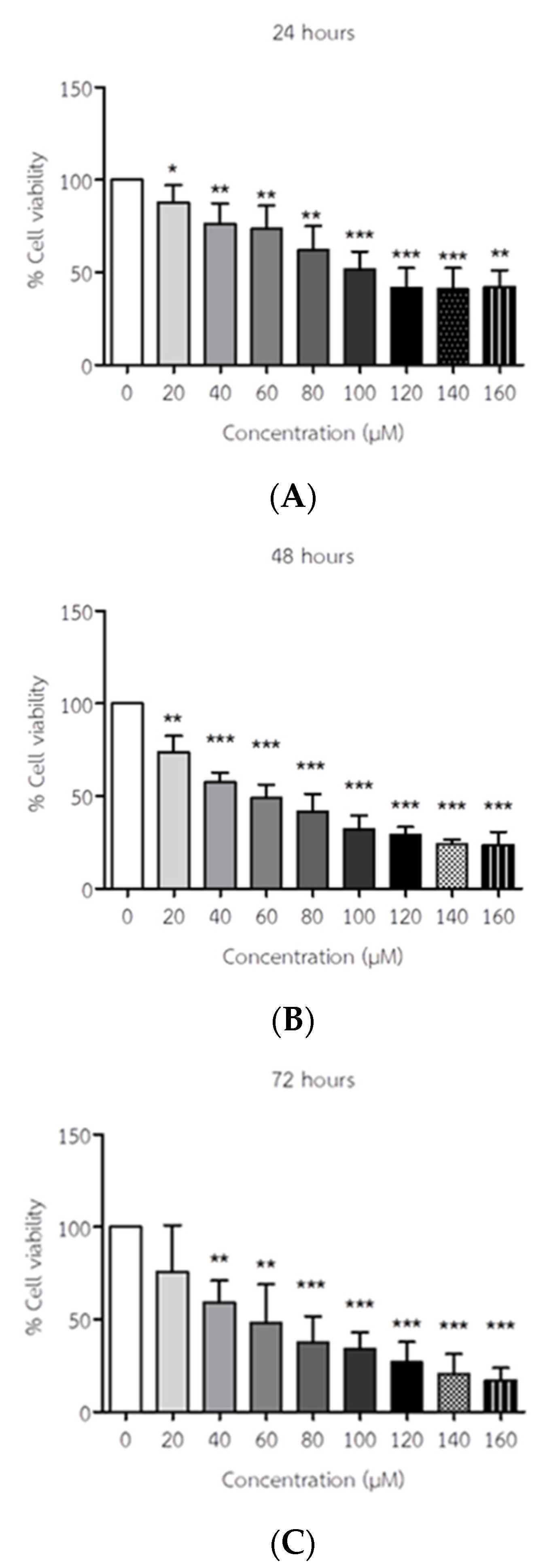
Figure 3.
The effect of API on LDH levels of ELT3 cells. The results represent mean± S.D. *p < 0.05 and **p < 0.005 significantly different from control and DMSO treatment.
Figure 3.
The effect of API on LDH levels of ELT3 cells. The results represent mean± S.D. *p < 0.05 and **p < 0.005 significantly different from control and DMSO treatment.
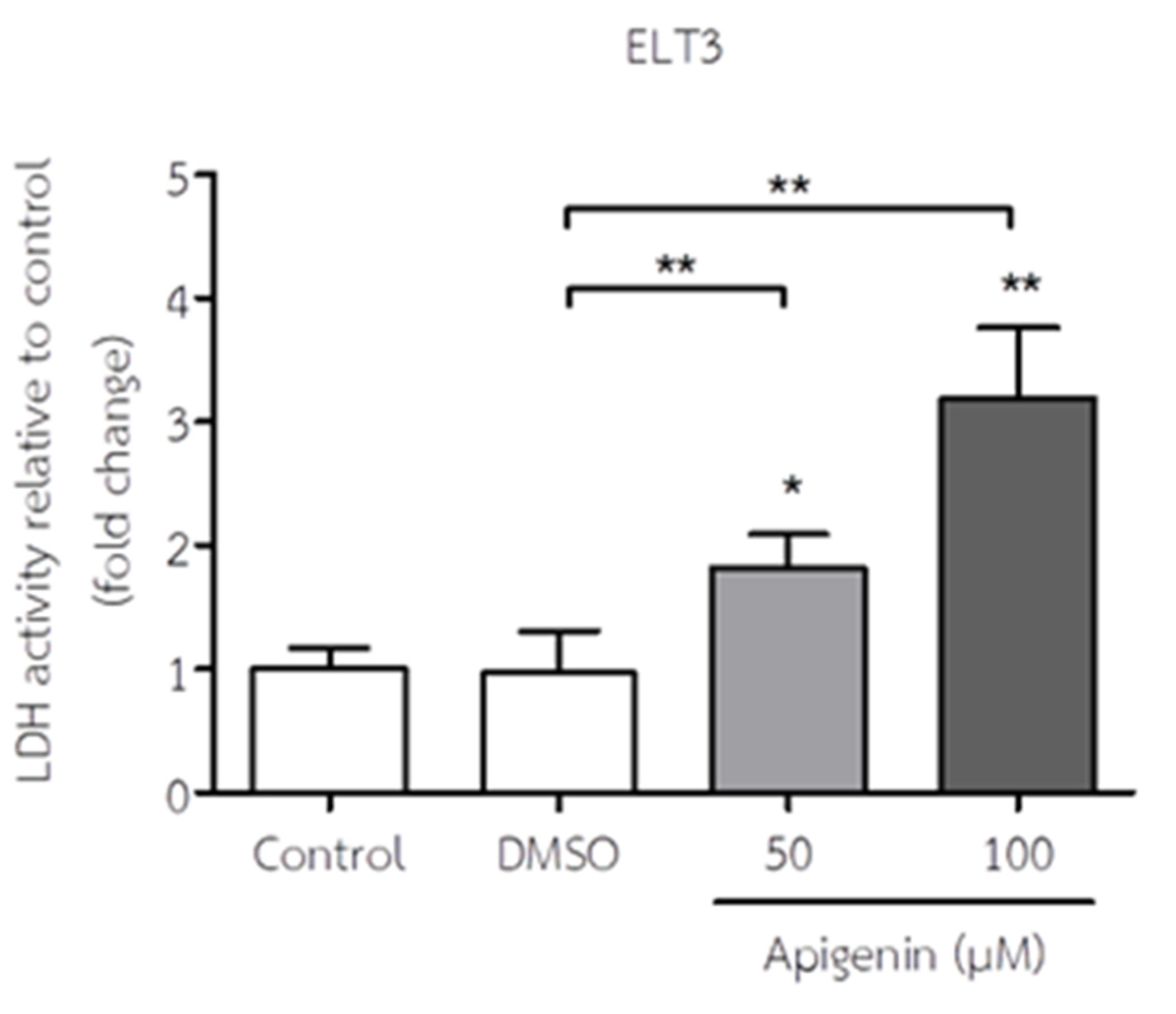
Figure 4.
The effect of API on ROS production of ELT3 cells. The results represent mean± S.D. **p < 0.005 significantly different from control. THBP, Tert-Butyl hydrogen peroxide, is a positive control.
Figure 4.
The effect of API on ROS production of ELT3 cells. The results represent mean± S.D. **p < 0.005 significantly different from control. THBP, Tert-Butyl hydrogen peroxide, is a positive control.
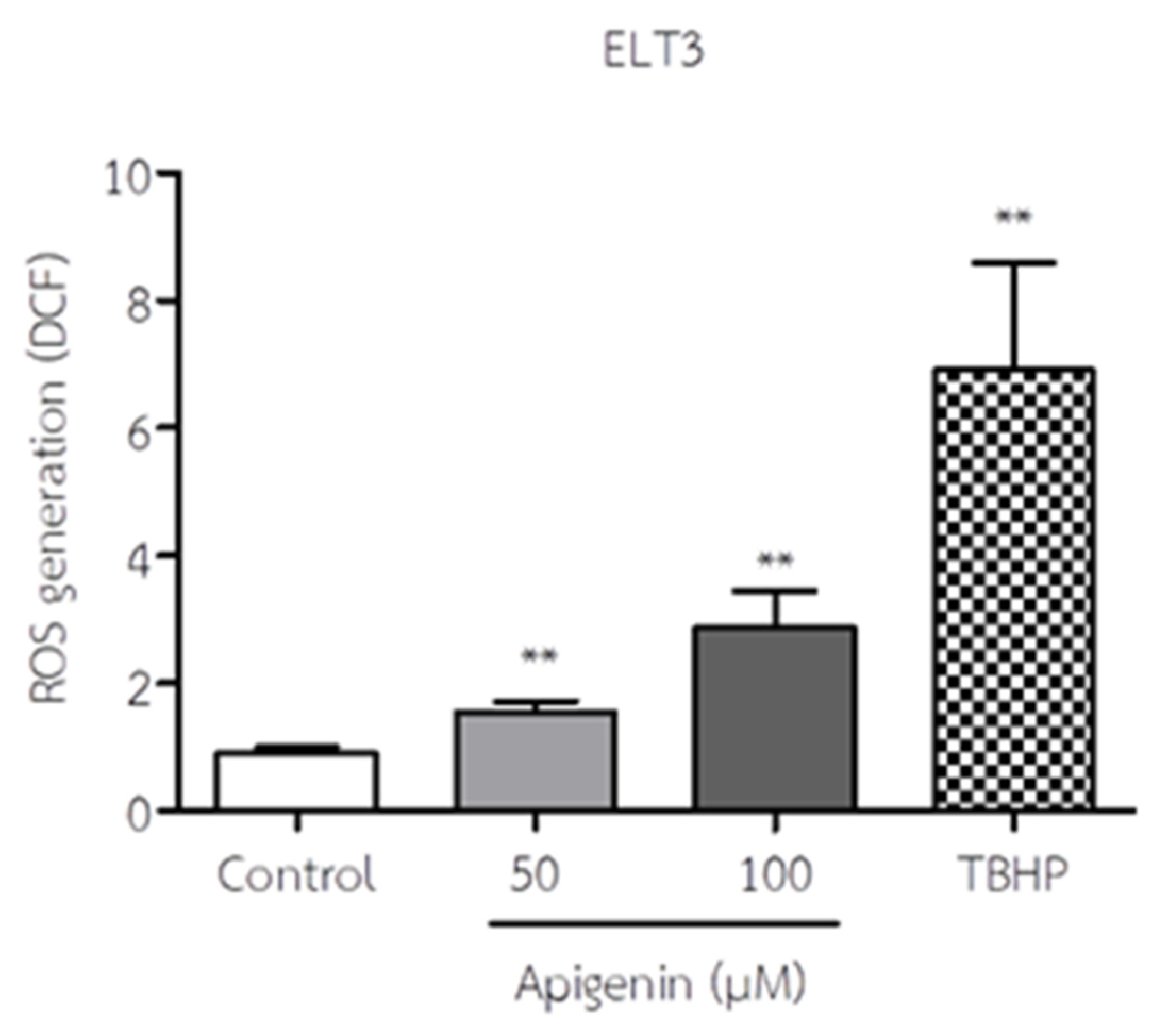
Figure 5.
The effect of API on MDA levels of ELT3 cells. The results represent mean± S.D. **p < 0.005 significantly different from control.
Figure 5.
The effect of API on MDA levels of ELT3 cells. The results represent mean± S.D. **p < 0.005 significantly different from control.
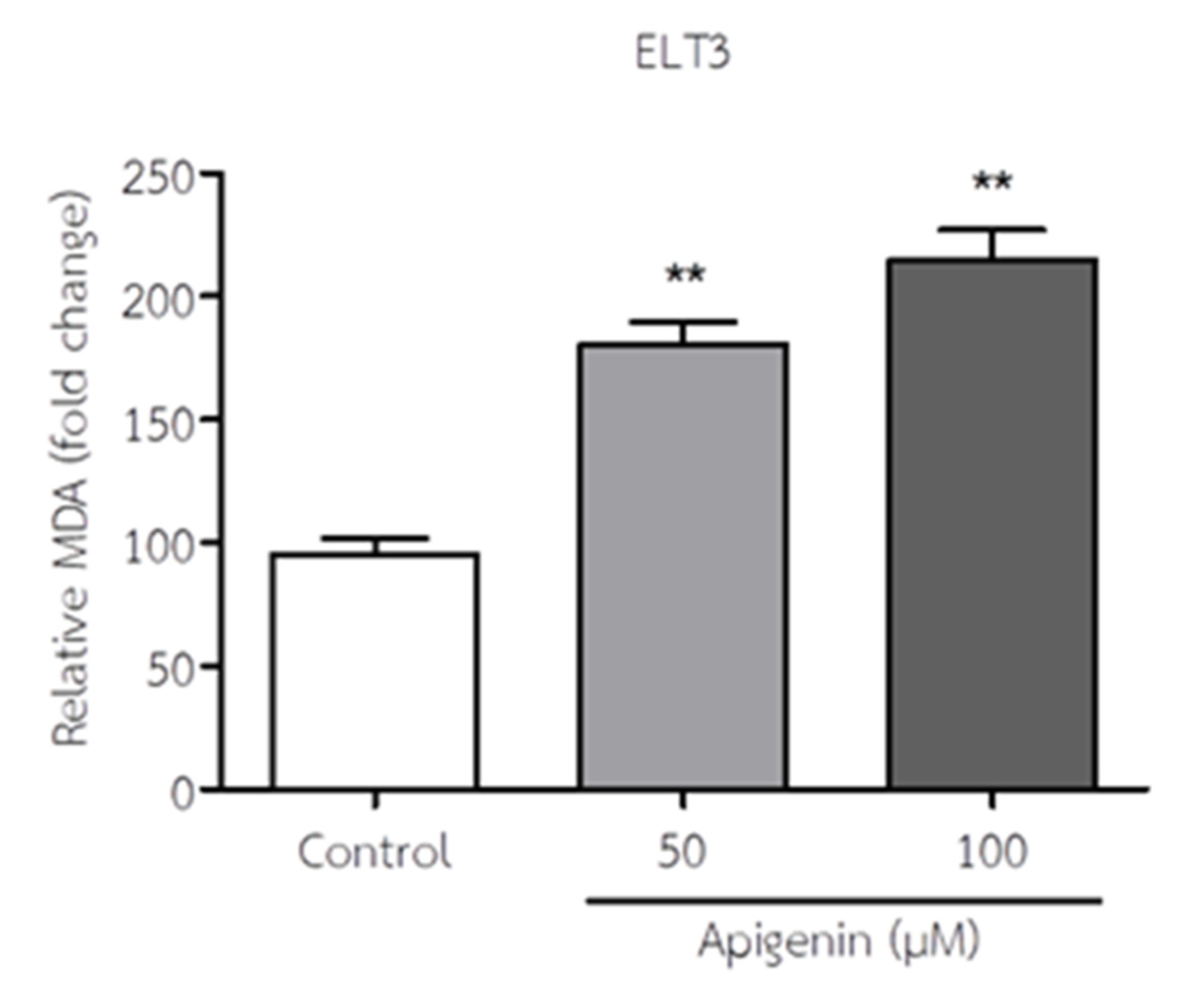
Figure 6.
The effect of API on GR activity of ELT3 cells. The results represent mean± S.D. *p < 0.05 and **p < 0.01 significantly different from control and DMSO treatment.
Figure 6.
The effect of API on GR activity of ELT3 cells. The results represent mean± S.D. *p < 0.05 and **p < 0.01 significantly different from control and DMSO treatment.
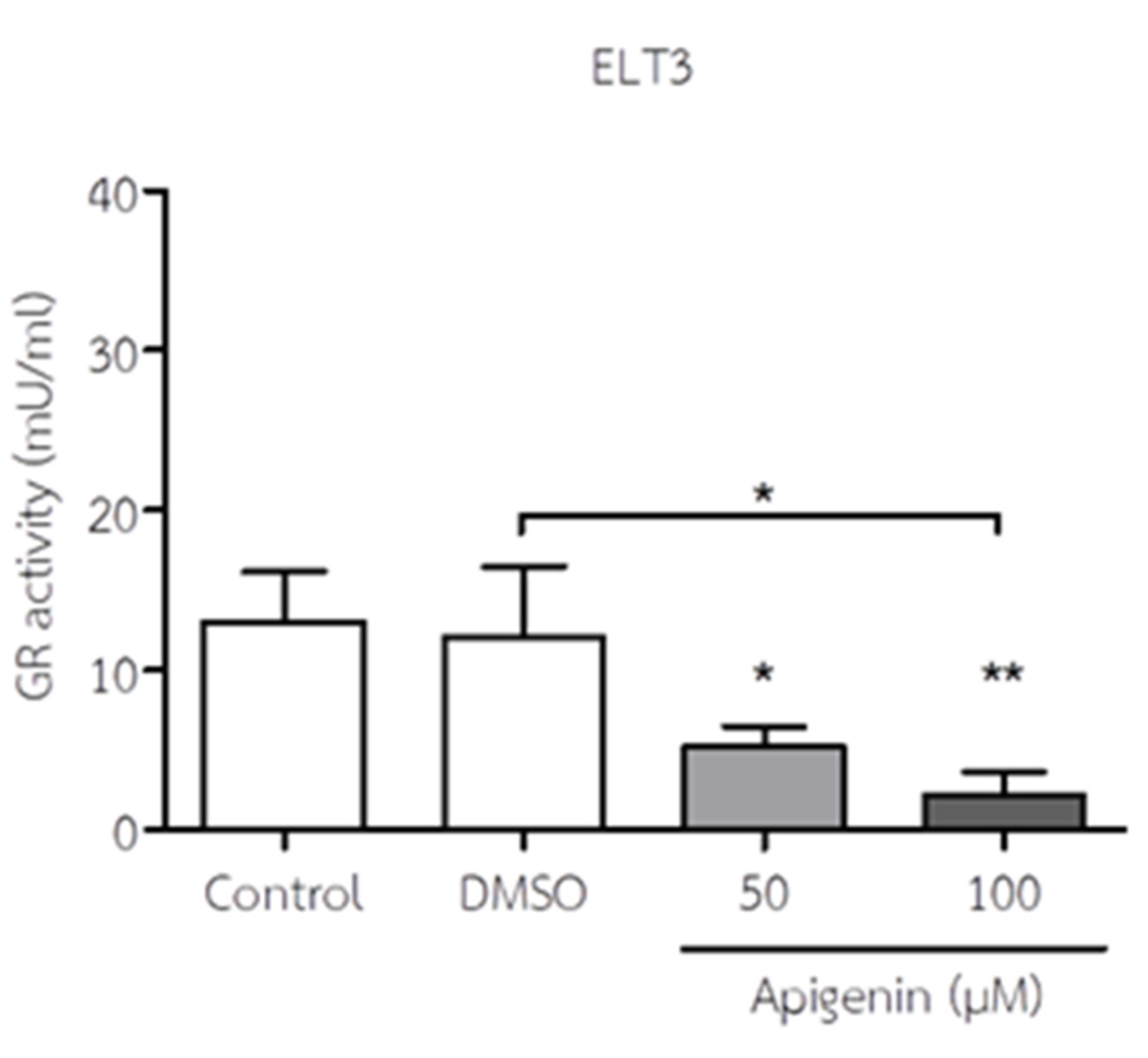
Figure 7.
The effect of API on apoptotic gene expression of ELT3 cells. The results represent mean ± S.D. *p < 0.005 and **p < 0.001 significantly different.
Figure 7.
The effect of API on apoptotic gene expression of ELT3 cells. The results represent mean ± S.D. *p < 0.005 and **p < 0.001 significantly different.
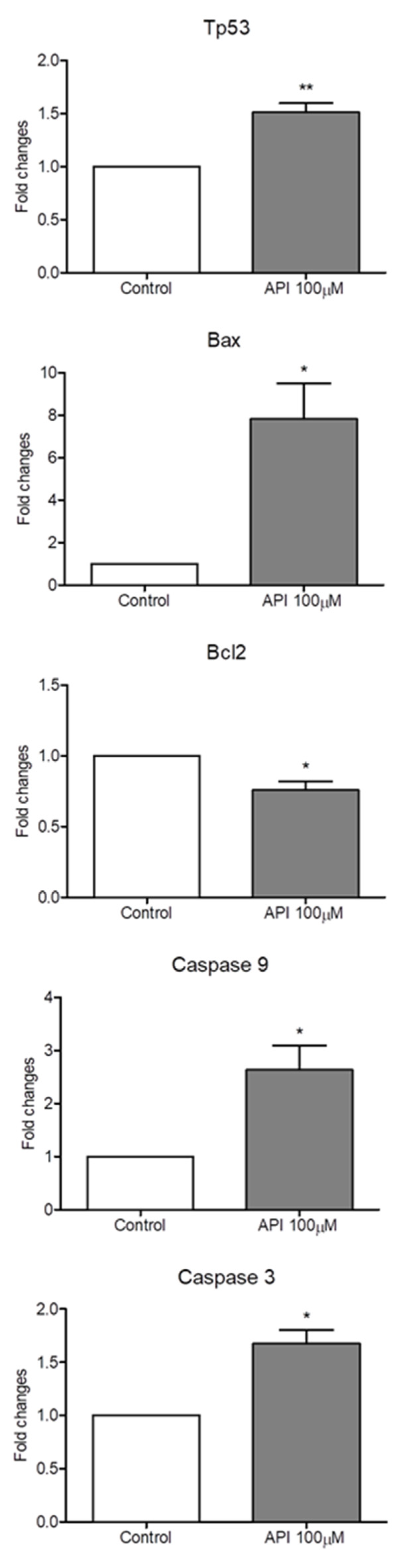

Table 1.
Primer used for qRT-PCR.
| Gene name | Forward primer | Reverse primer |
| GAPDH | ATGGGAAGCTGGTCATCAAC | GTGGTTCACACCCATCACAA |
| Tp53 | TCTCCCCAGCAAAAGAAAAA | TTTTATGGCGGGACGTAGAC |
| Bax | AAAGACATTGGAGCCACCAC | TATTGCCTGCCACAAACTCA |
| Bcl2 | GGGATGCCTTTGTGGAACTA | CATATTTGTTTGGGGCAGGT |
| Caspase3 | AGGGGCATGTTTCTGTTTTG | CATTGCAGGCAGTGGTATTG |
| Caspase9 | TCATTCTTGCAAAGCAGTGG | TGGGTGTTTCTGGTGTGAGA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



