
Submitted:
25 July 2024
Posted:
25 July 2024
You are already at the latest version
Abstract
Differences in metabolic regulation among obesity phenotypes, specifically metabolically healthy obese (MHO) and metabolically unhealthy obese (MUO) women, may lead to varied responses to interventions, which could be elucidated through metabolomics. Therefore, this study aims to investigate the differences in metabolite profiles between MHO and MUO women and the changes following a lifestyle intervention. Serum from 36 MHO and 34 MUO women, who participated in a lifestyle intervention for weight loss were analyzed using an untargeted proton nuclear magnetic resonance spectroscopy (1H NMR) at baseline and 6 months post-intervention. Anthropometric, clinical and dietary intake parameters were assessed at both time points. Both groups showed differential metabolites profile at baseline and sixth month. Seven metabolites including trimethylamine-N-oxide (TMAO), arginine, ribose, aspartate, carnitine, choline and tyrosine significantly changed between group post-intervention, in which all showed a decreasing pattern in MHO. Significant reduction in body weight and BMI in the MUO correlated with changes in carnitine and tyrosine levels. In conclusion, metabolite profiles differed significantly between MHO and MUO women before and after a lifestyle intervention. Changes in carnitine and tyrosine levels in MUO correlated with weight loss, suggesting potential targets for therapeutic intervention.
Keywords:
Metabolomics
; NMR
; metabolically healthy obese
; lifestyle intervention
; obesity
1. Introduction
Globally, there were 2.5 billion adults living with overweight and obesity [1]. In the last 12 years, Malaysia has seen a 10% rise in the prevalence of overweight and obesity [2] and this trend is not unique to Malaysia but is evident in many countries globally. The persistent rise in global obesity prevalence remains a significant public health concern worldwide, given its well-established association with an increased risk of developing chronic conditions, such as type 2 diabetes (T2D), hypertension (HPT) and cardiovascular diseases (CVD) [3,4]. Efforts to combat obesity through various weight loss interventions are widespread. However, the outcomes associated with weight loss intervention vary significantly among individuals.
Variations in the clinical manifestations of obesity could be one of the attributes that lead to differences in the responses towards obesity intervention regimens. Weight loss resulting from lifestyle interventions among overweight/obese women categorized as metabolically healthy has been linked to changes in lipid metabolism, activation of sulfation processes, and modulation of microbiota metabolism, potentially indicating a metabolically protective effect [5]. However, gaps persist in understanding disparities in metabolite profiles among obesity phenotypes and the underlying mechanisms that could contribute to differential responses to interventions. Comprehensive investigation is required to clarify the regulatory mechanisms that may vary across different obesity phenotypes.
Metabolomics provides a platform that allows us to capture the dynamic physiological conditions corresponding to current health conditions by analyzing low-molecular-weight metabolites present in tissues or biological fluids, such as lipids, amino acids, peptides, organic acids, and carbohydrates [6,7]. This comprehensive profiling of metabolites offers valuable insights into the etiology of obesity and other diseases, facilitating the discovery of potential biomarkers that could enhance our current knowledge of obesity pathophysiology and its related comorbidities.
Hence, the aims of our study were to explore the differences in serum metabolite profiles between women classified as metabolically healthy obese/overweight (MHO) and metabolically unhealthy obese/overweight (MUO) and examine the changes in these metabolite profiles following a 6-month lifestyle intervention for weight loss.
2. Materials and Methods
2.1. Study Design and Study Participants
This study utilized archived samples collected from an extension study of a lifestyle intervention for weight loss known as My Body is Fit and Fabulous at Home (MyBFF@home). The study was conducted in 2015 and the samples were stored at -80°C freezer. Information on the study design and recruitment details has been published elsewhere [8]. Participants in this study consisted of obese and overweight housewives aged 18 - 59 years old, living in the low-cost flats in Klang Valley, Malaysia. All the participants underwent six months lifestyle intervention for weight loss program consisting of dietary counselling, physical activity (PA) and self-monitoring tools (PA diary, food diary and pedometer). Fasting blood samples, anthropometric and clinical measurements were assessed at baseline and after 6 months of intervention. Biochemical profiles were analyzed in the laboratory according to methods described previously [9]. Dietary intakes were recorded using 3-day food diary and nutrient intakes were calculated using Nutritionist Pro TM version 2.4 (First Data Bank, The Hearst Corp, NY USA) as per described elsewhere [10]. A total of 70 (MHO=36, MUO=34) serum were randomly selected from the archived follow-up samples of MyBFF@home. Participants were considered as metabolically healthy obese (MHO) based on the following criteria: HbA1c <6.5% and systolic or diastolic blood pressure of <140mmHg, <90mmHg respectively, while participants were considered as metabolically unhealthy obese (MUO) by the following criteria: HbA1c ≥6.5%, systolic or diastolic blood pressure of ≥140mmHg, ≥90mmHg respectively, and self-reported of being diagnosed with T2D or HPT [11].
2.2. Sample Preparation for Metabolite Profiling
All samples were kept at -80°C prior to the analysis. Serum samples were thawed on ice to minimize metabolite degradation. Serum samples were vortexed and centrifuged (20,000g x 5min at 4°C). 200μL of the serum supernatant were mixed with 400μL of phosphate buffer (KH2PO4), pH 7.4 in deuterium oxide (D2O) and 0.1% TSP (Merck, Darmstadt, Germany) and 0.1% imidazole (Sigma-Aldrich, St. Louis, MO, USA) in a 1:1 ratio.
2.3. NMR Metabolomic Analysis
Untargeted metabolomic approach were used to analyze the metabolites present in the study. The NMR analysis method was adapted and optimized based on the work of Maulidani et al. [12]. 1D 1H-NMR spectra were collected at a temperature of 26°C using a 600 MHz Jeol NMR. The combination of PRESAT with CPMG pulse sequence were used to suppress water signals and wide protein resonances. NMR spectra with a spectral width of 10ppm were acquired using a total of 128 scans and a 660-second acquisition time. Spectra were processed using the Chenomx NMR suite version 9.0 software (Chenomx Inc., Edmonton, AB, Canada) with the following settings: 0.50 Hz line broadening, autophasing, baseline correction (Whittaker spline), referenced to TSP as an internal standard, and referenced to imidazole as a pH indicator. The spectral band between 0.50 and 10.00 ppm were divided into equal bins using intelligent binning (0.04 ppm). The peak ppm readings were calibrated against the 0 ppm TSP signal. The area of the spectrum associated with residual water and imidazole were eliminated prior to analysis. Subsequently, the corresponding spectra were transformed to a table of common integrals that has a non-negative value for multivariate data analysis (MVDA).
2.4. Statistical Analysis
Prior to MVDA, the spectral data were mean-centered and pareto-scaled to improve normality. Principal component analysis (PCA) and partial least squares discriminant analysis (OPLS-DA) were performed to detect outliers and observe trends and separation of metabolites between groups using the standard algorithm as implemented in the SIMCA® software version 17.0.2 (Sartorius, Göttingen, Germany). The validation of the OPLS-DA model was carried out using cross-validated analysis of variance (CV-ANOVA), with the results expressed as p-values for the model. Prior to univariate data analysis the metabolomics data were log-transformed and metabolites not found in at least 20% of the samples were removed. Statistical analysis was conducted using IBM SPSS Statistics for Windows, Version 27.0 (Armonk, NY, USA). The normality of continuous data was determined using Shapiro-Wilk test. Differences in the baseline parameters of the participants between the two groups were analyzed using independent t-tests. Generalized estimated equation (GEE) was used to assess significantly changed metabolites between groups (time X group). The analysis was adjusted for sociodemographic characteristics (age, education level and household income), baseline value of anthropometry parameters (BMI and WC), FPG, HbA1c, systolic blood pressure, the used of medication and baseline value of dietary intake (energy, carbohydrate, protein, total fat, saturated fat, sodium, potassium, and dietary fiber. The Benjamini–Hochberg (B-H) method was applied to correct for multiple testing with false discovery rate (FDR) set at 5%. To assess the extent of changes experienced after the intervention for all participants, delta values were calculated. These values were derived from the ratio of the sixth month measurements to the baseline values of significant metabolites, anthropometric measures, blood pressure, and biochemical variables. Pearson correlation analysis was then performed using the delta values to evaluate the relationship between the significant metabolites and anthropometry (weight, BMI and WC), biochemical (FPG, cholesterol, HDL, LDL and TG) and clinical data (systolic and diastolic blood pressure).
2.5. Pathway Analysis
To explore the metabolic pathways affected by changes in metabolites following the intervention, pathway analysis was conducted in the MetaboAnalyst 6.0 [13] web application, which utilizes databases for Homo sapiens from the Kyoto Encyclopedia of Genes and Genomes (KEGG) and the Human Metabolome Database (HMDB) as the pathway library. The Materials
3. Results
3.1. Baseline Characteristics of Study Participants
The demographic, anthropometric, and clinical characteristics of the study participants (MHO vs. MUO) are presented in Table 1. The mean age was 41.65 ± 8.46 years for MHO group and 50.01 ± 6.26 years for MUO group which indicates a highly significant different age gap between the two groups (p<0.001). Half of the participants in the MHO group were between 40 to 49 years old, while most of the participants in the MUO group (41.2%) were over 50 years old. The racial breakdown of participants in both groups was predominantly Malay (91.7% vs. 88.2%) and Indian (8.3% vs. 11.8%). The education level and household income of the participants were also significantly different (p<0.05) between the groups, with more than 90% of MHO participants achieving secondary/tertiary level of education and earning higher than the MUO group. BMI, waist circumference (WC), blood pressure, fasting plasma glucose (FPG) and HbA1c were significantly higher in the MUO compared to MHO (p<0.05). Conversely, there were no significant differences in lipid profiles between the groups. The significantly different parameters were accounted for in the subsequent statistical analysis.
3.2. Anthropometric, Clinical and Dietary Intake Changes Following 6-Month Intervention
The between-group analysis of the anthropometric and clinical data revealed body weight and WC showed significant changes post-intervention (p<0.05), as summarized in Table 2. A further within group analysis revealed the MUO showed significant reduction in body weight, BMI, WC and systolic blood pressure. All the dietary intake parameters showed significant decreased (p<0.05) following the intervention where decreased intake was evident in both groups.
3.3. Multivariate Analysis
An unsupervised principal component analysis (PCA) was performed using the binned data to observe potential separation between the MHO and MUO groups at baseline and the sixth month. The baseline PCA score plot (Figure S1) suggested that the metabolomic profiles of the two groups were similar, as indicated by the scattered and random scores on the plot. However, at the sixth month, the score plot (Figure S2) revealed some degree of separation between the groups, although there were visible overlapping scores. Further analysis then performed using the orthogonal projection latent square-discriminant analysis (OPLS-DA) to examine for a more distinct separation of the group at both baseline and sixth month. In contrast to the corresponding PCA score plot, the baseline OPLS-DA score plot shows a pattern of separation between MHO and MUO with the goodness of fit, R2 (cum) = 0.336 and predictive ability, Q2 (cum) = 0.207 by which the model could explain 33.6% variation in MHO and MUO metabolomic profiles. Likewise, the sixth month OPLS-DA score plot also revealed a distinct separation between MHO and MUO where the model attributed 62.7% variation in the metabolic profiles of the two groups with R2 (cum) = 0.627 and Q2 (cum) = 0.140. Cross validation analysis of variance (CV-ANOVA) performed on both baseline and sixth month model resulted in p<0.05, indicating that both models were significant. The anticipated significant clustering of MHO and MUO in the OPLS-DA model is likely attributed to the notable differences in clinical parameters between the two groups. A VIP score >1.0 on the loading plot (Figures S3 and S4) was utilized to pinpoint the regions that discriminate between MHO and MUO metabolic profiles. Regions with negative error bar values were excluded. The corresponding S-plot at baseline and at the sixth month, illustrated in Figure 1C and 1D, highlight the discriminating regions (ppm) in red. A detailed examination of the S-plot indicated that the regions with the highest magnitude and reliability in the OPLS-DA model were 3.42ppm and 3.46ppm at baseline, and 3.42ppm and 3.74ppm at the sixth month denoting that the significant role of these regions in differentiating the metabolic profiles between MHO and MUO.
3.4. Identification and Relative Quantification of Metabolites
The spectral data were profiled to 39 known metabolites, identified with reference to the Human Metabolome Database (HMDB) using Chenomx and the relative concentration of each metabolite was obtained by peak fitting with reference to TSP signal. The identified metabolites were listed in the Supplementary Table S2 along with their respective regions. To further understand the differences in metabolites levels between MHO and MUO at the two time points, the relative concentration data of the identified metabolites were exported from Chenomx. The concentration data were log-transformed to improve normality. Subsequently, an independent t-test was conducted to evaluate the disparity in metabolite levels between the groups at baseline and sixth month. The significant metabolites were summarized in Table 3. Three metabolites were found to be significantly different and higher (p<0.05) in the MUO group compared to MHO group at baseline, namely glucose, indole-3-acetate and τ-methylhistidine. At sixth month after the intervention, 15 metabolites were significantly different (p<0.05) between the MHO and MUO group. Fructose is the only metabolite that was significantly lower (p=0.044) in the MUO group. The remaining fourteen metabolites, namely acetate, arginine, aspartate, betaine, glucose, histidine, isobutyrate, isoleucine, leucine, N-acetylcysteine, phenylacetate, trimethylamine-N-oxide (TMAO), tyrosine and valine were significantly higher (p<0.05) in the MUO group compare to the MHO group.
The changes in metabolite levels from baseline to the sixth month between the groups (time x group effect) were evaluated using generalized estimating equation (GEE) analysis. This analysis was adjusted for sociodemographic characteristics (age, education level, and household income), baseline anthropometric parameters (BMI and WC), FPG, HbA1c, systolic blood pressure, medication use, and baseline dietary intake (energy, carbohydrate, protein, total fat, saturated fat, sodium, potassium, and dietary fiber). A Benjamini-Hochberg (B-H) correction was performed to account for false discovery rate (FDR), where the metabolites were ranked based on their significant p-values. As a result, seven metabolites were found to be significantly different between the groups after six months of intervention. Trimethylamine-N-oxide (TMAO), arginine, ribose, aspartate, carnitine, choline and tyrosine were the finalized significant metabolites. The details on the changes of these seven metabolites were summarized in Table 4 and illustrated in Figure 2.
As illustrated in Figure 2, the changes in metabolites levels in MHO group is greater as compared to the MUO group except for tyrosine where the changes were more prominent in MUO. It is also evident clearly that the changes for all the seven metabolites in MHO showed a decreasing pattern after sixth month of intervention, while in MUO only two metabolites (TMAO, ribose) showed decreasing pattern and the other seven metabolites displayed an increasing pattern. The most change was observed in the level of arginine in MHO where it decreased by 8.62%, p<0.05. In the MUO, tyrosine was the metabolite that displayed the most change in which the level decreased by 5.16%, p<0.05.
3.5. Pathway Analysis
Pathway analysis was conducted at two time points using concentration data from 40 metabolites. Initially, no significant pathways were identified at baseline. However, after six months of lifestyle intervention, significant changes in metabolic pathways were observed between MHO and MUO women, as shown in Figure 3. The most impactful pathways were phenylalanine, tyrosine, and tryptophan biosynthesis, with an impact value (piv) of 0.500 and a significant FDR-adjusted p-value of 0.016. Additionally, the starch and sucrose metabolism pathway were also significant and impactful, with a piv of 0.425 and a p-value of 0.009.
3.6. Correlation between the Significantly Changed Metabolites with Anthropometry and Clinical Variables
Delta values for each variable were computed by the ratio of the sixth month measurement to its corresponding baseline value. This approach was employed to accommodate the dynamic fluctuations of the variables over the intervention period. This delta values were then utilized to examine the correlation between the metabolites that exhibited significant changes with anthropometric, biochemical, and clinical variables using Pearson correlation analysis. Given the MUO's significant reduction in body weight, BMI, WC and systolic blood pressure, a correlation analysis was conducted to examine the relationship between the significant metabolites and these parameters. The results indicate that carnitine has a significant negative correlation with body weight (r= -0.367, p= 0.033) and BMI (r= -0.357, p= 0.038). Similarly, tyrosine shows a significant negative correlation with both body weight (r= -0.381, p= 0.038) and BMI (r= -0.401, p= 0.028) as presented in the heatmap correlation matrix in Figure 4.
A pooled correlation analysis was conducted irrespective of grouping to investigate the correlation between the significant metabolites with anthropometry and clinical variables among women with obesity. A heatmap correlation matrix was generated based on the correlation coefficient, r, as shown in Figure 5. No significant correlation was found with blood pressure parameters. Carnitine displayed a significant negative correlation with changes in weight (r= -0.364, p=0.002) and BMI (r= -0.353, p=0.003), while exhibiting a positive correlation with changes in TC (r= 0.280, p=0.023). Ribose showed significant positive correlation with changes in both FPG, (r=0.396, p=0.001) and HDL (r=0.298, p=0.014). TMAO demonstrated a negative correlation between changes in BMI (r= -0.257, p=0.032) and TG (r= -0.261, p=0.037). Both arginine and tyrosine showed significant correlation with age, r=0.248, p=0.040 and r=0.421, p<0.001 respectively.
4. Discussion
Our study demonstrated that prior to the intervention, the spectral regions 3.42ppm and 3.46ppm were highly predictive of differentiating between MHO and MUO. Following the intervention, regions 3.42ppm and 3.74ppm were highly predictive of the separation between MHO and MUO. Based on the spectral data generated and profiled in this study, these regions corresponded to glucose which was found to be significantly different between MHO and MUO both at baseline and sixth month of the intervention. Initial investigation of metabolite differences at baseline identified three metabolites were significantly different between the MHO and MUO groups. However, after six months of intervention a greater number of metabolites (15 metabolites) showed significant differences between the groups. Among these were the branched-chain amino acids (BCAAs): leucine, isoleucine, and valine which have been shown to be associated with insulin resistance, metabolic syndrome and T2D in various cohorts [14,15,16]. This aligns with our findings, where the levels of BCAAs were significantly higher in MUO women compared to MHO women. The proposed mechanisms linking BCAAs and T2D involve elevated BCAAs levels triggering activation of the mTOR pathway. This activation increases the production of lipid intermediates such as diacylglycerol (DAG), which subsequently results in the phosphorylation of insulin receptor substrate (IRS) proteins, impairing downstream insulin signalling [17,18]. The frequent association between BCAAs and obesity and T2D has led to suggestions that MHO and MUO could be characterized based on these metabolites [19].
Further analysis revealed that seven metabolites significantly changed following the intervention between MHO and MUO women namely TMAO, arginine, ribose, aspartate, carnitine, choline and tyrosine. All these metabolites displayed a significant decreasing trend in the MHO group. The substantial decrease in circulating choline, carnitine, and TMAO were likely be attributed to reduced intake of protein-rich foods such as red meat, which are high in choline and carnitine during the intervention. TMAO is produced by the gut microbiota from these precursors [20], hence a lower intake of choline- and carnitine-rich foods putatively leads to decreased TMAO levels. This direct relationship was demonstrated previously, showing that the consumption of fish and meat is associated with plasma concentrations of TMAO [21]. Additionally, the marked reduction in arginine levels in MHO could also be linked to the decreased protein intake as studies have shown that dietary intake of arginine is significantly associated with serum and plasma arginine levels, suggesting that a lower intake of arginine-rich foods, during the intervention may contribute to this observed decrease [22,23].
Tyrosine is an aromatic amino acid categorized as non-essential since it can be synthesized in our body from its precursor, phenylalanine. Our study does not only demonstrate that tyrosine levels were markedly elevated in MUO post-intervention but was also found to be higher in the MUO compared to MHO at baseline. Similar to BCAAs, elevated levels of tyrosine have been widely linked to insulin resistance and an increased risk of T2D [16,24,25]. This aligns with our findings, as some participants in the MUO group either had diagnosed T2D or exhibited elevated HbA1c levels beyond the normal range. Additionally, tyrosine has been linked to aging [26], which was also evident in our study as we found a positive correlation between tyrosine levels and age.
Despites the substantial changed observed in the metabolomic profile of MHO and MUO women in the present study, only minimal change was observed in the anthropometry parameter while no significant changes were observed in the clinical parameters. Although the changes in body weight and WC across the groups were minimal but the change were significant. In this study, carnitine levels decreased in MHO, but increased in MUO following the intervention which was accompanied by significant reduction in body weight and WC in the latter. Additionally, a significant correlation was found between carnitine and both body weight and BMI. A meta-analysis previously has revealed that supplementation with L-carnitine significantly reduced body weight [27]. This could pertain to the role of carnitine in fatty acid metabolism. Carnitine is crucial for transporting fatty acids into the mitochondria, where they undergo β-oxidation and are broken down into short and medium chain fatty acids to produce energy [28]. The increased energy expenditure through fatty acid metabolism is presumed to be the factor that promoted weight loss in the MUO women. Additionally, during periods of reduced calorie intake and weight loss, it is possible that more carnitine was released from the tissues [29]. This could potentially account for the high levels of carnitine observed in MUO individuals despite their reduced protein intake. The same pattern of changes and correlation with weight loss was also observed with tyrosine. Previous research has shown weight reduction is associated with decreased tyrosine levels in overweight/obese [30] and those with metabolic syndrome [31]. However, in the present study tyrosine levels was elevated in the MUO after the intervention and was significantly correlated with weight reduction. A study on 3-week weight loss program also reported significant weight loss with increase tyrosine levels following intervention. While elevated levels of tyrosine in the MUO have often been reported [32,33,34] and are associated with insulin resistance and T2D [16,24,25], the discrepancy in the findings could potentially be attributed to several factors including differences in the study population, participants’ dietary intake and the methods used to measure the metabolites.
We conducted a pooled correlation analysis to investigate the association between the significant metabolites and various anthropometric and clinical parameters using the delta data and found TMAO, arginine, carnitine and ribose were correlated with lipid parameters namely TG, LDL, TC and HDL respectively. These metabolites are mostly involved in the energy metabolism, lipid metabolism and vascular function. TMAO were found to be negatively correlated with both BMI and TG. These correlations were also reported in other studies involving overweight individuals, though they found a positive correlation [35,36]. The correlation might be related to the role of the gut microbiota, as it has been shown to also affect changes in TG levels and BMI [37]. As previously mentioned, the circulating levels of TMAO are influenced by the dietary intake of protein-rich food and its biosynthesis by the gut microbiota [20]. In this study, participants across all groups exhibited a significant reduction in protein intake and corresponding decreased in TMAO levels. This suggests that changes in dietary protein intake could influence circulating TMAO levels by modulating the metabolism of the gut microbiota [38,39]. Another factor that could also influence the circulating levels of TMAO is the activity of the flavin monooxygenase 3 (FMO3) enzyme that may link the relationship between TMAO and lipid regulation. FMO3 does not only responsible for the conversion of TMA to TMAO in the liver but it also involves in the regulation of lipid via the farnesoid X receptor (FXR) and liver X receptor (LXR) which resulted in reduced reverse cholesterol transport to the intestine and leads to accumulation of cholesterol [40,41].
In the context of the positive correlation between arginine and LDL, it's plausible that changes in arginine could be a consequence of obesity rather than the effect on arginine itself on LDL changes as it has been reported in a meta-analysis that supplementation with arginine did not have any significant impact on LDL [42]. Apart from excessive body weight, participants in the present study exhibited both impaired fasting glucose and hyperlipidemia. This aligns with the well-documented coexistence of obesity with insulin resistance, metabolic syndrome, inflammation, and oxidative stress [43]. Chronic inflammation and metabolic abnormalities associated with obesity are thought to increase the activity of arginase 1, an enzyme that competes with nitric oxide (NO) synthase for arginine [44,45]. This competition reduces NO production, which is crucial for maintaining healthy vascular function. The decrease in NO leads to increased production of reactive oxygen species (ROS), further promoting oxidative stress and endothelial dysfunction [44,45]. Consequently, this could result in lipid dysregulation, including elevated LDL levels [46,47], as observed in our study. As previously suggested, the significant reduction in carnitine may be attributed to decreased protein intake during the intervention. Therefore, the observed association between carnitine and total cholesterol (TC) might also be due to metabolic disturbances associated with obesity. It is worth noting that studies have shown that supplementation with L-carnitine significantly reduces TC levels [48,49] which contradicted with our findings.
We also managed to demonstrate correlation between ribose-FPG and ribose-HDL. Unfortunately, no previous human study has reported such correlation. While small amount of ribose can be sourced from dietary intake, our body primary source of ribose is synthesized through pentose phosphate pathway (PPP) [50]. Obesity is known to be associated with elevated activity of glucose-6-phosphate dehydrogenase (G6PD), the rate-limiting enzyme in the pentose phosphate pathway (PPP). The increase in G6PD activity led to enhanced oxidative PPP activity, resulting in higher NADPH production. The surplus NADPH fueled NADPH oxidase (NOX)-mediated reactive oxygen species (ROS) generation, which exacerbated inflammatory response and induced DNA damage [51,52,53,54,55]. The non-oxidative phase of the pentose phosphate pathway (PPP) produces ribose-5-phosphate (R5P), a crucial building block for nucleotide synthesis [56]. Obesity-induced DNA damage could putatively increase the non-oxidative PPP activity to boost nucleotide synthesis necessary for DNA repair leading to reduced circulating levels of ribose as observed in the present study.
This study has certain limitations. We did not measure the inflammatory cytokines associated with obesity; thus, we could not verify the association between obesity-related inflammation and the metabolites. Additionally, since the samples were obtained from a community-based intervention program, we lacked access to the medical history of participants with comorbidities (T2D and HPT), particularly the duration since their diagnosis. This is a notable limitation, as existing evidence indicates that metabolite levels significantly change as these diseases progress [57]. Consequently, we were unable to control for this factor in our analysis. Furthermore, dietary intake data were recorded using self-reported 3-day food diaries, which may lead to underreporting. However, to minimize this issue, each participant's food diary was reviewed one-on-one with a professional dietician or nutritionist at each visit.
5. Conclusion
In conclusion, our study highlights distinct metabolomics profiles between MHO and MUO individuals before and after a lifestyle intervention for weight loss, with glucose being a key differentiating metabolite. The intervention also led to significant changes in seven metabolites; TMAO, arginine, ribose, aspartate, carnitine, choline, and tyrosine with a decreasing pattern in these metabolites was observed in the MHO group, potentially linked to reduced protein intake, altered lipid metabolism, and gut microbiota modulation. Interestingly, we highlight the increased in carnitine and tyrosine levels following the intervention in MUO was associated with significant weight loss. This finding suggests that carnitine and tyrosine could be considered as potential therapeutic targets for weight loss, particularly in MUO women. However, a larger scale and long-term study is warranted to observe the full extent of these effects. Additionally, future studies should consider the potential interactions with other dietary components and the individual variability in response to interventions. Such research could provide valuable insights into personalized nutrition strategies for effective weight management, particularly for MUO women.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Table S1: List of metabolites identified at baseline and sixth month; Table S2: PCA model fit at baseline and sixth month; Table S3: OPLS-DA model fit at baseline and sixth month; Figure S1: PCA score plot at baseline; Figure S2: PCA score plot at sixth month; Figure S3: Loading plot at baseline; Figure S4: Loading plot at sixth month.
Author Contributions
Conceptualization, F.S., M.N.M.N., L.A.Z., N.A.W. and R.S.; Methodology, M.N.M.N. and A.M.; Formal Analysis, F.S.; Investigation, F.S.; Resources, M.F.M.N and F.M.; Data Curation, M.A.O and N.S.A.A; Writing – Original Draft Preparation, F.S.; Writing – Review & Editing, all authors; Supervision, R.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Ministry of Health Malaysia. Reference no. NMRR- 23-01417-1FD.
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki, and approved by the Medical Research Ethics Committee, Ministry of Health Malaysia (with approval code: NMRR ID-23-01417-1FD (IIR) and approval date: 8 June 2023).
Informed Consent Statement
Written informed consent was obtained from all subjects involved in the study.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors on reasonable request. A proposal with a detailed description of study objectives and a statistical analysis plan will be needed for assessment of requests. Additional materials might also be required during the process of assessment. Deidentified participant data will be provided after approval by the investigators.
Acknowledgments
The authors would like to thank the Director General of Health Malaysia for the permission to publish this paper. The authors would also like to thank all research team members from various institutions. Special thanks to all the participants in the MyBFF@home study, for their full co-operation and dedication.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- WHO. Obesity and Overweight Fact Sheets. Available online: https://www.who.int/news-room/fact-sheets/detail/obesity-and-overweight.
- IPH. National Health and Morbidity Survey (NHMS) 2023 Fact Sheet. Available online: https://iku.gov.my/nhms-2023.
- Hu, F.B. Globalization of diabetes: the role of diet, lifestyle, and genes. Diabetes Care 2011, 34, 1249–1257. [Google Scholar] [CrossRef] [PubMed]
- Poirier, P.; Giles, T.D.; Bray, G.A.; Hong, Y.; Stern, J.S.; Pi-Sunyer, F.X.; Eckel, R.H. Obesity and Cardiovascular Disease: Pathophysiology, Evaluation, and Effect of Weight Loss. Circulation 2006, 113, 898–918. [Google Scholar] [CrossRef] [PubMed]
- Palau-Rodriguez, M.; Garcia-Aloy, M.; Minarro, A.; Bernal-Lopez, M.R.; Brunius, C.; Gomez-Huelgas, R.; Landberg, R.; Tinahones, F.J.; Andres-Lacueva, C. Effects of a long-term lifestyle intervention on metabolically healthy women with obesity: Metabolite profiles according to weight loss response. Clin. Nutr. 2020, 39, 215–224. [Google Scholar] [CrossRef] [PubMed]
- Aleksandrova, K.; Egea Rodrigues, C.; Floegel, A.; Ahrens, W. Omics Biomarkers in Obesity: Novel Etiological Insights and Targets for Precision Prevention. Curr. Obes. Rep. 2020, 9, 219–230. [Google Scholar] [CrossRef] [PubMed]
- Rochfort, S. Metabolomics Reviewed: A New “Omics” Platform Technology for Systems Biology and Implications for Natural Products Research. Journal of Natural Products 2005, 68, 1813–1820. [Google Scholar] [CrossRef] [PubMed]
- Abdul Aziz, N.S.; Shahar, S.; Ambak, R.; Mohamad Nor, N.S.; Jamil, A.T.; Aris, T. Influence of co-morbidity on body composition changes after weight loss intervention among overweight housewives: a follow-up study of the MyBFF@home. BMC Womens Health 2018, 18, 115. [Google Scholar] [CrossRef] [PubMed]
- Liyana, A.Z.; Appannah, G.; Sham, S.Y.Z.; Fazliana, M.; Nor, N.S.M.; Ambak, R.; Samad, A.A.; Dahlan, N.Y.; Aris, T. Effectiveness of a community-based intervention for weight loss on cardiometabolic risk factors among overweight and obese women in a low socio-economic urban community: findings of the MyBFF@home. BMC Womens Health 2018, 18, 126. [Google Scholar] [CrossRef]
- Othman, F.; Mohamad Nor, N.S.; Appannah, G.; Zaki, N.A.M.; Ambak, R.; Omar, A.; Fazliana, M.; Salleh, R.; Yusof, B.N.M.; Muksan, N.; et al. Prediction of body fat loss in relation to change in nutrient intake among housewives participating in the MyBFF@home study. BMC Womens Health 2018, 18, 102. [Google Scholar] [CrossRef]
- Kim, M.; Yoo, H.J.; Ko, J.; Lee, J.H. Metabolically unhealthy overweight individuals have high lysophosphatide levels, phospholipase activity, and oxidative stress. Clin. Nutr. 2020, 39, 1137–1145. [Google Scholar] [CrossRef]
- Maulidiani; Abas, F.; Khatib, A.; Perumal, V.; Suppaiah, V.; Ismail, A.; Hamid, M.; Shaari, K.; Lajis, N.H. Metabolic alteration in obese diabetes rats upon treatment with Centella asiatica extract. J. Ethnopharmacol. 2016, 180, 60–69. [Google Scholar] [CrossRef]
- Pang, Z.; Lu, Y.; Zhou, G.; Hui, F.; Xu, L.; Viau, C.; Spigelman, A.F.; MacDonald, P.E.; Wishart, D.S.; Li, S.; et al. MetaboAnalyst 6.0: towards a unified platform for metabolomics data processing, analysis and interpretation. Nucleic Acids Res. 2024, 52, W398–W406. [Google Scholar] [CrossRef] [PubMed]
- Palmer, N.D.; Stevens, R.D.; Antinozzi, P.A.; Anderson, A.; Bergman, R.N.; Wagenknecht, L.E.; Newgard, C.B.; Bowden, D.W. Metabolomic profile associated with insulin resistance and conversion to diabetes in the Insulin Resistance Atherosclerosis Study. J. Clin. Endocrinol. Metab. 2015, 100, E463–468. [Google Scholar] [CrossRef] [PubMed]
- Taghizadeh, H.; Emamgholipour, S.; Hosseinkhani, S.; Arjmand, B.; Rezaei, N.; Dilmaghani-Marand, A.; Ghasemi, E.; Panahi, N.; Dehghanbanadaki, H.; Ghodssi-Ghassemabadi, R.; et al. The association between acylcarnitine and amino acids profile and metabolic syndrome and its components in Iranian adults: Data from STEPs 2016. Front Endocrinol (Lausanne) 2023, 14, 1058952. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.J.; Kwak, S.Y.; Jo, G.; Song, T.J.; Shin, M.J. Serum metabolite profile associated with incident type 2 diabetes in Koreans: findings from the Korean Genome and Epidemiology Study. Sci. Rep. 2018, 8, 8207. [Google Scholar] [CrossRef] [PubMed]
- Jang, C.; Oh, S.F.; Wada, S.; Rowe, G.C.; Liu, L.; Chan, M.C.; Rhee, J.; Hoshino, A.; Kim, B.; Ibrahim, A.; et al. A branched-chain amino acid metabolite drives vascular fatty acid transport and causes insulin resistance. Nat. Med. 2016, 22, 421–426. [Google Scholar] [CrossRef]
- Vanweert, F.; Schrauwen, P.; Phielix, E. Role of branched-chain amino acid metabolism in the pathogenesis of obesity and type 2 diabetes-related metabolic disturbances BCAA metabolism in type 2 diabetes. Nutr. Diabetes 2022, 12, 35. [Google Scholar] [CrossRef] [PubMed]
- Ottosson, F.; Smith, E.; Ericson, U.; Brunkwall, L.; Orho-Melander, M.; Di Somma, S.; Antonini, P.; Nilsson, P.M.; Fernandez, C.; Melander, O. Metabolome-Defined Obesity and the Risk of Future Type 2 Diabetes and Mortality. Diabetes Care 2022, 45, 1260–1267. [Google Scholar] [CrossRef] [PubMed]
- Kuhn, T.; Rohrmann, S.; Sookthai, D.; Johnson, T.; Katzke, V.; Kaaks, R.; von Eckardstein, A.; Muller, D. Intra-individual variation of plasma trimethylamine-N-oxide (TMAO), betaine and choline over 1 year. Clin. Chem. Lab. Med. 2017, 55, 261–268. [Google Scholar] [CrossRef]
- Kruger, R.; Merz, B.; Rist, M.J.; Ferrario, P.G.; Bub, A.; Kulling, S.E.; Watzl, B. Associations of current diet with plasma and urine TMAO in the KarMeN study: direct and indirect contributions. Mol. Nutr. Food Res. 2017, 61. [Google Scholar] [CrossRef]
- Birgitta Venho, S.V., Veli-Pekka Valkonen, Jyrki Virtanen, Timo A Lakka, Tiina H Rissanen, Marja-Leena Ovaskainen, Matti Laitinen, and Jukka T Salonen. Arginine intake, blood pressure, and the incidence of acute coronary events in men: the Kuopio Ischaemic Heart Disease Risk Factor Study. Am. J. Clin. Nutr. 2002, 359 - 364.
- Zell, J.A.; Taylor, T.H.; Albers, C.G.; Carmichael, J.C.; McLaren, C.E.; Wenzel, L.; Stamos, M.J. Phase IIa Clinical Biomarker Trial of Dietary Arginine Restriction and Aspirin in Colorectal Cancer Patients. Cancers (Basel) 2023, 15. [Google Scholar] [CrossRef]
- Li, J.; Cao, Y.F.; Sun, X.Y.; Han, L.; Li, S.N.; Gu, W.Q.; Song, M.; Jiang, C.T.; Yang, X.; Fang, Z.Z. Plasma tyrosine and its interaction with low high-density lipoprotein cholesterol and the risk of type 2 diabetes mellitus in Chinese. J. Diabetes Investig. 2019, 10, 491–498. [Google Scholar] [CrossRef]
- Chen, S.; Akter, S.; Kuwahara, K.; Matsushita, Y.; Nakagawa, T.; Konishi, M.; Honda, T.; Yamamoto, S.; Hayashi, T.; Noda, M.; et al. Serum amino acid profiles and risk of type 2 diabetes among Japanese adults in the Hitachi Health Study. Sci. Rep. 2019, 9, 7010. [Google Scholar] [CrossRef] [PubMed]
- Darst, B.F.; Koscik, R.L.; Hogan, K.J.; Johnson, S.C.; Engelman, C.D. Longitudinal plasma metabolomics of aging and sex. Aging 2019, 11, 1262–1282. [Google Scholar] [CrossRef]
- Talenezhad, N.; Mohammadi, M.; Ramezani-Jolfaie, N.; Mozaffari-Khosravi, H.; Salehi-Abargouei, A. Effects of l-carnitine supplementation on weight loss and body composition: A systematic review and meta-analysis of 37 randomized controlled clinical trials with dose-response analysis. Clin. Nutr. ESPEN 2020, 37, 9–23. [Google Scholar] [CrossRef]
- Adeva-Andany, M.M.; Calvo-Castro, I.; Fernandez-Fernandez, C.; Donapetry-Garcia, C.; Pedre-Pineiro, A.M. Significance of l-carnitine for human health. IUBMB Life 2017, 69, 578–594. [Google Scholar] [CrossRef] [PubMed]
- Schooneman, M.G.; Napolitano, A.; Houten, S.M.; Ambler, G.K.; Murgatroyd, P.R.; Miller, S.R.; Hollak, C.E.; Tan, C.Y.; Virtue, S.; Vidal-Puig, A.; et al. Assessment of plasma acylcarnitines before and after weight loss in obese subjects. Arch. Biochem. Biophys. 2016, 606, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Ceglarek, U.; Huang, T.; Li, L.; Rood, J.; Ryan, D.H.; Bray, G.A.; Sacks, F.M.; Schwarzfuchs, D.; Thiery, J.; et al. Weight-loss diets and 2-y changes in circulating amino acids in 2 randomized intervention trials. Am. J. Clin. Nutr. 2016, 103, 505–511. [Google Scholar] [CrossRef] [PubMed]
- Tochikubo, O.; Nakamura, H.; Jinzu, H.; Nagao, K.; Yoshida, H.; Kageyama, N.; Miyano, H. Weight loss is associated with plasma free amino acid alterations in subjects with metabolic syndrome. Nutr. Diabetes 2016, 6, e197. [Google Scholar] [CrossRef]
- Wiklund, P.K.; Pekkala, S.; Autio, R.; Munukka, E.; Xu, L.; Saltevo, J.; Cheng, S.; Kujala, U.M.; Alen, M.; Cheng, S. Serum metabolic profiles in overweight and obese women with and without metabolic syndrome. Diabetol. Metab. Syndr. 2014, 6, 40. [Google Scholar] [CrossRef]
- Chen, H.H.; Tseng, Y.J.; Wang, S.Y.; Tsai, Y.S.; Chang, C.S.; Kuo, T.C.; Yao, W.J.; Shieh, C.C.; Wu, C.H.; Kuo, P.H. The metabolome profiling and pathway analysis in metabolic healthy and abnormal obesity. Int J Obes (Lond) 2015, 39, 1241–1248. [Google Scholar] [CrossRef]
- Libert, D.M.; Nowacki, A.S.; Natowicz, M.R. Metabolomic analysis of obesity, metabolic syndrome, and type 2 diabetes: amino acid and acylcarnitine levels change along a spectrum of metabolic wellness. PeerJ 2018, 6, e5410. [Google Scholar] [CrossRef] [PubMed]
- Randrianarisoa, E.; Lehn-Stefan, A.; Wang, X.; Hoene, M.; Peter, A.; Heinzmann, S.S.; Zhao, X.; Konigsrainer, I.; Konigsrainer, A.; Balletshofer, B.; et al. Relationship of Serum Trimethylamine N-Oxide (TMAO) Levels with early Atherosclerosis in Humans. Sci. Rep. 2016, 6, 26745. [Google Scholar] [CrossRef] [PubMed]
- Xiong, X.; Zhou, J.; Fu, Q.; Xu, X.; Wei, S.; Yang, S.; Chen, B. The associations between TMAO-related metabolites and blood lipids and the potential impact of rosuvastatin therapy. Lipids Health Dis. 2022, 21, 60. [Google Scholar] [CrossRef] [PubMed]
- Fu, J.; Bonder, M.J.; Cenit, M.C.; Tigchelaar, E.F.; Maatman, A.; Dekens, J.A.; Brandsma, E.; Marczynska, J.; Imhann, F.; Weersma, R.K.; et al. The Gut Microbiome Contributes to a Substantial Proportion of the Variation in Blood Lipids. Circ. Res. 2015, 117, 817–824. [Google Scholar] [CrossRef] [PubMed]
- Beaumont, M.; Portune, K.J.; Steuer, N.; Lan, A.; Cerrudo, V.; Audebert, M.; Dumont, F.; Mancano, G.; Khodorova, N.; Andriamihaja, M.; et al. Quantity and source of dietary protein influence metabolite production by gut microbiota and rectal mucosa gene expression: a randomized, parallel, double-blind trial in overweight humans. Am. J. Clin. Nutr. 2017, 106, 1005–1019. [Google Scholar] [CrossRef] [PubMed]
- Bel Lassen, P.; Belda, E.; Prifti, E.; Dao, M.C.; Specque, F.; Henegar, C.; Rinaldi, L.; Wang, X.; Kennedy, S.P.; Zucker, J.D.; et al. Protein supplementation during an energy-restricted diet induces visceral fat loss and gut microbiota amino acid metabolism activation: a randomized trial. Sci. Rep. 2021, 11, 15620. [Google Scholar] [CrossRef] [PubMed]
- Warrier, M.; Shih, D.M.; Burrows, A.C.; Ferguson, D.; Gromovsky, A.D.; Brown, A.L.; Marshall, S.; McDaniel, A.; Schugar, R.C.; Wang, Z.; et al. The TMAO-Generating Enzyme Flavin Monooxygenase 3 Is a Central Regulator of Cholesterol Balance. Cell Rep. 2015, 10, 326–338. [Google Scholar] [CrossRef]
- Canyelles, M.; Tondo, M.; Cedo, L.; Farras, M.; Escola-Gil, J.C.; Blanco-Vaca, F. Trimethylamine N-Oxide: A Link among Diet, Gut Microbiota, Gene Regulation of Liver and Intestine Cholesterol Homeostasis and HDL Function. Int. J. Mol. Sci. 2018, 19. [Google Scholar] [CrossRef]
- Sepandi, M.; Abbaszadeh, S.; Qobady, S.; Taghdir, M. Effect of L-Arginine supplementation on lipid profiles and inflammatory markers: A systematic review and meta-analysis of randomized controlled trials. Pharmacol. Res. 2019, 148, 104407. [Google Scholar] [CrossRef]
- Cӑtoi, A.F.; Parvu, A.E.; Andreicut, A.D.; Mironiuc, A.; Crӑciun, A.; Cӑtoi, C.; Pop, I.D. Metabolically Healthy versus Unhealthy Morbidly Obese: Chronic Inflammation, Nitro-Oxidative Stress, and Insulin Resistance. Nutrients 2018, 10. [Google Scholar] [CrossRef]
- Yao, L.; Bhatta, A.; Xu, Z.; Chen, J.; Toque, H.A.; Chen, Y.; Xu, Y.; Bagi, Z.; Lucas, R.; Huo, Y.; et al. Obesity-induced vascular inflammation involves elevated arginase activity. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2017, 313, R560–R571. [Google Scholar] [CrossRef]
- Kovamees, O.; Shemyakin, A.; Eriksson, M.; Angelin, B.; Pernow, J. Arginase inhibition improves endothelial function in patients with familial hypercholesterolaemia irrespective of their cholesterol levels. J. Intern. Med. 2016, 279, 477–484. [Google Scholar] [CrossRef] [PubMed]
- Rui-Li Yang, Y.-H.S., Gang Hao, Wu Li, Guo-Wei Le. Increasing Oxidative Stress with Progressive Hyperlipidemia in Human: Relation between Malondialdehyde and Atherogenic Index. J. Clin. Biochem. Nutr. 2008, 154 - 158.
- Bian, F.; Cui, J.; Zheng, T.; Jin, S. Reactive oxygen species mediate angiotensin II-induced transcytosis of low-density lipoprotein across endothelial cells. Int. J. Mol. Med. 2017, 39, 629–635. [Google Scholar] [CrossRef]
- Asbaghi, O.; Kashkooli, S.; Amini, M.R.; Shahinfar, H.; Djafarian, K.; Clark, C.C.T.; Shab-Bidar, S. The effects of L-carnitine supplementation on lipid concentrations in patients with type 2 diabetes: A systematic review and meta-analysis of randomized clinical trials. J. Cardiovasc. Thorac. Res. 2020, 12, 246–255. [Google Scholar] [CrossRef] [PubMed]
- Musazadeh, V.; Alinejad, H.; Esfahani, N.K.; Kavyani, Z.; Keramati, M.; Roshanravan, N.; Mosharkesh, E.; Dehghan, P. The effect of L-carnitine supplementation on lipid profile in adults: an umbrella meta-analysis on interventional meta-analyses. Front. Nutr. 2023, 10, 1214734. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Yu, L.; Wang, Y.; Wei, Y.; Xu, Y.; He, T.; He, R. d-Ribose contributes to the glycation of serum protein. Biochim. Biophys. Acta Mol. Basis Dis. 2019, 1865, 2285–2292. [Google Scholar] [CrossRef] [PubMed]
- Schneider, A.M.; Rawat, D.; Weinstein, L.S.; Gupte, S.A.; Richards, W.O. Effects of laparoscopic Roux-en-Y gastric bypass on glucose-6 phosphate dehydrogenase activity in obese type 2 diabetics. Surgical Endoscopy 2011, 26, 823–830. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.J.; Choe, S.S.; Sohn, J.H.; Kim, J.B. The role of glucose-6-phosphate dehydrogenase in adipose tissue inflammation in obesity. Adipocyte 2017, 6, 147–153. [Google Scholar] [CrossRef]
- Ham, M.; Choe, S.S.; Shin, K.C.; Choi, G.; Kim, J.-W.; Noh, J.-R.; Kim, Y.-H.; Ryu, J.-w.; Yoon, K.-H.; Lee, C.-H.; et al. Glucose-6-Phosphate Dehydrogenase Deficiency Improves Insulin Resistance with Reduced Adipose Tissue Inflammation in Obesity. Diabetes 2016, 65, 2624–2638. [Google Scholar] [CrossRef]
- Jin, E.S.; Lee, M.H.; Murphy, R.E.; Malloy, C.R. Pentose phosphate pathway activity parallels lipogenesis but not antioxidant processes in rat liver. Am. J. Physiol. Endocrinol. Metab. 2018, 314, E543–E551. [Google Scholar] [CrossRef]
- Usman, M.; Woloshynowych, M.; Britto, J.C.; Bilkevic, I.; Glassar, B.; Chapman, S.; Ford-Adams, M.E.; Desai, A.; Bain, M.; Tewfik, I.; et al. Obesity, oxidative DNA damage and vitamin D as predictors of genomic instability in children and adolescents. International Journal of Obesity 2021, 45, 2095–2107. [Google Scholar] [CrossRef] [PubMed]
- Ge, T.; Yang, J.; Zhou, S.; Wang, Y.; Li, Y.; Tong, X. The Role of the Pentose Phosphate Pathway in Diabetes and Cancer. Frontiers in Endocrinology 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhao, H.; Liu, B.; Shu, H.; Zhang, L.; Bao, M.; Yi, W.; Tan, Y.; Ji, X.; Zhang, C.; et al. Human serum metabolomic analysis reveals progression for high blood pressure in type 2 diabetes mellitus. BMJ Open Diabetes Res. Care 2021, 9. [Google Scholar] [CrossRef] [PubMed]
Figure 2.
Metabolites significantly changed between MHO and MUO following intervention.
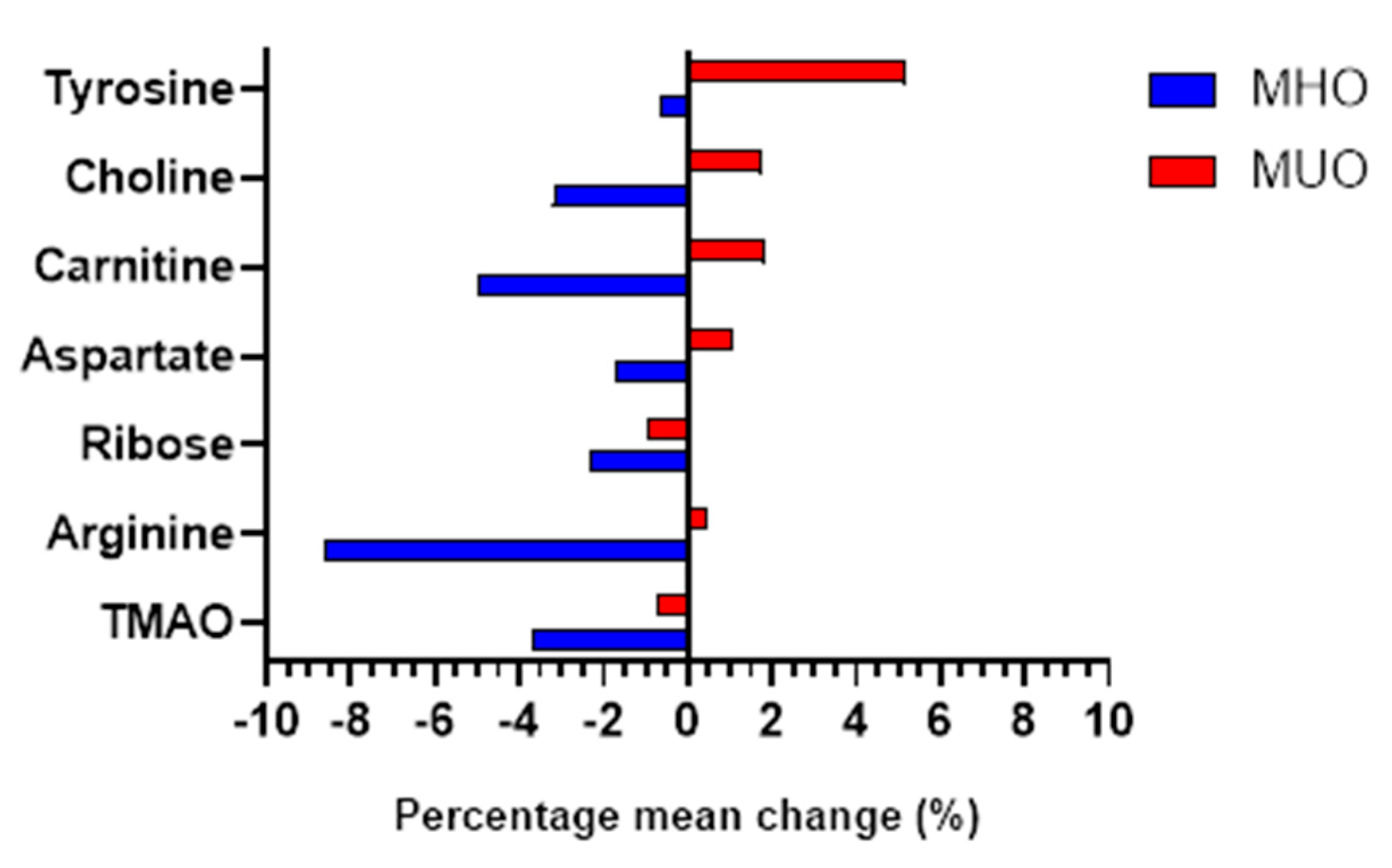
Figure 3.
Metabolic pathways affected at sixth month.
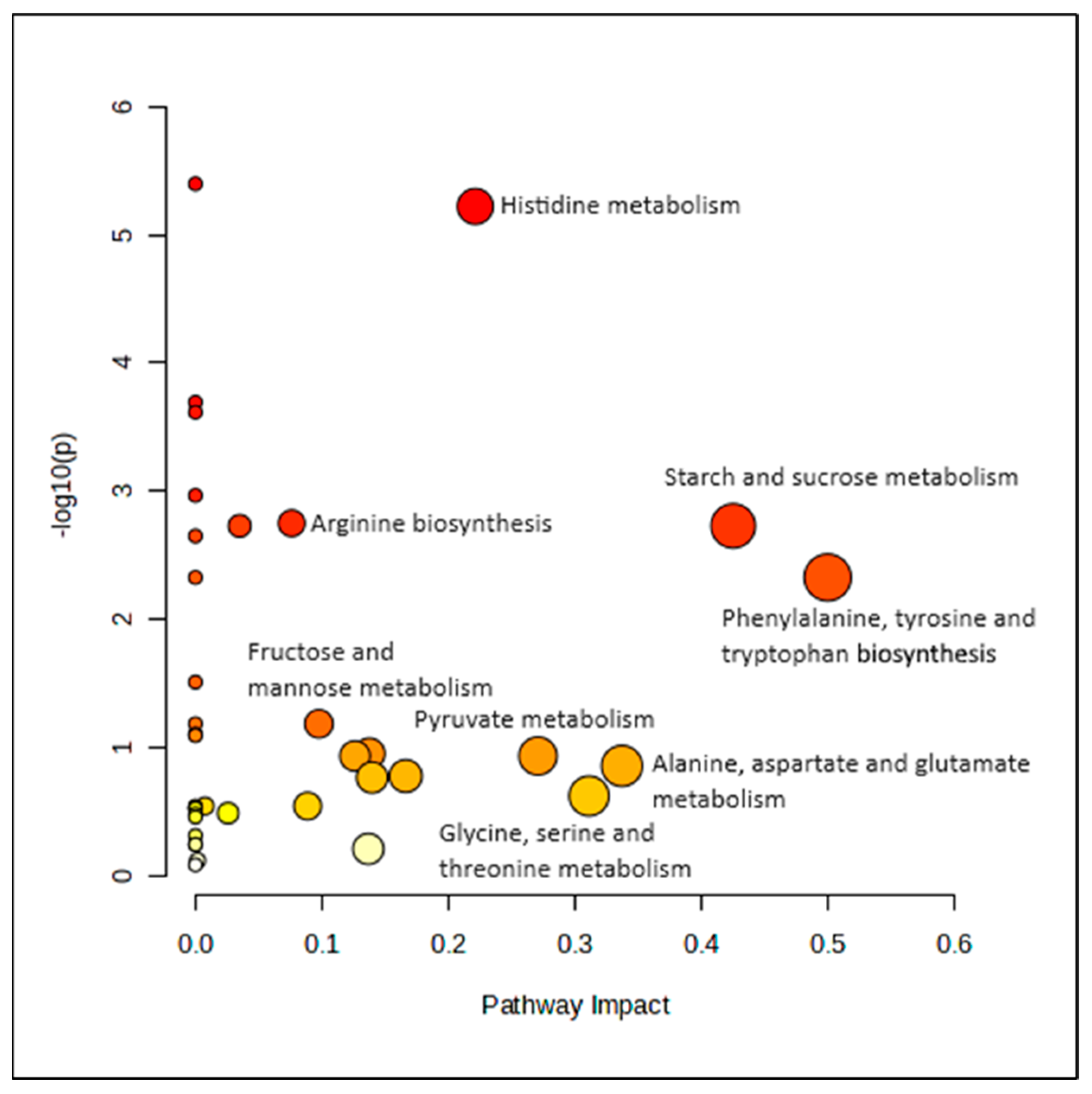
Figure 4.
Heatmap of correlation between the significantly changed metabolites with body weight, BMI, WC and systolic blood pressure in MUO. Correlation determined using Pearson correlation. Green indicates positive correlation, while red indicates negative correlation.
Figure 4.
Heatmap of correlation between the significantly changed metabolites with body weight, BMI, WC and systolic blood pressure in MUO. Correlation determined using Pearson correlation. Green indicates positive correlation, while red indicates negative correlation.
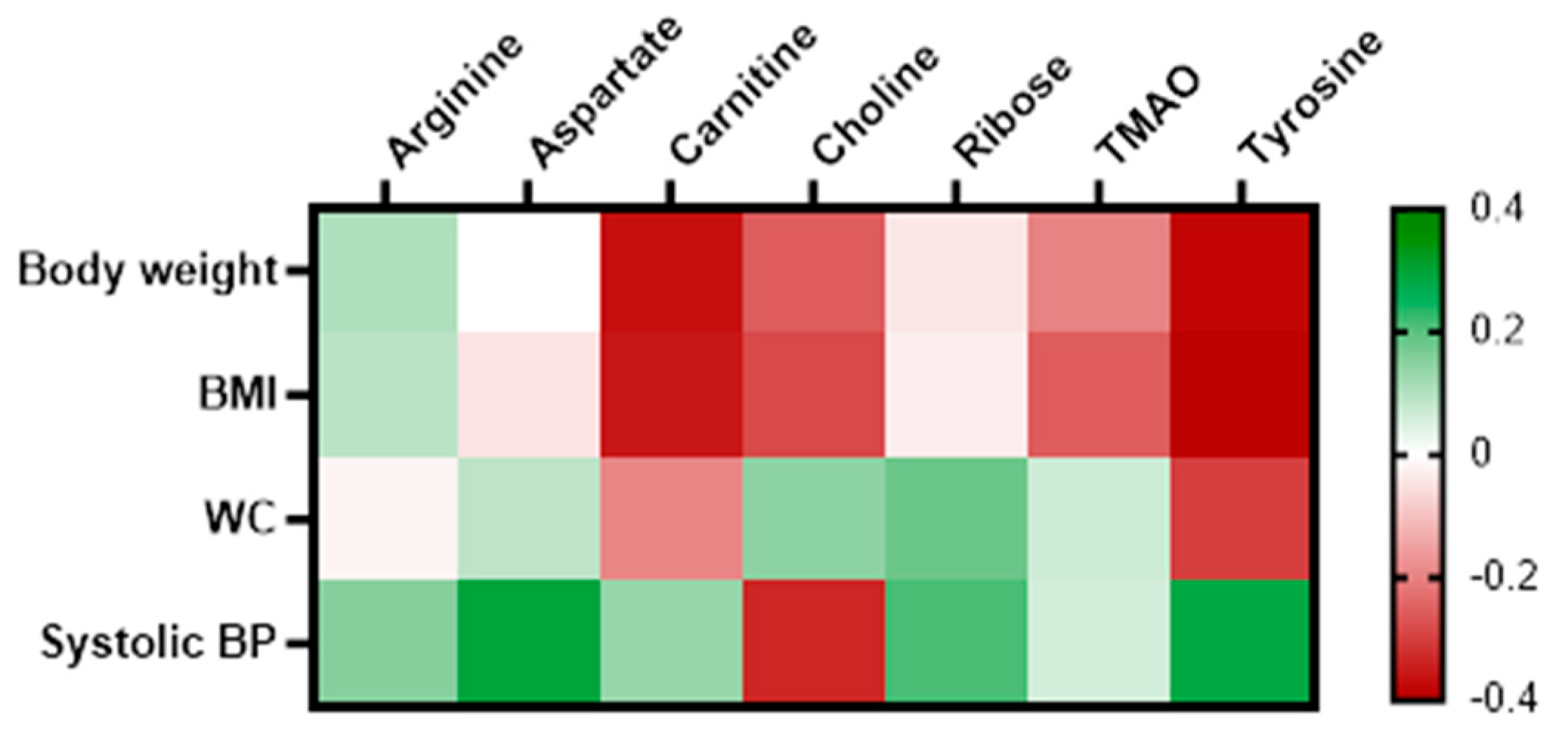
Figure 5.
Heatmap of correlation between the significantly changed metabolites with changes in anthropometry, clinical and biochemical variable. Correlation determined using Pearson correlation analysis in both groups combined. Blue indicates positive correlation, while red indicates negative correlation.
Figure 5.
Heatmap of correlation between the significantly changed metabolites with changes in anthropometry, clinical and biochemical variable. Correlation determined using Pearson correlation analysis in both groups combined. Blue indicates positive correlation, while red indicates negative correlation.
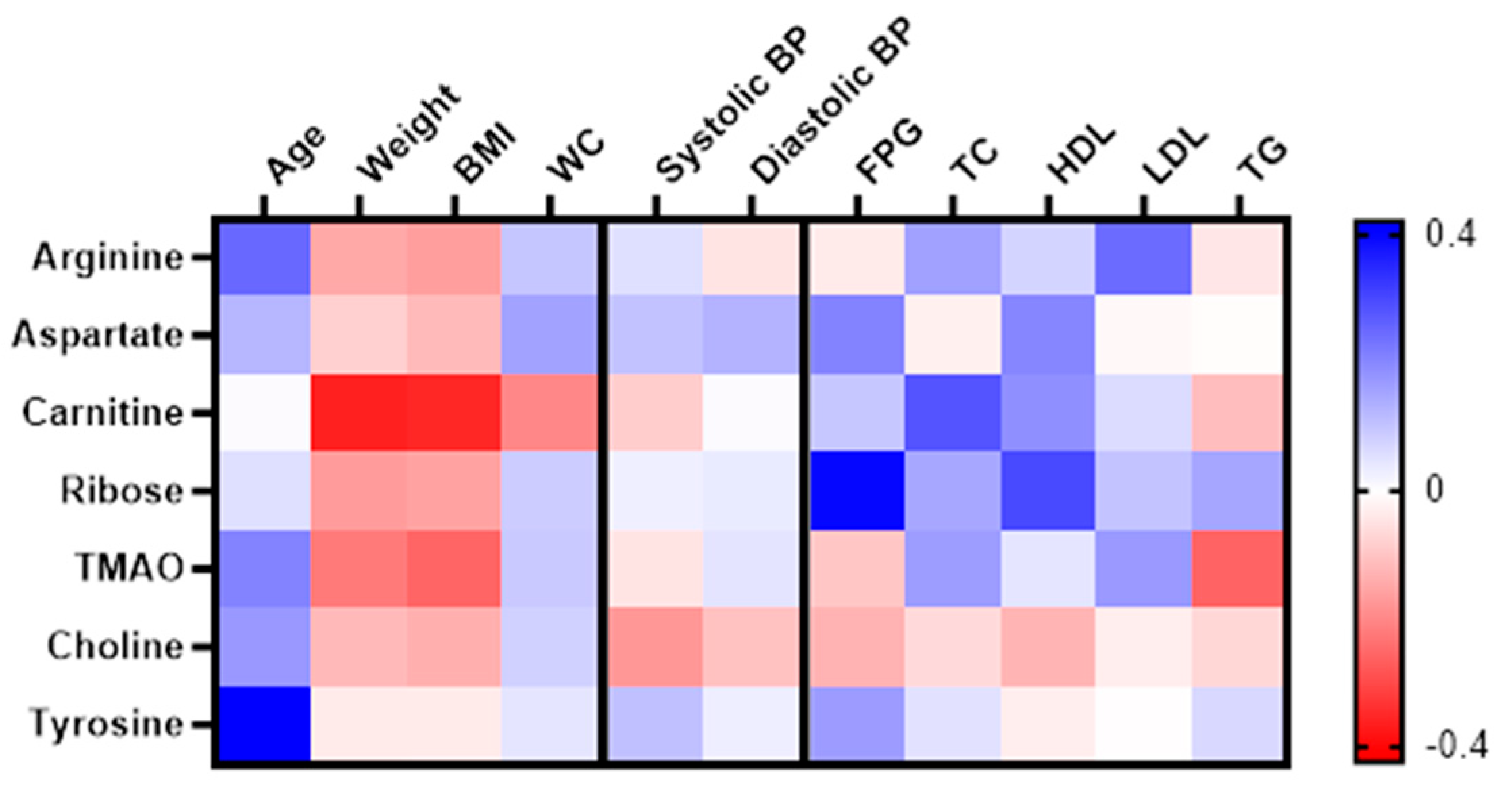

Table 1.
Demographic, anthropometric and clinical characteristics of the participants at baseline.
| MHO | MUO | p | |||
|---|---|---|---|---|---|
| n=36 | n=34 | ||||
| Age (year), mean ± SD | 41.65 ± 8.46 | 50.01 ± 6.26 | <0.001* | ||
| Age group (year), n, % | |||||
| 18 - 39 | 12 | 33.3 | 3 | 8.8 | 0.004* |
| 40 -49 | 18 | 50 | 14 | 41.2 | |
| ≥ 50 | 6 | 16.7 | 17 | 50 | |
| Race, n, % | |||||
| Malay | 33 | 91.7 | 30 | 88.2 | 0.706 |
| Indian | 3 | 8.3 | 4 | 11.8 | |
| Level of education, n, % | |||||
| Primary school | 3 | 8.6 | 13 | 39.4 | |
| Secondary school/ tertiary education | 32 | 91.4 | 20 | 60.6 | 0.003* |
| Household income (RM), n, % | |||||
| ≤ 1500 | 12 | 33.3 | 23 | 67.6 | |
| 1501 - 2500 | 15 | 41.7 | 7 | 20.6 | 0.016* |
| ≥ 2501 | 9 | 25 | 4 | 11.8 | |
| Family history, n, % | |||||
| Diabetes | 11 | 31.4 | 15 | 44.1 | 0.326 |
| Hypertension | 16 | 44.4 | 20 | 60.6 | 0.230 |
| Cardiovascular diseases | 3 | 8.3 | 6 | 18.2 | 0.294 |
| Body weight (kg) | 71.20 ± 11.39 | 75.32 ± 10.27 | 0.117 | ||
| BMI (kg/m2) | 29.66 ± 4.05 | 31.64 ± 3.45 | 0.031* | ||
| Waist circumference (cm) | 91.07 ± 9.11 | 96.93 ± 7.94 | 0.006* | ||
| Systolic BP (mmHg) | 113.76 ± 11.67 | 133.06 ± 20.13 | <0.001* | ||
| Diastolic BP (mmHg) | 73.06 ± 10.11 | 84.02 ± 12.98 | <0.001* | ||
| FPG (mmol/L) | 5.19 ± 0.55 | 6.21 ± 1.63 | 0.001* | ||
| HbA1c (%) | 5.35 ± 0.55 | 6.05 ± 1.05 | 0.002* | ||
| TC (mmol/L) | 5.44 ± 1.18 | 5.30 ± 0.89 | 0.590 | ||
| HDL-C (mmol/L) | 1.31 ± 0.30 | 1.32 ± 0.22 | 0.888 | ||
| LDL-C (mmol/L) | 3.95 ± 1.46 | 3.98 ± 0.87 | 0.922 | ||
| Triglyceride (TG) (mmol/L) | 1.15 ± 0.56 | 1.35 ± 0.47 | 0.129 | ||
Normally distributed variables are presented as mean (standard deviation) (SD) and categorical variables are presented as count (percentage). P-values were determined with independent t-test for continuous variables and Pearson’s Chi Square test for categorical variables. *Significant at p-value < 0.05. Abbreviation - MHO: Metabolically Healthy Obese, MUO: Metabolically Unhealthy Obese, kg: kilogram, cm: centimeters, kg/m2: kilogram per meter square, BMI: Body Mass Index, BP: Blood Pressure, FPG: Fasting Plasma Glucose, TC: Total Cholesterol, HDL-C: High Density Lipoprotein-Cholesterol, LDL-C: Low Density Lipoprotein-Cholesterol.
Table 2.
Anthropometry, clinical and dietary intake changes between MHO and MUO 6 months post-intervention.
Table 2.
Anthropometry, clinical and dietary intake changes between MHO and MUO 6 months post-intervention.
| Variables | Group | Estimated marginal means (95% CI) | Mean difference (%) |
Within group | Between group | ||
|---|---|---|---|---|---|---|---|
| Baseline | 6th month | p | Wald Chi-Square | p | |||
| Body weight (kg) | MHO | 69.68 (65.41, 73.94) | 69.52 (65.24, 73.80) | -0.156 (-0.23) | 0.719 | 8.125 | 0 .043 |
| MUO | 72.64 (68.79, 76.50) | 71.72 (67.93, 75.51) | -0.926 (-1.30) | 0.015 | |||
| BMI | MHO | 30.78 (30.52, 31.03) | 30.74 (30.33, 31.16) | -0.034 (-0.11) | 0.853 | 5.464 | 0.141 |
| MUO | 30.79 (30.54, 31.04) | 30.43 (30.06, 30.79) | -0.362 (-1.17) | 0.023 | |||
| WC (cm) | MHO | 90.01 (85.67, 94.35) | 92.77 (88.74, 96.81) | 2.761 (3.05) | 0.483 | 12.306 | 0 .006 |
| MUO | 93.18 (91.64, 94.73) | 90.86 (89.06, 92.67) | -2.322 (-2.48) | <0.001 | |||
| Systolic BP (mmHg) | MHO | 117.52 (111.38, 123.66) | 121.00 (111.45, 127.56) | 3.486 (2.91) | 0.105 | 7.321 | 0.062 |
| MUO | 123.95 (119.64, 128.27) | 114.65 (104.69, 124.61) | -9.309 (-7.36) | 0.049 | |||
| Diastolic BP (mmHg) | MHO | 75.24 (69.39, 81.09) | 76.57 (69.81, 83.34) | 1.333 (1.76) | 0.523 | 4.629 | 0.201 |
| MUO | 80.43 (75.43, 85.43) | 76.68 (68.91, 84.45) | -3.750 (-4.63) | 0.226 | |||
| Glucose (mmol/L) | MHO | 6.44 (5.75, 7.54) | 6.51 (5.94, 7.08) | -0.139 (-2.16) | 0.679 | 0.268 | 0.966 |
| MUO | 6.53 (5.69, 7.37) | 6.43 (5.48, 7.37) | -0.101 (-1.61) | 0.847 | |||
| TC (mmol/L) | MHO | 5.91 (4.65, 7.16) | 6.02 (5.17, 6.88) | 0.113 (1.93) | 0.802 | 0.444 | 0.931 |
| MUO | 6.14 (5.26, 7.03) | 6.21 (5.34, 7.07) | 0.064 (1.05) | 0.676 | |||
| HDL (mmol/L) | MHO | 1.64 (1.16, 2.13) | 1.51 (1.30, 1.72) | -0.130 (-8.18) | 0.518 | 3.656 | 0.301 |
| MUO | 1.49 (1.21, 1.77) | 1.54 (1.26, 1.82) | 0.053 (3.71) | 0.089 | |||
| LDL (mmol/L) | MHO | 4.66 (3.53, 5.78) | 4.96 (4.03, 5.89) | 0.305 (6.56) | 0.378 | 1.515 | 0.679 |
| MUO | 4.79 (3.98, 5.61) | 4.93 (4.19, 5.67) | 0.134 (2.80) | 0.461 | |||
| TG (mmol/L) | MHO | 2.18 (1.59, 2.77) | 1.95 (1.57, 2.33) | -0.229 (-10.13) | 0.358 | 3.84 | 0.279 |
| MUO | 1.70 (1.32, 2.09) | 1.77 (1.31, 2.23) | 0.062 (3.46) | 0.539 | |||
| Calorie intake | MHO | 1559.99 (1308.06, 1811.93) | 1148.70 (915.12, 1382.28) | -411.29 (-26.36) | <0.001 | 17.287 | <0.001 |
| MUO | 1315.36 (1105.33, 1525.39) | 1082.19 (855.40, 1308.98) | -233.17 (-17.73) | 0.014 | |||
| Carbohydrate | MHO | 201.41 (167.84, 234.97) | 152.50 (118.03, 186.98) | -48.90 (-24.28) | 0.005 | 12.353 | 0.006 |
| MUO | 169.12 (140.16, 198.08) | 142.85 (109.04, 176.66) | -26.27 (-15.53) | 0.046 | |||
| Cholesterol | MHO | 224.64 (173.93, 275.34) | 139.40 (83.97, 194.84) | -85.23 (-37.94) | 0.007 | 11.117 | 0.011 |
| MUO | 168.70 (125.87, 211.53) | 139.96 (93.60, 186.32) | -28.74 (-17.04) | 0.066 | |||
| Dietary fiber | MHO | 8.13 (5.11, 11.15) | 5.39 (2.85, 7.92) | -2.74 (-33.70) | 0.011 | 11.429 | 0.01 |
| MUO | 6.95 (4.41, 9.48) | 4.82 (2.16, 7.48) | -2.13 (-30.65) | 0.036 | |||
| Potassium | MHO | 1057.04 (836.51, 1277.57) | 846.48 (619.61, 1073.35) | -210.56 (-19.92) | 0.023 | 11.097 | 0.011 |
| MUO | 969.53 (778.77, 1160.29) | 773.54 (564.77, 982.31) | -195.99 (-20.21) | 0.016 | |||
| Protein | MHO | 60.37 (49.24, 71.49) | 43.84 (32.56, 55.12) | -16.53 (-27.38) | 0.003 | 19.53 | <0.001 |
| MUO | 54.29 (44.73, 63.84) | 40.25 (30.23, 50.28) | -14.04 (-25.86) | 0.001 | |||
| Saturated fat | MHO | 15.32 (11.65, 19.00) | 11.75 (8.09, 15.40) | -3.58 (-23.37) | 0.024 | 15.063 | 0.002 |
| MUO | 11.93 (9.10, 14.75) | 8.57 (5.18, 11.96) | -3.36 (-28.16) | 0.009 | |||
| Sodium | MHO | 2188.44 (1748.27, 2628.61) | 1685.29 (1314.10, 2056.48) | -503.15 (-22.99) | 0.021 | 8.721 | 0.033 |
| MUO | 1441.14 (1044.33, 1837.95) | 1370.37 (907.81, 1832.93) | -70.77 (-4.91) | 0.643 | |||
| Total fat | MHO | 58.76 (46.94, 70.58) | 41.61 (30.99, 52.22) | -17.16 (-29.20) | <0.001 | 16.557 | <0.001 |
| MUO | 46.12 (36.83, 55.40) | 37.76 (28.56, 46.95) | -8.36 (-18.13) | 0.038 | |||
Data presented as estimated marginal mean (95% CI). p<0.05 was considered significant after GEE analysis in which the analysis was adjusted for age, education level, household income, medication use, baseline BMI, WC, FPG, HbA1c and systolic blood pressure.
Table 3.
Serum metabolites resolved by 1H NMR at baseline and sixth month in MHO and MUO groups.
| Metabolites | MHO (n=36) | MUO (n=34) | p |
|---|---|---|---|
| Baseline | |||
| Glucose | 4.16 ± 0.06 | 4.25 ± 0.14 | 0.001 |
| Indole-3-acetate | 2.67 ± 0.16 | 2.75 ± 0.20 | 0.047 |
| τ-methylhistidine | 2.73 ± 0.15 | 2.82 ± 0.16 | 0.021 |
| 6th month | |||
| Acetate | 2.58 ± 0.26 | 2.71 ± 0.27 | 0.030 |
| Arginine | 2.57 ± 0.31 | 2.75 ± 0.27 | 0.014 |
| Aspartate | 3.00 ± 0.21 | 3.15 ± 0.11 | <0.001 |
| Betaine | 2.73 ± 0.12 | 2.85 ± 0.16 | 0.001 |
| Fructose | 3.49 ± 0.08 | 3.44 ± 0.08 | 0.044 |
| Glucose | 4.15 ± 0.08 | 4.23 ± 0.12 | 0.002 |
| Histidine | 2.77 ± 0.19 | 2.92 ± 0.12 | <0.001 |
| Isobutyrate | 2.69 ± 0.12 | 2.77 ± 0.17 | 0.025 |
| Isoleucine | 2.87 ± 0.19 | 2.98 ± 0.24 | 0.031 |
| Leucine | 2.67 ± 0.26 | 2.82 ± 0.35 | 0.049 |
| N-acetylcysteine | 2.63 ± 0.19 | 2.72 ± 0.18 | 0.030 |
| Phenylacetate | 2.82 ± 0.21 | 2.94 ± 0.25 | 0.024 |
| TMAO | 2.66 ± 0.12 | 2.77 ± 0.14 | 0.001 |
| Tyrosine | 3.12 ± 0.19 | 3.24 ± 0.18 | 0.014 |
| Valine | 2.93 ± 0.17 | 3.02 ± 0.18 | 0.037 |
Values are reported as mean ± SD (μM). Significant is at p<0.05 based on independent T-test.
Table 4.
Summary of the significantly changed metabolites.
| Metabolites | Group | Baseline | 6th month | Mean difference (%) |
Within group | Between group |
|---|---|---|---|---|---|---|
| p | p | |||||
| TMAO | MHO | 2.80 (2.72, 2.88) | 2.70 (2.62, 2.77) | -0.103 (-3.68) | <0.001 | < 0.001 |
| MUO | 2.79 (2.71, 2.87) | 2.77 (2.69, 2.85) | -0.021(-0.75) | 0.387 | ||
| Arginine | MHO | 2.89 (2.76, 3.02) | 2.64 (2.51, 2.77) | -0.249 (-8.62) | <0.001 | 0.002 |
| MUO | 2.80 (2.67, 2.94) | 2.81 (2.69, 2.94) | 0.012 (0.43) | 0.835 | ||
| Ribose | MHO | 3.64 (3.56, 3.71) | 3.55 (3.48, 3.62) | -0.084 (-2.31) | 0.008 | 0.002 |
| MUO | 3.67 (3.61, 3.72) | 3.63 (3.57, 3.70) | -0.034 (-0.93) | 0.079 | ||
| Aspartate | MHO | 3.09 (2.97, 3.21) | 3.04 (2.93, 3.15) | -0.054 (-1.75) | 0.152 | 0.005 |
| MUO | 3.20 (3.11, 3.28) | 3.23 (3.15, 3.31) | 0.034 (1.06) | 0.142 | ||
| Carnitine | MHO | 2.78 (2.68, 2.89) | 2.65 (2.53, 2.77) | -0.138 (-4.96) | 0.001 | 0.005 |
| MUO | 2.66 (2.54, 2.78) | 2.71 (2.60, 2.82) | 0.049 (1.84) | 0.301 | ||
| Choline | MHO | 2.75 (2.62, 2.88) | 2.67 (2.54, 2.79) | -0.088 (-3.20) | 0.066 | 0.006 |
| MUO | 2.55 (2.43, 2.67) | 2.60 (2.47, 2.73) | 0.045 (1.76) | 0.432 | ||
| Tyrosine | MHO | 2.93 (2.75, 3.10) | 2.91 (2.73, 3.09) | -0.019 (-0.65) | 0.821 | 0.008 |
| MUO | 3.08 (3.92, 3.25) | 3.24 (3.10, 3.38) | 0.159 (5.16) | 0.016 |
Metabolites listed according to the rank of p between group. Data presented as estimated marginal mean (95% CI) (µM). Significance is at p<0.05 after GEE analysis in which the analysis was adjusted for age, education level, household income, baseline BMI, WC, FPG, HbA1c, systolic blood pressure, medication use, and baseline dietary intake (energy, carbohydrate, protein, total fat, saturated fat, sodium, potassium, and dietary fiber).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



