
Submitted:
24 July 2024
Posted:
26 July 2024
You are already at the latest version
Abstract
Background: Approximately 10-20% of subjects vaccinated with HBsAg-based hepatitis B virus (HBV) vaccines are non-responders. BM32 is a recombinant grass pollen allergy vaccine containing the HBV-derived preS surface antigen as immunological carrier protein. PreS includes the binding site of HBV to its receptor on hepatocytes. We investigated if immunological non-responsiveness to HBV after repeated HBsAg-based vaccinations can be overcome by immunization with VVX001 (i.e., Alum-adsorbed BM325, a component of BM32). Methods: A subject failing to develop protective HBV-specific immunity after HBsAg-based vaccination received five monthly injections of 20 µg VVX001. PreS-specific antibody responses were measured by ELISA and micro-array technology. Serum reactivity to subviral particles of different HBV genotypes was determined by sandwich ELISA. PreS-specific T cell responses were monitored by CFSE staining and subsequent FACS analysis. HBV neutralization was assessed using cultured HBV-infected HepG2 cells. Results: Vaccination with VVX001 induced a strong and sustained preS-specific antibody response composed mainly of the IgG1 subclass. PreS-specific IgG antibodies were primarily directed to the N-terminal part of preS containing the NTCP attachment site. IgG reactivity to sub-viral particles as well as to the N-terminal preS-derived peptides was comparable for HBV genotypes A-H. A pronounced reactivity of CD3+CD4+ lymphocytes specific for preS after the complete injection course remaining up to one year after the last injection was found. Maximal HBV neutralization (98.4%) in vitro was achieved 1 month after the last injection which correlated with the maximal IgG reactivity to the N-terminal part of preS. Conclusion: Our data suggest that VVX001 may be used as a preventive vaccination against HBV even in non-responders to HBsAg-based HBV vaccines.
Keywords:
HBV vaccines
; preS surface antigen
; HBsAg
; non-responsiveness
1. Introduction
Hepatitis B virus (HBV) causes acute and chronic liver infections which often lead to long-term complications such as cirrhosis and liver cancer. Accordingly, HBV is one of the major public health threats. As reported by the WHO, there were 1.2 million of new HBV infections in 2022 and the estimated number of deaths caused by HBV was around 1.1 million. Ongoing chronic HBV infection is a main driver for hepatocellular carcinoma which represents 80% of all liver cancer cases and is the third most common cause of cancer deaths [1]. Vaccination against HBV was introduced as the most important tool for the control of HBV morbidity forty years ago, starting with the first generation of vaccines derived from plasma obtained from HBV-infected donors. Later, second-generation vaccines based on recombinant HBV surface antigen (HBsAg) produced in yeast became available and currently remain the basis for the internationally-accepted HBV immunization program which resulted in significant decrease of the hepatitis B burden in a number of countries with high vaccine coverage. However, at least 10% of vaccinees fail to mount a protective anti-HBs response, and the rate of non-responsiveness appears to be largely underestimated, since a significant proportion of the population has one or more low-response risk factors [2,3]. Thus, an alternative to the HBsAg-based standard (e.g. Engerix, Twinrix) or alternative (e.g. Fendrix, Heblisav) vaccines is needed, especially for individuals at high risk of HBV exposure.
Protection against HBV can be achieved by inducing antibodies against the viral envelope consisting of the three surface proteins encoded in one open reading frame (S-ORF) with different start codons: large hepatitis B surface protein (LHBs) composed of preS1, preS2 and S domains; middle hepatitis B surface protein (MHBs) composed of preS2 and S domains; and small hepatitis B surface protein (SHBs,) consisting of the S domain only. The term HBV surface antigen (HBsAg) is used to describe reactivity against the highly antigenic loop (AGL) of the S-domain (anti-HBs), especially for vaccines containing only SHBs (e.g. Engerix/Twinrix). While all three proteins serve as the fundamental components of the viral envelope, they also constitute the building blocks of subviral particles (SVP). These non-infectious particles are produced in surplus from HBV-infected cells and contain only the three surface proteins, with varying quantities. The surface protein domains preS (preS1 + preS2) and S are both involved in HBV entry to liver cells and therefore are reasonable vaccine candidates. Of those, the S domain, responsible for the initial low-specificity attachment to heparan sulphate proteoglycans, was discovered first and became the basis for conventional vaccination preventing the initial step of HBV entry [4]. It was reported that preS2 binds to polymerized human albumin and several studies have shown the virus neutralizing effect of anti-preS2 antibodies, however it was later concluded that MHBs and a specific sequence of preS2 is not essential for HBV infectivity [2]. As discovered later, preS1 and in particular its myristoylated N-terminus mediates the highly-specific binding to sodium taurocholate co-transporting polypeptide (NTCP), a bile salt transporter on hepatocytes, and contains highly conserved amino acids essential for infection. Thus, the preS1-NTCP interaction represents a potent target for HBV entry inhibition by blocking NTCP with, e.g., preS1-derived peptides or blocking the infectious viral particles with preS1-specific antibodies by passive or active immunization [4].
BM32 is a recombinant grass pollen allergy vaccine based on the peptide-carrier technology, consisting of preS (preS1 + preS2, GenBank: AAT28735) protein as an immunological carrier decorated with hypoallergenic peptides on both N- and C-terminus. Designed initially for allergen-specific immunotherapy of grass pollen allergy, BM32 has four compounds: BM321, BM322, BM325 and BM326 containing hypoallergenic peptides derived from major timothy grass pollen allergens Phl p 1, Phl p 2, Phl p 5 and Phl p 6, respectively [5]. Besides hypoallergenic activity, good safety profile, induction of allergen-specific blocking antibodies and clinical efficacy for treating grass pollen-induced rhinitis shown in a series of clinical trials (ClinicalTrials.gov IDs: NCT01350635, NCT01445002, NCT01538979, NCT02643641), BM32 elicited preS-specific antibodies, which are directed mainly against the N-terminal part of preS1 and neutralized HBV in vitro thus posing BM32 as an alternative HBV vaccine candidate, especially for non-responders to the standard HBsAg-based vaccination [6,7]. In this report we show that the lack of anti-HBV protection can be overcome by immunization with VVX001, a vaccine containing Alumn-adsorbed BM325, in a subject with a long history of unsuccessful vaccinations with HBsAg-based vaccines.
2. Materials and Methods
2.1. Vaccines and Immunization Schedule
The subject of the present report (male, born on 25.02.1963, medical doctor and researcher) has been vaccinated with licensed HBsAg-based vaccines six times as described in Figure 1. The basic immunization was performed with the plasma-derived vaccine Hevac B Pasteur [8]. Subsequent booster injections were combined hepatitis A and B vaccine Twinrix (20 µg HBsAg, yeast-derived, GlaxoSmithKline Biologicals, Belgium) and Engerix-B vaccine (20 µg HBsAg, yeast-derived, GlaxoSmithKline). After the last immunization with Engerix-B on 08.02.2018, anti-HBs levels were monitored at 1, 2, 4, 6, 9 months after vaccination followed by self-immunization with VVX001 as shown in Figure 2a.
BM325, a component of BM32 vaccine, is a recombinant fusion protein composed of preS (HBV genotype A2, GenBank: AAT28735) of LHBs and peptides derived from the grass pollen allergen Phl p 5. BM325 was produced by Biomay AG (Austria) according to GMP standards, aluminum hydroxide was used as adjuvant [5]. The vaccine VVX001 containing Alum-adsorbed BM325 was applied by the subject himself subcutaneously (20 µg per injection) in the non-dominant upper arm (deltoid muscle region) at each of the following time points: 19.11.2018, 14.12.2018, 11.01.2019, 07.02.2019, 08.03.2019. Blood samples were obtained at the time points depicted in Figure 2a.
2.2. Serological Assays
Anti-HBs antibody levels were determined using the Architect anti-HBs quantitative assay (Abbott Laboratories, Germany). Recombinant preS (preS1 + preS2) and preS-derived synthetic peptides spanning the whole preS sequence of HBV genotype A2 (GenBank: AAT28735), as well as peptides covering the NTCP attachment site of genotypes A-H (Figure S1) were produced as described [7]. The preS- and peptide-specific IgG, IgG1-4, IgM, IgA determination as well as IgG1 and IgG4 quantification were performed by enzyme-linked immunosorbent assay (ELISA) as described in [7]. If not stated otherwise, plots and charts were created using Prism 6.0 software (GraphPad, CA, USA).

Table S1.
Lists of synthetic preS-derived peptides. Shown are the amino acid sequences of the peptides used in the assessment of preS-specific immune response.
Table S1.
Lists of synthetic preS-derived peptides. Shown are the amino acid sequences of the peptides used in the assessment of preS-specific immune response.
| preS-derived peptides | Accession№. | Sequence |
| Peptides mapping the N-terminal epitopes of preS | ||
| Peptide A (aa 1-29) Peptide B (aa 30-61) Peptide C (aa 17-51) |
AAT28735 | CMGGWSSKPRKGMGTNLSVPNPLGFFPDHQ |
| CLDPAFGANSNNPDWDFNPIKDHWPAANQVGVG | ||
| CSVPNPLGFFPDHQLDPAFGANSNNPDWDFNPIKDH | ||
| Overlapping peptides spanning the whole preS sequence | ||
| P1 (aa 2-31) P2 (aa 22-51) P3 (aa 42-71) P4 (aa 62-91) P5 (aa 82-111) P6 (aa 102-131) P7 (aa 122-151) P8 (aa 142-174) |
AAT28735 | GGWSSKPRKGMGTNLSVPNPLGFFPDHQLD |
| LGFFPDHQLDPAFGANSNNPDWDFNPIKDH DWDFNPIKDHWPAANQVGVGAFGPGLTPPH AFGPGLTPPHGGILGWSPQAQGILTTVSTI QGILTTVSTIPPPASTNRQSGRQPTPISPP GRQPTPISPPLRDSHPQAMQWNSTAFHQAL WNSTAFHQALQDPRVRGLYFPAGGSSSGTV PAGGSSSGTVNPAPNIASHISSISARTGDPVTN | ||
| Peptides covering the NTCP attachment site of genotypes A-H | ||
| Genotype A Genotype B Genotype C Genotype D Genotype E Genotype F Genotype G Genotype H |
APD28359 | GTNLSVPNPLGFFPDHQLDPAFGANSNNPDWDFNPIKDH |
| BAA88276 | GTNLSVPNPLGFFPDHQLDPAFKANSENPDWDLNPHKDN | |
| BAA32833 | GTNLSVPNPLGFFPDHQLDPAFGANSNNPDWDFNPNKDH | |
| BAD02320 BAC65105 AAG49720 BAB64320 BAB69786 |
GQNLSTSNPLGFFPDHQLDPAFRANTANPDWDFNPNKDT GKNHSTTNPLGFFPDHQLDPAFRANTRNPDWDHNPNKDH GQNLSVPNPLGFFPDHQLDPLFRANSSSPDWDFNKNKDN GKNLSTSNPLGFLPDHQLDPAFRANTNNPDWDFNPKKDP GQNLSVPNPLGFFPDHQLDPLFRANSSSPDWDFNTNKDN |
|
2.3. preS Micro-Array Production and Sample Analysis
Glass slides containing 6 micro-arrays formed by oval epoxy frames (Paul Marienfeld GmbH & Co. KG, Germany) were coated with amino-reacting polymer MCP-2 (Lucidant Polymers, CA, USA). The recombinant preS and preS-derived synthetic peptides were diluted and re-buffered for the following spotting conditions: concentration 0.5-1 mg/mL, buffer 75 mM Na2HPO4, pH 8.4. Afterwards they were spotted in triplicates on the pre-activated glass slides with a SciFlexArrayer S12 (Scienion AG, Germany) as described in [9]. The micro-array layout is depicted in Figure S1.
Figure S1.
Layout and exemplary measurements of the preS micro-array. (a) Spotting scheme of a preS micro-array showing the order of dots (triplicates). (b, c) Scanned images depicting fluorescence of the (b) pre-immune and (c) and a post-vaccination serum sample.
Figure S1.
Layout and exemplary measurements of the preS micro-array. (a) Spotting scheme of a preS micro-array showing the order of dots (triplicates). (b, c) Scanned images depicting fluorescence of the (b) pre-immune and (c) and a post-vaccination serum sample.

For specific antibody detection, micro-arrays were washed for 1 min with phosphate-buffered saline containing 1% Tween 20 (PBST) and dried with a Sigma centrifuge and MTP-11113 rotor (both Sigma Laborzentrifugen GmbH, Germany). For IgG, IgG1, IgG4, IgM detection serum was diluted 1:100 and for IgA detection 1:20 with a sample diluent (ImmunoCAP Specific IgA/IgG Sample Diluent, Phadia Ab, Sweden). Aliquots of 30 µL were added per array and incubated for 2 h at 22°C. After another washing step, corresponding detection antibodies labelled with DyLight 550 (Cat. 62269, Pierce, IL, USA) with concentration 1.8 µg/mL targeting various types of immunoglobulins were applied: IgG (Cat. 009-000-003, Jackson ImmunoResearch, PA, USA), IgG1 (Cat.555868, BD Biosciences, NJ, USA), IgG4 (Cat. 555878, BD), IgA1/A2 (Cat. 555886, BD), IgM (Cat. 555856, BD), and incubated for 30 min at 22 ◦C. After incubation, the micro-arrays were washed with PBST and with ddH2O to remove unbound antibodies and salts and then scanned with a Tecan Powerscanner (Tecan Trading AG, Switzerland). Detection was performed as described in [10]. MAPIX version 8.5.0 (Innopsys, France) software was used to analyze the scanned images. The specific antibody levels are expressed in fluorescence intensity (FI) units, the FI of sample diluent alone was subtracted for each antigen. Depicted are means of triplicate measurements.
2.4. Determination of Antibody Reactivity to HBV Sub-Viral Particles
In order to produce subviral particles (SVPs) consisting of HBV envelope proteins, HuH7 cells were transfected with sub-genomic plasmids encoding HBsAg (LHBs, MHBs, SHBs) of isolates of genotypes A2 (N4879; WHO panel no. 3), B2 (N4214; WHO panel no. 4), C2 (N3825; WHO panel no. 9), D1 (N4203; WHO panel no. 10), E (N4881; WHO panel no. 13) described in Chudy et al., 2012 [11] F4 (GenBank: KY382411), H (V1688; WHO panel no. 15/H) described in Chudy et al., 2011 [12] as well as D3 (SHBs only, GenBank: NC_003977) under endogenous viral promoters. Cell culture supernatant was collected from day 2 to day 6 post transfection and cell-cleared supernatant was concentrated by a factor of 50 (Vivaspin 20, 100,000 MWCO, Sartorius, Germany). Total HBsAg content of concentrates was determined with the quantitative Architect HBsAg assay (Abbott Laboratories, Germany).
Antibody reactivity to SVPs was determined by sandwich ELISA in the subjects pre-immune serum, in a serum sample obtained on 05.04.2019 and in serum samples from two control subjects who had been vaccinated with HBsAg-based vaccines (subject 1: anti-HBsAg > 4000 IU/L); subject 2: anti-HBsAg > 6500 IU/L). Ninety six-well microplates (Cat. 675061, Greiner Bio-One, Austria) were coated with 0.2 µg/well HB1 mAb [13] in bicarbonate buffer (pH 9.6) overnight at 4°C, washed twice with 100 µl/well PBS 0.05% Tween 20 (PBST), residual binding sites were blocked with 100 µl/well 3% BSA/PBST for 5h at 37°C. After washing the plates twice, they were incubated with 0.1 µg/well SVPs in HuH7 cell culture supernatant diluted with 0.5% BSA/PBST overnight at 4°C, washed three times, incubated with 50 µl/well serum diluted 1:10 in 0.5% BSA/PBST overnight at 4°C. After washing three times with PBST, detection was performed by addition of 50 µl/well HRP-labeled anti-human IgG (Cat. 555788, BD Biosciences, NJ, USA) diluted 1:3000 in 0.5% BSA/PBST and incubated on the plate for 2h at 37°C. The plates were then washed three times and the reaction was developed by incubation with substrate solution: 1 mg/mL 2,2′-Azino-bis(3-ethylbenzothiazoline-6-sulfonic acid) di-ammonium salt (ABTS) (Cat. A1888, Sigma-Aldrich, MO, USA) in 70 mM citrate-phosphate buffer containing 0.003% H2O2 (Cat.H1009, Sigma-Aldrich); absorbance was measured at wavelength 405 nm (reference wavelength 492 nm) on Tecan Infinite F50 spectrophotometer (Tecan Trading AG, Switzerland). ELISA was controlled by omission of either part of the sandwich and buffer control, the background reactivity of HB1 antibody with each of the serum samples was subtracted, the values are displayed as means of duplicate measurements of optical density (OD) values with deviations of less than 5%.
2.5. Virus Neutralization Assay
A brief summary of the in vitro virus neutralization assay is depicted in Figure S2. In order to generate the infectious HBV preparation in vitro, a human hepatoma cell line (HepG2) was stably transfected with a plasmid encoding a 1.1 over-length HBV genome of genotype D3 (GenBank: NC_003977). Cells were incubated at 37°C, 5% CO2 in William’s E medium (Thermo Fisher Scientific, Germany) containing 2% DMSO (Carl Roth, Germany), 2% fetal calf serum (FCS; Thermo Fisher Scientific), 1 µg/mL doxycycline (Carl Roth), 1 µg/mL G418 (Carl Roth) and 1µg/mL puromycin (Carl Roth) for several weeks with continuous media exchanges. Supernatant was collected and ultra-filtrated to enrich viral particles. Viral load was determined to be 3.7x1011 Ge/mL by a highly-sensitive qPCR as previously described [14]. HBsAg in the viral stock was determined to be 4080 IU/mL by the quantitative Architect HBsAg assay (Abbott Laboratories, Germany).
Figure S2.
Scheme of the in vitro virus neutralization assay. A human hepatoma cell line (HepG2), stably expressing the HBV high affinity entry receptor NTCP, was infected with in vitro generated HBV (genotype D) after pre-incubation with immune sera at specified concentrations. HBV-infected cells secrete the soluble HBeAg, which is an accepted quantitative marker for productive HBV infection in cell culture experiments. To authenticate the generated HBV infectivity parameter HBeAg, the conventionally used qualitative immunofluorescence of HBV-infected cells was used. Here, newly produced intracellular HBV core protein (HBcAg) after HBV infection was stained and analyzed fully-automated to gain additional semi-quantitative data on the number of HBV-infected cells.
Figure S2.
Scheme of the in vitro virus neutralization assay. A human hepatoma cell line (HepG2), stably expressing the HBV high affinity entry receptor NTCP, was infected with in vitro generated HBV (genotype D) after pre-incubation with immune sera at specified concentrations. HBV-infected cells secrete the soluble HBeAg, which is an accepted quantitative marker for productive HBV infection in cell culture experiments. To authenticate the generated HBV infectivity parameter HBeAg, the conventionally used qualitative immunofluorescence of HBV-infected cells was used. Here, newly produced intracellular HBV core protein (HBcAg) after HBV infection was stained and analyzed fully-automated to gain additional semi-quantitative data on the number of HBV-infected cells.
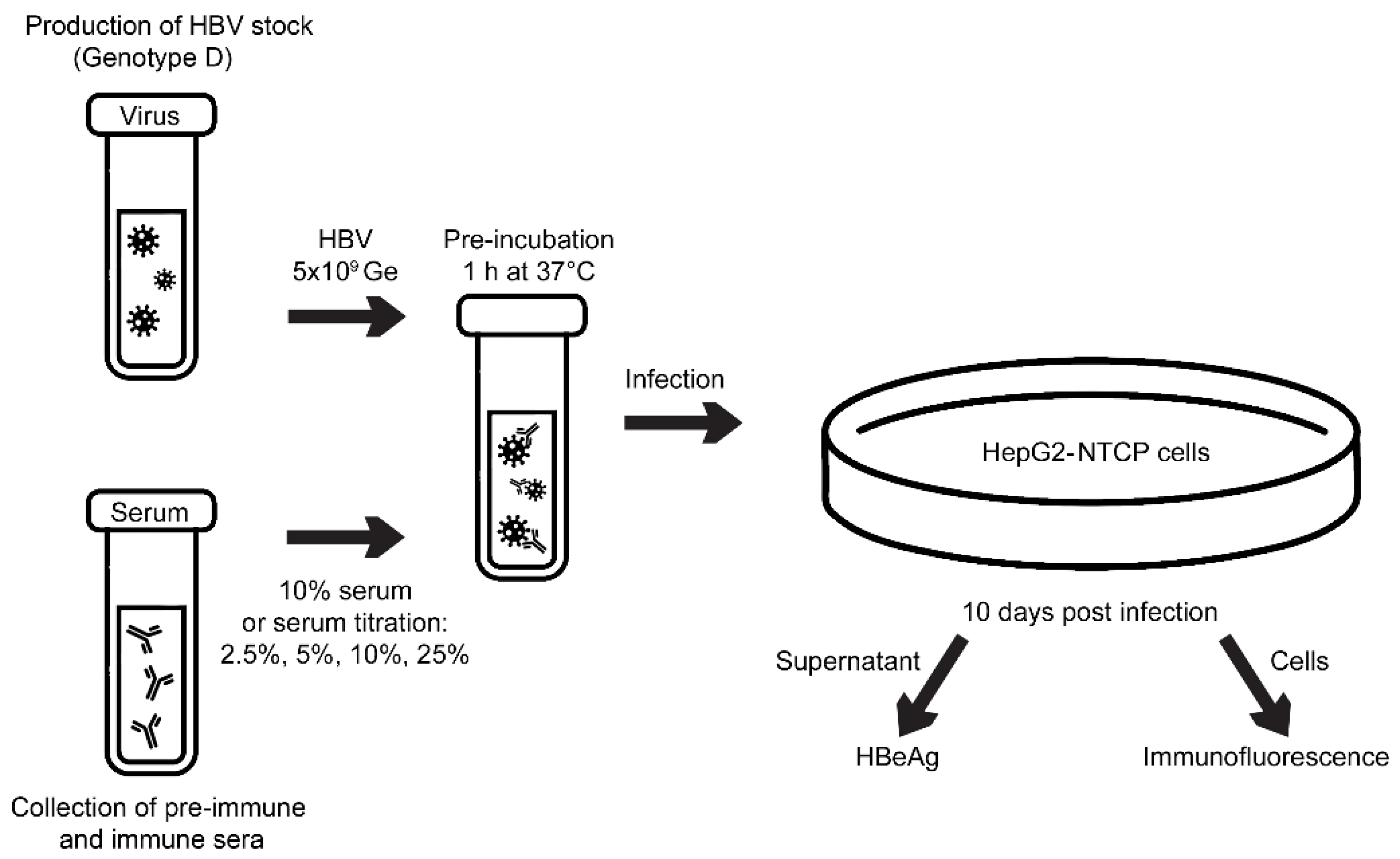
Next, HepG2-NTCP cells [15] were pre-treated with 6 µg/mL doxycycline to induce NTCP expression at least 3 days prior to infection and subsequently seeded into 24-well plates the day before infection. Cells were infected with virus inoculum (5x109 Ge; 55.9 IU HBsAg) in hepatocyte growth medium (HGM) consisting of William’s E medium supplemented with 1× insulin-transferrin-selenium (Thermo Fisher Scientific, Germany), 2 mM l-glutamine (Thermo Fisher Scientific), 100 µg/mL gentamicin (Thermo Fisher Scientific), 10 nM dexamethasone (Sigma-Aldrich, Germany), 1 mM sodium pyruvate (Thermo Fisher Scientific), 0.2% bovine serum albumin (BSA; Carl Roth, Germany) in the presence of 4% PEG-8000 (Sigma-Aldrich), 2% DMSO (Carl Roth) and 100 ng/mL EGF (PeproTech, Germany) as final concentration (f.c.) each. NTCP expression was routinely controlled by uptake of the green-fluorescent bile salt NBD-TC (4-nitrobenzo-2-oxa-1,3-diazole taurocholic acid) as previously described [16].
For neutralization, virus inoculum was pre-incubated with sera at defined concentrations in a dilution with HGM for 1 h, 37°C and under regular agitation. Pre-incubation with mouse-derived monoclonal anti-preS1 antibody MA18/7 detecting an epitope DPXF in the preS1 amino acids 20 to 23 (31 to 34 in genotype A) [17] was performed in the same manner and served as neutralization control. Twenty hours after virus inoculation, cells were washed three times with HGM and further incubated for 9 days in HGM supplemented with 2% DMSO with medium changed every 2 days. As a first outcome parameter of the HBV virus neutralization assay, HBeAg secreted from infected cells into the cell culture supernatant was determined in supernatants collected from day 7 to 10 post infection (p.i)., using the Architect HBeAg assay (Abbott Laboratories, Germany).
As a second outcome parameter, immunofluorescence of the infected cells was measured as follows: At day 10 p.i., cells were fixed by incubation with 3.7% formaldehyde in PBS for one hour at 4°C, washed with PBS and permeabilized using 0.2% Triton X-100 in PBS for 30 min at room temperature (RT). Cells were washed again, and blocked with 10% FCS in PBS for 60 min at RT. As primary antibody, a polyclonal α-HBc antiserum from an immunized guinea pig (Eurogentec, Belgium) was used in a 1:500 dilution for 1 h at 37°C and cells were then washed five times. For detection, cells were incubated with Alexa-488 coupled α-guinea-pig-IgG-antibodies (Thermo Fisher Scientific, Germany) in a 1:400 dilution for 30 min at 37°C in the dark and subsequently washed five times. Nuclei were stained by incubation of cells with 4′,6-diamidino-2-phenylindole (DAPI, 1 µg/mL f.c.) for one hour at 37°C in the dark. Analysis of immunofluorescence was performed with the ImageXpress Pico automated cell-imaging system (Molecular Devices, CA, USA). The total area analyzed was 12% of the total well size.
IC50 values were determined using the integrated “Absolute IC50, X is concentration” analysis integrated into Graphpad Prism version 9.4.1. Results are displayed as IC50 (± 95% CI). In the neutralization experiments serum from a subject immunized with HBsAg-based vaccines containing 2600 IU/L anti-HBs was included for control purposes.
2.6. CD4+/CD8+ T Cell Proliferation Assay and Cytokine Responses
Peripheral blood mononuclear cells (PBMCs) were freshly isolated from heparinized blood by Ficoll-Paque™ Plus density gradient (Cat. 17-1440-03, GE Healthcare, Sweden) and stained with carboxyfluorescein diacetate succinimidyl ester (CFSE) (Cat. C1157, Invitrogen, MA, USA) as described [18]. Aliquots of 2x105 PBMCs in 200 µL of UltraCulture™ Serum-free Medium (Cat. BE12-725F, Lonza, Switzerland) supplemented with 2 mM L-glutamine (Gibco, CA, USA) and 50 µM β-mercaptoethanol (Gibco) and 0.1 mg gentamicin per 500 mL (Gibco) were incubated for 7 days at 37 °C in a humidified atmosphere with 5% CO2 in Nuncclon 96-well plates (Cat. 163320, Thermo Fisher Scientific, MA, USA) in triplicates in the presence of 0.15 µg/well of recombinant preS, preS-derived peptides or mix of peptides P1-P8 equimolar to preS. Aliquots of 0.5 μL/well of Dynabeads Human T-Activator CD3/CD28 (Cat. 11161D, Thermo Fisher Scientific) were used as positive controls, medium alone served as a negative control. On the day 8, flow cytometry analysis and gating was performed as described [6]. The levels of IL-2, IFN-γ, IL-4, IL-5, IL-10, IL-13, GM-CSF and TNF-α were measured in supernatants of seven-day PBMC cultures by Luminex multiplex assay (Bio-Plex ProTM Human Cytokine 9-Plex Assay, Bio-Rad, CA, USA) according to the manufacturer's instructions using the Bio-Plex® 200 System (Bio-Rad). For analysis, the median of the medium-only wells was subtracted from each of the triplicates cultivated with antigen, the values displayed are means of the triplicates.
3. Results
3.1. Repeated Vaccination with HBsAg-Based Vaccines Induced Only Low Anti-HBs Responses in the Study Subject
After the basic immunization with the HBsAg-based vaccine Hevac B Pasteur and subsequent booster injections with Twinrix and Engerix-B (Figure 1) no robust anti-HBs antibody response could be established in the study subject. The subject was 23 years old at the time of completion of the first four-dose immunization schedule and completely healthy. The subject was also not affected by any medical condition or medication intake during the whole further observation period (i.e., until March 2020). The subsequent booster vaccinations with licensed HBsAg-based vaccines Twinrix (January 2000) and Engerix-B (February 2018) elicited only low levels of HBs-specific antibodies (anti-HBs; i.e., <100 IU/L after Engerix B) waning quickly during 2018 as depicted in Figure 2a and showing no virus neutralization before the VVX001 immunization (Figure 2c). Although no further HBsAg-based vaccines were administered, after February 2018 a sharp increase of anti-HBs antibodies to approximately 200 IU/L was found in January 2019 (Figure 2a). These anti-HBs antibodies declined quickly and were already below 50 IU/L one month later.
3.2. VVX001 Induces Robust HBV-Specific Antibody Responses Directed Mainly to the N-Terminus of the preS-Containing NTCP Binding Site
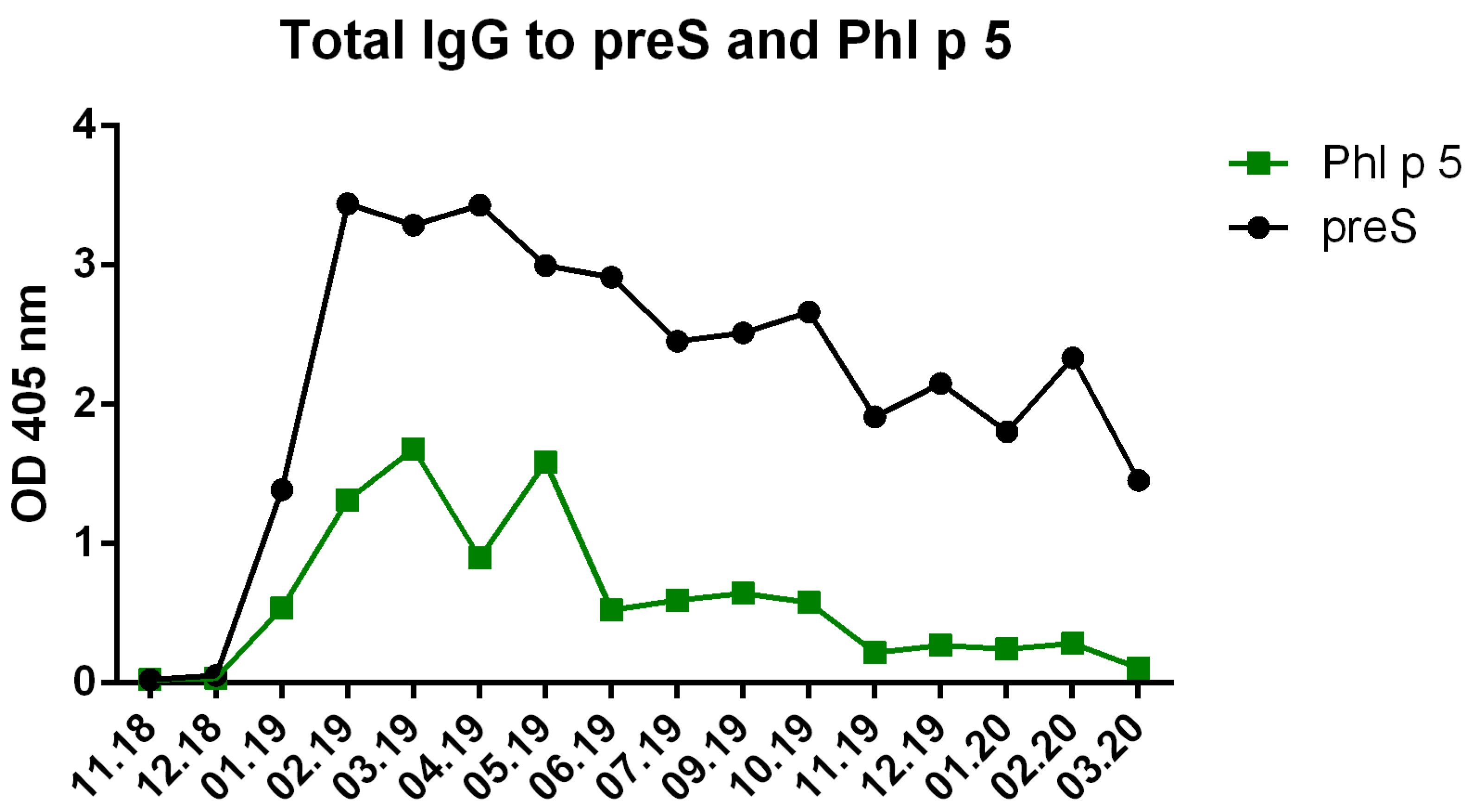
Subcutaneous immunization with VVX001 was started in November 2018. No relevant immediate or late-phase side effects were observed after each of the five injections containing 20 µg VVX001 adsorbed to Aluminum hydroxide when applied in monthly intervals. The vaccination with VVX001 induced a strong and sustained preS-specific IgG response peaking already after the third immunization (Figure 2b, Figure S3) as well as distinct increases of Phl p 5-specific IgG antibodies (Figure S4). During the ongoing VVX001 immunization we also observed a sharp increase of anti-HBs antibodies which vanished almost completely after one month, although VVX001 did not contain SHBs (Figure 2a).
Figure S3.
The preS-specific antibody response was dominated by IgG1. Shown are the OD values of preS-specific IgG, IgG1, IgG2, IgG3, IgG4, IgM, IgA measured by ELISA (y-axis) over the time.
Figure S3.
The preS-specific antibody response was dominated by IgG1. Shown are the OD values of preS-specific IgG, IgG1, IgG2, IgG3, IgG4, IgM, IgA measured by ELISA (y-axis) over the time.
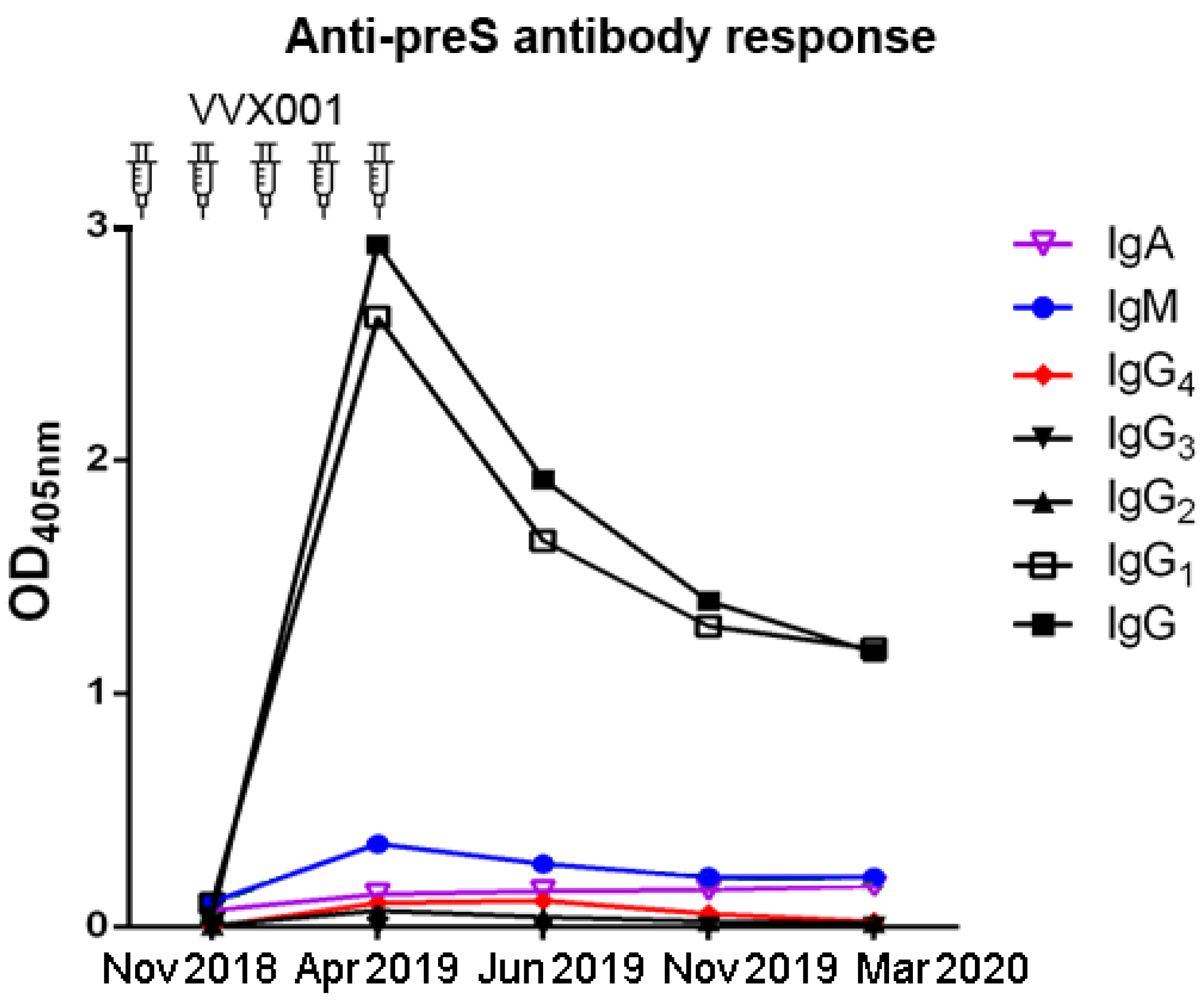
Figure S4.
Phl p 5-specific IgG were lower than preS-specific IgG levels. Shown are the OD values of preS- and Phl p 5-specific IgG levels measured by ELISA over the time.
Figure S4.
Phl p 5-specific IgG were lower than preS-specific IgG levels. Shown are the OD values of preS- and Phl p 5-specific IgG levels measured by ELISA over the time.
The analysis of preS-specific antibody isotypes and subclasses showed that the preS-specific antibody response was composed mainly of IgG1 (Figure S3). The epitope specificity of the induced antibodies assessed with overlapping synthetic peptides P1-P8 (Figure 3a) showed that IgG antibodies were mainly directed to the N-terminal part of preS which contains the liver cell attachment site of HBV (Figure 3b). Antibodies were measured by ELISA and by micro-array technology.
Figure 3.
VVX001-induced IgG antibodies react mainly with the N-terminus of preS1. Shown is (a) the localization scheme of preS-derived peptide within the preS sequence (amino acid numbering indicated for genotype A) and levels of preS/peptide-specific IgG measured by (b) ELISA (OD) and (c) by micro-array technology (fluorescence intensity, FI).
Figure 3.
VVX001-induced IgG antibodies react mainly with the N-terminus of preS1. Shown is (a) the localization scheme of preS-derived peptide within the preS sequence (amino acid numbering indicated for genotype A) and levels of preS/peptide-specific IgG measured by (b) ELISA (OD) and (c) by micro-array technology (fluorescence intensity, FI).
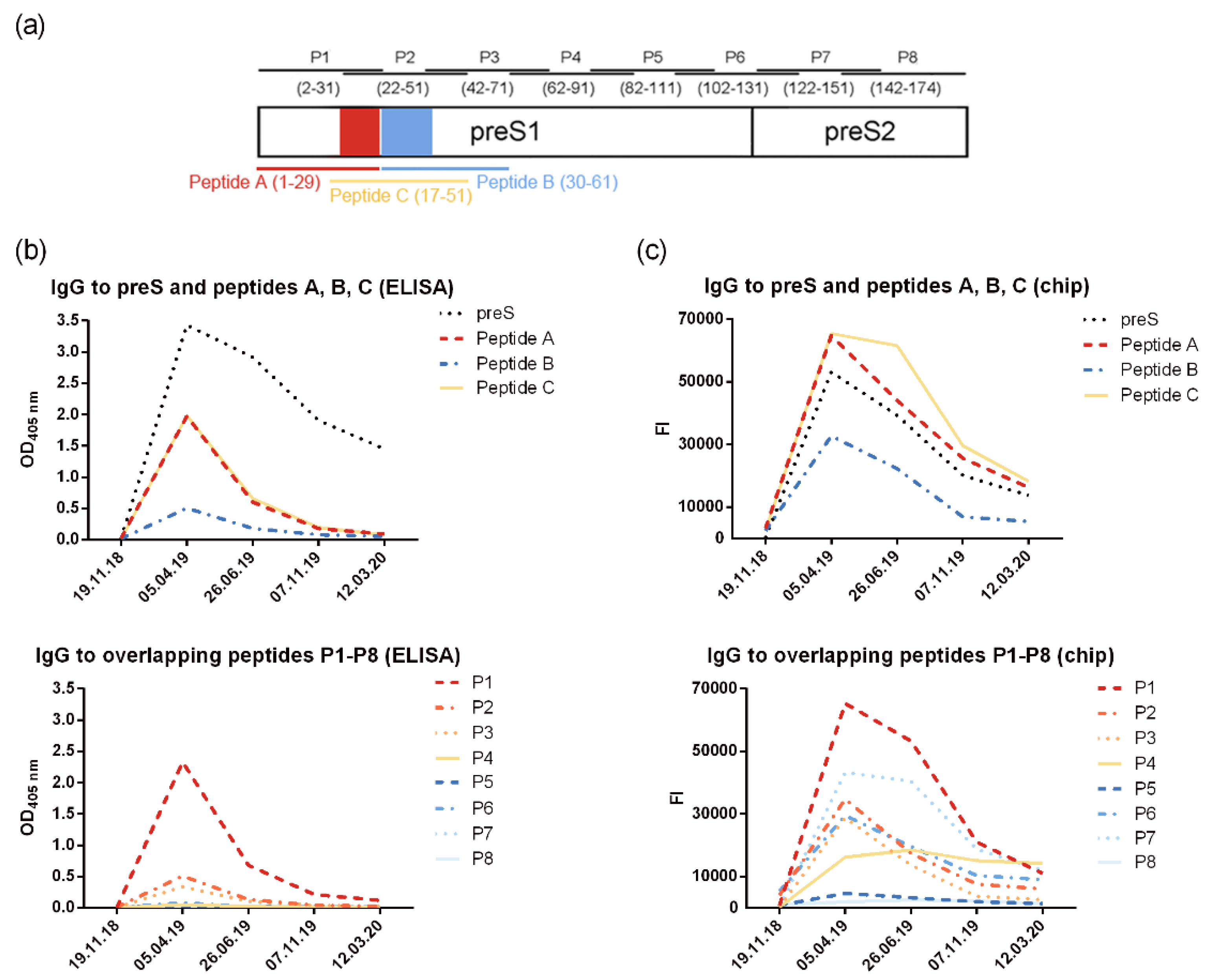
The design of the preS microarray containing preS and preS-derived peptides (six micro-arrays on each glass slide) and images of scanned images are depicted in Figure S1. The results from the micro-array measurements were in good agreement with results obtained by ELISA and showed a similar pattern of IgG reactivity focusing mainly to the N-terminus of preS. However, stronger signals were obtained with the micro-array for the peptides as compared to the complete preS protein (Figure 3b, 3c). The levels of preS-specific IgM and IgA (Figure S5) were low and also directed mainly to N-terminal peptides of preS.
Figure S5.
IgM and IgA levels specific for preS and preS-derived peptides. Shown are levels of preS/peptide-specific (a) IgM and (b) IgA (fluorescence intensity, FI) measured with the preS micro-array.
Figure S5.
IgM and IgA levels specific for preS and preS-derived peptides. Shown are levels of preS/peptide-specific (a) IgM and (b) IgA (fluorescence intensity, FI) measured with the preS micro-array.
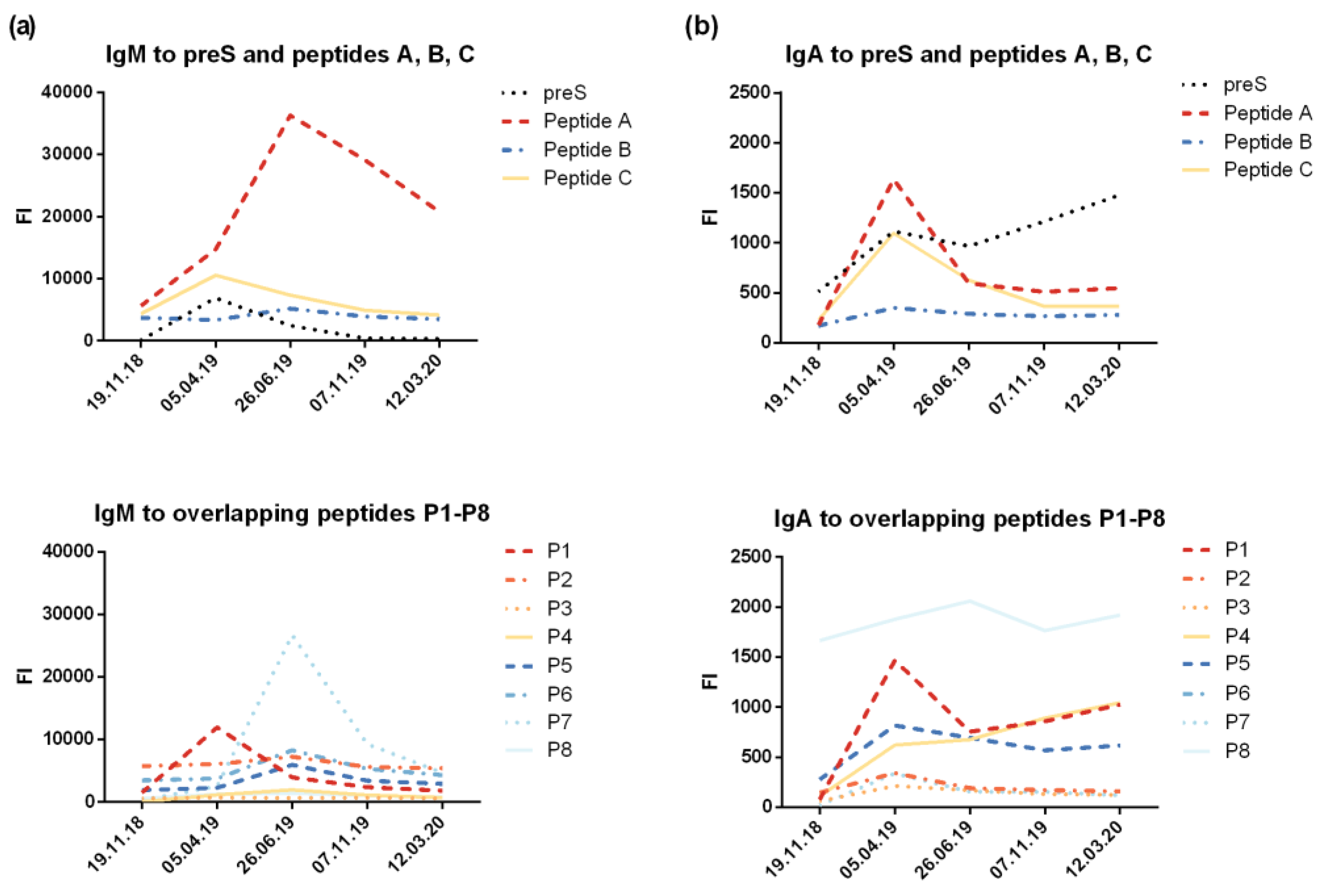
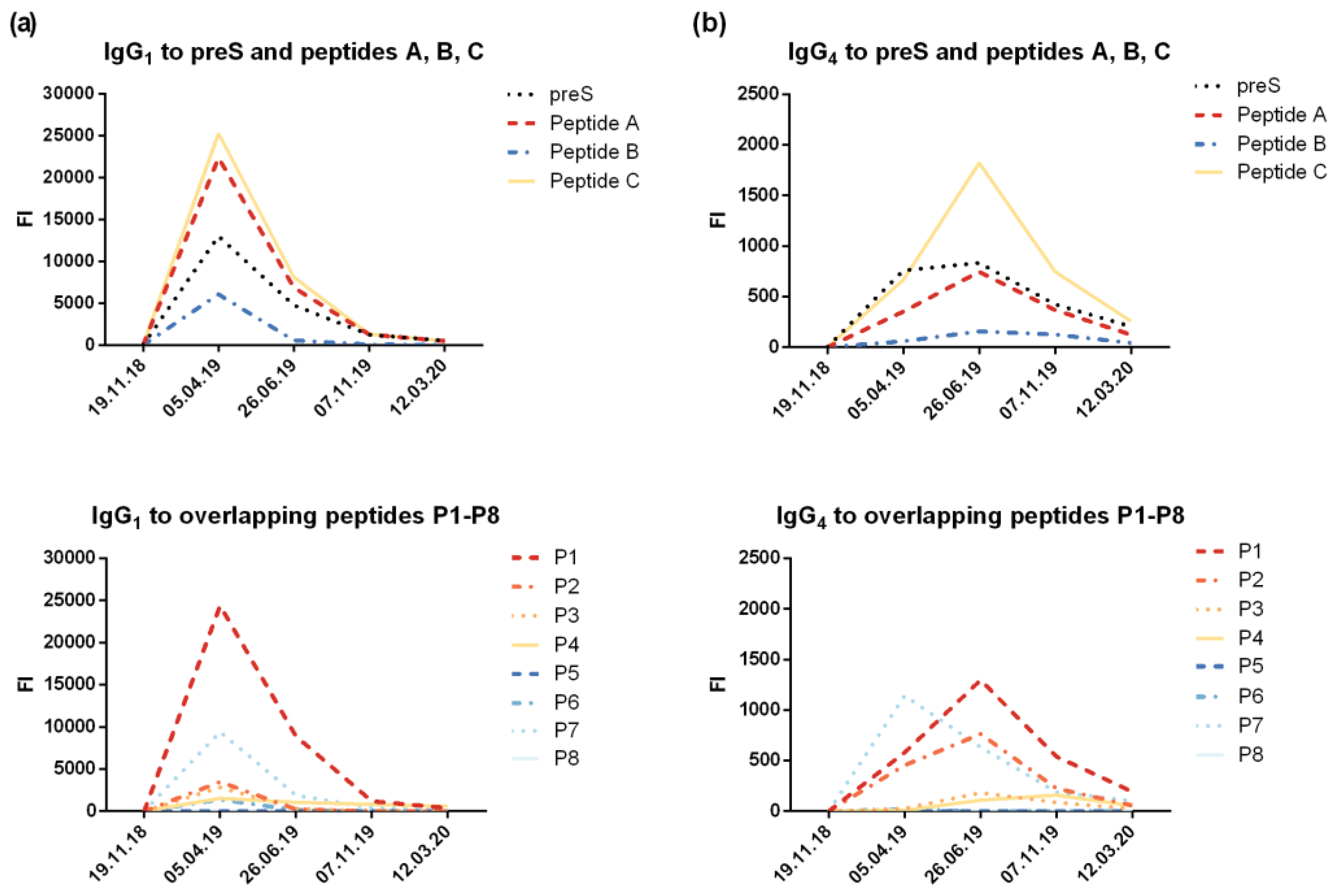
The micro-array measurements of preS- and peptide-specific IgG1 and IgG4 confirmed that IgG1 was the dominating IgG subclass with maximal levels present 1 month after the last injection of VVX001 (Figure S6a). The induction of IgG4 became also visible, but followed a different kinetic. The build-up phase of IgG4 was longer and maximal antibody levels were reached 3 months later than for IgG1 (i.e., 4 months after the last VVX001 injection) (Figure S6b). The quantification of IgG1 and IgG4 specific to preS showed that the preS-specific IgG1 serum concentration reached 1.9 mg/mL one month after the last injection and preS-specific IgG1 at a concentration of 0.68 mg/mL was still present one year after the course of immunizations (Table 1). The concentrations of preS-specific IgG4 were approximately 100-fold lower and, accordingly, could not be detected after the single immunization (Table 1). Nonetheless, preS-specific IgG4 were still detectable by micro-array measurements one year later (Figure S6b). The quantification of N-terminal peptide-specific antibody levels suggested that the induced antibodies reacted to both N-terminal epitopes of preS which are important for HBV infectivity with most of the reactivity directed towards the region defined by peptide A (Table 1).
Figure S6.
IgG1 and IgG4 reactivity to preS and preS-derived peptides. Shown are levels of preS/peptide-specific (a) IgG1 and (b) IgG4 (fluorescence intensity, FI) measured with the preS micro-array.
Figure S6.
IgG1 and IgG4 reactivity to preS and preS-derived peptides. Shown are levels of preS/peptide-specific (a) IgG1 and (b) IgG4 (fluorescence intensity, FI) measured with the preS micro-array.
3.3. VVX001-Induced Antibodies Cross-React with HBV Genotypes A-H
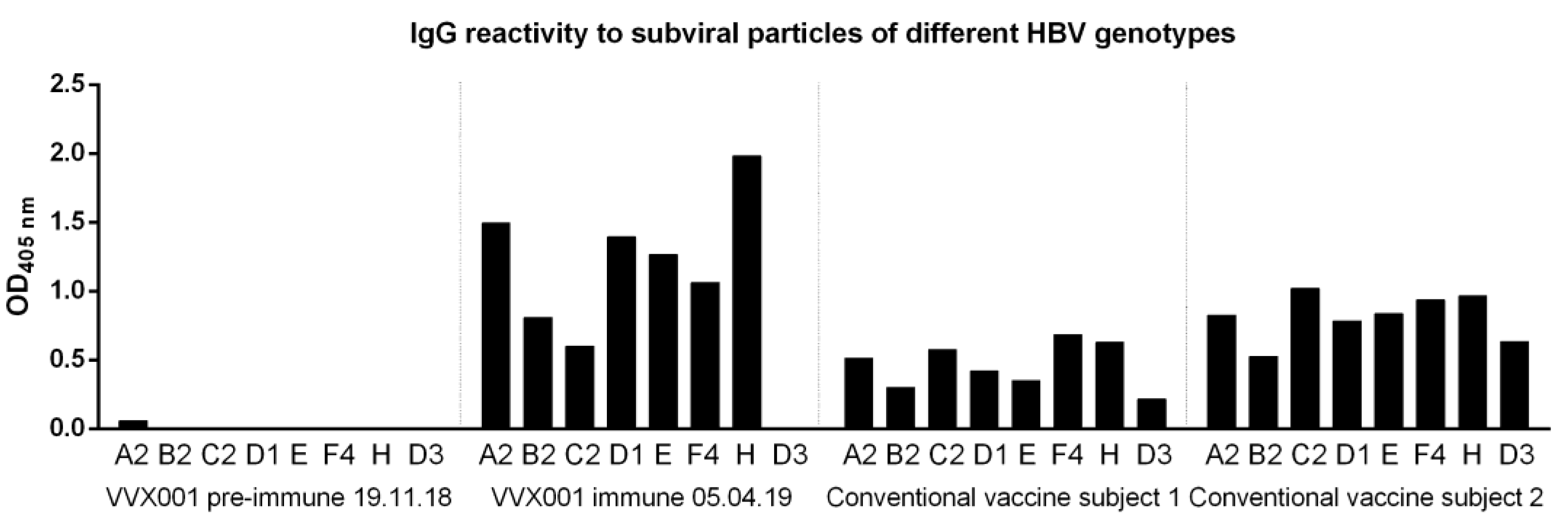
To address the question of genotype cross-reactivity, we have synthesized eight peptides representing the liver cell attachment site of HBV genotypes A-H (Table S1) as described [7]. According to both ELISA and micro-array measurements, IgG cross-reactivity to the eight peptides representing the NTCP attachment site was at all time points detectable for genotypes A-H with somewhat lower reactivity to genotypes D, E and G (Figure 4). The genotype cross-reactivity was also assessed using recombinant SVPs composed of large, middle and small HBV surface proteins (LHBs, MHBs, SHBs) of HBV genotypes A2, B2, C2, D1, E, F4, H as well as a control SVPs composed of genotype D3 SHBs only (Figure 5). The pre-immune serum of the subject of the current study did not show any SVP reactivity before VVX001 immunization. The sample obtained one month after the last immunization reacted to all SVPs tested with the exception of the control SVPs of genotype D3 which contained no preS. Both of the two conventionally vaccinated subjects who had received the standard HBsAg-based immunization and had high anti-HBs levels within two years prior to the blood donation (subject 1: anti-HBs > 4000 IU/L, subject 2: anti-HBs > 6500 IU/L) reacted to all SVPs including control genotype D3 comparably. Serum obtained from the study subject obtained one month (05.04.2019) after the last immunization with VVX001 showed higher reactivity to all LHBs-containing SVPs than the sera from the two control subjects (Figure 5).
Figure 4.
VVX001-induced antibodies cross-react to the NTCP binding site of preS of HBV genotypes A-H. Shown are the IgG levels to the synthetic peptides representing the NTCP attachment site of HBV genotypes A-H measured by (a) ELISA (OD) and (b) micro-array technology (fluorescence intensity, FI).
Figure 4.
VVX001-induced antibodies cross-react to the NTCP binding site of preS of HBV genotypes A-H. Shown are the IgG levels to the synthetic peptides representing the NTCP attachment site of HBV genotypes A-H measured by (a) ELISA (OD) and (b) micro-array technology (fluorescence intensity, FI).
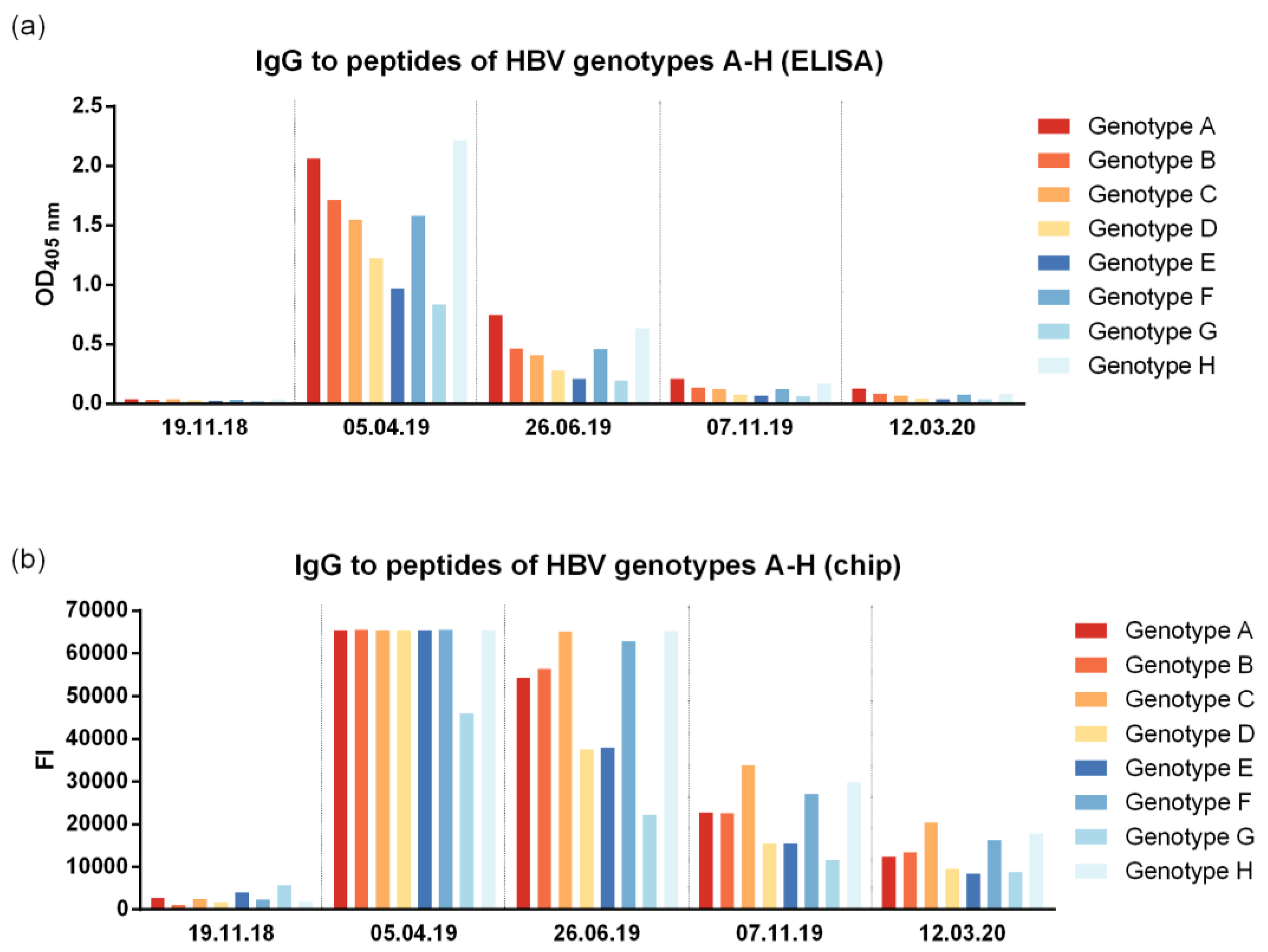
Figure 5.
VVX001-induced antibodies react to SVPs of different HBV genotypes. Shown are IgG levels (OD) to SVPs of HBV genotypes A2, B2, C2, D1, E, F4, H (LHBs, MHBs, SHBs) and D3 (SHBs only) in serum samples of the study subject before and after immunization as well as in two subjects successfully vaccinated with conventional HBsAg-based vaccines.
Figure 5.
VVX001-induced antibodies react to SVPs of different HBV genotypes. Shown are IgG levels (OD) to SVPs of HBV genotypes A2, B2, C2, D1, E, F4, H (LHBs, MHBs, SHBs) and D3 (SHBs only) in serum samples of the study subject before and after immunization as well as in two subjects successfully vaccinated with conventional HBsAg-based vaccines.
3.4. VVX001-Induced Antibodies Strongly Neutralize HBV Infection In Vitro
A neutralizing ability of the antibodies induced by the vaccination was assessed for the samples obtained from the subject after vaccination with Engerix (February 2018) as well as before, during and after immunization with VVX001 (Figure 2c). A well-established diagnostic HBeAg assay was used to determine in vitro virus neutralization with a high range between positive controls (i.e., HBV infection without addition of serum “PC” HBeAg: 158.82-166.09 S/CO (signal/cutoff) and negative control (i.e., uninfected cells HBeAg <1 S/CO). There was no relevant increase of virus neutralization after the last HBsAg booster injection with Engerix-B from February 2018 to November 2018 (Figure 2c). Immunization with VVX001 resulted in a strong increase in HBV neutralization with a strong neutralization observed between the time points February 2019 (1 month after the 3rd VVX001 injection) and May 2019 (2 months after the 5th VVX001 injection). The subsequent samples showed a decrease of neutralization potential but neutralization effects were observed until October 2019. After October 2019 no virus neutralization was observed in the in vitro virus neutralization test although anti-preS antibodies were still present (Figure 2b,c).
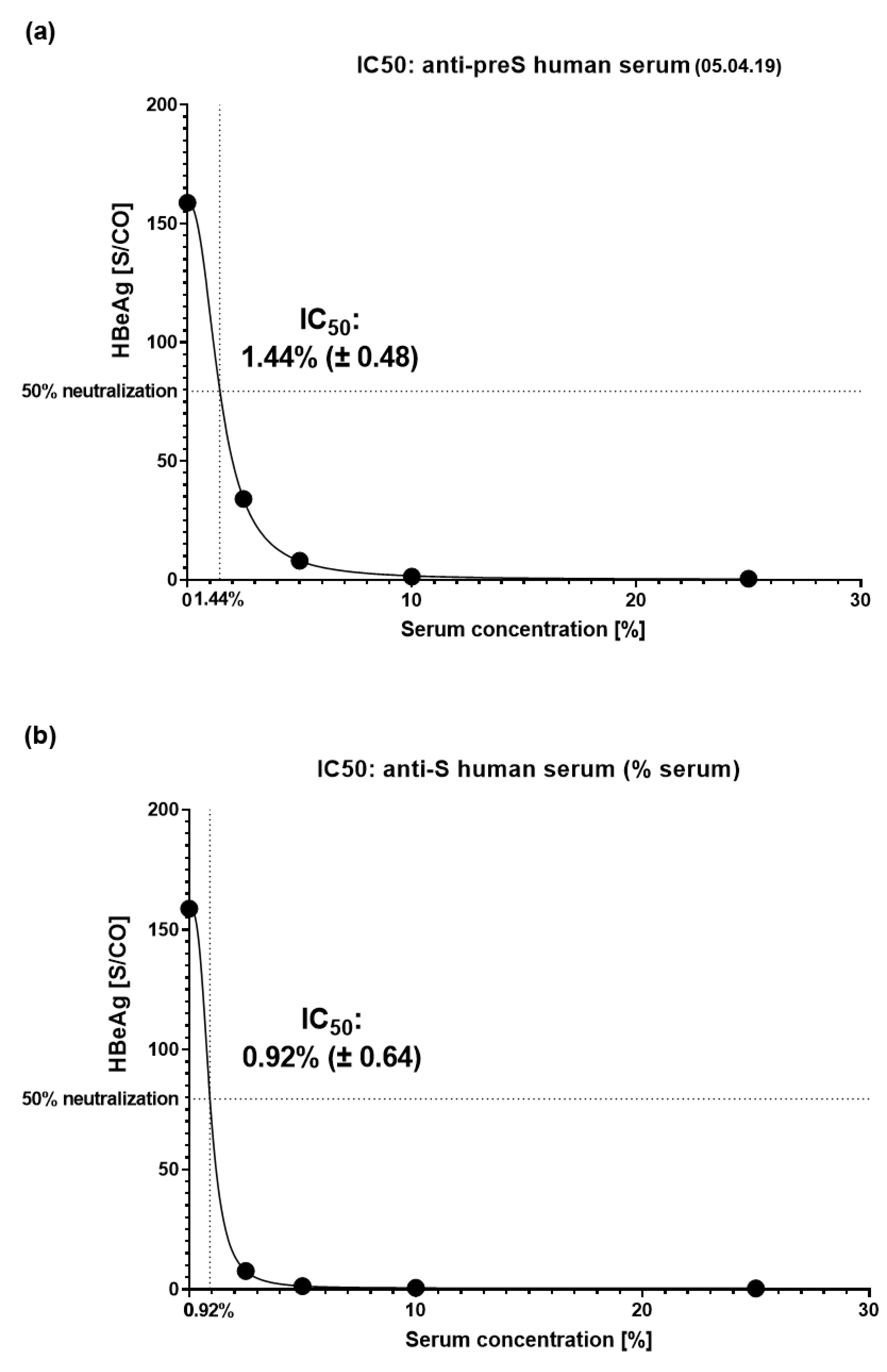
Next, titration experiments were performed to determine IC50 values (Figure S7). For titration the serum obtained 1 month after the last VVX001 injection on 05.04.2019 was used. It showed an IC50 corresponding to 1.44% (± 0.48) of serum in the virus pre-incubation mix (Figure 6a). As a control, an immune serum containing 2600 IU/L of anti-HBs antibodies after conventional vaccination was used which yielded an IC50 corresponding to 0.92% (± 0.64) of serum in the virus pre-incubation mix (Figure 6b). Based on this comparison we estimated that the subject´s serum taken in April 2019 contained “functionally” neutralizing antibodies corresponding to approximately 1661 IU/L of anti-HBs antibodies. The monoclonal anti-preS1 antibody MA18/7 was already 97% neutralizing at 1 µg/mL and fully neutralizing at all higher concentrations (Figure S7a,b). Therefore, the calculation of an IC50 of the MA18/7 could not be performed.
Figure S7.
Additional outcome parameters of in vitro HBV neutralization assays. (a) HBeAg secretion and (b) de novo expressed HBcAg (baseline correction of immunofluorescence was performed by subtracting the value of the uninfected control) of infected cells after pre-incubation of HBV inoculum with titrated sera after vaccination. Data are presented as % compared to positive control (PC, infection w/o serum pre-incubation). Neutralization: ≥90% (strong neutralization; green), ≥50% (partial neutralization; blue), ≥10% (weak neutralization; magenta); HBeAg below cut-off (grey). Ø = 0% HBcAg-positive cells after baseline-correction.
Figure S7.
Additional outcome parameters of in vitro HBV neutralization assays. (a) HBeAg secretion and (b) de novo expressed HBcAg (baseline correction of immunofluorescence was performed by subtracting the value of the uninfected control) of infected cells after pre-incubation of HBV inoculum with titrated sera after vaccination. Data are presented as % compared to positive control (PC, infection w/o serum pre-incubation). Neutralization: ≥90% (strong neutralization; green), ≥50% (partial neutralization; blue), ≥10% (weak neutralization; magenta); HBeAg below cut-off (grey). Ø = 0% HBcAg-positive cells after baseline-correction.
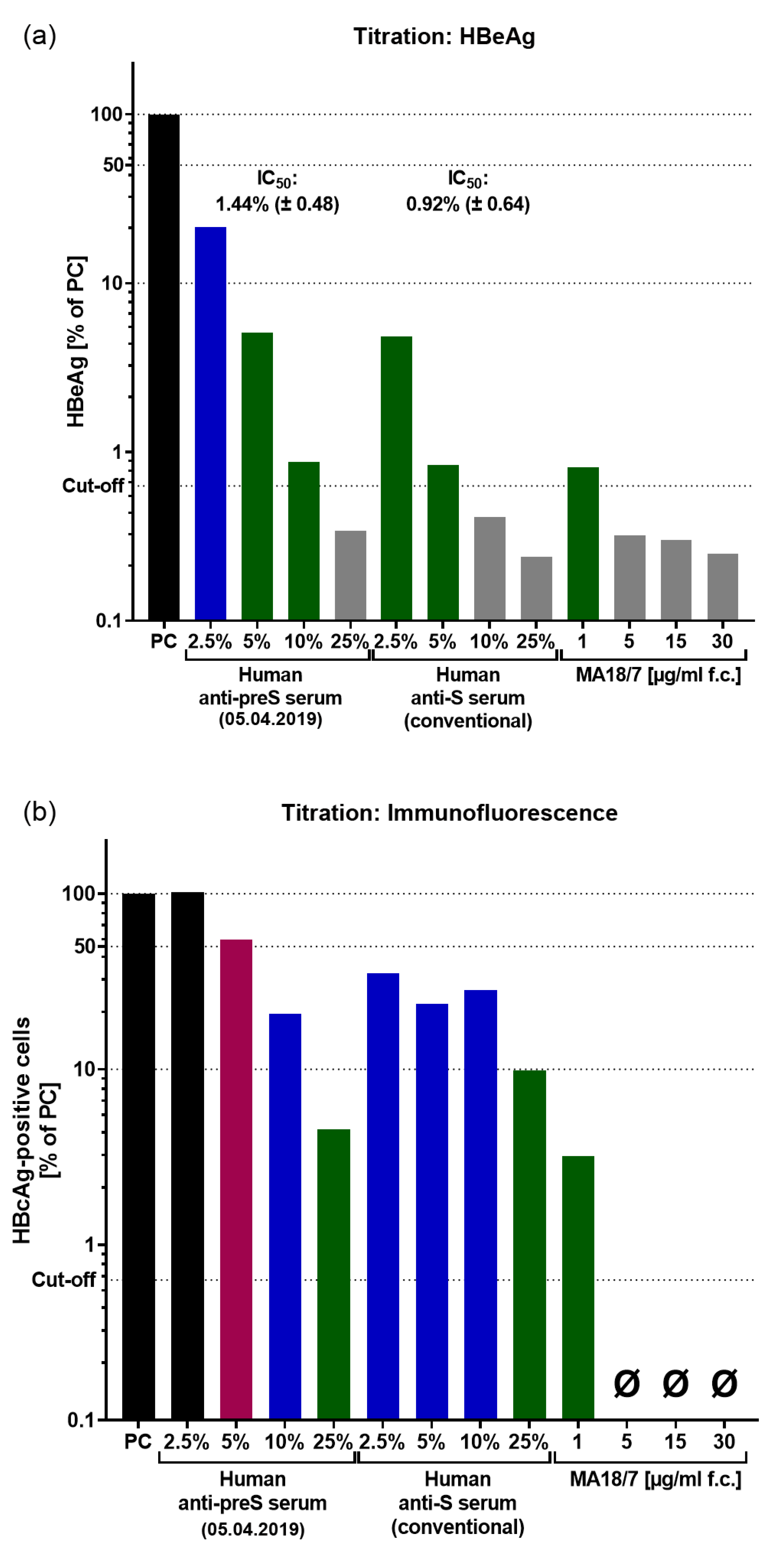
Figure 6.
IC50 determination. IC50 values for serum dilutions determined based on the HBeAg results are presented with 95% confidence interval for (a) the anti-preS-positive human serum obtained in April 2019 and (b) for the anti-S-positive human serum (conventional vaccine, 2600 IU/L anti-HBs).
Figure 6.
IC50 determination. IC50 values for serum dilutions determined based on the HBeAg results are presented with 95% confidence interval for (a) the anti-preS-positive human serum obtained in April 2019 and (b) for the anti-S-positive human serum (conventional vaccine, 2600 IU/L anti-HBs).
In fact, an even higher virus neutralization was measured in the subject of this study after another course of preS-based self-vaccination with a preS-RBD SARS-CoV-2 vaccine as described [19]. After the immunization with the preS-RBD vaccine, a 98.6% HBV neutralization (HBeAg 1.185 S/CO in the tested sample vs. HBeAg 83.63 S/CO at baseline) was observed indicating that it is possible to boost the preS-specific virus-neutralizing antibody response by repeated immunizations.
3.5. VVX001 Induces a Sustained preS-Specific CD3+CD4+ Cellular Response Which Is Accompanied by a Mixed Th2/Th1 Cytokine Response
Vaccination with VVX001 induced a growing and sustained preS-specific CD4+ T cell and to a much lower degree of a preS-specific CD8+ T cell response in the study subject (Figure 7a). Interestingly, peptide 1 derived from the N-terminus of preS, which was identified as a major B cell epitope, was also identified as a T cell epitope-containing peptide (Figure 7b). All peptides derived from the NTCP attachment site of HBV genotypes A-H induced proliferation of CD4+>CD8+ T cells from the vaccinated subject although proliferation stimulated with peptides from genotypes D, F, G, H was low (Figure 7c).
Figure 7.
PreS-specific T cell responses after vaccination with VVX001. Shown are percentages of proliferated СD3+CD4+ and CD3+CD8+ cells upon stimulation with (a) preS over time, (b) preS, P1-P8, equimolar mix of P1-P8 and (c) peptides representing the NTCP attachment site of genotypes A-H at time point 4 months after the last injection (26.06.2019).
Figure 7.
PreS-specific T cell responses after vaccination with VVX001. Shown are percentages of proliferated СD3+CD4+ and CD3+CD8+ cells upon stimulation with (a) preS over time, (b) preS, P1-P8, equimolar mix of P1-P8 and (c) peptides representing the NTCP attachment site of genotypes A-H at time point 4 months after the last injection (26.06.2019).
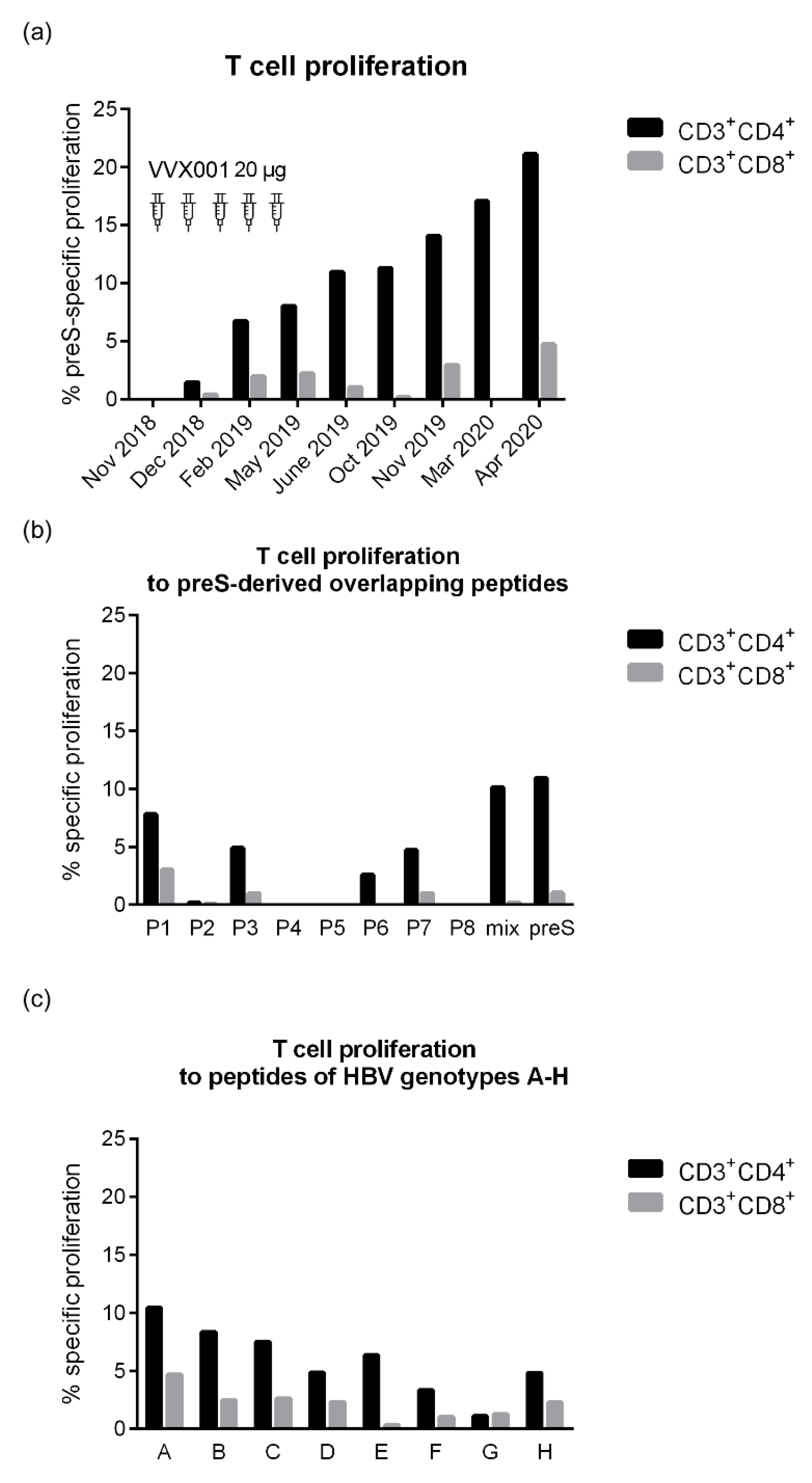
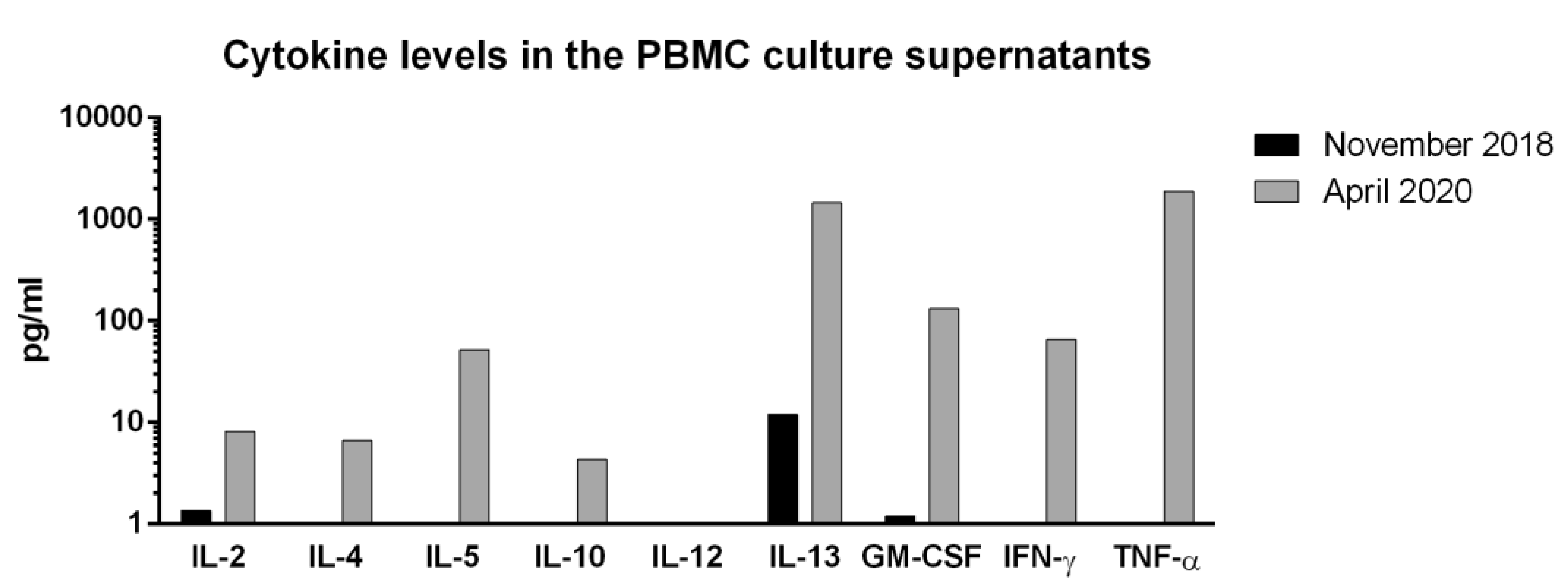
Figure S8 shows the induction of preS-specific cytokine responses induced in cultured PBMCs obtained before and after immunization with VVX001 in the study subject. We found that immunization with VVX001 induced a mixed preS-specific Th2/Th1 cytokine secretion profile, which was characterized by the induction of IL-4, IL-5, IL-13 and GM-CSF on the one hand and by the induction of IFN-γ on the other hand (Figure S8). We also noticed the development of preS-specific IL-2 and TNF-α responses and the development of an eventually tolerogenic IL-10 response (Figure S8).
Figure S8.
PreS-specific cytokine responses. Shown are the levels of IL-2, IFN-γ, IL-4, IL-5, IL-10, IL-13, GM-CSF and TNF-α measured in supernatants of seven-day PBMC cultures stimulated with preS at baseline (November 2018) and one year after the last injection (April 2020) after subtraction of medium control.
Figure S8.
PreS-specific cytokine responses. Shown are the levels of IL-2, IFN-γ, IL-4, IL-5, IL-10, IL-13, GM-CSF and TNF-α measured in supernatants of seven-day PBMC cultures stimulated with preS at baseline (November 2018) and one year after the last injection (April 2020) after subtraction of medium control.
4. Discussion
Previously, we developed a platform for recombinant allergen-specific immunotherapy vaccines which is based on recombinant fusion proteins containing HBV-derived preS as immunological carrier protein fused to hypoallergenic allergen derived peptides [20,21]. PreS was selected as carrier protein for these allergy vaccines because it contains the binding site of HBV to its cognate receptor NTCP on liver cells and hence was expected to generate antibodies upon immunization which may protect also against HBV infections. Indeed, we found that grass pollen allergic patients who had been immunized with BM32, a grass pollen allergy vaccine containing four preS-fusion proteins, BM321, BM322, BM325 and BM326, developed preS-specific antibodies, which were able to neutralize HBV infections in vitro [6,7]. Accordingly, BM32 and its components were considered as vaccine candidates against HBV infections. To the best of our knowledge, BM32 is the first and yet the only recombinant vaccine which has been used for active immunization of humans and is based solely on preS [5,22]. A number of vaccine candidates contained preS1 and/or preS2, mainly in combination with HBsAg or other HBV antigens, but very few advanced to the market, among which were the so-called third-generation vaccines PreHevbrio (also known as Sci-B-Vac, Bio-Hep-B, Hepimmune) and Hepacare (other name Hepagene, discontinued), both produced in mammalian cells containing a low portion of preS when compared to its contents of HBsAg [23,24,25].
The aim of the present self-experiment was to induce a protective immune response against HBV in a medical doctor and researcher with regular contact to blood products (R.V.). The study subject has been vaccinated six times with HBsAg-based vaccines without achieving protective HBV-specific immunity as documented by the lack of formation of HBV-neutralizing antibodies. Even after the last HBsAg-based vaccination only a very low and rapidly declining HBV-specific antibody response not exceeding 60 IU/L was achieved. Thus the achieved seroconversion at a low level (anti-HBs < 100 IU/L) was considered unsuccessful vaccination according to current guidelines [3,23,26].
Since conventional HBsAg-based vaccines did not seem to ensure protection against HBV, self-immunization with the preS-based recombinant vaccine VVX001 was performed by the volunteer in accordance to the Declaration of Helsinki. The vaccine was well tolerated. The maximal anti-preS level as well as strong increase in HBV neutralization was observed already after the third injection (Figure 2), which suggests that a short basic immunization might be sufficient. The increase in virus neutralization was associated with the induction of IgG antibodies to the N-terminal part of preS (represented by peptides A, B, C, P1, P2, P3 and peptides of genotypes A-H) (Figure 3, Table 1) and to the whole preS protein which were still strongly detectable 1 year after the injection course in the absence of virus neutralizing ability in the serum. Generally, mostly N-terminal preS1 peptides, but also preS2 N-terminus peptides (P6, P7), were recognized by both antibodies and T cells, corresponding to the regions with described HBV neutralizing epitopes [2].
We have already reported earlier the results from proof-of-principle work demonstrating the HBV neutralization capacity of BM32-induced antibodies in a random population of grass pollen allergic subjects [6] and a detailed report on its epitope mapping, cross-reactivity to NTCP binding site of genotypes A-H and quantification of serum antibody concentrations [7,27]. This study confirms our previous observations but also contains several new important findings.
First, we demonstrate that VVX001 can induce an HBV-neutralizing antibody response in a low-responder to classical HBV vaccines. Second, we show that the HBV-neutralizing antibody response can be obtained with a relatively low dose of BM antigen, while the doses administered in the previous allergy trials included two, four and eight times more preS than in our current study. Third, the HBV genotype cross-reactivity was assessed not only with synthetic linear peptides but also with SVPs containing all three HBV surface proteins in a three-dimensional conformation analogous to that of infectious virions. Furthermore, we report the development of a micro-array containing preS and preS-derived peptides which turned out to be useful for analyzing preS-specific antibody responses with small sample volumes (i.e., approx., 1 μl per test/array).
Immunization with VVX001 induced a robust anti-preS IgG response which mainly consisted of a preS-specific IgG1 subclass response, but also preS-specific IgG4 development was observed. It is quite likely that the preS-specific IgG4 response would be boosted by further immunizations to achieve a sustained specific IgG4 response as it was observed in earlier allergen-specific immunotherapy trials with BM32 [7]. Peculiarly, in the course of the immunization with VVX001 a short-term increase of anti-HBs up to 210 IU/L was observed although VVX001 does not contain HBsAg. At the moment we can only speculate why this increase occurred. There were no clinical signs of infection during the increase of anti-HBs antibodies indicative of an HBV infection in the study subject thereafter.
A comparison of the efficacy of the VVX001-induced preS-specific antibodies with successful conventional vaccination was obtained in two types of experiments, i.e., serum titration and IC50 determination in the virus neutralization assay and by demonstration of serum IgG reactivity to SVPs of different HBV genotypes. In both tests the VVX001 immunization has been comparable to standard vaccination yielding high anti-HBs levels (>2000 IU/L).
The protection against heterologous HBV genotypes is uneven and breakthrough symptomatic or occult HBV infections occur despite vaccination [23]. According to IgG and T cell reactivity to HBV preS1-peptides representing the NTCP attachment site, after VVX001 immunization, only the genotype G was notably less recognized, and this genotype was also not represented in the SVP experiment. However, infections with genotype G are rare and chronic infections are only observed during coinfection with another HBV genotype [28]. Otherwise, the tested cross-reactivity was comparable for the common HBV genotypes. Of note, the preS-specific T cell and cytokine response measured in the study subject indicates that the response consists mainy of a CD4+ and to a low extent of a CD8+ T cell response accompanied by a mixed Th2/Th1 cytokine response. This response is compatible with the induction of a robust blocking antibody response and may be modified on demand by the use of different adjuvants or modes of vaccination if a more cytotoxic responses is needed.
PreS-based HBV vaccines are of interest because they may be used as preventive and eventually also as therapeutic HBV vaccines. In fact, preS-specific antibodies may inhibit infection of liver cells by preventing HBV from binding to its receptor NTCP. The preS1-derived lipopeptide bulevirtide (also known as Myrcludex B) which inhibits the NTCP-preS1 interaction is now an approved drug [4]. Bulevirtide has the advantage of immediate action not requiring the functional immune response of the host, however, the disadvantages are the quick elimination time, interference with bile acid transportation in hepatocytes and, most importantly, lack of the long-term protection if the treatment is discontinued. Therapeutic vaccination with preS-based vaccines such as VVX001 may have advantages over entry inhibitors blocking the preS1-NTCP interaction. Firstly, preS fusion constructs can be inexpensively obtained in gram amounts under good manufacturing practice conditions by recombinant expression in E. coli as soluble, pure and stable proteins [29]. While the myristoylation of preS1-peptide was essential for the efficacy of bulevirtide, it does not seem to be important for the induction of HBV-neutralizing antibodies as illustrated by the fact that BM32 produced in E. coli lacking myristoylation could induce neutralizing antibodies and as also shown by various monoclonal antibody studies [30].
Another advantage of preS-based vaccination is that active post-vaccination immunity is achieved by relatively short immunization schedules and could be subsequently boosted with just one or few immunizations. In this context, preS-based vaccination may be also considered to block co-infection with hepatitis D virus, a satellite viroid requiring HBV envelope proteins for replication. Hepatitis D is considered the most severe chronic form of viral hepatitis due to quicker progression towards liver-related death and often requires liver transplantation [4]. Vaccination with preS induces polyclonal preS-specific antibodies which may prevent the formation of escape HBV mutants due to high conservation of the NTCP binding site between all common HBV genotypes A-J as evidenced in our study and in our previous work [7]. Finally, another important application of preS-based vaccines like VVX001 might be the prevention of mother-to-child transmission which is responsible for a considerable rate of break-through infections in neonates despite the application of both active and passive HBsAg-based vaccination [23].
Regarding preventive vaccination, preS-based vaccines may especially be considered to overcome low- and non-responsiveness in subjects to HBsAg-based vaccines. In fact, between 5-20% of the persons currently vaccinated with HBsAg-based vaccines represent low- or non-responders [2,3,23]. It is a limitation of our study that only one person was tested and more data is needed to test this approach. As a matter of fact, VVX001 is currently under evaluation in a clinical trial including, alongside healthy volunteers and HBV-infected individuals, a population of low- and non-responders to the HBsAg-based vaccination (NCT03625934).
In conclusion, our study showed that vaccination with the preS-based vaccine VVX001 was safe and induced a robust HBV-neutralizing antibody response in a low responder subject to HBsAg-based vaccine. VVX001 may therefore be considered as a promising candidate for HBV vaccination in low or non-responders to currently available HBsAg-based vaccines. Further clinical studies will be required to test this hypothesis.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Supplemental Figures are currently included in the manuscript to facilitate the reviewing process.
Author Contributions
Conceptualization, R.V. and I.T.; methodology, software, validation, formal analysis, investigation, data curation, I.T., F.L., N.G., M.T., D.T., C.C., M.F., M.W., U.W., D.G.; visualization, I.T., F.L., N.G., A.D.; writing—original draft preparation, I.T., R.V.; writing—review and editing, I.T., F.L., N.G., A.D., D.T., M.T., C.C., M.W., M.F., A.K., R.H., U.W., D.G. and R.V.; project administration, I.T., R.V.; supervision, resources, funding acquisition, R.V. All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by Viravaxx AG, by the Danube Allergy Research Cluster of the country of Lower Austria (FA648A0312) and was supported by a grant from the Russian Science Foundation (project no.: 23-75-30016: “Allergen micro-array-based assessment of allergic sensitization profiles in the Russian Federation as basis for personalized treatment and prevention of allergy (AllergochipRUS)”) regarding the development of antigen-specific IgG measurements by microarray technology.
Institutional Review Board Statement
The self-experiment performed by R.V. was conducted according to the Declaration of Helsinki, paragraph 37 (as on 23.07.2024, source: https://www.wma.net/policies-post/wma-declaration-of-helsinki-ethical-principles-for-medical-research-involving-human-subjects/). Sample analysis was approved by the Ethics Committee of Justus Liebig University Giessen (AZ 257/2019 and AZ 196/2020) and of Medical University of Vienna (EK 1641/2014).
Data Availability Statement
All the data supporting the findings of this study is available in the text, figures, tables, supplementary figures or from the corresponding author upon reasonable request.
Conflicts of Interest
Rudolf Valenta has received research grants from HVD Life-Sciences, Vienna, Austria, WORG Pharmaceuticals, Hangzhou, China and from Viravaxx AG, Vienna, Austria. He served as a consultant for Viravaxx AG. Dieter Glebe has received research grants from Viravaxx AG and served as a consultant for Viravaxx AG. Rainer Henning was an employee of Viravaxx AG. Rudolf Valenta, Carolin Cornelius, Margarete Focke-Tejkl and Milena Weber are authors of a patent application regarding the vaccine. The other authors have no conflicts of interest to declare. The authors with a Russian affiliation declare that they have prepared the article in their “personal capacity” and/or that they are employed at an academic/research institution where research or education is the primary function of the entity. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- World Health Organization Global Hepatitis Report: Action for Access in Low-and Middle-Income Countries, 2024. Global Hepatitis Programme; 2024; ISBN 9789240091672.
- Gerlich, W.H. Prophylactic Vaccination against Hepatitis B: Achievements, Challenges and Perspectives. Med Microbiol Immunol 2015, 204. [Google Scholar] [CrossRef]
- Gerlich, W.H.; Glebe, D. Hepatitis B-Impfung: Altbewährt, Doch Nicht Perfekt. Hepatitis & More 2010, 2, https://www.hepatitisandmore.de/archiv/2010–2/fobi_gerlichshtml. [Google Scholar]
- Liu, H.; Zakrzewicz, D.; Nosol, K.; Irobalieva, R.N.; Mukherjee, S.; Bang-Sørensen, R.; Goldmann, N.; Kunz, S.; Rossi, L.; Kossiakoff, A.A.; et al. Structure of Antiviral Drug Bulevirtide Bound to Hepatitis B and D Virus Receptor Protein NTCP. Nat Commun 2024, 15. [Google Scholar] [CrossRef]
- Zieglmayer, P.; Focke-Tejkl, M.; Schmutz, R.; Lemell, P.; Zieglmayer, R.; Weber, M.; Kiss, R.; Blatt, K.; Valent, P.; Stolz, F.; et al. Mechanisms, Safety and Efficacy of a B Cell Epitope-Based Vaccine for Immunotherapy of Grass Pollen Allergy. EBioMedicine 2016, 11. [Google Scholar] [CrossRef]
- Cornelius, C.; Schöneweis, K.; Georgi, F.; Weber, M.; Niederberger, V.; Zieglmayer, P.; Niespodziana, K.; Trauner, M.; Hofer, H.; Urban, S.; et al. Immunotherapy With the PreS-Based Grass Pollen Allergy Vaccine BM32 Induces Antibody Responses Protecting Against Hepatitis B Infection. EBioMedicine 2016, 11. [Google Scholar] [CrossRef]
- Tulaeva, I.; Cornelius, C.; Zieglmayer, P.; Zieglmayer, R.; Schmutz, R.; Lemell, P.; Weber, M.; Focke-Tejkl, M.; Karaulov, A.; Henning, R.; et al. Quantification, Epitope Mapping and Genotype Cross-Reactivity of Hepatitis B PreS-Specific Antibodies in Subjects Vaccinated with Different Dosage Regimens of BM32. EBioMedicine 2020, 59. [Google Scholar] [CrossRef]
- Salm, K. Herstellung von Hevac B Pasteur, Dtsch Arztebl. Dtsch Arztebl 1983, 80, A-75. [Google Scholar]
- Gattinger, P.; Niespodziana, K.; Stiasny, K.; Sahanic, S.; Tulaeva, I.; Borochova, K.; Dorofeeva, Y.; Schlederer, T.; Sonnweber, T.; Hofer, G.; et al. Neutralization of SARS-CoV-2 Requires Antibodies against Conformational Receptor-Binding Domain Epitopes. Allergy: European Journal of Allergy and Clinical Immunology 2022, 77. [Google Scholar] [CrossRef]
- Lupinek, C.; Wollmann, E.; Baar, A.; Banerjee, S.; Breiteneder, H.; Broecker, B.M.; Bublin, M.; Curin, M.; Flicker, S.; Garmatiuk, T.; et al. Advances in Allergen-Microarray Technology for Diagnosis and Monitoring of Allergy: The MeDALL Allergen-Chip. Methods 2014, 66. [Google Scholar] [CrossRef]
- Chudy, M.; Hanschmann, K.M.; Kress, J.; Nick, S.; Campos, R.; Wend, U.; Gerlich, W.; Nübling, C.M. First WHO International Reference Panel Containing Hepatitis B Virus Genotypes A-G For Assays Of The Viral DNA. J Clin Virol. 2012, 55. [Google Scholar] [CrossRef]
- Chudy, M; Hanschmann, KM; Scheiblauer, H; Wend, U; Schüttler, C; et al. Collaborative study to establish a World Health Organization international Hepatitis B virus genotype panel for HBsAG Assays. World Health Organization. 2011. Available at: https://iris.who.int/handle/10665/70785.
- Kucinskaite-Kodze, I.; Pleckaityte, M.; Bremer, C.M.; Seiz, P.L.; Zilnyte, M.; Bulavaite, A.; Mickiene, G.; Zvirblis, G.; Sasnauskas, K.; Glebe, D.; et al. New Broadly Reactive Neutralizing Antibodies against Hepatitis B Virus Surface Antigen. Virus Res 2016, 211. [Google Scholar] [CrossRef]
- Drosten, C.; Weber, M.; Seifried, E.; Roth, W.K. Evaluation of a New PCR Assay with Competitive Internal Control Sequence for Blood Donor Screening. Transfusion (Paris) 2000, 40. [Google Scholar] [CrossRef] [PubMed]
- König, A; Döring, B; Mohr, C; Geipel, A; Geyer, J; Glebe, D. Kinetics of the bile acid transporter and hepatitis B virus receptor Na+/taurocholate cotransporting polypeptide (NTCP) in hepatocytes. J Hepatol. 2014, 61. [Google Scholar] [CrossRef]
- de Carvalho Dominguez Souza, B.F.; König, A.; Rasche, A.; de Oliveira Carneiro, I.; Stephan, N.; Corman, V.M.; Roppert, P.L.; Goldmann, N.; Kepper, R.; Müller, S.F.; et al. A Novel Hepatitis B Virus Species Discovered in Capuchin Monkeys Sheds New Light on the Evolution of Primate Hepadnaviruses. J Hepatol 2018, 68. [Google Scholar] [CrossRef]
- Glebe, D.; Aliakbari, M.; Krass, P.; Knoop, E. V.; Valerius, K.P.; Gerlich, W.H. Pre-S1 Antigen-Dependent Infection of Tupaia Hepatocyte Cultures with Human Hepatitis B Virus. J Virol 2003, 77. [Google Scholar] [CrossRef] [PubMed]
- Quah, B.J.C.; Warren, H.S.; Parish, C.R. Monitoring Lymphocyte Proliferation in Vitro and in Vivo with the Intracellular Fluorescent Dye Carboxyfluorescein Diacetate Succinimidyl Ester. Nat Protoc 2007, 2. [Google Scholar] [CrossRef]
- Gattinger, P.; Kratzer, B.; Tulaeva, I.; Niespodziana, K.; Ohradanova-Repic, A.; Gebetsberger, L.; Borochova, K.; Garner-Spitzer, E.; Trapin, D.; Hofer, G.; et al. Vaccine Based on Folded Receptor Binding Domain-PreS Fusion Protein with Potential to Induce Sterilizing Immunity to SARS-CoV-2 Variants. Allergy: European Journal of Allergy and Clinical Immunology 2022, 77. [Google Scholar] [CrossRef] [PubMed]
- Valenta, R.; Campana, R.; Focke-Tejkl, M.; Niederberger, V. Vaccine Development for Allergen-Specific Immunotherapy Based on Recombinant Allergens and Synthetic Allergen Peptides: Lessons from the Past and Novel Mechanisms of Action for the Future. Journal of Allergy and Clinical Immunology 2016, 137. [Google Scholar] [CrossRef]
- Valenta, R.; Campana, R.; Niederberger, V. Recombinant Allergy Vaccines Based on Allergen-Derived B Cell Epitopes. Immunol Lett 2017, 189. [Google Scholar] [CrossRef]
- Niederberger, V.; Neubauer, A.; Gevaert, P.; Zidarn, M.; Worm, M.; Aberer, W.; Malling, H.J.; Pfaar, O.; Klimek, L.; Pfützner, W.; et al. Safety and Efficacy of Immunotherapy with the Recombinant B-Cell Epitope–Based Grass Pollen Vaccine BM32. Journal of Allergy and Clinical Immunology 2018, 142. [Google Scholar] [CrossRef]
- Gerlich, W.H. Hepatitis-B-Impfstoffe – Geschichte, Erfolge, Herausforderungen Und Perspektiven. Bundesgesundheitsblatt Gesundheitsforschung Gesundheitsschutz 2022, 65. [Google Scholar] [CrossRef]
- Zuckerman, J.N.; Zuckerman, A.J. Recombinant Hepatitis B Triple Antigen Vaccine: Hepacarer®. Expert Rev Vaccines 2002, 1. [Google Scholar] [CrossRef]
- Roggendorf, H.; Shouval, D.; Roggendorf, M.; Gerken, G. Longterm Outcome of Therapeutic Vaccination with a Third Generation Pre-S/S HBV Vaccine (PreHevbrioR) of Chronically HBV Infected Patients. J Pers Med 2024, 14. [Google Scholar] [CrossRef]
- Robert Koch-Institut Empfehlung Der Ständigen Impfkommission (STIKO) Beim Robert Koch-Institut - 2017/2019. Epidemiologischen Bulletin 2018.
- Gerlich, W.H. “Dual Use”: The Anti-Allergy Vaccine BM32 and Its HBV Carrier Protein. EBioMedicine 2020, 60. [Google Scholar] [CrossRef]
- Liu, Z.; Zhang, Y.; Xu, M.; Li, X.; Zhang, Z. Distribution of Hepatitis B Virus Genotypes and Subgenotypes. Medicine (United States) 2021, 100. [Google Scholar] [CrossRef]
- Focke-Tejkl, M.; Weber, M.; Niespodziana, K.; Neubauer, A.; Huber, H.; Henning, R.; Stegfellner, G.; Maderegger, B.; Hauer, M.; Stolz, F.; et al. Development and Characterization of a Recombinant, Hypoallergenic, Peptide-Based Vaccine for Grass Pollen Allergy. Journal of Allergy and Clinical Immunology 2015, 135. [Google Scholar] [CrossRef]
- Bremer, C.M; Sominskaya, I.; Skrastina, D.; Pumpens, P.; El Wahed, A.A.; Beutling, U.; Frank, R.; Fritz, H.J.; Hunsmann, G.; Gerlich, W.H.; Glebe, D. N-Terminal Myristoylation-Dependent Masking Of Neutralizing Epitopes In The Pres1 Attachment Site Of Hepatitis B Virus. J Hepatol. 2011, 55. [Google Scholar] [CrossRef]
Figure 1.
Vaccinations with conventional HBsAg vaccines in the study subject. Time points and administered vaccines are indicated.
Figure 1.
Vaccinations with conventional HBsAg vaccines in the study subject. Time points and administered vaccines are indicated.
Figure 2.
Development of antibodies to HBV surface proteins and their virus neutralization capacity. (a) Anti-HBs antibody levels (IU/L) measured after the last booster immunization with Engerix-B and after the preS-based vaccination. (b) preS-specific IgG levels (OD) measured by ELISA after VVX001 immunization. (c) Percentages reduction of HBeAg secretion of infected HepG2 cells after pre-incubation of HBV inocula with sera obtained in the course of immunization compared to the baseline. Neutralization: ≥90% (strong neutralization), ≥50-90% (partial neutralization), ≥10-50% (low neutralization). .
Figure 2.
Development of antibodies to HBV surface proteins and their virus neutralization capacity. (a) Anti-HBs antibody levels (IU/L) measured after the last booster immunization with Engerix-B and after the preS-based vaccination. (b) preS-specific IgG levels (OD) measured by ELISA after VVX001 immunization. (c) Percentages reduction of HBeAg secretion of infected HepG2 cells after pre-incubation of HBV inocula with sera obtained in the course of immunization compared to the baseline. Neutralization: ≥90% (strong neutralization), ≥50-90% (partial neutralization), ≥10-50% (low neutralization). .
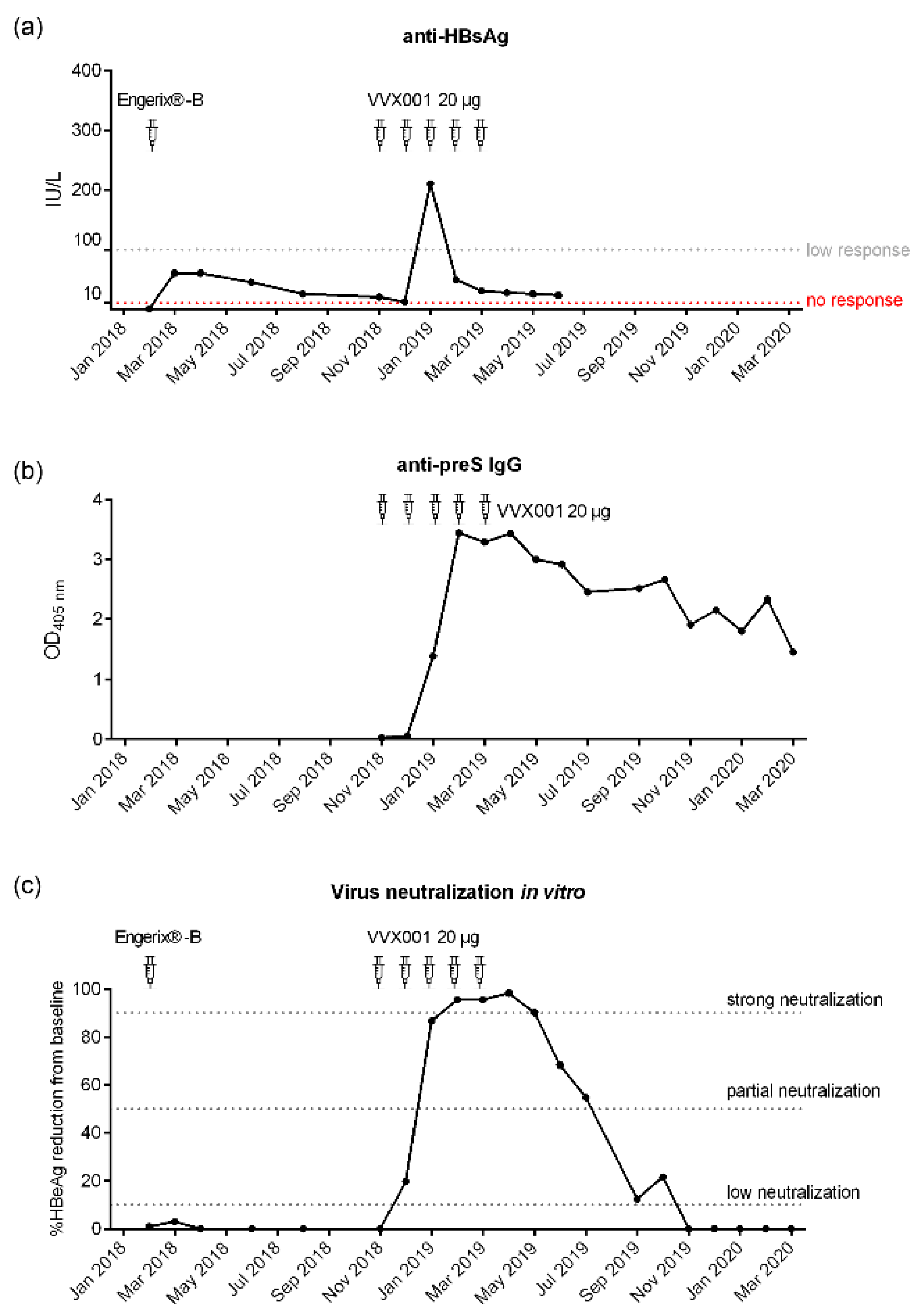
Table 1.
Quantification of the preS-specific IgG1 and IgG4 antibody concentrations.
| Time point | Date | IgG1, µg/mL | IgG4, µg/mL |
| preS-specific | |||
| Baseline (before VVX001 vaccination) 1 month after the last injection 4 months after the last injection 8 months after the last injection 12 months after the last injection |
19.11.2018 | 6.19 | 0 |
| 05.04.2019 | 1940.72 | 15 | |
| 26.06.2019 07.11.2019 12.03.2020 |
1347.55 868.05 679.51 |
16.4 4.4 0 |
|
| Peptide A-specific | |||
| Baseline (before VVX001 vaccination) 1 month after the last injection 4 months after the last injection 8 months after the last injection 12 months after the last injection |
19.11.2018 | 0 | 0 |
| 05.04.2019 26.06.2019 07.11.2019 12.03.2020 |
72.24 24.8 9.91 5.85 |
1.55 2.7 0 0 |
|
| Peptide B-specific | |||
| Baseline (before VVX001 vaccination) 1 month after the last injection 4 months after the last injection 8 months after the last injection 12 months after the last injection |
19.11.2018 | 0 | 0 |
| 05.04.2019 | 23.42 | 0 | |
| 26.06.2019 | 9 | 0 | |
| 07.11.2019 12.03.2020 |
4.45 2.85 |
0 0 |
|
| Peptide C-specific | |||
| Baseline (before VVX001 vaccination) 1 month after the last injection 4 months after the last injection 8 months after the last injection 12 months after the last injection |
19.11.2018 | 1.23 | 0 |
| 05.04.2019 26.06.2019 07.11.2019 12.03.2020 |
90.09 29.48 11.4 7.41 |
3.3 5 0 0 |
|
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



