
Submitted:
25 July 2024
Posted:
26 July 2024
You are already at the latest version
Abstract
Background: This study provides a comprehensive analysis of Acinetobacter baumannii in aquatic environments and fish microbiota by integrating culture-dependent methods, 16S metagenomics, and antibiotic resistance profiling. Methods: A total of 83 A. baumannii isolates were recovered using culture-dependent methods from intra-hospital infections (IHI), wastewater (WW) and surface water (SW) samples from two southern Romanian cities in august 2022. The antibiotic susceptibility was screened using disc diffusion, microdilution, PCR and Whole Genome Sequencing assays. Results: The highest microbial load in the analyzed samples was found in Glina, Bucharest for both WW and SW samples across all investigated phenotypes. For Bucharest isolates, the resistance levels corresponded to fluoroquinolones > aminoglycosides > β-lactam antibiotics. In contrast, A. baumannii from upstream SW samples in Targoviste showed the highest resistance to aminoglycosides. The blaOXA-23 gene was frequently detected in IHI, WW, and SW isolates in Bucharest but was absent in Targoviste. Molecular phylogeny revealed presence of ST10 in Târgoviște isolates and ST2 in Bucharest isolates, while other minor STs were not specifically correlated with a sampling point. Using 16S rRNA sequencing, significant differences in microbial populations between the two locations was identified. The low abundance of Alphaproteobacteria and Actinobacteria in both locations suggests environmental pressures or contamination events. Conclusions: These findings indicate significant fecal contamination and potential public health risks, emphasizing the need for improved water quality monitoring and management.
Keywords:
resistant Acinetobacter baumannii isolates
; wastewater and surface water microbial load
; surface water microbiome
; pangenome analysis
; Southern Romanian cities
1. Introduction
Increasing concerns about antibiotic resistance (AR) have led numerous groups of researchers to inquire about the effects of antibiotic resistant bacteria (ARB) and antibiotic resistance genes (ARGs) on human and animal health, agriculture, food production and waste management from the perspective of the One Health concept [1]. The One Health approach aims to achieve excellent health for people, animals, and the environment by addressing the spread of emerging infectious diseases at the animal-human-environment interface. Effective solutions require understanding and managing the complex interactions among these interconnected domains. The “One World – One Health” approach is based on four key components: ecological, geographic, human activities, and food-agricultural. Food processors, growers, and merchants are responsible for ensuring product quality. This quality encompasses not only the absence of pathogens but also the consideration of risks to humans at the top of the food chain. Preventive measures should begin at the start of the chain, with feed given to animals being free from contaminants such as mycotoxins and antibiotics [2]. AR is one of the major global healthcare crises of the 21st century. The imprudent use of antibiotics in both humans and animals has led to the emergence of antibiotic resistant bacteria (ARB). Wastewater treatment plants (WWTPs) and hospital environments, due to their high microbial load, have become reservoirs for ARGs and hotspots for the dissemination of AR into the environment. Conventional mechanical and biological wastewater treatment processes cannot completely eliminate all pollutants, leading to the release of pollutants into surface water bodies along with treated wastewater. Additionally, the disposal of waste and treated water from urban areas further increases the presence of resistance genes in surface water [3]. Transmission routes into the environment include: the use of raw or digested manure or sewage sludge as fertilizers on agricultural sectors, the use of treated wastewater for irrigation fields and discharging effluent from the WWTPs into the natural ecosystems [4]. During wastewater treatment, surplus sludge is produced, which highlights a high diversity of microorganisms, including pathogenic ones. Nosocomial infections are rapidly transmissible from one patient to another or even through medical employers. A. baumannii is an opportunistic nosocomial pathogen responsible for a wide range of infections, of which pneumonia and septicemia are the most common and included in the World Health Organization’s (WHO) priority list for research on drug-resistant bacteria and AR [5,6]. Multidrug resistance (MDR) is caused by several resistance markers in A. baumannii isolates, such as β-lactamase encoding genes (blaOXA-23, blaOXA-24, blaVIM, blaIMP, blaTEM, blaVEB, blaCTX-M, blaGES and blaPER). A. baumannii can develop resistance due to the chemical similarity between β-lactamase inhibitors and β-lactams [7].
However, the primary mechanism behind β-lactam resistance in A. baumannii involves the production of class D β lactamases. β-lactamases are bacterial hydrolases that bind and acylate β-lactam antibiotics. β-lactamases divide into four classes: the active-site serine β-lactamases (classes A, C and D) and the zinc-dependent or (metallo-β-lactamases, or MBLs) [8]. Class D β-lactamases (OXA enzymes) are narrow-spectrum lactamases that provide resistance to cefotaxime and ceftazidime. For A. baumannii the most common are class D oxacillinases (OXA type) β-lactamases (OXA-23, OXA-24, OXA-51, OXA-58, OXA-235, OXA-40) [9]. Class A β-lactamases identified in Acinetobacter spp. include β-lactamases such as TEM, SHV, CARB, CTX-M, PER, VEB, GES și KPC [10]. Among the MBLs detected in A. baumannii are IMP, VIM, SIM-1, SPM-1 and NDM [8]. Class C β-lactamases are cephalosporinases that hydrolyze most penicillins and narrow-spectrum cephalosporins [11]. Genotypic resistance may be the result of chromosomal mutations or may be due to the acquisition of genetic determinants. A. baumannii can acquire resistance to various antibiotic classes through chromosomal mutations and the horizontal transfer of ARGs. Mobile genetic elements are represented by transposons (Tn), insertion sequences (SI) integrons and resistance islands [12]. Four transposons carrying blaOXA-23 gene have been reported: Tn2006, Tn2007, Tn2008, and Tn2009. Tn2006 and Tn2008, associated with ISAba1 contribute to the dissemination of blaOXA-23. [13]. A variety of resistance islands were identified in A. baumannii, including AbaR1, AbaR3, AbaR5, AbaR6, AbaR7, AbaR8, AbaR9, and AbaR10. Specifically, the AbaR1 resistance island contains genes such as tet(A) a tetracycline efflux pump which confer tetracycline resistance, and respectively strA, strB, aphA1, and aac69, responsible for aminoglycosides resistance [14,15].
In recent years, international authorities have made significant efforts to enhance the monitoring of ARBs and ARGs across various environments. A key strategy in these efforts involves mapping the distribution of MDR nosocomial pathogens in different clinical settings and WWTPs [16]. Effluents from WWTPs are released to the surface water level. Therefore, a major risk factor that can affect human health is the contamination with antibiotic-resistant pathogens of plant crops irrigated with water from contaminated rivers [17].
For 2022, Romania reported very high levels of resistance in A. baumannii to fluoroquinolones, aminoglycosides, and carbapenems (ranked fourth and third, respectively, after countries such as Croatia, Greece, Cyprus, and Italy), according to the ECDC [18].
In this context, this study aims to provide a comprehensive analysis of Acinetobacter baumannii in aquatic environments and fish microbiota by integrating culture-dependent methods, 16S metagenomics, and antibiotic resistance profiling of recently isolated isolates from different sources in southern Romania.
2. Materials and Methods
2.1. Water Sampling Campaign
On 01.08.2022 and respectively 09.08.2022, 2 liters of wastewater and surface water samples were collected from two wastewater treatment plants (WWTPs) in southern Romania: Glina (n=4 samples), which collect wastewater from Bucharest (the capital city with 1.72 million inhabitants), and Târgoviște (n=4 samples, having 79,610 inhabitants). Samples were taken from the influent, IN; activated sludge, AS from the aeration tank; effluent, EF of both locations; and respectively upstream, UP (200 m) and downstream, DO (200 m) regions of the sampled WWTPs (Dâmbovița and Ialomița rivers, respectively) and transported at 4°C till the microbiology laboratory of the Faculty of Biology, University of Bucharest, Romania.
2.2. Strains Isolation, Quantification, Identification and Antimicrobial Susceptibility Profiles
The diluted samples up to a factor of 10^-5 were filtered through membrane filtration technique, and the filters were inoculated on chromogenic media (CHROMagar Acinetobacter, Paris, France) and on chromogenic media supplemented with carbapenem, cephalosporin and polymyxin antibiotics (CHROMagar CARBA; CHROMagar ESBL and CHROMagar Colistin, Paris, France), incubated at 37 °C for 24 h under aerobic conditions followed by determination of the colony-forming units number (CFU/100 mL) belonging to A. baumannii and to the Gram-negative non-fermenting bacilli (NF-GNB), considering filters with a number of white colonies ≤200 per culture medium and using the following relationship:
D- density or microbial load; N-total number of the colonies; V-volume x dilution
Next step was represented by the confirmation of carbapenemase (CP) and extended-spectrum β-lactamase (ESBL) producing isolates, and respectively colistin resistant ones by culturing up to 6 colonies for each phenotype on the same culture media and taxonomic identification of wastewater and surface water isolates, carried out using MALDI-TOF MS (Bruker, Germany). The isolates were preserved on broth (Mueller Hinton, Liofilchem, Italy) culture medium supplemented with 20% glycerol at -80°C.
During the same timeframe, a total of 17 A. baumannii isolates from intra-hospital infections (IHI) were isolated and identified using automated systems (VitekII Compact).
A total number of 83 A. baumannii isolates recovered from aquatic and clinical samples were tested for antibiotic susceptibility using the standard disc diffusion method, following the protocols outlined in the current editions of the Clinical and Laboratory Standards Institute (CLSI) guidelines pertinent to the isolation year [19]. The antibiotic susceptibility profiles of these isolates were tested to the following antibiotics: amikacin (30 µg); ampicillin-sulbactam (20 µg); aztreonam (30 µg); cefepime (30 µg); ceftazidime (30 µg); ciprofloxacin (5 µg); doripenem (10 µg); imipenem (10 µg); meropenem (10 µg); gentamicin (10 µg); and minocycline (30 µg). The antibiotic susceptibility results were interpreted according to the antibiotic classes, respectively for β-lactams, fluoroquinolones, aminoglycosides and tetracyclines.
For colistin susceptibility the microdilution method in Cation-Adjusted Mueller-Hinton Broth medium (CAMHB, OXOID, England) using standard 96-well microtiter plates by performing serial two-fold microdilutions of colistin sulfate (19.000 IU/mg, Sigma-Aldrich, Merck) in 75 μL of CAMHB medium (ranged between 128 - 0.25 μg/mL) according to the CLSI, 2022. The media was inoculated in the next step with a 75 μL of 0.5 McFarland suspension from 24-hour cultures grown at 37 °C on Plate Count Agar media. The positive (untreated cultures) and negative controls (sterility control) were included and the minimum inhibitory concentration (MIC) values were determined after incubating for 24 hours at 37 °C as being the last concentration for which no growth was recorded.
2.3. Characterization of Genotypic Resistance Profiles
The presence of carbapenem and cephalosporin encoding genes (blaVIM, blaIMP, blaNDM, blaOXA-23, blaOXA-24, blaOXA-58, blaOXA-235, blaOXA-51, blaKPC, blaGES, blaSHV, blaTEM, blaCTX-M, blaPER, and blaVEB) was investigated by simplex and multiplex PCR using DNA template extracted through an alkaline extraction method, specific primers and amplification programs and checked by gel electrophoresis [7,20].
2.4. Whole Genome Sequencing (WGS) and Bioinformatic Analyses of Clinical and Aquatic A. baumannii Isolates
From a total of 83 A. baumannii isolates recovered from two WWTPs and from Fundeni Bucharest Hospital in Romania, we performed WGS sequencing for a total of 20 isolates. The selection criteria were based on AR profiles (were selected isolates from all identified phenotypes) and isolation sources (from all sources: IN, EF, AS, UP and DO samples). Total DNA was extracted using DNeasy UltraClean Microbial Kit (Qiagen, Germany), followed by library preparations with Nextera DNA Flex Library Prep Kit (Illumina). The sequencing was performed on Illumina MiSeq and NextSeq platforms (V3, 600 cycles).
Hence, the raw reads were assembled de novo using Shovill v1.1.0 pipeline [21]. Furthermore, the resulting sequences were analyzed using ABRicate v1.0.0 [22] tool and the NCBI and VFDB [23] databases to determinate profiles of ARGs and virulence factors (VFs). The Multilocus Sequence Type (MLST) [24] method was utilized to determinate the sequence type (ST) of the isolates, in conformity with the Pasteur scheme. Moreover, Prokka v1.14.6 [25] tool was used to annotate the sequences of the selected isolates. The output generated by Prokka was then utilized as input for Roary v3.13.0 [26]. Newick tree, generated from Roary along with core and accessory genes, was illustrated using Phandango [27] online tool. Roary output was converted with the following script [29] for multidimensional scaling (MDS) and pangenome tree representation by FriPan [28]. Subsequently, the Heaps’ law was determined for data set, using Seth Commichaux’s Python script [30].
2.5. Metagenomic Analysis of Surface Water Samples and Fish Microbiota, to Highlight the Connection between the Environment and Fish Microbiota
To examine the microbiome from fish gut samples, DNA extraction was performed using the Pure Link Microbiome DNA Purification Kit (Invitrogen, Thermo Scientific, USA), following the manufacturer’s instructions. DNA extraction from water samples was carried out using the DNA Power Water Kit (Qiagen, Germany), according to the manufacturer’s instructions. The 16S rRNA sequences were then amplified using specific primer pairs for the V3-V4 hypervariable region of the 16S rRNA gene. The PCR products resulting from the amplification of the hypervariable regions of the 16S rRNA gene were purified using AmPure XP magnetic beads (Beckman Coulter, Inc.). Library preparation was conducted using the Ion Plus Fragment Library Kit (Life Technologies, USA), following the manufacturer’s instructions. The obtained amplicon libraries were sequenced on an Ion Torrent 316 chip using the Ion Torrent PGM system and the Ion Sequencing 400 Kit (Life Technologies, USA), adhering to the manufacturer’s instructions. The sequencing data obtained were processed using the Quantitative Insights Into Microbial Ecology (QIIME) pipeline, a tool used for microbiome sequencing data analysis, allowing the determination of microbiota composition and diversity. For calculating diversity measures, operational taxonomic units (OTUs) of the 16S rRNA gene were defined at a sequence similarity of at least 97%. The final analysis of the obtained sequences was performed using Ion Reporter software.
3. Results
3.1. Phenotypic and Genotypic AR Profiles of A. baumannii Isolates
3.1.1. Isolation and Quantification of A. baumannii from Romanian Wastewater and Surface Water Samples
The inoculation of diluted wastewater and surface water samples collected between 01-09.08.2022 from different sampling points of the investigated WWTPs (influent, active sludge, effluent) and surface water samples collected from the upstream and downstream region of the sampled WWTPs on chromogenic culture media (CHROMagar Acinetobacter for determining the total microbial load corresponding to A. baumannii) and chromogenic media supplemented with antibiotics (CHROMagar CARBA; CHROMagar ESBL and CHROMagar Colistin) allowed the determination of microbial load in the collected samples (Figure 1 and Figure 2; Supplementary Tables S3 and S4).
Analysis of the comparative levels of the microbial load in the analyzed samples from the receiving river revealed in decreasing order by the phenotype:
- CARBA phenotype: DO Glina, Bucharest > UP Targoviste > UP Glina, Bucharest > DO Targoviste;
- Colistin phenotype: DO Glina, Bucharest > DO Targoviste > UP Glina, Bucharest > UP Targoviste;
- ESBL and total Acinetobacter phenotype: DO Glina, Bucharest > DO Targoviste > UP; (Figure 1 and Supplementary Table S3).
Analysis of the comparative levels of the microbial load in the analyzed samples from the WWTPs revealed in decreasing order by the phenotype:
- 4)
- CARBA and ESBL phenotype: EF Glina, Bucharest > IN Glina, Bucharest > IN Targoviste >AS Glina, Bucharest > EF Targoviste;
- 5)
- total Acinetobacter phenotype: IN Glina, Bucharest > IN Targoviste > AS Glina, Bucharest > EF Glina, Bucharest > EF Targoviste; (Figure 2 and Supplementary Table S4).
3.1.2. Antimicrobial Susceptibilities Profiles of A. baumannii Isolates from Different Isolation Sources and by Geographical Location
In decreasing order, the resistance level of A. baumannii isolates recovered from intra-hospital infections (n=17) correspond to fluoroquinolones (100%), β-lactam (98% of the isolates), followed by aminoglycosides and tetracycline antibiotics (94 % of the isolates). The comparative study of the AR profiles according to the isolation sources and location demonstrated the following: for a total of 33 A. baumannii isolates recovered from Glina, Bucharest’s WWTP and the receiving river the highest resistance level corresponds to aminoglycosides in the case of the AS samples (57%); followed by the isolates isolated from EF (36%), and respectively IN and DO region (43%). For β-lactam antibiotics the resistance levels were in decreasing order as follows: AS, DO (31% of the isolates) > IN (25%) > EF (21%). In the case of fluoroquinolone antibiotics the resistance levels were attributed to the isolates in decreasing order as follows: AS samples (29%) > IN, DO samples (14%) > EF (9% of the isolates). The most susceptible A. baumannii correspond to tetracycline antibiotics (Figure 3 and Supplementary Table S5).
The resistance levels in the case of A. baumannii isolated from Targoviste shows that the aminoglycosides resistance was recorded at the top of the resistance level, with 54% of the isolates recovered from surface water sample from UP region, followed by A. baumannii isolated from IN, DO (50%) and isolates isolated from EF sample (36%). For β-lactam antibiotics the resistance level was observed to be on the decrease, in the following order: surface water sample from UP region (42% of the isolates) > IN (33%) > EF (31%) and DO (29% of the isolates). The resistance levels identified for fluoroquinolone were as follows: IN samples (33%) > UP samples (17% of the isolates). The most susceptible A. baumannii isolates were identified for tetracycline antibiotics, as illustrated in the Figure 4 and Supplementary Table S6.
Using the microdilution method in CAMHB were determined the MIC values for colistin susceptibility as following: in the case of one intra-hospital infection isolate (encoded 24 IHI Buc) within the international clone (IC) 2 (ST2) isolated from Fundeni hospital, was intermediate to colistin with a MIC value of 1 μg/mL. Resistance to colistin may be caused by mutation in genes encoding lipopolysaccharides (LPS), such as lpxA, lpxC and lpxD, as well as genes encoding for phosphoethanolamine transferase (PEtN), such as operon pmrCAB as previously demonstrated [31]. In this study, mutations were identified in lpxC (N286D), lpxD (E117K) and pmrB (A138T), the genes were compared to the A. baumannii ATCC 1906 genome (GenBank CP045110) [32]. The rest of the clinical and wastewater isolates belonging to epidemic ICs or non-IC were susceptible to colistin (MIC < 0.25 μg/mL) (see Table 1).
3.1.3. Genotypic Characterization of β-Lactam Resistance in Clinical, Wastewater and Surface Water A. baumannii Isolates
The comparative molecular study for a total of 33 A. baumannii stains recovered from Glina, Bucharest wastewater and surface water samples according to the isolation sources (Figure 4 and Supplementary Table S7) demonstrated the following: the most frequently encountered gene in investigated sources was blaOXA-23: AS (28% of the isolates) > EF (19%) and IN, DO (14% of the isolates). As anticipated, the blaOXA-51 gene was identified in all isolates isolated from all isolation sources. Nevertheless, isolates isolated from intra-hospital infections were positives for blaOXA-23 gene (62%) and blaOXA-24 gene (46% of the isolates). Additionally, the presence of the blaTEM gene was confirmed, although at a relatively lower prevalence (ranging from 15 to 14% in IHI and WWTP IF).
Figure 5.
β-lactamase producing A. baumannii isolates from Bucharest, Romania in 2022.
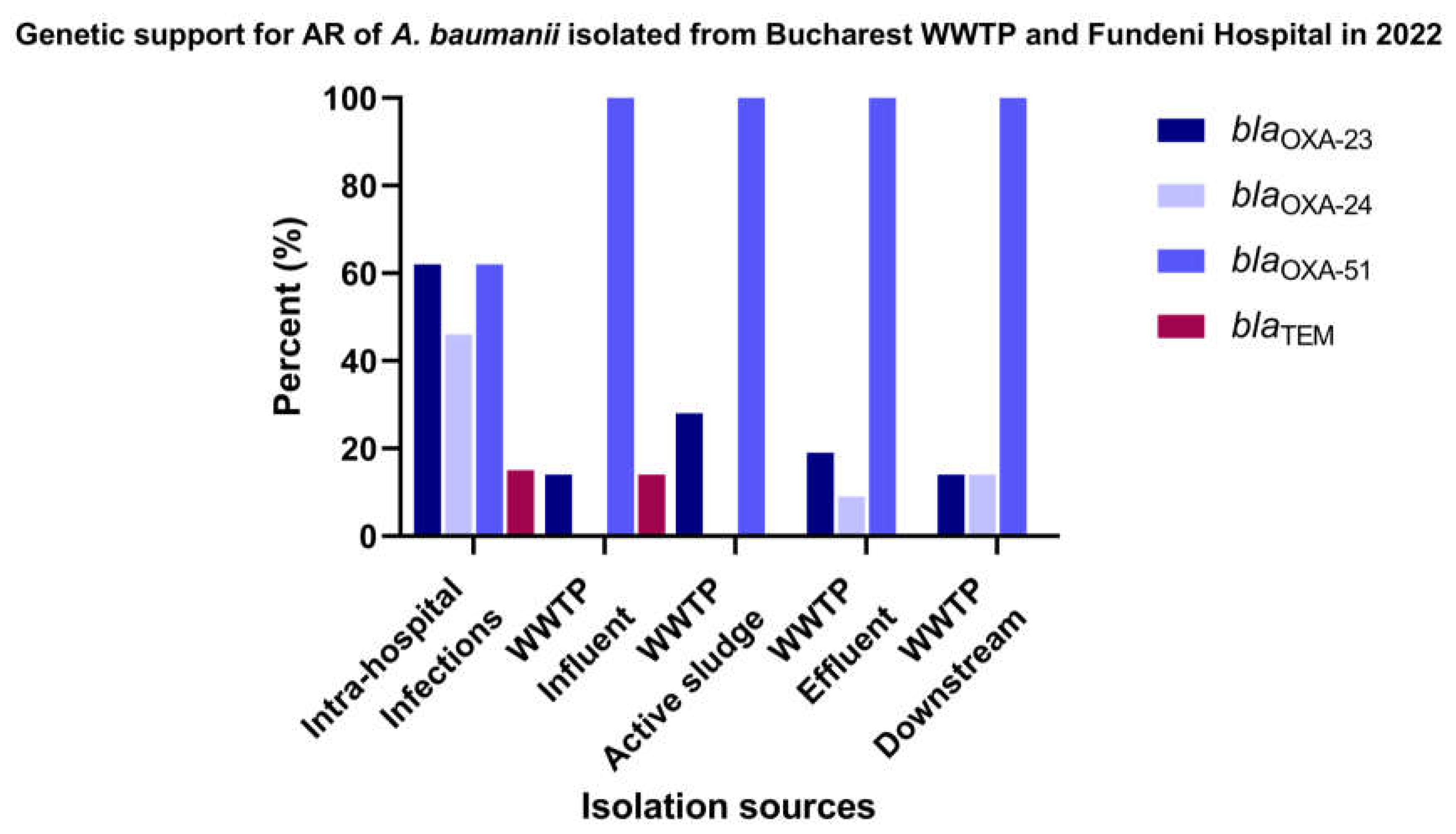
The genotypic characterization of β-lactam resistance performed in 33 isolates of A. baumannii isolated from Targoviste wastewater and surface water samples (Figure 6 and Supplementary Table S8) revealed that the blaCTX-M gene was identified in 13% of the isolates obtained from surface water samples collected from the DO region of the sampled WWTP. As expected, blaOXA-51 gene was identified in 100% of the isolates from all sources.
3.1.4. WGS Analysis in Clinical, Wastewater and Surface Water A. baumannii Isolates
WGS analysis of intra-hospital infections, wastewater surface water A. baumannii isolates revealed the presence of the following ARGs: ant(3’‘)-IIa, aph(3’‘)-Ib, aph(3’)-VIa, aph(6)-Id and armA for aminoglycosides resistance. Macrolide and tetracycline resistance was confirmed by the presence of mph(E), msr(E) and respectively tet(B) genes. In addition, sul1 and sul2 genes which encode resistance to sulfonamides, were also identified. Moreover, the genes encoding for chloramphenicol resistance, catA1 and cmlB1 genes, were identified in isolates from all sampled sources. Notably, blaOXA-23 and blaOXA-72 genes encoding for carbapenem resistance, were present in six of the tested isolates, including isolates from intra-hospital infections and wastewater samples from Bucharest. Furthermore, in one isolate belonging to IC2, isolated from Bucharest WWTP effluent (encoded 22014-CA2), both CP encoding genes mentioned above were identified. Nevertheless, the most prevalent genes, as expected, belonged to the blaADC and blaOXA-51 families, encoding for classes C and D β-lactamases. Additionally, the blaTEM gene, which encodes for class A broad spectrum β-lactamase, has also been identified in a wastewater A. baumannii isolate recovered from Bucharest WWTP effluent (Supplementary Table S9).
MLST analyses indicate that the most prevalent clones within the tested isolates from all isolation sources were IC2 (ST2, 25% of the sequenced isolates) followed by the IC8 (ST10, identified in the case of one isolate from wastewater and another one from a surface water samples from Targoviste) and IC7 (ST113 in the case of one wastewater isolate from Targoviste) according Shelenkov et al. classification [33]. Moreover, several other non-IC clones were identified, i.e., ST154 (surface water sample from Glina, Bucharest and wastewater isolate from Targoviste), ST150 (surface water sample from Targoviste), ST32 (wastewater isolate from Glina, Bucharest) (Figure 7).
Pangenome analysis was undertaken on 20 genomes of A. baumannii isolates from WW and SW in two cities in Romania, as well as from IHI. Notably, one cluster stands out from all 20 genomes (red color in Figure 8); it belongs to ST2 and was isolated from WW (n=2) and IHI (n=3) in Bucharest. The other isolates belong to various STs, such as ST150, ST10 and ST154, and correlations between isolations sources could not be established (Figure 7). Moreover, A. baumannii pangenome analysis identified a gene pool consisting of 7,644 genes in the sequences of all 20 isolates. Most of these genes (~60%) are classified as hypothetical proteins. The 5-isolate cluster has 224 unique genes, compared to the other 15 isolates in this study. Of these 224 genes, most of them are classified as hypothetical proteins (88%) but there are other genes that could be linked to antibiotic resistance (e.g., tetA, tetR (associated with Tn10) and other MGEs (Tn3 family transposase Tn2, Tn3 family transposase TnAs3, IS66 family transposase ISAba17). Following Heaps’ law, the pangenome of A. baumannii remains open, characterized by a γ = 0.26.
Using the VFDB database it was shown that circulating A. baumannii clones in Romanian intra-hospital infections from Bucharest and wastewater from two cities in southern Romania were associated with virulence factors encoding for adherence (ata, tufA), biofilm formation (algW, bap, csuA, csuB, csuC, csuD, csuE), effector delivery systems (tssA, B, C, D, E, F, G, K, L, M, tagX), exoenzymes (cpaA), immune modulation (cps4l, galE, galU, pseB, pseC, pseF, pseG, pseH, pseI, tviB, wbpD), nutrition/metabolic factors (bauA, hemO) [34]. The highest number of virulence factors were associated with clinical and wastewater isolates positives for OXA-23 and OXA23+OXA-72 CPs in the case of wastewater isolate isolated from Bucharest WWTP effluent and belonging to IC2: (40 and respectively 35 encoding genes). An interesting situation was observed in the case of the cpaA gene, identified in A. baumannii isolates recovered from wastewater and surface water samples from both investigated locations, and associated with ST154, ST903, ST32, and ST10 clones (Supplementary Table S10).
3.2. Metagenomic Analysis of Surface Water Samples and Fish Microbiota, to Highlight the Connection between the Environment and Fish Microbiota
The fish samples collected from Targoviste exhibited a microbiota predominantly composed of Aeromonadales species (28%), Flavobacteriaceae (10%), Enterobacteriaceae (5%), and Vibrionaceae (4%) (Figure 9). The phylum Alpha Proteobacteria was poorly represented, with an abundance of only 2%. The order Pseudomonadales was present in the gut samples with an abundance of 2% of the total number of sequences obtained. The fish samples collected from the Glina, Bucharest site were characterized by lower microbial diversity (based on Shannon diversity index values) (Figure 10). The microbiome at this site was predominantly composed of Aeromonadaceae species (56%) and a very high abundance of Fusobacteriaceae (15%). The percentage of Enterobacteriaceae (8%) was higher compared to that obtained for the samples collected from Targoviste, indicating a degree of fecal contamination, as this family of microorganisms is associated with the human microbiome (Figure 9).
The microbiota of the aquatic environment is typically composed of members of the phyla Alphaproteobacteria, Cytophaga / Flexibacter / Flavobacterium, and Actinobacteria. However, the samples collected from Targoviste and Glina, Bucharest exhibited a low abundance of the phyla Alphaproteobacteria and Actinobacteria.
The surface water samples from the UP region of Targoviste WWTP were characterized by a very high abundance of Burkholderiales (38%), Neisseriales (19%), and Pseudomonadales (9%). Additionally, Enterobacteriales, Aeromonadales, Alteromonadales, and Rhodobacterales were identified, albeit at lower abundances. The DO samples from Targoviste harbored Actinomycetales (6%), Aeromonadaceae (5%), and Flavobacteriaceae (4%), along with a high presence of Campylobacteraceae (6%), a family of microorganisms pathogenic to humans but commensal in some animals (e.g., chickens) (Figure 11 A).
In the UP water samples collected from Glina, Bucharest WWTP, 16S rRNA sequencing revealed high levels of Burkholderiales (11%), Oscillatoriales (6%), Flavobacteriales (4%), Actinomycetales (8%), and Cytophagales (3%). The DO samples were characterized by the presence of pathogenic microorganisms (Campylobacteraceae - 11%) and microorganisms typical of the human microbiota such as Enterobacteriaceae, Bacteroidaceae, Prevotellaceae, and Lactobacillales. Additionally, species of the families Pseudomonadaceae (4%) and Moraxellaceae (6%) were identified (Figure 11 B).
Furthermore the analysis of the 16S rRNA metagenomic data from surface water samples from DO regions of Glina, Bucharest and Targoviste WWTPs reveals significant insights into the microbial communities present in these environments. The focus on the Moraxellaceae family, particularly the genus Acinetobacter, provides a detailed understanding of bacterial diversity and prevalence.
In the water samples collected from DO region of Glina, Bucharest WWTPs Acinetobacter was identified as a significant component of the microbial community within the Moraxellaceae family. Specifically, Acinetobacter constituted 43% of the Moraxellaceae reads. This substantial presence highlights the potential environmental impact and resilience of Acinetobacter species in this downstream water ecosystem (Figure 12).
Similarly, the water samples from Targoviste revealed an even higher prevalence of Acinetobacter within the Moraxellaceae family. In this location, Acinetobacter accounted for 52% of the Moraxellaceae reads. This higher percentage indicates a robust population of Acinetobacter species in the Targoviste downstream waters, suggesting possible variations in environmental conditions or anthropogenic influences that favor the proliferation of this genus (Figure 12).
4. Discussion
This paper analyzes A. baumannii in aquatic environments and fish microbiota using culture-dependent methods, 16S metagenomics, and antibiotic resistance profiling of recent isolates from Glina, Bucharest and Targoviste WWTP and the receiving rivers (Dambovita and Ialomița) southern Romania. Two WWTPs were selected to represent different pollution sources: urbanized city, wastewater discharges in Bucharest, capital city of Romania and respectively anthropogenic activities and animal waste from a dog shelter în Targoviște. Eight water samples (four from each location) were collected for A. baumannii isolation, identified using MALDI-TOF mass spectrometry, further investigated for AR by phenotypic and genotypic assays and 16S metagenomics. Previously, monitoring the quality parameters for five sections of the Dâmboviţa river, both upstream and downstream of Bucharest, showed that the river’s overall ecological state falls into quality classes III–V (poor to bad quality). The worst conditions corresponded for the DO region of Bucharest, which received partially treated wastewater from the Bucharest WWTP [35]. The Ialomita River’s water quality, monitored along its length, ranged from very good to very poor (classes I to V). After 2010, the water quality improved, with only the DO region showing a moderate status [36]. Using culture-dependent assays, we demonstrated that the highest microbial load in the analyzed samples (wastewater from the IN, AS, EF sources, and surface water from the UP and DO regions of the Dambovita and Ialomita rivers) was found in Bucharest for both wastewater (WWTP EF and WWTP IN) and surface water samples (DO region of the WWTP Bucharest) across all investigated phenotypes (CARBA, ESBL, colistin and total Acinetobacter population). From both investigated locations a total of 66 A. baumannii isolates (33 for each location) were obtained through culture dependent assays from wastewater and surface water samples and compared with 17 isolates recovered in 2022 from IHI to evaluate the circulating clones in different isolation sources. In both WWTPs, the highest resistance levels were found for aminoglycoside antibiotics, followed by β-lactams and fluoroquinolones, with resistance levels varying by location and isolation source. In the case of IHI isolates the resistance levels in decreasing order corresponded to fluoroquinolones > β-lactam > aminoglycosides and tetracycline antibiotics. The investigated isolates for colistin resistance have demonstrated that only in the case of an IHI isolate belonging to ST2 isolated from a large hospital in Bucharest, where patients from all over the country are admitted, was intermediate, while the rest of the isolates were sensitive to colistin. Previous data have reported that ST2 is the most prevalent ST associated with colistin resistance in A. baumannii across Europe, Asia, Africa, and North and South America [32], [37,38,39,40,41,42,43,44,45,46,47,48].
In Romania, the most frequently detected bacterial isolates with clinical relevance include Klebsiella spp., A. baumannii, Escherichia coli, Staphylococcus aureus, and Pseudomonas aeruginosa, all showing MDR phenotype. A. baumannii, associated with nosocomial infections such as pneumonia, meningitis, and urinary tract infections, was a focus of a 2018 study aimed at identifying microorganisms responsible for pneumonia in patients at an emergency hospital in Bucharest. Antimicrobial susceptibility testing for A. baumannii revealed high resistance rates: 88% to fluoroquinolones (including ciprofloxacin), 86% to β-lactam antibiotics (meropenem), and 86% to aminoglycosides (including amikacin) [49]. The transmission of ARGs among human, animal, and environmental reservoirs is a significant concern, with WWTPs being critical reservoirs for the spread of these genes. For A. baumannii isolates isolated from Romanian WWTPs, the highest resistance rates were recorded for fluoroquinolones (87.5% to ciprofloxacin), followed by aminoglycosides (86% to gentamicin and amikacin), and β-lactam antibiotics (84% to aztreonam and meropenem) [17].Viable MDR and carbapenem-resistant A. baumannii were detected in urban wastewater, which included hospital wastewater, both before and after secondary wastewater treatment [50]. Other studies have highlighted the presence of putative carbapenem resistant Acinetobacter isolates detected in all WWTP samples, except the primary sludge. Also, studies have revealed that A. baumannii isolates were resistant to fluoroquinolones, aminoglycosides, β-lactams and polymyxins in different sampling points of urban WWTPs [51].
The WGS analysis of A. baumannii isolates from investigated locations revealed both shared and unique characteristics, i.e., ant(3’‘)-IIa gene in all isolation sources from both locations opposite to several CPs: OXA-23 (Bucharest IHI and wastewater); OXA-72 (Bucharest wastewater); OXA-23+OXA-72 (Bucharest wastewater); TEM-1 (Bucharest wastewater); OXA-121 (Targoviste surface water); OXA-120 (Targoviste wastewater). In a study carried out in Croatia on A. baumannii isolates recovered from wastewater samples, the carbapenem-resistant isolates were positive for blaOXA-23 gene and belonged similarly to our obtained results to IC2 and the susceptible ones to IC5. Furthermore, these isolates revealed resistance genes encoding for chloramphenicol, aminoglycosides and tetracycline antibiotics [52]. In another study, in eastern Poland using conventional methods and metagenomic assays were demonstrated the presence of Acinetobacter spp. and A. baumannii isolates carrying MBL (VIM2, NDM and IMP-1)) and class D β-lactamases (OXA-23, OXA-24, OXA-51, OXA-58) in wastewater and river water samples collected in June and September in 2019. High frequency of isolation of A. baumannii in IHI, positive for OXA-23 CP and belonging to ST2, was described also in two Bulgarian hospitals, Romania’s neighboring country. The CHLDs linked to IC2 were reported also in clinical A. baumannii in other neighboring countries of Romania: OXA-23 and OXA-72 in Serbia; and OXA-23 in Albania, OXA-23, OXA-58, and OXA-72 CPs in Croatia, Serbia, Bosnia and Herzegovina.
Molecular typing of A. baumannii isolates revealed the presence of four distinct clusters: the ST2 cluster was found in isolates from Bucharest, indicating a localized prevalence in this region, while the cluster containing the ST10 clone was primarily associated with isolates from Targoviște, suggesting a regional specificity for this sequence type. A. baumannii ST10 has been found in clinical and community acquired infections globally, including USA [53], Vietnam [54], Iran [55], Australia [56], Belgium [57] and Germany [58]. Moreover, pangenome analysis demonstrated that the genomes of A. baumannii are open, as indicated by Heaps’ law (γ = 0.26). An open genome indicates that the gene pool of a isolate has not reached an upper limit, thus allowing the acquisition of new genes through transposable elements [59]. This finding was corroborated by Gherghe-Barbu and collaborators, who used the same tool (e.g., Seth Commichaux’s Python script) to analyze pangenome of A. baumannii isolated from WW and clinical samples in Targoviste and Ramnicu Valcea, where the value was γ = 0.41 [60].
The low abundance of Alphaproteobacteria and Actinobacteria in both Targoviste and Glina, Bucharest samples contrasts with the typical composition of aquatic microbiota [61]. This deviation could be indicative of specific environmental pressures or contamination events affecting these communities. Factors such as pollution, nutrient loads, or other anthropogenic activities could be influencing the microbial balance.
The dominance of certain bacterial orders in UP versus DO samples highlights the impact of local environmental conditions and potential sources of contamination. The presence of pathogenic bacteria like Campylobacteraceae in both locations underscores public health concerns, especially regarding the use of these water bodies for recreational or agricultural purposes.
The elevated levels of Enterobacteriaceae and other human-associated bacteria in DO samples suggest fecal contamination, likely from sewage discharge or runoff from agricultural lands. The presence of pathogenic bacteria like Campylobacteraceae indicates a risk of waterborne diseases, necessitating stringent water quality monitoring and management strategies [62].
The combined analysis of 16S rRNA metagenomic data and chromogenic culture media findings underscores the significant presence of Acinetobacter in surface water samples from DO region of Glina, Bucharest and Targoviste WWTPs. This high prevalence is associated with elevated microbial loads and significant resistance phenotypes (CARBA, ESBL, and colistin), especially in the receiving river from DO regions. These insights are important for developing strategies to monitor and mitigate the spread of ARB in the environment, ensuring public health safety and effective wastewater treatment practices.
Limitations of this study may arise from the fact that samples were collected at a single point in time, which does not account for seasonal or temporal variations in microbial load and resistance patterns, that could affect the generalizability of the findings, as well as the fact that the study did not extensively analyze environmental factors such as water temperature, pH, or nutrient levels, which could influence the microbial communities and antibiotic resistance patterns.
5. Conclusions
The findings further underscore WWTPs as reservoirs for MDR A. baumannii, highlighting the potential for environmental dissemination and public health risks. Deviations in microbial community composition suggest specific environmental pressures, necessitating stringent water quality monitoring and integrated surveillance strategies to mitigate public health risks associated with fecal contamination and pathogenic bacteria.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Table S1: A. baumannii isolates from Bucharest wastewater, surface water and intra-hospital infection samples; Table S2: A. baumannii isolates from Targoviste wastewater and surface water samples; Table S3: CFU/100 mL in surface water samples; Table 4: CFU/100 mL in wastewater samples; Table 5: Antimicrobial resistance profiles of A. baumannii isolated from Bucharest WWTP and Fundeni Hospital in 2022; Table S6: Antimicrobial resistance profiles of A. baumannii isolated from Târgoviște WWTP in 2022; Table S7: Genetic support for AR of A. baumannii isolated from Bucharest WWTP and Fundeni Hospital in 2022; Table S8: Genetic support for the AR of A. baumanii isolated from Târgoviște WWTP în 2022; Table S9: ARGs profiles for A. baumannii isolated from WWTPs and Fundeni Hospital in Romania; Table S10: Virulence factors profiles for A. baumannii isolated intra-hospital infections and WWTPs in Romania.
Author Contributions
Conceptualization, I.G.-B., I.C.B., G.P.G., and R.I.D.; methodology, I.G.-B., I.C.B., G.P.G., M.S.; R.I.D.; I.A.D.; and M.D.G..; software, I.C.B.; R.I.D.; M.S.; and G.P.G.; ; validation, M.S.; R.I.D.; and I.C.B.; formal analysis, M.S.; I.A.D.; M.D.G.; R.I.D.; G.P.G.; I.C.B.; and I.G.-B.; investigation, M.S.; I.A.D.; M.D.G.; R.I.D.; G.P.G.; I.C.B.; and I.G.-B.; resources, I.G.-B.; and G.P.G; data curation, R.I.D.; and M.S.; writing—original draft preparation, M.S.; I.A.D.; M.D.G.; R.I.D.; G.P.G.; I.C.B.; and I.G.-B.; writing—review and editing, M.S.; R.I.D.; G.P.G.; I.C.B.; and I.G.-B.; project administration, I.G.-B. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by PN–III–P4-PCE-2021-1797 (PCE 96/2022); ERANET 243/2021 PN-III-P1-1.1-TE-2021-1515 (TE112/2022) and PN-III-P1-1.1-PD-2021-0540 (PD102/2022) research projects awarded by UEFISCDI.
Institutional Review Board Statement
Agreement of the Bioethics Commission: No. 9/8121; 2 October 2018.
Data Availability Statement
A. baumannii isolates and data sets regarding the results are available from the authors.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Drane, K.; Sheehan, M.; Whelan, A.; Ariel, E.; Kinobe, R. The Role of Wastewater Treatment Plants in Dissemination of Antibiotic Resistance: Source, Measurement, Removal and Risk Assessment. Antibiotics 2024, 13, 668. [Google Scholar] [CrossRef] [PubMed]
- Calistri, P.; Iannetti, S.L.; Danzetta, M.; Narcisi, V.; Cito, F.; Di Sabatino, D.; Bruno, R.; Sauro, F.; Atzeni, M.; Carvelli, A.; Giovannini, A. The Components of “One World - One Health” Approach. Transboundary and Emerging Diseases. November 2013, pp 4–1. [CrossRef]
- Amos, G.C.A.; Ploumakis, S.; Zhang, L.; Hawkey, P.M.; Gaze, W.H.; Wellington, E.M.H. The Widespread Dissemination of Integrons throughout Bacterial Communities in a Riverine System. ISME Journal 2018, 12, 681–691. [Google Scholar] [CrossRef]
- Pérez-Valera, E.; de Melo Rangel, W.; Elhottová, D. Cattle Manure Application Triggers Short-Term Dominance of Acinetobacter in Soil Microbial Communities. Applied Soil Ecology 2022, 176, 104466. [Google Scholar] [CrossRef]
- Poirel, L.; Naas, T.; Nordmann, P. Diversity, Epidemiology, and Genetics of Class D β-Lactamases. Antimicrob Agents Chemother 2010, 54, 24–38. [Google Scholar] [CrossRef] [PubMed]
- Blanco, N.; Harris, A.D.; Rock, C.; Johnson, J.K.; Pineles, L.; Bonomo, R.A.; Srinivasan, A.; Pettigrew, M.M.; Thom, K.A. Risk Factors and Outcomes Associated with Multidrug-Resistant Acinetobacter baumannii upon Intensive Care Unit Admission. Antimicrob Agents Chemother 2018, 62, 10–1128. [Google Scholar] [CrossRef] [PubMed]
- Gheorghe, I.; Cristea, V.C.; Marutescu, L.; Popa, M.; Murariu, C.; Trusca, B.S.; Borcan, E.; Ghita, C.; Lazar, V.; Chifiriuc, M.C. Resistance and Virulence Features in Carbapenem-Resistant Acinetobacter baumannii Community Acquired and Nosocomial Isolates in Romania. [CrossRef]
- Tooke, C.L.; Hinchliffe, P.; Bragginton, E.C.; Colenso, C.K.; Hirvonen, V.H.A.; Takebayashi, Y.; Spencer, J. β-Lactamases and β-Lactamase Inhibitors in the 21st Century. J Mol Biol 2019, 431, 3472–3500. [Google Scholar] [CrossRef] [PubMed]
- Nigro, S.J.; Hall, R.M. Structure and Context of Acinetobacter Transposons Carrying the Oxa23 Carbapenemase Gene. Journal of Antimicrobial Chemotherapy 2016, 71, 1135–1147. [Google Scholar] [CrossRef]
- Philippon, A.; Slama, P.; Dény, P.; Labia, R. A Structure-Based Classification of Class A β-Lactamases, a Broadly Diverse Family of Enzymes. Clin Microbiol Rev 2016, 29, 29–57. [Google Scholar] [CrossRef]
- Bouza, A.A.; Swanson, H.C.; Smolen, K.A.; VanDine, A.L.; Taracila, M.A.; Romagnoli, C.; Caselli, E.; Prati, F.; Bonomo, R.A.; Powers, R.A.; Wallar, B.J. Structure-Based Analysis of Boronic Acids as Inhibitors of Acinetobacter-Derived Cephalosporinase-7, a Unique Class C β-Lactamase. ACS Infect Dis 2018, 4, 325–336. [Google Scholar] [CrossRef]
- Vrancianu, C.O.; Pelcaru, C.F.; Alistar, A.; Gheorghe, I.; Marutescu, L.; Popa, M.; Czobor, I.; Gradisteanu, G.; Dobre, E.G.; Chifiriuc, M.C. Escaping from ESKAPE. Clinical Significance and Antibiotic Resistance Mechanisms in Acinetobacter baumannii: A Review. Biointerface Research in Applied Chemistry. AMG Transcend Association January 1, 2021, pp 8190–8203. [CrossRef]
- Chen, Y.; Gao, J.; Zhang, H.; Ying, C. Spread of the blaOXA-23-Containing Tn2008 in Carbapenem-Resistant Acinetobacter baumannii Isolates Grouped in CC92 from China. Front Microbiol 2017, 8. [Google Scholar] [CrossRef]
- Naderi, G.; Talebi, M.; Gheybizadeh, R.; Seifi, A.; Ghourchian, S.; Rahbar, M.; Abdollahi, A.; Naseri, A.; Eslami, P.; Douraghi, M. Mobile Genetic Elements Carrying Aminoglycoside Resistance Genes in Acinetobacter baumannii Isolates Belonging to Global Clone 2. Front Microbiol 2023, 14. [Google Scholar] [CrossRef] [PubMed]
- Fournier, P.-E.; Vallenet, D.; Barbe, V.; Audic, S.; Ogata, H.; Poirel, L.; Richet, H.; Robert, C.; Mangenot, S.; Abergel, C.; Nordmann, P.; Weissenbach, J.; Raoult, D.; Claverie, J.-M. Comparative Genomics of Multidrug Resistance in Acinetobacter baumannii. PLoS Genet 2006, 2, e7. [Google Scholar] [CrossRef] [PubMed]
- Barbu, I.C.; Gheorghe-Barbu, I.; Grigore, G.A.; Vrancianu, C.O.; Chifiriuc, M.C. Antimicrobial Resistance in Romania: Updates on Gram-Negative ESCAPE Pathogens in the Clinical, Veterinary, and Aquatic Sectors. Int J Mol Sci 2023, 24, 7892. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg Goldstein, R.E.; Micallef, S.A.; Gibbs, S.G.; George, A.; Claye, E.; Sapkota, A.; Joseph, S.W.; Sapkota, A.R. Detection of Vancomycin-Resistant Enterococci (VRE) at Four U.S. Wastewater Treatment Plants That Provide Effluent for Reuse. Science of The Total Environment 2014, 466–467, 404–411. [Google Scholar] [CrossRef] [PubMed]
- https://atlas.ecdc.europa.eu/public/index.aspx (Accessed 15 July 2024).
- CLSI, Performance Standards for Antimicrobial Susceptibility Testing. 32nd Ed. CLSI Supplement M100, Clinical Laboratory Standard Institute, 2022.
- Gheorghe-Barbu, I.; Corbu, V.M.; Vrancianu, C.O.; Marinas, I.C.; Popa, M.; Dumbravă, A. Ștefania; Niță-Lazăr, M.; Pecete, I.; Muntean, A.A.; Popa, M.I.; Marinescu, L.; Ficai, D.; Ficai, A.; Czobor Barbu, I. Phenotypic and Genotypic Characterization of Recently Isolated Multidrug-Resistant Acinetobacter baumannii Clinical and Aquatic Isolates and Demonstration of Silver Nanoparticle Potency. Microorganisms 2023, 11. [Google Scholar] [CrossRef]
- https://github.com/tseemann/shovill (Accessed 2 July 2024).
- https://github.com/tseemann/abricate (Accessed 2 July 2024).
- Chen, L.; Zheng, D.; Liu, B.; Yang, J.; Jin, Q. VFDB 2016: Hierarchical and Refined Dataset for Big Data Analysis—10 Years On. Nucleic Acids Res 2016, 44, D694–D697. [Google Scholar] [CrossRef]
- Seemann, T. Mlst (v2.19.0), 2024. Github. https://github.com/tseemann/mlst, (Accessed 1 July 2024).
- Seemann, T. Prokka: Rapid Prokaryotic Genome Annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef] [PubMed]
- Page, A.J.; Cummins, C.A.; Hunt, M.; Wong, V.K.; Reuter, S.; Holden, M.T.G.; Fookes, M.; Falush, D.; Keane, J.A.; Parkhill, J. Roary: Rapid Large-Scale Prokaryote Pan Genome Analysis. Bioinformatics 2015, 31, 3691–3693. [Google Scholar] [CrossRef] [PubMed]
- Hadfield, J.; Croucher, N.J.; Goater, R.J.; Abudahab, K.; Aanensen, D.M.; Harris, S.R. Phandango: An Interactive Viewer for Bacterial Population Genomics. Bioinformatics 2018, 34, 292–293. [Google Scholar] [CrossRef]
- https://drpowell.github.io/FriPan/ (Accessed 12 July 2024).
- https://github.com/kwongj/roary2fripan (Accessed 12 July 2024).
- https://github.com/SethCommichaux/Heap_Law_for_Roary (Accessed 3 July 2024).
- Novović, K.; Jovčić, B. Colistin Resistance in Acinetobacter baumannii: Molecular Mechanisms and Epidemiology. Antibiotics. MDPI March 1, 2023. [CrossRef]
- Jovcic, B.; Novovic, K.; Dekic, S.; Hrenovic, J. Colistin Resistance in Environmental Isolates of Acinetobacter baumannii. Microbial Drug Resistance 2021, 27, 328–336. [Google Scholar] [CrossRef]
- Shelenkov, A.; Akimkin, V.; Mikhaylova, Y. International Clones of High Risk of Acinetobacter Baumannii—Definitions, History, Properties and Perspectives. Microorganisms 2023, 11. [Google Scholar] [CrossRef] [PubMed]
- http://www.mgc.ac.cn/cgi-bin/VFs/genus.cgi?Genus=Acinetobacter. (Accessed 1 July 2024).
- Zaharia, L.; Ioana-toroimac, G.; Cocoş, O.; Ghiţă, F.A.; Mailat, E. Urbanization Effects on the River Systems in the Bucharest City Region (Romania). Ecosystem Health and Sustainability 2016, 2, e01247. [Google Scholar] [CrossRef]
- Manoiu, V.-M. Water Quality Changes in Ialomiţa River under the Influence of Human Settlements and Activities. In Aerul şi Apa: Componente ale Mediului; 2017, 2017, 317–324. [CrossRef]
- Ušjak, D.; Novović, K.; Filipić, B.; Kojić, M.; Filipović, N.; Stevanović, M.M.; Milenković, M.T. In Vitro Colistin Susceptibility of Pandrug-resistant Acinetobacter baumannii Is Restored in the Presence of Selenium Nanoparticles. J Appl Microbiol 2022, 133, 1197–1206. [Google Scholar] [CrossRef] [PubMed]
- Cafiso, V.; Stracquadanio, S.; Lo Verde, F.; Gabriele, G.; Mezzatesta, M.L.; Caio, C.; Pigola, G.; Ferro, A.; Stefani, S. Colistin Resistant Acinetobacter baumannii: Genomic and Transcriptomic Traits Acquired Under Colistin Therapy. Front Microbiol 2019, 9. [Google Scholar] [CrossRef]
- Kabic, J.; Novovic, K.; Kekic, D.; Trudic, A.; Opavski, N.; Dimkic, I.; Jovcic, B.; Gajic, I. Comparative Genomics and Molecular Epidemiology of Colistin-Resistant Acinetobacter baumannii. Comput Struct Biotechnol J 2023, 21, 574–585. [Google Scholar] [CrossRef] [PubMed]
- Mustapha, M.M.; Li, B.; Pacey, M.P.; Mettus, R.T.; McElheny, C.L.; Marshall, C.W.; Ernst, R.K.; Cooper, V.S.; Doi, Y. Phylogenomics of Colistin-Susceptible and Resistant XDR Acinetobacter baumannii. Journal of Antimicrobial Chemotherapy 2018, 73, 2952–2959. [Google Scholar] [CrossRef] [PubMed]
- Trebosc, V.; Gartenmann, S.; Tötzl, M.; Lucchini, V.; Schellhorn, B.; Pieren, M.; Lociuro, S.; Gitzinger, M.; Tigges, M.; Bumann, D.; Kemmer, C. Dissecting Colistin Resistance Mechanisms in Extensively Drug-Resistant Acinetobacter baumannii Clinical Isolates. mBio 2019, 10, 10–1128. [Google Scholar] [CrossRef]
- Fam, N.S.; Gamal, D.; Mohamed, S.H.; Wasfy, R.M.; Soliman, M.S.; El-Kholy, A.A.; Higgins, P.G. Molecular Characterization of Carbapenem/ Colistin-Resistant Acinetobacter baumannii Clinical Isolates from Egypt by Whole-Genome Sequencing. Infect Drug Resist 2020, 13, 4487–4493. [Google Scholar] [CrossRef]
- Palmieri, M.; D’Andrea, M.M.; Pelegrin, A.C.; Perrot, N.; Mirande, C.; Blanc, B.; Legakis, N.; Goossens, H.; Rossolini, G.M.; van Belkum, A. Abundance of Colistin-Resistant, OXA-23- and ArmA-Producing Acinetobacter baumannii Belonging to International Clone 2 in Greece. Front Microbiol 2020, 11. [Google Scholar] [CrossRef]
- Thadtapong, N.; Chaturongakul, S.; Soodvilai, S.; Dubbs, P. Colistin and Carbapenem-Resistant Acinetobacter baumannii Aci46 in Thailand: Genome Analysis and Antibiotic Resistance Profiling. Antibiotics 2021, 10, 1054. [Google Scholar] [CrossRef]
- Pournaras, S.; Poulou, A.; Dafopoulou, K.; Chabane, Y.N.; Kristo, I.; Makris, D.; Hardouin, J.; Cosette, P.; Tsakris, A.; Dé, E. Growth Retardation, Reduced Invasiveness, and Impaired Colistin-Mediated Cell Death Associated with Colistin Resistance Development in Acinetobacter baumannii. Antimicrob Agents Chemother 2014, 58, 828–832. [Google Scholar] [CrossRef]
- Agodi, A.; Voulgari, E.; Barchitta, M.; Quattrocchi, A.; Bellocchi, P.; Poulou, A.; Santangelo, C.; Castiglione, G.; Giaquinta, L.; Romeo, M.A.; Vrioni, G.; Tsakris, A. Spread of a Carbapenem- and Colistin-Resistant Acinetobacter baumannii ST2 Clonal Isolate Causing Outbreaks in Two Sicilian Hospitals. Journal of Hospital Infection 2014, 86, 260–266. [Google Scholar] [CrossRef] [PubMed]
- Lowe, M.; Singh-Moodley, A.; Ismail, H.; Thomas, T.; Chibabhai, V.; Nana, T.; Lowman, W.; Ismail, A.; Chan, W.Y.; Perovic, O. Molecular Characterisation of Acinetobacter baumannii Isolates from Bloodstream Infections in a Tertiary-Level Hospital in South Africa. Front Microbiol 2022, 13. [Google Scholar] [CrossRef]
- Nogbou, N.-D.; Ramashia, M.; Nkawane, G.M.; Allam, M.; Obi, C.L.; Musyoki, A.M. Whole-Genome Sequencing of a Colistin-Resistant Acinetobacter baumannii Isolate Isolated at a Tertiary Health Facility in Pretoria, South Africa. Antibiotics 2022, 11, 594. [Google Scholar] [CrossRef]
- Blejan, I.E.; Diaconu, C.E.; Arsene, A.L.; Udeanu, D.I.; Ghica, M.; Drăgănescu, D.; Dragomiroiu, G.T.A.B.; Rădulescu, M.; Maltezou, H.C.; Tsatsakis, A.M.; Papasavva, M.; Drakoulis, N.; Popa, D.E. Antibiotic Resistance in Community-Acquired Pneumonia. A Romanian Perspective. Farmacia 2020, 68, 512–520. [Google Scholar] [CrossRef]
- Goic-Barisic, I.; Hrenovic, J.; Kovacic, A.; Musić, M.Š. Emergence of Oxacillinases in Environmental Carbapenem-Resistant Acinetobacter baumannii Associated with Clinical Isolates. Microbial Drug Resistance 2016, 22, 559–563. [Google Scholar] [CrossRef]
- Pulami, D.; Kämpfer, P.; Glaeser, S.P. High Diversity of the Emerging Pathogen Acinetobacter Baumannii and Other Acinetobacter Spp. in Raw Manure, Biogas Plants Digestates, and Rural and Urban Wastewater Treatment Plants with System Specific Antimicrobial Resistance Profiles. Science of The Total Environment 2023, 859, 160182. [Google Scholar] [CrossRef]
- Higgins, P.G.; Hrenovic, J.; Seifert, H.; Dekic, S. Characterization of Acinetobacter baumannii from Water and Sludge Line of Secondary Wastewater Treatment Plant. Water Res 2018, 140, 261–267. [Google Scholar] [CrossRef]
- Jones, C.L.; Clancy, M.; Honnold, C.; Singh, S.; Snesrud, E.; Onmus-Leone, F.; McGann, P.; Ong, A.C.; Kwak, Y.; Waterman, P.; Zurawski, D.V.; Clifford, R.J.; Lesho, E. Fatal Outbreak of an Emerging Clone of Extensively Drug-Resistant Acinetobacter baumannii With Enhanced Virulence. Clinical Infectious Diseases 2015, 61, 145–154. [Google Scholar] [CrossRef] [PubMed]
- Schultz, M.B.; Pham Thanh, D.; Tran Do Hoan, N.; Wick, R.R.; Ingle, D.J.; Hawkey, J.; Edwards, D.J.; Kenyon, J.J.; Phu Huong Lan, N.; Campbell, J.I.; Thwaites, G.; Thi Khanh Nhu, N.; Hall, R.M.; Fournier-Level, A.; Baker, S.; Holt, K.E. Repeated Local Emergence of Carbapenem-Resistant Acinetobacter baumannii in a Single Hospital Ward. Microb Genom 2016, 2. [Google Scholar] [CrossRef]
- Abhari, S.S.; Badmasti, F.; Modiri, L.; Aslani, M.M.; Asmar, M. Circulation of Imipenem-Resistant Acinetobacter baumannii ST10, ST2 and ST3 in a University Teaching Hospital from Tehran, Iran. J Med Microbiol 2019, 68, 860–865. [Google Scholar] [CrossRef] [PubMed]
- Meumann, E.M.; Anstey, N.M.; Currie, B.J.; Piera, K.A.; Kenyon, J.J.; Hall, R.M.; Davis, J.S.; Sarovich, D.S. Genomic Epidemiology of Severe Community-Onset Acinetobacter baumannii Infection. Microb Genom 2019, 5. [Google Scholar] [CrossRef] [PubMed]
- Valcek, A.; Nesporova, K.; Whiteway, C.; De Pooter, T.; Coster, W. De; Strazisar, M.; der Henst, C. Van. Genomic Analysis of a Isolate Collection Containing Multidrug-, Extensively Drug-, Pandrug-, and Carbapenem-Resistant Modern Clinical Isolates of Acinetobacter baumannii. Antimicrob Agents Chemother 2022, 66, e00892–22. [Google Scholar] [CrossRef] [PubMed]
- Savin, M.; Sib, E.; Heinemann, C.; Eichel, V.M.; Nurjadi, D.; Klose, M.; Andre Hammerl, J.; Binsker, U.; Mutters, N.T. Tracing Clinically-Relevant Antimicrobial Resistances in Acinetobacter baumannii -calcoaceticus Complex across Diverse Environments: A Study Spanning Clinical, Livestock, and Wastewater Treatment Settings. Environ Int 2024, 186, 108603. [Google Scholar] [CrossRef]
- Park, S.C.; Lee, K.; Kim, Y.O.; Won, S.; Chun, J. Large-Scale Genomics Reveals the Genetic Characteristics of Seven Species and Importance of Phylogenetic Distance for Estimating Pan-Genome Size. Front Microbiol 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Gheorghe-Barbu, I.; Surleac, M.; Barbu, I.C.; Paraschiv, S.; Bănică, L.M.; Rotaru, L.I.; Vrâncianu, C.O.; Niță Lazăr, M.; Oțelea, D.; Chifiriuc, M.C. Decoding the Resistome, Virulome and Mobilome of Clinical versus Aquatic Acinetobacter baumannii in Southern Romania. Heliyon 2024, 10. [Google Scholar] [CrossRef]
- Sehnal, L.; Brammer-Robbins, E.; Wormington, A.M.; Blaha, L.; Bisesi, J.; Larkin, I.; Martyniuk, C.J.; Simonin, M.; Adamovsky, O. Microbiome Composition and Function in Aquatic Vertebrates: Small Organisms Making Big Impacts on Aquatic Animal Health. Front Microbiol 2021, 12. [Google Scholar] [CrossRef]
- Shayo, G.M.; Elimbinzi, E.; Shao, G.N.; Fabian, C. Severity of Waterborne Diseases in Developing Countries and the Effectiveness of Ceramic Filters for Improving Water Quality. Bull Natl Res Cent 2023, 47, 113. [Google Scholar] [CrossRef]
Figure 1.
The microbial load of Acinetobacter for the upstream and downstream sampling points of investigated WWTPs in the two locations in southern Romania.
Figure 1.
The microbial load of Acinetobacter for the upstream and downstream sampling points of investigated WWTPs in the two locations in southern Romania.
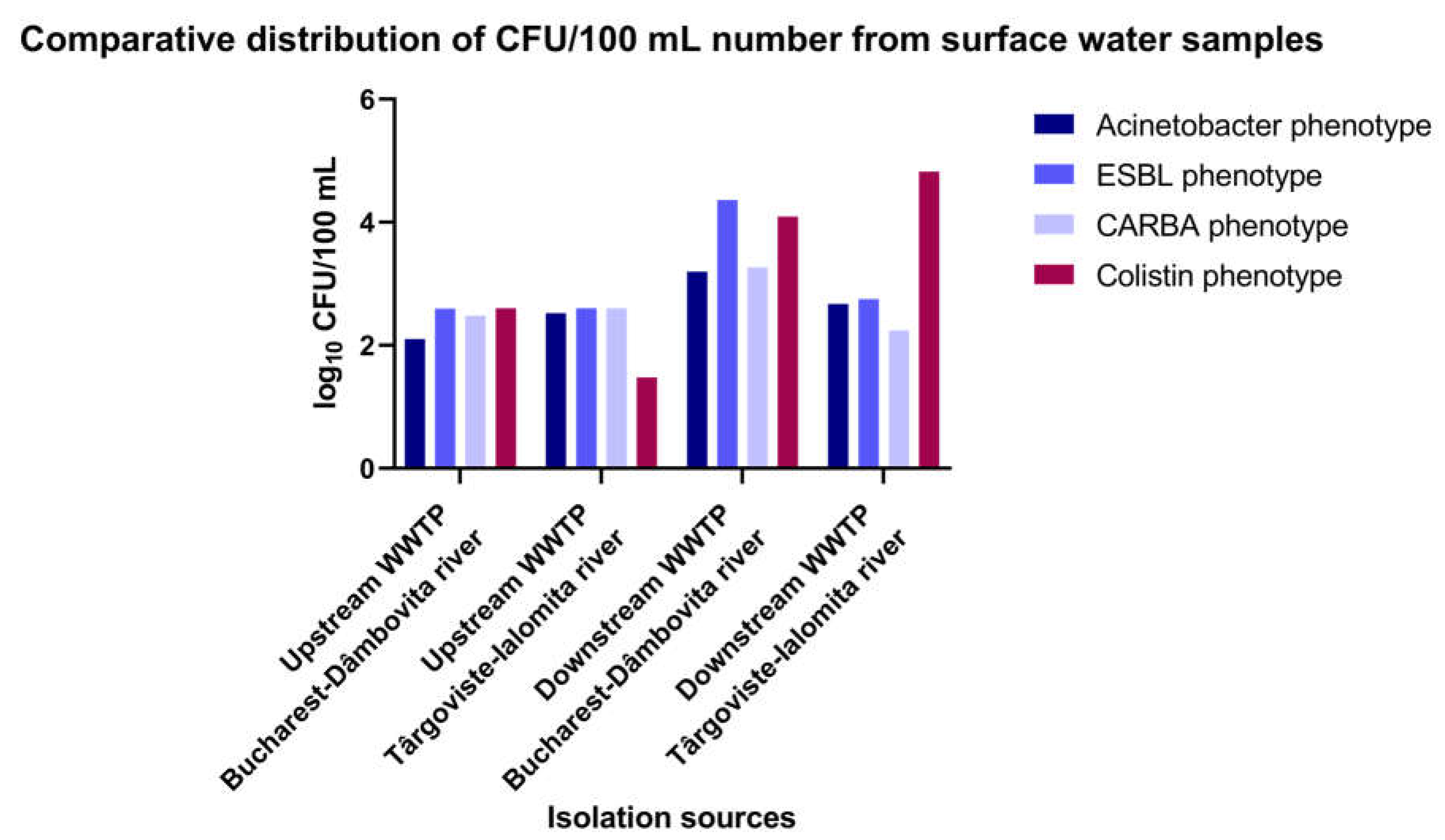
Figure 2.
The microbial load with Acinetobacter for the wastewater sample collection inside the investigated WWTPs in the two locations in southern Romania.
Figure 2.
The microbial load with Acinetobacter for the wastewater sample collection inside the investigated WWTPs in the two locations in southern Romania.
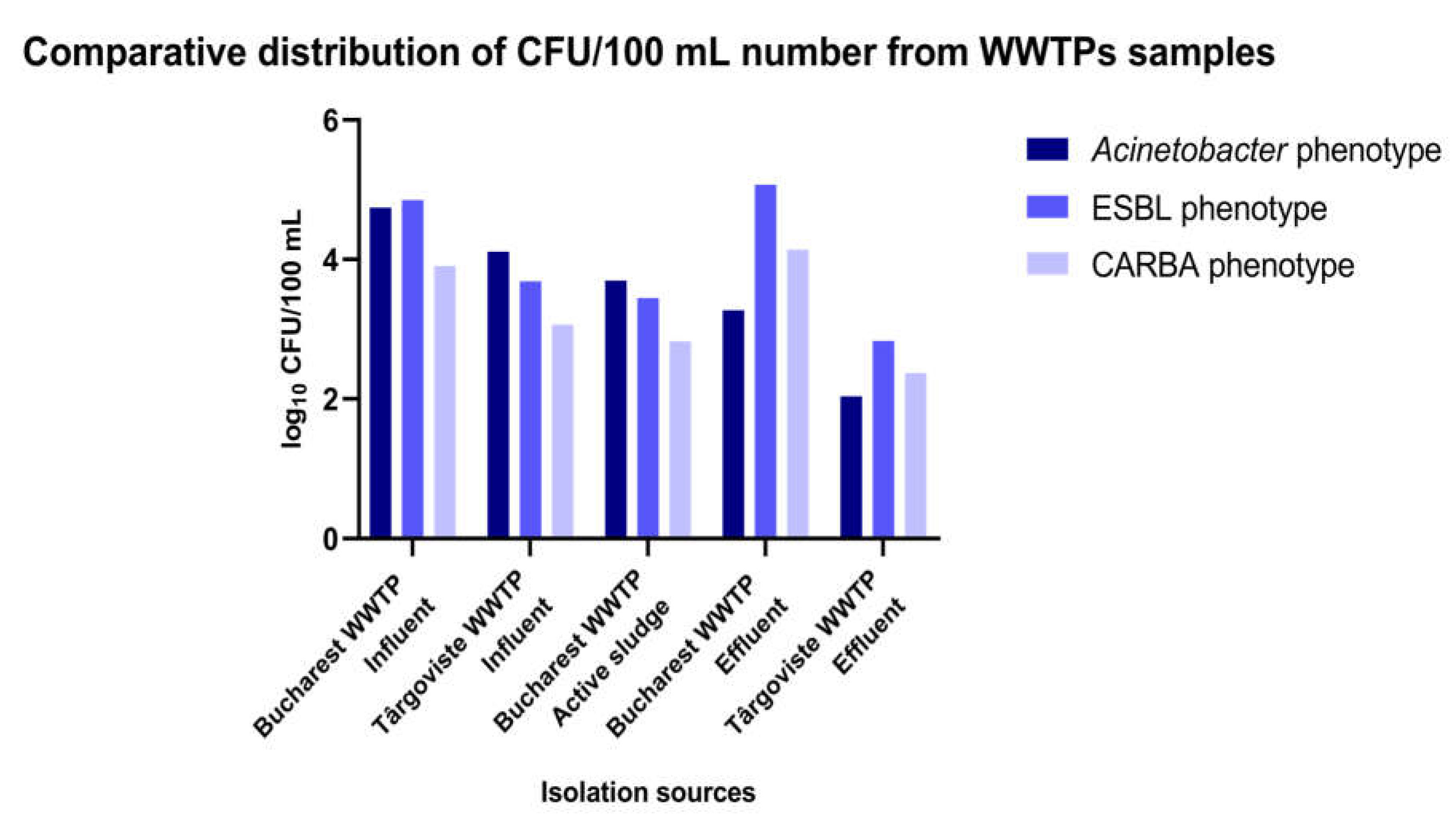
Figure 3.
Percentage of A. baumannii isolates isolated from IHIs, WWTP, and surface water samples in Bucharest, categorized according to their resistance profile to different antibiotic classes.
Figure 3.
Percentage of A. baumannii isolates isolated from IHIs, WWTP, and surface water samples in Bucharest, categorized according to their resistance profile to different antibiotic classes.
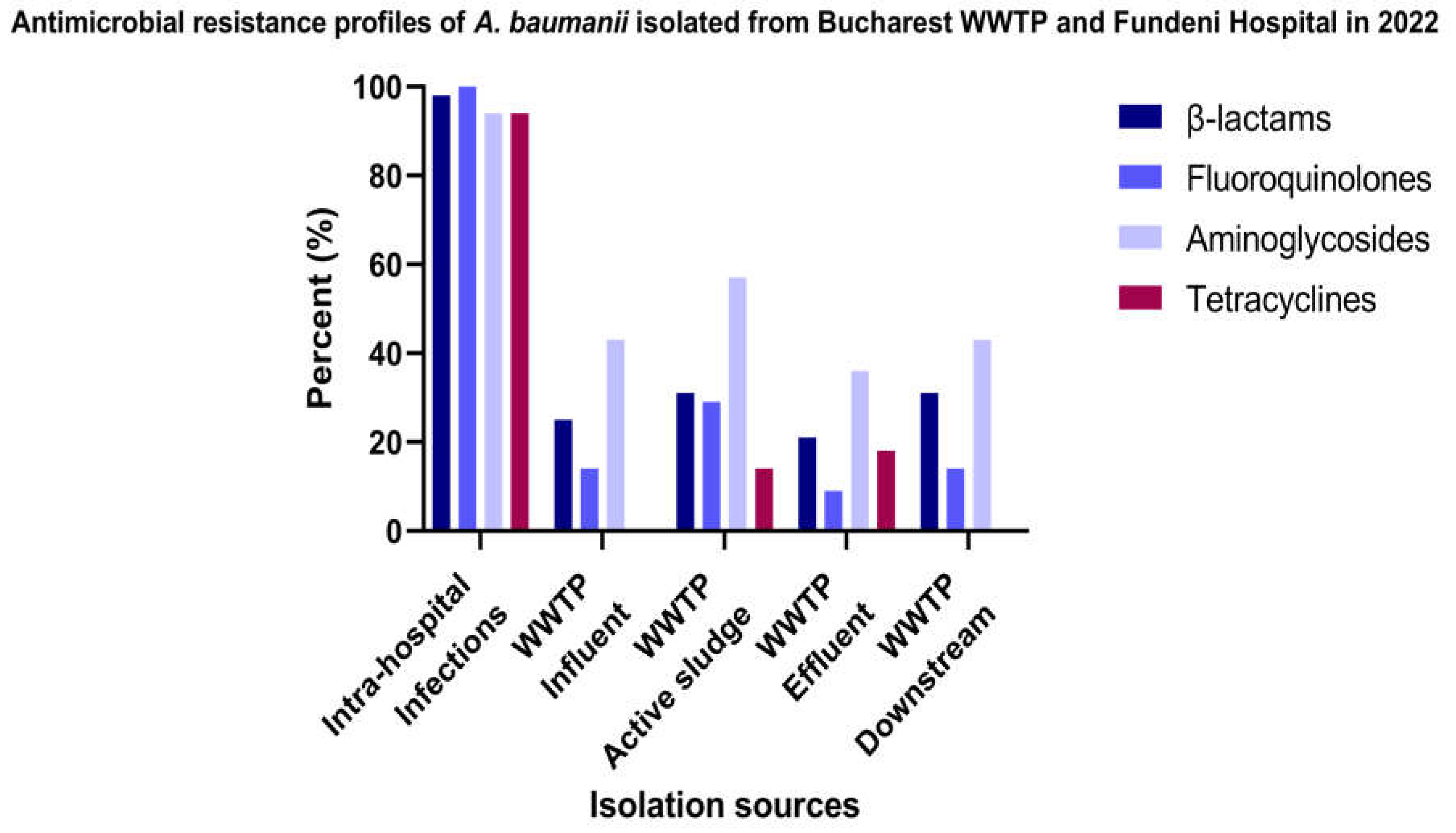
Figure 4.
Percentage of A. baumannii isolates recovered from WWTP and surface water samples in Targoviste, categorized according to their resistance profile to different antibiotic classes.
Figure 4.
Percentage of A. baumannii isolates recovered from WWTP and surface water samples in Targoviste, categorized according to their resistance profile to different antibiotic classes.
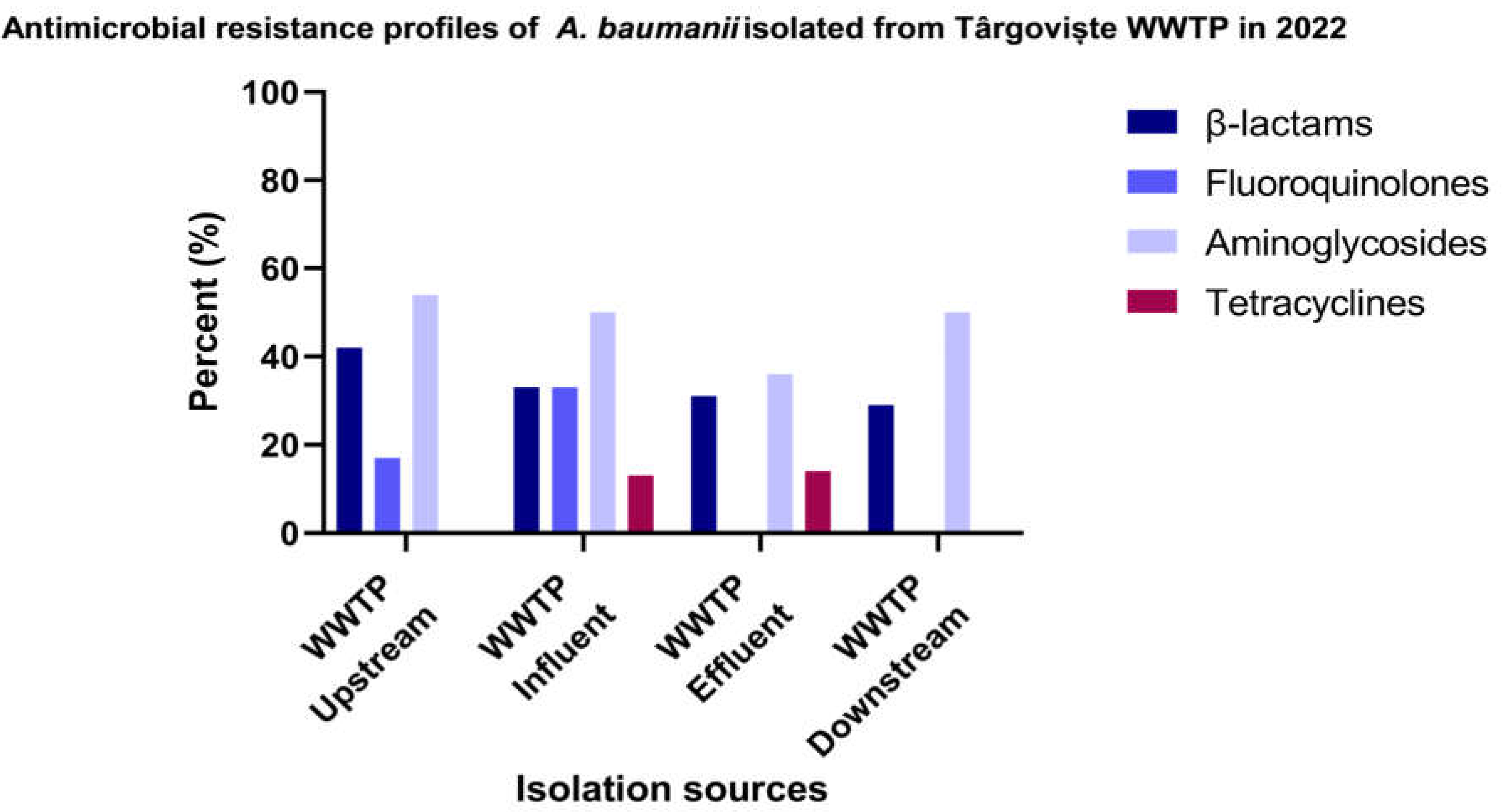
Figure 6.
β-lactamase producing A. baumannii isolates isolated from Targoviste, Romania in 2022.
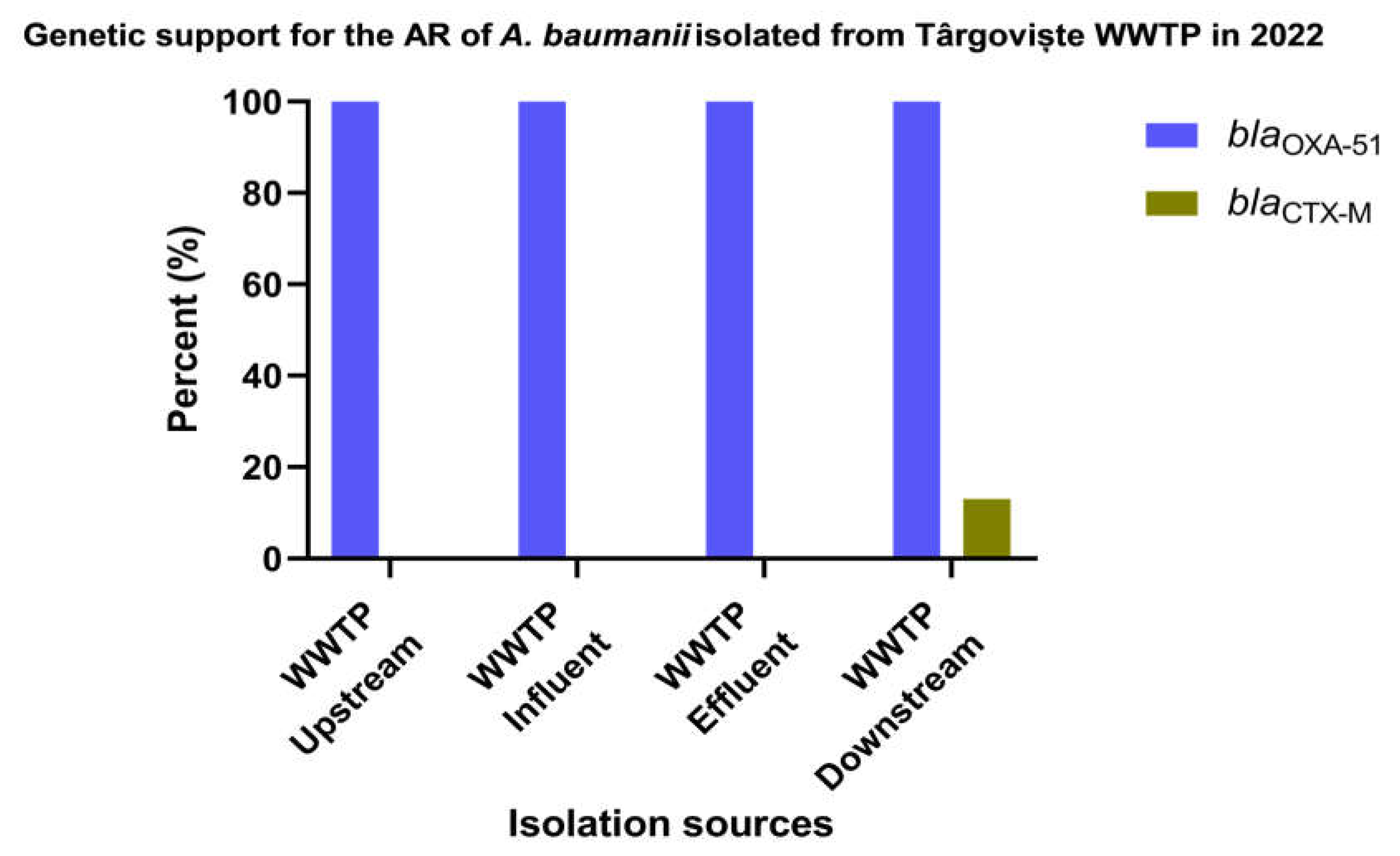
Figure 7.
Pangenome analysis of A. baumannii isolates from WW, SW and IHI samples in Southern Romania, based on accesory genes.
Figure 7.
Pangenome analysis of A. baumannii isolates from WW, SW and IHI samples in Southern Romania, based on accesory genes.
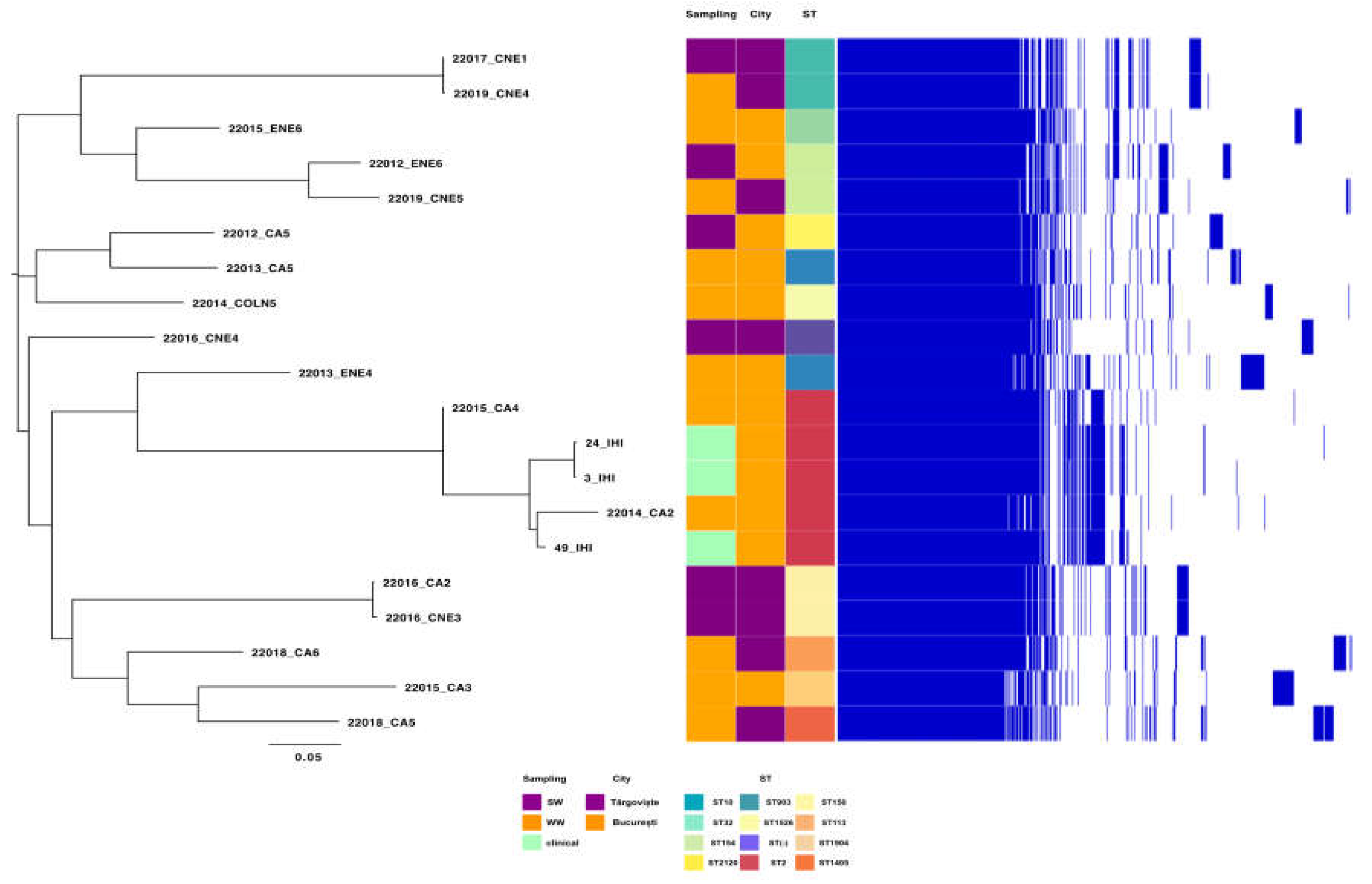
Figure 8.
A. baumannii isolates’ pangenome - FriPan MDS & pangenome tree representation based on accessory genes.
Figure 8.
A. baumannii isolates’ pangenome - FriPan MDS & pangenome tree representation based on accessory genes.
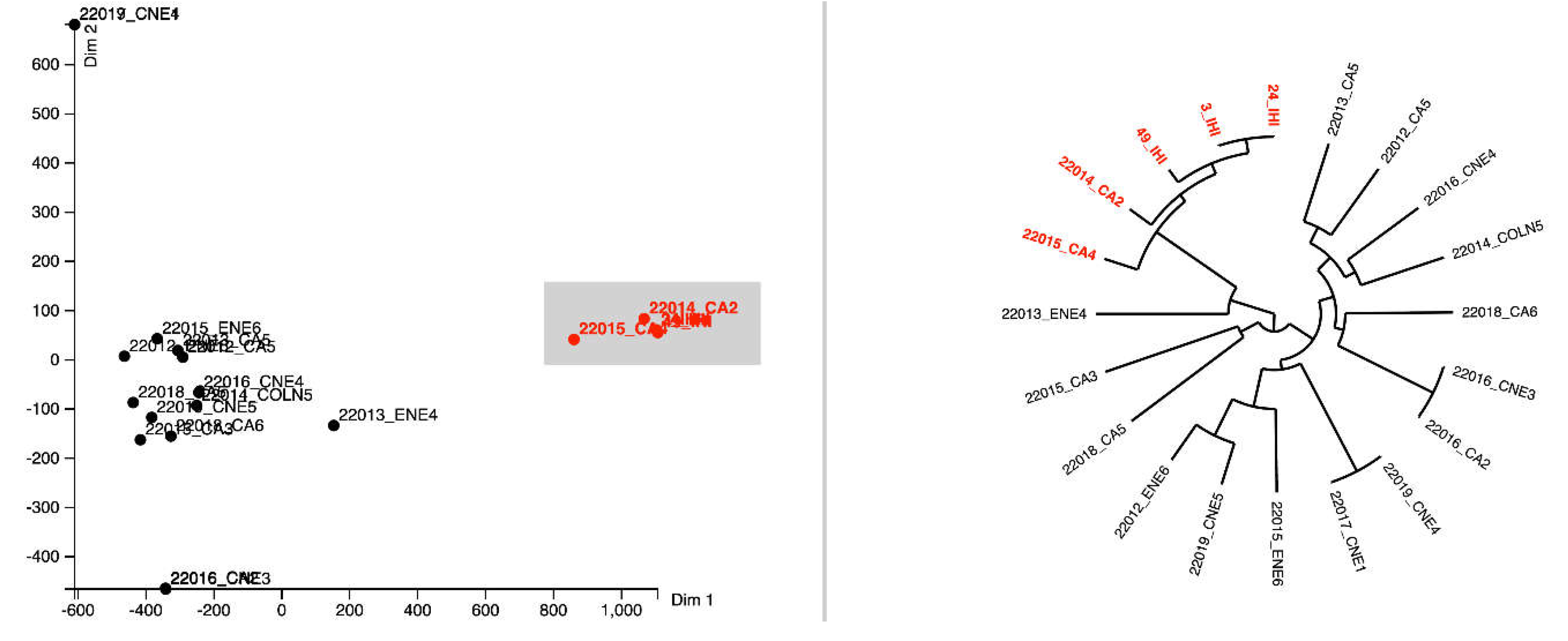
Figure 9.
Krona plots illustrating microbial community composition based on 16S rRNA sequencing of fish intestine samples from Glina, Bucharest and Targoviste. (A) Krona plot representing microbial taxa present in fish intestines from Glina, Bucharest. (B) Krona plot representing microbial taxa present in fish intestines from Targoviste. Each segment in the plots represents a taxonomic group at different levels (phylum, class, order, etc.), with the size of the segments corresponding to the relative abundance of that taxon within the sample. Taxonomic labels are color-coded for clarity.
Figure 9.
Krona plots illustrating microbial community composition based on 16S rRNA sequencing of fish intestine samples from Glina, Bucharest and Targoviste. (A) Krona plot representing microbial taxa present in fish intestines from Glina, Bucharest. (B) Krona plot representing microbial taxa present in fish intestines from Targoviste. Each segment in the plots represents a taxonomic group at different levels (phylum, class, order, etc.), with the size of the segments corresponding to the relative abundance of that taxon within the sample. Taxonomic labels are color-coded for clarity.
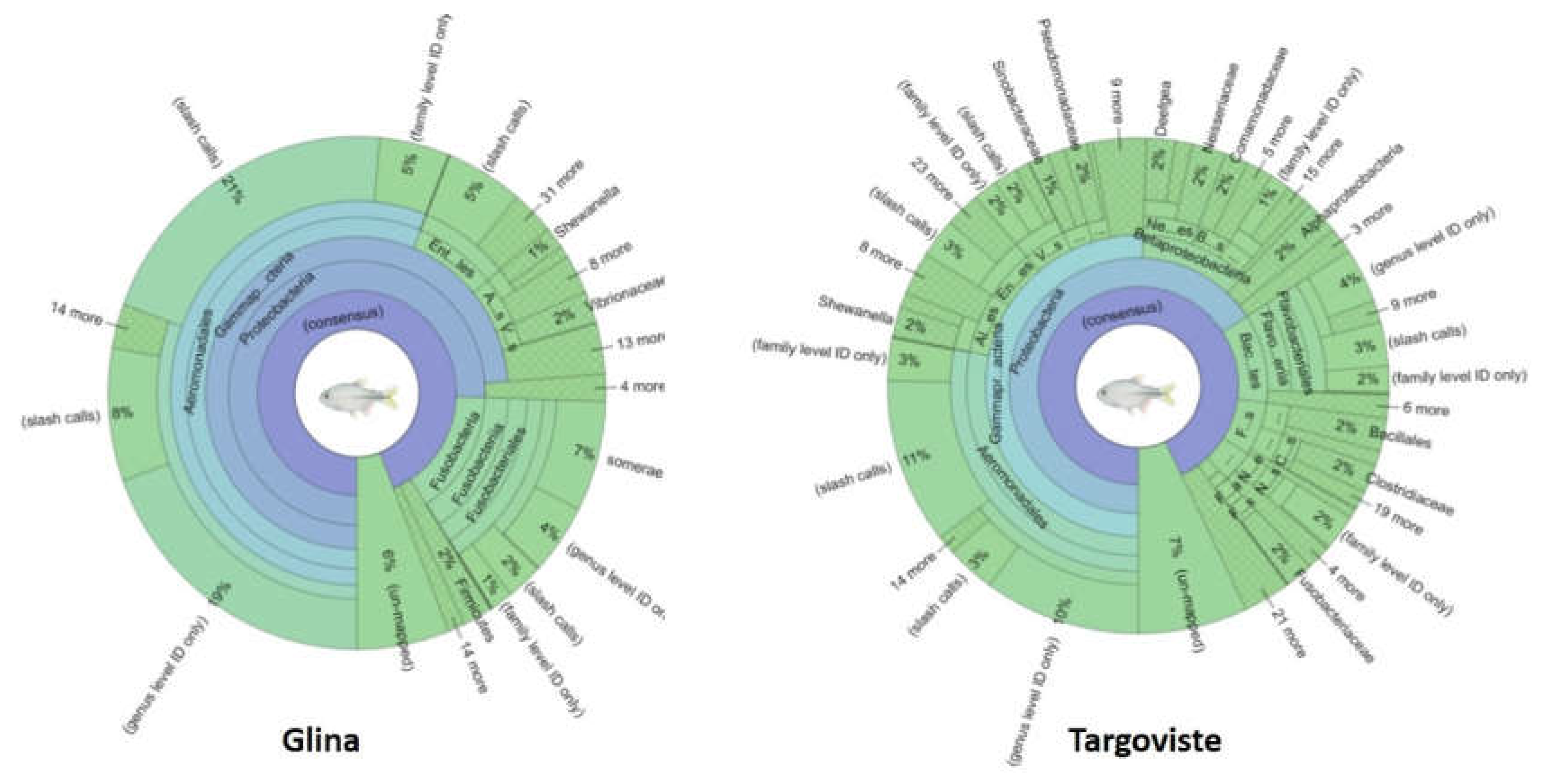
Figure 10.
Shannon Diversity Index of microbial communities in fish intestine samples from Glina and Targoviste.
Figure 10.
Shannon Diversity Index of microbial communities in fish intestine samples from Glina and Targoviste.
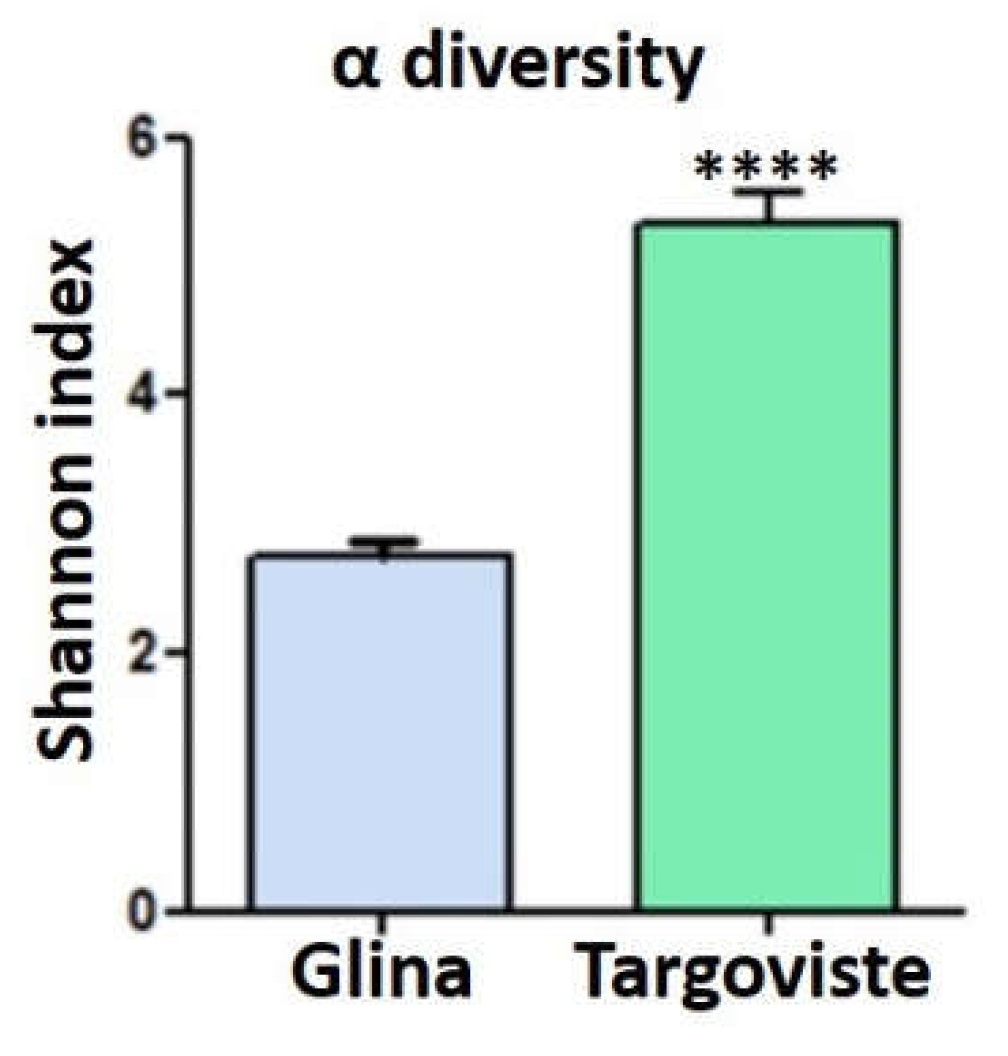
Figure 11.
Krona plots illustrating microbial community composition based on 16S rRNA sequencing of upstream and downstream water samples in Targoviste (A) and Glina (B).
Figure 11.
Krona plots illustrating microbial community composition based on 16S rRNA sequencing of upstream and downstream water samples in Targoviste (A) and Glina (B).
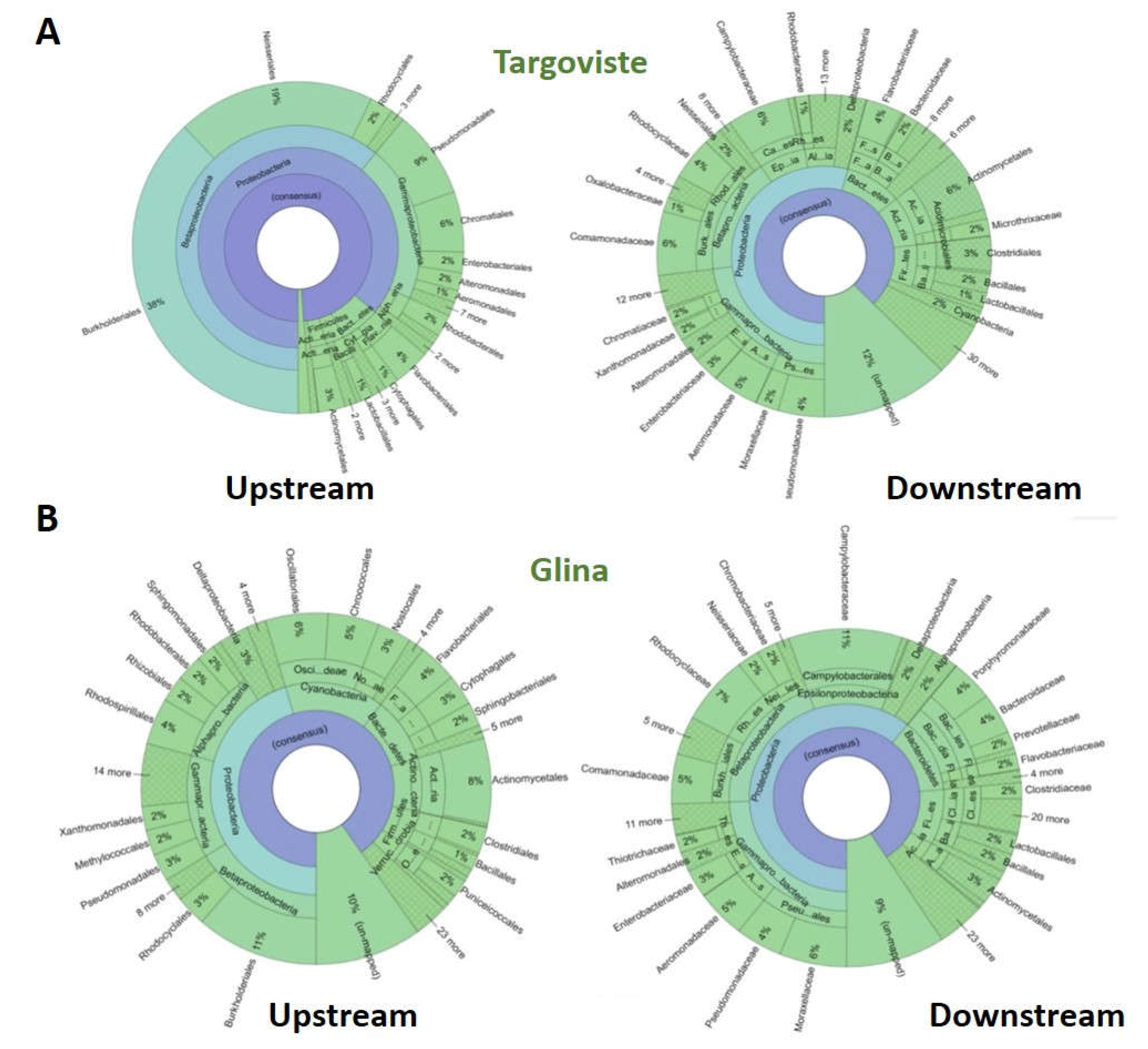
Figure 12.
Taxonomic composition of Moraxellaceae in surface water samples from DO regions of Glina, Bucharest and Targoviste WWTPs based on 16S rRNA metagenomic analysis.The inner circle represents the taxonomic classification at the family level, while the outer circle provides a more detailed view at the genus level. Glina, Bucharest: Acinetobacter (43%), unidentified genera within Moraxellaceae (28%), Moraxella (7%), Psychrobacter (3%), and Enhydrobacter (0.9%). Targoviste: Acinetobacter (52%), unidentified genera within Moraxellaceae (37%), Moraxella (7%), and Enhydrobacter (2%).
Figure 12.
Taxonomic composition of Moraxellaceae in surface water samples from DO regions of Glina, Bucharest and Targoviste WWTPs based on 16S rRNA metagenomic analysis.The inner circle represents the taxonomic classification at the family level, while the outer circle provides a more detailed view at the genus level. Glina, Bucharest: Acinetobacter (43%), unidentified genera within Moraxellaceae (28%), Moraxella (7%), Psychrobacter (3%), and Enhydrobacter (0.9%). Targoviste: Acinetobacter (52%), unidentified genera within Moraxellaceae (37%), Moraxella (7%), and Enhydrobacter (2%).
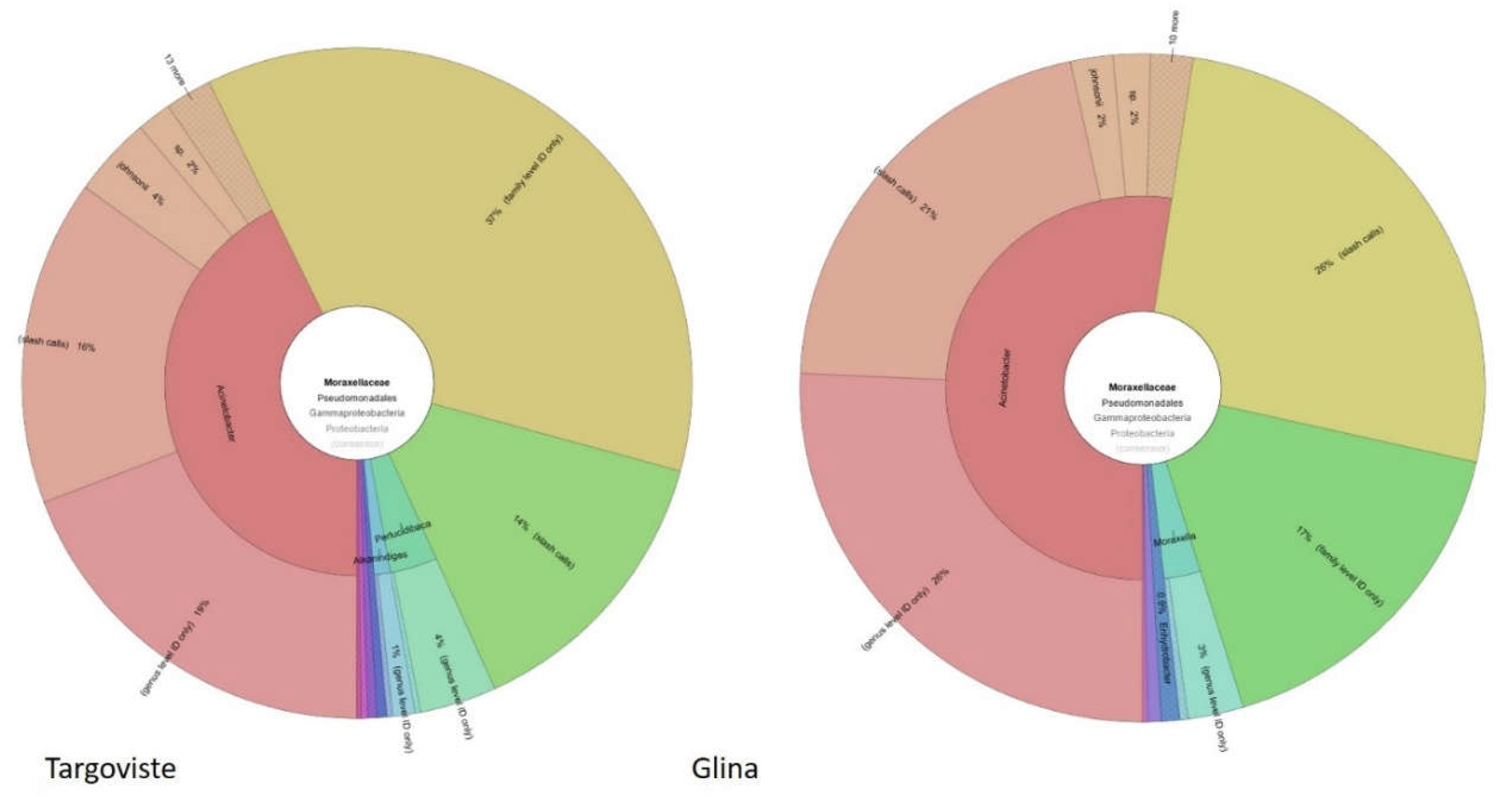

Table 1.
MIC values for colistin susceptibility in clinical and wastewater A. baumannii isolates from Southern Romania.
Table 1.
MIC values for colistin susceptibility in clinical and wastewater A. baumannii isolates from Southern Romania.
| ANTIBIOTIC/ ISOLATE | 22012-CA5 | 22012-ENE6 | 22013-CA5 | 22013-ENE4 | 22014-CA2 | 22014-COLN5 | 22015-CA3 | 22015-CA4 | 22015-ENE6 | 22016-CA2 | 22016-CNE3 | 22016-CNE4 | 22017-CNE1 | 22018-CA5 | 22018-CA6 | 22019-CNE4 | 22019-CNE5 | 24-IHI | 3-IHI | 49-IHI |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| COLISTIN (µG/ML) | 0.25 | <0.25 | <0.25 | <0.25 | <0.25 | <0.25 | <0.25 | <0.25 | <0.25 | <0.25 | <0.25 | <0.25 | <0.25 | <0.25 | <0.25 | <0.25 | <0.25 | 1 | <0.25 | <0.25 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



