
Submitted:
29 July 2024
Posted:
30 July 2024
You are already at the latest version
Abstract
Epigenetic changes regulate gene expression through histone modifications, chromatin remod-eling and proteins interpreter of these modifications. The PRC1 and PRC2 complexes shape gene repression via histone modifications. Specifically, the CBX protein family aids PRC1 recruitment to chromatin, impacting the progressive multistep process driving chromatin silencing. Among family members, CBX3 is a complex protein involved in aberrant epigenetic mechanisms that drive lung cancer progression. CBX3 promotes lung tumorigenesis by interacting with key pathways such as PI3K/AKT, Ras/KRAS, Wnt/β-catenin, MAPK, Notch, and p53, leading to in-creased proliferation, inhibition of apoptosis, and enhanced resistance to therapy. Given our current lack of knowledge, additional research is required to uncover the intricate mechanisms underlying CBX3 activity, as well as its involvement in molecular pathways and its potential biomarker evaluation. Specifically, the dissimilar roles of CBX3 could be reexamined in order to gain a greater insight into lung cancer pathogenesis. This review aims to provide a clear overview of the context-related molecular profile of CBX3, which could be useful for addressing clinical challenges and developing novel targeted therapies based on personalized medicine.
Keywords:
CBX3/ HP1γ
; lung cancer
; epigenetics
; chromatin and signaling pathways
1. Introduction
Modifications associated with chromatin structure and function, known as epigenetic changes, are responsible for the activation and repression of genes, thereby impacting the synthesis and production of specific proteins within cells and with variable expression patterns [1,2,3]. Gene transcription can be reversibly modulated via chromatin remodeling, which regulates the accessibility of promoter and enhancer regions to regulatory proteins. The remodeling process is controlled by more than 300 proteins and enzymes, which recognize, add, or remove chemical moieties from histone proteins. Such modifications, including acetylation, methylation, ubiquitination, and phosphorylation, occur at various lysine, arginine, and serine/threonine residues. The cumulative effect of these post-translational modifications results in the formation of a “histone code [4,5,6,7]. Importantly, the biological significance of these histone modifications is deciphered by a diverse group of histone code readers. Reader proteins with specialized domains identify and interpret histone and DNA chemical modifications. These specialized enzymes include histone and DNA readers. Histone readers containing chromodomains, bromodomains, and plant homeodomain zinc fingers recognize and interpret histone modifications, while DNA readers that possess methyl-CpG-binding domains recognize specific alterations in DNA [8,9,10]. Additionally, RNA-binding proteins recognize and interpret modifications in RNA, such as 5-methylcytosine and N6-methyladenosine which affect gene expression [8,11]. Various enzymes, known as epigenetic writers and erasers, regulates the dynamic of epigenetic modifications. Specifically, the writers introduce chemical modifications to histones and DNA [1]. These include histone and DNA methyltransferases, histone acetyltransferases, ubiquitin ligases, and histone kinases. While erasers remove epigenetic modifications including histone deacetylases and demethylases, phosphatases, deubiquitinating enzymes, and ten-eleven translocations [9]. These activities collectively regulate gene expression and cellular homeostasis [12,13]. The epigenetic regulatory complex known as Polycomb repressive complex (PRC) 1, is a chromatin-modifying complex that is responsible for keeping the chromatin in a repressed state by mono-ubiquitinating histone H2A, thus restricting target gene transcription [14,15]. PRC1 is completed via its association with the Chromobox (CBX) family members, which are responsible for its recruitment to chromatin whereas the RING1a/b subunit represent the catalytic subunits of the complex [16]. Likewise, the PRC1, the PRC2 is a multi-subunit protein complex that cooperate with PRC1 playing a vital role in the epigenetic regulation of gene expression. The principal elements of PRC2 include the catalytic subunit enhancer of zeste homolog 1/2 (EZH1/2), which is able to methylate histones, as well as embryonic ectoderm development, suppressor of zeste 12 protein homolog and retinoblastoma-binding protein (RBBP) 4/7 [17,18,19]. In various cancers, the function of PRC2 is disrupted as a result of mutations or changes in the expression of its components. This phenomenon contributes to tumor development by activating tumor growth-suppressing genes or by triggering pathways that promote the growth of cancer cells [20,21,22]. PRC2 has therefore become a highly promising target for therapeutic intervention. The CBX family is a large group of proteins, comprising eight members, crucial in transcriptional repression and memory [23] due to the presence of a single N-terminal chromodomain [24]. The CBX family can be classified into two distinct groups. The first group, the Polycomb group (PcG), consists of CBX2, CBX4, CBX6, CBX7, and CBX8, all of which contain a C-terminal Polycomb repressor box and a conserved N-terminal chromodomain. The second group, the Heterochromatin Protein 1 (HP1) group, comprises CBX1, CBX3, and CBX5, characterized by an N-terminal chromodomain and a Chromoshadow domain associated with HP1. CBX family proteins facilitate the recruitment of PRC1 to chromatin, thus playing a crucial role in the initiation, growth, and development of tumors by suppressing the differentiation of cancer stem cells and promoting their self-renewal [24,25]. In non-small cell lung cancer (NSCLC), CBX3 has emerged as a promising prognostic biomarker. Higher CBX3 expression levels have been linked to the control of cell cycle progression and its potential impact on the PI3K/ AKT and Ras signaling pathways [26,27]. However, the precise mechanisms by which CBX3 contributes to the development and progression of lung cancer, including its involvement in different signaling pathways, have not been thoroughly examined and require additional research. Currently, the literature does not provide sufficient information to fully understand the involvement of CBX3 in these diseases, underlining the need to develop more elaborate and precise studies to obtain a greater insight into the function of CBX3 in molecular pathways. This review aims to give a detailed overview of the therapeutic potential of CBX3. We describe the various pathways involving CBX3, highlighting its differential mechanisms of action as well as its significance as a potential therapeutic biomarker in lung cancer.
2. CBX3/HP1γ Protein in Cancer Proliferation
The HP1 family in mammals consists of three distinct yet remarkably conserved non-histone homologs, namely CBX1/HP1β, CBX3/HP1γ, and CBX5/HP1α [28,29]. The chromodomain proteins of HP1 and Polycomb group (PcG) exhibit a significant degree of amino acid sequence similarity, with over 60% identity [30]. The criticality of the CHD of HP1 lies in its association with chromatin, which is facilitated by the specific interaction between the CHD and histone H3 lysine K9 di/trimethylation (H3K9me2/3). The strength of the binding affinity between the CHD and H3K9me2/3 was found to be directly proportional to the higher levels of H3K9me2/3 [31,32]. The carboxyl-terminal region of the HP1 protein family contains a second conserved domain known as the chromo shadow domain (CSD) (Figure 1) [33]. Although the general architecture of the CSD resembles that of the CHD, these domains exhibit distinct functionalities. The CSD functions primarily as a dimeric domain, therefore HP1 proteins readily form homodimers and heterodimers via their CSDs [32,34,35]. As regards CBX3, its principal function is the establishment of heterochromatin, which represents the condensed state of chromatin. Within the chromatin structure, the “co-packed state” corresponding to heterochromatin is associated with gene transcriptional inactivation and/or gene silencing. Transcriptional inactivation is mediated by the binding of the CBX3 protein to regions of DNA that have undergone methylation at histone H3 lysine K9 (H3K9) via a positive feedback loop [36]. CBX3 is able to recognize and bind the H3K9me2 and H3K9me3 marks. Subsequently, these modifications facilitate the recruitment of the H3K9 methyltransferase known as histone-lysine N-methyltransferase SUV39H1 to methylate neighboring H3K9 residues [37]. The diffusion of H3K9me3 marks is concomitant with the recruitment of multiple proteins, which elicit chromatin compaction and transcriptional repression by sequestering genes, rendering them transcriptionally inactive [38,39,40].
CBX3, rendered as a soluble nuclear protein and HP1 family member, is encoded by the CBX3 gene and is localized on chromosome 7p15.2 [41]. In addition, at subcellular level, it is localized to the nucleoplasm and nuclear bodies. CBX3 links methylation marks to RNA splicing, DNA repair, and transcriptional silencing resulting involved in various cellular processes, such as gene regulation, DNA repair, and telomere function [42,43]. Importantly CBX3 is regarded as a multifaceted crystal-structured protein in humans that also has a function in transcriptional inhibition and activation, cell growth and differentiation, and epigenetic modifications [44,45]. The CBX3 chromodomain recognizes and binds with non-histone and histone methylated peptides and, based on comparable affinities, also binds with H1K26, H3K9, and G9aK185 methylated peptides [36]. The binding of the CBX3 chromodomain to methylated histones occurs via a conserved mechanism, which is enhanced by the chromodomain ARKS/T motif, allowing the chromodomain to recognize and specifically bind to methylated histones [43,46]. CBX3 also interacts with non-histone proteins, including PIM1, CBX5, CBX1, Ki-67, and Lamin B receptor, controlling gene expression [43]. CBX3 interacts directly with active genes, particularly within gene bodies, and facilitates the process of transcriptional elongation and RNA processing. It is also involved in recruiting splicing factors to enable efficient co-transcriptional splicing [47]. CBX3 interacts with the E2F1 transcription factor, a key player in regulating the cell cycle. Several studies report that cellular proliferation is enhanced by the increase in E2F1 transcriptional activity mediated by CBX3. E2F1 selectively directs its binding toward genes that encode proteins responsible for the regulation of cell cycle progression during transition from G1 to S phase, including cyclin D1 and CDK4 [48,49]. CBX3 is able to interact with the tumor suppressor protein p53, impeding its transcriptional activity and resulting in apoptosis reduction and a concomitant increase in cell survival. The primary role of p53 under cellular stress, such as oncogenic activation and DNA damage, is to induce apoptosis regulated by pro-apoptotic genes such as BCL2 Associated X, Apoptosis Regulator (BAX) and phorbol-12-myristate-13-acetate-induced protein 1 (PMAIP1, also known as NOXA), as well as transcriptional activation of p53 upregulated modulator of apoptosis (PUMA) [50]. CBX3 was found able to indirectly enhance transcriptional activation of genes involved in DNA repair, including RAD51 and breast cancer gene 1 (BRCA1). RAD51 plays a critical role in repairing DNA double-strand breaks (DSBs) via homologous recombination (HR) [51,52]. A recent study described an association between mutations in the BRCA1 protein, which has a role in HR repair, and a higher susceptibility to breast and ovarian cancers [53]. The interface between CBX3 and E2F1 enhances the expression of RAD51 and BRCA1, resulting in increased HR repair and resistance to chemotherapy. CBX3 also interacts with other DNA repair proteins, such as PARP1 and Ku70, which participate in the repair of DSBs through non-homologous end joining [54]. Through its interactions, CBX3 plays a crucial role in multiple DNA repair pathways either directly or indirectly by recruiting DNA repair proteins, thereby maintaining genomic stability [55,56,57].
3. CBX3 as a Multiplayer in Lung Cancer Progression
CBX3 has been found dysregulated showing an abnormal expression profile in various cancers. Expression levels of this gene are increased in several cancer types including gastric, prostate, breast, colorectal, and lung cancers. CBX3 expression is also dysregulated in osteosarcoma and hepatocellular carcinoma [44,54,58,59]. Conversely, expression levels of CBX3 are lower in colorectal cancer low-grade adenomas, hyperplastic and mucosal polyps [54]. Among the three HP1 proteins, CBX3 is the histone reader protein that is highly expressed in lung adenocarcinoma (LUAD). The expression level of messenger RNA (mRNA) encoding CBX3 exhibits a positive correlation with size of tumors, occurrence of lymph node metastasis, and unfavorable prognosis in LUAD patients. Interestingly, the in vivo inhibition of CBX3 results in a reduction of tumor size and an extension of the survival period in mice with KRASG12D-induced LUAD [60]. Increased expression of CBX3 is correlated with an unfavorable prognosis in NSCLC and LUAD [61,62] through mechanisms involving the promotion of tumor proliferation via regulatory pathways of signal transduction affecting the cell cycle, notably G1/S phase transition and the p53 pathway [27]. The prognostic value of CBX3 is further supported by its association with tumor diameter and lymph node metastasis, suggesting its involvement in tumor growth and metastasis [27]. The therapeutic potential of targeting CBX3 in lung cancer is underscored by its overexpression in NSCLC and its association with epigenetic modifications and cell differentiation [44]. The oncogenic role of CBX3 is also highlighted by the observation that CBX3 and H3K9me3 levels are increased in NSCLC tumor-initiating cells, where they inhibit DNA damage responses to antineoplastic agents [61]. The expression of CBX3 is markedly increased in LUAD tissues of smokers compared to non-smokers and it is also associated with unfavorable prognosis and advanced disease stage [63]. Interestingly, a study show that cigarette smoke causes an increase in CBX3 expression by promoting binding of the transcription factor YBX1 to the CBX3 promoter [44]. High CBX3 protein levels also enhance the growth, invasion, and spread of LUAD cells by controlling the cell cycle progression and activating Rho GTPases [64]. Notably, elevated expression of CBX3 in lung cancers linked to smoking is often caused by genetic changes, such as an increase in the number of copies of the gene, as well as in epigenetic dysregulation [65]. In smokers, it has also been observed that CBX3 interacts with tripartite motif-containing (TRIM) 28, TRIM24, and RBBP4 to create a repressor complex. This complex binds to the Rho GTPase-activating protein 24 (ARHGAP24) promoter and inhibits its transcription. Reducing levels of ARHGAP24 results in the overexpression of active Ras-related C3 botulinum toxin substrate 1 (RAC1) that in turn, triggers signaling pathways (see also Subsection 4.6) promoting the advancement of LUAD [80]. Outside the CBX3/ARHGAP24/RAC1 axis, CBX3 can also facilitate smoking-induced LUAD by inhibiting the tumor suppressor FBP1 and controlling glycolysis [63]. Intriguingly, CBX3 can also enhances the development of lung tumors by suppressing the transcriptional activity of nuclear receptor co-repressor 2 (NCOR2) and zinc finger and BTB domain-containing 7A (ZBTB7A). These transcriptional regulators have an impact on cell proliferation and migration [66]. Further, the expression of CBX3 triggers the development of stem cell-like characteristics in lung tumors, enhancing the presence of markers associated with cancer stem cells and targets of the oncogenic transcription factor c-Myc [67]. Mechanistically, CBX3 plays a crucial role in suppressing target genes through chromatin remodeling, leading to abnormal cell development and the inhibition of differentiation pathways [44,67]. Due to its cancer-causing properties, CBX3 shows potential as a reliable predictive biomarker and a possible target for treatment in NSCLC associated with smoking. Manipulating the expression or activity of CBX3 could potentially limit the development of lung tumors and improve patient prognosis. Studies suggest that exposure to cigarette smoke can lead to specific alterations in the histone organization of lung cells, and that these changes can affect how transcription factors bind to promoters of genes, including CBX3 [63,68]. Although there is no direct evidence to support this hypothesis, it is suggested that the transcription factor NF-κB, known to be activated by cigarette smoke, can potentially regulate the expression of target genes such as CBX3 [69,70].
The different interactors of CBX3, their molecular mechanisms, and the different pathways involved in lung cancer are listed in Table 1.
4. Involvement of CBX3 in Pathways Leading to Lung Cancer
Molecular pathways involved in the regulation of cell cycle, differentiation, death, and signaling are known to be altered in processes of tumorigenesis. To date, major efforts have been made to discover new cancer driver genes and unravel the molecular mechanisms in which they are involved by combining scientific data, such as multi-omics data, and knowledge obtained from the literature. For example, therapies based on molecular targets have transformed anticancer treatment approaches through personalized and/or precision medicine startegies. Based on this premise, CBX3 has been found implicated in a broad spectrum of human cancers, including NSCLC [27,71]. Remarkably, CBX3 is involved in several signaling pathways, critical for cell survival, proliferation, and differentiation, and plays a dominant role in lung cancer. However, further research is needed to better elucidate these pathways and deeper explore the potential of CBX3 as a therapeutic target. The following subsections describe the role of CBX3 in key cancer-associated pathways aim to shedding light its mechanistic role in lung cancer progression. The involvement of CBX3 in crucial lung cancer networks is schematically illustrated in Figure 2.
4.1. Role of CBX3 in PI3K-AKT and (K)Ras Signaling Pathways
Recent studies describe a significant connection between CBX3 and the activation of the PI3K-AKT pathway. This pathway regulates essential cellular processes such as growth, survival, and metabolism [73]. The aberrant activation of the PI3K-AKT pathway is a well-known characteristic of tumor development. In this context, CBX3 was found to play a role in PI3K-AKT dysregulation by facilitating the phosphorylation and subsequent activation of AKT. CBX3 may act as an oncogenic driver and is potentially involved in the Ras signaling pathway, one of the most crucial molecular mechanisms inducing oncogenic transformation. Enrichment analysis seems to support this hypothesis, but further research is needed to establish the specific role of CBX3 in this pathway in lung cancer [76]. A functional relationship is known to exist between CBX3 and EGFR or RAC1 in different human cancers [71], potentially impacting these signaling pathways. CBX3 has a significant effect on the KRAS signaling pathway in lung cancer. Specifically, CBX3 preferentially interacts with EZH2 triggering the transcription inhibition of microRNAs (miRNAs) such as let-7b, miR-31, and miR-128b. This results in the upregulation of target genes, including KRAS and MYC, that stimulate the growth and survival of tumor cells [77]. CBX3 aslo promotes oncogenic KRAS signaling, activating downstream effector pathways such as MAPK/ERK and PI3K/AKT. This activation enhances cancer cell proliferation, invasion, and metastasis [78]. Of note, a strong association is known to exist between elevated CBX3 levels and unfavorable prognosis in individuals with LUAD harboring KRAS mutations [79]. These findings highlight the potential of CBX3 as both a prognostic biomarker and a therapeutic target. Its association with these critical oncogenic signaling pathways further supports the importance of gaining a better insight into its role in cancer progression [72].
4.2. Role of CBX3 in Notch Signaling Pathway
In the context of lung cancer, the involvement of CBX3 in the Notch signaling pathway is intricate and diverse, underscoring the complex interplay between chromatin organization and signaling pathways in cancer progression [80]. The Notch signaling pathway is a crucial cell communication system in determining cell fate. CBX3 is able to interfere with the functioning of this pathway, and its effect may vary depending on the specific cellular environment and cancer type [81]. The Notch signaling pathway is highly conserved and involves the interaction of Notch receptors with their ligands, leading to cleavage of the Notch intracellular domain (NICD) and its translocation to the nucleus, where it influences gene expression [82,83]. However, the involvement of CBX3 in lung cancer, and particularly its relationship with the Notch signaling pathway, remains relatively understudied and seems to present a more complex scenario. CBX3 directly interacts with the NICD, recruiting the co-repressor complex, including histone deacetylases and DNA methyltransferases, to the promoter regions of Notch3 target genes, such as HES1 and HEY1[84]. This interaction leads to the epigenetic silencing of these genes through increased histone deacetylation and DNA methylation, ultimately resulting in the downregulation of the entire signaling pathway [84]. Thus, the chromatin remodeling may arise due to the ability of CBX3 to modify histones and change the chromatin state, consequently affecting the accessibility of Notch-responsive elements in the genome. In addition, since CBX3 has been linked to epigenetic changes such as H3K9me3, which plays a role in regulating the response to DNA damage, we speculate that it could potentially contribute to the resistance of tumor-initiating cells in NSCLC to antineoplastic drugs. This specific resistance is potentially conferred due to CBX3 binding to H3K9me3, resulting in the formation of transcriptionally repressive chromatin environments that can lead to the silencing of tumor suppressor genes in drug metabolism and efflux, contributing to drug resistance [61,85]. Interestingly, CBX3 expression has also been correlated to immune-related function regulation, also regulated by Notch signaling [86]. These interactions could additionaly modulate the tumor microenvironment, supporting tumor growth as well as resistance to various therapies. In the case of NSCLC, CBX3 might indirectly favor drug resistance by influencing immune evasion and immune cell infiltration mechanisms [72].
4.3. Role of CBX3 in Wnt Pathway
CBX3 regulates the Wnt/β-catenin signaling pathway, essential for cell proliferation, differentiation, and tumorigenesis in several types of cancer, including lung cancer [87]. The mode of action of CBX3 in lung cancer involves its function as a transcriptional regulator. CBX3 is able to bind gene promoters and influence gene expression. A recent study showed that CBX3 plays a crucial role in the transcriptional regulation of non-structural maintenance of chromosomes condensin I complex subunit G (NCAPG). This regulation, in turn, leads to activation of the Wnt/β-catenin signaling pathway. In colorectal cancer, activation of this pathway promotes cell proliferation and cell cycle progression, while inhibiting apoptosis. This mechanism likely operates in a similar manner in lung cancer, as the pathways involved in tumor formation are shared [88]. CBX3 overexpression has been linked to the advancement of lung adenocarcinoma via activation of the RAC1 pathway, a component of the Wnt signaling network [71,72]. Previous studies found a connection between CBX3 and cell cycle control [89]. Specifically, CBX3 was found to reduce the transition from G1 to S phase by influencing the activity of p21 [64,90]. This, in turn, may contribute to the growth of tumors. It is also reported that CBX3 might have an impact on tumor growth by engaging with cell cycle regulators [91,92]. To summarize, CBX3 functions as a transcriptional regulator in the Wnt/β-catenin signaling pathway, impacting the advancement of lung cancer by controlling gene expression, facilitating cell cycle progression, and interacting with pathways such as RAC1. In this context, the overexpression of CBX3 gene is associated with a negative outlook, suggesting its potential role as a target for treatment and a useful indicator of lung cancer prognosis [64,88].
4.4. Role of CBX3 in p53 Pathway
The tumor suppressor p53, a transcription factor responsible for initiating cell cycle arrest, apoptosis, and DNA repair when cells undergo stress, is involved in the multiple mechanisms that provide evidence supporting the role of CBX3 in lung cancer progression [93]. p53 activity is typically suppressed in normal cells by specific degradation regulated by the E3 ubiquitin ligase mouse double minute 2 homolog (MDM2) [93]. MDM2 primarily targets p53 via proteasomal degradation. In addition, its auto-ubiquitination activity does not directly affect MDM2 itself, but rather its interactions with other proteins [94]. It has been observed that CBX3 interacts with and potentially enhances the stability of MDM2 in lung cancer cells, inhibiting its auto-ubiquitination. However, the precise mechanism by which CBX3 enhances the stability of MDM2 is still unknown, and more research is required to elucidate this interaction [44,94]. CBX3 facilitates the survival and growth of lung cancer cells by indirectly impeding the activity of p53 through the MDM2 axis [95]. Additionally, CBX3 can directly suppress the expression of p53 target genes responsible for inhibiting growth, regardless of its impact on p53 protein levels [96]. Collectively, these findings show that CBX3 is able to bypass the tumor-suppressing effects of p53 and promote the development of lung tumors [97]. Thus, targeting the CBX3-MDM2-p53 pathway could potentially offer a novel therapeutic approach to restore the functionality of p53 in lung cancer.
4.5. Role of CBX3 in ErbB Pathway
The ErbB signaling pathway is crucial in controlling numerous cellular processes, such as differentiation, proliferation, migration, adhesion, and apoptosis [98]. This pathway is activated by EGF-like growth factor ligands’ attachment to the outer part of ErbB receptors [99]. This causes the receptors to form dimers, which can be either homodimers or heterodimers with other members of the same family. The ErbB family consists of four members: ErbB1 (HER1), ErbB2 (HER2), ErbB3 (HER3), and ErbB4 (HER4) [100]. When the receptors dimerize, their inherent tyrosine kinase activity is triggered, leading to autophosphorylation and activating the downstream signaling cascades, including the MAPK, Akt, and JAK/STAT pathways [99,100]. Although the precise mechanism by which CBX3 contributes to lung cancer is far to be completely elucidated, some evidence points to its potential interaction with EGFR signaling. Intriguingly, a co-occurrence of positive CBX3 expression and EGFR mutations was found in NSCLC samples [62]. However, in the same study, the expression of CBX3 remained unaltered in EGFR mutant NSCLC cell lines treated with the EGFR inhibitor gefitinib [101], suggesting that the downstream signaling of EGFR may not influence expression of CBX3 [98]. CBX3 is overexpressed in NSCLC and is associated with an unfavorable prognosis. Nonetheless, further investigation is required to clarify its exact function in interacting with the ErbB/EGFR pathway during the advancement of lung cancer. Specifically, future studies should focus on determining whether CBX3 governs the expression of ErbB pathway members via epigenetic regulation, investigating the impact of EGFR signaling on CBX3 activity, and elucidating the mechanisms by which CBX3-mediated gene regulation enhances lung cancer development and survival.
4.6. Role of CBX3 in MAPK Pathway
The MAPK/ERK pathway is a crucial signaling cascade that regulates cellular growth and survival [63]. CBX3 is able to inhibit transcription of the ARHGAP24 gene by interacting with H3K9me3 and binding to proteins that modify the structure of chromatin, such as TRIM28 and TRIM24, at the ARHGAP24 promoter. As a result, the expression of ARHGAP24 is suppressed, leading to an increase in RAC1 activity. Subsequently, RAC1 stimulates receptor tyrosine kinases on the cell membrane, propagating signals through the conventional MAPK cascade involving MEK1/2, ERK1/2, Ras, and Raf. Ultimately, activated ERK1/2 migrates to the nucleus and phosphorylates many transcription factors that control the proliferation and survival of cancer cells [102]. CBX3 also impacts other pathways such as PI3K/AKT and Ras signaling, which converge on MAPK. For example, AKT is able to phosphorylate and inhibit Raf proteins, providing crosstalk between these pathways. Ras functions directly upstream of Raf in the MAPK cascade [102]. In conclusion, targeting the CBX3-MAPK network may represent a potential treatment strategy for controlling lung cancer caused by an overactive CBX3 and MAPK pathway.
5. Conclusions
Lung cancer is one of the leading causes of cancer-related deaths in industrialized countries due to the extremely variable contributing factors such as detrimental lifestyle behavior, including smoking [103]. Therefore, aberration of epigenetic mechanism significantly impacts on gene expression and divert cellular signaling pathways that are crucial for coordinating the correct growth, regulation, and functioning of cells. In lung cancer, and especially in NSCLC, CBX3 appears to act primarily as an oncoprotein, promoting tumor growth and progression. Multiple studies show that CBX3 is overexpressed in NSCLC [104] and correlates with poor prognosis [27]. Mechanistically, CBX3 interacts with key oncogenic pathways such as PI3K/AKT, Ras, Wnt/β-catenin, and p53, thereby stimulating proliferation, inhibiting apoptosis, and increasing therapeutic resistance [81]. It may also contribute to a stem cell-like phenotype in lung cancer cells [105]. The upregulation of CBX3 in smoking-associated LUAD and its inhibition of tumor suppressors such as ARHGAP24 and NCOR2/ZBTB7A provide further evidence for its oncogenic role [63]. Carefully designed preclinical studies are essential before CBX3-based therapies can progress to the clinic. Additional mechanistic and functional studies of CBX3 hold the promise of uncovering new insights into disease pathogenesis and unveiling novel therapeutic opportunities. Thanks to the spread of increasingly advanced technologies such as those based on NGS techniques, it will be possible to extend studies aimed at clarifying the complex mechanisms that underlie the functioning of CBX3. Such studies could be particularly useful in the context of lung cancer, providing a more personalized molecular signature in high-risk patients.
Author Contributions
M.C. was responsible for the conceptualization, design of the study and image processing. L.A. supervised the study. M.A.W. and B.G. wrote and edited the manuscript. A.N., N.D.G., V.Q., and N.M. revised the manuscript. All authors read and approved the final manuscript.
Funding
This work was supported by PNRR-MAD-2022-12376723; PNRR-CN3, National Centre for Gene Therapy and Drugs Based on RNA Technology, cod:CN000000041. PNRR Project ANTHEM (AdvaNced Technologies for Human-cEntred Medicine) CUP: B53C22006540001. EPI- MET Fondo Crescita Sostenibile – Accordi per l’Innovazione D.M. 31.12.2021, D.D. 18.03.2022 no. 34; no. project F/310034/03/X56 (VANVITELLI). PRIN P2022F3YRF; Bando di Ateneo per il finanziamento di progetti di ricerca fondamentale ed applicata dedicato ai giovani Ricercatori D.R. no. 834 del 30/09/2022: IDEA (CUP: B63C22001470005). VALERE: Vanvitelli per la Ricerca Program: EPInhibitDRUGre (CUP B66J20000680005). NDG. was supported by PON Ricerca e Innovazione 2014–2020 Linea 1, AIM (AIM1859703); MUR-PRIN/PNRR2022: P2022KMP9K (MC); Programma V: ALERE 2020 Progetto competitivo “NETWINS” D.R. no. 138 of 17/02/2020. MAW is PhD student in translational medicine.
Data Availability Statement
Data sharing is not applicable.
Acknowledgments
We wish to thank C. Fisher for English-language editing of the manuscript. The figures in this review article were created with BioRender.com.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
| ARHGAP24 | rho GTPase Activating Protein 24 |
| BAX | bcl2-associated X protein |
| BRCA1 | breast cancer gene 1 |
| CBX | chromobox |
| CHD | chromodomain |
| CSD | chromoshadow domain |
| DSBs | DNA double-strand breaks |
| DSBs | double-strand breaks |
| EGFR | epidermal growth factor receptor |
| EZH1/2 | enhancer of zeste homologous 1/2 |
| HR | homologous recombination |
| LUAD | lung adenocarcinoma |
| MDM2 | mouse double minute 2 homolog |
| mRNA | messenger RNA |
| SMC | structural maintenance of chromosomes |
| NCAPG | non-SMC Condensin I Complex Subunit G |
| NICD | notch intracellular domain |
| NSCLC | non-small cell lung cancer |
| PI3K | phosphoinositide 3-kinase |
| PMAIP1 | phorbol-12-myristate-13-acetate-induced protein 1 |
| PRC1 | polycomb (Pc) inhibitory complex 1 |
| PRC2 | polycomb Repressive Complex 2 |
| PUMA p53 | upregulated modulator of apoptosis |
| RAC1 | active Ras-related C3 botulinum toxin substrate 1 |
| RBBP4/7 | retinoblastoma binding protein 4/7 |
| STAT3 | signal transducer and activator of transcription 3 |
| TRIM | tripartite motif-containing |
| ZBTB7A | zinc finger and BTB domain containing 7A |
References
- Tang, Q.; Cheng, J.; Cao, X.; Surowy, H.; Burwinkel, B. Blood-Based DNA Methylation as Biomarker for Breast Cancer: A Systematic Review. Clin Epigenetics 2016, 8, 115. [Google Scholar] [CrossRef]
- Chan, S.C.H.; Liang, J.Q. Advances in Tests for Colorectal Cancer Screening and Diagnosis. Expert Rev Mol Diagn 2022, 22, 449–460. [Google Scholar] [CrossRef]
- Tobi, E.W.; Slieker, R.C.; Luijk, R.; Dekkers, K.F.; Stein, A.D.; Xu, K.M.; Biobank-based Integrative Omics Studies Consortium; Slagboom, P. E.; van Zwet, E.W.; Lumey, L.H.; et al. DNA Methylation as a Mediator of the Association between Prenatal Adversity and Risk Factors for Metabolic Disease in Adulthood. Sci Adv 2018, 4, eaao4364. [Google Scholar] [CrossRef]
- Nicetto, D.; Zaret, K.S. Role of H3K9me3 Heterochromatin in Cell Identity Establishment and Maintenance. Curr Opin Genet Dev 2019, 55, 1–10. [Google Scholar] [CrossRef]
- Ho, L.; Crabtree, G.R. Chromatin Remodelling during Development. Nature 2010, 463, 474–484. [Google Scholar] [CrossRef]
- Goldberg, A.D.; Allis, C.D.; Bernstein, E. Epigenetics: A Landscape Takes Shape. Cell 2007, 128, 635–638. [Google Scholar] [CrossRef]
- Allis, C.D.; Jenuwein, T. The Molecular Hallmarks of Epigenetic Control. Nat Rev Genet 2016, 17, 487–500. [Google Scholar] [CrossRef]
- Wu, D.; Liu, T. Studying Reversible Protein Post-Translational Modification through Co-Translational Modification. Chembiochem 2023, 24, e202200716. [Google Scholar] [CrossRef]
- Biswas, S.; Rao, C.M. Epigenetic Tools (The Writers, The Readers and The Erasers) and Their Implications in Cancer Therapy. Eur J Pharmacol 2018, 837, 8–24. [Google Scholar] [CrossRef]
- Vann, K.R.; Klein, B.J.; Kutateladze, T.G. Mechanistic Similarities in Recognition of Histone Tails and DNA by Epigenetic Readers. Curr Opin Struct Biol 2021, 71, 1–6. [Google Scholar] [CrossRef]
- Beyer, J.N.; Raniszewski, N.R.; Burslem, G.M. Advances and Opportunities in Epigenetic Chemical Biology. ChemBioChem 2021, 22, 17–42. [Google Scholar] [CrossRef]
- Cao, G.; Xuan, X.; Hu, J.; Zhang, R.; Jin, H.; Dong, H. How Vascular Smooth Muscle Cell Phenotype Switching Contributes to Vascular Disease. Cell Commun Signal 2022, 20, 180. [Google Scholar] [CrossRef]
- Yang, A.Y.; Kim, H.; Li, W.; Kong, A.-N.T. Natural Compound-Derived Epigenetic Regulators Targeting Epigenetic Readers, Writers and Erasers. Curr Top Med Chem 2016, 16, 697–713. [Google Scholar] [CrossRef]
- Shukla, S.; Ying, W.; Gray, F.; Yao, Y.; Simes, M.L.; Zhao, Q.; Miao, H.; Cho, H.J.; González-Alonso, P.; Winkler, A.; et al. Small-Molecule Inhibitors Targeting Polycomb Repressive Complex 1 RING Domain. Nat Chem Biol 2021, 17, 784–793. [Google Scholar] [CrossRef]
- Vidal, M.; Starowicz, K. Polycomb Complexes PRC1 and Their Function in Hematopoiesis. Exp Hematol 2017, 48, 12–31. [Google Scholar] [CrossRef]
- Ma, R.; Zhang, Y.; Sun, T.; Cheng, B. Epigenetic Regulation by Polycomb Group Complexes: Focus on Roles of CBX Proteins. J Zhejiang Univ Sci B 2014, 15, 412–428. [Google Scholar] [CrossRef]
- Comet, I.; Riising, E.M.; Leblanc, B.; Helin, K. Maintaining Cell Identity: PRC2-Mediated Regulation of Transcription and Cancer. Nat Rev Cancer 2016, 16, 803–810. [Google Scholar] [CrossRef]
- Zoroddu, S.; Marchesi, I.; Bagella, L. PRC2: An Epigenetic Multiprotein Complex with a Key Role in the Development of Rhabdomyosarcoma Carcinogenesis. Clin Epigenetics 2021, 13, 156. [Google Scholar] [CrossRef]
- Ntziachristos, P.; Mullenders, J.; Trimarchi, T.; Aifantis, I. Mechanisms of Epigenetic Regulation of Leukemia Onset and Progression. Adv Immunol 2013, 117, 1–38. [Google Scholar] [CrossRef]
- Parreno, V.; Martinez, A.-M.; Cavalli, G. Mechanisms of Polycomb Group Protein Function in Cancer. Cell Res 2022, 32, 231–253. [Google Scholar] [CrossRef]
- Brockman, Q.R.; Scherer, A.; McGivney, G.R.; Gutierrez, W.R.; Voigt, A.P.; Isaacson, A.L.; Laverty, E.A.; Roughton, G.; Knepper-Adrian, V.; Darbro, B.; et al. PRC2 Loss Drives MPNST Metastasis and Matrix Remodeling. JCI Insight 2022, 7. [Google Scholar] [CrossRef]
- Duan, R.; Du, W.; Guo, W. EZH2: A Novel Target for Cancer Treatment. J Hematol Oncol 2020, 13, 104. [Google Scholar] [CrossRef]
- Kim, J.; Kingston, R.E. The CBX Family of Proteins in Transcriptional Repression and Memory. J Biosci 2020, 45. [Google Scholar] [CrossRef]
- Vincenz, C.; Kerppola, T.K. Different Polycomb Group CBX Family Proteins Associate with Distinct Regions of Chromatin Using Nonhomologous Protein Sequences. Proc Natl Acad Sci U S A 2008, 105, 16572–16577. [Google Scholar] [CrossRef]
- Jaensch, E.S.; Zhu, J.; Cochrane, J.C.; Marr, S.K.; Oei, T.A.; Damle, M.; McCaslin, E.Z.; Kingston, R.E. A Polycomb Domain Found in Committed Cells Impairs Differentiation When Introduced into PRC1 in Pluripotent Cells. Mol Cell 2021, 81, 4677–4691. [Google Scholar] [CrossRef]
- Zhang, X.; Zhou, W.; Zhang, Y.; Liu, Z. CBX3 Is a Prognostic Biomarker Correlated with ATR Activation and Immune Infiltration in Head and Neck Squamous Cell Carcinoma. Int J Gen Med 2022, 15, 1497–1508. [Google Scholar] [CrossRef]
- Xie, X.; Ning, Y.; Long, J.; Wang, H.; Chen, X. Diverse CBX Family Members as Potential Prognostic Biomarkers in Non-Small-Cell Lung Cancer. FEBS Open Bio 2020, 10, 2206–2215. [Google Scholar] [CrossRef]
- Nielsen, A.L.; Ortiz, J.A.; You, J.; Oulad-Abdelghani, M.; Khechumian, R.; Gansmuller, A.; Chambon, P.; Losson, R. Interaction with Members of the Heterochromatin Protein 1 (HP1) Family and Histone Deacetylation Are Differentially Involved in Transcriptional Silencing by Members of the TIF1 Family. EMBO J 1999, 18, 6385–6395. [Google Scholar] [CrossRef]
- Saunders, W.S.; Chue, C.; Goebl, M.; Craig, C.; Clark, R.F.; Powers, J.A.; Eissenberg, J.C.; Elgin, S.C.; Rothfield, N.F.; Earnshaw, W.C. Molecular Cloning of a Human Homologue of Drosophila Heterochromatin Protein HP1 Using Anti-Centromere Autoantibodies with Anti-Chromo Specificity. J Cell Sci 1993, 104, 573–582. [Google Scholar] [CrossRef]
- Paro, R.; Hogness, D.S. The Polycomb Protein Shares a Homologous Domain with a Heterochromatin-Associated Protein of Drosophila. Proc Natl Acad Sci U S A 1991, 88, 263–267. [Google Scholar] [CrossRef]
- Lachner, M.; O’Carroll, D.; Rea, S.; Mechtler, K.; Jenuwein, T. Methylation of Histone H3 Lysine 9 Creates a Binding Site for HP1 Proteins. Nature 2001, 410, 116–120. [Google Scholar] [CrossRef]
- Nielsen, A.L.; Oulad-Abdelghani, M.; Ortiz, J.A.; Remboutsika, E.; Chambon, P.; Losson, R. Heterochromatin Formation in Mammalian Cells: Interaction between Histones and HP1 Proteins. Mol Cell 2001, 7, 729–739. [Google Scholar] [CrossRef]
- Aasland, R.; Stewart, A.F. The Chromo Shadow Domain, a Second Chromo Domain in Heterochromatin-Binding Protein 1, HP1. Nucleic Acids Res 1995, 23, 3168–3173. [Google Scholar] [CrossRef]
- Cowieson, N.P.; Partridge, J.F.; Allshire, R.C.; McLaughlin, P.J. Dimerisation of a Chromo Shadow Domain and Distinctions from the Chromodomain as Revealed by Structural Analysis. Curr Biol 2000, 10, 517–525. [Google Scholar] [CrossRef]
- Brasher, S. V; Smith, B.O.; Fogh, R.H.; Nietlispach, D.; Thiru, A.; Nielsen, P.R.; Broadhurst, R.W.; Ball, L.J.; Murzina, N. V; Laue, E.D. The Structure of Mouse HP1 Suggests a Unique Mode of Single Peptide Recognition by the Shadow Chromo Domain Dimer. EMBO J 2000, 19, 1587–1597. [Google Scholar] [CrossRef]
- Ruan, J.; Ouyang, H.; Amaya, M.F.; Ravichandran, M.; Loppnau, P.; Min, J.; Zang, J. Structural Basis of the Chromodomain of Cbx3 Bound to Methylated Peptides from Histone H1 and G9a. PLoS One 2012, 7, e35376. [Google Scholar] [CrossRef]
- Lachner, M.; O’Carroll, D.; Rea, S.; Mechtler, K.; Jenuwein, T. Methylation of Histone H3 Lysine 9 Creates a Binding Site for HP1 Proteins. Nature 2001, 410, 116–120. [Google Scholar] [CrossRef]
- Ligresti, G.; Caporarello, N.; Meridew, J.A.; Jones, D.L.; Tan, Q.; Choi, K.M.; Haak, A.J.; Aravamudhan, A.; Roden, A.C.; Prakash, Y.S.; et al. CBX5/G9a/H3K9me-Mediated Gene Repression Is Essential to Fibroblast Activation during Lung Fibrosis. JCI Insight 2019, 5. [Google Scholar] [CrossRef]
- Casale, A.M.; Cappucci, U.; Fanti, L.; Piacentini, L. Heterochromatin Protein 1 (HP1) Is Intrinsically Required for Post-Transcriptional Regulation of Drosophila Germline Stem Cell (GSC) Maintenance. Sci Rep 2019, 9, 4372. [Google Scholar] [CrossRef]
- Azzaz, A.M.; Vitalini, M.W.; Thomas, A.S.; Price, J.P.; Blacketer, M.J.; Cryderman, D.E.; Zirbel, L.N.; Woodcock, C.L.; Elcock, A.H.; Wallrath, L.L.; et al. Human Heterochromatin Protein 1α Promotes Nucleosome Associations That Drive Chromatin Condensation. J Biol Chem 2014, 289, 6850–6861. [Google Scholar] [CrossRef]
- Lomberk, G.; Wallrath, L.; Urrutia, R. The Heterochromatin Protein 1 Family. Genome Biol 2006, 7, 228. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.; Li, H.; Liang, X.; Xiang, Z. CBX3 Promotes Colon Cancer Cell Proliferation by CDK6 Kinase-Independent Function during Cell Cycle. Oncotarget 2017, 8, 19934–19946. [Google Scholar] [CrossRef] [PubMed]
- Smallwood, A.; Hon, G.C.; Jin, F.; Henry, R.E.; Espinosa, J.M.; Ren, B. CBX3 Regulates Efficient RNA Processing Genome-Wide. Genome Res 2012, 22, 1426–1436. [Google Scholar] [CrossRef] [PubMed]
- Niu, H.; Chen, P.; Fan, L.; Sun, B. Comprehensive Pan-Cancer Analysis on CBX3 as a Prognostic and Immunological Biomarker. BMC Med Genomics 2022, 15, 29. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Su, T.; Xue, Y.; Cheng, C.; Lay, F.D.; McKee, R.A.; Li, M.; Vashisht, A.; Wohlschlegel, J.; Novitch, B.G.; et al. Cbx3 Maintains Lineage Specificity during Neural Differentiation. Genes Dev 2017, 31, 241–246. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Shilatifard, A. Epigenetic Modifications of Histones in Cancer. Genome Biol 2019, 20, 245. [Google Scholar] [CrossRef] [PubMed]
- Saldi, T.; Cortazar, M.A.; Sheridan, R.M.; Bentley, D.L. Coupling of RNA Polymerase II Transcription Elongation with Pre-MRNA Splicing. J Mol Biol 2016, 428, 2623–2635. [Google Scholar] [CrossRef] [PubMed]
- Sheldon, L.A. Inhibition of E2F1 Activity and Cell Cycle Progression by Arsenic via Retinoblastoma Protein. Cell Cycle 2017, 16, 2058–2072. [Google Scholar] [CrossRef] [PubMed]
- Pei, Y.; Banerjee, S.; Sun, Z.; Jha, H.C.; Saha, A.; Robertson, E.S. EBV Nuclear Antigen 3C Mediates Regulation of E2F6 to Inhibit E2F1 Transcription and Promote Cell Proliferation. PLoS Pathog 2016, 12, e1005844. [Google Scholar] [CrossRef]
- Marei, H.E.; Althani, A.; Afifi, N.; Hasan, A.; Caceci, T.; Pozzoli, G.; Morrione, A.; Giordano, A.; Cenciarelli, C. P53 Signaling in Cancer Progression and Therapy. Cancer Cell Int 2021, 21, 703. [Google Scholar] [CrossRef]
- Wang, Z.; Jia, R.; Wang, L.; Yang, Q.; Hu, X.; Fu, Q.; Zhang, X.; Li, W.; Ren, Y. The Emerging Roles of Rad51 in Cancer and Its Potential as a Therapeutic Target. Front Oncol 2022, 12. [Google Scholar] [CrossRef]
- Orhan, E.; Velazquez, C.; Tabet, I.; Sardet, C.; Theillet, C. Regulation of RAD51 at the Transcriptional and Functional Levels: What Prospects for Cancer Therapy? Cancers 2021, 13, 2930. [Google Scholar] [CrossRef] [PubMed]
- Li, E.; Xia, M.; Du, Y.; Long, K.; Ji, F.; Pan, F.; He, L.; Hu, Z.; Guo, Z. METTL3 Promotes Homologous Recombination Repair and Modulates Chemotherapeutic Response in Breast Cancer by Regulating the EGF/RAD51 Axis. Elife 2022, 11. [Google Scholar] [CrossRef]
- Wang, H.; Zhao, W.; Wang, J.; Zhang, Z. Clinicopathological Significance of CBX3 in Colorectal Cancer: An Intensive Expression Study Based on Formalin-fixed and Paraffin-embedded Tissues. Pathol Int 2022, 72, 107–116. [Google Scholar] [CrossRef]
- Li, E.; Xia, M.; Du, Y.; Long, K.; Ji, F.; Pan, F.; He, L.; Hu, Z.; Guo, Z. METTL3 Promotes Homologous Recombination Repair and Modulates Chemotherapeutic Response in Breast Cancer by Regulating the EGF/RAD51 Axis. Elife 2022, 11. [Google Scholar] [CrossRef]
- Wang, Z.; Jia, R.; Wang, L.; Yang, Q.; Hu, X.; Fu, Q.; Zhang, X.; Li, W.; Ren, Y. The Emerging Roles of Rad51 in Cancer and Its Potential as a Therapeutic Target. Front Oncol 2022, 12. [Google Scholar] [CrossRef]
- Orhan, E.; Velazquez, C.; Tabet, I.; Sardet, C.; Theillet, C. Regulation of RAD51 at the Transcriptional and Functional Levels: What Prospects for Cancer Therapy? Cancers 2021, 13, 2930. [Google Scholar] [CrossRef]
- CBX3 Promotes Gastric Cancer Progression and Affects Factors Related to Immunotherapeutic Responses.
- CBX3 Predicts an Unfavorable Prognosis and Promotes Tumorigenesis in Osteosarcoma.
- Alam, H.; Li, N.; Dhar, S.S.; Wu, S.J.; Lv, J.; Chen, K.; Flores, E.R.; Baseler, L.; Lee, M.G. HP1γ Promotes Lung Adenocarcinoma by Downregulating the Transcription-Repressive Regulators NCOR2 and ZBTB7A. Cancer Res 2018, 78, 3834–3848. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.; Zhao, X.; Xia, L.; Lian, J.; You, J. Clinicopathological and Prognostic Significance of CBX3 Expression in Human Cancer: A Systematic Review and Meta-Analysis. Dis Markers 2020, 2020, 1–11. [Google Scholar] [CrossRef]
- CBX3/Heterochromatin Protein 1 Gamma Is Significantly Upregulated in Patients with Non-Small Cell Lung Cancer.
- Jin, X.; Zhang, B.; Zhang, H.; Yu, H. Smoking-Associated Upregulation of CBX3 Suppresses ARHGAP24 Expression to Activate Rac1 Signaling and Promote Tumor Progression in Lung Adenocarcinoma. Oncogene 2022, 41, 538–549. [Google Scholar] [CrossRef]
- Wang, J.; Yang, B.; Zhang, X.; Liu, S.; Pan, X.; Ma, C.; Ma, S.; Yu, D.; Wu, W. Chromobox Proteins in Cancer: Multifaceted Functions and Strategies for Modulation (Review). Int J Oncol 2023, 62. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Chang, L.; Yao, Y.; Chao, C.; Ge, Z.; Fan, C.; Yu, H.; Wang, B.; Yang, J. Role of the CBX Molecular Family in Lung Adenocarcinoma Tumorigenesis and Immune Infiltration. Front Genet 2021, 12. [Google Scholar] [CrossRef] [PubMed]
- Hu, K.; Yao, L.; Xu, Z.; Yan, Y.; Li, J. Prognostic Value and Therapeutic Potential of CBX Family Members in Ovarian Cancer. Front Cell Dev Biol 2022, 10, 832354. [Google Scholar] [CrossRef] [PubMed]
- Czerwinska, P.; Mackiewicz, A.A. Mining Transcriptomic Data to Uncover the Association between CBX Family Members and Cancer Stemness. Int J Mol Sci 2022, 23. [Google Scholar] [CrossRef] [PubMed]
- Sundar, I.K.; Nevid, M.Z.; Friedman, A.E.; Rahman, I. Cigarette Smoke Induces Distinct Histone Modifications in Lung Cells: Implications for the Pathogenesis of COPD and Lung Cancer. J Proteome Res 2014, 13, 982–996. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Yu, H.; Ren, D.; Sun, Y.; Guo, F.; Cai, H.; Zhou, C.; Zhou, Y.; Jin, X.; Wu, H. CBX3 Regulated By YBX1 Promotes Smoking-Induced Pancreatic Cancer Progression via Inhibiting SMURF2 Expression. Int J Biol Sci 2022, 18, 3484–3497. [Google Scholar] [CrossRef] [PubMed]
- Haley, J.A.; Haughney, E.; Ullman, E.; Bean, J.; Haley, J.D.; Fink, M.Y. Altered Transcriptional Control Networks with Trans-Differentiation of Isogenic Mutant-KRas NSCLC Models. Front Oncol 2014, 4, 344. [Google Scholar] [CrossRef] [PubMed]
- Bosso, G.; Cipressa, F.; Tullo, L.; Cenci, G. Co-Amplification of CBX3 with EGFR or RAC1 in Human Cancers Corroborated by a Conserved Genetic Interaction among the Genes. Cell Death Discov 2023, 9, 317. [Google Scholar] [CrossRef]
- Xu, H.; Jiang, C.; Chen, D.; Wu, Y.; Lu, J.; Zhong, L.; Yao, F. Analysis of Pan-Cancer Revealed the Immunological and Prognostic Potential of CBX3 in Human Tumors. Front Med (Lausanne) 2022, 9, 869994. [Google Scholar] [CrossRef]
- Chen, J.; Lin, Y.; Zheng, S.; Chen, Q.; Tang, S.; Zhong, X. CBX3 Promotes Clear Cell Renal Carcinoma through PI3K/AKT Activation and Aberrant Immunity. J Transl Med 2023, 21, 600. [Google Scholar] [CrossRef]
- Chen, H.; Han, C.; Liu, D.; Wang, F.; Ha, C. CBX3 Promotes Ovarian Cancer Progression by Regulating P53/P21-Mediated Glucose Metabolism via Inhibiting NCOR2. Archives of Medical Science 2022. [Google Scholar] [CrossRef]
- Taguchi, Y.-H.; Wang, H. Genetic Association between Amyotrophic Lateral Sclerosis and Cancer. Genes (Basel) 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Loboda, A.; Nebozhyn, M.; Klinghoffer, R.; Frazier, J.; Chastain, M.; Arthur, W.; Roberts, B.; Zhang, T.; Chenard, M.; Haines, B.; et al. A Gene Expression Signature of RAS Pathway Dependence Predicts Response to PI3K and RAS Pathway Inhibitors and Expands the Population of RAS Pathway Activated Tumors. BMC Med Genomics 2010, 3, 26. [Google Scholar] [CrossRef] [PubMed]
- Pavan, A.; Bragadin, A.B.; Calvetti, L.; Ferro, A.; Zulato, E.; Attili, I.; Nardo, G.; Dal Maso, A.; Frega, S.; Menin, A.G.; et al. Role of next Generation Sequencing-Based Liquid Biopsy in Advanced Non-Small Cell Lung Cancer Patients Treated with Immune Checkpoint Inhibitors: Impact of STK11, KRAS and TP53 Mutations and Co-Mutations on Outcome. Transl Lung Cancer Res 2021, 10, 202–220. [Google Scholar] [CrossRef] [PubMed]
- de Jesus, V.H.F.; Mathias-Machado, M.C.; de Farias, J.P.F.; Aruquipa, M.P.S.; Jácome, A.A.; Peixoto, R.D. Targeting KRAS in Pancreatic Ductal Adenocarcinoma: The Long Road to Cure. Cancers (Basel) 2023, 15. [Google Scholar] [CrossRef] [PubMed]
- Kalungi, F.; Nsubuga, A.; Anywar, G. Network Analysis and Molecular Docking Studies of Quercetin as a Potential Treatment for Prostate Cancer. In Silico Pharmacol 2023, 11, 24. [Google Scholar] [CrossRef] [PubMed]
- Lim, J.S.; Ibaseta, A.; Fischer, M.M.; Cancilla, B.; O’Young, G.; Cristea, S.; Luca, V.C.; Yang, D.; Jahchan, N.S.; Hamard, C.; et al. Intratumoural Heterogeneity Generated by Notch Signalling Promotes Small-Cell Lung Cancer. Nature 2017, 545, 360–364. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Chen, D.; Maguire, E.M.; He, S.; Chen, J.; An, W.; Yang, M.; Afzal, T.A.; Luong, L.A.; Zhang, L.; et al. Cbx3 Inhibits Vascular Smooth Muscle Cell Proliferation, Migration, and Neointima Formation. Cardiovasc Res 2018, 114, 443–455. [Google Scholar] [CrossRef] [PubMed]
- Kopan, R. Notch Signaling. Cold Spring Harb Perspect Biol 2012, 4. [Google Scholar] [CrossRef]
- Zhou, B.; Lin, W.; Long, Y.; Yang, Y.; Zhang, H.; Wu, K.; Chu, Q. Notch Signaling Pathway: Architecture, Disease, and Therapeutics. Signal Transduct Target Ther 2022, 7, 95. [Google Scholar] [CrossRef]
- Ragot, H.; Monfort, A.; Baudet, M.; Azibani, F.; Fazal, L.; Merval, R.; Polidano, E.; Cohen-Solal, A.; Delcayre, C.; Vodovar, N.; et al. Loss of Notch3 Signaling in Vascular Smooth Muscle Cells Promotes Severe Heart Failure Upon Hypertension. Hypertension 2016, 68, 392–400. [Google Scholar] [CrossRef]
- Zou, B.; Zhou, X.-L.; Lai, S.-Q.; Liu, J.-C. Notch Signaling and Non-Small Cell Lung Cancer. Oncol Lett 2018, 15, 3415–3421. [Google Scholar] [CrossRef] [PubMed]
- Janghorban, M.; Xin, L.; Rosen, J.M.; Zhang, X.H.-F. Notch Signaling as a Regulator of the Tumor Immune Response: To Target or Not To Target? Front Immunol 2018, 9, 1649. [Google Scholar] [CrossRef]
- Koni, M.; Pinnarò, V.; Brizzi, M.F. The Wnt Signalling Pathway: A Tailored Target in Cancer. Int J Mol Sci 2020, 21, 7697. [Google Scholar] [CrossRef]
- Yang, H.; Pu, L.; Li, R.; Zhu, R. NCAPG Is Transcriptionally Regulated by CBX3 and Activates the Wnt/β-Catenin Signaling Pathway to Promote Proliferation and the Cell Cycle and Inhibit Apoptosis in Colorectal Cancer. J Gastrointest Oncol 2023, 14, 900–912. [Google Scholar] [CrossRef]
- Mauser, R.; Kungulovski, G.; Keup, C.; Reinhardt, R.; Jeltsch, A. Application of Dual Reading Domains as Novel Reagents in Chromatin Biology Reveals a New H3K9me3 and H3K36me2/3 Bivalent Chromatin State. Epigenetics Chromatin 2017, 10, 45. [Google Scholar] [CrossRef]
- Fan, Y.; Li, H.; Liang, X.; Xiang, Z. CBX3 Promotes Colon Cancer Cell Proliferation by CDK6 Kinase-Independent Function during Cell Cycle. Oncotarget 2017, 8, 19934–19946. [Google Scholar] [CrossRef]
- Fan, Y.; Li, H.; Liang, X.; Xiang, Z. CBX3 Promotes Colon Cancer Cell Proliferation by CDK6 Kinase-Independent Function during Cell Cycle. Oncotarget 2017, 8, 19934–19946. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.-Y.; Cheng, C.-S.; Qu, C.; Wang, P.; Chen, H.; Meng, Z.-Q.; Chen, Z. Overexpression of CBX3 in Pancreatic Adenocarcinoma Promotes Cell Cycle Transition-Associated Tumor Progression. Int J Mol Sci 2018, 19. [Google Scholar] [CrossRef] [PubMed]
- Saxena, K.; Konopleva, M. An Expert Overview of Emerging Therapies for Acute Myeloid Leukemia: Novel Small Molecules Targeting Apoptosis, P53, Transcriptional Regulation and Metabolism. Expert Opin Investig Drugs 2020, 29, 973–988. [Google Scholar] [CrossRef]
- Sandy, Z.; da Costa, I.C.; Schmidt, C.K. More than Meets the ISG15: Emerging Roles in the DNA Damage Response and Beyond. Biomolecules 2020, 10. [Google Scholar] [CrossRef] [PubMed]
- Carr, S.M.; Munro, S.; La Thangue, N.B. Lysine Methylation and the Regulation of P53. Essays Biochem 2012, 52, 79–92. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Yang, N.; Chen, Y.; Zhu, M.; Lian, Y.; Xiong, Z.; Wang, B.; Feng, L.; Jia, X. An Integrated Strategy for Effective-Component Discovery of Astragali Radix in the Treatment of Lung Cancer. Front Pharmacol 2020, 11, 580978. [Google Scholar] [CrossRef] [PubMed]
- Hirata, H.; Hinoda, Y.; Kikuno, N.; Kawamoto, K.; Suehiro, Y.; Tanaka, Y.; Dahiya, R. MDM2 SNP309 Polymorphism as Risk Factor for Susceptibility and Poor Prognosis in Renal Cell Carcinoma. Clin Cancer Res 2007, 13, 4123–4129. [Google Scholar] [CrossRef] [PubMed]
- Appert-Collin, A.; Hubert, P.; Crémel, G.; Bennasroune, A. Role of ErbB Receptors in Cancer Cell Migration and Invasion. Front Pharmacol 2015, 6, 283. [Google Scholar] [CrossRef] [PubMed]
- Fry, W.H.D.; Kotelawala, L.; Sweeney, C.; Carraway, K.L. Mechanisms of ErbB Receptor Negative Regulation and Relevance in Cancer. Exp Cell Res 2009, 315, 697–706. [Google Scholar] [CrossRef] [PubMed]
- Black, L.E.; Longo, J.F.; Carroll, S.L. Mechanisms of Receptor Tyrosine-Protein Kinase ErbB-3 (ERBB3) Action in Human Neoplasia. Am J Pathol 2019, 189, 1898–1912. [Google Scholar] [CrossRef] [PubMed]
- Cragg, M.S.; Kuroda, J.; Puthalakath, H.; Huang, D.C.S.; Strasser, A. Gefitinib-Induced Killing of NSCLC Cell Lines Expressing Mutant EGFR Requires BIM and Can Be Enhanced by BH3 Mimetics. PLoS Med 2007, 4, 1681–1689. [Google Scholar] [CrossRef] [PubMed]
- Uimari, O.; Rahmioglu, N.; Nyholt, D.R.; Vincent, K.; Missmer, S.A.; Becker, C.; Morris, A.P.; Montgomery, G.W.; Zondervan, K.T. Genome-Wide Genetic Analyses Highlight Mitogen-Activated Protein Kinase (MAPK) Signaling in the Pathogenesis of Endometriosis. Hum Reprod 2017, 32, 780–793. [Google Scholar] [CrossRef]
- Thandra, K.C.; Barsouk, A.; Saginala, K.; Aluru, J.S.; Barsouk, A. Epidemiology of Lung Cancer. Contemp Oncol (Pozn) 2021, 25, 45–52. [Google Scholar] [CrossRef]
- Chang, S.-C.; Lai, Y.-C.; Chen, Y.-C.; Wang, N.-K.; Wang, W.-S.; Lai, J.-I. CBX3/Heterochromatin Protein 1 Gamma Is Significantly Upregulated in Patients with Non-Small Cell Lung Cancer. Asia Pac J Clin Oncol 2018, 14, e283–e288. [Google Scholar] [CrossRef]
- Czerwinska, P.; Mackiewicz, A.A. Mining Transcriptomic Data to Uncover the Association between CBX Family Members and Cancer Stemness. Int J Mol Sci 2022, 23. [Google Scholar] [CrossRef]
Figure 1.
A. Ribbon diagram showing the chromodomain of HP1 complexed with histone H3 tail containing monomethyl lysine 9. B. Crystal structure of the HP1α, HP1β, HP1γ chromo shadow domains (left); schematic representation of HP1 isoform proteins (right). LR = linker region; CD = chromodomain; CSD = chomoshadow domain.
Figure 1.
A. Ribbon diagram showing the chromodomain of HP1 complexed with histone H3 tail containing monomethyl lysine 9. B. Crystal structure of the HP1α, HP1β, HP1γ chromo shadow domains (left); schematic representation of HP1 isoform proteins (right). LR = linker region; CD = chromodomain; CSD = chomoshadow domain.
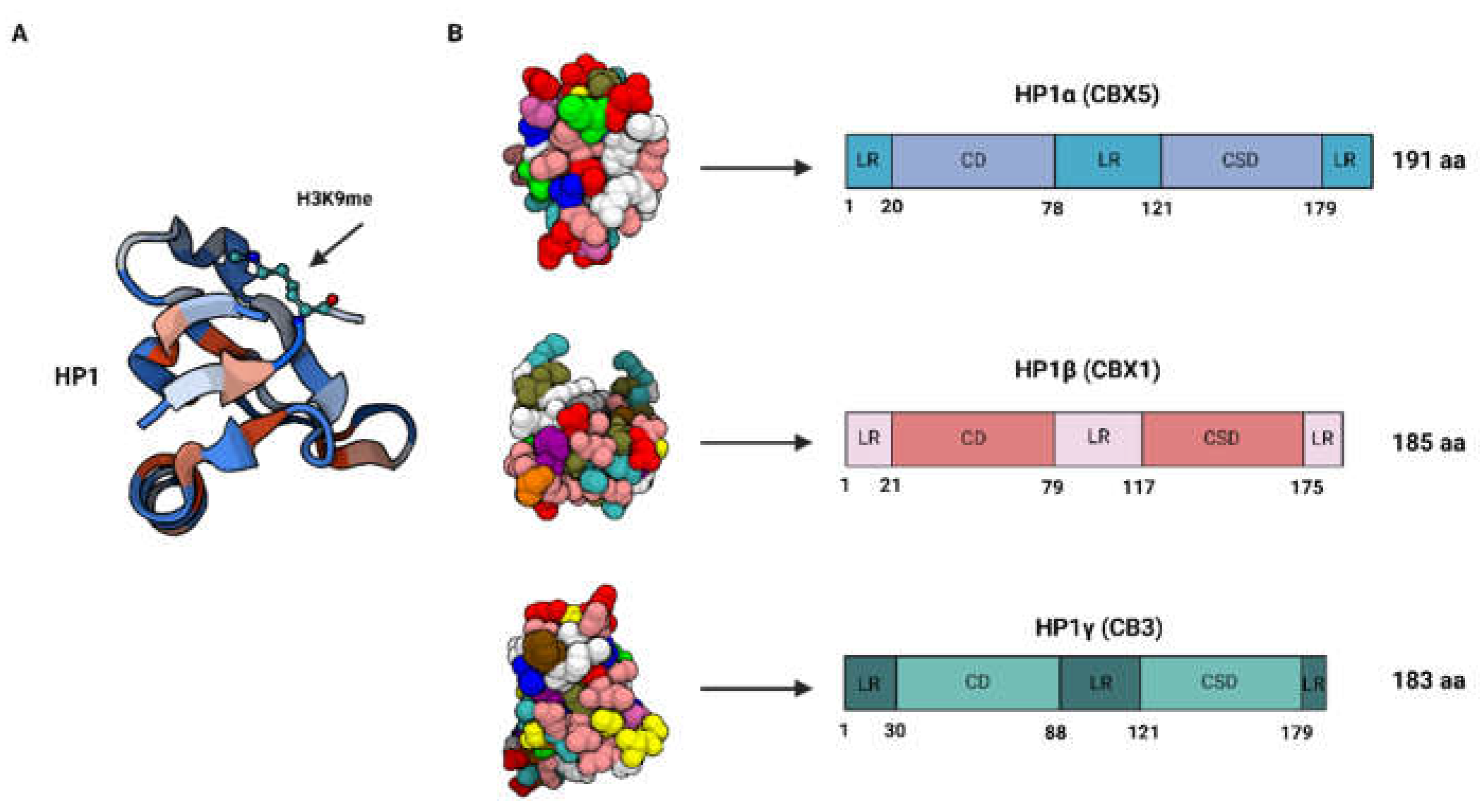
Figure 2.
Cellular pathways directly or indirectly modulated by CBX3 in lung cancer.
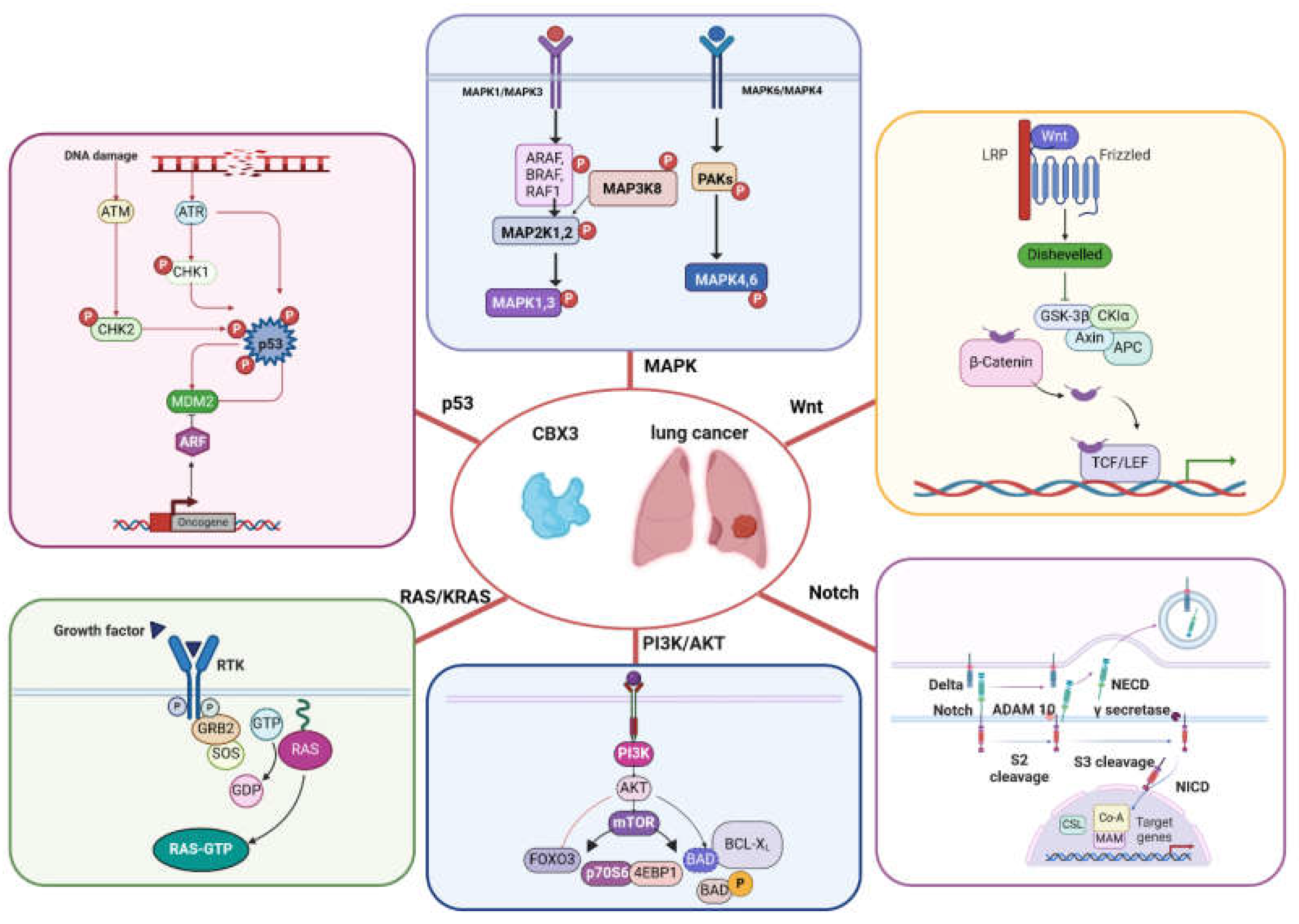

Table 1.
CBX3 interactors, their effects on lung cancer and pathways involvement via differential mechanisms of action.
Table 1.
CBX3 interactors, their effects on lung cancer and pathways involvement via differential mechanisms of action.
| Interactor | Type of Interaction | Effect on Lung Cancer | Pathway Involvement | Mechanism of Action |
|---|---|---|---|---|
| EGFR/RAC1 | Genetic Interaction | Co-amplification with CBX3 is associated with lung adenocarcinoma proliferation and poor prognosis | Not Specified | Increase in CBX3 mRNA leads to increased EGFR/RAC1 protein levels, promoting cancer cell proliferation [71] |
| CDK6/P21 | Transcriptional Regulation | CBX3 inhibits transcription of negative cell cycle regulators, promoting colorectal cancer cell proliferation; similar mechanisms may be involved in lung cancer | Cell Cycle Regulation |
CBX3 is able to inhibit transcription of CDK6 and p21, promoting cell proliferation [72] |
| CBX Molecular Family (CBX1/2/3/5/7) | Gene Expression | CBX3/5 expression is associated with poor prognosis in lung adenocarcinoma, while CBX7 shows the opposite effect | Tumorigenesis and Immune Infiltration |
Differential expression of CBX family members affects tumor progression and immune response [65] |
| PI3K/AKT Pathway | Activation | Although the study is on renal carcinoma, similar activation by CBX3 may occur in lung cancer, promoting metastasis and invasion | PI3K/AKT Pathway | CBX3 promotes cancer progression through PI3K/AKT activation, which regulates cell metastasis and invasion [73] |
| ARHGAP24 | Suppression | Smoking-associated upregulation of CBX3 suppresses ARHGAP24, activating RAC1 signaling and promoting tumor progression in lung adenocarcinoma | RAC1 Signaling | CBX3 overexpression leads to suppression of ARHGAP24, activating RAC1 and promoting tumor progression [63] |
| NCOR2 | Regulation | In ovarian cancer, CBX3 inhibits NCOR2, affecting p53/p21-mediated glucose metabolism; similar effects may occur in lung cancer | Glucose Metabolism | CBX3 inhibits NCOR2, affecting p53/p21-mediated pathways and potentially promoting cancer metabolism [74] |
| Immune System | Immunological Biomarker | CBX3 expression is related to immune cell infiltration and may serve as an immunological and prognostic biomarker in various cancers, including lung cancer | Immune Response |
CBX3 expression influences immune cell infiltration and tumor immunity, which varies based on tumor type [72] |
| Transcriptome/Metabolome | Biomarker Association | Possible association between CBX3 expression and transcriptome/metabolome changes in cancers, including lung cancer | Various Pathways | CBX3 expression may be linked to changes in the transcriptome and metabolome, affecting multiple cancer-related pathways [75] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



