
Submitted:
30 July 2024
Posted:
30 July 2024
You are already at the latest version
Abstract
The moist Afromontane forest of Ethiopia plays a crucial role in carbon storage and mitigating greenhouse gas (GHG) emissions. To guide effective conservation policies, it is essential to understand the relationship between species diversity and carbon storage in this unique ecosystem. Therefore, this study, aimed to evaluate the response of above-ground carbon (AGC) to species and functional diversity, and their effects on AGC in the Yayu Coffee Forest Biosphere Reserve (YCFBR). Using different plot sizes, we collected vegetation data from the core, buffer, and transitional zones of the reserve. We selected traits related to plant-life strategy, competition ability, response to disturbances, and climate change. Structural Equation Models (SEM) were employed to assess the effects of species diversity, functional diversity, and functional dominance on AGC. Data analysis was conducted using R software. Our results revealed that the effects of species richness on AGC were mediated through functional diversity and functional dominance, as demonstrated by both the full mediation and partial mediation models. This suggests that the selection effects and niche complementarity are mutually exclusive yet equally important for predicting AGC. The findings further indicate that functional diversity and functional dominance together accounted for a significant predictors of the AGC by 21% in the YCFBR. From a practical standpoint, findings highlight the importance of conserving species diversity as a means to maintain higher AGC in the tropical moist Afromontane forests. In conclusion, this research significantly contributes to scientific literature by providing valuable insights into the complex relationship between species diversity and carbon storage. The results emphasize the crucial role of functional diversity and dominance in predicting AGC, underscoring the importance of biodiversity conservation for climate mitigation efforts. These findings have practical implications for the development of effective conservation policies tailored to protect and sustain the carbon storage potential of this unique ecosystem.
Keywords:
above-ground carbon
; Ethiopia
; functional diversity
; moist Afromontane forest
; species diversity
; Yayu Coffee Forest Biosphere Reserve
1. Introduction
Understanding how biodiversity affects ecosystem functioning is a fundamental aim in an ecological research but demanding further investigation (Shen, 2016; Yi et al., 2020). There has been increasing interest in the relationship between functional diversity and carbon storage in the context of global climatic change over the past few decades, especially in natural multispecies forest ecosystems (Ding & Nunes, 2014). As both biodiversity and carbon storage are threatened by deforestation and climate change, it is imperative to understand the relationships between species diversity and carbon storage in tropical forest, which helps to inform conservation policies intended to mitigate GHG emissions (IPCC, 2023). Human activities, principally through emissions of GHG, have unequivocally caused global warming, with global surface temperature reaching 1.1 °C above 1850-1900 in 2011-2020 (IPCC, 2023). Adopting the trait-based approach, the relationship between species diversity and AGC, can be examined (Mensah et al., 2021). Studies have linked plant functional traits to carbon storage, since tree species with different characteristics have distinguishing abilities to capture, store, and release biomass carbon (Shen, 2016). Because biomass carbon is an important component of forest stand productivity, the relationship between biomass carbon and biodiversity can also be assimilated to the one of biodiversity and ecosystem function (Lasky et al., 2014).
Two non-mutually exclusive hypotheses have been proposed to explain the relationships between biodiversity and ecosystem functions (Yuan et al., 2018). These are the niche complementarity and mass ratio hypothesis. The concept of niche complementarity is frequently invoked to account for increased biomass yield and productivity within diverse ecosystems. Nonetheless, higher biomass can also arise due to a greater possibility of species, or specific traits, exhibiting robust reactions to resources or exerting significant impacts on the ecosystem (Reich et al., 2004). Some researchers suggest that both theories could influence the impact of diversity on ecosystem functions, particularly through competition and resource partitioning among species (Carroll et al., 2011; Mensah et al. 2018). Research support both the selection effects and niche complementarity hypothesis on forest ecosystems functioning (Rodríguez-Hernández, 2021). Sasaki & Lauenroth (2011) provide support for the selection effects hypothesis, while Michalet et al (2021) and Xu et al (2022) offer evidence in favor of niche complementarity hypothesis. The niche complementarity hypothesis suggests that high species diversity or functional diversity can enhance ecosystem functions, and hence biomass productivity increases through resource-use efficiency by interacting individuals due to the niche partitioning or facilitation (Mensah et al., 2018; Zhang et al., 2021).
Selection effects highlight the implication of dominant species in maintaining ecosystem function and enhancing the relationship between biodiversity and ecosystem function. This relationship is evident when the possibility of dominant species occurring increases with greater tree species diversity, often due to competitive interactions (Mensah et al., 2018). Both species richness (niche complementarity) and the community-weighted mean (CWM) of trait values (selection effects) support the beneficial effects of forest structural diversity on above-ground biomass (AGB); however, the relative importance of these effects differed depending on the type of forest (Noulèkoun et al., 2024). Also, the availability of resources (water and nutrients) limits the relationships between forest diversity and AGB. This knowledge can offer new understandings into the role of functional traits as key determinants of the effects of forest diversity on AGB in tropical forests (Mensah et al. 2018).
There is growing support for the idea that functional traits are indicative of the unique functional roles that species coexist in a given community, and how they are linked to differences in their ecological niches. Thus, functional traits may be more effective at predicting the relationship between biodiversity and productivity compared to taxonomic diversity (Mensah et al., 2016; Weisser et al., 2017; Yi et al., 2020). These indicated that functional diversity may be more helpful to maintaining ecosystem multi-functionality than species diversity (Kong et al., 2023). Therefore, understanding how the relative contributions of these two mechanisms change according to the context may inform about new strategies for the forest management and biodiversity conservation.
Findings shows, AGC had significant positive correlation with species richness and tree density (Mensah et al., 2021). Similarly, Mengistu et al. (2021) found that linking diversity to AGC provides prediction of ecosystem functions and the relationship is direct. The positive effects of species richness on AGC are significantly explained by non-mutually exclusive mechanisms such as complementarity and selection effects where resource availability and environmental filtering play a central role (Mensah et al., 2023). As such, AGC can increase due to a few highly productive and dominant species (selection effects), or a better performance of coexisting species through increased resource use efficiency (niche complementarity) (Mensah et al., 2023). For example, Mensah et al.(2023) found that shifting diversity-carbon relationship at local scale can be explained by the effects of stand structural complexity and large-size tree. On the other hand, there is also negative relationship between diversity and carbon, this is due to the influence of diversity on carbon is scale dependent and environmental variation (Sullivan et al., 2017).
In addition, environmental factors also have a significant influence on the relationship between ecosystem functioning and diversity (Mensah et al., 2023); however, if local environmental factors are ignored in large-scale studies, some significant connections may go unnoticed (Gourlet-Fleury et al., 2011; Mensah et al., 2023; Shen et al., 2016). Because, the shift in species richness-AGC relationships under contrasting environmental conditions suggests that larger scale patterns of diversity and carbon are indeed regulated by environmental conditions, because species distributions, composition and growth performance are governed by adaptations to physical conditions of the environment (Mensah et al., 2023). For example, Ma et al. (2010) found that large-scale environmental variation had significant effects in regulating the relationship between species diversity of grasses and above-ground productivity. From this, environmental variables are an important predictors of carbon and diversity-carbon relationship are scale dependent and environmental variation effects (Sullivan et al., 2017).
Among the various traits used to study the predictive role of species diversity for AGC and to clearly elucidate their relationships, forest ecologists commonly use tree maximum height (Hmax), wood density (WD) and specific leaf area (SLA). Also, functional diversity and community weighted mean of Hmax, WD and SLA are important traits used as predictor of AGC (Mensah et al., 2021). The thing is these attributes of functional traits are linked to hypotheses of complementarity and selection effect in ecological studies, based on the assumption that plant functional traits can explain differences in fitness and niches, resulting in competitive exclusion and coexistence, respectively (Cadotte, 2017; Carroll et al., 2011). This shows that, both complementarity and selection are complementary and can partly result from either trait dissimilarity or community weighted mean (Mensah et at., 2024). Accordingly, it has been shown that both complementarity and selection effects are described as non-mutually exclusive mechanisms interacting to regulate ecosystems functioning (Fargione et al., 2007; Mensah et al., 2018; Noulèkoun et al., 2024). Therefore, examining the relationship between traits and their diversity in different systems, including variations within species, is now essential in understanding function (Cadotte, 2017; Laughlin, 2014; Violle et al., 2012). Because, these traits provides a proxy for the potential tree growth performance, (Yuan et al., 2018), species tolerance to climate change (Poorter et al., 2015) and categorize species into fast growth (acquisitive species) and slow growth (conservative species) (Sainge et al., 2019).
Ecosystem functioning is related to taxonomic, functional diversity and functional dominance (Zhang et al., 2021). However, there is the lack of studies that have examined the relative importance of different metrics of biodiversity in relation to AGC levels in the moist Afromontane forest of Southwest Ethiopia, specifically within the YCFBR. Additionally, the influence of topographic variables (such as slope, aspect, and elevation) and anthropogenic disturbances on these relationships has not been thoroughly investigated at a country level. Furthermore, while it is widely recognized that both biodiversity and carbon storage are threatened by deforestation and climate change, there is limited knowledge about the quantitative and causal relationships between forest biodiversity and AGC (Brockerhoff et al., 2017). Therefore, it is crucial to understand the relationships between species diversity and AGC in the YCFBR, to effectively mitigate GHG emissions resulting from deforestation, primarily driven by anthropogenic impacts in the region. Based on this motivation, the study aimed to explore the response of AGC to species richness, functional diversity, and functional dominance, and to investigate the potential mediating effects of functional diversity and functional dominance on the relationship between species richness and AGC in the YCFBR. By addressing these research objectives, the study seeks to improve our understanding and prediction of the consequences of biodiversity loss on ecosystem functioning through the use of traits-based diversity measurement.
2. Materials and Methods
2.1. Sampling Design
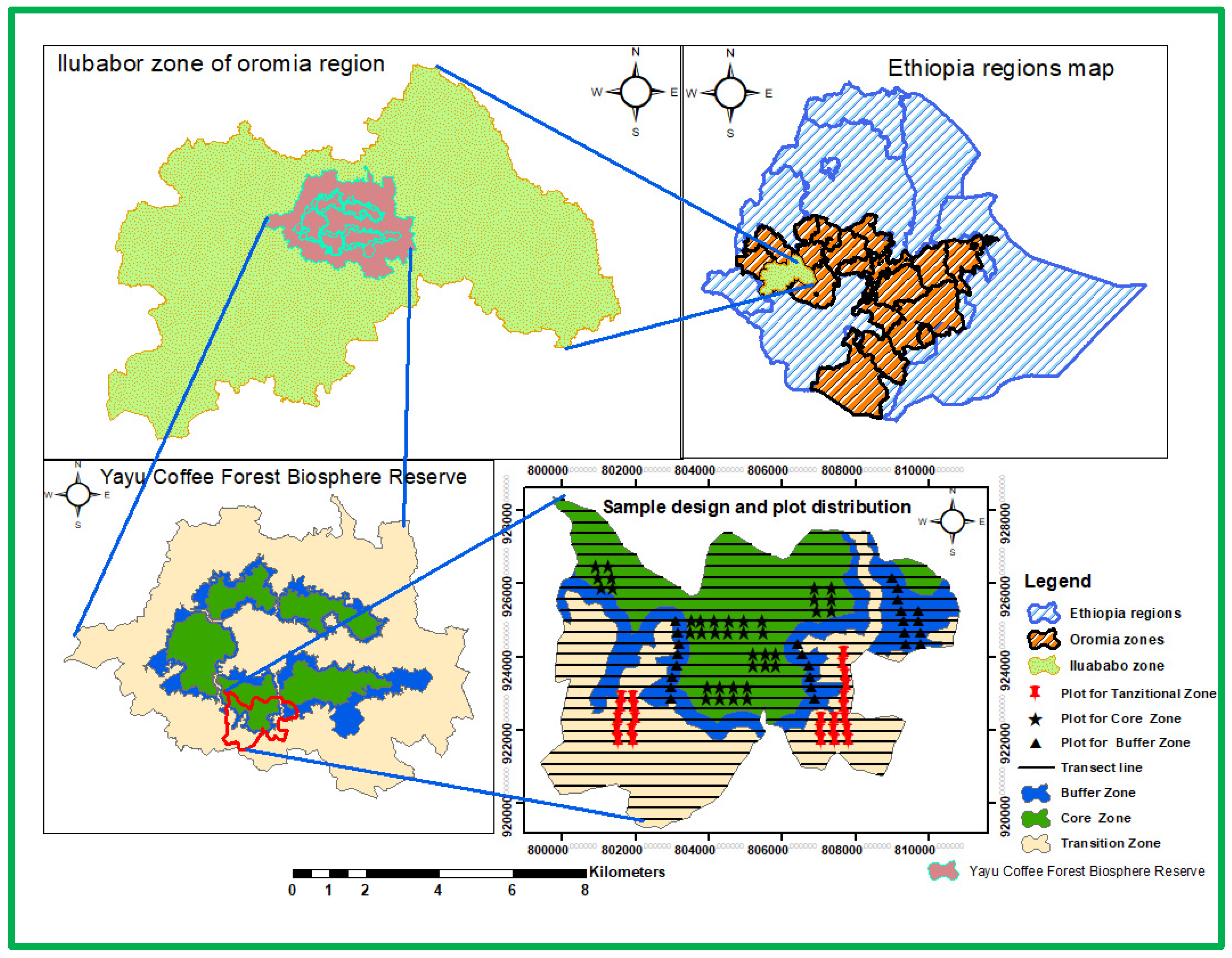
The study was conducted at Yayu Coffee Forest Biosphere Reserve (YCFBR) in Southwest Ethiopia. Multistage sampling techniques were used to select representative samples from the core, buffer, and transitional zones of the reserve following (Jeldu et al., 2023). Yayu and Hurumu districts were purposefully selected because they contain these three zones, unlike other districts. From these districts, Wabo and Gaba kebeles were selected in the final stage using the same criteria as the district selection (Figure 1).
Transect lines were distributed systematically in the core, buffer, and transitional zones. The lines were oriented northwards and established following a random start. Typical plot sizes used in forest inventory assessments are 200 m2, 400 m2, and 500 m2, but any size could possibly be used (UNFCCC, 2015). Following this, we designed a plot size of 20 m × 20 m (400 m2) to collect vegetation data from the core and buffer zones, while a 30 m × 30 m (900 m2) plot size was designed for the transitional zone (Jeldu et al., 2023). A total of 90 plots were distributed, with a 300 m gap between transect lines and sample plots.
Figure 1.
Map of the study area (Yayu Coffee Forest Biosphere Reserve), sample design and plot distributions within the Zones and transect line. Source: Jeldu et al. (2023)
Figure 1.
Map of the study area (Yayu Coffee Forest Biosphere Reserve), sample design and plot distributions within the Zones and transect line. Source: Jeldu et al. (2023)
2.2. Vegetation and Environmental Data Collection
The diameter at breast height (DBH) and height (H) of trees and shrubs with a DBH greater than 2.5 cm were measured. In two perpendicular directions, the DBH (cm) was measured and its average value was recorded. If multi-stemmed individuals had branching below the DBH, each stem was measured separately. Additionally, all woody lianas with a DBH greater than 1 cm at 1.30 m from the rooting point were recorded to assess diversity (Snowdon et al., 2002). For coffee plants, the diameter at stump height (40 cm height) was measured in two perpendicular directions and the average value was taken. In the case of multi-stemmed coffee plants, each stem in a single plant was measured independently, and its equivalent diameter (d 40 cm) was calculated (Snowdon et al., 2002). Plant identification was done at the National Herbarium (ETH), Addis Ababa University, Ethiopia.
Disturbance intensity levels were measured using indicators such as grazing intensity, number of tree stumps, and coffee dominance. Plots were categorized into four disturbance intensity levels: undisturbed, slightly disturbed, moderately disturbed, and disturbed forest (Wekesa et al., 2016). Data on elevation, aspect, slope, and geographical location of each plot was collected. Elevation and aspect were measured using an altimeter, while slope was measured using a Sunto clinometer and classified into five levels as: flat (0%–3%), gentle (3%–8%), moderate (8%–25%), hilly (25%–40%), and steep (> 40%). Aspect was classified into eight sub-directions as: North (N) for direction readings between (0-22.5°) and Northeast (NE) if it reads between (337.5-360°), Northwest (NW) if it reads from 22.5° to 67.5°, East (E) if it reads between 67.5 and 112.5°, Southeast (SE) if it reads between 112.5 and 157.5°, South (S) if the reads are between 157.5 and 202.5°, Southwest (SW) if it is between 202.5 and 292.5°, and Northwest (NW) if it is located between 292.5 and 337.5° (Hu et al., 2015).
2.3. Quantification of Functional Diversity, Functional Dominance and Taxonomic Diversity
For the functional diversity estimation, traits relevant to the life-history strategies of the species and ecosystem function were selected (McGill et al., 2006). These were WD, SLA and Hmax. The SLA and Hmax were extracted from the TRY database (www.try-db.org) (Kattge et al., 2020), whereas the WD was obtained from Ethiopia’s Forest Reference Level Submission to the UNFCCC (EFRL, 2017). Average values for the traits was used for multiple values available for a single species. Similarly, the average genus values was used when trait values were missing for a given species (Mendes et al., 2018; Mensah et al., 2016; Montesinos-López et al., 2018). Likewise, the mean values at the family level was used when genus data for the traits was missing. Additionally, a plot-level average values was used for missing family (Jeldu et al., 2023; Mensah et al., 2016).
For each individual plot, functional richness (FRic), functional evenness (FEve), functional divergence (FDiv) and functional dispersion (FDis) were estimated for functional diversity metrics (Zhang et al., 2021). A plot-based community-weighted mean (CWM) of traits was estimated for functional dominance metrics using the mean of the functional trait weighting by species' relative abundance for each plot (Cavanaugh et al., 2014). Then, CWM was estimated for Hmax, SLA and WD. Standardization was performed for multi-trait functional diversity indices, because, this study’s traits data shows differences in orders of magnitude and scale of measurement. Whereas functional dominance (a single functional trait index) was calculated without standardization for CWM (Teshome et al., 2020).
2.4. Estimation of Above-Ground Carbon
The study used a generic multispecies biomass equation developed by Chave et al. (2014) to calculate AGC (Table 1). For Diospyros abyssinica trees, a species-specific equation developed by Damena and Teshome (2019) was perfomed (Table 1). Total AGC was calculated by combining the above-ground biomass (AGB) of individual trees and coffee plants. The carbon fraction of dry biomass was applied to calculate AGC (IPCC, 2006). Then AGC was scaled up from tree to plot level and extrapolated to megagrams of carbon per hectare (Mg C ha-1).
2.5. Statistical Analysis
The structural equation models (SEM) was used to test the effects of species diversity, functional diversity and functional dominance on AGC. For this, two separate SEM were constructed: (1) a full mediation model (when diversity effects on AGC are fully transmitted through functional diversity and functional dominance metrics) and (2) a partial mediation model (when both direct and indirect diversity effects on AGC are transmitted through species richness, functional diversity and functional dominance). The SEM was started by fitting a full mediation i.e., all paths minus the direct path between species richness and AGC. Using the path modification indices, the model fitting was improved by performing a step-by-step inclusion of additional significant paths leading to partial mediation of SEM. Independent variables were selected using stepwise procedures to avoid over-fitting the model. The comparison between the full mediation and partial mediation models were performed based on the goodness-of-fit statistics (Zürich et al., 2005). For this, only functional diversity and functional dominance measures that significantly influenced AGC were considered for SEM fitting. All SEM were fitted using the lavaan package (Rosseel, 2012) and the overall fit of the models was evaluated using chi-square (P > 0.05 indicates an absence of significance), the comparative fit index (CFI), the goodness-of-fit index (GFI) and the standardized root-mean-square residual (SRMR) (Mensah et al., 2021). All the selected functional diversity indices was calculated by the FDiversity software (Casanoves et al., 2011).
The mixed effects model (MEM) was fitted with functional diversity and functional dominance as predictors of AGC to test the combined effects of functional diversity, and functional dominance metrics. By using the package ‘lme4’ (Bates et al., 2015), the MEM was run, and selection of variables was done by ‘backward selection’ using ‘cAIC4’package. The significant effect of fixed factors was determined using “lmerTest” package (Kuznetsova et al., 2017). Also, regression model was used to test the effects of disturbance, elevation, aspect, and slope (the fixed factors) on AGC (the response variable). As a result, only the elevation showed slightly significant effects on the AGC and considered for further analysis. For those variables that showed no significant effect on the AGC, further analyses, i.e., a multiple linear regressions model, were used to test their effects on the AGC (Bates et al., 2015). Then, we tested for multicollinearity by means of Breusch-Pagan tests, the Durbin-Watson autocorrelation statistic, and the variance inflation factor. All statistical analyses were performed using R statistical software (v4.1.2; R Core Team, 2021).
3. Results
3.1. Predictive Role of Species Diversity for Above-Ground Carbon
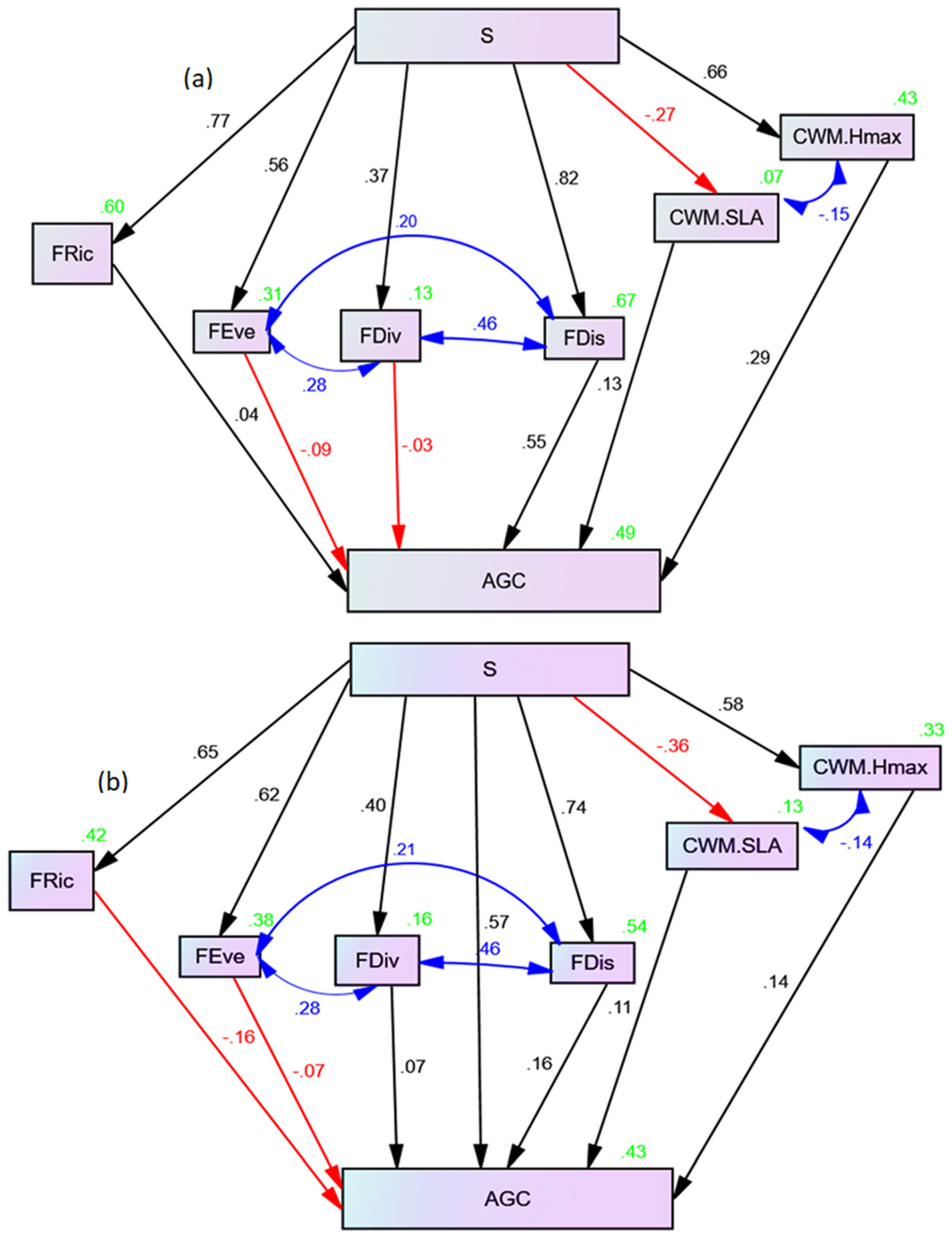
The effects of species diversity richness on AGC, as mediated via functional diversity and dominance, were revealed in Figure 2a,b and Table 2. In the "full mediation model," the chi-square value for the model was 63.789, with 12 degrees of freedom and a 'p' value of 0.001. Additionally, the standardized root-mean-square residual (SRMSR) was 0.223, indicating a satisfactory fit.
In the “full mediation model,” species richness showed a significant positive direct effect on FRic (Table 2). Consequently, the results showed that species richness mediated via FRic had a positive and significant effect on AGC (β = 0.77 ∗ 0.044 = 0.034; Figure 2a; Table 2). There was also significant influences of species richness on FEve, whereas, the species richness, through FEve, had a negative, but non-significant effect on the AGC (β = 0.56 ∗ −0.09 = −0.05). Species richness had a significant and positive influence on AGC via FDiv, as illustrated in the Table 2. Whereas, species richness had a positive significant effect on AGC passing via FDis. Similarly, the functional dominance indices for the community-weighted mean of maximum heights (CWM.Hmax) showed significant path responses to richness and then CWM.Hmax had significant influences on the AGC. Whereas, the species richness effects mediated via CWM.Hmax on AGC was significant and positive effects (Table 2). In contrast, the species richness had a negative and non-significant effect on the community-weighted mean of specific leaf area (CWM.SLA) (Table 2). In addition, the significant residual correlation between CWM.SLA and CWM.Hmax suggests that the mediated effects of species richness on AGC were transmitted by the two factors i.e., CWM.SLA and CWM.Hmax, which affects carbon storages.
The full mediation model implication revealed that the relationship between plant diversity and AGC is not independent; there is the functional diversity and dominance is responsible for the observed relationship between the fixed factors (diversity) and response variable (AGC). Also, the “partial mediation,” model was fitted only by adding a direct path from species richness to AGC to the “full mediation” model as revealed in the figure (Figure 2b). The chi-square value for the “partial mediation” model was 61.885 with 11 degrees of freedom and significant, also indicating good fit (Table 2). The results for the partial mediation model showed that the species richness has a positive with significant effect on AGC (Table 2). This suggests that an increase in species richness leads to an increase in AGC. Similarly, functional diversity indices for the FEve, FRic, and FDis have positive and significant influences on AGC. This implies that, the higher the functional diversity and dominance contribute to higher levels of AGC. From this partial mediation model, we found the significant influence of the added causal path from species richness to AGC, suggesting that an existing true direct influence of species diversity on the AGC (Figure 2b; Table 2). These implied that understanding the mechanisms controlling forest carbon storage is crucial to support “nature-based” solutions for climate change mitigation in the moist Afromontane tropical biosphere reserves.
There were similarities between the two models. Results for both models suggest that the effects of species richness on AGC are mediated via functional diversity and dominance, which confirm both the complementarity and selection effect hypothesis has contributions for the AGC enhancements in the YCFBR. Most of this study tended to compare the relative effects of species richness and other essential functional diversity metrics provided an additional example of exploring diversity effects on AGC, through the assumption that effects of species diversity richness mediated via functional diversity and dominance. Therefore, our findings confirmed this assumption and supported the need to explore beyond species richness to better elucidate the mechanisms that govern species diversity-ecosystem functioning link. Furthermore, these findings also revealed that, significant correlation between species richness and AGC was due to significant correlation between each functional diversity indices and correlations between functional identity indices. For example, findings shows positive correlation between FEve and FDiv, FEve and FDis as well as between FDiv and FDis, which suggests the mediated effects of species richness were mediated and transmitted via FEve, FDiv and FDis. Also, functional dominance showed that, there was negative correlations between CWM.SLA and CWM.Hmax, which implied that the mediated effects of species richness on AGC were transmitted by these two indices of functional dominance.
The positive links between species richness, functional diversity, and functional dominance with AGC highlight the potential role of diverse ecosystems in climate change mitigation. Therefore, management efforts should prioritize the conservation and restoration of biodiversity for maximizing carbon sequestration and contributing to global climate change policies.

Table 2.
The effects of species richness on above-ground carbon stock through functional diversity and functional dominance.
Table 2.
The effects of species richness on above-ground carbon stock through functional diversity and functional dominance.
| Predictor | Response variable | Partial mediation model | Full mediation model | ||||
| Est.std | SE | P value | Est.std | SE | P value | ||
| S | FDiv | 0.40 | 0.004 | *** | 0.37 | 0.004 | *** |
| S | FEve | 0.62 | 0.004 | *** | 0.56 | 0.004 | *** |
| S | FRic | 0.65 | 0.14 | *** | 0.77 | 0.15 | *** |
| S | FDis | 0.74 | 0.01 | *** | 0.82 | 0.01 | *** |
| S | CWM.Hmax | 0.58 | 0.15 | *** | 0.66 | 0.16 | *** |
| S | CWM.SLA | -0.36 | 0.91 | * | -0.27 | 0.94 | 0.084 |
| S | AGC | 0.57 | 26.64 | 0.081 | - | - | - |
| FRic | AGC | -0.16 | 20.07 | 0.466 | 0.04 | 14.80 | 0.814 |
| FEve | AGC | -0.07 | 469.22 | 0.666 | -0.09 | 483.46 | 0.569 |
| FDiv | AGC | 0.07 | 306.27 | 0.598 | -0.03 | 266.74 | 0.815 |
| FDis | AGC | 0.16 | 281.66 | 0.568 | 0.55 | 143.54 | *** |
| CWM.SLA | AGC | 0.11 | 1.76 | 0.379 | 0.13 | 1.76 | 0.293 |
| CWM.Hmax | AGC | 0.14 | 10.62 | 0.34 | 0.29 | 7.79 | 0.014 |
| Fit statistics | |||||||
| Chi-square = 61.885 (p = 0.001) | Chi-square = 63.789 (p = 0.001) | ||||||
| DF = 11 | DF = 12 | ||||||
| SRMSR: 0.231 | SRMSR: 0.223 | ||||||
| GFI: 0.883 | GFI: 0.817 | ||||||
Note: *** - significant at 0.001; ** - significant at 0.01; * - significant at 0.05; S = species richness (species abundance per ha); AGC = Above-ground carbon (Mg C ha-1); FRic - Functional richness; FEve - functional evenness; FDis - functional dispersion; FDiv - functional divergent; CWM.SLA - community-weighted mean of specific leaf area (cm2/g); CWM.Hmax - community-weighted mean of maximum heights (m); SE = standard error; Est. std = standardized estimate; Chisq = Chi-square statistic; DF = degree of freedom (indicating the number of paths omitted from the model); p = the probability of the data given the model (p > 0.05); GFI = goodness fit index; RMSREA = standardized root means square residual.
Figure 2.
Structural equation model for (a) Full mediation model, and (b) Partial mediation model for the effects of species diversity on the above-ground carbon via functional diversity and dominance. Note: S = species richness (species abundance per ha); the figures on the lines are the standardized path coefficients; the green figure numbers represent the coefficient of determination (R2); the blue arrows with blue color numbers represents residual errors to show correlations between each functions (Ɛ); red arrows with numbers denote negative effect paths (-β) and black arrows with numbers denote positive effect paths (β); AGC = Above-ground carbon (Mg C ha-1); FRic - Functional richness; FEve - functional evenness; FDis - functional dispersion; FDiv - functional divergent; CWM.SLA - community-weighted mean of specific leaf area (cm2/g); CWM.Hmax - community-weighted mean of maximum heights (m).
Figure 2.
Structural equation model for (a) Full mediation model, and (b) Partial mediation model for the effects of species diversity on the above-ground carbon via functional diversity and dominance. Note: S = species richness (species abundance per ha); the figures on the lines are the standardized path coefficients; the green figure numbers represent the coefficient of determination (R2); the blue arrows with blue color numbers represents residual errors to show correlations between each functions (Ɛ); red arrows with numbers denote negative effect paths (-β) and black arrows with numbers denote positive effect paths (β); AGC = Above-ground carbon (Mg C ha-1); FRic - Functional richness; FEve - functional evenness; FDis - functional dispersion; FDiv - functional divergent; CWM.SLA - community-weighted mean of specific leaf area (cm2/g); CWM.Hmax - community-weighted mean of maximum heights (m).
3.2. Functional Diversity Effects on Above-Ground Carbon
Our findings showed that the FRic index has a significant positive effect on the AGC, with an estimated coefficient of 14.32 (p < 0.05). This suggests that increasing functional richness can lead to higher levels of AGC in YCFBR. The individual effects of functional diversity metrics on the AGC was tested using linear mixed-effects model and the results showed that all functional diversity indices measured i.e., FRic, FEve, FDiv, and FDis were significant effect (Table 3). Among these three indices i.e., FEve, FDiv, and FDis had a negative effect on AGC. FEve, FDiv and FDis showed high values of probability which is varied from 0.001 to 0.006. Therefore, the effects of functional diversity on the AGC were thus shown by a significant positive effect of FRic and negative effect of FEve, FDiv and FDis. Also, the combined effects of functional diversity metrics, i.e., FRic combined with FEve had significant effects on the AGC enhancements after backward selection for the last model (Table 3). For example, the combined effects of FRic and FEve explained 0.234 (23.4%) of the variance of AGC (Table 3). The reason why the positive effect of FRic on the AGC, could be due to FRic being positively correlated with species richness (Table 2 of SEM results). Enhanced biomass carbon storage can be achieved by functional richness, evenness, divergence, and dispersion in the moist Afromontane forest of the YCFBR. This suggests the need for conservation efforts that maintain and enhance functional diversity to protect and sustain the carbon stock in the biosphere reserve.
Forest managers can use this information for climate change mitigation and biosphere reserve conservation by promoting and maintaining functional diversity in the forest. This can be achieved by preserving a variety of plant species and ensuring a balance in ecological functions. By doing so, they can enhance carbon sequestration and storage, which contributes to climate change mitigation. Policy recommendations can be made based on these findings. For example, if FRic has a significant positive effect on ecosystem services, it might be recommended to prioritize forest management practices that enhance forest resilience. Similarly, if FDiv has a significant positive effect, promoting biodiversity conservation could be encouraged. On the other hand, if FDis has a significant negative effect, it might suggest the need for strategies to reduce disturbances or increase connectivity in forest ecosystems. Also the local benefits of forest ecosystem services can be inferred from these findings. For instance, positive estimates for FEve suggest that increasing forest vegetation can have beneficial effects on local communities. On the other hand, negative estimates for FDis might indicate that disturbances such as deforestation or fragmentation could have negative impacts on local populations.
Table 3.
The effects of functional diversity on above-ground carbon stock of Yayu Coffee Forest Biosphere Reserve, Southwest Ethiopia.
Table 3.
The effects of functional diversity on above-ground carbon stock of Yayu Coffee Forest Biosphere Reserve, Southwest Ethiopia.
| Fixed effects | Random effects (variance) | |||||||
| Estimate | Std.E | df | Pr (>|t|) | Sites | Rsd. | Marg. R2 | AIC | |
| (Intercept) | 238.67 | 55.95 | 6.37 | ** | 31.48 | 0.058 | 1244.3 | |
| FRic | 14.32 | 5.85 | 85.77 | * | ||||
| (Intercept) | -64.30 | 143.40 | 85.80 | 0.650 | 12.98 | 0.080 | 1233.4 | |
| FEve | 619.30 | 221.00 | 86.00 | ** | ||||
| (Intercept) | -153.32 | 164.79 | 85.98 | 0.350 | 22.95 | 0.080 | 1232.8 | |
| FDiv | 633.51 | 219.89 | 84.05 | ** | ||||
| (Intercept) | -39.61 | 81.40 | 68.07 | 0.630 | 21.34 | 0.209 | 1221.1 | |
| FDis | 325.95 | 65.24 | 85.90 | *** | ||||
| (Intercept) | -212.19 | 134.47 | 84.67 | 0.118 | 16.17 | 0.234 | 1207.9 | |
| FEve | 344.9 | 210.17 | 84.85 | 0.104 | ||||
| FDis | 293.99 | 67.52 | 84.96 | *** | ||||
Note: *** - significant at 0.001; ** - significant at 0.01; * - significant at 0.05; Estimate = coefficient estimates; Std.E = standard errors; Rsd. = residual variance; Marg. R2 = marginal R square; AGC = above-ground carbon (Mg C ha-1); p = the probability of the data given the model (p > 0.05); FRic - Functional richness; FEve - functional evenness; FDis - functional dispersion; FDiv - functional divergent; AIC = Akaike Information Criterion.
3.3. Functional Dominance Effects on Above-Ground Carbon Stock
This finding showed that, CWM.Hmax had significant effects on the AGC (Table 4). In contrast, community-weighted mean of wood density (CWM.WD) and CWM.SLA had no significant effect on AGC. Furthermore, the combined effects of CWM.WD with CWM.Hmax on AGC showed significant (Table 4). For instance, the CWM.Hmax has an estimate of 21.11, implying that for each unit increase in CWM.Hmax, the AGC is expected to increase by 21.11 MgCha-1. The CWM.SLA has an estimate of -1.11, suggesting that for each unit increase in CWM.SLA, the AGC is expected to decrease by 1.11 MgCha-1. These results highlight the complexity of factors influencing AGC stock in a biosphere reserve and suggest that multiple fixed factors, beyond functional dominance alone, are likely at play. In terms of policy implications, these results suggest that both fixed and random effects have an impact on the AGC. These findings implied that AGC increased with CWM values that reflect greater dominance of species exhibiting acquisitive traits (i.e. greater maximum plant height, lower wood density and earlier onset of growth) rather than conservative traits.
3.4. The Combined Effects of Functional Diversity and Functional Dominance on Above-Ground Carbon Stock
The result in Table 5 shows that the coefficients of the combined effects of functional diversity and dominance for FDis: CWM.Hmax was significant (p < 0.001), indicating that functional diversity is strongly related to functional dominance. Specifically, for every one-unit increase in "FDis: CWM.Hmax", the AGC is expected to increase by 9.09 Mg Cha-1. The marginal R-squared value of 0.21 suggests that the model explains 21% of the variation in the AGC due to the combined effects of functional diversity indices (Table 5). This was because of tree height being closely related to tree diameter, the significant and positive relationship between CWM.Hmax and AGC reflects the potential importance of characteristics of dominant and large girth trees for ecosystem functioning.
The mass ratio thinking, often known as the selection effect hypothesis, postulates that the characteristics of the majority of dominant species primarily affect how an ecosystem performs at a given period. Also the results implied that, Hmax have the biggest influence on AGC in forests. Because disturbances do not occur in isolation and the effect of multiple drivers on an ecosystem must be considered. Therefore, the combined effects of environmental changes and direct human impacts are likely to reduce greatly overall species richness and trait diversity by filtering out species. As a new understanding, knowing how the relative contributions of these two mechanisms and their interaction affects AGC of the forests may help develop new strategies for managing forests and preserving plant diversity. Although complementarity effects (niche differentiation) and selection effects (mass ration theory) appear to be more significant for biomass productivity, they are not mutually exclusive in the context of forest ecosystem function. A combination of the mass ratio and niche complementarity effects was found to be positively associated with tree diversity; however, the relative significance of these effects varied depending on the trait and range of environmental conditions.
Table 5.
The combined effects of functional diversity and functional dominance on above-ground carbon stock of YCFBR, Southwest Ethiopia.
Table 5.
The combined effects of functional diversity and functional dominance on above-ground carbon stock of YCFBR, Southwest Ethiopia.
| Model | Est. | SE | df | Pr (>|t|) | ||
| Functional diversity + Functional dominance | Fixed effect | (Intercept) | 94.63 | 61.68 | 43.81 | 0.130 |
| FDis: CWM.Hmax | 9.09 | 1.87 | 85.65 | 0.001 | ||
| Random effects (variance) | Sites | |||||
| Rsd. | 12.82 | |||||
| Marg. R2 | 0.21 | |||||
Note: Est. - coefficient of estimates; SE - standard errors; Rsd. - residual variance; df - degree of freedom; Marg. R2 - marginal R square; FDis - functional dispersion; CWM.Hmax - community-weighted mean of maximum heights (m); FDis: CWM.Hmax-interaction of functional dispersion with community-weighted mean of maximum heights.
3.5. Topographic Variables and Site Level Disturbance Effects on Above-Ground Carbon
There were significant effects of elevation and slope on AGC as results for linear regression model revealed, however there was no significant effects due to aspect, except for N (Table 6). For instance, the coefficient estimate of -0.95 suggests that for every unit increase in elevation, there is an expected decrease of 0.95 units in the AGC (assuming all other factors are held constant). The t-value of -5.28 indicates that this effect is statistically significant (Table 6). This implies that higher elevation tends to be associated with lower values of the AGC. However, there is no significant relationship found between aspect and the AGC in this analysis. The coefficient estimate of 40.43 suggests that for every unit increase in species richness, there is an expected increase of 40.43 units in the AGC. Similarly, the t-value indicates that species richness effect is statistically significant. This implies that higher species richness is associated with higher values of the dependent variable. The coefficients for the moderately disturbed and slightly disturbed categories indicate the expected difference in the AGC compared to the disturbed forest sites. However, none of these coefficients is statistically significant, as indicated by the t-values and high p-values. The coefficient for the undisturbed category, however, is statistically significant, suggesting that undisturbed areas tend to have higher values of the AGC compared to the undisturbed forest sites. Both elevation and species richness coefficients are statistically significant (Table 6). This implies that the combined effect of elevation and species richness has a significant impact on the AGC.
The results showed that both elevation and slope had significant effects on AGC, with p-values of 0.002 and 0.047, respectively. However, aspect did not show a significant effect (p = 0.47). Overall, these topographic variables explained 30.3% of the variation in AGC due to environmental factors. Additionally, anthropogenic factors at the site level also had a significant effect on AGC, with 34.1% of the variation explained by disturbance gradients (Table 7). The study found that undisturbed and slightly disturbed sites had higher regression coefficients for AGC than moderately disturbed and disturbed sites. The increase in altitude was found to have a significant effect on AGC, with biomass carbon stocks decreasing at higher altitudes. Also, the integration of biodiversity and considering both traits and diversity helps to improve our understanding of the effects of biotic factors on ecosystems (Table 7).
In general, the results indicate that elevation and species richness have a significant impact and important predictors of the AGC, while aspect and disturbance do not. This information can be valuable for understanding and managing forest ecosystem functions and carbon storage. Therefore, conservation efforts should prioritize on areas with high species richness and accounts elevation gradients to promote biodiversity and support climate mitigation actions.
Table 6.
Results of a linear regression analysis for the individual effects of elevation, slope, aspects and disturbance gradients on above-ground carbon stock using linear regression model.
Table 6.
Results of a linear regression analysis for the individual effects of elevation, slope, aspects and disturbance gradients on above-ground carbon stock using linear regression model.
| Factors | Estimate | Std.Error | tvalue | Pr(>|t|) | BP | DW | VIF | |
| (Intercept) | 1840.59 | 280.42 | 6.56 | *** | 2.51 | 1.60 | 1.83 | |
| Elevation | -0.95 | 0.18 | -5.28 | *** | ||||
| Adjusted R2 (%) | 0.233 | |||||||
| Aspec | (Intercept) | 377.94 | 67.81 | 5.57 | *** | 4.89 | 1.43 | 1.34 |
| N | 291.11 | 115.47 | 2.52 | * | ||||
| NE | -51.11 | 8.12 | -0.59 | 0.554 | ||||
| NW | -9.67 | 162.60 | -0.06 | 0.953 | ||||
| SE | -39.16 | 162.60 | -0.24 | 0.810 | ||||
| SW | -96.60 | 97.22 | -0.99 | 0.323 | ||||
| S | -90.41 | 183.62 | -0.49 | 0.624 | ||||
| Adjusted R2 (%) | 0.072 | |||||||
| S + Dist | (Intercept) | -167.01 | 83.99 | -1.99 | 0.050 | 14.34 | 1.74 | 1.95 |
| Species richness | 40.43 | 7.92 | 5.10 | *** | ||||
| Moderately Dist. | 84.25 | 79.88 | 1.06 | 0.290 | ||||
| Slightly Dist. | 116.99 | 101.82 | 1.15 | 0.254 | ||||
| Undisturbed | 230.43 | 89.49 | 2.58 | * | ||||
| Adjusted R2 (%) | 0.405 | |||||||
| E + S | (Intercept) | 837.55 | 298.93 | 2.80 | ** | 8.57 | 1.79 | 1.92 |
| Elevation (E) | -0.56 | 0.17 | -3.34 | ** | ||||
| Species richness | 39.72 | 7.02 | 5.66 | *** | ||||
| Adjusted R2 (%) | 0.44 |
Note: *** - significant at 0.001; ** - significant at 0.01; * - significant at 0.05; Estimate - estimates of regression coefficients; Std.Error - standard errors; BP - p-values for Breusch-Pagan tests; DW - Durbin-Watson autocorrelation statistic; VIF - Value inflation factor; Adjusted R2 - coefficient of determination; N - north; NE - northeast; NW - northwest; SE - southeast; SW - southwest; S - south; E - elevation; S - species richness (number of individual tree species per hectare); Dist - disturbance.
Table 7.
Results of a multiple non-linear regression analysis with various factors, including elevation, slope, aspect, and disturbance, on above-ground carbon.
Table 7.
Results of a multiple non-linear regression analysis with various factors, including elevation, slope, aspect, and disturbance, on above-ground carbon.
| Factors | Estimate | Std.Error | tvalue | Pr(>|t|) | Ad.R2 | P | |
| (Intercept) | 1260.41 | 389.60 | 3.24 | ** | 0.303 | P < 0.001 | |
| Topography | Elevation | -0.61 | 0.24 | -2.59 | ** | ||
| Slope | -5.55 | 2.75 | -2.02 | * | |||
| c(Aspect) | -11.99 | 16.60 | -0.72 | 0.472 | |||
| Disturbance level | Moderately disturbed | 207.77 | 91.04 | 2.28 | * | 0.341 | P < 0.001 |
| Slightly disturbed | 334.80 | 105.78 | 3.17 | ** | |||
| Undisturbed | 352.24 | 106.68 | 3.30 | *** |
Note: *** - significant at 0.001; ** - significant at 0.01; * - significant at 0.05; Estimate-coefficient of estimates; Ad.R2 - coefficient of determination; Std.Error - standard errors.
4. Discussions
4.1. The Predictive Role of Species Diversity Contributions for the Above-Ground Carbon (AGC)
Full mediation model showed species richness mediated via FRic had a positive and significant effect on AGC as shown under result sections on the Table 2, whereas, species richness effects mediated via CWM.Hmax on AGC was significant and positive effects. This implied that the relationship between plant diversity and AGC is not independent; there is the functional diversity and dominance is responsible for the observed relationship between the fixed factors (diversity) and response variable (AGC). Furthermore, the findings of this study have important implications for biodiversity conservation and tropical biosphere reserve management by considering the contributions of species richness for AGC enhancement, which is consistence with the previous findings (Gautam and Mandal, 2016; Yuan et al., 2018). Therefore, to effectively manage and enhance AGC stock, it is crucial to consider and understand the role of this mediator (species richness).
This study results further supports that both complementarity and selection effects are not exclusively affecting AGC (Mensah et al., 2020). Species diversity richness boosts AGC stock through effects of both functional diversity and functional dominance, partly because these diversity components are based on specific functional traits, which would reflect functional differences among the species (Fei et al., 2017; Parron et al., 2019). This finding can also be due to the fact that increased species richness indirectly accounted for differences among species, in terms of ecological niche and resource use efficiency. Besides to this, various studies in subtropical forests have reported biomass productivity have the same patterns with increasing species diversity richness i.e., AGC increased with increasing species diversity richness (Gautam and Mandal, 2016; Yuan et al., 2018) and tropical forests (Mensah et al., 2017) have also reported increases in biomass productivity with increasing species diversity richness. This partial mediation model suggest that understanding the mechanisms controlling forest carbon storage is crucial to support “nature-based” solutions for climate change mitigation.
It is usually expected that increasing species diversity would increase AGC storage, because higher taxonomic diversity would enhanced higher stem density and forest ecosystem productivity (Mensah et al., 2020). The positive effect of species diversity can also be explained through the benefits of plant-plant interactions such as facilitation, where by some species could enhance soil fertility (by fixing nitrogen) for the productivity of other species (Mensah et al., 2020). This can be suggested that mixed species communities of forests are more productive than homogeneous species diversity stands. Also this may be increasing species richness increases the probability of inclusion of highly productive and dominant species which had consistence with the previous findings (Mengistu et al., 2021; Teshome et al., 2020).
This study findings of moist Afromontane forest promotes both the positive and negative species richness with AGC relationship, however, there was also inverse effect existed. For example, Mengistu et al., (2021) reported the positive effects of species richness on AGC. Furthermore, various studies have reported null or negative relationships between AGC and species diversity in forest ecosystems (Alfaro et al., 2014). These inconsistencies and provocative results pointed out that the effects of species diversity on AGC might be vary with other factors such as forest sites conditions, topographic factors and anthropogenic factors, as well as the dimension of the diversity metrics performed (Wu et al., 2016; Mengistu et al., 2021).
4.2. Functional Diversity and Functional Dominance Effects on Above-Ground Carbon (AGC)
Conservation efforts aimed at preserving and increasing functional diversity are necessary for the protection and maintenance of the carbon stock in the moist Afromontane forest of the YCFBR, which may help improve biomass carbon storage. This is because factors such as functional richness, evenness, divergence, and dispersion have been found to play a significant role in enhancing biomass carbon storage in this ecosystem. This implies that in order to safeguard and sustain the carbon stock in the biosphere reserve, conservation initiatives must preserve and improve functional diversity. These findings are consistent with similar studies conducted before, indicating the importance of functional diversity in promoting AGC in forest ecosystems. For instances, FRic measures the amount of trait space occupied by the community (Clark et al., 2014); thus increase the carbon storage because species with various traits would more efficiently use the resources for growth, which confirmed the niche complementarity effects on the AGC (Mensah et al., 2020). Policy recommendations can be made based on these findings. For example, if FRic has a significant positive effect on ecosystem services, it might be recommended to prioritize forest management practices that enhance forest resilience. Similarly, if FDiv has a significant positive effect, promoting biodiversity conservation could be encouraged. On the other hand, if FDis has a significant negative effect, it might suggest the need for strategies to reduce disturbances in forest ecosystems.
Also, studies in Dindin dry Afromontane forest (Mengistu et al., 2021), and Mistbelt forests (Mensah et al., 2020) found that functional diversity metrics, such as FRic, FEve, and FDis, significantly predict variation in AGC. Positive relationships between FDis and AGC were found in previous studies (Ziter et al., 2013), consistent with our current findings. Similarly, negative effects of FDiv on biomass carbon were observed in subtropical forests in China (Wong et al., 2015; Xin et al., 2018) and temperate mixed forests (Yuan et al., 2018).
Additionally, the study revealed that, AGC increased as the CWM values increased, which indicates a higher prevalence of species with traits that prioritize acquiring resources (such as taller maximum plant height, lower wood density, and earlier growth initiation), rather than traits that prioritize resource conservation. Although contradicting findings was found as selection effects were more important for the AGC stock in tropical forests (Ruiz-Jaen et al., 2011). Furthermore, Wasof et al. (2018) found that the abundance and traits of the most dominant species matter more than taxonomic, functional or phylogenetic diversity in explaining its biomass, supporting the ‘biomass ratio hypotheses’ and ‘diversity hypothesis’ had a negative impact on biomass. These contrary results was obtained due to the methodology and weighting variables varied among the authors. For instance, Yuan et al. (2018) and Phillips et al. (2019) used basal area and biomass as weighting variables for functional dominance metrics, whereas in this study, the species relative abundance was used as weighting variable. From this, relationship between CWM of traits and the ecosystem function of interest could depend on the weighting variable accounted. Consequently, the magnitudes effects of functional diversity and functional dominance contributions for biomass carbon also varied. This is because of selection effects was strongly transmitted through specific Hmax, which supports the importance of forest vertical stratification for species diversity-AGC relationship. The AGC increased with CWM values that reflect greater dominance of species exhibiting acquisitive traits rather than conservative traits which is consistent with Ariel et al. (2021). For instance, Shen et al. (2016) found that, dominance and diversity explained 34% and 32% of variation in carbon storage, whereas the environments explained 26-44% of variation in dominance and diversity. This showed that the proportions of variation in carbon storage explained by dominance and diversity were improved after considering environments (Mensah et al., 2023).
The CWM.WD combined with CWM.Hmax implied that integrating biodiversity into ecosystem ecology and assessing both traits and diversity improves our mechanistic understanding of biotic effects on ecosystems (Hong et al., 2021). The positive effects of biodiversity were mainly driven by interspecific complementarity and that these effects increased over time in both ambient and manipulated environments (Hong et al., 2021). This showed that biodiversity conservation as a key strategy for sustainable ecosystem management in the face of global environmental change.
Functional dominance effects varied depending on the functional trait used (Sainge et al., 2019; Mensah et al., 2020). For example, WD and SLA are associated with population persistence, species tolerance to climate change, and they are used to categorize species as fast or slow growth and resistance to disturbances (Sainge et al., 2019). This might be because of leaf area is crucial for the radiant energy intercepted by the plant and facilitate the exchange of CO2 and H2o between foliage and atmosphere (Mensah et al., 2020). Thus, mass ratio hypothesis are suggesting ecosystem functions at a given time is mainly determined by the traits of most dominant species (Grime, 1998). Also, it was revealed, Hmax have the greatest impact on forest AGC (Mensah et al., 2020). Because disturbances do not occur in isolation and the effect of multiple drivers on an ecosystem must be considered.
As a new message, understanding the relative contributions and interaction effects of these mechanisms on changes in forest AGC can provide insights for developing new strategies in forest management and conserving plant diversity. This hypothesis proposes that selection effects (mass ratio hypothesis), are more influential than complementarity effects (niche differentiation), in terms of biomass productivity. However, it is important to note that these mechanisms are not mutually exclusive and both play a role in forest ecosystem function. Tree diversity was positively associated with a combination of mass ratio and niche complementarity effects, but the relative importance of these effects depended on the trait and range of environmental conditions (Lammerant et al., 2023).
4.3. Topographic Variables and Site Level Disturbance Effects on Above-Ground Carbon
Integrating biodiversity into ecosystem ecology and assessing both traits and diversity improves our mechanistic understanding of biotic effects on ecosystems. Thus, the results suggest that elevation and species richness are important predictors of the AGC, while aspect and disturbance do not have a significant relationship with AGC in this analysis. Also consistently to previous findings, here slope had significant influence, and contributed for AGC variations, evidencing that differences in carbon stocks can result from topological constraints (Mensah et al., 2016; Mensah et al., 2020). Because, the influence of slope can be seen as prior impacts of environment on resources availability and AGC is intrinsically related to wood and biomass production, which in turn affect forest ecosystem services dynamics (Clark et al., 2014; Durán et al., 2015; Mensah et al., 2016). This informed us, tree growth and biomass production can be potentially reduced at higher slope sites. Because, steeper slope speed up nutrients and water run-off and constrain trees, as consequence of nutrient and moisture stress there, which has consistence with Clark et al. (2014). Whereas, flat and gentle slope sites would allow for more water and relevant resources availability for tree growth and biomass productivity, facilitating species would likely respond positively (Mensah et al., 2016; Wu et al., 2014).
Our findings detected significant influences of elevation on biomass carbon stock, which is in consistence with Cavanaugh et al. (2014), but in contrast with (Mensah et al., 2016), who reported a lack of significant relationships between altitude and forest carbon stock. In addition, previous studies found that, forest biomass carbon increased up to a certain a altitudinal points, as suggested up to 3000 meters above sea levels (m.a.s.l.) and then declined (Ensslin et al., 2015; Singh et al., 1994). The significant influence of altitude on forest biomass was due to altitudinal ranges of our study covered greater ranges, which have been considerable enough to detect substantial variation on growth performance (Teshome et al., 2020) and ultimately AGC (in our cases up to 2600m.a.s.l). However, in contrary to our findings, effects of altitude on forest biomass was unseen due to smaller altitudinal range, which is not considerable enough to detected the significant variation on forest carbon stocks (Mensah et al., 2016).
Regarding the effects of disturbance relations with AGC, the findings showed significant and negative relationship with AGC. Consistently, previous findings also showed AGC in highly disturbed plots was lower than plots with medium and low disturbance levels (Yuan et al., 2018), which is in line with the negative relationship between disturbance and AGC found in the current study. These all detected and visible significant effect of topographic variables and anthropogenic factors confirmed that forest ecosystem productivity in general and AGC in particular, are environment-structured, which is similar with the previous findings (Mensah et al., 2016; Wu et al., 2014). This helps to underscore the importance of considering environmental factors and disturbances in understanding and predicting AGC levels in forest ecosystems.
5. Conclusions
This study finding showed that, woody richness effects on AGC were mediated via functional diversity and dominance. Recognizing the pivotal role of niche differences and selection effects disparities unveils the elaborate relationship shaping biodiversity within ecosystems. Niche differences foster coexistence by enabling species to shape out specialized ecological niches and exploit resources uniquely. Conversely, selection effects variations boost competitive exclusion, benefitting the persistence and increase of select species while disregarding others. Understanding the relative contributions of functional diversity, functional dominance, and their combined effects on AGC inform about new strategies for forest management and plant diversity conservation for climate mitigation. The effect of species richness on AGC was significant even after accounting the effects of topographic variables and anthropogenic disturbance. Furthermore, elevation and species richness are important predictors of the AGC. To fully comprehend the extent to which weighting variables for functional diversity and functional dominance contribute to forest ecosystem functions, future studies should focus on assessing their impact on AGC and other climate mitigation measures.
Authors’ Contributions
Ferede Abuye Jeldu, Motuma Tolera and Teshale Woldeamanuel conceived the presented idea and developed the theory and analytical methodologies. Ferede Abuye Jeldu also carried out the data collections and analysis. All author contributed to interpretation and discussions of results. Ferede Abuye Jeldu wrote the first draft and received substantial contributions from other authors. The authors read and approved the final manuscript.
Funding Statement
The authors gratefully acknowledge the financial support provided by the Intergovernmental Panel on Climate Change (IPCC) and African Forest Forum (AFF), which was instrumental in facilitating this research work.
Data Availability
Data are available on request from the authors.
Conflicts of interest
The authors declare no conflict of interest.
References
- Alfaro, R.I.; Fady, B.; Vendramin, G.G.; Dawson, I.K.; Fleming, R.A.; Sáenz-Romero, C.; Lindig-Cisneros, R.A.; Murdock, T.; Vinceti, B.; Navarro, C.M.; Skrøppa, T.; Baldinelli, G.; El-Kassaby, Y.A.; Loo, J. The role of forest genetic resources in responding to biotic and abiotic factors in the context of anthropogenic climate change. Forest Ecology and Management 2014. [Google Scholar] [CrossRef]
- Ayma-Romay, A.I.; Bown, H.E.; Pérez-Harguindeguy, N.; Enrico, L. Trait similarity among dominant highly-competitive species rather than diversity increases. Forest Ecology and Management 2021, 486. [Google Scholar] [CrossRef]
- Bates, D.; Maechler, M.; Bolker, B.; Walker, S. lme4: Linearlinear mixed-effects models using Eigeneigen and S4 R package version R. Package Version 1. Journal of Statistical Software 2015, 1, 1–48. [Google Scholar] [CrossRef]
- Brockerhoff, E.G.; Barbaro, L.; Castagneyrol, B.; Forrester, D.I.; Gardiner, B.; González-Olabarria, J.R.; Lyver, P.O.B.; Meurisse, N.; Oxbrough, A.; Taki, H.; Thompson, I.D.; van der Plas, F.; Jactel, H. Forest biodiversity, ecosystem functioning and the provision of ecosystem services. SpringerPlus 2017, 26, 3005–3035. [Google Scholar] [CrossRef]
- Cadotte, M.W. Functional traits explain ecosystem function through opposing mechanisms. Ecology Letters 2017, 20, 989–996. [Google Scholar] [CrossRef]
- Cadotte, M.W.; Carscadden, K.M.N. Beyond species: functional diversity and the maintenance of ecological processes and services. Journal of Applied Ecology 2011, 48, 1079–1087. [Google Scholar] [CrossRef]
- Carroll, I.T.; Cardinale, B.J.; Nisbet, R.M. Niche and fitness differences relate the maintenance of diversity to ecosystem function. Ecology 2011, 92, 1157–1165. [Google Scholar] [CrossRef] [PubMed]
- Casanoves, F.; Pla, L.; Di Rienzo, J.; Diaz, S. FDiversity: A software package for the integrated analysis of functional diversity, Methods. Methods in Ecology and Evolution 2011, 2, 233–237. [Google Scholar] [CrossRef]
- Cavanaugh, K.C.; Gosnell, J.S.; Davis, S.L.; Ahumada, J.; Boundja, P.; Clark, D.B.; Mugerwa, B.; Jansen, P.A.; O’Brien, T.G.; Rovero, F.; Sheil, D.; Vasquez, R.; Andelman, S. Carbon storage in tropical forests correlates with taxonomic diversity and functional dominance on a global scale. Global Ecology and Biogeography 2014, 23, 563–573. [Google Scholar] [CrossRef]
- Chave, J.; Réjou-Méchain, M.; Búrquez, A.; Chidumayo, E.; Colgan, M.S.; Delitti, W.B.; Duque, A.; Eid, T.; Fearnside, P.M.; Goodman, R.C.; Henry, M. Improved allometric models to estimate the aboveground biomass of tropical trees. Global Change Biology 2014, 20, 3177–3190. [Google Scholar] [CrossRef]
- Chave, J.; Andalo, C.; Brown, S.; Cairns, M.A.; Chambers, J.Q.; Eamus, D.; Fölster, H.; Fromard, F.; Higuchi, N.; Kira, T.; Lescure, J.P.; Nelson, B.W.; Ogawa, H.; Puig, H.; Riéra, B.; Yamakura, T. Tree allometry and improved estimation of carbon stocks and balance in tropical forests. Oecologia 2005. [Google Scholar] [CrossRef] [PubMed]
- Cavanaugh, K.C.; Gosnell, J.S.; Davis, S.L.; Ahumada, J.; Boundja, P.; Clark, D.B.; Mugerwa, B.; Jansen, P.A.; O’Brien, T.G.; Rovero, F.; Sheil, D.; Vasquez, R.; Andelman, S. Carbon storage in tropical forests correlates with taxonomic diversity and functional dominance on a global scale. Global Ecology and Biogeography 2014, 23, 563–573. [Google Scholar] [CrossRef]
- Damena, E.D.; Teshome, S. Allometric equations for aboveground biomass estimation of Diospyros abyssinica (Hiern) F. White tree species. Ecosystem Health and Sustainability 2019. [Google Scholar] [CrossRef]
- Díaz, S.; Fargione, J.; Chapin, F.S., III; Tilman, D. Biodiversity loss threatens human well-being. PLoS Biology 2006, 4, e277. [Google Scholar] [CrossRef] [PubMed]
- Díaz, S.; Purvis, A.; Cornelissen JH, C.; Mace, G.M.; Donoghue, M.J.; Ewers, R.M.; Jordano, P.; Pearse, W.D. Functional traits, the phylogeny of function, and ecosystem service vulnerability. Ecology and Evolution 2013, 3, 2958–2975. [Google Scholar] [CrossRef]
- Díaz, S.C.M. Vive la différence: plant functional diversity matters to ecosystem processes. Trends in Ecology and Evolution 2011, 16, 646–655. [Google Scholar] [CrossRef]
- Ding, H.; Nunes, P.A.L.D. Modeling the links between biodiversity, ecosystem services and human wellbeing in the context of climate change: Results from an econometric analysis of the European forest ecosystems. Ecological Economics 2014. [Google Scholar] [CrossRef]
- Durán, S.M.; Sánchez-Azofeifa, G.A.; Rios, R.S.; Gianoli, E. The relative importance of climate, stand variables and liana abundance for carbon storage in tropical forests. Global Ecology and Biogeography 2015. [Google Scholar] [CrossRef]
- EFRL. Ethiopia’s Forest Reference Level Submission To The UNFCCC. 2017.
- Ensslin, A.; Rutten, G.; Pommer, U.; Zimmermann, R.; Hemp, A.; Fischer, M. Effects of elevation and land use on the biomass of trees, shrubs and herbs at Mount Kilimanjaro. Ecosphere 2015, 6. [Google Scholar] [CrossRef]
- Fargione, J.; Tilman, D.; Dybzinski, R.; Lambers, J.H.R.; Clark, C.; Harpole, W.S.; Knops, J.M.; Reich, P.B.; Loreau, M. From selection to complementarity: shifts in the causes of biodiversity-productivity relationships in a long-term biodiversity experiment. Proceedings of the Royal Society B: Biological Sciences 2007, 274, 871–876. [Google Scholar] [CrossRef]
- Fei, S.; Desprez, J.M.; Potter, K.M.; Jo, I.; Knott, J.A.; Oswalt, C.M. Divergence of species responses to climate change. Science Advances 2017. [Google Scholar] [CrossRef] [PubMed]
- Finegan, B.; Peña-Claros, M.; de Oliveira, A.; Ascarrunz, N.; Bret-Harte, M.S.; Carreño-Rocabado, G.; Casanoves, F.; Díaz, S.; Eguiguren Velepucha, P.; Fernandez, F.; Licona, J.c.; Lorenzo, L.; Salgado, N.B.; Vaz, M.; Poorter, L.; Canham, C. Does functional trait diversity predict above-ground biomass and productivity of tropical forests? Testing three alternative hypotheses. Journal of Ecology 2015, 103, 191–201. [Google Scholar] [CrossRef]
- Gautam, T.P.; Mandal, T.N. Effect of disturbance on biomass, production and carbon dynamics in moist tropical forest of eastern Nepal. Forest Ecosystems 2016. [Google Scholar] [CrossRef]
- Gourlet-Fleury, S.; Rossi, V.; Rejou-Mechain, M.; Freycon, V.; Fayolle, A.; Saint-André, L.; Cornu, G.; Gérard, J.; Sarrailh, J.M.; Flores, O.; Baya, F.; Billand, A.; Fauvet, N.; Gally, M.; Henry, M.; Hubert, D.; Pasquier, A.; Picard, N. Environmental filtering of dense-wooded species controls above-ground biomass stored in African moist forests. Journal of Ecology 2011, 99, 981–990. [Google Scholar] [CrossRef]
- Grime, J.P. Benefits of plant diversity to ecosystems: immediate, filter and founder effects. Journal of Ecology 1998, 86, 901–1076. [Google Scholar] [CrossRef]
- Hong, P.; Schmid, B.; De Laender, F.; Eisenhauer, N.; Zhang, X.; Chen, H.; Craven, D.; De Boeck, H.J.; Hautier, Y.; Petchey, O.L.; Reich, P.B.; Steudel, B.; Striebel, M.; Thakur, M.P.; Wang, S.D. Biodiversity promotes ecosystem functioning despite environmental change. Ecolgical Letters 2021, 25, 555–569. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.; He, Z.; Liu, J.; Zheng, C. Method for measuring the information content of terrain from digital elevation models. Entropy 2015, 17, 7021–7051. [Google Scholar] [CrossRef]
- IPCC. Generic Methodologies Applicable to Multiple Landuse: Volume 4: Agriculture, Forestry and Other Land Use Categories. 2006.
- IPCC. Climate Change 2023: Synthesis Report. Contribution of Working Groups I, II and III to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change. Core Writing Team, Lee, H., Romero, J., Eds.; 2023. [Google Scholar] [CrossRef]
- Jeldu, F.A.; Tolera, M.; Woldeamanuel, T. Functional diversity along disturbance and environmental gradients in Ethiopian moist Afromontane forest. Biosytems Diversity 2023, 31, 460–469. [Google Scholar] [CrossRef]
- Kattge, J.; Bönisch, G.; Díaz, S.; Lavorel, S.; Prentice, I.C.; Leadley, P.; Tautenhahn, S.; Werner, G.D.A.; Aakala, T.; Abedi, M.; et al. TRY plant trait database enhanced coverage and open access. Global Change Biology 2020, 26, 119–188. [Google Scholar] [CrossRef] [PubMed]
- Kong, L.; Xiong, K.; Zhang, S.; Zhang, Y.; Deng, X. Review on Driving Factors of Ecosystem Services: Its Enlightenment for the Improvement of Forest Ecosystem Functions in Karst Desertification Control. Forests 2023, 14, 582. [Google Scholar] [CrossRef]
- Kuznetsova, A.; Brockhoff, P.B.; Christensen, R.H.B. lmerTest Package: Tests in linear mixed effects models. Journal of Statistical Software 2017, 82, 1–26. [Google Scholar] [CrossRef]
- Lammerant, R.; Rita, A.; Borghetti, M.; Muscarella, R. Water-limited environments affect the association between functional diversity and forest productivity. Ecology and Evolution 2023, 13. [Google Scholar] [CrossRef] [PubMed]
- Lasky, J.R.; Uriarte, M.; Boukili, V.K.; Erickson, D.L.; John Kress, W.C.R. The relationship between tree biodiversity and biomass dynamics changes with tropical forest succession. Ecolgical Letters 2014, 17, 1158–1167. [Google Scholar] [CrossRef]
- Laughlin, D.C. Applying trait-based models to achieve functional targets for theory-driven ecological restoration. Ecology Letters 2014, 17, 771–784. [Google Scholar] [CrossRef] [PubMed]
- Ma, W.; He, J.; Yang, Y.; Wang, X.; Liang, C.; Anwar, M.; Zeng, H.; Fang, J.; Schmid, B. Environmental factors covary with plant diversity–productivity relationships among Chinese grassland sites. Global Ecology and Biogeography 2010, 19, 233–243. [Google Scholar] [CrossRef]
- Sullivan, M.J.; Talbot, J.; Lewis, S.L.; Phillips, O.L.; Qie, L.; Begne, S.K.; Chave, J.; Cuni-Sanchez, A.; Hubau, W.; Lopez-Gonzalez, G.; Miles, L. Diversity and carbon storage across the tropical forest biome. Scientific Reports 2017, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Mason, N.W.H.; Mouillot, D.; Lee, W.G.; Wilson, J.B. Functional richness, functional evenness and functional divergence: The primary components of functional diversity. Oikos 2005, 111, 112–118. [Google Scholar] [CrossRef]
- McGill, B.J.; Enquist, B.J.; Weiher, E.W.M. Rebuilding community ecology from functional traits. Trends in Ecology and Evolution 2006, 21, 178–185. [Google Scholar] [CrossRef]
- Mendes, F.K.; Fuentes-Gonzalez, J.A.; Schraiber, J.G.; Hahn, M.W. A multispecies coalescent model for quantitative traits. Elife 2018, 7, e36482. [Google Scholar] [CrossRef]
- Wondimu, M.T.; Nigussie, Z.A.; Yusuf, M.M. Tree species diversity predicts aboveground carbon storage through functional diversity and functional dominance in the dry evergreen Afromontane forest of Hararghe highland, Southeast Ethiopia. Ecological Processes 2021, 10, 1–15. [Google Scholar] [CrossRef]
- Mensah, S.; Dimobe, K.; Noulèkoun, F.; van der Plas, F.; Seifert, T. Phylogenetic diversity and community wide-trait means offer different insights into mechanisms regulating aboveground carbon storage. Science of Total Environmnet 2024, 907. [Google Scholar] [CrossRef]
- Mensah, S.; du Toit, B.; Seifert, T. Diversity–biomass relationship across forest layers: implications for niche complementarity and selection effects. Oecologia 2018, 187, 783–795. [Google Scholar] [CrossRef] [PubMed]
- Mensah, S.; Van der Plas, F.; Noulèkoun, F. Do functional identity and divergence promote aboveground carbon differently in tropical semi-arid forests and savannas? Ecosphere 2021, 12, 1–13. [Google Scholar] [CrossRef]
- Mensah, S.; Noulèkoun, F.; Dimobe, K.; Seifert, T.; Glèlè Kakaï, R. Climate and soil effects on tree species diversity and aboveground carbon patterns in semi-arid tree savannas. Scientific Reports 2023, 13. [Google Scholar] [CrossRef] [PubMed]
- Mensah, S.; Veldtman, R.; Assogbadjo, A.E.; Glèlè Kakaï, R.; Seifert, T. Tree species diversity promotes aboveground carbon storage through functional diversity and functional dominance. Ecology and Evolution 2016, 6, 7546–7557. [Google Scholar] [CrossRef]
- Mensah, S.; Salako, K.V.; Assogbadjo, A.; Kakaï, R.G.; Sinsin, B.; Seifert, T. Functional trait diversity is a stronger predictor of multifunctionality than dominance: Evidence from an Afromontane forest in South Africa. Ecological Indicators 2020, 115, 1–11. [Google Scholar] [CrossRef]
- Mensah, S.; Veldtman, R.; Assogbadjo, A.E.; Ham, C.; Kakaï, R.G.; Seifert, T. Ecosystem service importance and use vary with socio-environmental factors: A study from household-surveys in local communities of South Africa. Ecosystem Services 2017, 23, 1–8. [Google Scholar] [CrossRef]
- Negash, M.; Starr, M.; Kanninen, M.; Berhe, L. Erratum to: Allometric equations for estimating aboveground biomass of Coffea arabica L. grown in the Rift Valley escarpment of Ethiopia. Agroforestry Systems 2013, 87, 953–966. [Google Scholar] [CrossRef]
- Michalet, R.; Delerue, F.; Liancourt, P.; Pugnaire, F.I. Are complementarity effects of species richness on productivity the strongest in species-rich communities? Journal of Ecology 2021, 109, 2038–2046. [Google Scholar] [CrossRef]
- Montesinos-López, O.A.; Montesinos-López, A.; Crossa, J.; Kismiantini Ramírez-Alcaraz, J.M.; Singh, R.; Mondal, S.; Juliana, P. A singular value decomposition Bayesian multiple-trait and multiple-environment genomic model. Heredity 2018, 122, 381–401. [Google Scholar] [CrossRef]
- Noulèkoun, F.; Mensah, S.; Dimobe, K.; Birhane, E.; Kifle, E.T.; Naab, J.B.; Son, Y.; Khamzina, A. Both the selection and complementarity effects underpin the effect of structural diversity on aboveground biomass in tropical forests. Global Ecology and Biogeography 2024, 33, 325–340. [Google Scholar] [CrossRef]
- Parron, L.M.; Fidalgo, E.C.C.; Luz, A.P.; Campanha, M.M.; Turetta AP, D.; Pedreira, B.C.C.G.; Prado, R.B. Research on ecosystem services in Brazil: A systematic review. Revista Ambiente e Agua 2019, 14. [Google Scholar] [CrossRef]
- Phillips, O.L.; Sullivan, M.J.; Baker, T.R.; Monteagudo Mendoza, A.; Vargas, P.N.; Vásquez, R. Species Matter: Wood Density Influences Tropical Forest Biomass at Multiple Scales. Surveys in geophysics 2019, 40, 913–935. [Google Scholar] [CrossRef]
- Poorter, L.; van der Sande, M.T.; Thompson, J.; Arets, E.J.; Alarcón, A.; Álvarez-Sánchez, J.; Ascarrunz, N.; Balvanera, P.; Barajas-Guzmán, G.; Boit, A.; Bongers, F. Diversity enhances carbon storage in tropical forests. Global Ecology and Biogeography 2015, 24, 1314–1328. [Google Scholar] [CrossRef]
- Reich, P.B.; Tilman, D.; Naeem, S.; Ellsworth, D.S.; Knops, J.; Craine, J.; Wedin, D.; Trost, J. Species and functional group diversity independently influence biomass accumulation and its response to CO 2 and N. Proceedings of the National Academy of Sciences 2004, 101, 10101–10106. [Google Scholar] [CrossRef]
- Rodríguez-Hernández, D.I.; Deane, D.C.; Wang, W.; Chen, Y.; Li, B.; Luo, W.; Chu, C. Direct effects of selection on aboveground biomass contrast with indirect structure-mediated effects of complementarity in a subtropical forest. Oecologia 2021, 196, 249–261. [Google Scholar] [CrossRef] [PubMed]
- Rosseel, Y. Lavaan: An R package for structural equation modeling and more, Version 0.5–12 (BETA). Journal of Statistical Software 2012, 48, 1–36. [Google Scholar] [CrossRef]
- Ruiz-Jaen, M.C.; Potvin, C. Can we predict carbon stocks in tropical ecosystems from tree diversity? Comparing species and functional diversity in a plantation and a natural forest. New Phytologist 2011, 189, 978–987. [Google Scholar] [CrossRef]
- Sainge, M.N.; Lyonga, N.M.; Mbatchou, G.; Kenfack, D.; Nchu, F.; Peterson, A.T. Vegetation, floristic composition and structure of a tropical montane forest in Cameroon. Bothalia-African Biodiversity & Conservation 2019, 49, 1–12. [Google Scholar]
- Sasaki, T.; Lauenroth, W.K. Dominant species, rather than diversity, regulates temporal stability of plant communities. Oecologia 2011, 166, 761–768. [Google Scholar] [CrossRef]
- Segura, M.; Kanninen, M. Allometric models for tree volume and total aboveground biomass in a tropical humid forest in Costa Rica 1. Biotropica. The Journal of Biology and Conservation 2005, 37, 2–8. [Google Scholar]
- Shen, Y.; Yu, S.; Lian, J.; Shen, H.; Cao, H.; Lu, H.; Ye, W. Tree aboveground carbon storage correlates with environmental gradients and functional diversity in a tropical forest. Scientific Reports 2016, 6, 25304. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.P.; Adhikari, B.S.; Zobel, D.B. Biomass, Productivity, Leaf Longevity, and Forest Structure in the Central Himalaya. Ecological Monographs 1994, 64, 401–421. [Google Scholar] [CrossRef]
- Snowdon, P.; Raison, J.; Keith, H.; Montagu, K.; Bi, H.; Ritson, P.; Grierson, P.; Adams, M.; Burrows, W.; Eamus, D. Protocol for sampling tree and stand biomass. National Carbon Accounting System, Technical Report No. 31. Australian Greenhouse Office: Canberra, 2002. [Google Scholar]
- Sutherland, W.J.; Adams, W.M.; Aronson, R.B.; Aveling, R.; Blackburn, T.M.; Broad, S.; Ceballos, G.; Côté, I.M.; Cowling, R.M.; DA Fonseca, G.A.B.; et al. One hundred questions of importance to the conservation of global biological diversity. Conservation biology 2009, 23, 557–567. [Google Scholar] [CrossRef]
- R: A language and environment for statistical computing; R Foundation for Statistical Computing: Vienna, Austria, 2021.
- Teshome, M.; Asfaw, Z.; Mohammed, M. Pattern of Functional Diversity along the Elevation Gradient in the Dry Evergreen Afromontane Forest of Hararghe Highland, Southeast Ethiopia. Biosytems Diversity 2020, 28, 257–264. [Google Scholar] [CrossRef]
- Violle, C.; Enquist, B.J.; McGill, B.J.; Jiang, L.; Albert, C.H.; Hulshof, C.; Jung, V.; Messier, J. The return of the variance: intraspecific variability in community ecology. Trends in Ecology & Evolution 2012, 27, 244–252. [Google Scholar] [CrossRef]
- Wasof, S.; Lenoir, J.; Hattab, T.; Jamoneau, A.; Gallet-Moron, E.; Ampoorter, E.; Saguez, R.; Bennsadek, L.; Bertrand, R.; Valdès, A.; Verheyen, K. Dominance of individual plant species is more importantthan diversity in explaining plant biomass in the forestunderstorey. Journal of Vegation Science 2018, 29, 521–531. [Google Scholar] [CrossRef]
- Weisser, W.W.; Roscher, C.; Meyer, S.T.; Ebeling, A.; Luo, G.; Allan, E.; Beßler, H.; Barnard, R.L.; Buchmann, N.; Buscot, F.; Engels, C. Biodiversity effects on ecosystem functioning in a 15-year grassland experiment: Patterns, mechanisms, and open questions. Basic and Applied Ecology 2017, 23, 1–73. [Google Scholar] [CrossRef]
- Wekesa, C.; Leley, N.; Maranga, E.; Kirui, B.; Muturi, G.; Mbuvi, M.; Chikamai, B. Effects of Forest Disturbance on Vegetation Structure and Above-Ground Carbon in Three Isolated Forest Patches of Taita Hills. Open Journal of Forestry 2016, 6, 142–161. [Google Scholar] [CrossRef]
- Wong, C.P.; Jiang, B.; Kinzig, A.P.; Lee, K.N.; Ouyang, Z. Linking ecosystem characteristics to final ecosystem services for public policy. Ecology Letters 2015, 18. [Google Scholar] [CrossRef]
- Wu, X.; Wang, X.; Tang, Z.; Shen, Z.; Zheng, C.; Xia, X.; Fang, J. The relationship between species richness and biomass changes from boreal to subtropical forests in China. Ecography 2014, 38, 602–613. [Google Scholar] [CrossRef]
- Wu, X.; Lu, Y.; Zhou, S.; Chen, L.; Xu, B. Impact of climate change on human infectious diseases: Empirical evidence and human adaptation. Environment International 2016, 86, 14–23. [Google Scholar] [CrossRef]
- Xin, Z.; Li, C.; Liu, H.; Shang, H.; Ye, L.; Li, Y.; Zhang, C. Evaluation of temporal and spatial ecosystem services in Dalian, China: Implications for urban planning. Sustainability 2018, 10. [Google Scholar] [CrossRef]
- Xu, Y.; Chen, H.Y.H.; Xiao, Z.; Wan, D.; Liu, F.; Guo, Y.; Qiao, X.; Jiang, M. Species Richness Promotes Productivity through Tree Crown Spatial Complementarity in a Species-Rich Natural Forest. Forests 2022, 13, 106. [Google Scholar] [CrossRef]
- Yi, S.; Wu, P.; Peng, X.; Bai, F.; Gao, Y.; Zhang, W.; Du, N.; Guo, W. Functional identity enhances aboveground productivity of a coastal saline meadow mediated by Tamarix chinensis in Laizhou Bay, China. Scientific Reports 2020, 10, 5826. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Z.; Wang, S.; Ali, A.; Gazol, A.; Ruiz-Benito, P.; Wang, X.; Lin, F.; Ye, J.; Hao, Z.; Loreau, M. Aboveground carbon storage is driven by functional trait composition and stand structural attributes rather than biodiversity in temperate mixed forests recovering from disturbances. Annals of Forest Science 2018, 75, 1–13. [Google Scholar] [CrossRef]
- Zanne, A.E.; Lopez-Gonzalez, G.; Coomes David, A.; Ilic, J.; Jansen, S.; Lewis Simon, L.S.; Miller, R.B.; Swenson, N.G.; Wiemann, M.C.; Chave, J. Data from: Towards a worldwide wood economics spectrum [Dataset]. Dryad. 2009. [Google Scholar] [CrossRef]
- Zhang, J.-T.; Bai, X.; Shao, D. Functional Diversity And Its Attribute Factors In The Yunmeng Mountain National Forest Park In Beijing, China. CERNE 2017, 23, 75–84. [Google Scholar] [CrossRef]
- Zihao, Z.; Jihua, H.; Nianpeng, H. Predictability of Functional Diversity Depends on the Number of Traits. Journal of Resources and Ecology 2021, 12, 332–345. [Google Scholar] [CrossRef]
- Ziter, C.; MacDougall, A. Nutrients and defoliation increase soil carbon inputs in grassland. Ecology 2013, 94, 106–116. [Google Scholar] [CrossRef]
- Zürich, F.; Zingg Æ, A.; Biber Æ, P.; Zimmerman, D.L.; Zimble D a Evans, D.L.; Carlson, G.C.; Parker, R.C.; Grado, S.C.; Gerard, P.D.; Zianis, D.; Muukkonen, P.; Mäkipää, R.; Zhao, Y.; Krzic, M.; Bulmer, C.E.; Schmidt, M.G.; Simard, S.W.; Zhao, D.; Leuven, K.U. A Structural Equation Model Analysis of Postfire Plant Diversity. Forest Ecology and Management 2005, 10. [Google Scholar]
Table 1.
Equations for analysis of above-ground biomass of YCFBR, Southwest Ethiopia.
| No. | Equations | References |
|---|---|---|
| 1 | (Chave et al., 20014) | |
| 2 | (Mesele et al., 2013) | |
| 3 | Diospyros abyssinica | (Damena and Teshome, 2019) |
Table 4.
The effects of functional dominance on above-ground carbon stock of Yayu Coffee Forest Biosphere Reserve, Southwest Ethiopia.
Table 4.
The effects of functional dominance on above-ground carbon stock of Yayu Coffee Forest Biosphere Reserve, Southwest Ethiopia.
| Fixed effects | Random effects (variance) | |||||||
| Estimate | Std.E | df | Pr (>|t|) | Site | Rsd. | Marg. R2 | AIC | |
| (Intercept) | 139.27 | 133.25 | 34.49 | 0.300 | 118.20 | 0.006 | 1240.9 | |
| CWM.WD | 117.92 | 134.21 | 85.93 | 0.380 | ||||
| (Intercept) | 411.65 | 125.39 | 21.40 | 0.003 | 39.12 | 0.013 | 1250.3 | |
| CWM.SLA | -1.11 | 0.98 | 85.03 | 0.260 | ||||
| (Intercept) | -120.31 | 117.42 | 85.74 | 0.310 | 4.21 | 0.165 | 1233.2 | |
| CWM.Hmax | 21.11 | 5.09 | 85.02 | *** | ||||
| (Intercept) | -49.26 | 110.85 | 55.55 | 0.660 | 88.43 | 0.077 | 1238.3 | |
| CWM.WD:CWM.Hmax | 17.47 | 5.47 | 82.52 | ** | ||||
Note: *** - significant at 0.001; ** - significant at 0.01; * - significant at 0.05; Estimate = coefficient estimates, Std.E = s tandard errors, Rsd. = residual variance, Marg. R2 = marginal R square, p = the probability of the data given the model (p > 0.05); AIC = Akaike Information Criterion; CWM.SLA - community-weighted mean of specific leaf area (cm2/g); CWM.WD - community-weighted mean of wood density (g/cm3); CWM.Hmax - community-weighted mean of maximum heights (m); CWM.WD: CWM.Hmax-interaction of community-weighted mean of wood density with community-weighted mean of maximum heights.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



