Submitted:
29 July 2024
Posted:
30 July 2024
You are already at the latest version
Abstract
Demographic studies have revealed a strong association between exposure to high ambient temperatures during pregnancy and increased risks of preterm birth (PTB). The mechanism underlying this association is unclear, but it is plausible that altered placental function may contribute. In this study, we conducted differential gene expression analysis, gene set enrichment analysis (GSEA), and gene ontology (GO) analysis on bulk RNA-seq data from human placentas delivered at term and preterm during the warmer months compared to placentas delivered at term and preterm during the colder months in the UK. We detected 48 differentially expressed genes in preterm placentas delivered during the warmer months compared to preterm placentas delivered during the colder months, the majority of which were inflammatory cytokines and chemokines, including SERPINA1, IL1B, CCL3, CCL3L3, CCL4, CCL4L2, CCL20, and CXCL8. The GSEA positively enriched 17 signalling pathways, including the NF-κB, IL17, toll-like receptor, and chemokine signalling pathways, in warm-exposed preterm placentas, which were not observed in warm-exposed term placentas. The GO analysis revealed several biological processes, including neutrophil, granulocyte, monocyte, and lymphocyte chemotaxis, as well as inflammatory and humoral immune responses in warm-exposed preterm placentas, but not in warm-exposed term placentas. We conclude that maternal exposure to warm environmental temperatures during pregnancy alters the placental transcriptome towards inflammation and immune regulation, potentially leading to PTB.
Keywords:
Thermal effects on placental transcriptome
; Climate change and preterm birth
; Placental inflammation in heat exposure
1. Introduction
Increased global environmental temperature owing to climate change poses significant health risks for the vulnerable population including pregnant women with limited physiological adaptability to extreme temperatures [1,2]. The heat-associated detrimental effects likely affect women in low- and middle-income countries (LMICs) who often continue their agricultural works throughout the pregnancies [3,4]. Epidemiological studies provide strong evidence of increased risks of pregnancy complications such as preterm birth (PTB) (<37 gestational week) from exposure to high ambient temperature and heatwaves during pregnancy [5,6,7,8,9,10].
Approximately 15 million babies are born preterm globally every year, and over a million of them die due to prematurity-associated complications [11]. The rate of PTB is significantly higher in women in LMICs compared to those in high-income countries (HICs). Over 81% of all preterm births occur in Asia and Sub-Saharan Africa [5,11]. Nearly 70% of preterm births occur spontaneously without any detectable causes. Spontaneous preterm birth has strong links with exposure to extreme temperatures during pregnancy, though the underpinning mechanisms remain elusive [9,12].
A systematic review of 70 studies from 27 countries revealed a strong link between high environmental temperature and adverse pregnancies including PTB. The study estimated an overall 5% increased risk of PTB per 1 °C increase in temperature and 16% increase during heatwaves [5]. In longer exposure to high heat for over a trimester or more, the PTB risk increased to 14%, whereas exposure for less than 4 weeks was associated with a rate of 7%, suggesting a dose-response association of high temperature with PTB. A cohort study on Chinese singleton pregnancies showed a significant association of both heat and cold exposure with increased risk of PTB [9]. The 2nd and 3rd trimesters of pregnancy seem most vulnerable to the effect of ambient heat exposure causing PTB and low birth weight [8].
A moderate level of heat exposure (>24.8 °C) for a shorter duration can significantly increase maternal body temperature resulting in both maternal and fetal stress potentially driven by reduction of placental blood flow [13]. However, how continuous heat exposure for longer periods during gestation affects placental function, and how it causes PTB is unclear.
The placenta plays a pivotal role in maintaining the gestation in health and is critically involved in fetal growth. A study showed that maternal exposure to 29 °C during late pregnancy reduced the average size of the placenta compared to exposure to 20 °C ambient temperature [14]. Animal model studies also showed that excessive heat exposure during pregnancy significantly reduces placental weight and diminishes placental transport [15,16]. Together, these indicate that exposure to high temperatures during pregnancy impairs placental development and function.
However, there is a paucity of data on placental molecular changes due to heat exposure during pregnancy, some of which may be linked to preterm birth. A clear understanding of genetic changes in the placenta due to exposure to high environmental temperatures during pregnancy is essential for developing effective interventions. These interventions could include environmental and behavioural modifications, targeted therapeutics, and biomarker prediction tests for preterm birth to address climate change-associated neonatal deaths, particularly in the LMICs.
To address this need, we conducted a differential gene expression analysis and comparison of RNA-seq data from human placentas (chorionic villi) delivered at term and preterm during the warmer time of the year and RNA-seq data from placentas delivered at term and preterm during the colder time of the year. Our differential gene expression analysis did not identify any transcriptomic differences in term placentas between the warm and cold groups. However, in preterm placentas, 48 genes were differentially expressed in the warm group compared to the cold group, and these genes were predominantly immune and inflammatory in nature. Our gene set enrichment analysis (GSEA) revealed the positive enrichment of signalling pathways predominantly associated with inflammation, immune regulation, and graft rejection. To our knowledge, this is the first report on the human placenta showing environmental temperature effects on the placental transcriptome linked to preterm birth. Nevertheless, further studies with larger sample sizes from diverse geographic regions are required to better understand this phenomenon.
2. Results
2.1. Study Subjects and Temperature Exposures
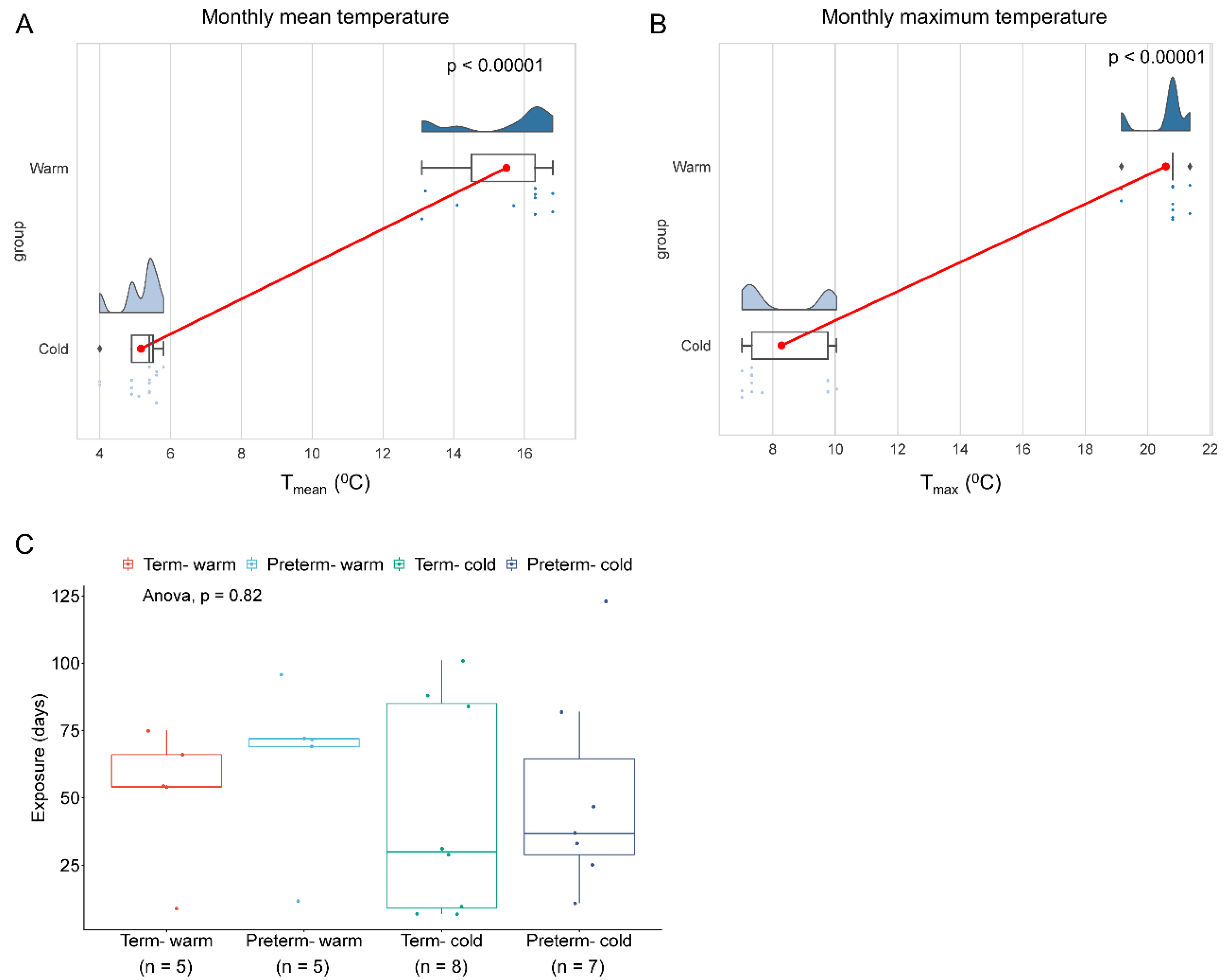
To investigate how warm environmental temperatures influence placental molecular functions, we analysed bulk RNA-seq data from our previously published research on human placentas (chorionic villi) from spontaneous term and preterm births [17]. A total of 25 raw FASTQ RNA-seq files out of 40 sequence datasets were downloaded from the NCBI Gene Expression Omnibus depository using GEO accession number GSE211927 (Supplemental Table S1). Datasets were divided into four groups: (i) Preterm placentas delivered during the summertime in the South Yorkshire region of the UK with a monthly mean weather temperature of 14.92 °C (Tmean = 14.92 °C) and a mean maximum temperature of 20.47 °C (Tmax = 20.47 °C) (n = 5) (Preterm-warm group), (ii) Preterm placentas delivered during the wintertime with a Tmean = 5.14 °C and a Tmax = 8.37 °C (n = 7) (Preterm-cold group), (iii) Term placentas delivered during the summertime with a Tmean = 16.06 °C and a Tmax = 20.69 °C (n = 5) (Term-warm group), and (iv) Term placentas delivered during the wintertime with a Tmean = 5.18 °C and a Tmax = 8.20 °C (n = 8) (Term-cold group) (Table 1, Figure 1, Supplemental Table S1). The pregnant women were exposed to warm or cold temperatures for an average of 8.3 and 6.8 weeks, respectively, preceding delivery (Table 1, Figure 1C).
We compared gene expression and signalling pathways between Preterm-warm vs. Preterm-cold, and Term-warm vs. Term-cold placentas to evaluate the molecular changes due to warm environmental temperature exposure on placentas in preterm and term births, respectively. This strategy eliminated the potential gestational age (GA)-associated confounding effects on the placental transcriptome. Our subjects were mostly of Caucasian background with predominantly male-sex placentas from singleton pregnancies in each group. There were no significant differences in maternal ages, maternal BMI, gestational ages, placental weights, baby birth weights, and exposure durations between the warm and cold groups (Table 1, Figure 1C). None of the pregnant women had diabetes mellitus, hypertension, or signs of genitourinary infection at labour [17].

Table 1.
The study participants’ criteria whose RNA-seq data were analysed (n = 25). Data are presented as mean ± standard deviation. p values were determined by either Mann-Whitney U test or # Fisher’s Exact Test. ns = not significant.
Table 1.
The study participants’ criteria whose RNA-seq data were analysed (n = 25). Data are presented as mean ± standard deviation. p values were determined by either Mann-Whitney U test or # Fisher’s Exact Test. ns = not significant.
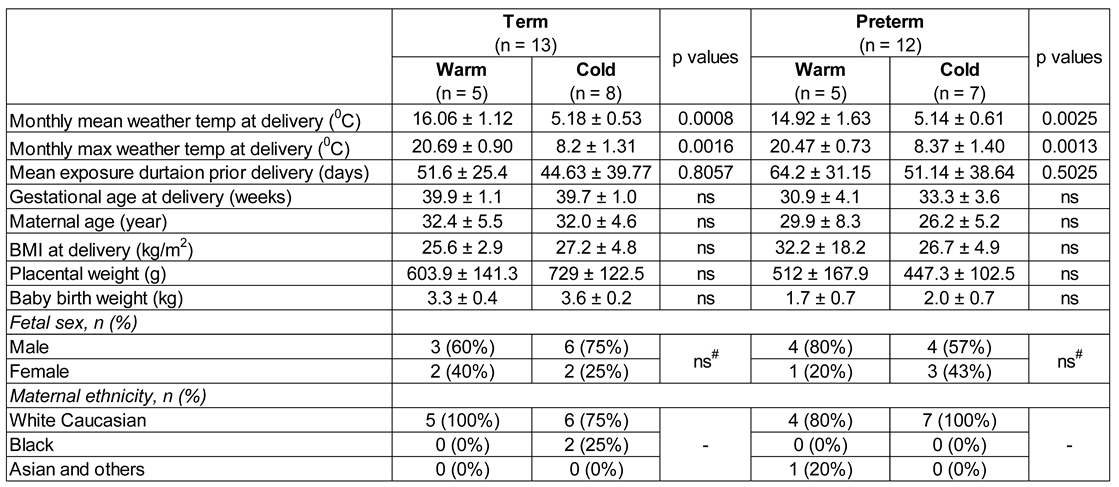 |
Figure 1.
Environmental temperature exposure during pregnancy. The rain-cloud plots showing monthly mean temperature (A) and average maximum temperature (B) when the women delivered. The clouds show the data kernel density, red dots with connectors show the mean, box plots show the median with IQR, and each dot represents an individual subject. n = 10 warm and n = 15 cold group (A, B). p values were calculated by the Mann-Whitney U test. (C) Box blots with jitter showing duration of exposure to warm or cold weather temperatures to four groups of pregnant women prior delivery. Data are presented as median with IQR, and each dot represents an individual subject. p = 0.82, determined by One-way ANOVA.
Figure 1.
Environmental temperature exposure during pregnancy. The rain-cloud plots showing monthly mean temperature (A) and average maximum temperature (B) when the women delivered. The clouds show the data kernel density, red dots with connectors show the mean, box plots show the median with IQR, and each dot represents an individual subject. n = 10 warm and n = 15 cold group (A, B). p values were calculated by the Mann-Whitney U test. (C) Box blots with jitter showing duration of exposure to warm or cold weather temperatures to four groups of pregnant women prior delivery. Data are presented as median with IQR, and each dot represents an individual subject. p = 0.82, determined by One-way ANOVA.
2.2. Warm Environmental Temperature Alters Placental Transcriptome in Preterm Birth
To evaluate the molecular effects of warm temperatures on placentas, we conducted a differential gene expression (DE) analysis between the Preterm-warm vs. Preterm-cold, and Term-warm vs. Term-cold groups separately using edgeR. Our Principal Component Analysis (PCA) demonstrated clearer clustering between the warm and cold groups in preterm placentas compared to term placentas (Figure 2A).
The DE analysis identified a total of 48 significantly differentially expressed genes (DEGs) (log2FC > ±1, FDR < 0.05), with 40 genes upregulated and 8 genes downregulated (Figure 2B, Table 2, Supplemental Table S2). The majority of the top-hit upregulated genes were inflammatory cytokines and chemokines, including SERPINA1, IL1B, CCL3, CCL3L3, CCL4, CCL4L2, CCL20, and CXCL8 (FDR < 0.05) (Figure 2B, Table 2). In contrast, the downregulated DEGs were non-immune in nature (Figure 2B, Table 2). Heatmap analysis showed clustering of these upregulated and downregulated genes between the warm and cold-exposure groups (Figure 2C). Conversely, the DE analysis between the Term-warm and Term-cold groups did not identify any significantly differentially expressed genes (Supplemental Table S3).
Figure 2.
Differential gene expression analysis between warm and cold groups of placentas. (A) PCA plots showing group separation between warm and cold groups in preterm and term placentas. (B) Volcano plots showing DEGs in Preterm-warm placentas compared to Preterm-cold group. (C) Heat map showing expression of 48 significant DEGs from Preterm-warm vs. Preterm-cold placentas. (D) Signalling pathways in Preterm-warm placentas positively enriched by GSEA. (E) A selected set of gene expression comparison between term and preterm placentas.
Figure 2.
Differential gene expression analysis between warm and cold groups of placentas. (A) PCA plots showing group separation between warm and cold groups in preterm and term placentas. (B) Volcano plots showing DEGs in Preterm-warm placentas compared to Preterm-cold group. (C) Heat map showing expression of 48 significant DEGs from Preterm-warm vs. Preterm-cold placentas. (D) Signalling pathways in Preterm-warm placentas positively enriched by GSEA. (E) A selected set of gene expression comparison between term and preterm placentas.
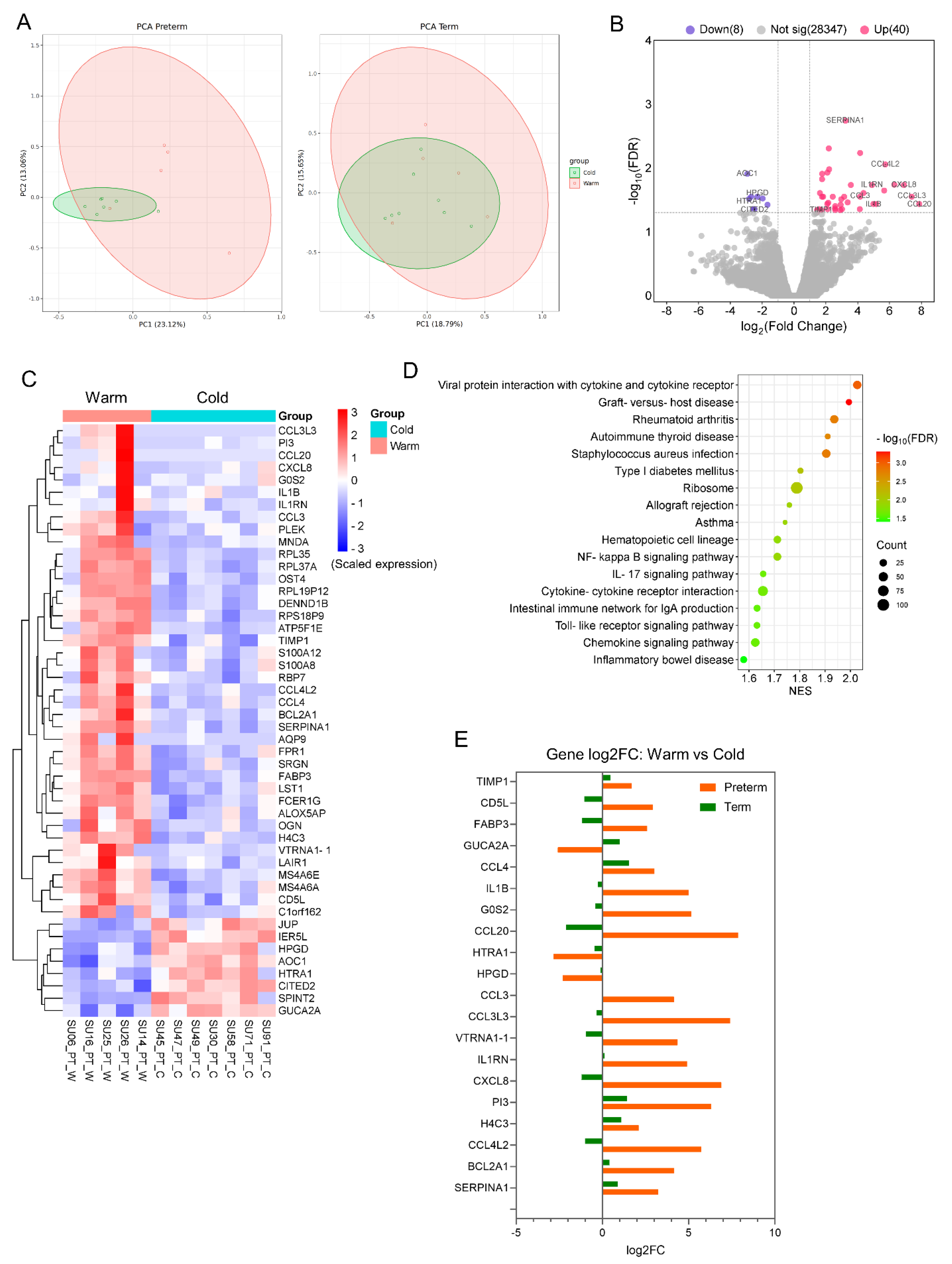
Next, we performed a GSEA analysis on the differentially expressed genes from preterm placentas using the KEGG Pathway functional database [18]. The upregulated genes positively enriched 17 signalling pathways, which were mostly related to inflammation, infection, immune regulation, and graft rejection (FDR < 0.05), including the NF-kappa B signalling (NES = 1.71), IL17 signalling (NES = 1.65), Toll-like receptor signalling (NES = 1.63), chemokine signalling (NES = 1.62), and allograft rejection (NES = 1.76) pathways (Figure 2D, Supplemental Table S4). These signalling pathways have implications for various adverse pregnancy outcomes. No signalling pathway was significantly enriched with the downregulated genes. Our GSEA analysis of differentially expressed genes from the Term-warm placentas positively enriched only one signalling pathway, which was the steroid hormone biosynthesis pathway (NES = 1.96, FDR = 0.001), but no negative enrichment was noted (Figure S1, Supplemental Table S5).
We also compared 20 selected DEGs from the Preterm-warm placentas with those genes from the Term-warm placentas (Figure 2E). The expression patterns of the inflammation- and chemokine-associated genes in the preterm placentas were opposite to the expression patterns of those genes in the term placentas (Figure 2E). For instance, IL1B, CXCL8, CCL4L2, CCL3, CCL3L3, and CCL20 genes were upregulated in the Preterm-warm group compared to the Preterm-cold group, whereas these genes were downregulated in the Term-warm group compared to the Term-cold group (Figure 2E).
Together, our data revealed a distinct transcriptomic signature associated with upregulated inflammation, immune modulation, and chemokine signalling in preterm placentas exposed to warm environmental temperatures compared to preterm placentas exposed to cold temperatures. This distinct signature was not observed in term placentas under similar thermal exposure conditions.
2.3. Warm Environmental Temperature Enriches Inflammatory and Immune Cells and Their Functions in Preterm Placenta
Our functional cell-type enrichment analysis identified significantly positive enrichment of various inflammatory and immune cells in preterm placentas exposed to warm temperatures compared to the cold exposure group (FDR < 0.05) (Figure 3A). No negative enrichment was detected in this analysis. The positively enriched cells within the preterm placentas (chorionic villi) were predominantly antigen-presenting cells/dendritic cells, neutrophils, monocytes, T and B cells, and M2 macrophages (Figure 3A). The major DEGs associated with the enriched cell types are illustrated in the chord plot (Figure 3B). Besides immune cells, sinusoidal endothelial cells, fetal fibroblasts, and mesenchymal progenitor cells were also positively enriched in the Preterm-warm placentas (Figure 3A). No cell-type enrichment was observed in the Term-warm placentas when compared with the Term-cold group (data not shown).
Our gene ontology (GO) functional analysis on the upregulated DEGs from the Preterm-warm placenta group identified several significant biological processes (FDR < 0.05), predominantly associated with neutrophil, granulocyte, monocyte, and lymphocyte chemotaxis/migration, inflammatory responses, chemokine/cytokine-mediated signalling pathways, and humoral immune responses (Figure 3C, Supplemental Table S6). The major cellular components (CC) involved in these biological processes were identified as secretory granules, secretory vesicles, vesicle lumens, and secretory granule lumens, suggesting a secretory phenotype of the immune cells potentially active within the preterm placentas delivered during the warm seasons (FDR < 0.05, Figure 3C, Table S6). Downregulated DEGs in the Preterm-warm group did not significantly identify any biological process via the GO analysis. As there were no significant DEGs detected in term placentas, GO analysis was not conducted.
Table 2.
Table showing 48 significantly differentially expressed genes (DEGs) (FDR < 0.05) in preterm placentas exposed to warm environmental temperature, of which 40 DEGs were upregulated and 8 DEGs were downregulated. Genes are ranked based on log2FC in descending order.
Table 2.
Table showing 48 significantly differentially expressed genes (DEGs) (FDR < 0.05) in preterm placentas exposed to warm environmental temperature, of which 40 DEGs were upregulated and 8 DEGs were downregulated. Genes are ranked based on log2FC in descending order.
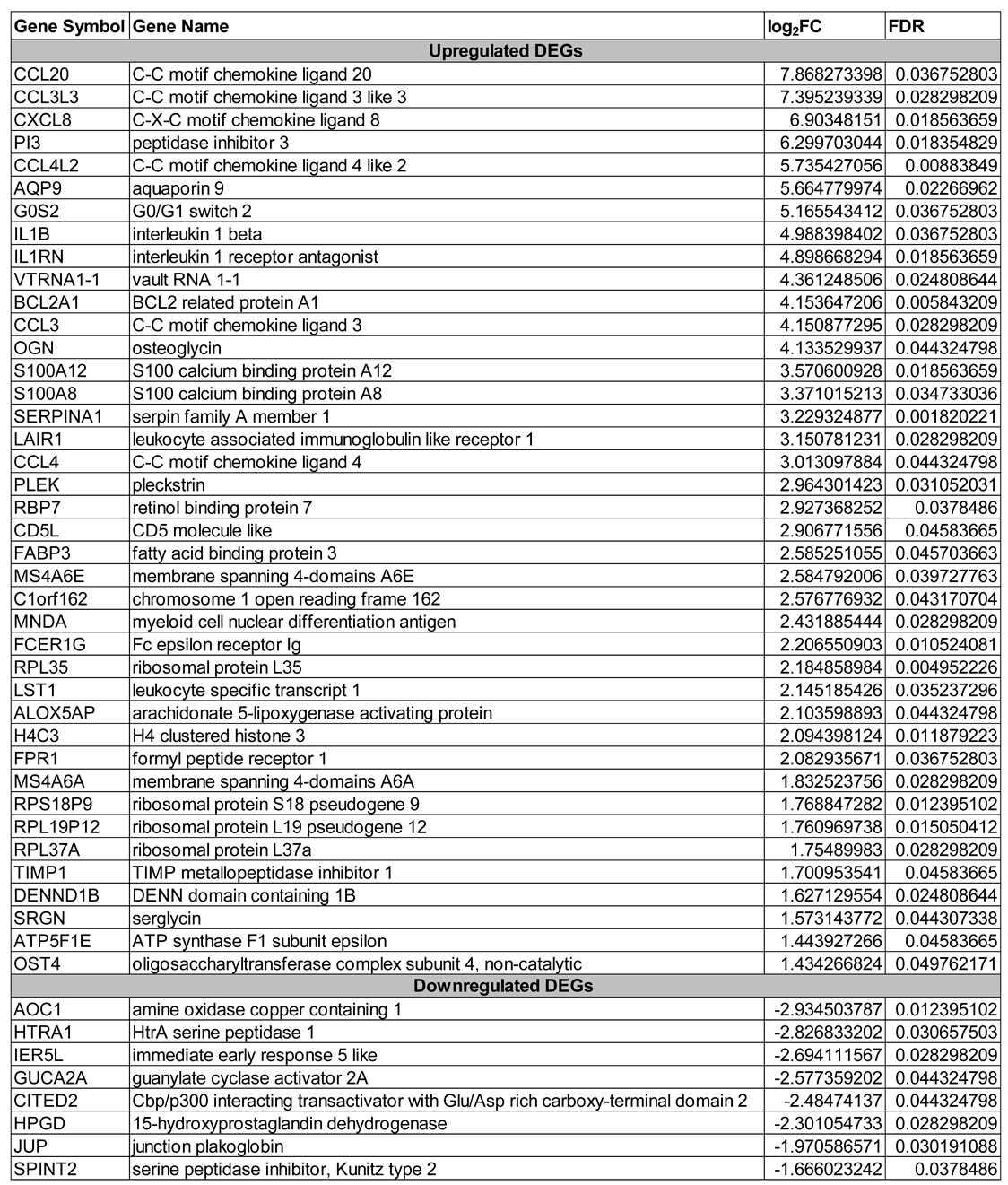 |
Figure 3.
Cell-type enrichment and Gene Ontology analysis. (A) Bar plot showing positive enrichment of cell-types in Preterm-warm placentas identified by GSEA. (B) Chord plot showing links between individual DEGs and the enriched cell-types. (C) Bar plot showing biological processes and cell components significantly enriched with the upregulated DEGs in Preterm-warm placentas. (D) Bar plot showing disease phenotypes enrichment with the upregulated DEGs in the Preterm-warm placentas.
Figure 3.
Cell-type enrichment and Gene Ontology analysis. (A) Bar plot showing positive enrichment of cell-types in Preterm-warm placentas identified by GSEA. (B) Chord plot showing links between individual DEGs and the enriched cell-types. (C) Bar plot showing biological processes and cell components significantly enriched with the upregulated DEGs in Preterm-warm placentas. (D) Bar plot showing disease phenotypes enrichment with the upregulated DEGs in the Preterm-warm placentas.
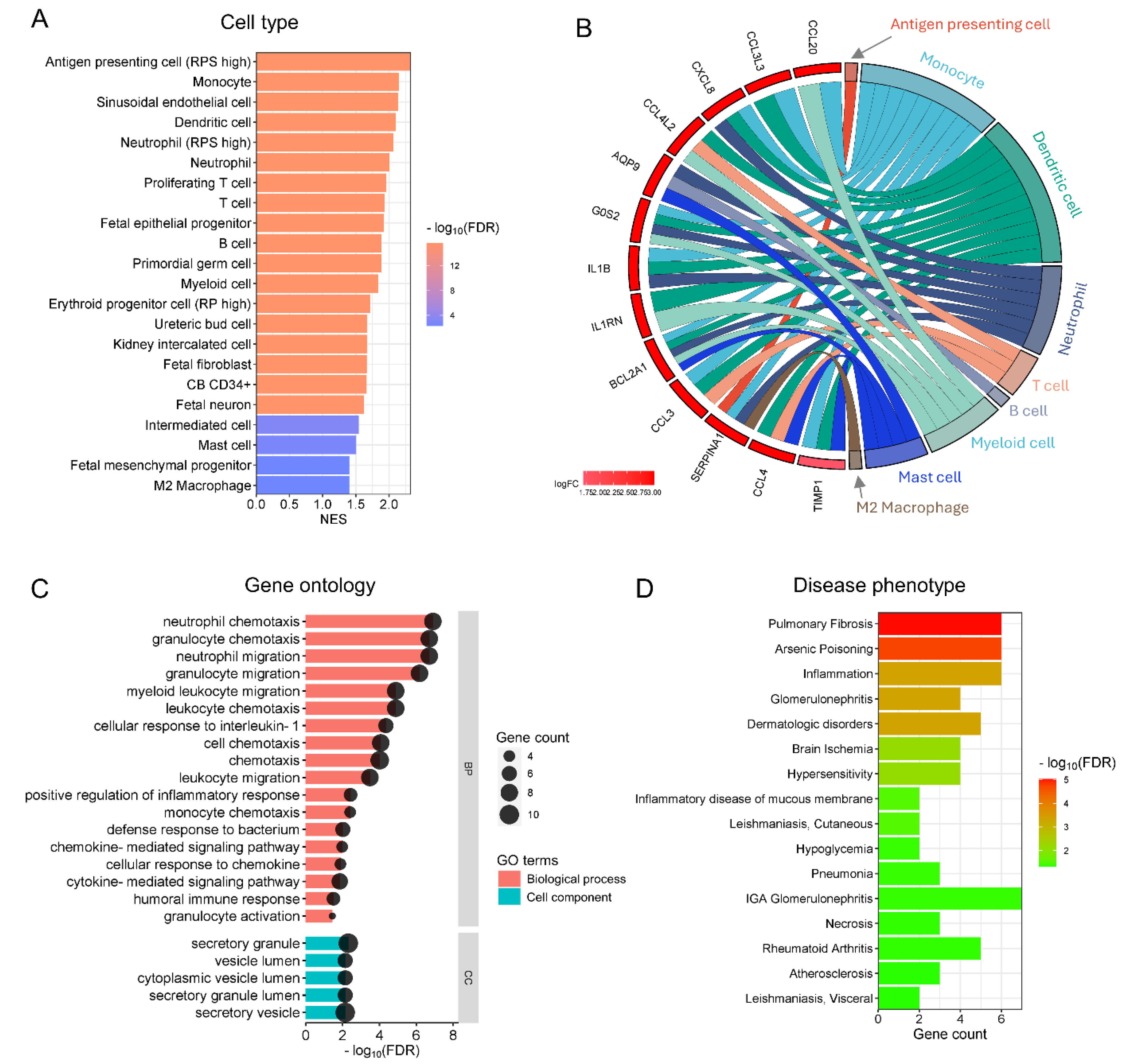
Next, we performed a functional enrichment analysis for disease phenotypes using the DisGeNET disease database on the differentially expressed genes from the Preterm-warm placenta group. Our analysis positively enriched several inflammatory and immune-related disease conditions (FDR < 0.05), including pulmonary fibrosis, glomerulonephritis, rheumatoid arthritis, inflammation, and hypersensitivity (Figure 3D).
Together, our data suggest that exposure to warm temperatures in preterm placentas potentially favours immune cell migration, chemotaxis, and vesicular secretion, facilitating inflammatory and immune responses, which were not observed in the term placentas delivered during the summer months.
2.4. Key Genes Linked to Placental Inflammatory and Immune Responses Secondary to Increased Environmental Temperature
Next, we selected a panel of genes based on their biological associations with inflammation, immune modulation, and placental functions, which were significantly upregulated or downregulated in the Preterm-warm placentas (Figure 4). We performed the Wilcoxon signed-rank test to compare the normalized gene expression (determined by edgeR) between Preterm-warm and Preterm-cold groups.
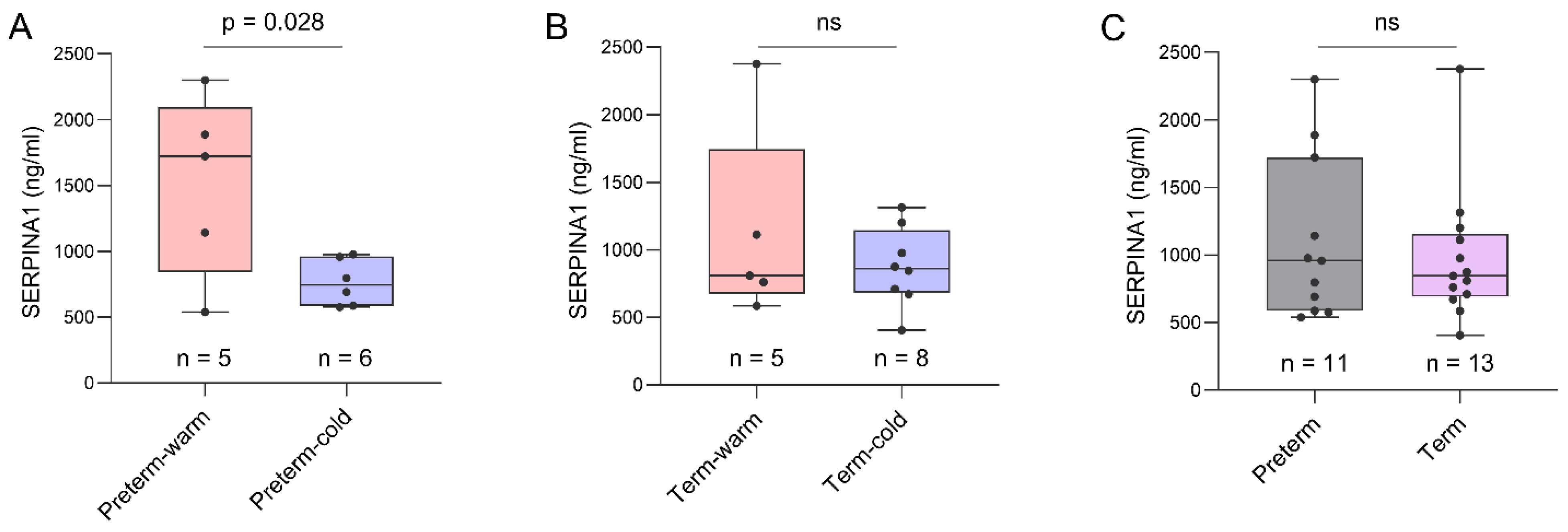
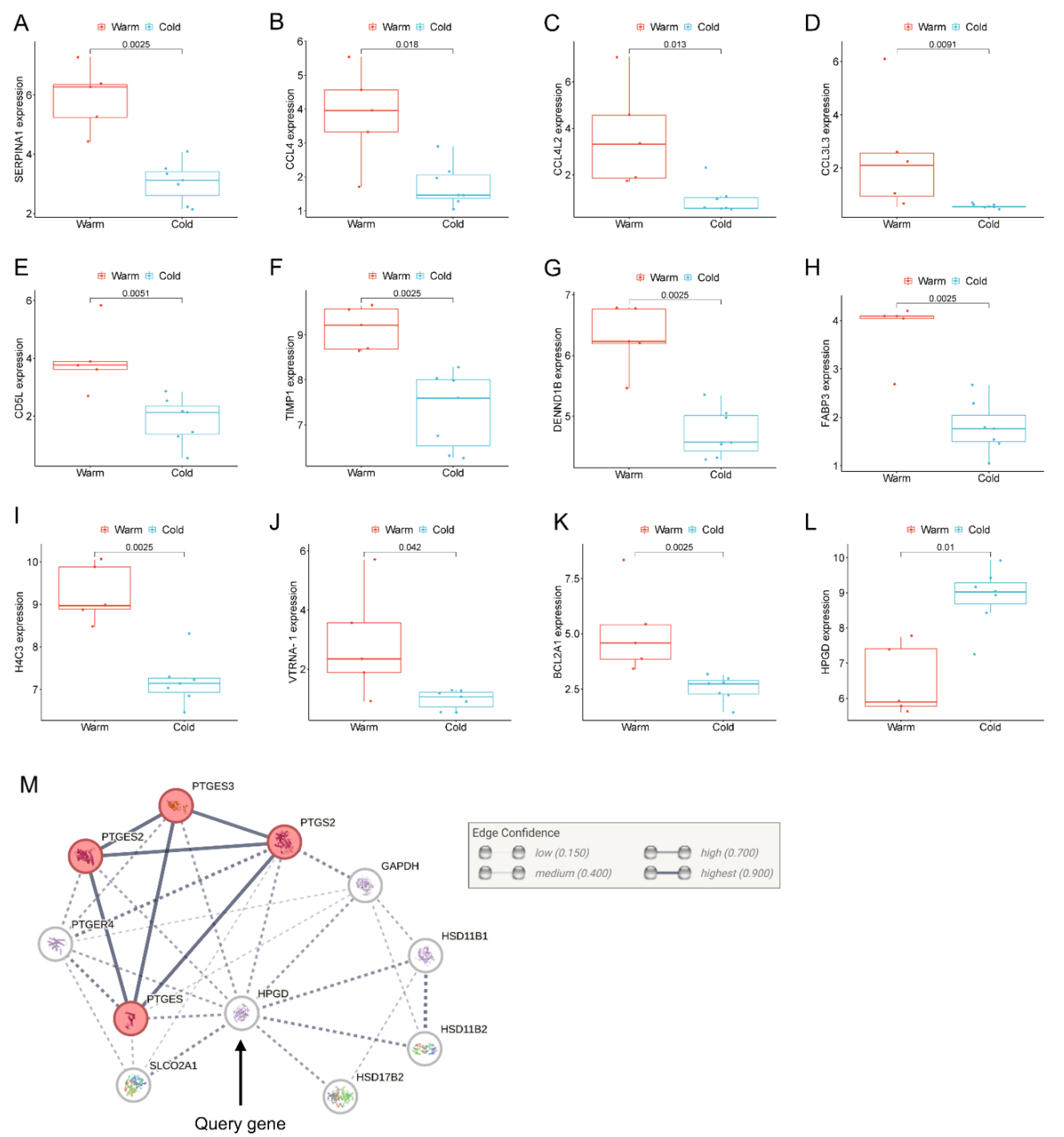
SERPINA1, which encodes for α1-antitrypsin protein, is an acute-phase response protein with anti-inflammatory and immunomodulatory properties. It is predominantly produced by hepatocytes but also produced by neutrophils, monocytes, and macrophages in response to inflammation [19]. SERPINA1 was the most significantly upregulated gene in the Preterm-warm placentas compared to the Preterm-cold group as determined by our DE analysis (FDR = 0.001, Figure 2B, Table 2). The Wilcoxon test also confirmed its significant expression (p = 0.002, Figure 4A). This gene was linked with the regulation of APCs, neutrophils, monocytes, and M2 macrophages as determined by functional analysis (Figure 3A,B).
We conducted an ELISA assay on the same placental tissue samples used for RNA-seq analysis to confirm the expression of SERPINA1 at the protein level. The mean SERPINA1 protein concentration was ~2-fold higher in Preterm-warm placentas compared to Preterm-cold placentas (1518 ng/ml vs. 764.2 ng/ml; p = 0.028) (Figure 5A). The SERPINA1 protein concentration was 29% higher in Term-warm placentas compared to Term-cold placentas, although this difference was not statistically significant (Figure 5B). Additionally, we compared the SERPINA1 protein concentrations between term (n = 13) and preterm placentas (n = 11), finding no significant difference (Figure 5C). Cumulatively, these protein data are consistent with our RNA-seq findings.
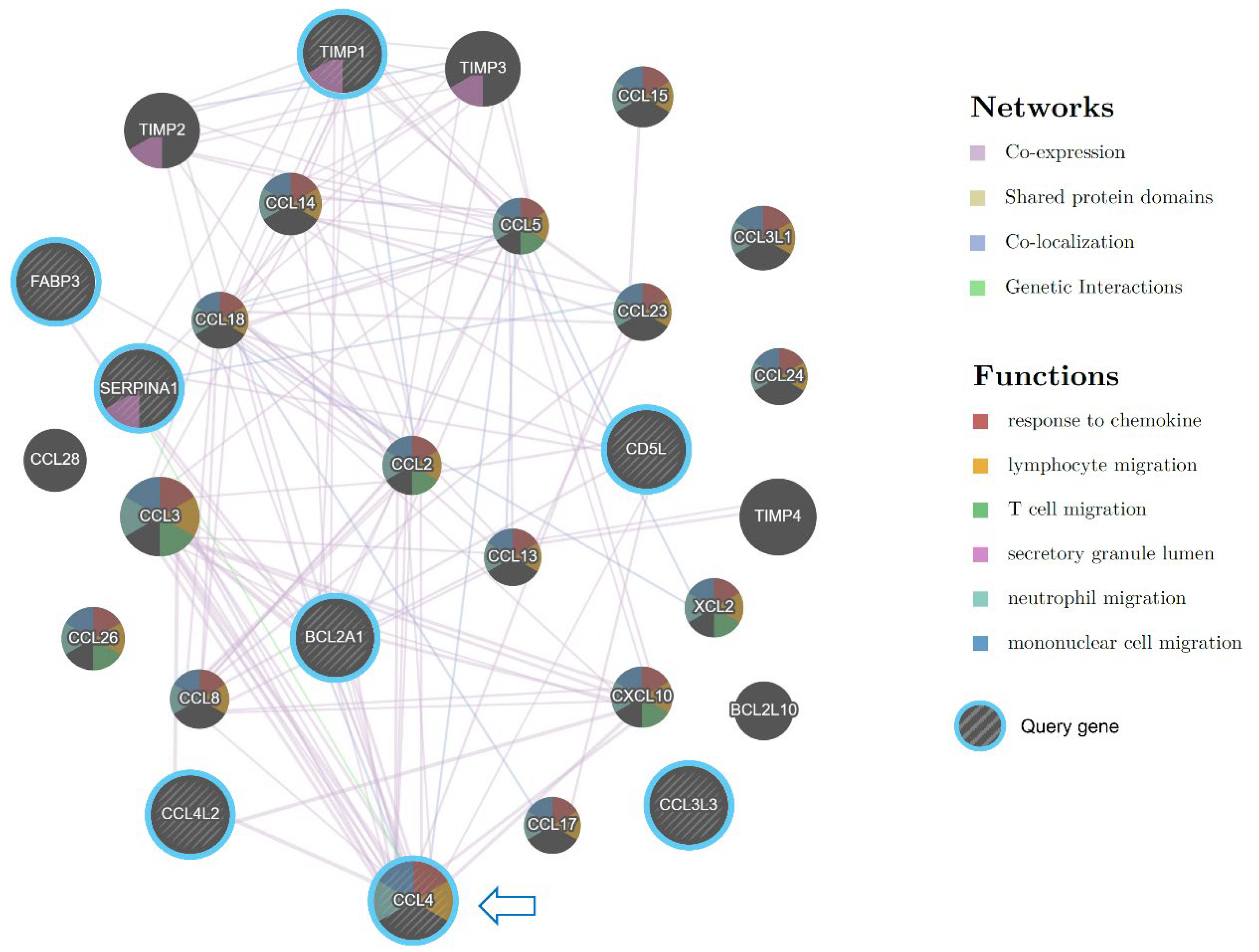
Among the upregulated chemokines in the Preterm-warm placentas, CCL4 was the most highly upregulated chemokine (log2FC = 3; Figure 4B). CCL4 is a potent chemokine released by monocytes, lymphocytes, neutrophils, fibroblasts, and endothelial cells. It critically regulates immune and inflammatory responses by attracting a diverse population of immune cells [20,21,22]. The gene network analysis identified CCL4 as one of the most functionally connected genes with other immune regulatory genes (Figure 6).
CD5L, encoding a soluble glycoprotein of the SRCR superfamily, modulates macrophage activities in inflammatory processes via macrophage polarization towards the M2 phenotype [23,24]. CD5L was significantly upregulated in the Preterm-warm placentas, where M2 macrophage enrichment was also noted (FDR = 0.005) (Figure 2, Table 2, cell-type enrichment, Figure 3A and Figure 4E). Another emerging immunomodulator, TIMP1, which fosters the inflammatory process via its cytokine activity, was significantly overexpressed in the Preterm-warm placentas (Figure 4F) [25]. DENND1B, a T-cell receptor (TCR) internalization regulator in TH2 cells, was also upregulated in Preterm-warm placentas (Figure 4G, Table 2).
Fatty acid binding proteins (FABPs) are involved in fatty acid metabolism and trafficking in the placenta. FABP3 and FABP6 proteins have been found to be increased within the placental barrier in obese pregnant mice, enhancing fatty acid transport to the fetus [26]. In our analysis, FABP3 was significantly upregulated in Preterm-warm placentas compared to the Preterm-cold group (Figure 4H, Table 2). In the Term-warm placenta group, this gene was downregulated (though not significantly) (Figure 2E).
Figure 5.
ELISA assay on placenta tissue. Box plots showing SERPINA1 protein concentrations in Preterm-warm vs. Preterm-cold placentas (A) and Term-warm vs Term-cold placentas (B). (C) Cumulative comparison of SERPINA1 protein concentrations between all preterm and all term placentas. A protein concentration of 300 µg/ml was used as the loading concentration for each sample in the ELISA assay. Data are presented as median with IQR. Each dot represents an individual subject. The p value was determined by a two-tailed unpaired Student’s t-test. ns = not significant.
Figure 5.
ELISA assay on placenta tissue. Box plots showing SERPINA1 protein concentrations in Preterm-warm vs. Preterm-cold placentas (A) and Term-warm vs Term-cold placentas (B). (C) Cumulative comparison of SERPINA1 protein concentrations between all preterm and all term placentas. A protein concentration of 300 µg/ml was used as the loading concentration for each sample in the ELISA assay. Data are presented as median with IQR. Each dot represents an individual subject. The p value was determined by a two-tailed unpaired Student’s t-test. ns = not significant.
One of the four core histone genes, H4C3, encodes a protein that forms an octamer and regulates chromatin organization. It was upregulated in the preterm placentas exposed to warm temperatures (Figure 4I, Table 2). Among other non-immune-associated genes, VTRNA1-1 and BCL2A1 were significantly upregulated in the Preterm-warm placentas compared to the Preterm-cold group, which are associated with impaired autophagy and apoptosis, respectively (Figure 4J,K).
15-hydroxyprostaglandin dehydrogenase (HPGD) catalyses prostaglandins, resulting in their functional inactivation [27]. In our DE analysis and Wilcoxon test, HPGD was significantly downregulated in the Preterm-warm placentas compared to Preterm-cold placentas (Figure 2C and Figure 4L, Table 2). Due to its diverse actions on prostaglandin E synthases (STRING analysis, Figure 4M), inactivation of this gene may augment prostaglandin-driven mechanisms, including inflammation and the initiation of labour [28].
3. Discussion
During pregnancy, physiological and anatomical adaptations challenge maternal thermoregulation in response to excessive external heat. As pregnancy progresses, internal heat production rises due to fetal and placental metabolism; however, dissipation of this excess heat is limited by increased maternal body mass and a relative reduction in body surface area [29]. A growing body of evidence strongly links maternal exposure to heat, particularly during the second and third trimesters, with increased risks of spontaneous preterm birth across diverse geographical regions, including the West, Sub-Saharan Africa, and Asia [3,4,5,6,7,8,9,10]. The underlying mechanisms of heat exposure-induced preterm birth (PTB) remain elusive. Animal model studies have postulated that high ambient temperatures alter maternal systemic thermoregulation capacity, resulting in cortisol release and subsequent oxytocin-induced uterine contractions and labour [5,8,30,31]. However, the effects of maternal heat exposure on placental molecular functions are not yet fully understood.
Here, we provide the first report on the human placenta showing that maternal exposure to warm environmental temperatures predominantly altered the placental transcriptome in cases of spontaneous preterm birth, a phenomenon not as prominent in term births. These transcriptomic alterations were primarily associated with placental inflammation, immune regulation, and allograft rejection signalling pathways. The dominant driver cells in these signalling pathways were inflammatory and immune cells, including neutrophils, monocytes, M2 macrophages, T and B lymphocytes, and dendritic cells, likely originating from the villous placenta.
This observation was noted in preterm placentas from pregnant women who delivered during the British summer period, with a mean temperature Tmean = 14.92 °C and an average maximum temperature Tmax = 20.47 °C. For the comparable cold group, the mean temperature was 5.14 °C, with a maximum temperature of 8.37 °C. It was estimated that the women experienced these temperatures for an average of 9 weeks during the warmer months and 7.3 weeks during the colder months preceding parturition. Although these weather temperatures might be perceived as within a comfortable range, in colder geographical regions such as South Yorkshire, where people are naturally acclimatized to colder temperatures, a rise of 10-12 °C during the summertime could cause discomfort or heat exhaustion for pregnant women, as documented by several studies [5,8,32]. This evidence and assumption provided the basis for comparing the transcriptomic data between these two thermal groups (warm vs. cold) to evaluate the effects of temperature.
The placenta plays a major role in maintaining fetal growth and gestation. Placental dysfunction has been implicated in various adverse pregnancy outcomes, including preeclampsia, fetal growth restriction, recurrent miscarriage, stillbirth, and preterm birth. The key functions of the placenta are driven by the cellular elements within the villous compartment, including cytotrophoblasts, syncytiotrophoblasts, extravillous trophoblasts, mesenchymal cells, endothelial cells, and immune cells [33].
The RNA-seq datasets used in this study were generated from the villous tissue (devoid of decidua and maternal blood) of the placentas containing the aforementioned cell populations, as demonstrated by our previous work [34]. Our functional enrichment analysis for ‘cell-type’ categories did not show enrichment for trophoblast cells but did show positive enrichment for inflammatory and immune cells, as well as endothelial, mesenchymal, and fetal epithelial progenitor cells in the preterm-warm placentas. This differential enrichment suggests an immunomodulatory response of the preterm placenta to exposure to warm temperatures during pregnancy. This inference is supported by our Gene Ontology (GO) analysis, which identified the enrichment of biological processes involving immune/inflammatory cell migration, chemotaxis, and humoral immune responses. Interestingly, the enriched cellular components (CC) were associated with vesicular secretion but not locomotion. Therefore, we speculate that the placental resident immune and parenchymal cells released chemokines and inflammatory cytokines, including SERPINA1, IL1B, CCL3, CCL3L3, CCL4, CCL4L2, CCL20, and CXCL8 (as determined by DE analysis), which likely attract maternal immune cells migrating into the villous placenta, thereby altering the normal immune balance. This hypothesis, however, requires confirmation through exploratory ex vivo and in vitro experiments.
While there is a lack of research on the molecular effects of heat exposure on the human placenta, an interesting study has shown that maternal stress due to climate-related disasters, such as hurricanes, during pregnancy altered the placental transcriptome, which was linked to infant temperament. This altered transcriptome identified an enrichment of functional pathways related to inflammation, extracellular matrix integrity, and sensory perception [35]. A mouse model study demonstrated that maternal exposure to cold ambient temperatures (18 °C vs. 28 °C) was associated with the upregulation of genes involved in antigen processing and presentation, cytokine receptor interaction, and complement activation, and the downregulation of genes involved in oxidative phosphorylation, myofibril assembly, and muscle contraction [36]. Cold exposure-induced differentially expressed genes also enriched several biological processes, including prostaglandin biosynthesis, regulation of the humoral immune response, regulation of the acute inflammatory response, Staphylococcus aureus infection, and macrophage migration/chemotaxis [36].
These findings have similarities with what we observed in our Preterm-warm placentas, where pregnant women were exposed to warm summer temperatures during their third trimester (Figure 2 and Figure 3). These findings suggest that placental molecular responses to thermal exposure follow similar patterns that are predominantly inflammatory and immunomodulatory in nature. However, it is important to note that we did not detect any significant transcriptomic alterations in the term placentas exposed to similar thermal conditions. The reasons for this difference are not apparent but could be due to the presence of certain maternal or placental predisposing factors that prevent the thermal modulation of placental transcriptomes.
We identified 48 differentially expressed genes in the preterm placentas delivered during the summertime. The majority of the top 25 upregulated genes were associated with inflammation, immune regulation, and graft rejection. These biological processes are crucial for the immune tolerance of the placenta toward the allogeneic fetus and the labour process [34,37,38,39,40]. Our GSEA analysis positively enriched several signalling pathways in the Preterm-warm placentas, including NF-κB signalling, Toll-like receptor signalling, and IL-17 signalling.
The placental NF-κB signalling pathway remains downregulated to maintain pregnancy. Its activation initiates a cascade of events that facilitate the onset of labour through the production of pro-inflammatory cytokines, chemokines, and prostaglandin-synthesizing enzymes [41]. Infiltrating immune cells in the villous placenta secrete chemokines and cytokines, leading to the activation of NF-κB [42]. Premature aberrant activation of NF-κB signalling has been implicated in PTB [43]. This putative activation of NF-κB signalling by the infiltrating immune cells aligns with our assumption that heat-exposed preterm placentas may favour the chemotaxis of immune cells from the maternal circulation to the placenta.
Toll-like receptor (TLR) signalling and its functional activation have been shown to increase in PTB [44]. Stimulation of TLRs induces the release of pro-inflammatory cytokines in the fetal membrane and placenta [45,46]. Activation of TLR signalling leads to uterine contractions, cervical ripening, and preterm premature rupture of membranes (PPROM) in PTB [44]. Therefore, it is likely that heat exposure during pregnancy could amplify the risk of preterm labour via upregulation of TLR signalling, as we have demonstrated. Additionally, studies have shown that IL17, produced by T cells, promotes inflammation at the fetomaternal interface of the placenta in PTB [47]. Our analyses showed that a warm environment positively enriched both T cells and IL-17 signalling in the preterm placenta. Therefore, this ‘T cell-IL-17 signalling axis’ could be linked to the detrimental effects on placental function associated with thermal exposure.
Out of the 48 differentially expressed genes, we identified 12 genes as important mediators in thermal responses based on their expression patterns and relevance to pregnancy and placental functions (Figure 4). Among them, SERPINA1, which encodes for α1-antitrypsin, stands out as a potential biomarker target. Increased levels of α1-antitrypsin have been detected in the villous placenta of preeclampsia patients, where it was associated with syncytiotrophoblast destruction and secretion into urine [48]. Detection of placenta-originated α1-antitrypsin in urine has been suggested as a potential biomarker for assessing preeclampsia severity [48]. Interestingly, α1-antitrypsin expression at the gene and protein levels was decreased in spontaneous PTB compared to term birth [49]. Conversely, a recent study demonstrated a significant increase in α1-antitrypsin levels in maternal blood from spontaneous PTB compared to term birth during the 1st and 2nd trimesters, though no difference was observed at delivery [50].
In our analysis, SERPINA1 was the most significantly upregulated gene in the Preterm-warm group of placentas compared to the corresponding cold group, which was also true for its protein level expression (Figure 5), suggesting its potential as a biomarker for assessing placental response to high-heat exposure in high-risk pregnancies, particularly in cervicovaginal fluid (CVF) and maternal blood. Additionally, CCL4 and TIMP1 are also promising candidates worth exploring as serum and CVF biomarkers to assess placental responses to heat exposure.
Heat stress and heat acclimation can induce epigenetic modifications such as DNA methylation and histone modifications, which help protect cells from thermal damage by adjusting the transcriptional levels of heat-responsive genes [51]. In our study, although no classical heat-responsive genes like heat-shock proteins were differentially expressed, we observed significant upregulation of the core histone gene H4C3 in Preterm-warm placentas. Interestingly, H4C3 was also upregulated in Term-warm placentas, although this increase was not statistically significant (Figure 2E). The H4C3 protein plays a critical role in nucleosome formation and chromatin organization, thereby regulating DNA replication and transcription. Variants of this gene have been associated with various pathologies, including neurodevelopmental disorders [52]. The thermal responsiveness of H4C3 in the context of placental pathology remains unclear and warrants further investigation.
Prostaglandins play pivotal roles in initiating human parturition by promoting uterine contraction and cervical ripening, and their inhibition can delay labour [28,53]. The enzyme HPGD catalyses the functional inactivation of prostaglandins [27]. In our study, HPGD was significantly downregulated in preterm placentas exposed to warm environmental temperatures. Placental HPGD has been reported to be markedly downregulated in both animal models and human preterm birth [54,55]. Therefore, it is plausible that the suppression of HPGD induced by warm temperatures may predispose to premature labour, although this hypothesis requires validation through further exploratory research.
Study limitations: Although our findings align with the transcriptomic data from controlled thermal-exposure animal studies, it should be noted with caution that our conclusions are based on a relatively small sample size. The temperature exposure for our study participants was estimated using regional weather data. Including measurements of body and domiciliary temperatures of the pregnant women during their third trimester, in addition to regional weather data, could provide a more accurate assessment of temperature exposure. However, there is a growing body of evidence indicating that pregnant women are more likely to experience heat exhaustion during summer compared to winter across various geographical regions, including high-income countries [5,8,32]. Therefore, it is likely that our selected cohort of participants, who delivered during warmer and colder months in the South Yorkshire region, experienced the respective thermal conditions.
4. Conclusion
Maternal exposure to elevated environmental temperatures during the third trimester of pregnancy significantly altered the placental transcriptomes, predominantly in cases of spontaneous preterm birth, but not in term births. This transcriptomic alteration was primarily associated with placental inflammation, immune regulation, and allograft rejection. The predominant cells driving these signalling pathways were inflammatory and immune cells, including neutrophils, monocytes, M2 macrophages, T and B lymphocytes, and dendritic cells. However, it remains uncertain whether these placental responses were a direct effect of temperature on the placenta or secondary to maternal systemic responses to warm exposure. Our data revealed a potential detrimental impact on placental functions due to long-term exposure to moderately elevated temperatures during pregnancy. These findings warrant further large-scale molecular studies across diverse geographical regions, particularly in low- and middle-income countries (LMICs), to better understand the thermal effects on placental function. This research is crucial for combating the impacts of global climate change on vulnerable pregnant women to save maternal and neonatal lives.
Figure 6.
Gene interaction network analysis by Cytoscape plug-in GeneMANIA. Striped bigger nodes indicate DEGs upregulated in Preterm-warm placentas. Colour codes inside each node indicate their biological function as stated in the legends. Coloured edges and their connections with other genes indicate the nature of interactions.
Figure 6.
Gene interaction network analysis by Cytoscape plug-in GeneMANIA. Striped bigger nodes indicate DEGs upregulated in Preterm-warm placentas. Colour codes inside each node indicate their biological function as stated in the legends. Coloured edges and their connections with other genes indicate the nature of interactions.
5. Methods
5.1. Study Participants and Thermal Exposure Groups Stratification
A total of 25 raw FASTQ RNA-seq files out of 40 RNA-seq datasets were downloaded from the NCBI Gene Expression Omnibus (GEO) repository using accession number GSE211927 (Supplemental Table S1). The datasets were divided into four groups based on the time of delivery and gestation. Women who delivered between June and September and spent at least one week in this period before spontaneous delivery at term (>37 weeks gestation) or preterm (<37 weeks gestation) were categorized as the warm-exposure group. Similarly, women who delivered between November and March and spent at least one week in this period before delivery were categorized as the cold-exposure group (Supplemental Table S1). Temperatures and exposure durations are tabulated in Table 1 and Table S1 and are presented in Figure 1. All participants resided in the South Yorkshire region in the UK during the pregnancy and immediately preceding labour. Regional weather temperatures (monthly mean and maximum temperatures) were collected from the UK Met Office websites: (https://www.metoffice.gov.uk/research/climate/maps-and-data/uk-and-regional-series and https://www.metoffice.gov.uk/research/climate/maps-and-data/uk-climate-averages/).
5.2. Differential Gene Expression (DE) Analysis
FASTQ RNA-seq files (Nanopore long reads) (NCBI SRA files) were downloaded to the Galaxy web server (Galaxy version 24.1.0.dev0) [56], and adapters were removed using the Porechop tool. Reads were further trimmed with the Trim Galore tool to extract high-quality reads with a Phred score ≥ 10, which is considered good quality in Nanopore sequencing. The final read quality was assessed using pycoQC (v2.5.2). Quality-passed reads were mapped to the reference human genome (hg38) using minimap2 (v2.28) [57]. Post-mapping BAM files were used for feature counting with the featureCounts tool [58].
Next, differential gene expression (DE) analyses between warm and cold groups in term and preterm placentas were conducted using the edgeR tool (v3.36.0), with TMM data normalization and Benjamini-Hochberg false discovery rate (FDR) correction [59]. To identify differentially expressed genes (DEGs), an FDR (adjusted p-value) threshold of <0.05 and a log2FC cutoff of > ±1 were applied.
5.3. Gene Set Enrichment Analysis (GSEA) and Gene Ontology (GO) Analysis
To explore the underlying signalling pathways, GSEA analysis was performed on the differentially expressed genes table using the KEGG database via the WebGestalt 2024 web tool [18,60,61]. GSEA was also conducted to determine cell-type and disease phenotype enrichment using the Human Cell Landscape and DisGeNET databases, respectively (WebGestalt). To explore the underlying biological processes, Gene Ontology (GO) analysis was performed on the significantly upregulated and downregulated DEGs using the Over-Representation Analysis (ORA) enrichment method [18]. An FDR value of <0.05 was considered statistically significant for all enrichment analyses.
5.4. ELISA Assay
Protein extraction from placental villous tissue and the ELISA assay were conducted following our previously published protocol [34]. Briefly, total protein was extracted from 100 mg of snap-frozen villous tissue (biopsied from an identical location from where tissue samples were harvested for the RNA-seq analysis [17]) using SigmaFAST/IgePal lysis buffer (Sigma-Aldrich, Cat No: I3021) with SigmaFAST protease inhibitor cocktail (Sigma-Aldrich, Cat No: S8830) according to the manufacturer’s instructions. The tissue was homogenised using a TissueLyser LT for 10 minutes at 50 Hz at room temperature, followed by incubation at 4 °C for 30 minutes. The fully lysed tissue homogenate was centrifuged at 16,000× g for 20 minutes at 4 °C. The supernatant was collected, and the total protein concentration was determined using the Qubit Protein Assay Kit (Thermo Fisher Scientific, Cat No: Q33211). For the ELISA assay, total protein was diluted to a concentration of 300 μg/ml in PBS for each sample. Quantification of SERPINA1 protein was performed using the Human SERPINA1 ELISA Kit (ProteinTech, Cat No: KE00178) according to the manufacturer’s instructions. A sigmoidal four-parameter logistic curve was used to generate a standard curve for quantifying protein concentration using GraphPad Prism (version 10.2.3).
5.5. Statistical Analysis
The statistical analyses and generation of plots were performed using GraphPad Prism (v10), the Galaxy web server, Cytoscape-GeneMANIA [62], and the SRplot web server [63]. The Mann-Whitney U test, Wilcoxon signed-rank test and Student t test were utilised to compare two groups, while the One-way ANOVA was employed for comparisons involving more than two groups. A p-value < 0.05 was considered statistically significant. Data are presented as mean ± standard deviation (SD) or as median and interquartile ranges (IQR), as specified in the respective figure legends.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Conceptualisation: KA and DA; Study design: KA; RNA-seq and weather data acquisition: KA and ED; Bioinformatics and data analyses: KA and ED; ELISA assay and analysis: KA; Figures and Tables: KA; Original first draft preparation: KA; Manuscript revision: DA; Funding acquisition: KA and DA; Project supervision: KA and DA; All authors have read and agreed to the published version of the manuscript.
Funding
The study was funded by the United Kingdom’s National Institute for Health Research (NIHR) Global Health Research Scheme to DA (GHR 17/63/26). Additional funding by The University of Sheffield Postgraduate Research funds.
Ethical Approval
This study involving human participants was approved by London-Fulham Research Ethics Committee, NHS Health Research Authority (18/LO/2044). The patients provided their written informed consent to participate in this study.
Data Availability Statement
Additional data can be found in the supplemental files. The previously published RNA-seq datasets used in this manuscript can be found in the NCBI Gene Expression Omnibus (GEO) repository using accession number GSE211927 (Supplemental Table S1). Any additional data from this study are available from the corresponding author upon reasonable request.
Acknowledgement
The authors wish to thank all the researchers of the PRIME research group at the University of Sheffield and the dedicated midwives, research nurse and doctors at the Jessop Wing Maternity Hospital for assisting in consenting patients, placenta collections, tissue harvesting and the Site File management.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Watts, N.; Amann, M.; Arnell, N.; Ayeb-Karlsson, S.; Beagley, J.; Belesova, K.; Boykoff, M.; Byass, P.; Cai, W.; Campbell-Lendrum, D.; et al. The 2020 report of the Lancet Countdown on health and climate change: responding to converging crises. Lancet 2021, 397, 129–170. [Google Scholar] [CrossRef] [PubMed]
- Hoegh-Guldberg, O.; Jacob, D.; Taylor, M.; Bolaños, T.G.; Bindi, M.; Brown, S.; Camilloni, I.A.; Diedhiou, A.; Djalante, R.; Ebi, K.; et al. The human imperative of stabilizing global climate change at 1.5°C. Science 2019, 365, aaw6974. [Google Scholar] [CrossRef] [PubMed]
- Judge, C.M.; Chasan-Taber, L.; Gensburg, L.; Nasca, P.C.; Marshall, E.G. Physical exposures during pregnancy and congenital cardiovascular malformations. Paediatr. Périnat. Epidemiology 2004, 18, 352–360. [Google Scholar] [CrossRef] [PubMed]
- Suarez, L.; Felkner, M.; Hendricks, K. The effect of fever, febrile illnesses, and heat exposures on the risk of neural tube defects in a Texas-Mexico border population. Birth Defects Res. A Clin. Mol. Teratol. 2004, 70, 815–819. [Google Scholar] [CrossRef] [PubMed]
- Chersich, M.F.; Pham, M.D.; Areal, A.; Haghighi, M.M.; Manyuchi, A.; Swift, C.P.; Wernecke, B.; Robinson, M.; Hetem, R.; Boeckmann, M.; et al. Associations between high temperatures in pregnancy and risk of preterm birth, low birth weight, and stillbirths: systematic review and meta-analysis. BMJ 2020, 371, m3811. [Google Scholar] [CrossRef] [PubMed]
- Bekkar, B.; Pacheco, S.; Basu, R.; DeNicola, N. Association of Air Pollution and Heat Exposure With Preterm Birth, Low Birth Weight, and Stillbirth in the US. JAMA Netw. Open 2020, 3, e208243–e208243. [Google Scholar] [CrossRef] [PubMed]
- Gong, Y.; Chai, J.; Yang, M.; Sun, P.; Sun, R.; Dong, W.; Li, Q.; Zhou, D.; Yu, F.; Wang, Y.; et al. Effects of ambient temperature on the risk of preterm birth in offspring of adolescent mothers in rural henan, China. Environ. Res. 2021, 201, 111545. [Google Scholar] [CrossRef] [PubMed]
- Syed, S.; O’sullivan, T.L.; Phillips, K.P. Extreme Heat and Pregnancy Outcomes: A Scoping Review of the Epidemiological Evidence. Int. J. Environ. Res. Public Heal. 2022, 19, 2412. [Google Scholar] [CrossRef] [PubMed]
- Ren, M.; Wang, Q.; Zhao, W.; Ren, Z.; Zhang, H.; Jalaludin, B.; Benmarhnia, T.; Di, J.; Hu, H.; Wang, Y.; et al. Effects of extreme temperature on the risk of preterm birth in China: A population-based multi-center cohort study. Lancet Reg. Heal. - West. Pac. 2022, 24, 100496. [Google Scholar] [CrossRef]
- Wang, Q.; Yin, L.; Wu, H.; Ren, Z.; He, S.; Huang, A.; Huang, C. Effects of gestational ambient extreme temperature exposures on the risk of preterm birth in China: A sibling-matched study based on a multi-center prospective cohort. Sci. Total. Environ. 2023, 887, 164135. [Google Scholar] [CrossRef]
- Chawanpaiboon, S.; Vogel, J.P.; Moller, A.-B.; Lumbiganon, P.; Petzold, M.; Hogan, D.; Landoulsi, S.; Jampathong, N.; Kongwattanakul, K.; Laopaiboon, M.; et al. Global, regional, and national estimates of levels of preterm birth in 2014: a systematic review and modelling analysis. Lancet Glob. Heal. 2018, 7, e37–e46. [Google Scholar] [CrossRef] [PubMed]
- Goldenberg, R.L.; Culhane, J.F.; Iams, J.D.; Romero, R. Epidemiology and causes of preterm birth. Lancet 2008, 371, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Bonell, A.; Sonko, B.; Badjie, J.; Samateh, T.; Saidy, T.; Sosseh, F.; Sallah, Y.; Bajo, K.; A Murray, K.; Hirst, J.; et al. Environmental heat stress on maternal physiology and fetal blood flow in pregnant subsistence farmers in The Gambia, west Africa: an observational cohort study. Lancet Planet. Heal. 2022, 6, e968–e976. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Liu, X.; Dong, M.; Sun, X.; Xiao, J.; Zeng, W.; Hu, J.; Li, X.; Guo, L.; Rong, Z.; et al. Associations of maternal ambient temperature exposures during pregnancy with the placental weight, volume and PFR: A birth cohort study in Guangzhou, China. Environ. Int. 2020, 139, 105682. [Google Scholar] [CrossRef] [PubMed]
- Galan, H.L.; Hussey, M.J.; Barbera, A.; Ferrazzi, E.; Chung, M.; Hobbins, J.C.; Battaglia, F.C. Relationship of fetal growth to duration of heat stress in an ovine model of placental insufficiency. Am. J. Obstet. Gynecol. 1999, 180, 1278–1282. [Google Scholar] [CrossRef] [PubMed]
- Limesand, S.; Regnault, T.; Hay, W. Characterization of Glucose Transporter 8 (GLUT8) in the Ovine Placenta of Normal and Growth Restricted Fetuses. Placenta 2004, 25, 70–77. [Google Scholar] [CrossRef] [PubMed]
- Akram, K.M.; Kulkarni, N.S.; Brook, A.; Wyles, M.D.; Anumba, D.O.C. Transcriptomic analysis of the human placenta reveals trophoblast dysfunction and augmented Wnt signalling associated with spontaneous preterm birth. Front. Cell Dev. Biol. 2022, 10, 987740. [Google Scholar] [CrossRef] [PubMed]
- Elizarraras, J.M.; Liao, Y.; Shi, Z.; Zhu, Q.; Pico, A.R.; Zhang, B. WebGestalt 2024: faster gene set analysis and new support for metabolomics and multi-omics. Nucleic Acids Res. 2024, 52, W415–W421. [Google Scholar] [CrossRef] [PubMed]
- Greene, C.M.; Marciniak, S.J.; Teckman, J.; Ferrarotti, I.; Brantly, M.L.; Lomas, D.A.; Stoller, J.K.; McElvaney, N.G. α1-Antitrypsin deficiency. Nat. Rev. Dis. Primers 2016, 2, 6051. [Google Scholar] [CrossRef]
- Sindhu, S.; Kochumon, S.; Shenouda, S.; Wilson, A.; Al-Mulla, F.; Ahmad, R. The Cooperative Induction of CCL4 in Human Monocytic Cells by TNF-α and Palmitate Requires MyD88 and Involves MAPK/NF-κB Signaling Pathways. Int. J. Mol. Sci. 2019, 20, 4658. [Google Scholar] [CrossRef]
- Mantovani, A.; Sica, A.; Sozzani, S.; Allavena, P.; Vecchi, A.; Locati, M. The chemokine system in diverse forms of macrophage activation and polarization. Trends Immunol. 2004, 25, 677–686. [Google Scholar] [CrossRef] [PubMed]
- Bystry, R.S.; Aluvihare, V.; Welch, K.A.; Kallikourdis, M.; Betz, A.G. B cells and professional APCs recruit regulatory T cells via CCL4. Nat. Immunol. 2001, 2, 1126–1132. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Moral, L.; Paul, T.; Martori, C.; Font-Díaz, J.; Sanjurjo, L.; Aran, G.; Téllez, T.; Blanco, J.; Carrillo, J.; Ito, M.; et al. Macrophage CD5L is a target for cancer immunotherapy. EBioMedicine 2023, 91, 104555. [Google Scholar] [CrossRef] [PubMed]
- Sanjurjo, L.; Aran, G.; Téllez, É.; Amézaga, N.; Armengol, C.; López, D.; Prats, C.; Sarrias, M.-R. CD5L Promotes M2 Macrophage Polarization through Autophagy-Mediated Upregulation of ID3. Front. Immunol. 2018, 9, 480. [Google Scholar] [CrossRef] [PubMed]
- Schoeps, B.; Frädrich, J.; Krüger, A. Cut loose TIMP-1: an emerging cytokine in inflammation. Trends Cell Biol. 2022, 33, 413–426. [Google Scholar] [CrossRef] [PubMed]
- Díaz, P.; Harris, J.; Rosario, F.J.; Powell, T.L.; Jansson, T.; Mohan, R.; Baumann, D.; Alejandro, E.U.; Gandhi, K.; Li, C.; et al. Increased placental fatty acid transporter 6 and binding protein 3 expression and fetal liver lipid accumulation in a mouse model of obesity in pregnancy. Am. J. Physiol. Integr. Comp. Physiol. 2015, 309, R1569–R1577. [Google Scholar] [CrossRef] [PubMed]
- Cathcart, M.-C.; O'Byrne, K.J.; Reynolds, J.V.; O'Sullivan, J.; Pidgeon, G.P. COX-derived prostanoid pathways in gastrointestinal cancer development and progression: Novel targets for prevention and intervention. Biochim. Biophys. Acta (BBA) Rev. Cancer 2012, 1825, 49–63. [Google Scholar] [CrossRef] [PubMed]
- Olson, D.M. The role of prostaglandins in the initiation of parturition. Best Pr. Res. Clin. Obstet. Gynaecol. 2003, 17, 717–730. [Google Scholar] [CrossRef] [PubMed]
- Wells, J.C. Thermal Environment and Human Birth Weight. J. Theor. Biol. 2002, 214, 413–425. [Google Scholar] [CrossRef]
- Basu, R.; Malig, B.; Ostro, B. High Ambient Temperature and the Risk of Preterm Delivery. Am. J. Epidemiology 2010, 172, 1108–1117. [Google Scholar] [CrossRef]
- Yackerson, N.; Piura, B.; Sheiner, E. The influence of meteorological factors on the emergence of preterm delivery and preterm premature rupture of membrane. J. Perinatol. 2008, 28, 707–711. [Google Scholar] [CrossRef] [PubMed]
- Jiao, A.; Sun, Y.; Avila, C.; Chiu, V.; Slezak, J.; Sacks, D.A.; Abatzoglou, J.T.; Molitor, J.; Chen, J.-C.; Benmarhnia, T.; et al. Analysis of Heat Exposure During Pregnancy and Severe Maternal Morbidity. JAMA Netw. Open 2023, 6, e2332780–e2332780. [Google Scholar] [CrossRef]
- Turco, M.Y.; Moffett, A. Development of the human placenta. Development 2019, 146. [Google Scholar] [CrossRef]
- Akram, K.M.; Frost, L.I.; Anumba, D.O. Impaired autophagy with augmented apoptosis in a Th1/Th2-imbalanced placental micromilieu is associated with spontaneous preterm birth. Front. Mol. Biosci. 2022, 9, 897228. [Google Scholar] [CrossRef] [PubMed]
- Buthmann, J.; Huang, D.; Casaccia, P.; O’neill, S.; Nomura, Y.; Liu, J. Prenatal Exposure to a Climate-Related Disaster Results in Changes of the Placental Transcriptome and Infant Temperament. Front. Genet. 2022, 13, 887619. [Google Scholar] [CrossRef]
- Ghosh, S.; Park, C.-H.; Lee, J.; Lee, N.; Zhang, R.; Huesing, C.; Reijnders, D.; Sones, J.; Münzberg, H.; Redman, L.; et al. Maternal cold exposure induces distinct transcriptome changes in the placenta and fetal brown adipose tissue in mice. BMC Genom. 2021, 22, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Thomson, A.J.; Telfer, J.F.; Young, A.; Campbell, S.; Stewart, C.J.; Cameron, I.T.; Greer, I.A.; Norman, J.E. Leukocytes infiltrate the myometrium during human parturition: Further evidence that labour is an inflammatory process. Hum. Reprod. 1999, 14, 229–236. [Google Scholar] [CrossRef]
- Romero, R.; Dey, S.K.; Fisher, S.J. Preterm labor: One syndrome, many causes. Science 2014, 345, 760–765. [Google Scholar] [CrossRef]
- Gomez-Lopez, N.; StLouis, D.; Lehr, M.A.; Sanchez-Rodriguez, E.N.; Arenas-Hernandez, M. Immune cells in term and preterm labor. Cell Mol. Immunol. 2014, 11, 571–581. [Google Scholar] [CrossRef]
- Abu-Raya, B.; Michalski, C.; Sadarangani, M.; Lavoie, P.M. Maternal Immunological Adaptation During Normal Pregnancy. Front. Immunol. 2020, 11, 575197. [Google Scholar] [CrossRef]
- Gómez-Chávez, F.; Correa, D.; Navarrete-Meneses, P.; Cancino-Diaz, J.C.; Cancino-Diaz, M.E.; Rodríguez-Martínez, S. NF-κB and Its Regulators During Pregnancy. Front. Immunol. 2021, 12, 679106. [Google Scholar] [CrossRef] [PubMed]
- Elliott, C.; Allport, V.; Loudon, J.; Wu, G.; Bennett, P. Nuclear factor-kappa B is essential for up-regulation of interleukin-8 expression in human amnion and cervical epithelial cells. Mol. Hum. Reprod. 2001, 7, 787–790. [Google Scholar] [CrossRef] [PubMed]
- Lindström, T.M.; Bennett, P.R. The role of nuclear factor kappa B in human labour. Reproduction 2005, 130, 569–581. [Google Scholar] [CrossRef]
- Noguchi, T.; Sado, T.; Naruse, K.; Shigetomi, H.; Onogi, A.; Haruta, S.; Kawaguchi, R.; Nagai, A.; Tanase, Y.; Yoshida, S.; et al. Evidence for Activation of Toll-Like Receptor and Receptor for Advanced Glycation End Products in Preterm Birth. Mediat. Inflamm. 2010, 2010, 1–10. [Google Scholar] [CrossRef]
- Zarember, K.A.; Godowski, P.J. Tissue Expression of Human Toll-Like Receptors and Differential Regulation of Toll-Like Receptor mRNAs in Leukocytes in Response to Microbes, Their Products, and Cytokines. J. Immunol. 2002, 168, 554–561. [Google Scholar] [CrossRef]
- Schaefer, T.M.; Fahey, J.V.; Wright, J.A.; Wira, C.R. Innate Immunity in the Human Female Reproductive Tract: Antiviral Response of Uterine Epithelial Cells to the TLR3 Agonist Poly(I:C). 2005, 174, 992–1002. [Google Scholar] [CrossRef] [PubMed]
- Ito, M.; Nakashima, A.; Hidaka, T.; Okabe, M.; Bac, N.D.; Ina, S.; Yoneda, S.; Shiozaki, A.; Sumi, S.; Tsuneyama, K.; et al. A role for IL-17 in induction of an inflammation at the fetomaternal interface in preterm labour. J. Reprod. Immunol. 2010, 84, 75–85. [Google Scholar] [CrossRef] [PubMed]
- Starodubtseva, N.; Nizyaeva, N.; Baev, O.; Bugrova, A.; Gapaeva, M.; Muminova, K.; Kononikhin, A.; Frankevich, V.; Nikolaev, E.; Sukhikh, G. SERPINA1 Peptides in Urine as A Potential Marker of Preeclampsia Severity. Int. J. Mol. Sci. 2020, 21, 914. [Google Scholar] [CrossRef]
- Tiensuu, H.; Haapalainen, A.M.; Tissarinen, P.; Pasanen, A.; Määttä, T.A.; Huusko, J.M.; Ohlmeier, S.; Bergmann, U.; Ojaniemi, M.; Muglia, L.J.; et al. Human placental proteomics and exon variant studies link AAT/SERPINA1 with spontaneous preterm birth. BMC Med. 2022, 20, 1–23. [Google Scholar] [CrossRef]
- Tissarinen, P.; Tiensuu, H.; Haapalainen, A.M.; Ronkainen, E.; Laatio, L.; Vääräsmäki, M.; Öhman, H.; Hallman, M.; Rämet, M. Maternal serum alpha-1 antitrypsin levels in spontaneous preterm and term pregnancies. Sci. Rep. 2024, 14, 1–9. [Google Scholar] [CrossRef]
- Wu, J.; Zhang, W.; Li, C. Recent Advances in Genetic and Epigenetic Modulation of Animal Exposure to High Temperature. Front. Genet. 2020, 11, 653. [Google Scholar] [CrossRef] [PubMed]
- Tessadori, F.; Duran, K.; Knapp, K.; Fellner, M.; Smithson, S.; Meireles, A.B.; Elting, M.W.; Waisfisz, Q.; O’donnell-Luria, A.; Nowak, C.; et al. Recurrent de novo missense variants across multiple histone H4 genes underlie a neurodevelopmental syndrome. Am. J. Hum. Genet. 2022, 109, 750–758. [Google Scholar] [CrossRef] [PubMed]
- Wood, E.M.; Hornaday, K.K.; Slater, D.M. Prostaglandins in biofluids in pregnancy and labour: A systematic review. PLOS ONE 2021, 16, e0260115. [Google Scholar] [CrossRef] [PubMed]
- Palliser, H.K.; Kelleher, M.A.; Welsh, T.N.; Zakar, T.; Hirst, J.J. Mechanisms Leading to Increased Risk of Preterm Birth in Growth-Restricted Guinea Pig Pregnancies. Reprod. Sci. 2014, 21, 269–276. [Google Scholar] [CrossRef] [PubMed]
- Phillips, R.J.; A Fortier, M.; Bernal, A.L. Prostaglandin pathway gene expression in human placenta, amnion and choriodecidua is differentially affected by preterm and term labour and by uterine inflammation. BMC Pregnancy Childbirth 2014, 14, 241–241. [Google Scholar] [CrossRef] [PubMed]
- Afgan, E.; Baker, D.; Batut, B.; van den Beek, M.; Bouvier, D.; Čech, M.; Chilton, J.; Clements, D.; Coraor, N.; Grüning, B.A.; et al. The Galaxyplatform for accessible, reproducible and collaborative biomedical analyses: 2018 update. Nucleic Acids Res. 2018, 46, W537–W544. [Google Scholar] [CrossRef] [PubMed]
- Li, H. Minimap2: Pairwise alignment for nucleotide sequences. Bioinformatics 2018, 34, 3094–3100. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.; Smyth, G.K.; Shi, W. feature Counts: An efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 2014, 30, 923–930. [Google Scholar] [CrossRef] [PubMed]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. EdgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef]
- Liao, Y.; Wang, J.; Jaehnig, E.J.; Shi, Z.; Zhang, B. WebGestalt 2019: Gene Set Analysis Toolkit with Revamped UIs and APIs. Nucleic Acids Res. 2019, 47, W199–W205. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Zhang, B.; Kirov, S.; Snoddy, J. WebGestalt: an integrated system for exploring gene sets in various biological contexts. Nucleic Acids Res. 2005, 33, W741–W748. [Google Scholar] [CrossRef] [PubMed]
- Montojo, J.; Zuberi, K.; Rodriguez, H.; Kazi, F.; Wright, G.; Donaldson, S.L.; Morris, Q.; Bader, G.D. GeneMANIA Cytoscape plugin: fast gene function predictions on the desktop. Bioinformatics 2010, 26, 2927–2928. [Google Scholar] [CrossRef] [PubMed]
- Tang, D.; Chen, M.; Huang, X.; Zhang, G.; Zeng, L.; Zhang, G.; Wu, S.; Wang, Y. SRplot: A free online platform for data visualization and graphing. PLOS ONE 2023, 18, e0294236. [Google Scholar] [CrossRef] [PubMed]
Figure 4.
Individual gene expression in preterm placentas. (A-L) Box plots with jitters showing expressions of a selected set of DEGs in the warm and cold groups of preterm placentas. Data are presented as median with IQR. Each dot represents an individual subject. p values were determined by the Wilcoxon signed-rank test. (M) STRING protein-protein interaction analysis between the HPGD and related proteins (STRING v12).
Figure 4.
Individual gene expression in preterm placentas. (A-L) Box plots with jitters showing expressions of a selected set of DEGs in the warm and cold groups of preterm placentas. Data are presented as median with IQR. Each dot represents an individual subject. p values were determined by the Wilcoxon signed-rank test. (M) STRING protein-protein interaction analysis between the HPGD and related proteins (STRING v12).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



