Submitted:
29 July 2024
Posted:
31 July 2024
You are already at the latest version
Abstract
Sodium hypochlorite (NaOCl) is widely used in public healthcare facilities; this exposure can result in the development of bacterial tolerance to disinfectant, which has known links to antibiotic cross-resistance. However, the mechanism through which cross-resistance to antibiotics and disinfectants develops remains ambiguous. Therefore, this study aimed to examine the phenotypic, and transcriptomic changes caused by disinfectant exposure in gram-negative bacteria and determine the cause of cross-resistance to antibiotics. The results demonstrated that the misuse of disinfectants plays an important role in the emergence of disinfectant resistance and in the increase in antibiotic resistance. Antibiotic resistance may occur from exposure of gram-negative bacteria to subminimal inhibitory concentrations (MICs) of NaOCl. Ten passages of gram-negative bacteria in increasingly higher subMICs of NaOCl disinfectant was sufficient to increase the MIC to >2,500 µg/ml NaOCl, particularly in K. pneumoniae and P. aeruginosa. To determine the development of cross-resistance to antibiotics due to NaOCl exposure, the MICs for each antibiotic before and after exposure of each strain to sublethal concentrations of NaOCl were compared. After overnight incubation with a sublethal concentration of NaOCl, a statistically significant increase in MIC was only observed for imipenem (P<0.01). Investigation of the mechanism of cross-resistance by means of transcriptome analysis revealed that 1,250 µg/ml NaOCl-adapted K. pneumoniae and P. aeruginosa strains increased resistance to imipenem due to increased expression of the resistance-nodulation-cell division (RND) efflux pumps, such as AcrAB-TolC and MexAB/XY-OprM. Therefore, we suggest that exposure to NaOCl can influence the expression of RND efflux pump genes, contributing to imipenem cross-resistance.
Keywords:
cross-resistance
; imipenem antibiotics
; NaOCl disinfectant
; resistance-nodulation-cell division (RND) efflux pump
1. Introduction
Due to the ongoing COVID-19 pandemic, the use of disinfectants has increased markedly. The most commonly used disinfectants for combating COVID-19 include quaternary ammonium compounds (QACs), sodium hypochlorite (NaOCl), hydrogen peroxide (H2O2), and ethanol [1]. The use of disinfectants has rapidly increased worldwide and has been associated with the accelerated emergence of antimicrobial resistance (AMR) in pathogenic microbes in the post-COVID-19 pandemic era [2,3,4]. The biocide tolerance and antibiotic resistance by one common mechanism exhibited by a certain microorganism can be attributed to cross-resistance between biocides and antibiotics [5]. In general, if a microorganism is intrinsically tolerant to certain QACs or acquires such tolerance after exposure, it is likely to exhibit cross-tolerance to various antimicrobial agents [3,6]. Among the 654 disinfectants listed by the Environmental Protection Agency of the USA for use against severe acute respiratory syndrome virus-2, QACs comprise 45.1% and chlorines (NaOCl and HOCl) comprise 17.3% of the active ingredients.
Chlorine-based compounds, particularly sodium hypochlorite (NaOCl) or bleach in households (4–5%), are the most frequently used disinfectants. NaOCl is widely used in healthcare facilities in diverse settings for spot disinfection of surfaces, such as countertops and floors. Chlorination is widely used as a disinfection treatment for water or wastewater, to remove pathogens and potential antibiotic-resistant bacteria [7]. This general use of chlorine products is attributable to several factors: their broad-spectrum antimicrobial activity, absence of toxic residues, effectiveness even when used with hard water, cost-effectiveness, rapid action, and ability to remove dried or fixed organisms and biofilms from surfaces [8]. The exact mechanism by which free chlorine destroys microorganisms has not been elucidated; however, several factors have been proposed, including oxidation of intracellular content, inhibition of protein synthesis, decreased oxygen uptake, oxidation of respiratory components, decreased ATP production, breaks in DNA, and inhibition of DNA synthesis [9]. However, commonly used chlorine disinfectants are less effective against highly chlorine-resistant waterborne bacteria, such as pathogenic Pseudomonas aeruginosa, which can exist in drinking tap water and resist disinfection.
Several shared resistance mechanisms have been reported for disinfectants and antibiotics [10,11,12]. Cross-resistance between antibiotics and disinfectants may occur via cellular mechanisms that protect against multiple classes of antimicrobial agents, or by the selection of genetic determinants for resistance to non-antibiotic agents that are linked to genes for antibiotic resistance [9]. Previous studies have shown that many Proteobacterial species that are considered to be critical priority AMR-pathogens (e.g., Enterobacteriales, Acinetobacter spp., and Pseudomonas spp.) are intrinsically tolerant to higher concentrations of chlorine [13,14]. These species have demonstrated the ability to adapt to NaOCl upon prolonged or repeated exposure to sublethal NaOCl concentrations, ultimately leading to increased disinfectant tolerance, biocide cross-tolerance, and cross-resistance to clinically relevant antibiotics [2]. The annual global use of disinfectants being more prevalent than that of antibiotics remains a major concern [13,15]. Such overuse of disinfectants has made these compounds common pollutants in ecosystems.
In gram-negative bacteria, the most clinically relevant efflux pumps are resistance–nodulation–division (RND) family members, which recognize a broad range of substrates, including antibiotics and disinfectants such as QACs/chlorhexidine [16]. This family includes well-characterized members of the Enterobacteriaceae multidrug-resistant (MDR) efflux pumps AcrAB-TolC, MexAB-OprM from Pseudomonas aeruginosa, and AdeABC in Acinetobacter baumannii. These strains often demonstrate upregulation of MDR efflux pumps, such as AcrAB-TolC. While efflux is linked to increased biocide tolerance, little is known about the contributions of these individual efflux pumps to biocide tolerance.
Due to society’s reliance on and overuse of disinfectants, understanding how sodium hypochlorite (NaOCl) may drive antimicrobial resistance is crucial. Sustained exposure to sublethal levels of disinfectants can lead to MDR; however, the mechanism through which cross-resistance to antibiotics and disinfectants develops remains ambiguous. Therefore, we hypothesized that disinfectant-induced tolerance mechanisms (i.e., specific efflux pumps) to antibiotic cross-resistance could be more clearly elucidated by transcriptomic analysis. Therefore, this study tested the phenotypic, and transcriptomic changes caused by disinfectant exposure in gram-negative bacteria and to determine the cause of cross-resistance to antibiotics.
2. Results
2.1. Effect of Exposure to Disinfectant on Gram-Negative Bacteria
The MICs for NaOCl in 121 wild-type isolates of gram-negative bacteria were determined using the broth microdilution method. The NaOCl disinfectant MIC values of Enterobacteriaceae (including carbapenem-resistant) and carbapenem-resistant A. baumannii ranged from 250–500 µg/ml and 250–1,000 µg/ml, respectively. In particular, the MICs for P. aeruginosa strains were above the median of that for other species and ranged from 250 to 1,000 µg/ml of NaOCl.
Among the 121 strains (including reference strains), the MIC for NaOCl of 10 strains increased by a fold-change ≥ 2.5 after NaOCl exposure (Table 1). All E. coli and A. baumannii isolates showed a non-adaptive response (MIC increase < 2-fold). A strong and stable MIC change was observed for isolates of K. pneumoniae and P. aeruginosa. In the control experiment (passages without disinfectant exposure), no significant changes in the MICs were observed. As shown in Table 1, 10 passages of gram-negative bacteria cultured in gradually higher subMICs of the disinfectant was sufficient to increase the MIC for NaOCl up to 2,500 µg/ml. The stability of the NaOCl-adapted strains was tested in the absence of each disinfectant over five additional passages. The MIC of NaOCl was again tested at the end of these passages, and this value was still higher than the initial value.
2.2. Antibiotic Susceptibility Was Reduced According to the NaOCl Exposure
The MIC values before and after exposure to disinfectant were compared using the Mann–Whitney U test to determine the effect of overnight incubation of gram-negative bacteria with 1250 μg/ml of NaOCl on the MICs of antibiotics. As shown in Table 1, in isolates adapted to NaOCl, a statistically significant increase in MIC was observed for imipenem (P = 0.010, 83.3% of all the NaOCl-adapted strains). Particularly in the case of Enterobacteriaceae, NaOCl significantly increased the MIC for imipenem only (P = 0.006). We did not observe any changes in the MIC of P. aeruginosa with high levels of carbapenem resistance for imipenem.
2.3. Cross-resistance between NaOCl disinfectant and β-lactams in Klebsiella pneumoniae
The KEGG pathway analysis was performed to explore the biological functions and pathways of DEGs. In the KEGG analysis of adapted and wild-type K. pneumoniae Z0318KP0159, 951 DEGs were identified and assigned to 93 pathways. Between adapted and wild-type K. pneumoniae Z0318KP0107, 1,472 DEGs in 110 pathways were identified. The significantly upregulated gene-enriched pathways included the two-component system (Bonferroni corrected P = 7.7E-16), phosphotransferase system (PTS, Bonferroni corrected P = 1.6E-16), and oxidative phosphorylation (Bonferroni corrected P = 9.2E-9), which includes energy metabolism (Figure S1). The most notable result involved the up-regulation genes annotated in the “β-lactam-resistance” pathway (Bonferroni corrected P = 0.01). The β-lactam-resistance pathway included 8 and 10 DEGs in Z0317KP0159 and Z0318KP0107, respectively (Figure S1).
The overexpression of RND efflux pump genes in K. pneumoniae Z0317KP0159 and Z0318KP0107 after exposure to 1,250 μg/ml NaOCl promoted resistance to β-lactams (Table 2). Expression of AcrAB-TolC genes, associated with the RND efflux pump, was increased by at least two-fold, except for tolC in K. pneumoniae Z0318KP0107. Moreover, the β-lactam-resistance pathway of Z0317KP0107 was significantly upregulated (P < 0.001). Treatment of K. pneumoniae Z0318KP0107 with NaOCl induced overexpression of the MarRAB operon (67.7-fold increase in marA expression), a global antibiotic-resistance regulator involved in the production of the AcrAB-TolC efflux pump that extrudes antibiotics. As shown in Table 1, antibiotic phenotypic characteristics after exposure to 1,250 μg/ml NaOCl also significantly reduced susceptibility to imipenem by using Mann-Whitney U test (P = 0.006). This implied that increased resistance to β-lactam antibiotics (including carbapenem) and overexpression of AcrAB-TolC were associated with NaOCl exposure. In K. pneumoniae, β-lactamase and carbapenemase genes showed strain-specific differences in expression after NaOCl exposure.
2.4. Cross-Resistance between NaOCl Disinfectant and β-Lactams in Pseudomonas aeruginosa
In the KEGG pathway analysis of adapted and wild-type P. aeruginosa ATCC 27853, 1,328 DEGs were identified and assigned to 107 pathways, while 1,332 DEGs, assigned to 110 pathways, were identified in P. aeruginosa Z0219PA0007, and 1,309 DEGs assigned to 104 pathways were identified in P. aeruginosa Z0217PA0020. As shown in Figure S2, the expression of genes was significantly related to biofilm formation in P. aeruginosa Z0219PA0007, in which the upregulated gene-enriched pathways were associated with the two-component system (Bonferroni corrected P = 2.3E-25), biofilm formation (Bonferroni corrected P = 2.2E-23), bacterial secretion system (Bonferroni corrected P = 8.2E-23), and quorum sensing (Bonferroni corrected P = 5.5E-5). Likewise, the most notable change in Pseudomonas, as shown in Figure S2, was the upregulation of the pathway related to β-lactam-resistance (Bonferroni corrected P = 9.7E-4 in P. aeruginosa Z0219PA0007). β-lactam-resistance was associated with many DEGs: ATCC 27853, Z0219PA0007, and Z0217PA0020 showed upregulation of 12, 16, and 17 DEGs, respectively.
Overexpression of the RND efflux pump genes in P. aeruginosa ATCC 27853, Z2019PA0007, and Z0217PA0020 after exposure to 1,250 μg/ml NaOCl also promoted resistance to β-lactam antibiotics (Table 3). The expression of MexAB-XY efflux pump genes associated with the RND efflux pump was increased by at least two-fold, except for mexAB-oprM in P. aeruginosa ATCC 27853. As shown in Figure S2, the β-lactam-resistance pathways of Z2019PA0007 and Z0217PA0020 were significantly upregulated (P < 0.001). In particular, NaOCl treatment of the three adapted P. aeruginosa strains commonly induced overexpression of armZ-mexXY-oprM (2.09–21.02-fold increase), which encode a global antibiotic-resistance regulator/modulator involved in the production of the MexXY-OprM efflux pump that extrudes antibiotics. In P. aeruginosa ATCC 27853, MexEF-OprN efflux pump gene expression increased by at least 2-fold, whereas expression of ParRS regulator genes decreased.
NaOCl exposure (adaptation) in P. aeruginosa also altered the gene expression related to the AmpC-AmpR-AmpG pathway. However, AmpC overexpression for β-lactamase production was not observed in this study. The genes associated with β-lactamase resistance in P. aeruginosa also exhibited strain-specific differential expression upon exposure to NaOCl exposure. This confirmed that increased resistance to β-lactams (including carbapenem) and overexpression of the RND efflux pump were linked to NaOCl tolerance in P. aeruginosa and K. pneumoniae.
3. Discussion
Authors Household items that contain disinfectants may be used “inadequately” by consumers, and diluted products and/or residues may allow for the growth of multidrug efflux pump-hyper-expressing strains that are concomitantly multidrug-resistant, which may pose a pressing epidemiological issue. Processes that are demonstrated in a laboratory may also be reproduced by humans and in the environment. Thus, in this study, the initial MIC values of unadapted strains for disinfectant were compared with the MIC values of NaOCl-adapted strains. As shown in Table 1, NaOCl-adapted gram-negative bacteria were not killed by sublethal or recommended disinfectant concentrations (500–5,000 μg/ml). We demonstrated that 10 passages of gram-negative bacteria in increasingly higher sublethal MICs of NaOCl disinfectant was sufficient to increase the MIC for NaOCl to > 2,500 µg/ml, particularly in K. pneumoniae and P. aeruginosa. Moreover, the MICs of each of the tested strains for a range of antibiotics before and after exposure to sublethal concentrations of NaOCl were compared. A statistically significant increase in MIC was only observed for imipenem (P < 0.010).
In a previous study, 5,000 μg/ml of NaOCl showed a lethal effect on 94.1% of P. aeruginosa isolates [17]. Ni et al. recommended that disinfectant concentrations of chorine-containing disinfectants for carbapenem-resistant K. pneumoniae (CRKP) disinfection be set at 2,000–5,000 μg/ml in China [18]. In addition, Kanamori et al. demonstrated that disinfectants commonly used in healthcare facilities would likely be effective (> 3log10 reduction) against carbapenem/colistin-resistant Enterobacteriaceae when used at appropriate concentrations, such as ≥ 5,000 μg/ml NaOCl [19]. These results indicated that NaOCl should not be used at sublethal concentrations in order to lower the risk of development of bacterial tolerance and resistance to antibiotics.
Whole genome sequencing (WGS) was conducted to elucidate the gene differences among wild and NaOCl-adapted K. pneumoniae and P. aeruginosa strains, respectively (data not shown). The average nucleotide identity (OrthoANI) analysis of the draft genomes (>99.9% similarity) suggests that wild and NaOCl-adapted K. pneumoniae or P. aeruginosa strains are the same, respectively. The draft genomes of wild- and NaOCl-adaptive K. pneumoniae contained genes conferring resistance to β-lactam (blaSHV) and disinfectant (qacC) on the chromosome. In addition, wild and NaOCl-adapted K. pneumoniae strains carried carbapenem resistance genes (blaKPC-2) on the IncX3 plasmid as well as various β-lactam resistance genes (blaTEM,/blaCTX-M-1/blaOXA-1) on the IncFIIK plasmid. Wild- and adaptive-strains in Pseudomonas spp., multidrug resistance genes (blaOXA-133, cat, van, etc.) were only detected on the chromosomes. As a result of comparative genomic analysis of the wild and the NaOCl-adapted strain, mutations were observed only in the cording region of the almost hypothetical proteins and the partial ribosomal RNA. However, transcriptome analysis revealed that 1,250 µg/ml NaOCl-adapted K. pneumoniae and P. aeruginosa strains increased resistance to β-lactam antibiotics (particularly imipenem) due to increased expression of the RND superfamily efflux pumps, such as AcrAB-TolC and MexAB/XY-OprM. In the case of NaOCl exposed, only blaKPC-2 gene was overexpressed among the β-lactam resistance genes on the plasmid of K. pneumoniae Z0318KP0107 (Table 2). Besides, all wild- and adaptive-strains of Pseudomonas don’t have any plasmid. Therefore, our data might mean that disinfectant (NaOCl) raised cross-resistance more than co-resistance with antibiotics (β-lactams).
Bacterial efflux pumps with inherent/acquired biocide tolerance can reduce susceptibility to other biocides and induce cross-resistance to specific antibiotics [12]. On the contrary, mechanisms of tolerance to biocides, such as permeability, degradation, and mutation can also result in an increase in tolerance or lead to cross-resistance. This report also mentioned that increased resistance to other biocides and cross-resistance to certain antibiotics is possible if phenotypic changes and induction occur due to biocide exposure. Some efflux pumps are upregulated by cationic disinfectants, which contributes to the antimicrobial resistance phenotype, and the role of these efflux pumps in cross-resistance to other disinfectants and antibiotics has been explored [16]. This study aimed to investigate the tolerance of gram-negative bacteria isolated from humans and the environment to NaOCl and evaluate cross-resistance to antibiotics after exposure to this disinfectant. Adaptation and tolerance to QACs and chlorhexidine are attributed to the presence and upregulation of specific efflux pumps, such as the small multidrug resistance pump [2,20], while NaOCl induces the expression of many functional gene families, including those associated with responses to oxidative stress, DNA repair, energy metabolism, membrane damage, and efflux pumps [21].
In particular, the RND family of efflux pumps strongly promote inherent antibiotic-resistant gram-negative bacteria. These pumps are composed of three components spanning the inner and outer membranes and export a broad spectrum of antibiotics and biopharmaceuticals [22,23]. The components of the RND efflux pump include an inner membrane pump protein specific to a particular substrate, an outer membrane protein, and a periplasmic accessory protein that binds to both the inner and outer membrane proteins, allowing extrusion of substrates from the cell. AcrAB-TolC and MexAB-OprM are the major RND efflux systems present in E. coli and P. aeruginosa and are essential for their survival, colonization, and virulence [24]. In the present study, we revealed the effect of NaOCl disinfectant on the promotion of microbial tolerance to disinfectant and antibiotic-resistance in K. pneumoniae and P. aeruginosa.
Among RND pumps in the Enterobacteriaceae, AcrAB-TolC is the most clinically important antibiotic efflux pump [25]. Our results, summarized in Figure 1/S1 and Tables 1/2/S1, show that 1,250 μg/ml NaOCl-adapted K. pneumoniae Z0317KP0159 and Z0317KP0107 showed increased resistance to β-lactam antibiotics (particularly imipenem) due to increased expression of the AcrAB-TolC efflux pump system. The AcrAB-TolC efflux pump has three global regulators: MarA, SoxS, and Rob [25]. Multiple regulators play important roles in promoting the expression of acrA/B, tolC, and micF, which are genes in the marA-soxS-rob regulon. The micF transcript inhibits the translation of ompF porin mRNA, which provides an entry channel for small hydrophilic antibiotics (β-lactams, aminoglycosides, and colistin). The multiple antibiotic-resistance (mar) locus mediates resistance primarily by up-regulating efflux of some antibiotics, disinfectants, and organic solvents via the AcrAB-TolC efflux pump and down-regulating influx through the outer membrane protein F [26]. Encoded by a mar locus containing marR/A/B, MarA positively regulates the expression of marR/A/B and many other genes (acrA/B, tolC, micF, etc.). SoxS and the Rob activator also stimulate the expression of many genes under the mar regulon. In addition, SoxR is activated by superoxide compounds, such as NaOCl and H2O2 [11]. Oxidation of SoxR induces activation of a second redox sensor, SoxS, which induces the transcription of several genes (manganese superoxide dismutase, ferredoxin, micF, etc.).
We confirmed that AcrAB-TolC efflux pump-related genes were overexpressed in 1,250-μg/ml NaOCl-adapted K. pneumoniae, by using qRT-PCR (Table S1). In particular, the regulatory gene marR showed decreased expression, whereas the expression of the positive regulatory gene marA, which serves downstream of these regulators, was increased. Consequently, the expression of genes encoding multidrug efflux pumps (acrA/B, tolC) was increased. In contrast, expression levels of acrR and soxR were upregulated or had different regulatory levels, depending on the strain. Therefore, we concluded that the overexpression of marA in K. pneumoniae strains made the adapted strains less sensitive to the effects of NaOCl. Our results also confirmed that the overexpression of marRAB led to cross-resistance between NaOCl and imipenem. These results are similar to the previous hypothesis by Randall and Woodward and Chetri et al. [26,29]. Although the level of antibiotic resistance conferred by marRAB is relatively low, increasing evidence suggests that marRAB and related systems are important for clinical antibiotic resistance, likely serving as a ‘stepping stone’ to achieve higher levels of resistance such as those of carbapenems.
Under various conditions, these multiple regulatory mechanisms can induce cross-resistance to NaOCl and imipenem by allowing simultaneously decreased influx (via the OmpF porin) and increased efflux (via AcrAB-TolC) of antimicrobial agents. Recent studies have suggested that chlorhexidine-adapted strains of K. pneumoniae are cross-resistant to other biocides and antibiotics, presumably because upregulation of acrAB and ramA in turn activate the AcrAB-TolC efflux pump [30,31]. In both these studies, the activation of AcrAB-TolC resulted in reduced susceptibility of K. pneumoniae to several antibiotics and biocides, including chlorhexidine, triclosan, and QACs. This result is consistent with our findings. In summary, we hypothesized that NaOCl exposure could influence gene expression, particularly those related to the AcrAB-TolC efflux pump of the RND family in K. pneumoniae, contributing to imipenem resistance. Based on this hypothesis, we described the gene expression levels of NaOCl–imipenem cross-resistance involving (1) a regulator (especially MarA), (2) a drug transporter and efflux pump, (3) cell membrane structure and transporter protein, and (4) loss of porin, in Figure 1.
Among the RND pumps in P. aeruginosa, the most clinically important antibiotic efflux pumps are MexAB-OprM, MexXY-OprM, MexCD-OprJ, and MexEF-OprN [25]. Verdial et al. described the overexpression of RND efflux pump systems as a common intrinsic or acquired resistance trait in P. aeruginosa [32]. They reported that overexpression of MexAB-OprM and MexXY-OprM results in P. aeruginosa resistance to aminoglycosides and β-lactams, and that mexAB-oprM, mexCD-oprJ, and mexEF-oprN are among the most studied genes encoding regulators of QACs, chlorhexidine, and trichlosan tolerance. As summarized in Figure 2 and Table 1 and Table 3 our results showed that NaOCl-adapted P. aeruginosa Z0219PA0007 and Z0217PA0020 had reduced sensitivity to β-lactam antibiotics (particularly imipenem), due to increased expression of the MexAB-OprM and MexXY-OprM efflux pump systems. In the case of 1,250 μg/ml NaOCl-adapted P. aeruginosa ATCC 27853, decreased sensitivities to β-lactam antibiotics were due to an increase in the expression MexEF-OprN and MexXY-OprM. Hou et al. showed that chlorine-injured P. aeruginosa cells that were exposed to sublethal concentrations (4 μg/ml) of NaOCl developed increased resistance, by 1.4–5.6 fold, to ceftazidime, ampicillin, and chloramphenicol [33]. These results were confirmed by quantitative PCR, which showed that genes related to the MexEF-OprN efflux pump were overexpressed. Bubonja-Sonje et al. revealed that approximately 30% of 62 isolates (mostly obtained from intensive care unit patients, and with reduced carbapenem susceptibility) showed increased production of transcripts related to MexEF-OprN (from 4- to 19-fold in mexF mRNA transcripts as compared with a wild-type reference isolate) [34]. Li et al. reported that the MexEF-OprN efflux system was not well-expressed in wild-type P. aeruginosa, and thus, its inactivation led to little or no change in antibiotic susceptibility [25]. In P. aeruginosa, where even small antibiotics must slowly diffuse across the outer membrane (OM) via its slow porin, the active efflux of its major RND pump is very effective in increasing the MICs of antibiotics [28]. In addition, imipenem can penetrate the OM much more rapidly than can other antibiotics, by utilizing a specific channel, OprD. Dulyayangkul et al. reported that hypochlorite triggers overexpression of major facilitator superfamily (MFS) pumps in Pseudomonas aeruginosa [22]. They also reported that increasing the production of MexXY-mediated by ArmZ reduces antibiotic susceptibility. Our results did not indicate loss of OprD or other porin proteins related to porin transcription and β-lactam-resistance (Table 3). Based on our results, we suggest that the MexXY-OprM efflux pump of the RND family is involved in the cross-resistance to NaOCl and imipenem. In addition, as shown in Figure 2, NaOCl and imipenem cross-resistance involved local regulators and gene expression related to the RND (MexAB, MexXY, and MexEF) efflux pumps of P. aeruginosa. Taken together, our results suggest that NaOCl disinfectant exposure influences expression of genes that contribute to β-lactam (carbapenem, particularly imipenem) cross-resistance, and particularly those related to the RND efflux pump in gram-negative bacteria. This can provide useful information to identify efflux pump-related gene mutations and elucidate the molecular mechanism of cross-resistance to NaOCl and imipenem.
4. Materials and Methods
4.1. Isolation and Identification of Bacterial Strains
Overall, 117 bacterial strains were included in this study (Table S2). Of them, 91 isolates were obtained from humans, including 41 Escherichia coli, 9 Klebsiella pneumoniae, 26 A. baumannii, and 15 P. aeruginosa strains. Further, 26 isolates were obtained from hospital and livestock environments: 7 E. coli, 4 K. pneumoniae, 11 A. baumannii, and 4 P. aeruginosa. All isolates from humans were MDR, including resistance to carbapenems. Environmentally obtained strains of K. pneumoniae and A. baumannii are known to be MDR, including resistance to carbapenems. The standard strains included E. coli ATCC 25922 and ATCC 10536, A. baumannii ATCC 19606, and P. aeruginosa ATCC 27853.
4.2. Determination of Antibiotics and Disinfectant Minimum Inhibitory Concentrations (MICs)
Antimicrobial susceptibility testing was performed using the broth microdilution method with a customized Sensititre KRCDC2F panel (TREK Diagnostic Systems, East Grinstead, United Kingdom) in accordance with the guidelines established by the Clinical and Laboratory Standards Institute [36]. The following antimicrobial agents were tested: amikacin, ampicillin, azithromycin, cefotaxime, cefoxitin, ceftazidime, ceftriaxone, chloramphenicol, ciprofloxacin, colistin, imipenem, gentamicin, nalidixic acid, streptomycin, tetracycline, and trimethoprim/sulfamethoxazole. All experiments in this study were carried out with independent biological replicates at least 3 times.
For NaOCl, a household chlorine bleach (Clorox ≥ 4%, Yuhan Corp, Seoul, Korea) was used. To determine the initial exposure concentrations of the disinfectant, the MIC of NaOCl was tested. Briefly, an overnight bacterial culture was diluted with sterile 0.85% NaCl solution to a 0.5 McFarland standard. Then, 96-well plates were prepared and 90 μl of bacterial suspension (1.5 × 106 CFU/ml), containing 10 μl of serially, two-fold diluted NaOCl disinfectant in cation-adjusted Mueller–Hinton Broth (CA-MHB) medium, was added to each well. For the growth control group, 90 μl bacterial suspension was added to 10 μl MHB medium without NaOCl disinfectant. For the negative control group, 90 μl sterile 0.85% NaCl solution was used instead of bacterial suspension.
4.3. NaOCl Disinfectant Exposure and Bacterial Subculturing
To determine the adaptation to disinfectant, the strains were exposed to increasing sublethal concentrations of NaOCl. Each disinfectant concentration was continuously increased two-fold and the cells were subcultured for 2 weeks, using 96-well plates. Specifically, 10 μl overnight culture of wild-type gram-negative bacteria was first added to each of at least eight wells containing in 90 μl of CA-MHB medium with 2× sublethal concentration of NaOCl. These plates were incubated at 37 °C with shaking at 150 rpm for 24 h. When growth was observed at the highest inhibitory concentration, this culture suspension was inoculated into a well containing medium with a NaOCl concentration two-fold higher than the previous concentration. Finally, we selected disinfectant-adapted strains that increased the disinfectant MIC of the wild-type strain by at least two-fold and maintained the highest MICs for 10 continuous subcultures.
4.4. RNA Extraction, Transcriptome Sequencing, and Quantitative reverse Transcription Polymerase Chain Reaction
Wild-type strains were cultured in CA-MHB for 16–18 h at 37 ℃. Adapted isolates were cultured in CA-MHB supplemented with 1,250 μg/ml NaOCl for 24 h at 37 °C. The adapted strains included K. pneumoniae Z0317KP0159, K. pneumoniae Z0318KP0107, P. aeruginosa ATCC 27853, P. aeruginosa Z2017PA0020, and P. aeruginosa Z2019PA0007. Cell pellets derived from exponential phase cultures of each bacterial strain was stored at -80 °C in a 10X volume of the stabilization reagents RNALater (ThermoFisher Scientific, Waltham, MA, USA). DNA and RNA were extracted using the methods described below and sequenced.
Total RNA was extracted using the TRIzol reagent (Invitrogen, Waltham, MA, USA) according to the manufacturer’s protocol. RNA samples were purified using a RNeasy Mini Kit (Qiagen, Munchen, Germany), including on-column DNase digestion, according to the manufacturer’s protocol. RNA purity was assessed using a NanoDrop spectrophotometer (Thermo Fisher Scientific). The RNA concentration was measured using a Qubit RNA BR Assay Kit and a Qubit 4 Fluorometer (Invitrogen). The extracted RNA was used for strand-specific cDNA library construction and Illumina paired-end sequencing (HiSeq 2500; Illumina Inc., San Diego, CA, USA) at Macrogen Co. (Seoul, Korea). mRNA expression levels were normalized to fragments per kb of transcript per million mapped reads (FPKM). The Database for Annotation, Visualization, and Integrated Discovery (DAVID) database analysis used edgeR to identify genes that were differentially expressed (DEGs) between the wild-type and NaOCl-adapted strains. Potential targets of the DEGs were analyzed using the Gene Ontology term (GOTERM) and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway maps.
AcrAB-TolC pump-related gene expression (regulator/transporter) was quantified by modification of the quantitative reverse transcription polymerase chain reaction (qRT-PCR) method described by Jia et al. [27]. A significant effect on gene expression was deduced when the corresponding ratios exceeded 2.0. All reactions were performed in triplicate.
4.5. Statistical Analysis
The statistical significance of the MIC data (disinfectant-wild versus -adapted strain upon exposure to a specific antibiotic) was evaluated using the nonparametric Mann–Whitney U test. Significance was set at P < 0.05. KEGG enrichment analysis of the disinfectant-adapted strain was performed using Fisher’s exact test with the Bonferroni correction (P < 0.05). All statistical calculations were performed using SPSS version 24 (IBM Corp., Armonk, NY, USA).
5. Conclusions
Disinfectants have seen a rapid increase in use, particularly in recent years, due to the ongoing COVID-19 pandemic. The most used disinfectants for combatting COVID-19 include NaOCl, QACs, H2O2, and ethanol. The mode of action of these disinfectants, except for triclosan that has a single specific target, is nonspecific. The mechanism of resistance to disinfectants involves several transcriptional regulators, and no single gene has been linked to NaOCl resistance. NaOCl often leads to the overexpression of the efflux pump, which may confer resistance to multiple antimicrobials. Although RND efflux pumps are overexpressed in the presence of NaOCl, they are associated with increased cross-resistance to some β-lactam antibiotics (carbapenems, particularly imipenem) and other disinfectants.
In general, if recommended guidelines for the use of disinfectants are followed in a way that limits the exposure of bacteria to sublethal doses of disinfectants, the risk of developing resistance and cross-resistance could be eliminated. Therefore, the use of disinfectants that carry a high risk of antimicrobial resistance, such as NaOCl and QACs, in household products and over-the-counter medications should be reevaluated. The use of the disinfectant has increased substantially since the onset of the COVID-19 pandemic. Our findings highlight a need for monitoring the cross-resistance between different disinfectants (e.g., chlorhexidine, H2O2) and clinically important antibiotics (e.g., tigecycline, colistin). Additionally, the findings emphasize the importance of disinfectants in the development and spread of antibiotic-resistant bacteria.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figure S1: Kyoto Encyclopedia of Genes and Genomes enrichment histogram of Klebsiella pneumoniae of genes differentially expressed after exposure to 1,250 μg/ml NaOCl; Figure S2: Kyoto Encyclopedia of Genes and Genomes enrichment histogram of Pseudomonas aeruginosa of genes differentially expressed after exposure to 1,250 μg/ml NaOCl; Table S1: Fold-change in expression of genes related to the AcrAB-TolC efflux pump in 1,250 μg/ml NaOCl-adapted Klebsiella pneumoniae; Table S2: Results of antibiotic susceptibility tests of 117 strains of Gram-negative bacteria.
Author Contributions
Conceptualization, J.H.N.; validation, J.H.N.; formal analysis, J.H.N.; investigation, J.H.N.; data curation, J.H.N.; writing—original draft preparation, J.H.N.; writing—review and editing, J.H.N. and J.S.Y..; visualization, J.H.N.; supervision, J.S.Y..; project administration, J.H.N.; funding acquisition, J.S.Y. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by the “Korea National Institute of Health (KNIH)” Research Project (2020-NI-023-00).
Informed Consent Statement
Not applicable.
Data Availability Statement
All data generated or analyzed during this study are included in this published article and its supplementary information files.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- EPA, 2022. List N Advanced Search Page: Disinfectant for Coronavirus (COVID-19). Accessed 24 May 2022.
- Chen, B.; Han, J.; Dai, H.; Jia, P. Biocide-tolerance and antibiotic-resistance in community environments and risk of direct transfers to humans: Unintended consequences of community-wide surface disinfecting during COVID-19? Environ. Pollut. 2021, 283, 117074. [Google Scholar] [CrossRef] [PubMed]
- Lobie, T.A. ; Roba, A A.; Booth, J.A.; Kristiansen, K.I.; Aseffa, A.; Skarstad, K.; Bjørås, M. Antimicrobial resistance: A challenge awaiting the post-COVID-19 era. Int. J. Infect. Dis. 2021, 111, 322–325. [Google Scholar] [CrossRef] [PubMed]
- Mahoney, A.R.; Safaee, M.M.; Wuest, W.M.; Furst, A.L. The silent pandemic: Emergent antibiotic resistances following the global response to SARS-CoV-2. IScience 2021, 24, 102304. [Google Scholar] [CrossRef] [PubMed]
- Elekhnawy, E.; Sonbol, F.; Abdelaziz, A.; Elbanna, T. Potential impact of biocide adaptation on selection of antibiotic resistance in bacterial isolates. Future J. Pharm. Sci. 2020, 6, 1–10. [Google Scholar] [CrossRef]
- Mohapatra, S.; Yutao, L.; Goh, S.G.; Ng, C.; Luhua, Y.; Tran, N.H.; Gin, K.Y.H. Quaternary ammonium compounds of emerging concern: Classification, occurrence, fate, toxicity and antimicrobial resistance. J. Hazard. Mater. 2023, 445, 130393. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Wang, Y.; Lu, J.; Yu, Z.; Song, H.; Bond, P.L.; Guo, J. Chlorine disinfection facilitates natural transformation through ROS-mediated oxidative stress. ISME J. 2021, 15, 2969–2985. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention, 2016. National Center for Emerging and Zoonotic Infectious Diseases (NCEZID), Division of Healthcare Quality Promotion (DHQP). Page last reviewed September 18, 2016. https://www.cdc.gov/infectioncontrol/guidelines/disinfection/ disinfection-methods/chemical.html.
- Nasr, A.M.; Mostafa, M.S.; Arnaout, H.H.; Elshimy, A.A.A. The effect of exposure to subinhibitory concentrations of hypochlorite and quaternary ammonium compounds on antimicrobial susceptibility of Pseudomonas aeruginosa. Am. J. Infect. Control. 2018, 46, e57–e63. [Google Scholar] [CrossRef] [PubMed]
- Knauf, G.A.; Cunningham, A.L.; Kazi, M.I.; Riddington, I.M.; Crofts, A.A.; Cattoir, V.; Trent, M.S. , Davies, B.W. Exploring the antimicrobial action of quaternary amines against Acinetobacter baumannii. MBio. 2018, 9, e02394-17. [Google Scholar] [CrossRef]
- da Cruz Nizer, W.S.; Inkovskiy, V.; Overhage, J. Surviving reactive chlorine stress: Responses of gram-negative bacteria to hypochlorous acid. Microorganisms 2020, 8, 1220. [Google Scholar] [CrossRef]
- Scientific Committee on Emerging and Newly Identified Health Risks 2009. Assessment of the Antibiotic Resistance Effects of Biocides. Avalaible online at: http://ec.europa.eu/health/archive/.
- Jones, I.A.; Joshi, L.T. Biocide use in the antimicrobial era: a review. Molecules. 2021, 26, 2276. [Google Scholar] [CrossRef]
- Khan, S.; Beattie, T.K.; Knapp, C.W. Relationship between antibiotic-and disinfectant-resistance profiles in bacteria harvested from tap water. Chemosphere. 2016, 152, 132–141. [Google Scholar] [CrossRef]
- Lu, J.; Guo, J. Disinfection spreads antimicrobial resistance. Science. 2021, 371, 474–474. [Google Scholar] [CrossRef] [PubMed]
- Wand, M.E; Sutton, J.M. Efflux-mediated tolerance to cationic biocides, a cause for concern? Microbiology 2022, 168, 001263. [Google Scholar] [CrossRef] [PubMed]
- Mitiku, M.; Ali, S.; Kibru, G. Antimicrobial drug resistance and disinfectants susceptibility of Pseudomonas aeruginosa isolates from clinical and environmental samples in Jimma University specialized hospital, Southwest Ethiopia. Am J Biomed Life Sci. 2014, 2, 40–45. [Google Scholar] [CrossRef]
- Ni, L.; Zhang, Z.; Shen, R.; Liu, X.; Li, X.; Chen, B.; Wu, X.; Li, H.; Xie, X.; Huang, S. Disinfection Strategies for Carbapenem-Resistant Klebsiella pneumoniae in a Healthcare Facility. Antibiotics 2022, 11, 736. [Google Scholar] [CrossRef]
- Kanamori, H.; Rutala, W.A.; Gergen, M.F.; Sickbert-Bennett, E.E.; Weber, D.J. Germicidal activity against carbapenem/colistin-resistant Enterobacteriaceae using a quantitative carrier test method. Antimicrob. Agents Chemother. 2018, 62, 10–1128. [Google Scholar] [CrossRef] [PubMed]
- Bland, R.; Waite-Cusic, J.; Weisberg, A.J.; Riutta, E.R.; Chang, J.H.; Kovacevic, J. Adaptation to a commercial quaternary ammonium compound sanitizer leads to cross-resistance to select antibiotics in Listeria monocytogenes isolated from fresh produce environments. Front. Microbiol. 2022, 12, 4059. [Google Scholar] [CrossRef] [PubMed]
- Xiao, X.; Bai, L.; Wang, S.; Liu, L.; Qu, X.; Zhang, J.; Xiao, Y.; Tang, B.; Li, Y.; Yang, H.; Wang, W. Chlorine tolerance and cross-resistance to antibiotics in poultry-associated Salmonella isolates in China. Front. Microbiol. 2022, 12, 4363. [Google Scholar] [CrossRef]
- Dulyayangkul, P.; Satapoomin, N.; Avison, M.B.; Charoenlap, N.; Vattanaviboon, P.; Mongkolsuk, S. Over-expression of hypochlorite inducible Major Facilitator Superfamily (MFS) pumps reduces antimicrobial drug susceptibility by increasing the production of MexXY Mediated by ArmZ in Pseudomonas aeruginosa. Front. Microbiol. 2021, 11, 592153. [Google Scholar] [CrossRef]
- Seukep, A.J.; Mbuntcha, H.G.; Kuete, V.; Chu, Y.; Fan, E.; Guo, M.Q. What Approaches to Thwart Bacterial Efflux Pumps-Mediated Resistance? Antibiotics. 2022, 11, 1287. [Google Scholar] [CrossRef]
- Puzari, M.; Chetia, P. RND efflux pump mediated antibiotic resistance in Gram-negative bacteria Escherichia coli and Pseudomonas aeruginosa: a major issue worldwide. World J. Microbiol. Biotechnol. 2017, 33, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Li, X.Z., Elkins, C.A.; Zgurskaya, H.I. (Eds.). 2016. Efflux-mediated antimicrobial resistance in bacteria: mechanisms, regulation and clinical implications. Springer.
- Randall, L.P.; Woodward, M.J. The multiple antibiotic resistance (mar) locus and its significance. Res. Vet. Sci. 2002, 72, 87–93. [Google Scholar] [CrossRef] [PubMed]
- Jia, Y.; Lu, H.; Zhu, L. Molecular mechanism of antibiotic resistance induced by mono-and twin-chained quaternary ammonium compounds. Sci. Total Environ. 2022, 832, 155090. [Google Scholar] [CrossRef] [PubMed]
- Li, X.Z.; Plésiat, P.; Nikaido, H. The challenge of efflux-mediated antibiotic resistance in Gram-negative bacteria. Clin. Microbiol. Rev. 2015, 28, 337–418. [Google Scholar] [CrossRef] [PubMed]
- Chetri, S.; Das, B.J.; Bhowmik, D.; Chanda, D.D.; Chakravarty, A.; Bhattacharjee, A. Transcriptional response of mar, sox and rob regulon against concentration gradient carbapenem stress within Escherichia coli isolated from hospital acquired infection. BMC Res. Notes. 2020, 13, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Abbood, H.M.; Hijazi, K.; Gould, I.M. Chlorhexidine resistance or Cross-resistance, that is the question. Antibiotics. 2023, 12, 798. [Google Scholar] [CrossRef] [PubMed]
- Wand, M.E.; Darby, E.M.; Blair, J.M.; Sutton, J.M. Contribution of the efflux pump AcrAB-TolC to the tolerance of chlorhexidine and other biocides in Klebsiella spp. J. Med. Microbiol. 2022, 71, 001496. [Google Scholar] [CrossRef] [PubMed]
- Verdial, C.; Serrano, I.; Tavares, L.; Gil, S.; Oliveira, M. Mechanisms of Antibiotic and Biocide Resistance That Contribute to Pseudomonas aeruginosa Persistence in the Hospital Environment. Biomedicines. 2023, 11, 1221. [Google Scholar] [CrossRef]
- Hou, A.M.; Yang, D.; Miao, J.; Shi, D.Y.; Yin, J.; Yang, Z.W.; Shen, Z.Q.; Wang, H.R.; Qiu, Z.G.; Liu, W.L.; Li, J.W.; Jin, M. Chlorine injury enhances antibiotic resistance in Pseudomonas aeruginosa through over expression of drug efflux pumps. Water Res. 2019, 156, 366–371. [Google Scholar] [CrossRef]
- Bubonja-Sonje, M.; Matovina, M.; Skrobonja, I.; Bedenic, B.; Abram, M. Mechanisms of carbapenem resistance in multidrug-resistant clinical isolates of Pseudomonas aeruginosa from a Croatian hospital. Microb. Drug Resist. 2015, 21, 261–269. [Google Scholar] [CrossRef]
- Moradali, M.F.; Ghods, S.; Rehm, B.H. Pseudomonas aeruginosa lifestyle: a paradigm for adaptation, survival, and persistence. Front. Cell. Infect. Microbiol. 2017, 7, 39. [Google Scholar] [CrossRef]
- Clinical and Laboratory Standards Institute (CLSI). Performance standards for antimicrobial susceptibility testing. 31st ed. CLSI Supplement M100. Clinical and Laboratory Standards Institute, USA, 2021.
Figure 1.
Acquisition of cross-resistance to imipenem antibiotic and NaOCl disinfectant by ArcAB-TolC efflux pump-related gene expression in NaOCl-exposed Klebsiella pneumoniae. Gene expression prediction based on RNA-Seq and quantitative reverse-transcription polymerase chain reaction datasets of K. pneumoniae strains (Z0318KP0107 and Z0317KP0159). The depiction of the AcrAB-TolC efflux system was modified from Li et al., Jia et al., and Li et al. [25,27,28]. AcrAB-TolC is a tripartite complex formed by AcrA, a membrane fusion protein, AcrB, a cytoplasmic-membrane protein, and TolC, an outer membrane protein. acrA and acrB are part of the same operon, which is negatively regulated by the local repressor (blue semicircle) AcrR. The genes are represented as arrows, and their translated proteins are represented as green ovals (repressors) and red ovals (activators). The activation of acrAB-tolC and micF transcription occurs primarily because of the global regulatory proteins MarA, SoxS, and Rob (red ovals). The arrows in parenthesis represent down/up-regulation of gene expression. The bold red arrowed lines indicate that MarA increases its own transcription and activates the expression of acrAB-tolC and micF by NaOCl exposure in our study. Overall, under conditions of NaOCl exposure, these multiple regulation mechanisms can create cross-resistance by simultaneously allowing both decreased influx (via OmpF porin) and increased efflux (via AcrAB-TolC) of imipenem antibiotics.
Figure 1.
Acquisition of cross-resistance to imipenem antibiotic and NaOCl disinfectant by ArcAB-TolC efflux pump-related gene expression in NaOCl-exposed Klebsiella pneumoniae. Gene expression prediction based on RNA-Seq and quantitative reverse-transcription polymerase chain reaction datasets of K. pneumoniae strains (Z0318KP0107 and Z0317KP0159). The depiction of the AcrAB-TolC efflux system was modified from Li et al., Jia et al., and Li et al. [25,27,28]. AcrAB-TolC is a tripartite complex formed by AcrA, a membrane fusion protein, AcrB, a cytoplasmic-membrane protein, and TolC, an outer membrane protein. acrA and acrB are part of the same operon, which is negatively regulated by the local repressor (blue semicircle) AcrR. The genes are represented as arrows, and their translated proteins are represented as green ovals (repressors) and red ovals (activators). The activation of acrAB-tolC and micF transcription occurs primarily because of the global regulatory proteins MarA, SoxS, and Rob (red ovals). The arrows in parenthesis represent down/up-regulation of gene expression. The bold red arrowed lines indicate that MarA increases its own transcription and activates the expression of acrAB-tolC and micF by NaOCl exposure in our study. Overall, under conditions of NaOCl exposure, these multiple regulation mechanisms can create cross-resistance by simultaneously allowing both decreased influx (via OmpF porin) and increased efflux (via AcrAB-TolC) of imipenem antibiotics.
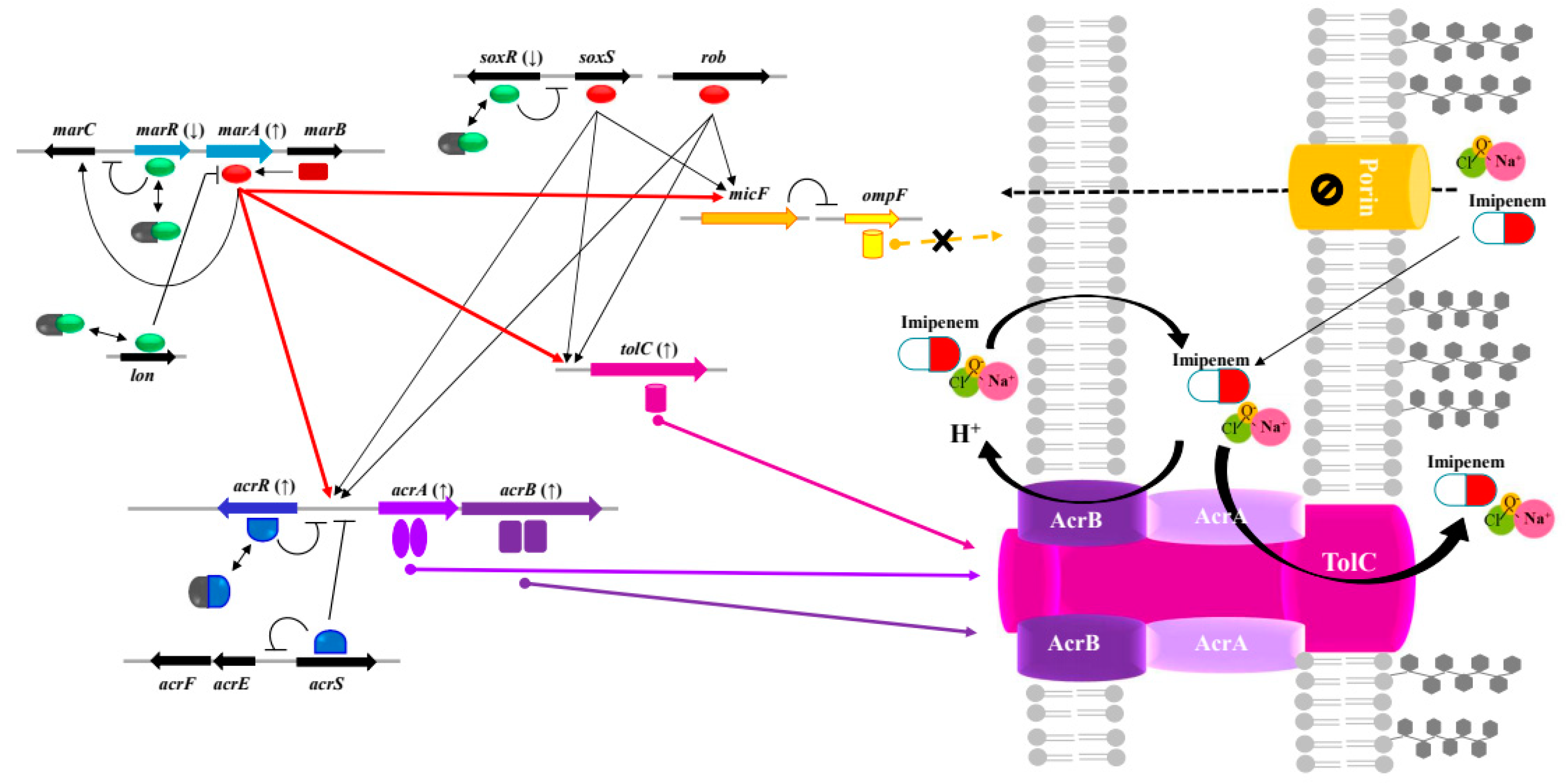
Figure 2.
Induction of cross-resistance to imipenem antibiotics by overexpression of the Mex-Opr efflux pump system in NaOCl-exposed Pseudomonas aeruginosa. Gene expression prediction was based on transcriptome analysis of P. aeruginosa (ATCC 27853, Z0219PA0007, and Z0217PA0020). The depiction of the efflux system was modified from Li et al. and Moradali et al. [25,28,35]. The genes are represented as arrows, and their translated proteins are represented as blue semicircles (repressors) and red ovals (activators). The arrows in parenthesis represent down/up-regulation of gene expression. The bold red arrowed lines indicates that AmrR and ArmZ increase their own transcription and activate the expression of mexAB/XY-oprM upon NaOCl exposure. Overall, under conditions of NaOCl exposure, these multiple regulation mechanisms can create cross-resistance by simultaneously allowing increased efflux of imipenem antibiotics.
Figure 2.
Induction of cross-resistance to imipenem antibiotics by overexpression of the Mex-Opr efflux pump system in NaOCl-exposed Pseudomonas aeruginosa. Gene expression prediction was based on transcriptome analysis of P. aeruginosa (ATCC 27853, Z0219PA0007, and Z0217PA0020). The depiction of the efflux system was modified from Li et al. and Moradali et al. [25,28,35]. The genes are represented as arrows, and their translated proteins are represented as blue semicircles (repressors) and red ovals (activators). The arrows in parenthesis represent down/up-regulation of gene expression. The bold red arrowed lines indicates that AmrR and ArmZ increase their own transcription and activate the expression of mexAB/XY-oprM upon NaOCl exposure. Overall, under conditions of NaOCl exposure, these multiple regulation mechanisms can create cross-resistance by simultaneously allowing increased efflux of imipenem antibiotics.
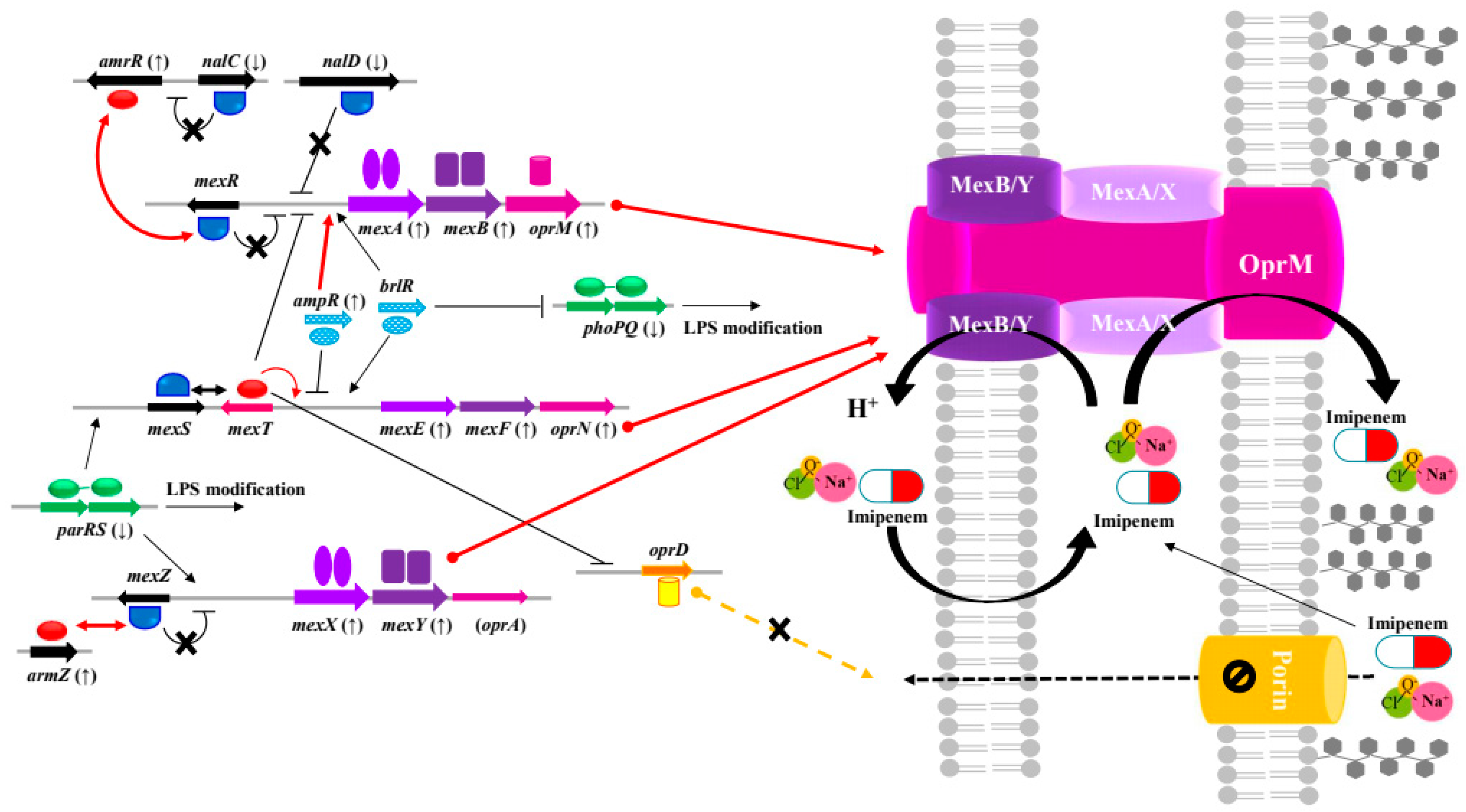

Table 1.
Evaluation of NaOCl minimal inhibitory concentration and antibiotic susceptibility changes with disinfectant exposure
Table 1.
Evaluation of NaOCl minimal inhibitory concentration and antibiotic susceptibility changes with disinfectant exposure
| Species | Strains | MICs (mg/l) |
Antibiotic susceptibility (Exposure to 1,250 µg/ml NaOCl) |
|||
| Before1 | After | Increased | Reduced | |||
|
K. pneumoniae |
CRKP | Z0317KP0159 | 500 | 2,500 | FOX, IMI, CHL | ND2 |
| Z0317KP0181 | 500 | 2,500 | IMI | GEN | ||
| Z0318KP0099 | 500 | 2,500 | IMI | STR | ||
| Z0318KP0107 | 500 | 2,500 | IMI, CHL, AZI | ND | ||
| Z0318KP0236 | 500 | 2,500 | IMI, CHL | GEN, AZI | ||
|
P. aeruginosa |
CSPA | ATCC 27853 | 500 | 2,500 | AXO, IMI, STR, COL |
TAZ, CIP, NAL, TET, GEN, AMI, SXT |
| Z0219PA0007 | 500 | 1,250 |
AXO, IMI, COL |
CIP, NAL, STR, SXT |
||
| CRPA | I0020PA0021 (HL-IMI) |
500 | 2,500 | X3 | X | |
| I0020PA0028 | 500 | 1,250 | IMI, CIP, NAL, TET, AMI, STR |
FOT, TAZ, AXO, GEN | ||
| Z0217PA0020 (HL-IMI) |
500 | 2,500 |
FOT, TAZ, CIP, CHL, GEN, STR, SXT |
NAL, AMI | ||
1 “Before” indicates no exposure to disinfectants (wild-type strain), and “After” indicates exposure to disinfectants (adapted-type strains). 2 ND: not detected. 3X: Exposure to disinfectant does not alter antibiotic susceptibility profiles.* Red words: Antibiotic susceptibility profiles changed “susceptible” to “resistant” (S→R). * Blue words: Antibiotic susceptibility profiles changed “resistant” to “susceptible” (R→S). * CRKP, carbapenem-resistant Klebsiella pneumoniae; CSPA, carbapenem-susceptible Pseudomonas aeruginosa; CRPA, carbapenem-resistant P. aeruginosa; FOT, cefotaxime; AXO, ceftriaxone; FOX, cefoxitin; TAZ, ceftazidime; AMP, ampicillin; IMI, imipenem; HL-IMI, high-level imipenem resistant; CIP, ciprofloxacin; NAL nalidixic acid; TET, tetracycline, CHL, chloramphenicol; GEN, gentamicin; AMI, amikacin; STR, streptomycin; AZI, azithromycin; COL, colistin; SXT, trimethoprim/sulfamethoxazole.
Table 2.
Fold-change of expression of genes related to β-lactam antibiotic-resistance in Klebsiella pneumoniae after exposure to 1,250 μg/ml NaOCl
Table 2.
Fold-change of expression of genes related to β-lactam antibiotic-resistance in Klebsiella pneumoniae after exposure to 1,250 μg/ml NaOCl
| Group | Function |
Gene locus tag (Gene name) |
Gene expression fold-change* |
||
|
Z0317 KP0159 |
Z0318 KP0107 |
||||
| Enzymatic degradation |
Class A carbapenemase Kpc-2 | KPHS_p200360 (blaKPC-2) |
-2.26 | 3.40 | |
| β-lactamase | KPHS_25220 (blaSHV) |
2.43 | -1.44 | ||
| KPHS_13880 | - | 2.71 | |||
| RND efflux pump |
Repression of porin OmpF (mar-sox-rob regulon activator) |
KPHS_25470 (marA) |
-1.44 | 67.73 | |
| Multidrug efflux membrane fusion protein | KPHS_11890 (acrA) |
1.46 | 5.18 | ||
| Multidrug efflux transporter | KPHS_11880 (acrB) |
2.19 | 2.30 | ||
| Multidrug efflux transporter (permease EefB) | KPHS_52090 (acrB) |
2.07 | 2.65 | ||
| Outer membrane channel protein |
KPHS_45760 (tolC) |
2.83 | 1.30 | ||
| Multidrug efflux transport outer membrane protein EefC | KPHS_52100 (adeK) |
2.13 | -1.16 | ||
| Porin | Outer membrane protein 1A/OmpK35 porin | KPHS_18370 (ompF) |
-2.45 | 1.52 | |
| Peptidoglycan synthetase, penicillin-binding protein 2 | KPHS_27710 (mrdA) |
2.54 | 4.02 | ||
* Difference in fold-change to antibiotic-resistance gene expression between before and after exposure to 1,250 µg/ml of NaOCl.
Table 3.
Fold-change in β-lactam-resistance gene expression in 1,250-µg/ml NaOCl-adapted Pseudomonas aeruginosa
Table 3.
Fold-change in β-lactam-resistance gene expression in 1,250-µg/ml NaOCl-adapted Pseudomonas aeruginosa
| Group | Function |
Gene locus tag (Gene name) |
Gene expression fold-change* |
|||
| ATCC 27853 | Z0219 PA0007 | Z0217 PA0020 | ||||
| β- lactamase |
AmpC beta-lactamase | PA4110 (ampC) |
- | -2.32 | -1.07 | |
| Beta-hexosaminidase | PA3005 (nagZ) |
4.30 | - | - | ||
| Transport of degraded muropeptides (GlcNAc-anhMurNAc) | PA4218 (ampG) |
-6.95 | 13.74 | 1.92 | ||
| RND efflux pump |
MexAB-OprM | MexR antirepressor ArmR | PA3719 (armR) |
- | 2.29 | 3.87 |
| Transcriptional regulator AmpR |
PA4109 (ampR) |
- | 2.31 | 2.62 | ||
| Transcriptional regulator |
PA3721 (nalC) |
-2.65 | - | - | ||
| Transcriptional regulator |
PA3574 (nalD) |
-2.39 | -2.33 | -1.75 | ||
| Multidrug resistance protein MexA | PA0425 (mexA) |
- | 2.50 | 2.03 | ||
| Multidrug resistance protein MexB | PA0426, PA4375 (mexB) |
- | 2.75 | 3.09 | ||
| Outer membrane protein OprM |
PA0427 (oprM) |
- | 2.09 | 3.06 | ||
| MexEF-OprN | Multidrug efflux membrane fusion protein MexE | PA2493 (mexE) |
11.88 | - | - | |
| Multidrug efflux transporter MexF |
PA2494 (mexF) |
4.11 | - | - | ||
| Transcriptional regulator AmpR |
PA2495 (oprN) |
9.75 | - | - | ||
| MexXY-(OprM) | Transcriptional regulator |
PA2020 (mexZ) |
- | 2.84 | -2.21 | |
| Two-component response regulator ParR | PA1799 (parR) |
-7.73 | -2.53 | -2.06 | ||
| Multidrug efflux membrane fusion protein | PA2019 (mexX) |
4.04 | 20.45 | 8.88 | ||
| Multidrug efflux transporter | PA2018 (mexY) |
2.32 | 21.02 | 8.00 | ||
| Outer membrane protein |
PA4144 (oprM) |
2.64 | 1.11 | -2.13 | ||
| MexZ antirepressor | PA5471 (armZ) |
9.59 | 4.59 | 10.34 | ||
| MexVW-OprM | Multidrug efflux membrane fusion protein MexV | PA4374 (mexV) |
- | - | - | |
| Multidrug efflux membrane protein | PA4375 (mexW) |
- | 1.31 | 2.62 | ||
| Outer membrane protein OprM | PA4974 (oprM) |
- | -2.63 | 5.65 | ||
| Porin | Porin D (imipenem) | PA0958 (oprD) |
-1.19 | 2.91 | ||
* The difference in fold-change to antibiotic-resistance gene expression between before and after exposure to 1,250 µg/ml of NaOCl.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



